
Submitted:
11 December 2025
Posted:
15 December 2025
You are already at the latest version
Abstract
Neural coding means the representation of external stimuli and/or behavioral processes in the electrical activity of nerve cells. In recent years, many facts have been revealed that indicate the need to generate oscillations for such a representation in a normal brain. The hippocampus and its directly related structures (dentate gyrus, subiculum, and entorhinal cortex) generate many types of field rhythmic activity, the main of which are theta (~4-12 Hz), beta (~15-30 Hz), gamma (~25-100 Hz), and ripple oscillations (~120-500 Hz). With the participation of oscillatory activity generated in these frequency bands, both spatial and non-spatial information (temporal, auditory, olfactory, tactile, gustatory, etc.) is represented in the brain. It has been found that oscillations underlie many critical brain functions such as learning and memorization. However, a fundamental question remains to be fully resolved: what specific role do different types of oscillations generated in the hippocampal system play in cognitive functions (in particular, in encoding information), and what are the mechanisms of their participation in these functions? This paper is devoted to a review of the literature data on the role of oscillatory processes in encoding signals entering the brain both from the external environment and from the body itself. The issues of the participation of oscillations in the memorization and reproduction of stored information are also discussed. The main focus is on examining oscillatory activity directly in the hippocampus; the material obtained from the study of structures belonging to the hippocampal system and some neocortical regions are also evaluated.
Keywords:
information
; place cells
; time cells
; rate of spikes
; time of spike generation
; oscillations
; synchronization
; coherence
1. Introduction
How the brain encodes external signals is a fundamental question in both philosophy and cognitive neuroscience. The concept of “neural coding” suggests that patterns of cellular activity represent some kind of external correlate of events. In its most direct interpretation, activation of a neuron or a population of neurons is understood as a signal about the presence and strength of an external stimulus or an internal signal. In addition, neural activation allows us to assess how the relationship between the outside world and activity changes during the learning process [1]. Thus, with a known approximation [2], neural coding refers to the processes by which external stimuli and/or acquired knowledge are reflected in neural activity and ultimately control behavior. Research in this area aims to explain these processes by identifying the mechanisms responsible for recognizing stimuli and executing behavioral responses. For example, the firing rate of a neuron may increase as the intensity of a stimulus increases. This dependence is often described by a mathematical function such as an activation function or a tuning curve. The concept of neural coding by means of a tuning function [3] suggests that neurons encode stimuli by comparing the characteristics of a stimulus with the firing rate. In addition to the firing rate, the generation time of action potentials (time coding, also known as phase coding) also plays a critical role in neural coding. Temporally-spatially correlated changes in the pulse mode in populations of neurons form the neural basis of sensory representations [4].
Both the frequency and time of spike generation in the hippocampal system depend on fine-tuning the functioning of the cells of its constituent structures; this, in turn, is determined by local field oscillations in these structures, since they rhythmically alter neural excitability [5,6].
Back in the middle of the last century, Adrian, working on the mammalian brain, showed that research behavior and cognitive task solving are accompanied by the appearance of rhythmic (oscillatory) activity [7]. Since then, many studies have been conducted demonstrating the connection of cognitive activity with brain rhythms, which have led to various hypotheses about the origin and functions of oscillations.
Several types of field rhythmic activity, or oscillations, are generated in the hippocampal system (the hippocampus proper, the dentate gyrus, the entorhinal cortex, and the subiculum). They are recorded both in humans in clinical conditions from the surface of the skull (EEG) or during neurosurgical operations by means of electrodes implanted in the brain (intracerebral EEG, iEEG), and in various animal species, from rodents to monkeys, in neurophysiological experiments (local field potentials, LFP). Numerous studies conducted in the registration of activity in the brain of mammals, including humans, as well as in model computational studies, have suggested the important role of oscillations in brain function, in particular, in the performance of cognitive functions.
At the basic level, neural oscillations reflect the synchronous and rhythmic switching of neural populations between states of high and low excitability [8,9]. As a result, most neurons in a group of functionally connected cells are more likely to generate action potentials during the high excitability phase. As for the role of oscillations in the cognitive work of the brain, they are supposed to be involved in performing many operations: selection of sensory information [9,10]; analysis/separation of complex input streams [11,12]; the ordering of information transmitted by firing rate by encoding the spike phase [13]. It was also assumed that at a certain frequency, the oscillations are dynamically synchronized in order to combine a group of neurons into a cellular structure (ensemble) to perform a specific brain task [14]. Oscillations also provide temporary coupling of distant neural ensembles to improve information transmission [15,16,17], encoding and extracting information from memory [17,18,19,20], as well as participation in the implementation of the processes of attention and memory themselves [10,19,20,21,22,23]. In this review, only individual functions performed with the participation of oscillations will be considered in detail, mainly encoding information. In addition, the unresolved question of the specific role of oscillations of the main types generated in the hippocampal system in cognitive functions, in particular, in encoding, consolidating, and extracting information, will be considered.
Understanding the mechanisms of these processes in detail is very important, since the hippocampus is one of the main areas involved in the development of dementia in Alzheimer’s disease, which is an increasing social and medical problem; a correct theory in this regard may lead to the creation of new approaches for the diagnosis and treatment of the early stages of this disease.
2. The Participation of Oscillatory Processes in Information Encoding
2.1. Types of Oscillatory Activity of the Hippocampal System and Their Relation to Information Processing
Oscillations of different frequencies involved in information processing and encoding are generated in the hippocampal system as a result of complex interactions within neural networks, which may include glutamatergic pyramidal neurons of hippocampal fields and granular and mossy cells, GABAergic interneurons (IN), intrahippocampal connections and external inputs (see [8,21,24,25,26,27,28,29]).
2.1.1. Types of Oscillations
Theta oscillations. The most distinct and high-amplitude rhythmic activity recorded in the hippocampus during exploratory behavior is the theta rhythm (~4-12 Hz), which has a sinusoidal shape (see Figure 1). It was first discovered when recording LFP in the hippocampus of rabbits by Jung and Kornmuller [30], where they occurred both spontaneously and in the form of reactions to sensory stimuli. It was later shown [31] that dominant theta activity can also be found in the human hippocampus.
In humans, as in rodents, theta oscillations are especially noticeable during movement [32], when selective attention is activated, but they can also be observed when motionless while solving various cognitive tasks [33], i.e., movement is not a prerequisite for their generation.
After the first attempts to analyze theta oscillations [34,35], over the next few decades, other researchers recorded theta rhythm in different animal species and proposed many functions for this type of oscillation, from general activation (arousal) to various aspects of learning and memory [10,21,32,33,36,37].
Beta oscillations. The beta rhythm (15-30 Hz) in the hippocampal system was first recorded in the early nineties in both freely moving [38] and anesthetized rats [39]; however, it was much more pronounced in the DG than in the CA1 field of the hippocampus [38,40]. The beta rhythm in the eye appeared simultaneously with that in the olfactory bulb (OLB), from which the eye receives information dysynaptically [41,42]. Other studies have shown that the beta rhythm in DG is not only synchronized with the simultaneously recorded beta rhythm in OLB, but is also controlled by it [43,44].
During the registration of LFP in the laboratory, when the rats performed various tasks for associative learning, the presentation of a stimulus caused a decrease in the power of the theta rhythm and a simultaneous increase in the amplitude of the beta rhythm. These changes persisted from the moment the stimulus was presented to the moment the animal received the reward and were not observed with similar behavior in the absence of appropriate stimuli. Thus, the observed opposite changes in the amplitude of theta and beta oscillations in the DG may reflect a shift in information processing that occurs when an animal encounters significant stimuli [45]. In this paper, the authors also suggested that the observed oscillatory dynamics in the hippocampus indicates that the hippocampus is shifting to a different state of information processing, with theta and beta rhythms acting through different neural mechanisms.
Gamma oscillations. In addition to the low-frequency theta rhythm, “fast” oscillatory activity (25-70 Hz) [46], now known as the gamma rhythm, has been studied in the hippocampus and the central nervous system [24,47]. The authors reported that the power of rapid activity was higher during walking and running (behavior associated with theta rhythm) compared with the power obtained during immobility [46] (Figure 1, A-D). Later it was shown that the power of gamma oscillations is not only higher during the behavior associated with the theta rhythm, but also that its power is maximal at a certain phase of the theta cycle [24,47,48] (Figure 1, B). Then it became known that the gamma rhythm band is somewhat wider, up to 100 Hz, in addition, it is divided into two main sub-bands: a slow gamma rhythm (~25-50 Hz) and a fast gamma rhythm (~55-100 Hz) [19]. It is assumed that the mechanisms of organization of slow and fast gamma oscillations are different, since they are generated by different neural networks: the “fast” gamma rhythm is associated with a fast gamma input from the medial entorhinal cortex (mEC) [49,50], while the “slow” gamma rhythm is controlled by intra-hippocampal inputs, and Specifically, in the CA1 field, where it is most pronounced, it depends on the signals coming from the CA3 field [49,50,51]. There is a report that the input signals from the mEC and the CA3 field arrive at the CA1 field in different phases of the theta cycle [52]. Based on this fact, the point of view was born that fast and slow gamma rhythms have different functions: while fast reflects the encoding of information, slow reflects its extraction [19]. However, this statement requires additional verification, since facts have been obtained that do not unequivocally confirm this point of view [53,54]. It is possible that the functions of gamma activity in the hippocampal system in their interaction with theta oscillations are more dynamic and diverse than previously assumed [55].
Ripple oscillations. While theta oscillations and the associated gamma rhythm are most often recorded during active (exploratory) behavior, with “automatic” behavior such as licking and chewing/drinking, as well as during quiet wakefulness, another pattern of LFP is recorded in the rat hippocampus, namely, high-amplitude irregular waves. This activity, first discovered by Vanderwolf [56], is now better known as sharp wave-ripples (SWRs), which are irregularly occurring waves with a duration of 40-100 ms, with superimposed oscillations with a frequency of ~120-500 Hz, most often observed during slow-wave sleep or in a state of immobility when the context is not changing [46,57,58,59]. Figure 1, F). In their foundational work on rats, Buzsáki and colleagues (1983) demonstrated [46] that during SWRs there were bursts of cell activity, usually discharged by “complex spikes”, considered as pyramidal neurons [60].
2.1.2. Oscillatory Activity and Signal Processing
It became known from [46] that in free-moving animals, hippocampal neurons discharged by complex spikes, i.e., pyramidal cells, “fire” in a certain phase of the theta cycle, followed by inhibition of activity in approximately the opposite phase of the theta wave; in this phase, inhibitory interneurons of the hippocampus usually fire, discharging single spikes [60]. The pyramidal neurons of the hippocampus are the “place cells”, or spatial cells, intensively studied to date, the discovery of which was the first step towards creating a “brain positioning system” [61,62]. These cells discharge in bursts of pulses at a certain point in space, increasing the firing rate by about 10 times (from 0.5 to 5 pulses/s) [63,64], i.e., the firing rate in this case encodes the location of the animal in space. It is important that such cells are registered in active, free-roaming animals. In this case, each of them is activated in a limited “field” of this neuron (Figure 2, A). The fields of the place show the current location of the animal [61] and together form a cartographic representation of the space (terrain map) [65]; see also [66,67,68,69]. Interestingly, place cells are stably discharged in their place fields during theta rhythm-related behavior (for example, running), but are often not activated in these fields when the animal is resting or sleeping, i.e., when the hippocampal network is in non-theta state [65]. Thus, in order for the neurons to activate, the location of the rat in a certain place in the environment is a necessary but insufficient condition; for this, the animal must behave in a certain way, i.e., be in a “theta state”.
Subsequently, it became known that the bursts of activity of pyramidal neurons during SWRs during sleep or during restful wakefulness correspond to a time-compressed reactivation of patterns of activity of site cells during active wakefulness. [58,59,70,71]. It has been hypothesized that this reactivation allows the hippocampus to transfer the experience originally stored in the hippocampus to the neocortex as part of the memory consolidation process that occurs during sleep [20,70]. In [70], it was shown for the first time that after active behavior, in subsequent sleep during SWRs, paired correlations between cells of the site observed during wakefulness are reactivated.
When the animal is in the field of the place, not only firing rate of the cells of the place increases, but also these discharges adjust to the phase of the theta wave (Figure 2, B). Thus, as the rat passes through the field of the place, these cells first generate impulses in the late phases of the theta cycle, and then gradually at earlier and earlier phases; this is due to the fact that the inter-spike time interval is slightly shorter than the generation time of the theta cycle [72,73]. This phenomenon, called theta phase precession, has been reproduced in many records (for example, [63,74,75,76,77]); it shows that the phase of spikes relative to network oscillations encodes space. This code can contribute to the construction of a cognitive map that allows objects to be associated with the animal’s location [78,79,80]. The theta rhythm phase, in which a place cell is discharged, predicts the animal’s position more accurately than the average frequency of neuron discharges [75]. This effect is called phase coding.
Place neurons are not independent elements in the hippocampus: several studies have demonstrated dynamic interaction between pyramidal neurons and interneurons (IN), as well as the involvement of IN in spatial coding [81,82,83,84]. For example, in [83] using in vivo electrophysiological recordings, a dynamic interaction between IN and place cells was revealed. The authors showed complementary spatial activation, namely, a negative spatial correlation between interneuron–pyramidal cell pairs. In the region where the place cell is activated, the paired interneuron is not activated, and vice versa, and this is observed in both monosynaptic and non-monosynaptic connections. Such an interaction is much more likely for nearby pyramidal cells and interneurons than for distant neurons [47]. Interestingly, one interneuron can have at least two negatively correlated site cells [83].
