
Submitted:
20 November 2025
Posted:
24 November 2025
You are already at the latest version
Abstract
Life is defined by self-governing networks of molecules changing conformation cyclically to convert thermodynamic motion into directional work that creates structure. A spectrum of scale, from nanoscopic to macroscopic, involves a shift from intracellular thermody-namically driven processes (thermal agitation ultimately rooted in quantum phenomena) to intercellular bulk flows described by classical physics; from short-distance transport involving diffusion and cytoskeletal transport to long-distance pressure fluxes in hydrau-lic networks. A review of internal transport systems in macroscopic eukaryotes suggests that a key evolutionary step favoring large size and multicellularity involved exploiting molecular-scale stochasticity to generate organized bulk flows (e.g. motor proteins collec-tively generating mechanical pressures in metazoan tissues such as cardiac muscle; within tracheophytes, active and passive phloem loading/unloading inducing pressure gradients, and active regulation enabling passive xylem function and hydraulic reliability; sieve-like conduction in heterokonts; peristaltic shuttle streaming in myxogastrian plas-modia). All macroscopic physiologies are underpinned by Brownian dynamics and thus quantum mechanics. Although well documented separately, acknowledgment of the role of quantum mechanics as the foundation of physiology unites the smallest single cells with the largest multicellular organisms across the tree of life; demonstrating how all large biological organisms represent an outgrowth of the smallest scales of physics.
Keywords:
systems biology
; transport networks
; diffusion
; bulk flow
; multicellularity
; cytoplasmic streaming
; evolutionary transition
1. Introduction
Living systems are self-governing networks of components that function collectively to create structure; networks of macromolecules that change conformation cyclically and dependably along one plane, exploiting thermodynamic gradients to induce directional motion that does work to consolidate matter (reduce entropy) [1−4]. A fundamental requirement for the operation of these networks is motion; not only the characteristic uniplanar motions (ratcheting or spinning [4−7]) of the single macromolecules and multi-subunit complexes, but also the fluidity of membranes and the internal transport of substrates and products to and from different regions within the organism. Constraints to the transport of material within organisms determine the capacity to scale in size and complexity [8,9] and are a key issue that different taxonomic groups have evidently overcome in contrasting ways during the evolutionary history of life on Earth.
Indeed, the evolution of large size, in both unicellular and multicellular organisms, involved surmounting the excessive time involved for transport over relatively large internal distances when relying on processes driven solely by thermodynamics (i.e., physical work/energy relationships), a critical limitation to the supply of substrates to metabolism and the elimination of waste products and thus physiological integration [10]. In terms of the physical forces that drive transport processes, the common underlying problem faced by all groups of organisms is that of a transition from extremely small-scale nanoscopic networks of molecules (in which thermodynamics and its causal quantum mechanical processes determine particle motion and flows) to the macroscopic, in which movement of bulk matter obeys the rules of classical physics and is essentially determined by pressure gradients. This essentially represents a transition from quantum to classical mechanics.
2. The Quantum to Classical Boundary and Its Relevance to Biology
Why is a rather abstract concept from physics – the quantum to classical boundary – of relevance to biology? Quantum mechanics describes particle superpositions, interference and entanglement [11,12,13,14], while classical mechanics depicts definite states of matter and trajectories. Yet evidently the classical world emerges from the quantum realm. For example, an electron (a localised excitation in the quantum field permeating the universe) is described by a wavefunction that gives the probability of finding it in a particular state or position [14]. But until it is observed (i.e. interacts with something) the electron effectively exists in superpositions of states - in multiple places or momentums at once - of which we cannot be certain. The same ‘quantum uncertainty’ applies to the subatomic particles making up atomic nuclei, and even to the state of the entire atom, meaning that in practice even when atoms are at rest they oscillate and vibrate in place (even at 0° K; hence the term ‘zero-point energy’ [15]). Atoms also possess kinetic energy and, especially when in solid lattices, transfer this to one another by virtual particles called phonons (i.e. they jostle one another, the phonon representing the interaction). In real terms, atoms and molecules spontaneously jiggle (an informal but serious term often used by Richard Feynman to explain a phenomenon that is difficult encapsulate in English [16]) with a combination of thermal (kinetic) and zero-point energy. This is the quantum mechanical basis of emergent random particle motion (thermal agitation or Brownian motion [17]) and is ultimately the physical driving force of classical physical processes such as diffusion along concentration gradients [18,19]).
Another link between quantum and classical worlds, and how this relates to the physical scale and size of organisms, is decoherence, or the cancellation of quantum effects when particles interact with (are ‘observed by’) their environment [20,21,22]. Decoherence is faster for systems of larger mass because larger numbers of atoms participate in more interactions with atoms and photons in the environment. Thus as mass increases decoherence is more likely (coherence time and quantum effects decrease), and a system moves along a continuum from quantum to classical. Single macromolecules in the ‘warm-wet’ environments inside organisms are at the upper mass limit for decoherence. Nonetheless, and despite the energetic environmental context, the atoms comprising them can exhibit coherence and quantum mechanical effects [e.g. 23-26], so much so that ‘quantum biology’ is now a recognized subdiscipline [27,28]. Essentially, their motion is an emergent effect of their own inherent thermal agitation but particularly of bombardment by smaller thermally agitated molecules such as those of water. In this thermal bath, macromolecules and molecular sub-unit complexes such as ribosomes, RNA polymerase, motor proteins and other enzymes continuously explore a broad conformational energy landscape (i.e. change shape semi-randomly depending on interactions with the environment), with fluctuations and transitions that are essential for their function. Rather than being hindered by thermal noise they harness it, converting random motion into directional motion and reliable activity [2−6]. Thermal agitation of molecules and diffusion gradients (especially when energetic barriers are overcome by energy inputs such as excitation donated by photons during photosynthesis) are the fundamental physical processes driving biology and ultimately emerge from quantum mechanical effects. Note that the ‘energy carrier’ nucleotide adenosine triphosphate (ATP) is created by trans-membrane electrochemical gradients driven mainly by diffusion; it carries energy derived from thermodynamic gradients, and so from thermal agitation, Brownian motion and thus ultimately from the quantum uncertainty that underpins atomic and molecular motion (Figure 1). Note also that other nucleotides, such as guanosine triphosphate (GTP) are synthesized by enzymes driven by ATP hydrolysis, and so essentially carry energy previously carried by ATP and thus again ultimately from thermodynamic disequilibria.
3. Biological Phenomena Driven by Quantum Mechanical Effects
Enzymatic catalysis exemplifies the role of thermal agitation in function. Enzymes undergo thermally driven conformational changes that align catalytic substrates into productive geometries, transiently stabilizing transition states and lowering activation barriers [29,30]. ATP is required to overcome energy barriers, inducing suitable conformation changes that essentially activate the enzyme, but enzyme motion during function is also driven directly by thermal agitation and bombardment [4,5,6]. Dependence on thermal agitation appears to be true of all catalytic globular protein complexes [31−33]. For example, RNA polymerase relies on stochastic conformational changes for its operational movement along the DNA template during transcription; local thermal fluctuations enable forward and backward motion of the polymerase, with nucleotide incorporation (of ATP, but also GTP, UTP or CTP depending on the DNA template) biasing the process toward net elongation (i.e. RNA polymerization) [34]. Note that in this case nucleotides bias the direction of the process, but ATP is a component incorporated into RNA rather than simply being an energy carrier – the main source of energy is thermal agitation and bombardment (and thus quantum mechanical effects).
Similarly, the ribosome, a large ribonucleoprotein complex, exhibits extensive Brownian motions during translation, including inter-subunit rotation [35−39]. These thermally accessible conformational changes, coordinated by GTP hydrolysis, ensure the fidelity and directionality of protein synthesis.
Eukaryotic motor proteins such as myosin, kinesin and dynein exemplify the conversion of energy into mechanical work under conditions dominated by thermal bombardment. Their movement along actin filaments or microtubules is inherently stochastic, consisting of cycles of binding, conformational change, and diffusion. ATP hydrolysis biases these thermally driven fluctuations, contributing to net directed motion. The coupling between random molecular motion and energy rectification, often modelled as a Brownian ratchet, underscores the fundamental principle of molecular biophysics that thermal noise is not suppressed but harnessed to perform work with high efficiency [3,5,6,34,35,36,37,38,39]. Note that within myocytes (contractile muscle cells) Brownian motion and ATP hydrolysis induce the single myosin molecules to pull against actin, evident collectively at the macroscopic scale literally as the physical force with which the muscle contracts [40].
