
Submitted:
06 August 2025
Posted:
07 August 2025
You are already at the latest version
Abstract
The origin and stability of life remain among the most profound scientific enigmas. While Darwinian evolution explains adaptation via random mutations and natural selection, the sheer improbability of abiogenesis and the emergence of complex biological structures suggests that additional principles may be at work. This article proposes a hypothesis of entropy-guided evolution, in which quantum phenomena—especially non-locality, coherence, and tunneling in large ensembles of particles—play a macroscopic role in biological systems. These mechanisms may allow life to traverse otherwise insurmountable barriers in the evolutionary fitness landscape and maintain its remarkable stability across geological timescales. Within this framework, genes act not as complete blueprints but as developmental parameters, interacting with emergent quantum archetypes that encode organizing principles beyond local molecular dynamics. Analogies from physics and engineering illustrate how a purely reductionist view may overlook hidden informational and structural layers. Though speculative, the hypothesis makes testable predictions regarding quantum coherence, mutation patterns, and evolutionary transitions. This work aims to inspire interdisciplinary research bridging quantum physics, biology, and complexity theory, potentially revealing deeper organizing laws behind life’s emergence and resilience.
Keywords:
emergent phenomena
; complex systems
; quantum physics
; origin of life
; evolution
; quantum non-locality
; fitness landscape
; stability of life
; interdisciplinary approach
; philosophy of science
1. Introduction
The existence of life presents a fundamental paradox: extraordinary complexity arising from processes with seemingly vanishingly small probabilities [1]. Standard evolutionary theory, while robust, faces mathematical challenges when explaining several key phenomena [2,3]:
- Abiogenesis probability crisis: The spontaneous emergence of self-replicating systems from inanimate matter appears mathematically implausible given current models [2]
- Punctuated equilibrium: Rapid evolutionary “explosions” like the Cambrian diversification suggest mechanisms beyond gradual mutation and selection [4]
- Non-random mutagenesis: Experimental evidence suggests environmental influence on mutation patterns, challenging purely random models [5]
2. Mathematical Framework: Quantum-Enhanced Evolution
2.1. Quantum Fitness Landscape
We propose that evolutionary dynamics may be governed by a quantum-classical hybrid model [5,13] where the probability of reaching a genetic configuration x is:
where:
P(x) ∝ |ψ(x)|² · F(x)
- ψ(x) is the quantum amplitude associated with genetic configuration x,
- |ψ(x)|² is the probability density in Hilbert space,
- F(x) is the classical fitness function over the configuration space.
This formulation merges quantum probability (superposition of states) with Darwinian selection (fitness weighting), enabling parallel exploration of high-fitness basins in a multidimensional landscape. The model implies that quantum evolution enhances convergence speed toward stable or functional configurations, potentially counteracting entropy-driven drift. ψ(x) represents the quantum amplitude for configuration x, and F(x) is its classical fitness [14]. Such a formulation suggests that quantum superposition could enable parallel exploration of genetic space, potentially explaining the apparent efficiency of evolutionary search processes [15].
2.2. System Stability Dynamics
The temporal evolution of system stability S can be expressed as [4,16]:
where:
dS/dt = -∇mut V(x) + Q_tunnel + N_non-local
- S is the system’s macro-stability,
- V(x) is the effective fitness potential landscape,
- ∇mut is the mutational gradient—representing how mutations move the system along or against the fitness landscape,
- Q_tunnel models the rate of quantum tunneling events, which allow systems to bypass classical barriers [5],
- N_non-local represents entanglement-driven or field-based coherence effects, potentially enabling synchronized adaptation across system components [17].
While only conceptual and illustrative at this stage, this equation formalizes how quantum contributions may modify the entropy flow, injecting stability and organization into otherwise noisy, unstable systems. Such a framework suggests that biological systems might exploit quantum coherence to maintain stability against entropic degradation while enabling directed evolutionary exploration [18,19].
In classical thermodynamics, all complex systems are expected to degrade over time due to entropy increase, especially in warm, noisy biological environments. However, living systems exhibit a persistent ability to maintain order and function, resisting thermal noise and stochastic degradation.
Quantum coherence and tunneling may enable life to momentarily ‘shortcut’ entropic trajectories by accessing low-probability, high-fitness configurations that are otherwise unreachable by classical means. These mechanisms act as locally entropy-defying processes, not violating the second law globally, but channeling information and energy through highly selective pathways.