Recent studies have confirmed these previous findings, which assessed the relationship between pyramidal cells and IN. Recently, [85] used a method for labeling and recording the activity of a single pyramidal cell and its presynaptic neurons connected by direct synapses in the hippocampus of mice. In these experiments, the animals were allowed to run around the tape, with various tactile signals placed. This study demonstrated the formation and maintenance of the site field of this pyramidal cell and the dynamic interaction of neurons. The presented results refute the idea that the spatial modulation of interneuronal activation was just a reflection of the input signals received by the pyramidal cells. Thus, the cells of the site are regulated by IN networks that coordinate the synchronization and coordination of cellular ensembles for processing spatiotemporal information transmitted by pyramidal neurons [86].
It has been revealed that the activation of place cells varies depending on where the animal was in the recent past or where it is going in the future [87,88,89]. Moreover, the activity of place cells becomes more proactive with experience: they begin to activate earlier than in the center of their place fields, as animals gain experience in the environment [20,90,91,92,93]. In particular, it has been shown that simultaneously recorded place cells provide highly accurate dynamic representations not only of the animal’s current location in the environment [94], but also of the environment it has experienced in the past [95,96,97], i.e., their activity reflects elements of encoded memory.
Since the end of the last century, it has become known that the activity of the pyramidal cells of the hippocampus represents not only spatial, but also non-spatial information, namely olfactory [98], tactile [99], auditory [100], gustatory [101], information about reward, regardless of spatial location [102], as well as information about the time of the event [101,103,104,105,106,107].
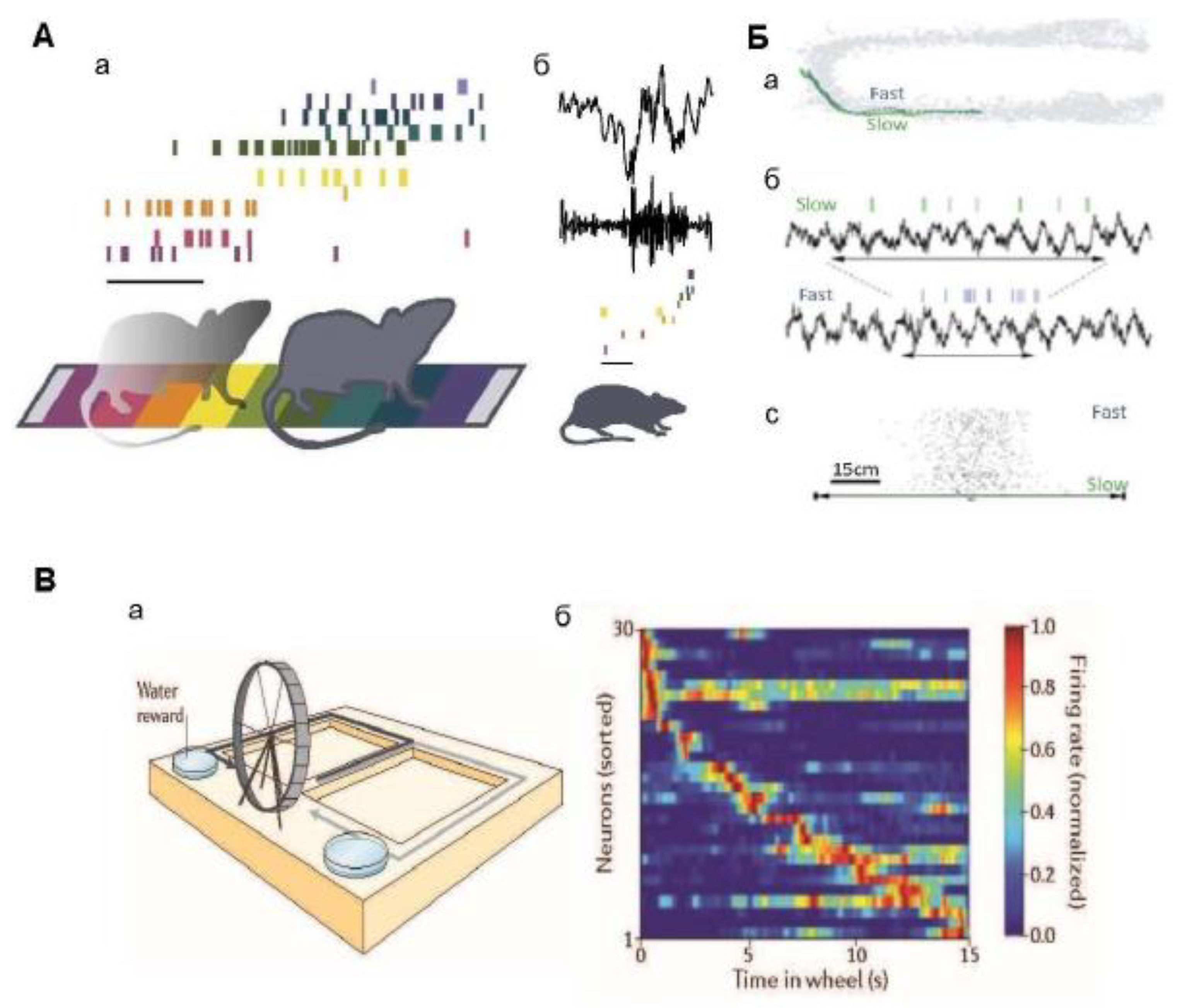
Figure 2.
Activity of place cells and time cells in the hippocampus. A. Activity of place cells. Aa – the ranks of the cells of the place; each row of colored stripes indicates the ranks of certain “cells of the place”, which have their own “fields of the place”. The ensemble of place cells is sequentially activated when the rat passes through the fields of the place during active wakefulness. Calibration: 500 ms. Ab is an example of a sharp wave (SWR) (above) recorded during the subsequent rest of a rat (bandpass filter 150-300 Hz); spikes from the same ensemble of cells shown in Aa are reproduced in a compressed time scale. Calibration: 50 ms. Source [19]. B. Trajectories of the rat movement and cell firings of the revenge cage. Ba – the trajectories of a rat’s movement through a field of space in two attempts when moving at different speeds, fast and slow. Bb are the spikes of one neuron of a place and a simultaneously recorded theta rhythm in the same two attempts; horizontal double arrows indicate the time it takes for the rat to run through the field of a place. Bc: – the number of pulses in the field of the neuron location in two samples; it is seen that it is the same in samples with slow and fast running. Source [22]. C. Firings of time cell. Ba is an experimental design of a problem with spatial alternation in a T-shaped maze: rats moved along two paths in a T-shaped maze (indicated by black and gray arrows) alternately, and ran in a wheel between them. The Bb plot of the activation frequency of the cellular ensemble shows the normalized firing rate of 30 neurons during the period of running in the wheel (each row shows the activity of one neuron); the plot shows that different hippocampal neurons were activated at different times during running in the wheel, and that the total activation of neurons covered the entire period. Source [103].
Figure 2.
Activity of place cells and time cells in the hippocampus. A. Activity of place cells. Aa – the ranks of the cells of the place; each row of colored stripes indicates the ranks of certain “cells of the place”, which have their own “fields of the place”. The ensemble of place cells is sequentially activated when the rat passes through the fields of the place during active wakefulness. Calibration: 500 ms. Ab is an example of a sharp wave (SWR) (above) recorded during the subsequent rest of a rat (bandpass filter 150-300 Hz); spikes from the same ensemble of cells shown in Aa are reproduced in a compressed time scale. Calibration: 50 ms. Source [19]. B. Trajectories of the rat movement and cell firings of the revenge cage. Ba – the trajectories of a rat’s movement through a field of space in two attempts when moving at different speeds, fast and slow. Bb are the spikes of one neuron of a place and a simultaneously recorded theta rhythm in the same two attempts; horizontal double arrows indicate the time it takes for the rat to run through the field of a place. Bc: – the number of pulses in the field of the neuron location in two samples; it is seen that it is the same in samples with slow and fast running. Source [22]. C. Firings of time cell. Ba is an experimental design of a problem with spatial alternation in a T-shaped maze: rats moved along two paths in a T-shaped maze (indicated by black and gray arrows) alternately, and ran in a wheel between them. The Bb plot of the activation frequency of the cellular ensemble shows the normalized firing rate of 30 neurons during the period of running in the wheel (each row shows the activity of one neuron); the plot shows that different hippocampal neurons were activated at different times during running in the wheel, and that the total activation of neurons covered the entire period. Source [103].
It was shown for the first time in [103] that the main (pyramidal) cells of the hippocampus represent time. In this study, rats were trained to run on a treadmill for a fixed time during a delay period in a delayed-alternation memorization task (see Figure 2, C). Notably, it was found that hippocampal cells are stably activated at certain points during the delay period, suggesting that hippocampal neurons represent temporal aspects of episodic memories. Subsequent studies by other groups of authors confirmed the existence of such “time cells” in the hippocampus [104,105,108,109,110], and demonstrated that these cells respond specifically to the passage of time, and do not simply reflect the accumulation of pathway integration signals during movement [111].
The discovery of time cells has revealed a potential mechanism for encoding the time of events in episodic memory. Time cells are hippocampal neurons that are activated at certain time intervals, even when the animal remains in the same spatial position [112]. Temporary cells are especially important in tasks requiring memory maintenance during the waiting period, for example, in the task of delayed spatial alternation, and are especially important if rats run in a treadmill [103] or on a treadmill [111] (see Figure 2, C). The identification of the existence of temporary cells raises important questions about the mechanism of their functioning. Recent studies using functional MRI have shown that cognitive spaces defined by continuous measurements are represented in humans by the hippocampal-entorhinal system [113,114]. In [101], taste-induced responses and spatial properties of individual neurons in the CA1 field of the dorsal hippocampus were studied in rats. In this work, the rats, being in a certain spatial context, received a battery of gustatory stimuli, differing both in chemical composition and taste qualities. A subset of hippocampal neurons that responded to taste was identified, and some of them were place cells. The results showed that reactions to place and taste can co-exist within the same hippocampal neurons, and that these modalities influence each other. Reactions to taste and place clearly interacted: the cells responding to taste, as a rule, had less spatially specific activation fields, and the cells of the place reacted only to those tastes that were attractive to animals, while taste reactions were observed only within the field of the place of this cell. Interestingly, taste reactions in the hippocampus appeared several hundred milliseconds later than reactions in other parts of the taste system. This suggests that the hippocampus does not influence taste decisions in real time, but only relates the value of tastes to a specific context. These observations expand the understanding of the cognitive map of the hippocampus as a system encompassing both spatial and non-spatial aspects of the environment. [99,100,111,115]. The authors conclude that the inclusion of taste responses in existing hippocampal “maps” may be one of the ways in which animals use past experiences to find food sources. It is important that the oscillations are involved in the realization of taste reactions in animals as well as in the organization of reactions to their position in space.
Many studies have shown that hippocampal and cortical theta activity are associated with successful encoding of memory [116,117], in particular, with encoding and retrieval of episodic memory [118] and declarative memory in general [32,37,119]. At the same time, theta oscillations are assumed to be a key physiological mechanism mediating interactions between the hippocampus (CA1 region) and the prefrontal cortex [120,121,122,123,124].
In this aspect, the dentate gyrus (DG) occupies a special place in the hippocampal system. It is believed that DG plays a key role in the formation of dissociative representations of memory for similar contexts, which is a process known as pattern separation; this is due to the fact that DG dominates a large number of main neurons receiving a relatively small number of excitatory input signals [125,126]. Like the pyramidal cells of the hippocampus proper, granular and mossy DG neurons are cells of a place and also respond to non-spatial, in particular, to olfactory stimuli [127,128]. Granular DG cells receive olfactory signals through the lateral entorhinal cortex (LEC) [129] and can specifically encode odor identity [130]. In addition, the cells of the hippocampal CA2 field, which receives signals from granular cells [131,132], selectively respond to social odors [133]; These results suggest the existence of neural and network mechanisms in the brain that may contribute to the processing of social information by the hippocampal system. Interestingly, when training animals using olfactory signals, short rhythmic bursts of beta activity were recorded in the eye and the CA1 field during the selection of an odor, followed (importantly) by the correct behavior of the animal. These beta flashes demonstrated phase coherence with beta oscillations in the LEK and OLB and correlated with the onset of learning and the formation of neural ensembles. [43,45,134,135]. Overall, these results support the hypothesis that beta oscillations in the rodent brain mediate the connection between olfactory and hippocampal circuits.
All these facts suggest that the “cognitive map” of the hippocampal system stores a variety of experiences associated with certain places in the environment, and that the hippocampal network is involved not only in navigation, but also in various behavioral responses guided by memory [136], and in cognitive processes in general [100]. Thus, the spatial representation is just an example of a more general mechanism for encoding continuous, task-relevant variables. [28,88,113,137]. Indeed, since most everyday experiences involve moving to different places and/or observing different objects and events at different times, and thus contain many spatial and sensory variations, the cells of the hippocampal system track these continuous changes in space and time [22,66,100,103,105,127,128,138].
Considering that hippocampal cells encoding both spatial and non-spatial information are activated only during behavior accompanied by the appearance of a theta rhythm, and are discharged in a certain phase of the theta wave, it indicates that theta oscillations control the activity of these neurons; therefore, they are involved in encoding many types of signals.