Another example of a thermally coupled molecular machine is the rotary enzyme ATP synthase. This complex, found in mitochondria, chloroplasts and bacterial membranes, operates as a reversible rotary motor (Figure 1). Diffusion of protons through the membrane-embedded F0 motor generates torque driving rotation of the central stalk [41], which pushes against subunits of the catalytic F1 region, inducing “hinged” [42] conformational changes central to synthesizing ATP from ADP and inorganic phosphate. Conversely, hydrolysis of ATP can drive reverse rotation, with the enzyme functioning as an ATP-driven proton pump. Thermal fluctuations facilitate subunit rotation and conformational transitions across energy barriers. In essence, the proton-motive force does not impose deterministic motion but rather biases the probability distribution of rotational states, leading to net directional rotation along one plane and broadly linear conformation cycles in catalytic F1 subunits. A similar conformational and energetic mechanism is apparent for ATPase, which typically uses ATP to drive rotational motion, setting up electrochemical gradients [43,44] (N.B. structurally, ATPsynthase is considered a subclass of ATPases, the suffixes referring rather to different primary functions; essentially tending to produce or to use ATP). The operation of ATP synthase and ATPase enzymes exemplify the principle that biological nanomachines exploit thermal agitation (derived from underlying quantum mechanical effects) as an integral component of their mechanochemical cycles.
4. Diffusion Limitation to Organism Size
Diffusion, governed by Brownian motion and ultimately quantum mechanics, dominates at small scales. But as volume increases, the time for diffusion of a molecule across a body grows approximately with the square of the distance, while the capacity for uptake via surface area grows linearly, thus imposing a surface-area-to-volume constraint on size. For example, in plants, manipulation of Fick’s second law [45] demonstrates that beyond a certain size passive diffusion is insufficient and bulk-flow systems become necessary. Indeed, without the evolution of systems for the translocation of volumes of fluids (bulk flow), organisms would have remained only a few millimetres tall [46,47,48]. Conversely, advection and bulk flow allow much larger size by distributing nutrients, signalling molecules and wastes more rapidly than diffusion alone.
Before true bulk-flow vascular systems can evolve, organisms exploit intermediate mechanisms. For instance, cytoplasmic streaming is a large-scale movement of cytoplasm in eukaryotic cells, induced by molecular motor systems working on the cytoskeleton, to overcome diffusion limitations in cells of unusually large size. Cytoplasmic streaming and bulk intercellular flow become necessary when cell diameters exceed ~100 µm [49]. At even smaller scales, intracellular transport on the cytoskeleton blends diffusion and directed motion: cytoskeletal network morphology strongly regulates intracellular transport dynamics and times [50,51]. Thus, within cells we see a gradient from diffusion, motor-driven transport through to streaming/advection in the liquid cytoplasm, which is essentially a small-scale bulk flow driven not by pressure gradients but propelled by the collective motions of hundreds of thousands (to millions, depending on cell type) of working molecules.
Crucially, the evolution of multicellularity extends these cellular transport modes beyond the single-cell scale. A key stage in the evolution of multicellularity was the evolution of colonial life, and this involves intermediate transport mechanisms external to single cells. In green algae of the order Volvocales (e.g., Volvox carteri F.Stein; Figure 1), the coordinated beating of flagella at the colony surface generates fluid flow that enhances nutrient uptake: in this system advection (transport via bulk motion of fluid) produces a nutrient-exchange rate that is quadratic in radius, overcoming the diffusion limitation locally. This illustrates an early form of bulk flow at the colony scale (i.e. ‘quasi’ multicellular). The transition to full-blown hydraulic (pressurized) systems in various clades also involves the intermediate evolutionary step of pseudo-circulatory mechanisms inducing local bulk flow in internal volumes of liquids.
5. Internal Long-Distance Transport Systems in Fungi
Of the eukaryotes, the first taxa known to use long distance, cell-to-cell transport were the fungi, evolving these phenotypes (and by inference these physiological systems) probably as early as ~1.4 Ga (billion years ago) [52]. In filamentous fungi, hyphae are composed of cells connected end-to-end and compartmentalised by perforated septa that allow the cytoplasm to remain largely continuous across the colony. This configuration produces a multinucleate, ‘syncytial’ cytoplasm (a continuous intercellular protoplast) through which water, ions, metabolites, and organelles can move freely [53,54]. In many fungi, anastomoses (hyphal fusions) further increase cytoplasmic connectivity, enabling extensive internal transport and resource sharing across the network. Passive transport processes within the fungal syncytium are primarily based on diffusion and pressure-driven bulk flow. Small solutes such as sugars, amino acids and ions can move passively through the cytoplasm and across septal pores following concentration gradients [55]). In addition, differences in turgor pressure between hyphal compartments may generate cytoplasmic streaming and bulk flow, which facilitate long-distance transport of solutes and water [56]). This is somewhat analogous to the pressure-flow mechanism of phloem transport in plants (see Section 8), although in fungi the driving forces are localized and transient rather than maintained by specialized source-sink relations.
Active transport processes provide regulation and directionality within the syncytium. Membrane transport proteins, including proton pumps (H+-ATPases), maintain electrochemical gradients that energize the secondary active uptake or efflux of nutrients such as sugars, amino acids, phosphate and metal ions [57,58]. This energy-dependent ion transport contributes to the maintenance of internal pH and turgor pressure homeostasis, both of which are essential for hyphal growth and intracellular circulation [53]. Furthermore, vesicle trafficking and organelle movement within hyphae rely on cytoskeletal transport using motor proteins such as myosins, kinesins and dyneins, which actively transport cargo along actin filaments and microtubules [59,60]). These mechanisms facilitate the targeted delivery of enzymes, lipids and wall-building materials to growing hyphal tips, as well as the redistribution of metabolites from older to actively extending regions of the colony. The integration of active and passive transport ensures that the fungal syncytium functions as a coherent and adaptive physiological system. Energy-demanding ion transport and motor-driven movement (based on quantum mechanical-driven biomolecular activity) establish and maintain turgor and concentration gradients; active generation of conditions that induce passive diffusion.
6. Internal Transport in Plasmodial Myxogastria (Slime Molds)
The crown-group of the Amoebozoan class Myxogastria (plasmodial slime-moulds) diverged at an estimated 661–823 Ma [61]. The vegetative or plasmodial stage of the slime mold Physarum polycephalum Schwein exhibits an exceptional form of internal transport that enables coordination and resource distribution across its macroscopic body, which may span several centimetres to meters in size (networks of ≥ 10 cm² have been investigated in the laboratory). In the plasmodial stage, Physarum exists as a single, multinucleate cell organized into a dynamic network of tubular veins through which cytoplasm is rhythmically shuttled in alternating directions in a process known as ‘shuttle streaming’. Unlike cytoplasmic streaming in the organisms considered above, movement is driven by periodic contractions of an actomyosin cortex in the vein walls, producing peristaltic waves propelling cytoplasm back and forth throughout the network at speeds of up to 0.048 mm s⁻¹ [62,63]; slow compared to transport systems in other eukaryotes, but nonetheless an order of magnitude faster than diffusion for large molecules such as proteins.
The mechanism underlying this transport is biophysical and biochemical: contractions of the actin–myosin system are regulated by calcium concentrations, forming a Ca²⁺-dependent contractile oscillator that sets the frequency and amplitude of streaming [64]. Oscillations propagate as phase waves along the network, to create peristaltic motion ensuring efficient advection of solutes and organelles over macroscopic distances [65]. Experimental and theoretical studies have demonstrated that the plasmodium regulates its contraction wavelength and phase relationships depending on its current size, to optimize fluxes [62]. The resulting peristaltic flows achieve long-range transport far more effectively than diffusion alone, mixing and distributing metabolites across the entire organism [65,67].
Internal flows also serve a signalling function. Changes in environmental conditions such as nutrient availability or light stimuli induce localized alterations in contraction amplitude and frequency, which can propagate as waves of enhanced or suppressed contraction through the network, effectively transmitting information about the external environment [62]. Thus, the same physical processes that drive nutrient transport also underlie a primitive form of distributed physiological coordination.
The macroscopic size of plasmodial slime molds is fundamentally constrained by the mechanics of their internal transport system. Experimental and theoretical studies demonstrate that the wavelength and frequency of contractions during shuttle streaming must scale with organism size to maintain effective advective transport; mismatches between contraction patterns and body geometry reduce transport efficiency and can limit the functional size of the plasmodium [62,64]. Furthermore, physical constraints such as vein diameters and viscous dissipation set limits on transport rates, and thus on how large a single, integrated plasmodium can grow without altering its network architecture [64,65]. Computational and analytical models further support the conclusion that organismal size is limited by transport optimization, as network geometry and flow dynamics determine the maximum scale at which shuttle streaming can maintain physiological integration [62,64]. Thus, this system represents the upper size limit and greatest sophistication of long-distance transport within single-celled organisms, beyond which large size requires dedicated cell types and multicellular tissues.