This behavior resembles Maxwell’s demon-like selection, where quantum correlations could play the role of an internal information-processing agent, distinguishing and preserving useful configurations. In this view, biological evolution becomes a thermodynamically open system with quantum-informational feedback loops that locally reduce effective entropy over biologically relevant timescales.
3. The Newton’s Radio Analogy: Limits of Reductionism
Consider Isaac Newton examining a transistor radio in 1700 [1]. Despite his genius, he would conclude that sound originates locally within the components, missing the non-local electromagnetic transmission. Similarly, contemporary biology may overlook non-local organizing principles in living systems [18,19].
Figure 1.
Thought Experiment: Isaac Newton Examining a Transistor Radio in 1700 Despite his extraordinary intellect, Newton—lacking the conceptual framework of electromagnetism—would likely infer that sound emerges locally from the device’s components. He would miss the non-local transmission of electromagnetic signals, illustrating how paradigm limitations constrain interpretation of complex systems.
Figure 1.
Thought Experiment: Isaac Newton Examining a Transistor Radio in 1700 Despite his extraordinary intellect, Newton—lacking the conceptual framework of electromagnetism—would likely infer that sound emerges locally from the device’s components. He would miss the non-local transmission of electromagnetic signals, illustrating how paradigm limitations constrain interpretation of complex systems.

This analogy illustrates how:
4. Quantum vs Classical Evolution Models
4.1. Classical Model Limitations
4.2. Quantum-Enhanced Model
Figure 2.
Hypothetical Quantum Optimization in Evolution: Overcoming Classical Model Limitations. The classical model of evolution is constrained by local fitness maxima, where random mutations enable only gradual optimization. Probability barriers may remain insurmountable within geological timescales. In contrast, the quantum-enhanced model leverages quantum tunneling to traverse fitness barriers, coherent superposition to explore multiple genetic configurations in parallel, and non-local correlations to enable coordinated multi-level changes.