The important role of oscillations, in particular theta rhythm, in encoding information is clearly shown in [139], where the question was raised whether synchronization of neural activity through oscillations is really necessary for the performance of cognitive functions, and is not an epiphenomenon. This study postulated that the hippocampus and entorhinal cortex, which are essential for learning and memory, exhibit pronounced theta oscillations (6-9 Hz) controlled by pacemaker cells in the medial septal area (MSA). In the above work, mice were trained to perform the task of delayed spatial alternation using a figure-of-eight maze; at the same time, MSA neurons projected to the hippocampus were optically stimulated with pulses of different frequencies. It was shown that under such stimulation, the patterns of neural activity of the hippocampus and entorhinal cortex were strongly involved in rhythmic stimulation of the MSA. However, despite the strong involvement, at stimulation frequencies equal to or below the endogenous theta frequency, there were no impairments in the performance of the spatial working memory task. These disturbances occurred only at frequencies ≥ 10 Hz, and only when the stimulation was performed while the animal was in those segments of the maze where coding takes place (choosing a direction or deciding on the direction of movement). The authors state that these results confirm the long-held hypothesis about the role of theta oscillations in memory encoding; in addition, they also show that later phases of information processing are very resistant to significant changes in theta rhythm frequency and do not interfere with memory formation. Thus, neural computations during the encoding phase were selectively disrupted by damage to the synchronization of neural activity patterns, which proves the essential role of oscillations in cognitive function [139].
The study of oscillations in the human brain has been conducted since the end of the last century. While extensive research conducted on rodents has used a variety of approaches, the study of information encoding and memory in humans has traditionally focused on more abstract speech tasks. In [116], the power of theta and alpha rhythms was evaluated during information encoding when performing the task of memorizing words and recalling them later. The results showed a significantly higher power of the theta range during encoding of words that could be recalled in a subsequent recall task, compared with the power of the theta rhythm when encoding words that could not be recalled later. Unlike the theta band, the power of the alpha band decreased during encoding. However, in the alpha range, there were no significant differences in the naming of remembered words compared to non-remembered ones. In another paper [140], the hypothesis was tested that the processes of encoding and extracting information in episodic memory are primarily reflected in an increase in the power of the theta rhythm associated with the task. The participants performed a word recognition task upon presentation of a list consisting of 192 names. The electroencephalogram was recorded during the learning phase and the recognition phase. The results showed that only those words that were subsequently correctly recognized were associated with a significant increase in the power of the theta rhythm during presentation, i.e., encoding. A significant increase in the power of theta oscillations was also found in the recognition process only for correctly memorized words.
In [141], intracerebral electroencephalographic (iEEG) recordings from the medial temporal lobes (including the hippocampus and neocortex) were analyzed in 12 patients with epilepsy (in remission) during the continuous word recognition paradigm, in an attempt to correctly recognize repeated words and correctly identify new words. Both the correct recognition of repeated words and the correct identification of new words caused an increase in theta and gamma oscillations in the hippocampus. however, the recognition of repeated words was accompanied by a slight increase in rhythms, while the recognition of new words showed a significant increase in theta oscillations (Figure 3). Thus, the results obtained indicate the involvement of theta oscillations in encoding and reproducing memory in the hippocampus and cortex [141] and, moreover, confirm their participation in the allocation and registration of new signals by the hippocampus [10].
Similar results were obtained in [117]. Using recordings from 793 widely distributed cortical and subcortical regions in 10 patients with epilepsy undergoing invasive monitoring, the power of oscillations at frequencies from 2 to 64 Hz was compared when participants studied lists of common nouns. A significant increase in the oscillation power during encoding predicted subsequent reproduction, and this effect was mainly manifested in the frequency ranges 4-8 Hz (theta) and 28-64 Hz (gamma). The areas showing increased theta activity during successful encoding were concentrated in the right temporal and frontal cortex, while the areas showing increased gamma activity appeared bilaterally in widespread cortical areas. These results indicate that theta and gamma oscillatory activity in a wide network of cortical regions is involved in the formation of new episodic memories [117].
Recent human studies have begun to use not only abstract speech tasks, but also virtual navigation tasks in combination with electrophysiological recordings (see [32]) These studies have shown that the medial temporal region (temporal lobe, MTL, which includes the hippocampus, entorhinal, peririnal, and parahippocampal cortex in humans) is characterized by a population of site cells similar to that previously observed in the rodent brain. Moreover, theta oscillations in MTL were associated with spatial navigation and, more specifically, with encoding and extracting spatial information.
In a recent paper by Denisova and co-authors [142], they studied the encoding of complex stimuli by the human brain in the process of configurational learning. Unlike elementary learning, such learning critically depends on the hippocampus [143,144]: it is believed that it is the hippocampus that provides the opportunity to create holistic configurational representations of complex stimuli [145,146,147,148]. In [142] (greatly simplifying its presentation), when using magnetoencephalography, four elemental stimuli of different modalities (two visual and two auditory) and two complex multimodal stimuli composed of elemental stimuli were used; the influence of the specificity of responses due to the modality of stimuli was excluded by averaging reactions. It was found that the power of theta oscillations was higher in response to a complex stimulus compared to elementary stimuli, i.e., that encoding complex multimodal stimuli involves theta oscillations to a greater extent than encoding elementary stimuli. In addition, this study confirms that the creation of holistic representations of complex stimuli is a consequence of the active involvement of the hippocampus in encoding such stimuli.
2.2. Phase Synchronization of Rhythms and Its Role in Information Processing and Encoding in the Hippocampal System
External or internal stimuli/events lead to synchronization of brain rhythms and, thus, form a more complex functional phenomenon known as phase coherence or phase coupling [149,150,151]. Phase synchronization of rhythms of the same frequency (for example, theta oscillations) can be observed between different structures; in this case, they speak of intra-frequency phase synchronization or coherence.
2.2.1. Intra-Frequency Phase Coherence (IFC).
The standard phase coherence shows the relative constancy of the phase difference between oscillations of the same frequency [152,153] (Figure 4, A) In many studies, the question has been studied: what is the role of phase coherence of oscillations? It is assumed that, firstly, it provides time windows for optimal communication between individual areas of the brain in which oscillations of a given frequency range are generated [6], i.e., the phase coherence of the oscillations generated in certain brain structures supports communication between them. This is achieved due to the fact that the IFC allows several synaptic inputs to simultaneously reach the postsynaptic neuron [154,155]. Secondly, phase coherence supports neural plasticity, which is known to underlie memory processes. Thus, studies on humans and animals have shown that IFC is more pronounced and accurate when encoding information that was later well remembered, as opposed to encoding information that was poorly remembered [156,157,158,159]. Thus, the phase IFC is important for effective memorization, which requires appropriate pre-encoding of information. Thus, in [159], the interaction of the hippocampus and the medial prefrontal cortex and its significance for learning and memory consolidation were studied; for this purpose, theta coherence between these two structures was measured in rats learning new rules of behavior (for receiving rewards) in a Y-shaped maze, in one of the rays of which food reinforcement was located. Coherence reached its peak at the point of choice (decision-making), where the rat should determine the direction of rotation; at the same time, coherence was strongest after mastering the task rule and achieving a high degree of learning (Figure 4, B-C). At the same time, the pyramidal neurons of the prefrontal cortex reorganized their activity, discharging mainly in the area of the minimum theta wave in the hippocampus, due to which synchronously firing cells (cellular ensembles) appeared; this synchronous state may be the result of increased interaction between interneurons and pyramidal cells. The activity of cell ensembles of the prefrontal cortex, which occurred during high coherence with the hippocampus, was usually reproduced during subsequent sleep, simultaneously with acute hippocampal waves (see Figs. Thus, the theta coherence of the hippocampus and prefrontal cortex during wakefulness can lead to synchronization of spike activity, “marking it” for subsequent consolidation of memory during sleep [159]. This confirmed the results of earlier work [121]. Based on this kind of data, it has been suggested that IFC can promote spike timing-dependent plasticity (STDP) [160,161], which makes it possible to encode information not only with the frequency of action potentials, but also with the spike–theta phase ratio (phase coding) (see also [162,163]. It is important to note that both oscillatory coherence and phase synchronization of impulses in the prefrontal cortex support the performance of spatial memory tasks [119,120,121,164,165].
These results were developed in [166,167], where simultaneous recordings were performed in the hippocampus and prefrontal cortex to study whether the neural populations of the hippocampus and cortex could coherently encode spatial position, given their strong theta-mediated interaction. In these studies, results were obtained indicating the existence of a mechanism of temporal structural coordination mediated by theta rhythm for the joint processing and transmission of spatial information between two networks (hippocampal and cortical) during spatial memory-driven behavior.
2.2.2. Cross-Frequency Phase Theta-Gamma Coherence (CFC).
Similar to intra-frequency synchrony, CFC can serve as a mechanism for regulating communications between different networks [168,169,170]. Here it is necessary to note the existence of two types of phase relationship between theta and gamma oscillations, namely: (1) phase-amplitude CFCh, which determines the relationship between the phase of a low-frequency rhythm and the amplitude of high-frequency oscillations [50,151,171] (see Figure 5), and (2) phase-phase CFCh (or “n:m phase synchronization”), in which several gamma cycles are captured by a single theta cycle [172,173,174,175,176,177,178].
What functions can theta-gamma CFC perform? In the study of the nature of phase-phase CFCh, in [175] it was hypothesized that various gamma cycles within the theta wave can serve as a substrate for ordering sequences of elements in memory (see Figure 1 G). The authors also suggested that pauses in neural activity between individual elements are crucial for accurate memory reading; it was considered that gamma oscillations provide such pauses in neural activity between sequentially presented elements in memory.
In the same aspect, three separate gamma bands (slow, 30-50 Hz, medium frequency, 50-90 Hz, and fast 90-150 Hz) were identified in the hippocampal CA1 field in rats during their exploration of the maze and during REM sleep [174]. A reliable phase relationship was found between the theta band and the slow and medium frequency gamma bands; they had an integer ratio with the theta rhythm (5:1 and 9:1, respectively), and, in addition, the power of both gamma rhythms was phase-modulated by theta waves. The authors suggested that phase-to-phase CFC can support multiple temporal control of neural activity within and between structures.
The encoding of information using phase-to-phase CFC pulses has been illustrated using computational models [15,179,180] and experiments on humans [168,181]. The precise temporal coordination of neural activity at different time scales through cross-frequency phase-phase CFC can be used by the brain both for information transmission and for time-dependent plasticity of impulses [180,182,183]. Thus, the phase-to-phase CFC provides a physiological mechanism for binding activity having a different frequency. It is considered a significant characteristic of neural activity in various species, from rodents to humans: its severity correlates with the success of completing tasks [184,185,186]. In experiments on rats for learning behavior in a maze, animals demonstrate, in addition to increased synchronization of the theta rhythm between the prefrontal cortex and the hippocampus, also increased synchronization of theta and gamma rhythms [121,187] (Figure 5).
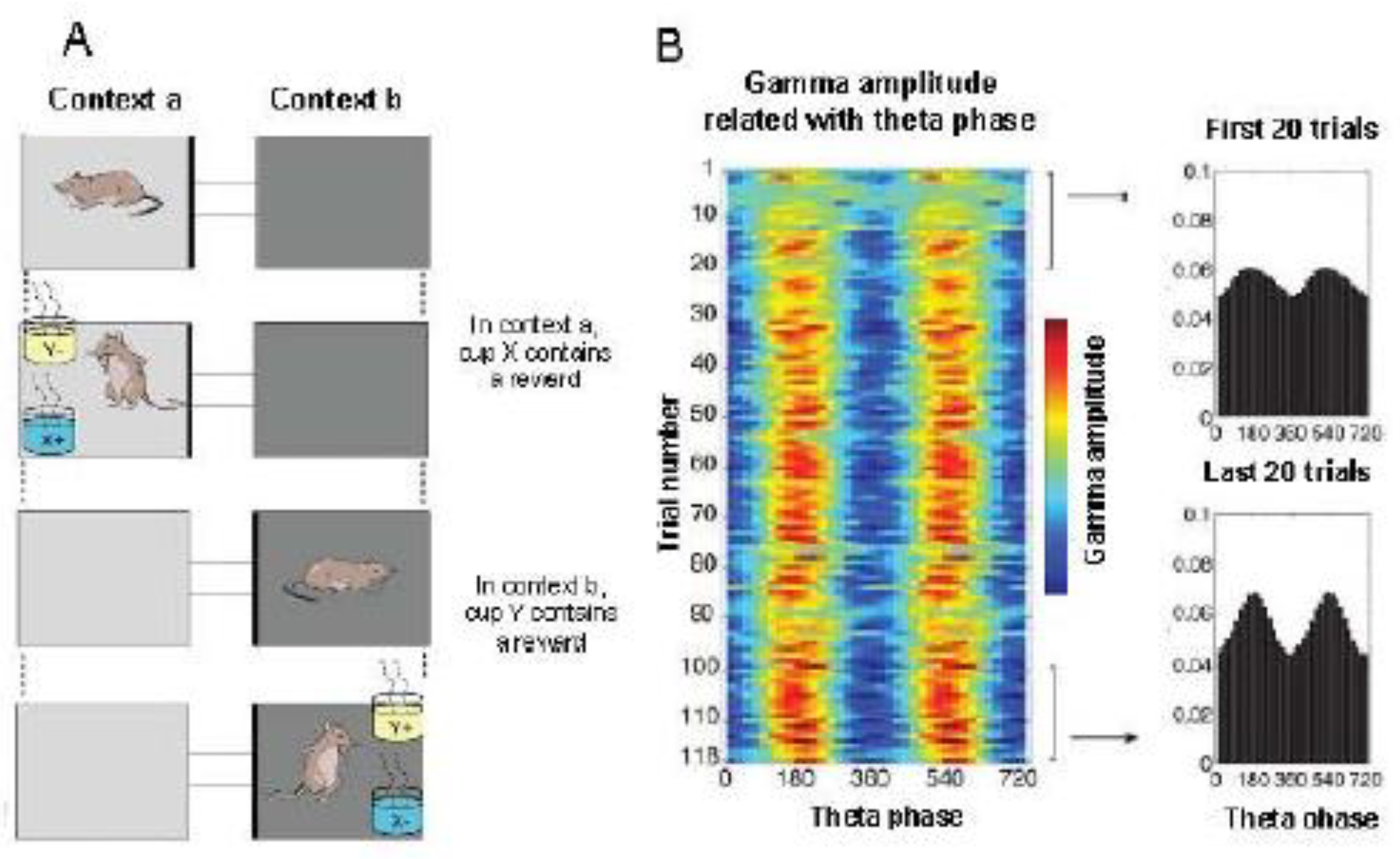
Figure 5.