Crucially, the shuttle streaming system may seem unique and unusual, but is fundamentally based on motion by thermodynamically responsive molecules (motor proteins, and in particular the actin/myosin cytoskeletal system also employed by other eukaryotes). It thus has a basis in the underlying quantum mechanical effects on the motion of matter, which then become evident as transport processes at the macroscopic scale.
7. Origin of Circulatory/Vascular Systems in Metazoa
The earliest multicellular animals (e.g., sponges, simple cnidarians), diverging 613.2 to 593.4 Ma in the Ediacaran period [67], lacked a dedicated vascular system; their tissues remained in direct contact with seawater or water within cavities, allowing diffusion-driven exchange [68,69,70]. With the evolution of triploblasty (i.e. three germ layers giving rise to associated tissue systems) and larger body size, animals required more effective internal transport. Comparative analyses suggest that a rudimentary blood vascular system first appeared in an ancestral bilaterian (581.8-569 Ma [66]), as a strategy to overcome the diffusion limit over larger transport distances [70,71]. This ancestral system likely consisted of coelomic cavities or channels lined by mesothelium rather than a true endothelium, combining features of lacunar and vessel-lined systems [68]. Multiple independent acquisitions of closed or semi-closed vascular systems are inferred, consistent with parallel evolution of circulatory architectures in diverse phyla [68]. The evolution of an endothelial lining occurred later in the vertebrate lineage (~420 Ma [66]), a step that allowed higher perfusion pressure (i.e. the driving force for blood flow), greater barrier control and functional specialization [69,70,72]. Thus, vascular evolution may be viewed as a sequence from diffusion-based transport, through lacunar/coelomic transport, primitive vascular networks, to closed endothelial-lined systems.
Specifically, over distances greater than around 100 µm diffusion, particularly of oxygen, becomes limiting; although cells such as Amoeba and Paramecium (cell diameter ~10 – 100 µm) are small enough to rely solely on diffusion, at the scale of nematodes (such as Caenorhabditis elegans Maupas, 1900; ~0.5 – 1 mm) diffusion is insufficient and a circulatory mechanism for pseudocoelomic fluid is required, based on muscular movement of the trunk which induces internal fluid movement. True, closed circulatory systems later evolved in annelids, mollusks, arthropods and vertebrates (Table 1).
Invertebrates exhibit a wide variety of circulatory systems, broadly categorized as open (hemocoelic) or closed (vessel-based) systems, though this distinction is often gradational [72]. In many arthropods (insects, crustaceans) and non-cephalopod mollusks, a contractile dorsal vessel pumps hemolymph into a hemocoel bathing the organs before returning to the heart via ostial openings. This arrangement is sufficient for small organisms or moderate activity levels [73]. In contrast, annelids possess a closed vascular system with dorsal and ventral longitudinal vessels and segmental interconnections [70]. In some species, such as Nereis japonica Izuka, 1908, the vessel walls lack a discrete endothelial layer and resemble interstitial spaces [73]. Pan-arthropods (Panarthropoda) such as onychophorans (“velvet worms”) show open vascular systems with peristaltic hearts and segmental ostia; probably the ancestral condition for arthropods [74].
Across invertebrates, the shift from diffusion to vessel-based transport correlates with increases in body size, metabolic rate, activity level and body plan complexity [70]. Furthermore, vascular evolution is intertwined with respiratory pigment evolution. The marine annelid Platynereis dumerilii Audouin & Milne-Edwards, 1834 exhibits a closed vascular system with extracellular hemoglobins; genomic analyses reveal that the ancestral bilaterian possessed at least five globin gene clades, indicating multiple independent recruitments of respiratory pigments alongside vascular innovations [75]. Thus, invertebrate vascular systems illustrate diverse evolutionary solutions, from open hemocoelic systems to rudimentary vessel networks, each adapted to the physiological demands of ecological circumstances.
Within deuterostomes (bilateral animals in which embryo development is characterized by the anus preceding gut and mouth development), the evolution of closed blood-vascular systems culminated in vertebrates with endothelial-lined vessels, muscularized arterial walls, capillary beds for exchange and venous return systems [69]. In the vertebrate stem, the appearance of endothelial cells permitted higher intravascular pressures and controlled permeability [70]. Subsequent vertebrate diversification involved progressive cardiac specialization: fish generally exhibit a single-circuit, two-chambered heart; amphibians and most reptiles exhibit a partially divided three-chambered heart; and birds and mammals evolved a fully separated four-chambered heart supporting dual high- and low-pressure circuits [2,76]. Additional innovations include capillary exchange networks and arteriolar smooth muscle regulation, and lymphatic vasculature.
Crucially, the pressure within animal circulatory systems is generated by muscular activity, either within the cells (of cardiac tissue) within a heart or the muscles responsible for moving the main trunk of the body. Muscles operate due to cytoskeletal activity in which motor proteins are driven by thermodynamic gradients (both direct thermal agitation and from ATP or GTP which essentially carry energy derived from thermodynamic gradients); these are ultimately driven by Brownian processes and thus quantum mechanical phenomena.
8. Viridiplantae - Active Regulation of Passive Fluxes
Photosynthetic organisms using chlorophylls a and b (Viridiplantae) include the green algae (Chlorophyta) and the land plants (Embryophyta; with differentiated embryos). Giant coenocytic (i.e. single-celled multinucleate) green algae, such as Caulerpa species, can reach sizes of up to 2.8 metres in length [77] but the entire thallus, including stolons, upright fronds and rhizoids, is a continuous cytoplasmic mass enclosed by one plasma membrane and a cell wall, but lacking any internal dividing cell walls. It uses cytoskeletal transport to induce cytoplasmic streaming, essentially driven by myosin motor proteins acting on actin filaments. In large specimens, streaming ensures physiological integration between photosynthetic and absorptive regions of the alga. In effect, cytoplasmic streaming in Caulerpa serves a role analogous to the vascular transport of plants, maintaining metabolic coordination and enabling rapid redistribution of resources across its coenocytic body, but it occurs within a single general-purpose, multi-role cell, lacking a dedicated translocation cell-type or tissue. As with animals, the macroscopic movement is ultimately generated by quantum mechanical effects operating on the atoms comprising biomolecules (and water and associated thermal bombardment).
The earliest embryophytes colonized the terrestrial surface probably around 515.5 to 469.0 Ma (middle Cambrian–Early Ordovician; [78]). They lacked vascular tissues and were physically limited to the soil/atmosphere interface by a reliance on diffusion. The evolution of simple vascular cell elements in some polysporangiophytes (branching plants) allowed spore-bearing bodies to be held aloft to facilitate spore dispersal [79], leading to the development of photosynthetic stems and true roots, and granting access to water and nutrients at depth within the soil. These plants were characterized by specialized translocation tissues, the xylem and phloem, which are retained in extant vascular plants (Tracheophyta or tracheophytes), which diverged ≈450.8–431.2 Ma [78].
During xylem function, water movement from the soil, through roots and stems to the atmosphere, occurs essentially along gradients of water concentration moderated by surface contact (matric) interactions; expressed as the tendency of water to move (or ‘water potential’; Ψ). Movement is driven largely passively by classical physical forces. However, water potential is subject to biological regulation at key cellular and tissue levels which actively improves function and performance, and is crucial to ‘kickstarting’ the process following stress events or damage. Water diffuses from the soil, passes through the root tissues into the xylem, ascends the stem, enters leaf tissues and finally evaporates into the atmosphere (transpiration). The overall flux proceeds from regions of higher water potential to regions of lower potential, and accordingly is most often described as a passive process. Indeed, the ascent of sap in the xylem is due to the cohesion–tension mechanism, which relies on transpiration to generate negative pressure (tension) on a cohesive column of water in the xylem vessels, pulling water upwards against gravity. Water enters root hair cells and the root cortex by osmosis and matric potential gradients, with negligible requirement for metabolic energy. Once water enters the xylem, its movement upward is largely passive.