Figure 2.
Hypothetical Quantum Optimization in Evolution: Overcoming Classical Model Limitations. The classical model of evolution is constrained by local fitness maxima, where random mutations enable only gradual optimization. Probability barriers may remain insurmountable within geological timescales. In contrast, the quantum-enhanced model leverages quantum tunneling to traverse fitness barriers, coherent superposition to explore multiple genetic configurations in parallel, and non-local correlations to enable coordinated multi-level changes.
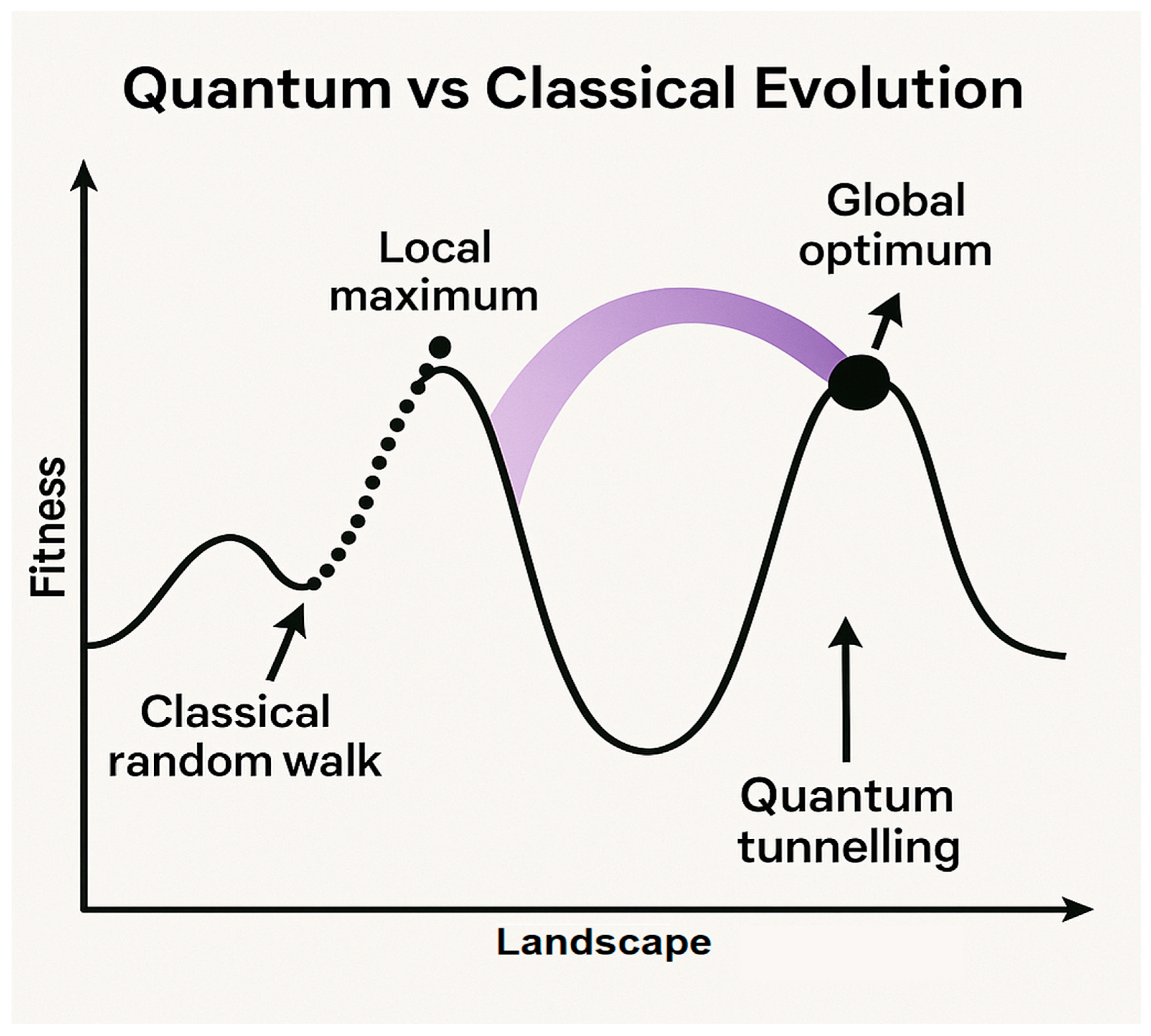
Recent computational studies suggest that quantum search algorithms can significantly outperform classical random walks in complex fitness landscapes [5], potentially explaining the efficiency of evolutionary processes.
5. Archetypal Structures: Universal Design Principles
Both natural organisms (mouse) and artificial systems (Mars rover) exhibit convergent functional architectures [3,22].
Figure 3.
Illustrative comparison between a biological organism (mouse) and an artificial system (Mars rover), highlighting convergent functional architecture. Despite vast differences in origin and material composition, both systems exhibit analogous components for sensing, processing, locomotion, and energy management—such as eyes vs. cameras, brain vs. computer, legs vs. wheels, and digestion vs. solar panels. The image emphasizes shared structural logic across natural and engineered domains.
Figure 3.
Illustrative comparison between a biological organism (mouse) and an artificial system (Mars rover), highlighting convergent functional architecture. Despite vast differences in origin and material composition, both systems exhibit analogous components for sensing, processing, locomotion, and energy management—such as eyes vs. cameras, brain vs. computer, legs vs. wheels, and digestion vs. solar panels. The image emphasizes shared structural logic across natural and engineered domains.