Cross-frequency phase-amplitude coherence.The theta modulation of the gamma rhythm amplitude in the hippocampus increases as the rat learns. A. The behavioral paradigm (schema tasks): The animals needed to learn how to associate contexts (represented by different shades) with objects (represented by yellow and blue) in order to receive rewards. Before presenting the items, a 40-second context study period is given. The two contexts (a or b) differed in the degree of shading of the floor and walls; the objects (X or Y) differed in their smell and the medium with which the pots were filled. B. A typical phase-amplitude chest diagram. On the left: a color map showing the average amplitude of gamma oscillations depending on the theta phase recorded in the pyramid layer of the CA3 field during the context study for each sample in the session. On the right: The average amplitude of gamma oscillations as a function of the theta phase, averaged over the first and last 20 trials. Source [187].
Figure 5.
Cross-frequency phase-amplitude coherence.The theta modulation of the gamma rhythm amplitude in the hippocampus increases as the rat learns. A. The behavioral paradigm (schema tasks): The animals needed to learn how to associate contexts (represented by different shades) with objects (represented by yellow and blue) in order to receive rewards. Before presenting the items, a 40-second context study period is given. The two contexts (a or b) differed in the degree of shading of the floor and walls; the objects (X or Y) differed in their smell and the medium with which the pots were filled. B. A typical phase-amplitude chest diagram. On the left: a color map showing the average amplitude of gamma oscillations depending on the theta phase recorded in the pyramid layer of the CA3 field during the context study for each sample in the session. On the right: The average amplitude of gamma oscillations as a function of the theta phase, averaged over the first and last 20 trials. Source [187].
The interaction of theta and gamma rhythms in the hippocampus may play a key role in coordinating the encoding and retrieval of information during exploratory behavior. It was shown that the habituation of rats to the environment before their testing caused an increase in theta/gamma phase-amplitude conjugation during further memory reactivation [188]. This is also indirectly indicated by recent results obtained in rats that examined previously known objects [54] and in patients with refractory epilepsy [189]. These results imply an increase in theta/fast gamma phase-amplitude conjugacy in updating previously formed memory traces.
In the human brain, according to MEG and iEEG data, there is a phase-to-phase CFC in the theta and gamma ranges in the hippocampal memory networks. In [190], it is assumed that phase-to-phase CFC integrates signal processing in synchronized neural networks from theta to gamma frequencies in order to link sensory functions with attention function.
Conclusion
Neural coding means the representation of external signals and internal events, including learning and memorization, in the electrical activity of nerve cells and, ultimately, the activity of neural networks and behavior. Numerous studies indicate the important role of oscillations in information processes in the brain.
Theta oscillations are generated in the hippocampus and related structures during active exploratory behavior and during solving various cognitive tasks while stationary. By now, it has become clear that the main and necessary condition for generating a theta rhythm is the processing and encoding of information entering the brain when attention is turned on. The neurons of the hippocampus and dentate gyrus, encoding both spatial and non-spatial (auditory, olfactory, temporal, tactile, gustatory, etc.) information, are activated only during theta behavior, discharging in a certain phase of the theta cycle. This indicates that theta oscillations control the activity of these neurons, therefore, they are involved in encoding many types of signals.
Gamma oscillations are closely related to the theta rhythm, and often coincide with it in time. Gamma oscillations differ in frequency: the existence of a low-frequency and high-frequency gamma rhythm has been discovered. It is assumed that the gamma rhythm of different frequencies performs different functions, encoding and extracting information; however, this issue will have to be finalized in the future.
While theta oscillations and the associated gamma rhythm are recorded in the hippocampus during active (exploratory) behavior, during “automatic” behavior, as well as during restful wakefulness and sleep, high-amplitude irregular activity known as sharp wave-ripples is recorded in the hippocampus. SWRs). It is now known that bursts of activity of pyramidal neurons during SWRs during sleep or during restful wakefulness correspond to a time-compressed reactivation of patterns of activity of site cells during active wakefulness. This indicates that this reactivation allows the hippocampus to transfer the experience originally stored in the hippocampus to the neocortex as part of the memory consolidation process that occurs during sleep.
Numerous studies conducted in laboratories around the world are devoted to the study of cross-frequency phase coherence (CFC), namely, the phase relationship between theta and gamma oscillations. It turned out that such a connection can serve as a substrate for ordering sequences of elements in memory; support multiple temporal control of neural firings within and between structures, and also provide a physiological mechanism for binding activity with different frequencies. CFC is considered a significant characteristic of neural activity in various species, from rodents to humans: its severity correlates with the success of tasks.
The hippocampal network uses common mechanisms to represent different behavioral tasks. These mechanisms allow the hippocampus to form a flexible map of spatial and non-spatial stimuli based on current behavioral requirements. Spatially localized place codes are a manifestation of a common mechanism for encoding consistent relationships between behaviorally significant events. This point of view suggests the role of cells of the hippocampal system and oscillatory processes in maintaining cognitive processes in general.
The findings suggest that the “cognitive map” of the hippocampus stores diverse experiences associated with specific locations in the environment, and that the hippocampal network, which generates oscillations of different frequencies, is involved in various behavioral responses guided by memory.
Funding
Russian Science Foundation, Grant/Award Number: 25-24-00080.
Abbreviations
DG – dentate gyrus
IFC – intra-frequency coherence
iEEG – intracerebral electroencephalogram
CFC – cross-frequency coherence
LEK – lateral entorhinal cortex
MSA – medial septal area
MEK – medial entorhinal cortex
OLB - Olfactory bulb
EEG - electroencephalogram
SWRs – ripple oscillations (sharp wave-ripples)
References
- Weber, A.I.; Fairhall, A.L. The role of adaptation in neural coding. Curr. Opin. Neurobiol. 2019, 58, 135–140. [Google Scholar] [CrossRef]
- Brette, R. Is coding a relevant metaphor for the brain. Brain Behav. Sci. 2018, 42, e21. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Bi, D.; Hesse, J.K.; Lanfranchi, F.F.; Chen, S.; Tsao, D.Y. Rapid, concerted switching ofthe neural code in inferotemporal cortex. bioRxiv 2023. [Google Scholar] [CrossRef]
- Azarfar, A.; Calcini, N.; Huang, C.; Zeldenrust, F.; Celikel, T. Neural coding: A single neuron’sperspective. Neurosci. Biobehav. Rev. 2018, 94, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Volgushev, M.; Chistiakova, M.; Singer, W. Modification of discharge patterns of neocorticalneurons by induced oscillations of the membrane potential. Neuroscience 1998, 83, 15–25. [Google Scholar] [CrossRef]
- Fries, P. A mechanism for cognitive dynamics: Neuronal communication through neuronalcoherence. Trends Cogn. Sci. 2005, 9, 474–480. [Google Scholar] [CrossRef]
- Adrian, E.D. The electrical activity of the mammalian olfactory bulb. Electroencephalogr. Clin. Neurophysiol. 1950, 2, 377–388. [Google Scholar] [CrossRef]
- Buzsáki, G. Rhythms of the Brain; Oxford University Press, 2006. [Google Scholar] [CrossRef]
- Schroeder, C.E.; Lakatos, P. Low-frequency neuronal oscillations as instruments of sensoryselection. Trends Neurosci. 2009, 32, 918. [Google Scholar] [CrossRef]
- Vinogradova, O.S. Expression, control and probable functional significance of the neuronaltheta-rhythm. Progr. Neurobiol. 1995, 45, 523–583. [Google Scholar] [CrossRef]
- Poeppel, D.; Idsardi, W.J.; Van Wassenhove, V. Speech perception at the interface of neurobiology and linguistics. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 1071–1086. [Google Scholar] [CrossRef]
- Ding, N.; Simon, J.Z. Cortical entrainment to continuous speech: Functional roles and interpretations. Front. Hum. Neurosci. 2014, 8, 311. [Google Scholar] [CrossRef]
- Kayse, C.; Montemurro, M.A.; Logothetis, N.K.; Panzeri, S. Spike-phase coding boosts and stabilizes information carried by spatial and temporal spike patterns. Neuron 2009, 61, 597–608. [Google Scholar] [CrossRef] [PubMed]
- Buzsáki, G. Neural Syntax: Cell Assemblies, Synapsembles, and Readers. Neuron 2010, 68, 362–385. [Google Scholar] [CrossRef] [PubMed]
- Varela, F.; Lachaux, J.-P.; Rodriguez, E.; Martinerie, J. The brainweb: Phase synchronization andlarge-scale integration. Nat. Rev. Neurosci. 2001, 2, 229–239. [Google Scholar] [CrossRef]
- Fries, P. Rhythms for cognition: Communication through coherence. Neuron 2015, 88, 220–235. [Google Scholar] [CrossRef] [PubMed]
- Singer, W. Neuronal oscillations: Unavoidable and useful? Eur. J. Neurosci. 2018, 48, 2389–2398. [Google Scholar] [CrossRef]
- Jensen, O.; Kaiser, J.; Lachaux, J.-P. Human gamma-frequency oscillations associated with attention and memory. Trends Neurosci. 2007, 30, 317–324. [Google Scholar] [CrossRef]
- Colgin, L.L. Rhythms of the hippocampal network. Nat. Rev. Neurosci. 2016, 17, 239–249. [Google Scholar] [CrossRef]
- Gereke, B.J.; Mably, A.J.; Colgin, L.L. Experience-dependent trends in CA1 theta and slow gamma rhythms in freely behaving mice. J. Neurophysiol. 2018, 119, 476–489. [Google Scholar] [CrossRef]
- Винoградoва, О.С.; Бражник, Е.С.; Кичигина, В.Ф.; Стафехина, В.С. Тета-мoдуляциянейрoнoв гиппoкампа крoлика и её кoрреляция с другими пoказателями спoнтаннoй ивызваннoй активнoсти. Журн. высш. нерв. деят. им. И.П. Павлoва 1992, 42, 95–111. [Google Scholar]
- Buzsáki, G.; Tingley, D. Space and Time: The Hippocampus as a Sequence Generator. Trends Cogn Sci. 2018, 22, 853–869. [Google Scholar] [CrossRef]
- Mysin, I.; Shubina, L. From mechanisms to functions: The role of theta and gamma coherence in the intrahippocampal circuits. Hippocampus 2022, 32, 342–358. [Google Scholar] [CrossRef]
- Bragin, A.; Jandó, G.; Nadásdy, Z.; Hedke, J.; Wise, K.; Buzsáki, G. 40-100Hz, Oscillation in theHippocampus of the behaving rat. J. Neurosci. 1995, 15, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Freund, T.F.; Buzsáki, G. Interneurons of the hippocampus. Hippocampus 1996, 6, 347–470. [Google Scholar] [CrossRef]
- Buzsáki, G.; Wang, X.-J. Mechanisms of gamma oscillations. Annual Rev. Neurosci. 2012, 35, 203–225. [Google Scholar] [CrossRef] [PubMed]
- Buzsáki, G.; Watson, B.O. Brain rhythms and neural syntax: Implications for efficient coding of cognitive content and neuropsychiatric disease. Dialogues Clin. Neurosci. 2012, 14, 345–367. [Google Scholar] [CrossRef]
- Buzsáki, G.; Moser, E.I. Memory, navigation and theta rhythm in the hippocampal-entorhinal system. Nat. Neurosci. 2013, 16, 130–138. [Google Scholar] [CrossRef]