However, although the bulk of the water flux is passive, plants do exert metabolic control at two principal regulatory sites: the root and the leaf. Root cells actively transport ions (e.g. K⁺, NO₃⁻) into the vascular tissues, thereby lowering the local water potential and promoting osmotic water uptake. While the water movement itself remains passive, the gradient that induces it is metabolically generated (especially so after periods of drought, when cavitated xylem vessels must be re-filled to re-establish the soil-plant-atmosphere continuum and restart the cohesion-tension mechanism). Moreover, the presence of apoplastic barriers (i.e. obstacles to water movement within the structure of the cell wall) such as the endodermis and corresponding hydrophobic Casparian strip forces water and solutes to traverse membranes, which are key points of control enabling selective uptake of beneficial nutrient ions and exclusion of deleterious ions such as salt or toxic metals. Aquaporins (water-channels) function passively via osmosis, but are regulated actively by phosphorylation, pH, and [Ca²⁺], which are under metabolic control. This in turn allows regulation of root hydraulic conductivity in response to environmental and internal cues. Additionally, at the leaf surface the rate of water loss (and hence the driving force for ascent) is regulated by the active opening and closing of stomata. Guard cells use ATP to pump ions (via H⁺-ATPases, K⁺ channels) and adjust their turgor to determine stomatal aperture: an active process modulating the passive flux of water vapour.
Thus while the driving force sensu stricto for xylem operation is passive, it is actively regulated, and this has a basis in thermodynamics and the ‘quantum mechanical machinery’ of the cell. The plant invests metabolic energy to establish osmotic gradients in the root, to modulate membrane and channel properties, and to regulate stomatal aperture. Water fluxes are embroiled in a regulated framework ensuring homeostasis, preventing uncontrolled influx or efflux and allowing responsiveness to environmental changes (e.g., soil moisture, salinity, vapour-pressure deficits). This duality of passive transport with active regulatory input is critical: purely passive systems would lack selectivity or dynamic adjustment, while purely active pumping of water would be energetically unfeasible given the volumes and heights evident in plants.
Additionally, note that plant height does not have a simple relationship with xylem conduction, and complicating factors can contribute to large size. As an extreme example, the tallest tree, a Sequoia sempervirens (D.Don) Endl. individual known as ‘Hyperion’ growing in Redwood National Park, California, USA, is 116.07 m tall. However, the relationship between height and internal water translocation is complicated by the fact that the upper leaves of this species (particularly on the main axial branches) can absorb water from mist and rain (indeed, they grow in cool humid coastal climates where fog and low cloud readily form), and have a relatively large volume of parenchyma tissue dedicated to water storage. This decreases reliance of photosynthesis and growth (and thus of size) on transport from the roots [80,81]. The canopy of such trees can also accumulate detritus, encouraging soil formation, which is potentially a further source of nutrients independent of the xylem.
Crucially, within root and stomatal cells, the membrane-associated molecules involved in active regulation of xylem sap translocation are exposed to Brownian and thus quantum mechanical-derived motions, and indeed rely on these for their operation. This classical, large-scale and (for the most part) passive translocation system can only function effectively due to molecular systems operating on underlying quantum mechanical effects.
In plant phloem, the translocation of organic solutes (notably sucrose) from sources (photosynthetic organs) to sinks (growth points and storage tissues/organs) involves both active and passive processes. Active processes are primarily associated with the loading and unloading phases of phloem transport. At source tissues, sucrose is actively transported into companion cells or sieve elements against a concentration gradient via membrane-bound sucrose/proton symporters, driven by ATP-dependent proton-pumps [82]. For example, in apoplastic phloem loading the proton-motive force established by H⁺-ATPases energises sucrose uptake into phloem loading cells, resulting in high phloem solute concentrations and concomitant osmotic water influx from accompanying xylem vessels, generating turgor pressure [83,84,85]. During unloading, sink tissue uptake may also involve energy-dependent transport across plasma membranes [86].
Passive processes operate during both loading and long-distance transport phases. In certain species, particularly woody plants, sucrose and other small solutes diffuse symplastically (in the continuous cytoplasm running between cells via plasmodesmata) into the sieve-element/companion cell complex; not requiring direct metabolic energy input at the loading interface [87]. Once a hydrostatic (turgor) pressure gradient between source (high turgor pressure) and sink (low turgor pressure) is established, bulk mass flow of phloem sap occurs along the sieve tubes by pressure-flow (known as the Münch mechanism) [85]. Additionally, unloading into sink tissues may occur via diffusion or bulk-flow through plasmodesmata (“convective unloading”) without exclusive reliance on energy-driven steps [88].
Thus, the active components of phloem functioning (loading/unloading via transporters and proton-pumps) establish the solute and pressure gradients driving passive bulk flows. The passive components (symplastic diffusion, bulk flow under pressure gradients) carry assimilates between source and sink once those gradients exist. Together, these mechanisms enable the regulated and efficient transport of carbon and other solutes essential for plant metabolism and growth.
Neither xylem or phloem is entirely passive (nor entirely active), and vascular translocation in plants involves bulk-flows described by classical physics, either initiated or regulated by active mechanisms involving the Brownian (quantum mechanical-derived) functioning of molecular machinery, particularly enzymatic proton-pumps.
9. Translocation Within Large Heterokonts
Heterokonts (formally, Stramenopiles) are fundamentally different from plants because, aside from being defined by cells possessing two different types of flagella, photosynthetic taxa possess chloroplasts with four bounding membranes, indicating two endosymbiosis events during their evolutionary history (plant chloroplasts have two membranes and also differ in terms of photosynthetic pigments, lacking chlorophyll c and fucoxanthin). The plastid-bearing and thus photosynthetic stramenopiles (the ochrophytes) diverged between 1298 and 622 Ma [89]. Within the brown algae (Phaeophyceae), kelps (order Laminariales; ~31 Ma [90]) evolved complex systems to meet the demands of their extremely large size. They do not possess the true vascular tissues of tracheophytes but do exhibit convergent evolution of specialized vascular-like tissues that facilitate the transport of water, nutrients and photosynthates. These tissues include trumpet-shaped sieve elements, analogous to the phloem in vascular plants, involved in the long-distance transport of organic compounds. Indeed, vascular optimisation in kelp is considered convergent with tracheophytes [91], highlighting that although kelps are only distantly related, they have evolved an equivalent transport system.
In the species Nereocystis luetkeana (K.Mertens) Postels & Ruprecht, 1840 the movement of fluorescent dyes through sieve-tube networks demonstrates source-to-sink bulk flow and even pressure-driven reversal of flow across sieve plates, mechanistically equivalent to phloem transport in tracheophytes (sources are photosynthetically active blades, sinks are growth zones at stipe bases or holdfasts) [91,92]. These sieve elements are embedded in a gelatinous extracellular matrix composed largely of alginate polysaccharides, which isolates them from neighbouring cells and thus provides a biomechanical buffer. Indeed, the internal tissues are organised such that the meristoderm (photosynthetic outer layer) and cortex overlie a central medulla where the sieve-element network resides. Transport velocities in kelp sieve elements have been reported in the range of tens to hundreds of mm h⁻¹ (for example, a mean of 430 mm h-1 determined by following the motion of fluorescent dye markers [91]; this study also discussed estimates of 50 – 780 mm h⁻¹ in the wider literature, determined using methods such as 14C-labelling of photoassimilates).
The sieve tubes form branched networks with junctions and lateral connections between parallel sieve tubes; some appear active while others are not, suggestive of flow regulation within the network. Unlike vascular plants, kelps lack water-conducting vessels equivalent to xylem and companion cells associated with sieve elements; the structural analogue is more basic.
Further research by Knoblauch et al. [92] revealed that the cell walls of sieve tubes in kelps can swell reversibly in response to changes in turgor pressure; this wall swelling may act as a turgor-generation and buffering mechanism, providing a regulatory dimension to flow control. While organic-carbon transport is one major function, kelps must also assimilate nutrients (nitrogen, phosphorus, trace metals) from seawater and distribute these internally to growth zones and can take up multiple forms of nitrogen (nitrate, ammonium, urea) and vary their uptake strategy depending on internal nitrogen status, thus linking external nutrient dynamics to internal transport needs.
Thus, analogous to plant phloem, the translocation system of kelp involves pressure flows operating over scales often measured in meters, but the physical forces generating pressure gradients are essentially due to energy demanding cellular activity involving biomolecules that are ultimately driven by thermodynamics and quantum mechanical effects – again, macroscopic ‘classical’ physics driven by nanoscopic quantum processes.
10. Honourable Mentions
Aside from fungi, slime molds, animals, green plants and heterokonts there are a few other major eukaryotic lineages in which some large organisms and multicellular-like forms have evolved, although these are exceptions rather than the rule within their lineages.
Marine xenophyophores (SAR clade: Rhizaria: Formanifera: Xenophyophorea) include one of the largest single celled organisms (at least in terms of having a diameter measured in the tens of centimetres, even though the volume of the body or ‘test’ is mainly comprised of sediment material): Syringammina fragilissima Brady, 1883. This has a very particular biology, as a fragile benthic analog to a slime mold, and, functionally, can similarly be summarized as coenocytic [93].