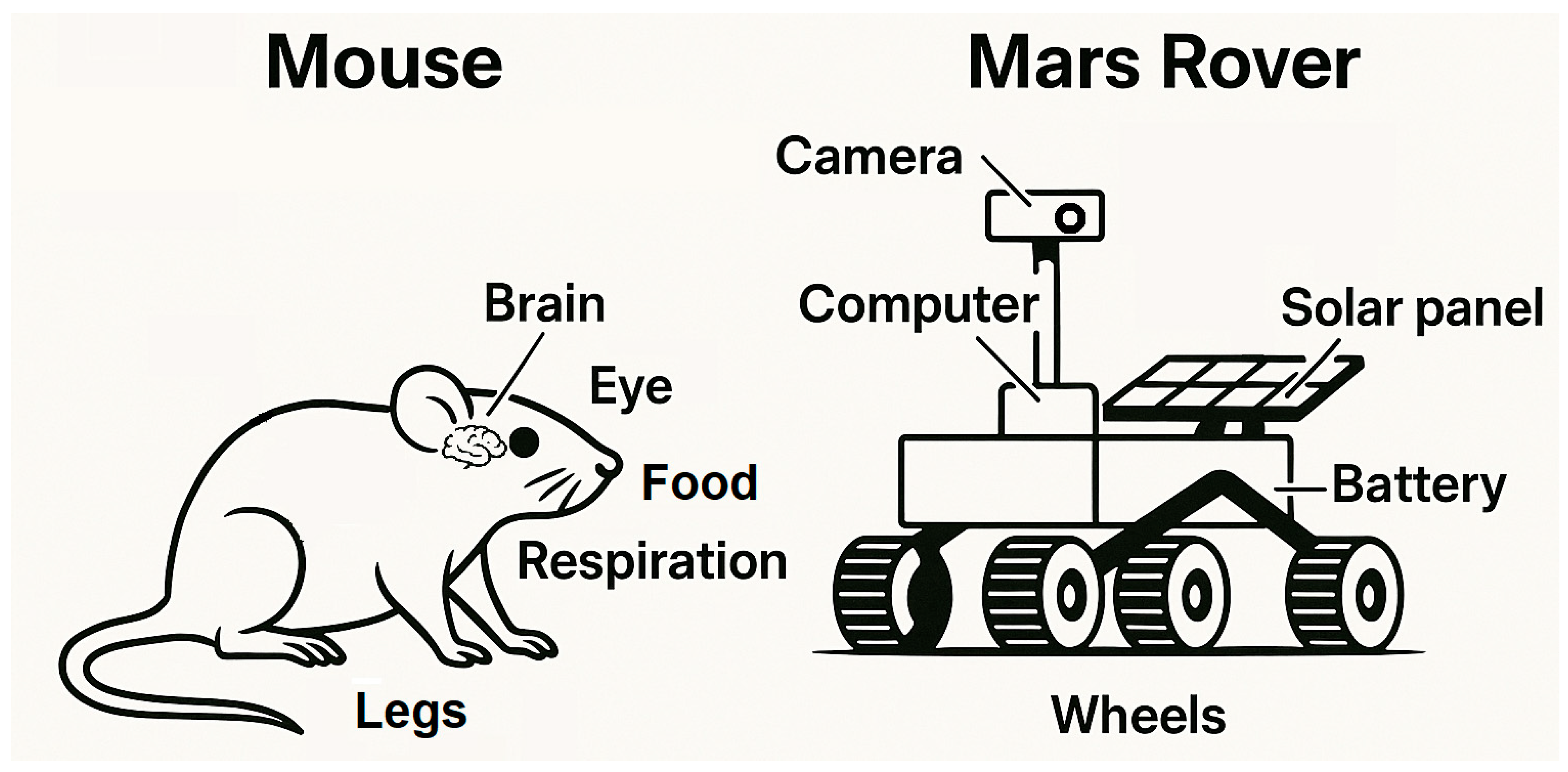

Table 1.
Functional parallels between biological organisms and artificial systems. Each row highlights a core function and its respective implementation in natural and engineered contexts, emphasizing convergent design principles across domains.
Table 1.
Functional parallels between biological organisms and artificial systems. Each row highlights a core function and its respective implementation in natural and engineered contexts, emphasizing convergent design principles across domains.
| Function | Biological | Artificial |
|---|---|---|
| Sensing | Eyes | Camera |
| Processing | Brain | Computer |
| Locomotion | Legs | Wheels |
| Energy | Digestion | Solar panels |
This convergence suggests a “space of possibilities” - optimal forms that transcend specific material implementations [22]. Such archetypal structures imply fundamental organizational principles that may be encoded in the physical laws themselves, potentially accessible through quantum coherence mechanisms [15,19].
6. Quantum Phenomena in Biology: Current Evidence and Predictions
Table 2.
This table summarizes key quantum phenomena and their hypothesized roles in biological systems. While some, like coherence in photosynthesis, are well-supported, others—such as non-locality and entanglement—remain largely theoretical. Suggested testing methods highlight paths for future exploration at the intersection of quantum physics and biology.
Table 2.