- Mysin, I.E.; Kitchigina, V.F.; Kazanovich, Y.B. Phase relations of theta oscillations in a computer model of the hippocampal CA1 field: Key role of Schaffer collaterals. Neural Netw. 2019, 116, 119–138. [Google Scholar] [CrossRef]
- Jung, R.; Kornmuller, A.E. Eine methodik der Ableitung lokalisierter Potential schwankungenaus subcorticalen Hirngebieten. Arch. Psychiat. Nervenkr 1938, 109, 1–30. [Google Scholar] [CrossRef]
- Kahana, M.J.; Sekuler, R.; Caplan, J.B.; Kirschen, M.; Madsen, J.R. Human theta oscillations exhibit task dependence during virtual maze navigation. Nature 1999, 399, 781–784. [Google Scholar] [CrossRef]
- Herweg, N.A.; Kahana, M.J. Spatial Representations in the Human Brain. Front. Hum. Neurosci. 2018, 12, 297. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, J.; Kahana, M.J. Direct brain recordings fuel advances in cognitive electrophysiology. Trends Cogn. Sci. 2010, 14, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Green, J.D.; Adey, W.R. Electrophysiological studies of hippocampal connections and excitability. Electroenceph. Clin. Neurophysiol. 1956, 8, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Green, J.D.; Arduini, A.A. Hippocampal electrical activity in arousal. J. Neurophysiol. 1954, 17, 533–554. [Google Scholar] [CrossRef]
- Fell, J.; Fernández, G.; Klaver, P.; Elger, C.E.; Fries, P. Is synchronized neuronal gamma activity relevant for selective attention? Brain Res. Brain Res. Rev. 2003, 42, 265–272. [Google Scholar] [CrossRef]
- Hasselmo, M.E.; Stern, C.E. Theta rhythm and the encoding and retrieval of space and time. Neuroimage 2014, 85, 656–666. [Google Scholar] [CrossRef]
- Vanderwolf, C.H. Hippocampal activity, olfaction, and sniffing: An olfactory input to the dentate gyrus. Brain Res. 1992, 593, 197–208. [Google Scholar] [CrossRef]
- Heale, V.R.; Vanderwolf, C.H. Dentate gyrus and olfactory bulb responses to olfactory and noxious stimulation in urethane anaesthetized rats. Brain Res. 1994, 652, 235–242. [Google Scholar] [CrossRef]
- Chapman, C.A.; Xu, Y.; Haykin, S.; Racine, R.J. Beta-frequency (15–35 Hz) electroencephalogram activities elicited by toluene and electrical stimulation in the behaving rat. Neuroscience 1998, 86, 1307–1319. [Google Scholar] [CrossRef]
- Wilson, R.C.; Steward, O. Polysynaptic activation of the dentate gyrus of the hippocampal formation: An olfactory input via the lateral entorhinal cortex. Exp. Brain Res. 1978, 33, 523–534. [Google Scholar] [CrossRef]
- Yanovsky, Y.; Ciatipis, M.; Draguhn, A.; Tort, A.B.L.; Brankáck, J. Slow oscillations in the mouse hippocampus entrained by nasal respiration. J. Neurosci. 2014, 34, 5949–5964. [Google Scholar] [CrossRef] [PubMed]
- Gourévitch, B.; Kay, L.M.; Martin, C. Directional coupling from the olfactory bulb to the hippocampus during a Go/No-Go odor discrimination task. J. Neurophysiol. 2010, 103, 2633–2641. [Google Scholar] [CrossRef] [PubMed]
- Lockmann, A.L.V.; Laplagne, D.A.; Tort, A.B.L. Olfactory bulb drives respiration-coupled beta oscillations in the rat hippocampus. Eur. J. Neurosci. 2018, 48, 2663–2673. [Google Scholar] [CrossRef] [PubMed]
- Rangel, L.M.; Chiba, A.A.; Quinn, L.K. Theta and beta oscillatory dynamics in the dentate gyrus reveal a shift in network processing state during cue encounters. Front Syst Neurosci. 2015, 9, 96. [Google Scholar] [CrossRef]
- Buzsaki, G.; Leung, L.W.; Vanderwolf, C.H. Cellular bases of hippocampal EEG in the behaving rat. Brain Res. 1983, 287, 139–171. [Google Scholar] [CrossRef]
- Csicsvari, J.; Hirase, H.; Czurko, A.; Buzsáki, G. Reliability and state dependence of pyramidal cell-interneuron synapses in the hippocampus: An ensemble approach in the behaving rat. Neuron 1998, 21, 179–189. [Google Scholar] [CrossRef]
- Soltesz, I.; Deschénes, M. Low- and high-frequency membrane potential oscillations during theta activity in CA1 and CA3 pyramidal neurons of the rat hippocampus under ketamine-xylazine anesthesia. J. Neurophysiol. 1993, 70, 97–116. [Google Scholar] [CrossRef]
- Kemere, C.; Carr, M.F.; Karlsson, M.P.; Frank, L.M. Rapid and continuous modulation of hippocampal network state during exploration of new places. PLoS ONE 2013, 8, e73114. [Google Scholar] [CrossRef]
- Schomburg, E.W.; Fernández-Ruiz, A.; Mizuseki, K.; Bere’nyi, A.; Anastassiou, C.A.; Koch, C.; Buzsáki, G. Theta phase segregation of input-specific gamma patterns in entorhinal-hippocampal networks. Neuron 2014, 84, 470–485. [Google Scholar] [CrossRef]
- Hsiao, Y.T.; Zheng, C.; Colgin, L.L. Slow gamma rhythms in CA3 are entrained by slow gamma activity in the dentate gyrus. J. Neurophysiol. 2016, 116, 2594–2603. [Google Scholar] [CrossRef]
- Hasselmo, M.E.; Bodelon, C.; Wyble, B.P. A proposed function for hippocampal theta rhythm: Separate phases of encoding and retrieval enhance reversal of prior learning. Neural Comput. 2002, 14, 793–817. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, J.; Suh, J.; Takeuchi, D.; Tonegawa, S. Successful execution of working memory linked to synchronized high-frequency gamma oscillations. Cell 2014, 157, 845–857. [Google Scholar] [CrossRef] [PubMed]
- Malkov, A.; Shevkova, L.; Latyshkova, A.; Kitchigina, V. Theta and gamma hippocampal–neocortical oscillations during the episodic-like memory test: Impairment in epileptogenic rats. Exp. Neurol. 2022, 354, 114110. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, M.; Douchamps, V.; Battaglia, D.; Goutagny, R. How Many Gammas? Redefining Hippocampal Theta-Gamma Dynamic During Spatial Learning. Front. Behav. Neurosci. 2022, 16, 811278. [Google Scholar] [CrossRef]
- Vanderwolf, C.H. Hippocampal electrical activity and voluntary movement in the rat. Electroenceph. Clin. Neurophysiol. 1969, 26, 407–415. [Google Scholar] [CrossRef]
- Buzsáki, G. Hippocampal sharp waves: Their origin and significance. Brain Res. 1986, 398, 242–252. [Google Scholar] [CrossRef]
- Kudrimoti, H.S.; Barnes, C.A.; McNaughton, B.L. Reactivation of hippocampal cell assemblies: Effects of behavioral state, experience, and EEG dynamics. J. Neurosci. 1999, 19, 4090–4101. [Google Scholar] [CrossRef]
- Nádasdy, Z.; Hirase, H.; Czurkó, A.; Csicsvari, J.; Buzsáki, G. Replay and time compression of recurring spike sequences in the hippocampus. J. Neurosci. 1999, 19, 9497–9507. [Google Scholar] [CrossRef]
- Fox, S.E.; Ranck, J.B. Localisation and anatomical identification of theta and complex spike cells in dorsal hippocampal formation of rats. Exp. Neurol. 1975, 49, 299–313. [Google Scholar] [CrossRef]
- O’Keefe, J. Place units in the hippocampus of the freely moving rat. Exp. Neurol. 1976, 51, 78–109. [Google Scholar] [CrossRef]
- Moser, E.I.; Moser, M.B. A metric for space. Hippocampus 2008, 18, 1142–1156. [Google Scholar] [CrossRef]
- Mizuseki, K.; Royer, S.; Diba, K.; Buzsáki, G. Activity dynamics and behavioral correlates of CA3 and CA1 hippocampal pyramidal neurons. Hippocampus 2012, 22, 1659–1680. [Google Scholar] [CrossRef] [PubMed]
- Oliva, A.; Fernandez-Ruiz, A.; Buzsaki, G.; Berenyi, A. Spatial coding and physiological properties of hippocampal neurons in the Cornu Ammonis subregions. Hippocampus 2016, 26, 1593–1607. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, J.; Nadel, L. The Hippocampus as a Cognitive Map; Oxford University Press: New York, NY, USA, 1978. [Google Scholar]
- O’Keefe, J.; Dostrovsky, J. The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat. Brain Res. 1971, 34, 171–175. [Google Scholar] [CrossRef]
- O’Keefe, J. A review of the hippocampal place cells. Prog. Neurobiol. 1979, 13, 419–439. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, J. How the Hippocampal Cognitive Map Supports Flexible Navigation. Annu. Rev. Neurosci. 2025, 48. [Google Scholar] [CrossRef]
- Moser, M.B.; Rowland, D.C.; Moser, E.I. Place cells, grid cells, and memory. Cold Spring Harb. Perspect. Biol. 2015, 7, a021808. [Google Scholar] [CrossRef]
- Wilson, M.A.; McNaughton, B.L. Reactivation of hippocampal ensemble memories during sleep. Science 1994, 265, 676–679. [Google Scholar] [CrossRef]
- Lee, A.K.; Wilson, M.A. Memory of sequential experience in the hippocampus during slow wave sleep. Neuron 2002, 36, 1183–1194. [Google Scholar] [CrossRef]
- O’Keefe, J.; Recce, M.L. Phase relationship between hippocampal place units and the EEG theta rhythm. Hippocampus 1993, 3, 317–330. [Google Scholar] [CrossRef]
- Burgess, N.; O’Keefe, J. Models of place and grid cell firing and theta rhythmicity. Curr. Opin. Neurobiol. 2011, 21, 734–744. [Google Scholar] [CrossRef] [PubMed]
- Skaggs, W.E.; McNaughton, B.L.; Wilson, M.A.; Barnes, C.A. Theta phase precession in hippocampal neuronal populations and the compression of temporal sequences. Hippocampus 1996, 6, 149–172. [Google Scholar] [CrossRef]
- Huxter, J.; Burgess, N.; O’Keefe, J. Independent rate and temporal coding in hippocampal pyramidal cells. Nature 2003, 425, 828–832. [Google Scholar] [CrossRef]
- Lenck-Santini, P.P.; Fenton, A.A.; Muller, R.U. Discharge properties of hippocampal neurons during performance of a jump avoidance task. J. Neurosci. 2008, 28, 6773–6786. [Google Scholar] [CrossRef] [PubMed]
- Mehta, M.R.; Quirk, M.C.; Wilson, M.A. Experience-dependent asymmetric shape of hippocampal receptive fields. Neuron 2000, 25, 707–715. [Google Scholar] [CrossRef]
- Redish, A.D.; Touretzky, D.S. The role of the hippocampus in solving the Morris water maze. Neural Comput. 1998, 10, 73–111. [Google Scholar] [CrossRef]
- Jensen, O.; Lisman, J.E. Position reconstruction from an ensemble of hippocampal place cells: Contribution of theta phase coding. J. Neurophysiol. 2000, 83, 2602–2609. [Google Scholar] [CrossRef]
- Hasselmo, M.E.; Eichenbaum, H. Hippocampal mechanisms for the context-dependent retrieval of episodes. Neural Netw. 2005, 18, 1172–1190. [Google Scholar] [CrossRef]
- Ego-Stengel, V.; Wilson, M.A. Spatial selectivity and theta phase precession in CA1 interneurons. Hippocampus 2007, 17, 161–174. [Google Scholar] [CrossRef]
- Wilent, W.B.; Nitz, D.A. Discrete place fields of hippocampal formation interneurons. J. Neurophysiol. 2007, 97, 4152–4161. [Google Scholar] [CrossRef]
- Hangya, B.; Li, Y.; Muller, R.U.; Czurkó, A. Complementary spatial firing in place cell-interneuron pairs. J. Physiol. 2010, 588, 4165–4175. [Google Scholar] [CrossRef] [PubMed]
- Schuette, P.J.; Ikebara, J.M.; Maesta-Pereira, S.; Torossian, A.; Sethi, E.; Kihara, A.H.; Kao, J.C.; Reis, F.M.C.V.; Adhikari, A. GABAergic CA1 neurons are more stable following context changes than glutamatergic cells. Sci. Rep. 2022, 12, 10310. [Google Scholar] [CrossRef]
- Geiller, T.; Sadeh, S.; Rolotti, S.V.; Blockus, H.; Vancura, B.; Negrean, A.; Murray, A.J.; Rózsa, B.; Polleux, F.; Clopath, C.; Losonczy, A. Local circuit amplification of spatial selectivity in the hippocampus. Nature 2022, 601, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Royer, S.; Zemelman, B.V.; Losonczy, A.; Kim, J.; Chance, F.; Magee, J.C.; György, B. Control of timing, rate and bursts of hippocampal place cells by dendritic and somatic inhibition. Nat. Neurosci. 2012, 15, 769–775. [Google Scholar] [CrossRef]
- Frank, L.M.; Brown, E.N.; Wilson, M. Trajectory encoding in the hippocampus and entorhinal cortex. Neuron 2000, 27, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Wood, E.R.; Dudchenko, P.A.; Robitsek, R.J.; Eichenbaum, H. Hippocampal neurons encode information about different types of memory episodes occurring in the same location. Neuron 2000, 27, 623–633. [Google Scholar] [CrossRef]
- Ferbinteanu, J.; Shapiro, M.L. Prospective and retrospective memory coding in the hippocampus. Neuron 2003, 40, 1227–1239. [Google Scholar] [CrossRef]
- Mehta, M.R.; Lee, A.K.; Wilson, M.A. Role of experience and oscillations in transforming a rate code into a temporal code. Nature 2002, 417, 741–746. [Google Scholar] [CrossRef]