Haplozoon protists such as Haplozoon axiothellae Siebert are dinoflagellate (SAR clade: Alveolata: Dinoflagellata: Haplozoonaceae) endoparasites of polychaetes. They give the appearance of being multicellular, to the extent that they were originally misclassified as a ‘simple’ ‘animal’ (literally ‘Haplozoon’). The apparent multicellularity actually represents a single multinucleate cell that develops to create internal septa (membranes) and specialized compartments, resembling a string of several cells with slightly different forms and functions. Interestingly, the protoplasm remains continuous throughout the compartments, and from a functional point of view this has been described as syncytial [94].
Crucially, in the cases mentioned here internal long-distance or compartment-to-compartment transport is managed by the cytoskeleton and cytoplasmic streaming, and so uses the same physical processes as other eukaryotes and is not fundamentally different (although the lifestyles and evolutionary histories of these organisms certainly are).
11. Discussion
The view that emerges here is that living systems scale up by superimposing dedicated transport systems upon an underlying ‘Brownian’ system of molecular motion: from purely Brownian/diffusion, to motor-driven transport on the cytoskeleton, cytoplasmic streaming, long-distance advection (sometimes including peristaltic mechanisms), up to organ- or organism-level bulk-flow systems. Vascular systems in multicellular organisms fundamentally use sub-cellular, quantum-driven processes to either induce or regulate classical pressure gradients, with macroscopic classical processes ultimately emerging from quantum mechanical forces. Thus a key evolutionary stage beyond simple multicellularity was the emergence of bulk-flow transport networks that extended the influence of Brownian and active transport processes into the macroscopic domain, thereby enabling large size and integrated physiology.
While network integration is a universal concept across scales, the specific challenge for size and physiology is to maintain transport rates and coupling across distances. The evolution of bulk-flow networks allowed metabolic units (cells or tissues) to be aggregated into large coherent organisms by providing a high-capacity means of connecting modules. This makes bulk flow a key ‘next step’ following multicellularity.
However, several limitations and open questions remain. First, the detailed mechanisms by which bulk-flow networks evolve remain under-explored. For example, how do vascular systems or circulatory networks emerge from simpler transport systems? What regulatory and developmental changes are required? Second, while systems-biology frameworks have addressed network integration and dynamics, they rarely incorporate explicit transport flows (especially advection) in large-scale models of organismal physiology. There is a need for models that integrate transport networks with regulatory and metabolic networks in evolutionary contexts. Third, size is still constrained: even organisms with bulk flow face limits (e.g., diffusion from vessels into tissues, constraints of pump power, network maintenance costs). Exploring the trade-offs between transport capacity, metabolic cost and organism size remains fertile ground.
Future research directions include: (a) comparative studies across lineages of the evolution of transport networks (e.g., plants vs animals vs colonial algae) to identify common design principles; (b) modelling frameworks in systems biology that explicitly integrate advection and network flows with regulation and metabolism; (c) synthetic biology or bio-engineering of model organisms or tissues to test how transport network topology and capacity influence size and integration; (d) palaeobiological and phylogenetic analyses that explore the timing and constraints of transport-network emergence in major evolutionary transitions.
12. Conclusions
This review collates evidence and ideas from systems biology, biophysics and evolutionary biology to propose that the emergence of bulk-flow transport networks, a key systems-biology transition enabling large organismal size and integrated physiology, are subject to classical physics but nonetheless underpinned by quantum mechanical phenomena. This provides a conceptual link between the physical functioning of matter and macroscopic life processes, and unites disparate evolutionary solutions to large size and physiology with a common fundamental mechanism. It also helps explain the observation that a universe characterized at the smallest scales by quantum effects gives rise to biological organisms that obey classical rules of physics.
Author Contributions
Conceptualization, S.P.; Writing – Original Draft Preparation, S.P.; Writing – Review & Editing, S.P.; Visualization, S.P.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The author thanks G.Wilson Fernandes (Department of General Biology, Federal University of Minas Gerais) for encouraging the development of these ideas via an invitation to talk to the Postgraduate Programme in Ecology, Conservation and Wildlife on 16 November 2016 (Programa de Pós-Graduação em Ecologia, Conservação e Manejo de Vida Silvestre Ecologia Evolutiva e Biodiversidade, Departamento de Biologia Geral, Universidade Federal de Minas Gerais, Belo Horizonte, MG, Brazil).
Conflicts of Interest
The author declares no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ATP | Adenosine-5'-triphosphate |
| CTP | Cytidine-5'-triphosphate |
| DNA | Deoxyribonucleic acid |
| GTP | Guanosine-5'-triphosphate |
| RNA | Ribonucleic acid |
| UTP | Uridine-5'-triphosphate |
References
- Schrödinger, E. What is Life? Cambridge University Press: Cambridge, UK, 1944; p. 194.
- Pross, A. What is Life? How Chemistry becomes Biology. Oxford University Press: Oxford, UK, 2016; p. 0198. [Google Scholar]
- Hoffmann, P.M. Life’s ratchet: how molecular machines extract order from chaos. Basic Books: New York, USA, 2012; p. 0465. [Google Scholar]
- Pierce, S. Life’s Mechanism. Life 2023, 13, 1750. [Google Scholar] [CrossRef]
- Astumian, R.D. The role of thermal activation in motion and force generation by molecular motors. Philos. Trans. Royal Soc. B 2000, 355(1396), 511–522. [Google Scholar] [CrossRef]
- Astumian, R.D.; Hänggi, P. Brownian motors. Phys. Today 2002, 55, 33–39. [Google Scholar] [CrossRef]
- Kodera, N.; Yamamoto, D.; Ishikawa, R.; Ando, T. Video imaging of walking myosin V by high-speed atomic force microscopy. Nature 2010, 468, 72–77. [Google Scholar] [CrossRef]
- Grosberg, R.K.; Strathmann, R.R. The evolution of multicellularity: A minor major transition? Annu. Rev. Ecol. Evol. Syst. 2007, 38, 621–654. [Google Scholar] [CrossRef]
- Czarnoleski, M.; Verberk, W.C.E.P. Cell size matters: A unifying theory across the tree of life. Trends Ecol. Evol. 2025, 40(11), p1113–1125. [Google Scholar] [CrossRef]
- West, G.B.; Brown, J.H.; Enquist, B.J. A general model for the origin of allometric scaling laws in biology. Science 1997, 276, 122–126. [Google Scholar] [CrossRef] [PubMed]
- de Broglie, L. Recherches sur la théorie des quanta. Ann. Phys. [CrossRef]
- Davisson, C.; Germer, L.H. The scattering of electrons by a single crystal of nickel. Phys. Rev. 1927, 30, 705–740. [Google Scholar] [CrossRef]
- Schrödinger, E. Die gegenwärtige Situation in der Quantenmechanik. Naturwissenschaften 1935, 23, 807–812. [Google Scholar] [CrossRef]
- Born, M. Zur Quantenmechanik der Stoßvorgänge. Z. Phys. 1926, 37, 863–867. [Google Scholar] [CrossRef]