This table summarizes key quantum phenomena and their hypothesized roles in biological systems. While some, like coherence in photosynthesis, are well-supported, others—such as non-locality and entanglement—remain largely theoretical. Suggested testing methods highlight paths for future exploration at the intersection of quantum physics and biology.
| Physical Phenomenon | Potential Biological Role | Empirical Support | Testing Possibilities |
|---|---|---|---|
| Quantum tunneling | Overcoming fitness barriers | Enzymatic processes [9] | Quantum simulations, femtosecond spectroscopy [23] |
| Coherence | Efficient energy transfer | Photosynthesis [6,7], FMO complex | Optical measurements, interferometry [24] |
| Non-locality | Hidden information channels | Hypothetical | Bell tests at molecular scale [17] |
| Entanglement | Coordinated mutations | Avian magnetoreception [8] | Correlation studies in genetic networks [25] |
7. Two-Component System Hypothesis
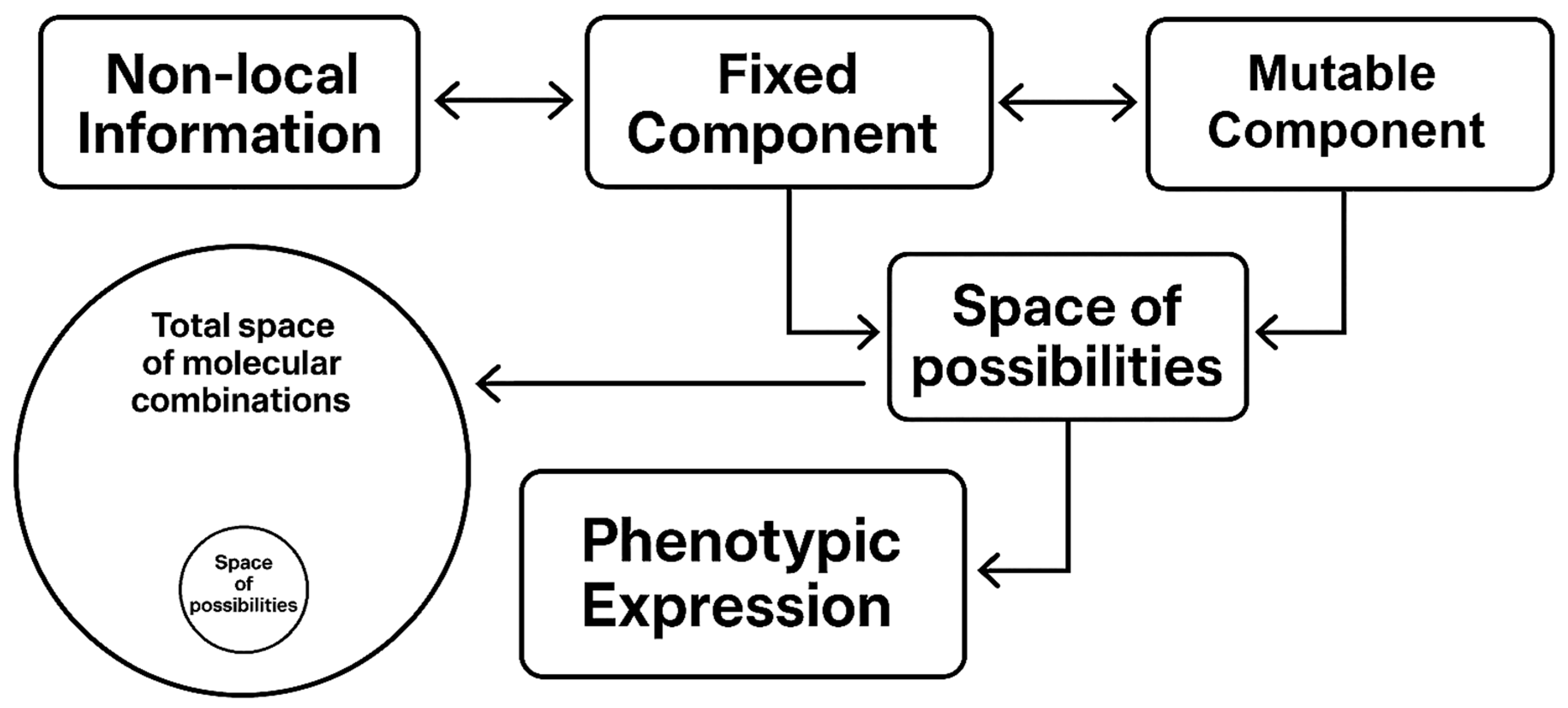
7.1. System Architecture
-
Fixed component: Evolutionarily stable structures (cell walls, ribosomes)
-
Mutable component: Evolutionary parameters (genes)
This division reflects the hierarchical organization observed in complex systems, where different levels operate on distinct timescales and follow different dynamical rules [4,18].
Figure 4.