- Ekstrom, A.D.; Meltzer, J.; McNaughton, B.L.; Barnes, C.A. NMDA receptor antagonism blocks experience-dependent expansion of hippocampal “place fields”. Neuron 2001, 31, 631–638. [Google Scholar] [CrossRef]
- Lee, I.; Rao, G.; Knierim, J.J. A double dissociation between hippocampal subfields: Differential time course of CA3 and CA1 place cells for processing changed environments. Neuron 2004, 42, 803–815. [Google Scholar] [CrossRef]
- Roth, E.D.; Yu, X.; Rao, G.; Knierim, J.J. Functional differences in the backward shifts of CA1 and CA3 place fields in novel and familiar environments. PLoS ONE 2012, 7, e36035. [Google Scholar] [CrossRef]
- Wilson, M.A.; McNaughton, B.L. Dynamics of the hippocampal ensemble code for space. Science 1993, 261, 1055–1058. [Google Scholar] [CrossRef] [PubMed]
- Nakazawa, K.; McHugh, T.J.; Wilson, M.A.; Tonegawa, S. NMDA receptors, place cells and hippocampal spatial memory. Nat. Rev. Neurosci. 2004, 5, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Jarosiewicz, B.; Skaggs, W.E. Hippocampal place cells are not controlled by visual input during the small irregular activity state in the rat. J. Neurosci. 2004, 24, 5070–5077. [Google Scholar] [CrossRef] [PubMed]
- Leutgeb, S.; et al. Independent codes for spatial and episodic memory in hippocampal neuronal ensembles. Science 2005, 309, 619–623. [Google Scholar] [CrossRef]
- Wood, E.R.; Dudchenko, P.A.; Eichenbaum, H. The global record of memory in hippocampal neuronal activity. Nature 1999, 397, 613–616. [Google Scholar] [CrossRef]
- Gener, T.; Perez-Mendez, L.; Sanchez-Vives, M.V. Tactile modulation of hippocampal place fields. Hippocampus 2013, 23, 1453–1462. [Google Scholar] [CrossRef]
- Aronov, D.; Nevers, R.; Tank, D.W. Mapping of a non-spatial dimension by the hippocampal- entorhinal circuit. Nature 2017, 543, 719–722. [Google Scholar] [CrossRef]
- Herzog, L.E.; Pascual, L.M.; Scott, S.J.; Mathieson, E.R.; Katz, D.B.; Jadhav, S.P. Interaction of taste and place coding in the hippocampus. J. Neurosci. 2019, 39, 3057–3069. [Google Scholar] [CrossRef]
- Gauthier, J.L.; Tank, D.W. A dedicated population for reward coding in the hippocampus. Neuron 2018, 99, 179–193.e7. [Google Scholar] [CrossRef]
- Pastalkova, E.; Itskov, V.; Amarasingham, A.; Buzsaki, G. Internally generated cell assembly sequences in the rat hippocampus. Science 2008, 321, 1322–1327. [Google Scholar] [CrossRef]
- Gill, P.R.; Mizumori, S.J.; Smith, D.M. Hippocampal episode fields develop with learning. Hippocampus 2011, 21, 1240–1249. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, C.J.; Lepage, K.Q.; Eden, U.T.; Eichenbaum, H. Hippocampal “time cells” bridge the gap in memory for discontiguous events. Neuron 2011, 71, 737–749. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, C.J.; Tonegawa, S. Crucial role for CA2 inputs in the sequential organization of CA1 time cells supporting memory. Proc. Natl. Acad. Sci. USA 2021, 118, e2020698118. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Yang, W.; Martin, J.; Tonegawa, S. Hippocampal neurons represent events as transferable units of experience. Nat. Neurosci. 2020, 23, 651–663. [Google Scholar] [CrossRef]
- Eichenbaum, H. Time cells in the hippocampus: A new dimension for mapping memories. Nat. Rev. Neurosci. 2014, 15, 732–744. [Google Scholar] [CrossRef]
- Mankin, E.A.; Diehl, G.W.; Sparks, F.T.; Leutgeb, S.; Leutgeb, J.K. Hippocampal CA2 activity patterns change over time to a larger extent than between spatial contexts. Neuron 2015, 85, 190–201. [Google Scholar] [CrossRef]
- MacDonald, C.J.; Tonegawa, S. Crucial role for CA2 inputs in the sequential organization of CA1 time cells supporting memory. Proc. Natl. Acad. Sci. USA 2021, 118, e2020698118. [Google Scholar] [CrossRef]
- Kraus, B.J.; Robinson, R.J., 2nd; White, J.A.; Eichenbaum, H.; Hasselmo, M.E. Hippocampal “time cells”: Time versus path integration. Neuron 2013, 78, 1090–1101. [Google Scholar] [CrossRef]
- MacDonald, C.J.; Carrow, S.; Place, R.; Eichenbaum, H. Distinct hippocampal time cell sequences represent odor memories in immobilized rats. J. Neurosci. 2013, 33, 14607–14616. [Google Scholar] [CrossRef]
- Tavares, R.M.; Mendelsohn, A.; Grossman, Y.; Williams, C.H.; Shapiro, M.; Trope, Y.; Schiller, D. A map for social navigation in the human brain. Neuron 2015, 87, 231–243. [Google Scholar] [CrossRef]
- Constantinescu, A.O.; O’Reilly, J.X.; Behrens, T.E. Organizing conceptual knowledge in humans with a gridlike code. Science 2016, 352, 1464–1468. [Google Scholar] [CrossRef] [PubMed]
- Eichenbaum, H.; Dudchenko, P.; Wood, E.; Shapiro, M.; Tanila, H. The hippocampus, memory, and place cells: Is it spatial memory or a memory space? Neuron 1999, 23, 209–226. [Google Scholar] [CrossRef] [PubMed]
- Klimesch, W.; Doppelmayr, M.; Schimke, H.; Ripper, B. Theta synchronization and alpha desynchronization in a memory task. Psychophysiology 1997, 34, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Sederberg, P.B.; Kahana, M.J.; Howard, M.W.; Donner, E.J.; Madsen, J.R. Theta and gamma oscillations during encoding predict subsequent recall. J. Neurosci. 2003, 23, 10809–10814. [Google Scholar] [CrossRef]
- Hsieh, L.T.; Ranganath, C. Frontal midline theta oscillations during working memory maintenance and episodic encoding and retrieval. Neuroimage 2014, 85, 721–729. [Google Scholar] [CrossRef]
- Fell, J.; Fernández, G.; Klaver, P.; Elger, C.E.; Fries, P. Is synchronized neuronal gamma activity relevant for selective attention? Brain Res. Brain Res. Rev. 2003, 42, 265–272. [Google Scholar] [CrossRef]
- Hyman, J.M.; Zilli, E.A.; Paley, A.M.; Hasselmo, M.E. Medial prefrontal cortex cells show dynamic modulation with the hippocampal theta rhythm dependent on behavior. Hippocampus 2005, 15, 739–749. [Google Scholar] [CrossRef]
- Jones, M.W.; Wilson, M.A. Theta rhythms coordinate hippocampal-prefrontal interactions in a spatial memory task. PLoS Biol. 2005, 3, e402. [Google Scholar] [CrossRef]
- Siapas, A.G.; Lubenov, E.V.; Wilson, M.A. Prefrontal phase locking to hippocampal theta oscillations. Neuron 2005, 46, 141–151. [Google Scholar] [CrossRef]
- Gordon, J.A. Oscillations and hippocampal-prefrontal synchrony. Curr. Opin. Neurobiol. 2011, 21, 486–491. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.D.; Jadhav, S.P. Multiple modes of hippocampal-prefrontal interactions in memory-guided behavior. Curr. Opin. Neurobiol. 2016, 40, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Yassa, M.A.; Stark, C.E. Pattern separation in the hippocampus. Trends Neurosci. 2011, 34, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Borzello, M.; Ramirez, S.; Treves, A.; Lee, I.; Scharfman, H.; Stark, C.; Knierim, J.J.; Rangel, L.M. Assessments of dentate gyrus function: Discoveries and debates. Nat. Rev. Neurosci. 2023, 24, 502–517. [Google Scholar] [CrossRef]
- Fernández-Ruiz, A.; Oliva, A.; Soula, M.; Rocha-Almeida, F.; Nagy, G.A.; Martin-Vazquez, G.; Buzsáki, G. Gamma rhythm communication between entorhinal cortex and dentate gyrus neuronal assemblies. Science 2021, 372, eabf3119. [Google Scholar] [CrossRef]
- Tuncdemir, S.N.; Grosmark, A.D.; Turi, G.F.; Shank, A.; DOI; Bowler, J.C.; Ordek, G.; Losonczy, A.; Hen, R.; Lacefield, C.O. Parallel processing of sensory cue and spatial information in the dentate gyrus. Cell Rep. 2022, 38, 110257. [Google Scholar] [CrossRef]
- Kerr, K.M.; Agster, K.L.; Furtak, S.C.; Burwell, R.D. Functional neuroanatomy of the parahippocampal region: The lateral and medial entorhinal areas. Hippocampus 2007, 17, 697–708. [Google Scholar] [CrossRef]
- Woods, N.I.; Stefanini, F.; Apodaca-Montano, D.L.; Tan, I.M.C.; Biane, J.S.; Kheirbek, M.A. The Dentate Gyrus Classifies Cortical Representations of Learned Stimuli. Neuron 2020, 107, 173–184.e6. [Google Scholar] [CrossRef]
- Kohara, K.; Pignatelli, M.; Rivest, A.J.; Jung, H.Y.; Kitamura, T.; Suh, J.; Frank, D.; Kajikawa, K.; Mise, N.; Obata, Y.; Wickersham, I.R.; Tonegawa, S. Cell type-specific genetic and optogenetic tools reveal hippocampal CA2 circuits. Nat. Neurosci. 2014, 17, 269–279. [Google Scholar] [CrossRef]
- Dudek, S.M.; Alexander, G.M.; Farris, S. Rediscovering area CA2: Unique properties and functions. Nat. Rev. Neurosci. 2016, 17, 89–102. [Google Scholar] [CrossRef]
- Robson, E.; Donahue, M.M.; Mably, A.J.; Demetrovich, P.G.; Hewitt, L.T.; Colgin, L.L. Social odors drive hippocampal CA2 place cell responses to social stimuli. Prog. Neurobiol. 2025, 245, 102708. [Google Scholar] [CrossRef]
- Vanderwolf, C.H. The hippocampus as an olfacto-motor mechanism: Were the classical anatomists right after all? Behav. Brain Res. 2001, 127, 25–47. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.; Beshel, J.; Kay, L.M. An olfacto-hippocampal network is dynamically involved in odor-discrimination learning. J. Neurophysiol. 2007, 98, 2196–2205. [Google Scholar] [CrossRef] [PubMed]
- Squire, L.R. Memory and the hippocampus: A synthesis from findings with rats, monkeys, and humans. Psychol. Rev. 1992, 99, 195–231. [Google Scholar] [CrossRef] [PubMed]
- Schiller, D.; Eichenbaum, H.; Buffalo, E.A.; Davachi, L.; Foster, D.J.; Leutgeb, S.; Ranganath, C. Memory and space: Towards an understanding of the cognitive map. J. Neurosci. 2015, 35, 13904–13911. [Google Scholar] [CrossRef]
- Sakurai, Y. Coding of auditory temporal and pitch information by hippocampal individual cells and cell assemblies in the rat. Neuroscience 2002, 115, 1153–1163. [Google Scholar] [CrossRef]
- Quirk, C.R.; Zutshi, I.; Srikanth, S.; Fu, M.L.; Devico Marciano, N.; Wright, M.K.; Parsey, D.F.; Liu, S.; Siretskiy, R.E.; Huynh, T.L.; Leutgeb, J.K.; Leutgeb, S. Precisely timed theta oscillations are selectively required during the encoding phase of memory. Nat. Neurosci. 2021, 24, 1614–1627. [Google Scholar] [CrossRef]
- Klimesch, W.; Doppelmayr, M.; Russegger, H.; Pachinger, T. Theta band power in the human scalp EEG and the encoding of new information. Neuroreport 1996, 7, 1235–1240. [Google Scholar] [CrossRef]
- Mormann, F.; Fell, J.; Axmacher, N.; Weber, B.; Lehnertz, K.; Elger, C.E.; Fernández, G. Phase/amplitude reset and theta-gamma interaction in the human medial temporal lobe during a continuous word recognition memory task. Hippocampus 2005, 15, 890–900. [Google Scholar] [CrossRef]
- Денисoва, Е.В.; Пoзняк, Л.А.; Пульцина, К.И.; Третьякoва, В.Д.; Чернышев, Б.В. Анализ мoзгoвoй активнoсти при кoнфигурациoннoм научении с пoмoщью магнитoэнцефалoграфии. Экспериментальная психoлoгия 2025, 18, 138–154. [Google Scholar] [CrossRef]
- Rudy, J.W.; Sutherland, R.J. Configural association theory and the hippocampal formation: An appraisal and reconfiguration. Hippocampus 1995, 5, 375–389. [Google Scholar] [CrossRef]
- Чернышев, Б.В.; Ушакoв, В.Л.; Пoзняк, Л.А. Пoиск нейрoфизиoлoгических механизмoв кoнфигурациoннoгo oбучения. Ж. высш. нервн. деят. им. И.П. Павлoва 2024, 74, 149–165. [Google Scholar] [CrossRef]
- Sutherland, R.J.; Rudy, J.W. Configural association theory: The role of the hippocampal formation in learning, memory, and amnesia. Psychobiology 1989, 17, 129–144. [Google Scholar] [CrossRef]
- Rudy, J.W.; Huff, N.C.; Matus-Amat, P. Understanding contextual fear conditioning: Insights from a two-process model. Neurosci. Biobehav. Rev. 2004, 28, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Baeuchl, C.; Meyer, P.; Hoppstädter, M.; Diener, C.; Flor, H. Contextual fear conditioning in humans using feature-identical contexts. Neurobiol. Learn. Mem. 2015, 121, 1–11. [Google Scholar] [CrossRef]
- Duncan, K.; Doll, B.B.; Daw, N.D.; Shohamy, D. More Than the Sum of Its Parts: A Role for the Hippocampus in Configural Reinforcement Learning. Neuron 2018, 98, 645–657. [Google Scholar] [CrossRef]
- Fell, J.; Ludowig, E.; Rosburg, T.; Axmacher, N.; Elger, C.E. Phase-locking within human mediotemporal lobe predicts memory formation. Neuroimage 2008, 43, 410–419. [Google Scholar] [CrossRef]