- Sciama, D.W. The physical significance of the vacuum state of a quantum field. In The Philosophy of Vacuum. Saunders. S., Brown, H.R., Eds. Oxford University Press: Oxford, UK; 1991. pp. 9780. [Google Scholar]
- Feynman, R.; Leighton, R.; Sands, M. The Feynman lectures on physics. Volume I: mainly mechanics, radiation, and heat. Basic Books: New York, USA, 1963; p. 560. 0465. [Google Scholar]
- Brown, R. A brief account of microscopical observations made in the months of June, July and August 1827, on the particles contained in the pollen of plants; and on the general existence of active molecules in organic and inorganic bodies. The Philosophical Magazine 1828, 4, 161–173. [Google Scholar] [CrossRef]
- Einstein, A. Über die von der molekularkinetischen Theorie der Wärme geforderte Bewegung von in ruhenden Flüssigkeiten suspendierten Teilchen. Annalen der Physik 1905, 322, 549–560. (in German). [Google Scholar] [CrossRef]
- von Smoluchowski, M. Zur kinetischen Theorie der Brownschen Molekularbewegung und der Suspensionen. Annalen der Physik 1906, 326(14), 756–780. (in German). [Google Scholar] [CrossRef]
- Zurek, W.H. Pointer Basis of Quantum Apparatus: Into What Mixture Does the Wave Packet Collapse? Phys. Rev. D 1981, 24, 1516–1525. [Google Scholar] [CrossRef]
- Joos, E.; Zeh, H.D. The Emergence of Classical Properties Through Interaction with the Environment. Z. Phys. B Condens. Matter 1985, 59, 223–243. [Google Scholar] [CrossRef]
- Ghirardi, G.C.; Rimini, A.; Weber, T. Unified Dynamics for Microscopic and Macroscopic Systems. Phys. Rev. D 1986, 34, 470–491. [Google Scholar] [CrossRef]
- Tegmark, M. The importance of quantum decoherence in brain processes. Phys. Rev. E 2000, 61, 4194–4206. [Google Scholar] [CrossRef]
- Hornberger, K.; Gerlich, S.; Haslinger, P.; Nimmrichter, S.; Arndt, M. Colloquium: Quantum interference of clusters and molecules. Rev. Mod. Phys. 2012, 84, 157–173. [Google Scholar] [CrossRef]
- Gerlich, S.; Eibenberger, S.; Tomandl, M.; Nimmrichter, S.; Hornberger, K.; Fagan, P.J.; Tüxen, J.; Mayor, M.; Arndt, M. Quantum interference of large organic molecules. Nat. Commun. 2011, 2, 263. [Google Scholar] [CrossRef]
- Lambert, N.; Chen, Y.N.; Cheng, Y.C.; Li, C.M.; Chen, G.Y.; Nori, F. Quantum biology. Nat. Phys. 2013, 9, 10–18. [Google Scholar] [CrossRef]
- Cao, J.; Cogdell, R.J.; Duan, H.-G.; Hauer, J.; Kleinekathöfer, U.; Jansen, T.L.C.; Mančal, T.; Miller, R.J.D.; Ogilvie, J.P.; Prokhorenko, V.I.; Renger, T.; Tan, H.-S.; Tempelaar, R.; Thorwart, M.; Thyrhaug, E.; Westenhoff, S.; Zigmantas, D. Quantum biology revisited. Sci. Adv. 4888; 6. [Google Scholar] [CrossRef]
- Kim, Y.; Bertagna, F.; D’Souza, E.M.; Heyes, D.J.; Johannissen, L.O.; Nery, E.T.; Pantelias, A.; Sanchez-Pedreño Jimenez, A.; Slocombe, L.; Spencer, M.G.; et al. Quantum Biology: An Update and Perspective. Quantum Rep. 2021, 3, 80–126. [Google Scholar] [CrossRef]
- Arcus, V.L.; Mulholland, A.J. Temperature, Dynamics, and Enzyme-Catalyzed Reaction Rates. Annu. Rev. Biophys. 2020, 49, 163–180. [Google Scholar] [CrossRef]
- Tame, J.R.H. Enzyme dynamics — a brief review. Biophys. Rev. 2023, 15, 317–320. [Google Scholar] [CrossRef]
- Narayanan, C.; Bernard, D.N.; Doucet, N. Role of Conformational Motions in Enzyme Function: Selected Methodologies and Case Studies. Catalysts 2016, 6, 81. [Google Scholar] [CrossRef]
- Zhao, X.; Gentile, K.; Mohajerani, F.; Sen, A. Powering motion with enzymes. Acc. Chem. Res. 2018, 51(10), 2373–2381. [Google Scholar] [CrossRef]
- Slochower, D.R.; Gilson, M.K. Motor-like properties of nonmotor enzymes. Biophys. J. 2018, 114, 2174–2179. [Google Scholar] [CrossRef]
- Bai, L.; Fulbright, R.M.; Wang, M.D. Mechanochemical Kinetics of Transcription Elongation. Phys. Rev. Lett. 2007, 98, 068103. [Google Scholar] [CrossRef] [PubMed]
- Ratje, A.H.; Loerke, J.; Mikolajka, A.; Brünner, M.; Hildebrand, P.W.; Starosta, A.L.; Dönhöfer, A.; Connell, S.R.; Fucini, P.; Mielke, T.; Whitford, P.C.; Onuchic, J.N.; Yu, Y.; Sanbonmatsu, K.Y.; Hartmann, R.K.; Penczek, P.A.; Wilson, P.N.; Spahn, C.M.T. Head swivel on the ribosome facilitates translocation by means of intra-subunit tRNA hybrid sites. Nature 2010, 468, 13–718. [Google Scholar] [CrossRef] [PubMed]
- Zaccai, G.; Natali, F.; Peters, J.; Řihová, M.; Zimmerman, E.; Ollivier, J.; Combet, J.; Maurel, M.-C.; Bashan, A.; Yonath, A. The fluctuating ribosome: thermal molecular dynamics and function. Sci. Rep. 2016, 6, 37138. [Google Scholar] [CrossRef]
- Leighton, M.P. Flow of Energy and Information in Molecular Machines. Annu. Rev. Phys. Chem. 2025, 76, 379–403. [Google Scholar] [CrossRef]
- Caban, K.; Gonzalez, R.L. The Emerging Role of Rectified Thermal Fluctuations in Initiator aa-tRNA- and Start Codon Selection during Translation Initiation. Biochimie 2015, 114, 30–38. [Google Scholar] [CrossRef]
- Spirin, A.S.; Finklestein, A.V. The ribosome as a Brownian ratchet machine. In Molecular Machines in Biology. Frank, J., Ed.; Cambridge University Press: Cambridge, UK, 2011; pp. 158–190. [Google Scholar] [CrossRef]
- Marcucci, L.; Yanagida, T. From single molecule fluctuations to muscle contraction: a Brownian model of A.F. Huxley's hypotheses. PLoS ONE, 4004. [Google Scholar] [CrossRef]
- Guo, H.; Rubinstein, J.L. Structure of ATP synthase under strain during catalysis. Nat. Commun. 2022, 13, 2232. [Google Scholar] [CrossRef]
- Nakanishi-Matsui, M.; Kashiwagi, S.; Kojima, M.; Nonaka, T.; Futai, M. Roles of the β-subunit hinge domain in ATP synthase F₁ sector: hydrophobic network formed by introduced βPhe174 inhibits subunit rotation. Biochem. Biophys. Res. Commun. 2010, 395, 173–7. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Ikeguchi, M.; Kidera, A. Mechanism of the αβ Conformational Change in F1-ATPase after ATP Hydrolysis: Free-Energy Simulations. Biophys. J. 2015, 108, 85–97. [Google Scholar] [CrossRef]
- Uchihashi, T.; Iino, R.; Ando, T.; Noji, H. High-speed atomic force microscopy reveals rotary catalysis of rotorless F1-ATPase. Science, /: https, 1126. [Google Scholar]
- Berg, H.C. Random Walks in Biology; Princeton University Press: Princeton, NJ, USA, 1993; ISBN 9780691000640. [Google Scholar]
- Vogel, S. Dealing honestly with diffusion. Am. Sci. /: 246–249. https, 4449. [Google Scholar]
- Pittermann, J. The evolution of water transport in plants: an integrated approach. Geobiology 2010, 8, 112–139. [Google Scholar] [CrossRef]
- Fick, A. Über Diffusion. Annalen der Physik und Chemie 1855, 170(1), 59–86. (in German). [Google Scholar] [CrossRef]
- Goldstein, R.E.; van de Meent, J. A physical perspective on cytoplasmic streaming. Interface Focus 2015, 5, 20150030. [Google Scholar] [CrossRef]
- Ando, D. , Korabel N., Huang K.C., Gopinathan A. Cytoskeletal network morphology regulates intracellular transport dynamics. Biophys. J. 2015, 109, 1574–1582. [Google Scholar] [CrossRef] [PubMed]
- Lu, W. , Gelfand V.I. Go with the flow – bulk transport by molecular motors. J. Cell Sci. 2603. [Google Scholar] [CrossRef]
- Szánthó, L.L.; Merényi, Z.; Donoghue, P.; Gabaldón, T.; Nagy, L.G.; Szöllősi, G.J.; Ocaña-Pallarès, E. A timetree of Fungi dated with fossils and horizontal gene transfers. Nat. Ecol. Evol. 2025, 9, 1989–2001. [Google Scholar] [CrossRef]
- Lew, R.R. Mass flow and pressure-driven hyphal extension in Neurospora crassa. Microbiol. 2005, 151, 2685–2692. [Google Scholar] [CrossRef]
- Bago, B.; Pfeffer, P.E.; Shachar-Hill, Y. Carbon Metabolism and Transport in Arbuscular Mycorrhizas. Plant Physiol. 2002, 128, 615–624. [Google Scholar] [CrossRef]
- Lew, R.R. How Does a Hypha Grow? The Biophysics of Pressurized Growth in Fungi. Nat. Rev. Microbiol. 2011, 9, 509–518. [Google Scholar] [CrossRef]