A symbolic representation of a system containing both fixed and mutable components. Phenotypic expression is influenced by both types of components, while the fixed component is hypothetically shaped by non-local information. The space of possibilities is significantly smaller than the space of all possible molecular combinations, favoring functional structures within the system.
Figure 4.
A symbolic representation of a system containing both fixed and mutable components. Phenotypic expression is influenced by both types of components, while the fixed component is hypothetically shaped by non-local information. The space of possibilities is significantly smaller than the space of all possible molecular combinations, favoring functional structures within the system.
8. Testable Predictions and Experimental Approaches
8.1. Quantum Coherence in DNA Replication
- Prediction: Quantum coherence during critical replication steps
- Test: Femtosecond spectroscopy of DNA polymerase activity [23]
- Expected outcome: Evidence of coherent superposition in base selection
8.2. Non-Random Mutation Patterns
- Prediction: Spatially or temporally correlated mutations beyond classical expectations
- Test: Statistical analysis of mutation patterns in isolated vs. connected populations
- Expected outcome: Enhanced correlation in connected systems
8.3. Fitness Landscape Tunneling
- Prediction: Evolutionary transitions that bypass expected intermediate forms
- Test: Computational modeling with quantum vs. classical search algorithms [5]
- Expected outcome: Quantum models better match observed evolutionary jumps
9. Implications and Future Directions
9.1. For Evolutionary Biology
These ideas invite a shift from viewing evolution as a purely stochastic walk on a rugged landscape to a guided, information-driven process, where quantum effects reduce the effective entropy of search. Genes may be better understood not as rigid blueprints, but as compressed descriptors shaped by and interacting with deeper organizing fields or structures.
The observed convergence in biological and artificial systems (see Section 5) suggests a hidden attractor structure in the fitness landscape, where evolution gravitates toward low-entropy, functionally optimal configurations. This points toward self-organizing principles driven not just by thermodynamics but also by quantum-informational symmetry-breaking mechanisms.
- Reframe genes as developmental parameters rather than complete blueprints
- Investigate quantum effects in mutation and selection processes
- Develop new mathematical frameworks combining quantum mechanics with population genetics [27]
9.2. For Quantum Biology
Understanding how living systems maintain quantum coherence and resist decoherence is not just a biochemical question, but one tied to informational entropy management. Organisms may actively shape internal conditions—via compartmentalization, coherent vibrational modes, or network redundancies—to suppress entropy increase and preserve quantum states longer than expected.
Future work should explore whether mutations, regulatory decisions, or developmental pathways correlate with information-theoretic entropy gradients, suggesting that life harnesses entropy not just as a constraint, but as a guide toward complexity and resilience.
- Study decoherence resistance mechanisms in biological environments [16]
9.3. For Technology
10. Conclusions
The hypothesis presented here suggests that life’s remarkable stability and complexity may arise from quantum phenomena operating at biological scales [11,12]. While speculative, this framework:
The concept of entropy-guided evolution integrates thermodynamic constraints with non-local quantum processes, proposing a deeper level of organization underlying biological dynamics. In this view, emergent quantum archetypes may serve as attractors in the fitness landscape—patterns that constrain evolution toward robust and functionally optimal configurations.
Such a model invites us to rethink not only the mechanisms of evolution, but also the nature of biological information itself—as something not solely encoded in DNA, but distributed across hierarchical and possibly non-local layers of organization. This shift has implications for how we understand development, heredity, adaptation, and even the origin of consciousness.
The “message of life” may indeed point toward fundamental physical principles operating beyond our current understanding [29]. Future research bridging quantum physics, biology, complexity theory, and information science may reveal the deeper organizing forces that make life not just possible, but—given the right conditions—an entropically favored outcome of the universe [30]. Recent work in Entropy has explored advanced entropy-based methodologies for analyzing complex systems [31], which could provide new tools for understanding the information dynamics underlying biological evolution. A more philosophical perspective on related informational structures in biological and artificial systems is explored in paper [32].
Acknowledgments
The author thanks colleagues for discussions that shaped this work. This research was conducted independently without institutional funding. Some passages of this manuscript were prepared or refined with the assistance of a large language model (LLM). The author takes full responsibility for the content and conclusions presented herein.
References
- Schrödinger, E. (1944). What is Life? The Physical Aspect of the Living Cell. Cambridge University Press.