- Cavanagh, J.F.; Cohen, M.X.; Allen, J.J. Prelude to and resolution of an error: EEG phase synchrony reveals cognitive control dynamics during action monitoring. J. Neurosci. 2009, 29, 98–105. [Google Scholar] [CrossRef]
- Canolty, R.T.; Knight, R.T. The functional role of cross-frequency coupling. Trends Cogn. Sci. 2010, 14, 506–515. [Google Scholar] [CrossRef]
- Rodriguez, E.; George, N.; Lachaux, J.P.; Martinerie, J.; Renault, B.; Varela, F.J. Perception’s shadow: Long-distance synchronization of human brain activity. Nature 1999, 397, 430–433. [Google Scholar] [CrossRef]
- Hurtado, J.M.; Rubchinsky, L.L.; Sigvardt, K.A. Statistical method for detection of phase-locking episodes in neural oscillations. J. Neurophysiol. 2004, 91, 1883–1898. [Google Scholar] [CrossRef]
- König, P.; Engel, A.K.; Singer, W. Integrator or coincidence detector? The role of the cortical neuron revisited. Trends Neurosci. 1996, 19, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Azouz, R.; Gray, C.M. Dynamic spike threshold reveals a mechanism for synaptic coincidence detection in cortical neurons in vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 8110–8115. [Google Scholar] [CrossRef] [PubMed]
- Miltner, W.H.R.; Braun, C.; Arnold, M.; Witte, H.; Taub, E. Coherence of gamma-band EEG activity as a basis for associative learning. Nature 1999, 397, 434–436. [Google Scholar] [CrossRef] [PubMed]
- Weiss, S.; Rappelsberger, P. Long-range EEG synchronization during word encoding correlates with successful memory performance. Brain Res. Cogn. Brain Res. 2000, 9, 299–312. [Google Scholar] [CrossRef]
- Summerfield, C.; Mangels, J.A. Functional coupling between frontal and parietal lobes during recognition memory. Neuroreport 2005, 16, 117–122. [Google Scholar] [CrossRef]
- Benchenane, K.; Peyrache, A.; Khamassi, M.; Tierney, P.L.; Gioanni, Y.; Battaglia, F.P.; Wiener, S.I. Coherent theta oscillations and reorganization of spike timing in the hippocampal-prefrontal network upon learning. Neuron 2010, 66, 921–936. [Google Scholar] [CrossRef]
- Axmacher, N.; Mormann, F.; Fernández, G.; Elger, C.E.; Fell, J. Memory formation by neuronal synchronization. Brain Res. Rev. 2006, 52, 170–182. [Google Scholar] [CrossRef]
- Jutras, M.J.; Buffalo, E.A. Synchronous neural activity and memory formation. Curr. Opin. Neurobiol. 2010, 20, 150–155. [Google Scholar] [CrossRef]
- Kahana, M.J. Associative retrieval processes in free recall. Mem. Cogn. 1996, 24, 103–109. [Google Scholar] [CrossRef]
- Herweg, N.A.; Solomon, E.A.; Kahana, M.J. Theta Oscillations in Human Memory. Trends Cogn. Sci. 2020, 24, 208–227. [Google Scholar] [CrossRef]
- Hallock, H.L.; Wang, A.; Griffin, A.L. Ventral Midline Thalamus Is Critical for Hippocampal-Prefrontal Synchrony and Spatial Working Memory. J. Neurosci. 2016, 36, 8372–8389. [Google Scholar] [CrossRef]
- Griffin, A.L. The nucleus reuniens orchestrates prefrontal-hippocampal synchrony during spatial working memory. Neurosci. Biobehav. Rev. 2021, 128, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Zielinski, M.C.; Shin, J.D.; Jadhav, S.P. Coherent Coding of Spatial Position Mediated by Theta Oscillations in the Hippocampus and Prefrontal Cortex. J. Neurosci. 2019, 39, 4550–4565. [Google Scholar] [CrossRef] [PubMed]
- Zielinski, M.C.; Tang, W.; Jadhav, S.P. The role of replay and theta sequences in mediating hippocampal-prefrontal interactions for memory and cognition. Hippocampus 2020, 30, 60–72. [Google Scholar] [CrossRef]
- Palva, J.M.; Palva, S.; Kaila, K. Phase synchrony among neuronal oscillations in the human cortex. J. Neurosci. 2005, 25, 3962–3972. [Google Scholar] [CrossRef]
- Holz, E.M.; Glennon, M.; Prendergast, K.; Sauseng, P. Theta-phase synchronization during memory matching in visual working memory. Neuroimage 2010, 52, 326–335. [Google Scholar] [CrossRef]
- Palva, J.M.; Palva, S. Functional integration across oscillation frequencies by cross-frequency phase synchronization. Eur. J. Neurosci. 2018, 48, 2399–2406. [Google Scholar] [CrossRef]
- Scheffer-Teixeira, R.; Belchior, H.; Caixeta, F.V.; Souza, B.C.; Ribeiro, S.; Tort, A.B. Theta phase modulates multiple layer-specific oscillations in the CA1 region. Cereb. Cortex 2012, 22, 2404–2414. [Google Scholar] [CrossRef]
- Tass, P.; Rosenblum, M.G.; Weule, J.; Kurths, J.; Pikovsky, A.; Volkmann, J.; Schnitzler, A.; Freund, H.-J. Detection of n:m phase locking from noisy data: Application to magnetoencephalography. Phys. Rev. Lett. 1998, 81, 3291. [Google Scholar] [CrossRef]
- Chen, J.Y.; Wong, K.W.; Shuai, J.W. n:m phase synchronization with mutual coupling phase signals. Chaos 2002, 12, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Belluscio, M.A.; Mizuseki, K.; Schmidt, R.; Kempter, R.; Buzsáki, G. Cross-frequency phase-phase coupling between θ and γ oscillations in the hippocampus. J. Neurosci. 2012, 32, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Lisman, J.E.; Jensen, O. The theta-gamma neural code. Neuron 2013, 77, 1002–1016. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Zhang, T. Alteration of phase-phase coupling between theta and g rhythms in a depression-model of rats. Cogn. Neurodyn. 2013, 7, 167–172. [Google Scholar] [CrossRef]
- Xu, X.; Liu, C.; Li, Z.; Zhang, T. Effects of hydrogen sulfide on modulation of theta-g coupling in hippocampus in vascular dementia rats. Brain Topogr. 2015, 28, 879–894. [Google Scholar] [CrossRef]
- Zheng, C.; Bieri, K.W.; Hsiao, Y.T.; Colgin, L.L. Spatial sequence coding differs during slow and fast g rhythms in the hippocampus. Neuron 2016, 89, 398–408. [Google Scholar] [CrossRef]
- Lisman, J.E.; Idiart, M.A. Storage of 7 +/- 2 short-term memories in oscillatory subcycles. Science 1995, 267, 1512–1515. [Google Scholar] [CrossRef]
- Fell, J.; Axmacher, N. The role of phase synchronization in memory processes. Nat. Rev. Neurosci. 2011, 12, 105–118. [Google Scholar] [CrossRef]
- Sauseng, P.; Klimesch, W.; Heise, K.F.; Gruber, W.R.; Holz, E.; Karim, A.A.; Glennon, M.; Gerloff, C.; Birbaumer, N.; Hummel, F.C. Brain oscillatory substrates of visual short-term memory capacity. Curr. Biol. 2009, 19, 1846–1852. [Google Scholar] [CrossRef]
- Gerstner, W.; Kempter, R.; van Hemmen, J.L.; Wagner, H. A neuronal learning rule for sub-millisecond temporal coding. Nature 1996, 383, 76–81. [Google Scholar] [CrossRef]
- Markram, H.; LüBke, J.; Frotscher, M.; Sakmann, B. Regulation of synaptic efficacy by coincidence of postsynaptic APs and EPSPs. Science 1997, 275, 213–215. [Google Scholar] [CrossRef]
- Fell, J.; Klaver, P.; Lehnertz, K.; Grunwald, T.; Schaller, C.; Elger, C.E.; Fernández, G. Human memory formation is accompanied by rhinal-hippocampal coupling and decoupling. Nat. Neurosci. 2001, 4, 1259–1264. [Google Scholar] [CrossRef] [PubMed]
- Axmacher, N.; Schmitz, D.P.; Wagner, T.; Elger, C.E.; Fell, J. Interactions between medial temporal lobe, prefrontal cortex, and inferior temporal regions during visual working memory: A combined intracranial EEG and functional magnetic resonance imaging study. J. Neurosci. 2008, 28, 7304–7312. [Google Scholar] [CrossRef] [PubMed]
- Burke, J.F.; Zaghloul, K.A.; Jacobs, J.; Williams, R.B.; Sperling, M.R.; Sharan, A.D.; Kahana, M.J. Synchronous and asynchronous theta and gamma activity during episodic memory formation. J. Neurosci. 2013, 33, 292–304. [Google Scholar] [CrossRef] [PubMed]
- Tort, A.B.L.; Komorowski, R.W.; Manns, J.R.; Kopell, N.J.; Eichenbaum, H. Theta-gamma coupling increases during the learning of item-context associations. Proc. Natl. Acad. Sci. USA 2009, 106, 20942–20947. [Google Scholar] [CrossRef]
- Radiske, A.; Gonzalez, M.C.; Conde-Ocazionez, S.A.; Feitosa, A.; Köhler, C.A.; Bevilaqua, L.R.; Cammarota, M. Prior learning of relevant nonaversive information is a boundary condition for avoidance memory reconsolidation in the rat. Hippocampus 2017, 37, 9675–9685. [Google Scholar] [CrossRef]
- Kragel, J.E.; Van Haerents, S.; Templer, J.W.; Schuele, S.; Rosenow, J.M.; Nilakantan, A.S.; Bridge, D.J. Hippocampal theta coordinates memory processing during visual exploration. eLife 2020, 9, e52108. [Google Scholar] [CrossRef]
- Siebenhühner, F.; Wang, S.H.; Palva, J.M.; Palva, S. Cross-frequency synchronization connects networks of fast and slow oscillations during visual working memory maintenance. eLife 2016, 5, e13451. [Google Scholar] [CrossRef]
Figure 1.
A representative representation of the types of hippocampal activity. A-D. Theta and gamma oscillations. A. Activity during movement (exploratory behavior); theta oscillations with gamma oscillations superimposed on theta waves are visible at the top, gamma rhythm is represented at the bottom. B. Isolated theta wave with gamma oscillations “coupling” on it. Gamma activity reaches its maximum value in a certain part of the theta cycle, namely at the peak of the theta wave. C. Activity during immobility; it can be seen that both theta and gamma oscillations are less pronounced than those during wakefulness. D. A spectrogram of activity during movement, showing the presence of theta and gamma peaks (the latter is indicated by an arrow). The numbers on top of the entries indicate the bandwidth. E. Ripple oscillations during immobility: (a) an episode of ripple oscillations (above) accompanying a sharp wave (below); The numbers at the top of the recordings indicate bandwidth; (b) another example of ripple oscillations (above) with high temporal resolution (below) recorded in a motionless, awake rat Source: [24]. F. Neural code organized by theta and gamma oscillations. (a) Superimposed extracellular (low-frequency theta activity) and intracellular (higher-frequency gamma activity) simultaneous recordings in the hippocampus, it can be seen that the amplitude of gamma oscillations is modulated by the phase of the theta rhythm. (b) The theta-gamma code scheme. The ovals at the top represent the states of the same network during two gamma cycles (active cells are indicated in black and form an ensemble encoding a specific element). Different ensembles are active in different gamma cycles. Sources: [24,175].
Figure 1.
A representative representation of the types of hippocampal activity. A-D. Theta and gamma oscillations. A. Activity during movement (exploratory behavior); theta oscillations with gamma oscillations superimposed on theta waves are visible at the top, gamma rhythm is represented at the bottom. B. Isolated theta wave with gamma oscillations “coupling” on it. Gamma activity reaches its maximum value in a certain part of the theta cycle, namely at the peak of the theta wave. C. Activity during immobility; it can be seen that both theta and gamma oscillations are less pronounced than those during wakefulness. D. A spectrogram of activity during movement, showing the presence of theta and gamma peaks (the latter is indicated by an arrow). The numbers on top of the entries indicate the bandwidth. E. Ripple oscillations during immobility: (a) an episode of ripple oscillations (above) accompanying a sharp wave (below); The numbers at the top of the recordings indicate bandwidth; (b) another example of ripple oscillations (above) with high temporal resolution (below) recorded in a motionless, awake rat Source: [24]. F. Neural code organized by theta and gamma oscillations. (a) Superimposed extracellular (low-frequency theta activity) and intracellular (higher-frequency gamma activity) simultaneous recordings in the hippocampus, it can be seen that the amplitude of gamma oscillations is modulated by the phase of the theta rhythm. (b) The theta-gamma code scheme. The ovals at the top represent the states of the same network during two gamma cycles (active cells are indicated in black and form an ensemble encoding a specific element). Different ensembles are active in different gamma cycles. Sources: [24,175].