- Money, N.P. Wishful Thinking of Turgor Revisited: The Mechanics of Fungal Growth. Fungal Genet. Biol. 1997, 21, 173–187. [Google Scholar] [CrossRef]
- Peñalva, M.A.; Tilburn, J.; Bignell, E.; Arst, H.N. Ambient pH Gene Regulation in Aspergillus nidulans: Molecular Control of Fungal Physiology. Microbiol. Mol. Biol. Rev. 1998, 62, 426–446. [Google Scholar] [CrossRef]
- Pérez-Bermúdez, P.; García-Carmona, M.; Gil, M.; Requena, M.E.; González-Mula, A.; Pardo, E.; Martínez-Expósito, F.; Nicolás, M.; Ponce-Noy, M.; López-Pérez, J.L. Proton Gradients and Nutrient Transport in Filamentous Fungi. J. Fungi 2020, 6, 185. [Google Scholar] [CrossRef]
- Steinberg, G. Hyphal Growth: A Tale of Motors, Lipids, and the Spitzenkörper. Eukaryot. Cell 2007, 6, 351–360. [Google Scholar] [CrossRef]
- Riquelme, M. Tip Growth in Filamentous Fungi: A Road Trip to the Apex. Annu. Rev. Microbiol. 2013, 67, 587–609. [Google Scholar] [CrossRef]
- Fiz-Palacios, O.; Romeralo, M.; Ahmadzadeh, A.; Weststrand, S.; Ahlberg, P.E.; Baldauf, S.L. Did terrestrial diversification of amoebas (Amoebozoa) occur in synchrony with land plants? PLoS ONE 2013, 8, e74374. [Google Scholar] [CrossRef]
- Alim, K.; Amselem, G.; Peaudecerf, F.; Brenner, M.P.; Pringle, A. Random Network Peristalsis in Physarum polycephalum Organizes Fluid Flows across an Individual. Proc. Natl. Acad. Sci. USA 2013, 110, 13306–13311. [Google Scholar] [CrossRef]
- Alim, K.; Andrew, N.; Pringle, A.; Brenner, M.P. Mechanism of Signal Propagation in Physarum polycephalum. Proc. Natl. Acad. Sci. USA 2017, 114, 5136–5141. [Google Scholar] [CrossRef]
- Smith, D.A.; Saldana, R. Model of the Ca²⁺ Oscillator for Shuttle Streaming in Physarum polycephalum. Biophys. J. 1992, 61, 368–380. [Google Scholar] [CrossRef]
- Iima, M.; Nakagaki, T. Peristaltic Transport and Mixing of Cytosol through the Whole Body of Physarum Plasmodium. Math. Med. Biol. 2012, 29, 263–281. [Google Scholar] [CrossRef]
- Goldstein, R.E.; van de Meent, J.-W. A Physical Perspective on Cytoplasmic Streaming. Interface Focus 2015, 5, 20150030. [Google Scholar] [CrossRef]
- Carlisle, E.M.; Yin, Z.; Pisani, D.; Donoghue, P.C.J. Ediacaran origin and Ediacaran-Cambrian diversification of Metazoa. Sci. Adv. 2024, 10, eadp7161. [Google Scholar] [CrossRef]
- Schmidt-Rhaesa, A. The Evolution of Organ Systems. Oxford University Press, Oxford, UK, 2007. [CrossRef]
- Ribatti, D. A Darwinian approach to the development of the vascular system in the vertebrates. Clin. Exp. Med. [CrossRef]
- Monahan-Earley R, Dvorak AM, Aird WC. Evolutionary origins of the blood vascular system and endothelium. J. Thromb. Haemost. 2013, 11 (Suppl 1), 46–66. [Google Scholar] [CrossRef]
- Bussolino, F. , Mantovani, A., & Persico, G. Molecular mechanisms of blood vessel formation. Trends in Biochemical Sciences 1997, 22(7), 251–256. [Google Scholar] [CrossRef]
- Hillyer, J.F. & Pass, G. The circulatory and immune systems of insects. Annu. Rev. Entomol. 2020, 65, 121–143. [Google Scholar] [CrossRef]
- Nakao, T. An electron microscopic study of the circulatory system in Nereis japonica. J. Morphol. /: https, 3031. [Google Scholar]
- Jahn H, Hammel JU, Göpel T, Wirkner CS, Mayer G. A multiscale approach reveals elaborate circulatory system and intermittent heartbeat in velvet worms (Onychophora). Commun. Biol. /: 468. https, 3711.
- Song, S. , Starunov, V., Bailly, X. et al. Globins in the marine annelid Platynereis dumerilii shed new light on hemoglobin evolution in bilaterians. BMC Evol. Biol. [CrossRef]
- Panara, V.; Varaliová, Z.; Wilting, J.; Koltowska, K.; Jeltsch, M. The relationship between the secondary vascular system and the lymphatic vascular system in fish. Biol. Rev. 2024, 99, 2108–2133. [Google Scholar] [CrossRef] [PubMed]
- Meinesz, A. (1999). Killer Algae: The True Tale of a Biological Invasion, U: Trans.). Chicago, 5192. [Google Scholar]
- Morris, J.L.; Puttick, M.N.; Clark, J.W.; Edwards, D.; Kenrick, P.; Pressel, S.; Wellman, C.H.; Yang, Z.; Schneider, H.; Donoghue, P.C.J. The timescale of early land plant evolution. Proc. Natl. Acad. Sci. USA 2018, 115, E2274–E2283. [Google Scholar] [CrossRef] [PubMed]
- Pierce, S.; Vianelli, A.; Cerabolini, B. From ancient genes to modern communities: the cellular stress response and the evolution of plant strategies. Funct. Ecol. 2005, 19, 763–776. [Google Scholar] [CrossRef]
- Chin, A. R. O.; Guzmán-Delgado, P.; Sillett, S.C.; Orozco, J.; Kramer, R.D.; Kerhoulas, L.P.; Moore, Z.J.; Reed, M.; Zwieniecki, M.A. Shoot dimorphism enables Sequoia sempervirens to separate requirements for foliar water uptake and photosynthesis. Am. J. Bot. /: https, 1002. [Google Scholar]
- Ishii, H.R.; Azuma, W.; Kuroda, K.; Sillett, S.C. Pushing the limits to tree height: could foliar water storage compensate for hydraulic constraints in Sequoia sempervirens? Funct. Ecol. /: https, 1093. [Google Scholar]
- Patrick, J.W.; Zhang, W.; Tyerman, S.D.; Offler, C.E.; Walker, N.A. Role of membrane transport in phloem translocation of assimilates and water. Aust. J. Plant Physiol. 2001, 28(7), 697–709. [Google Scholar] [CrossRef]
- Turgeon, R.; Oparka, K. Phloem Transport: Cellular Pathways and Molecular Trafficking. Annu. Rev. Plant Biol. 2009, 60, 207–221. [Google Scholar] [CrossRef]
- Turgeon, R. The role of phloem loading reconsidered. Plant Physiol. 2010, 152(4), 1817–1823. [Google Scholar] [CrossRef]
- Braun, D. M. Phloem Loading and Unloading of Sucrose: What a Long, Strange Trip from Source to Sink. Annu. Rev. Plant Biol. 2022, 73, 553–584. [Google Scholar] [CrossRef]
- Comtet, J.; Jensen, K.H.; Stroock, A.D.; Turgeon, R. Phloem loading through plasmodesmata: a biophysical analysis. Plant Physiol. 2017, 175(2), 904–915. [Google Scholar] [CrossRef]
- De Schepper, V.; De Swaef, T.; Bauweraerts, I.; Steppe, K. Phloem transport: a review of mechanisms and controls. J. Exp. Bot. 2013, 64(16), 4839–4850. [Google Scholar] [CrossRef]
- Ross-Elliott, T.J.; Jensen, K.H.; Haaning, K.S.; Wager, B.M.; Knoblauch, J.; Howell, A.G.; Mullendore, D.L.; Monteith, A.G.; Paultre, D.; Yan, D.; Otero-Perez, S.; Bourdon, M.; Sager, R. ; Lee, J-Y.; Helariutta, Y.; Knoblauch, M.; Oparka, K. Phloem unloading in Arabidopsis roots is convective and regulated by the phloem-pole pericycle. eLife. [CrossRef]
- Strassert, J.F.H.; Irisarri, I.; Williams, T.A.; Burki, F. A molecular timescale for eukaryote evolution with implications for the origin of red algal-derived plastids. Nat. Commun. 2021, 12, 1879. [Google Scholar] [CrossRef] [PubMed]
- Starko, S.; Martone, P.T.; Donoghue, P.C.J. A comprehensive kelp phylogeny sheds light on the radiation of kelps (Order Laminariales, Phaeophyceae). Mol. Phylogenet. Evol. 2019, 136, 138–150. [Google Scholar] [CrossRef]
- Knoblauch, J.; Peters, W.S.; Knoblauch, M. The gelatinous extracellular matrix facilitates transport studies in kelp: visualization of pressure-induced flow reversal across sieve plates. Ann. Bot. [CrossRef]
- Knoblauch, J.; Tepler Drobnitch, S.; Peters, W.S.; Knoblauch, M. In situ microscopy reveals reversible cell wall swelling in kelp sieve tubes: one mechanism for turgor generation and flow control? Plant Cell Environ. 1727. [Google Scholar] [CrossRef]
- Pawlowski, J.; Holzmann, M.; Fahrni, J.; Richardson, S.L. Small subunit ribosomal DNA suggests that the xenophyophorean Syringammina corbicula is a foraminiferan. J. Eukaryot. Microbiol. 2003, 50, 483–487. [Google Scholar] [CrossRef] [PubMed]
- Leander, B.S.; Saldarriaga, J.F.; Keeling, P.J. Surface morphology of the marine parasite Haplozoon axiothellae Siebert (Dinoflagellata). Eur. J. Protistol. 2002, 38, 287–297. [Google Scholar] [CrossRef]
Figure 1.