- Totani, T. (2020). Emergence of life in an inflationary universe. Nature Scientific Reports. [CrossRef]
- Wagner, A. (2014). Arrival of the Fittest: Solving Evolution’s Greatest Puzzle. Current.
- Marković, D., & Gros, C. (2014). Power laws and self-organized criticality in theory and nature. Physics Reports, 536(2), 41-74. [CrossRef]
- Vattay, G., Kauffman, S., & Niiranen, S. (2014). Quantum biology on the edge of quantum chaos. PLoS ONE, 9(3), e89017. [CrossRef]
- Engel, G. S., et al. (2007). Evidence for wavelike energy transfer through quantum coherence in photosynthetic systems. Nature, 446(7137), 782-786. [CrossRef]
- Fleming, G. R., & Scholes, G. D. (2004). Quantum mechanics in photosynthesis. Physics Today, 57(7), 38-43. [CrossRef]
- Ritz, T., et al. (2000). A model for photoreceptor-based magnetoreception in birds. Biophysical Journal, 78(2), 707-718. [CrossRef]
- Brookes, J. C. (2017). Quantum effects in biology: Golden rule in enzymes, olfaction, photosynthesis and magnetoreception. Proceedings of the Royal Society A, 473(2201), 20160822. [CrossRef]
- McFadden, J., & Al-Khalili, J. (2018). The origins of quantum biology. Proceedings of the Royal Society A, 474(2220), 20180674. [CrossRef]
- Ball, P. (2011). Physics of life: The dawn of quantum biology. Nature, 474(7351), 272-274. [CrossRef]
- Mohseni, M., et al. (2014). Quantum Effects in Biology. Cambridge University Press. [CrossRef]
- Kim, Y., et al. (2021). Quantum biology: An update and perspective. Quantum Reports, 3(1), 80-126. [CrossRef]
- Cao, F., et al. (2020). Quantum biology revisited. Science Advances, 6(14), eaaz4888. [CrossRef]
- Abbott, D., et al. (2008). Quantum aspects of life: An introduction. Imperial College Press. [CrossRef]
- Weiss, U. (2012). Quantum Dissipative Systems. World Scientific. [CrossRef]
- Penrose, R. (1989). The Emperor’s New Mind: Concerning Computers, Minds, and the Laws of Physics. Oxford University Press.
- Prigogine, I. (1980). From Being to Becoming: Time and Complexity in the Physical Sciences. W. H. Freeman.
- Davies, P. C. (2004). Does quantum mechanics play a non-trivial role in life? Biosystems, 78(1-3), 69-79. [CrossRef]
- Dawkins, R. (1989). Cumulative selection in relation to complex adaptation. BioEssays, 10(7), 163-169. [CrossRef]
- Glass, D. H. (2014). Parameter dependence in cumulative selection. Springer Nature. [CrossRef]
- Kauffman, S. A. (1993). Origins of Order: Self-Organization and Selection in Evolution. Oxford University Press.
- O’Reilly, E. J., & Olaya-Castro, A. (2014). Non-classicality of molecular vibrations activating electronic dynamics at room temperature. Nature Communications, 5, 3012. [CrossRef]
- Arndt, M., et al. (2014). Testing the limits of quantum mechanical superpositions. Nature Physics, 10(4), 271-277. [CrossRef]
- Marais, A., et al. (2018). The future of quantum biology. Journal of the Royal Society Interface, 15(148), 20180640. [CrossRef]
- Tegmark, M. (2000). Importance of quantum decoherence in brain processes. Physical Review E, 61(4), 4194-4206. [CrossRef]
- Lambert, N., et al. (2013). Quantum biology. Nature Physics, 9(1), 10-18. [CrossRef]
- Cao, J., et al. (2009). Quantum coherence and sensitivity in the avian compass. Proceedings of the National Academy of Sciences, 106(50), 21002-21007. [CrossRef]
- Hameroff, S., & Penrose, R. (2014). Consciousness in the universe: A review of the ‘Orch OR’ theory. Physics of Life Reviews, 11(1), 39-78. [CrossRef]
- Collective of Authors (2022). The complete sequence of a human genome. Science. [CrossRef]
- Wang, C., et al. (2024). Major Role of Multiscale Entropy Evolution in Complex Systems and Data Science. Entropy, 26(4), 330. [CrossRef]
- Straňák, P. (2025). A Hypothetical Informational Field Underlying Biological and Artificial Complexity. Preprints 202507.2653.v1. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