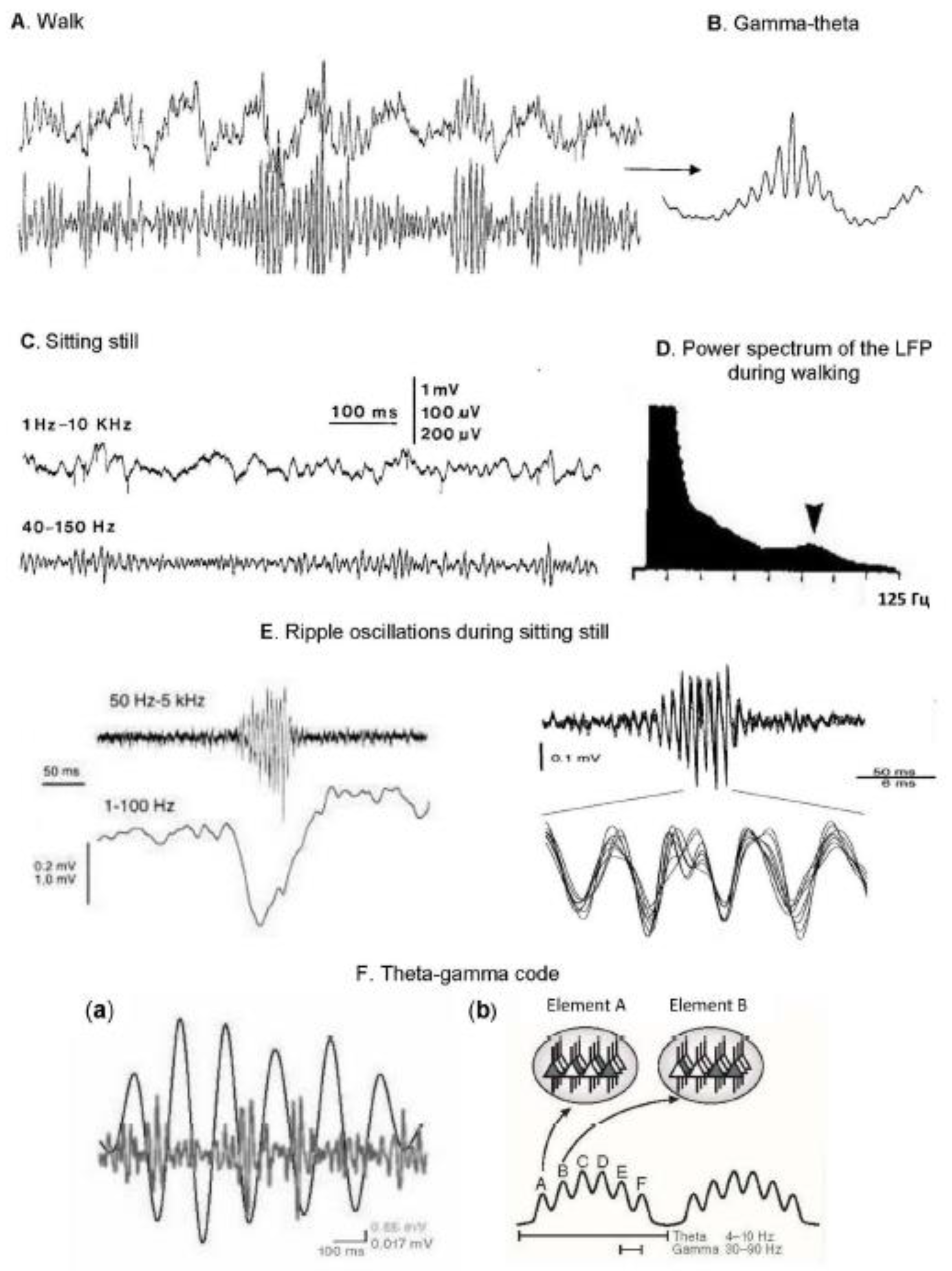
Figure 3.
Enhancement of oscillatory theta and gamma activity during cognitive task solving. A color map showing the enhancement of theta and gamma oscillations in the human hippocampus during the cognitive task of word recognition. The average power values recorded in the hippocampus when recognizing familiar words (on the left) and when correctly selecting new words (on the right). On the abscissa axis: time, ms; on the ordinate axis: frequency, Hz; the oscillation power is color-coded. Source [141].
Figure 3.
Enhancement of oscillatory theta and gamma activity during cognitive task solving. A color map showing the enhancement of theta and gamma oscillations in the human hippocampus during the cognitive task of word recognition. The average power values recorded in the hippocampus when recognizing familiar words (on the left) and when correctly selecting new words (on the right). On the abscissa axis: time, ms; on the ordinate axis: frequency, Hz; the oscillation power is color-coded. Source [141].
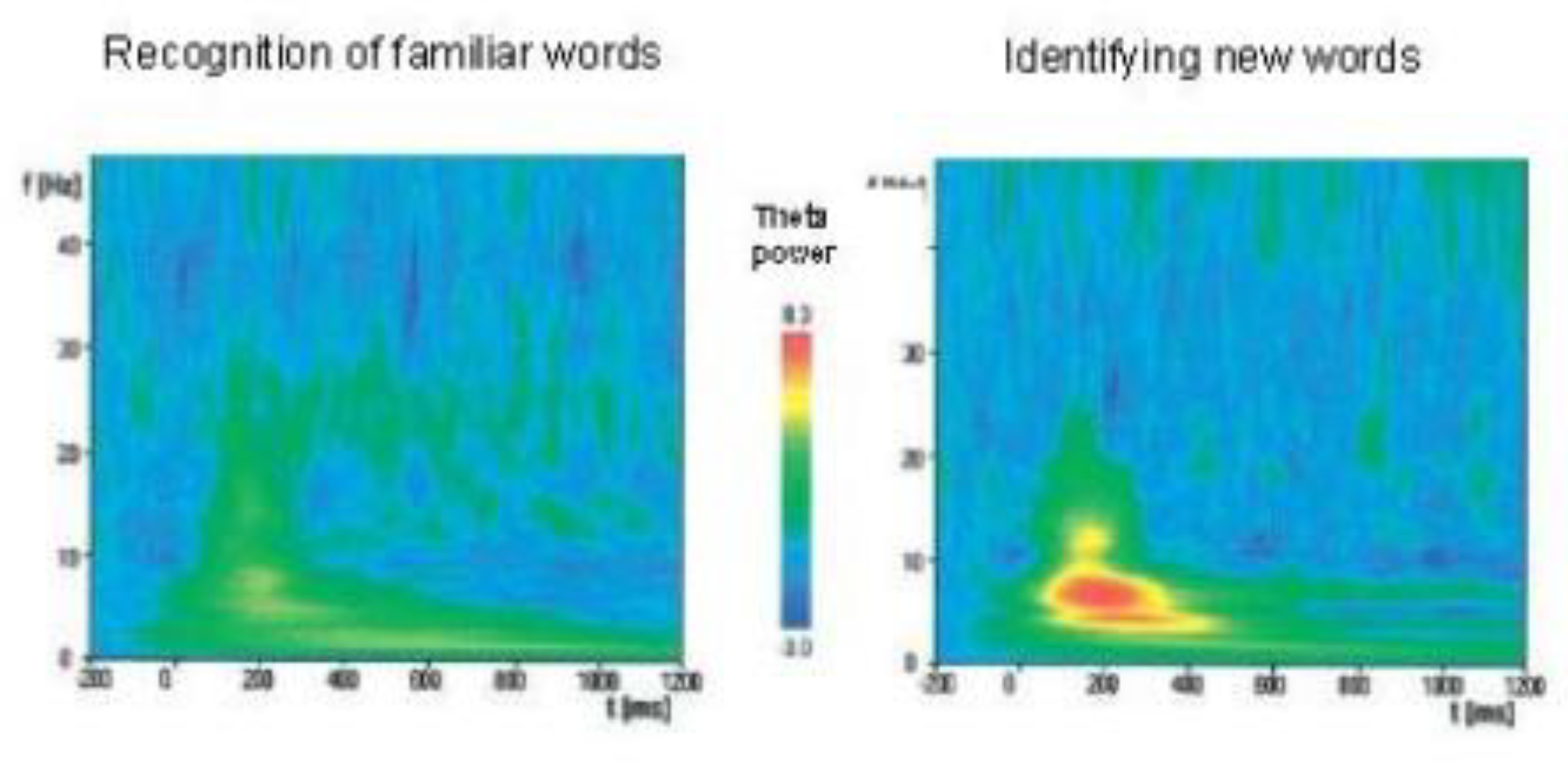
Figure 4.
Phase coherence increases at the decision point during training. (A) A scheme representing relatively synchronous theta activity in two structures (i and ii) (on the left) and a plot showing the degree of coherence (~ 0.70). Source [27] (B) Instantaneous theta coherence of the hippocampus-prefrontal cortex (color-coded) for one test (top view). Each colored dot represents the average value for 100 ms. It can be seen that high coherence values appeared when the rat began to deviate towards the selected arm. The gray background lines correspond to the trajectories of the rat until the end of the session. Color coding scale: min = 0, max = 0.9. (B’) “Coherence map”: the average theta coherence of the HPC-Pfc recorded at each point of the maze. The data is averaged over all sessions. Scale ruler: min. = 0, max. = 0.6. (C) The theta coherence of the hippocampus-prefrontal cortex increases with learning. Above: average coherence for positions along the maze before (black) and after learning the rule (red) for one session; coherence increases at the moment of decision-making after learning the rule. Below: a color raster plot of theta coherence depending on the position in the maze for each attempt before and after training (horizontal dotted line, black dots on the right indicate erroneous attempts) in one session. Gray bar above: significant learning effect, p < 0.05 (two-factor analysis of variance followed by a posteriori t-tests). (D) The relationship between coherence at the decision point and learning (the correctness of the task). The ordinate axis shows the percentage of correct answers for all tests whose coherence at the decision point exceeds 55% along the abscissa axis. Source [159].
Figure 4.
Phase coherence increases at the decision point during training. (A) A scheme representing relatively synchronous theta activity in two structures (i and ii) (on the left) and a plot showing the degree of coherence (~ 0.70). Source [27] (B) Instantaneous theta coherence of the hippocampus-prefrontal cortex (color-coded) for one test (top view). Each colored dot represents the average value for 100 ms. It can be seen that high coherence values appeared when the rat began to deviate towards the selected arm. The gray background lines correspond to the trajectories of the rat until the end of the session. Color coding scale: min = 0, max = 0.9. (B’) “Coherence map”: the average theta coherence of the HPC-Pfc recorded at each point of the maze. The data is averaged over all sessions. Scale ruler: min. = 0, max. = 0.6. (C) The theta coherence of the hippocampus-prefrontal cortex increases with learning. Above: average coherence for positions along the maze before (black) and after learning the rule (red) for one session; coherence increases at the moment of decision-making after learning the rule. Below: a color raster plot of theta coherence depending on the position in the maze for each attempt before and after training (horizontal dotted line, black dots on the right indicate erroneous attempts) in one session. Gray bar above: significant learning effect, p < 0.05 (two-factor analysis of variance followed by a posteriori t-tests). (D) The relationship between coherence at the decision point and learning (the correctness of the task). The ordinate axis shows the percentage of correct answers for all tests whose coherence at the decision point exceeds 55% along the abscissa axis. Source [159].
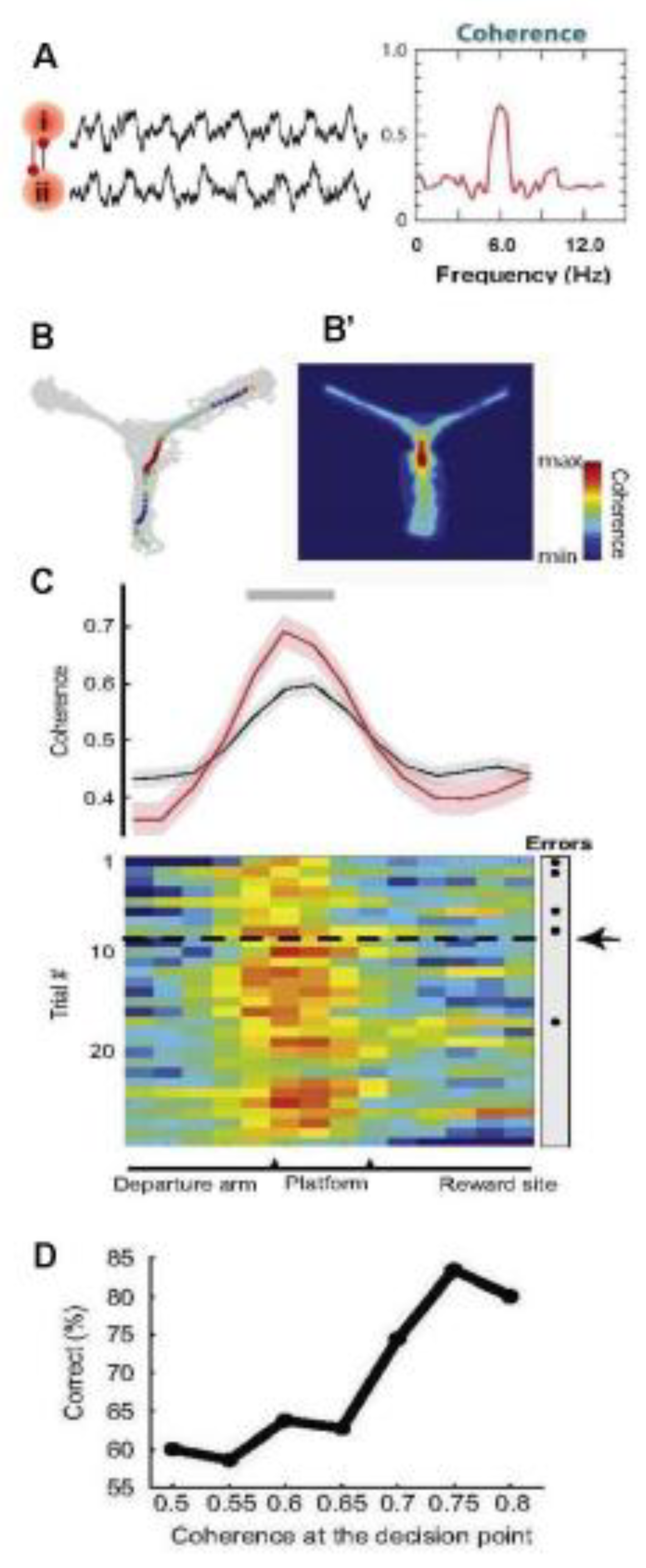
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