The motion and transport of material within eukaryotes at the macroscopic scale depends on nanoscopic thermodynamic processes, ultimately driven by quantum mechanical phenomena, that induce or actively regulate bulk pressure flows obeying the rules of classical physics. Disparate adaptations, cell types and tissue systems in disparate taxa share the same quantum mechanical underpinnings, with molecular motions and conformation state changes translating quantum motion into classical motion. Images are not AI generated and are either public domain or published under a Creative Commons Attribution-Share Alike 3.0 or 4.0 license at commons.wikimedia.org (accessed on 4 November 2025) and compiled using CorelDraw 9 (Corel Corporation). Individual image credits (from top to bottom of page): backlit Ficus elastica leaf (Julian Herzog); Nereocystis luetkeana (Chris Teague); human organ systems (Loneshieling); slime mold (Nativeplants garden); Nematoda (Vyzhdova V); external streaming of Volvox aureus (Matthew Herron); internal streaming of Elodea canadensis cells (Daemon Canchig); molecular model of the sarcomere in the A band (Dirk Gently PI); kinesin on a cytoskeletal microtubule (Tomoki Fukushima); membrane proteins (Eunice Laurent); ATPsynthase (Bone117).
Figure 1.
The motion and transport of material within eukaryotes at the macroscopic scale depends on nanoscopic thermodynamic processes, ultimately driven by quantum mechanical phenomena, that induce or actively regulate bulk pressure flows obeying the rules of classical physics. Disparate adaptations, cell types and tissue systems in disparate taxa share the same quantum mechanical underpinnings, with molecular motions and conformation state changes translating quantum motion into classical motion. Images are not AI generated and are either public domain or published under a Creative Commons Attribution-Share Alike 3.0 or 4.0 license at commons.wikimedia.org (accessed on 4 November 2025) and compiled using CorelDraw 9 (Corel Corporation). Individual image credits (from top to bottom of page): backlit Ficus elastica leaf (Julian Herzog); Nereocystis luetkeana (Chris Teague); human organ systems (Loneshieling); slime mold (Nativeplants garden); Nematoda (Vyzhdova V); external streaming of Volvox aureus (Matthew Herron); internal streaming of Elodea canadensis cells (Daemon Canchig); molecular model of the sarcomere in the A band (Dirk Gently PI); kinesin on a cytoskeletal microtubule (Tomoki Fukushima); membrane proteins (Eunice Laurent); ATPsynthase (Bone117).
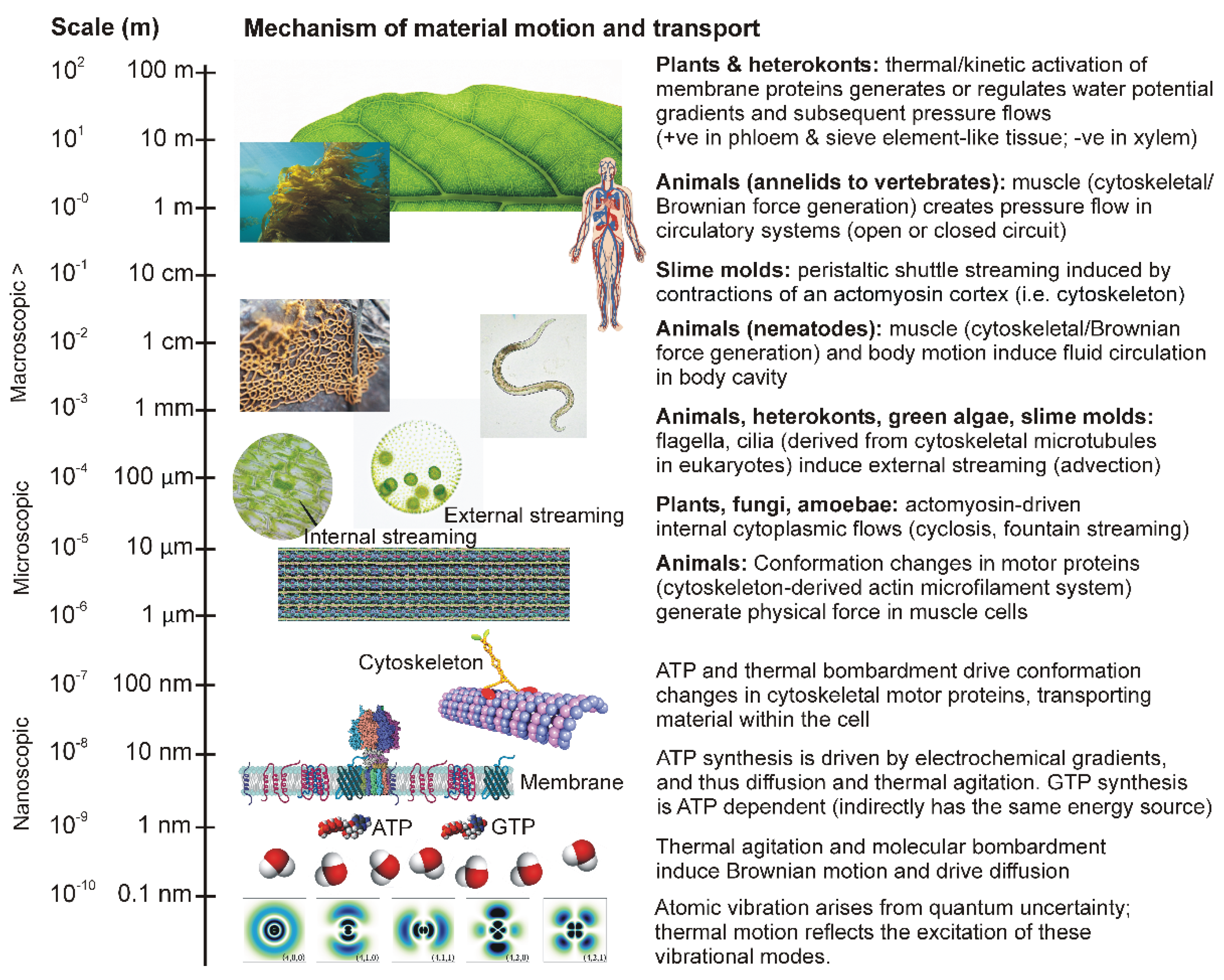

Table 1.
The physical scale at which internal transport shifts from quantum mechanical-determined (thermodynamic gradients) to classical (pressure flow) systems in Metazoa (note that this is not a simple, direct evolutionary transition via anagenesis; circulatory systems have evolved independently via convergent evolution in different metazoan taxa).
Table 1.
The physical scale at which internal transport shifts from quantum mechanical-determined (thermodynamic gradients) to classical (pressure flow) systems in Metazoa (note that this is not a simple, direct evolutionary transition via anagenesis; circulatory systems have evolved independently via convergent evolution in different metazoan taxa).
|
Physical scale (mm) |
Example organisms | Body cavity type | Traits | Circulatory system |
| 0.1 - 10 | Flatworms (Planaria; Platyhelminthes) |
Acoelomate | No internal cavity (solid tissue) |
Diffusion only |
| 0.5 - 500 | Nematodes (Nematoda), Rotifers | Pseudo-coelomate | Cavity not fully lined with mesoderm | Circulation by body movement (pseudocoelomic fluid) |
| 1 > | Earthworms (Annelida), Mollusks, Arthropods, Vertebrates | Coelomate | Cavity fully lined with mesoderm | True circulatory system (open or closed) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



