Submitted:
03 November 2025
Posted:
04 November 2025
You are already at the latest version
Abstract
Fruit and coffee industries are responsible for huge quantities of agro-industrial wastes which is of great environmental and public health concern. Therefore, the aim of this work involves the use of such wastes for production of β-D-glucan from basidiomycete strains which are powerful biological response modifiers in several clinical disorders. Experimental planning for optimization of several parameters was carried out by a full factorial of two levels of three factors for production of be-ta-glucans and basidiomycete species, waste concentration and interaction between species and agro-industrial waste were the most important factors. The best conditions involved a basidiomy-cete strain of Lentinula edodes in a culture medium containing 400 g/L of waste coffee grounds which revealed the production of extracellular β-glucans (141,16 mg/L), at 3rd day of fermentation. Intrinsic fluorescence properties of mushroom β-D-glucan was investigated by fluorescence spec-troscopy as well as fluorescence microtiter plate reader exhibiting emission peaks at 492 and 528 nm .
Differential chromatographic behavior of β-D-glucan was investigated by immobilized metal affinity chromatography (IMAC) by using epoxy-activated Sepharose 6B containing different chelating agents, spacer arms and metal ions. One-step purification of β-D-glucan was devised by using a column of epoxy-activated Sepharose 6B-IDA-Cu (II). FTIR analysis of several β-D-glucan from the chromatographic fractions was carried out to investigate their structural properties.
Keywords:
fruit and coffee industry wastes
; experimental planning
; extracellular of β-D-glucan from basidiomycete mushrooms
; immobilized metal affinity chromatography
; IMAC
; intrinsic fluorescence of of β-D-glucan
; FTIR
1. Introduction
Coffee and fruit industries produce annually a huge amount of agro-industrial wastes which are highly polluted and a disposal problem in the environment [1,2]. These wastes are very rich in a large variety of bioactive compounds which could be valorised into higher value products either by extraction and purification or by fermentation to produce fine chemicals [3,4]. Basidiomycete mushroom strains have been known, for many centuries, to grow on agro-industrial wastes producing high value products such as enzymes, β-D-glucans, lectins, vitamins, lipids and several secondary metabolites [5,6]. It has been well-established that natural or processed foods containing safe amounts of such bioactive compounds are found to provide clinically validated and documented health benefits by using specific biomarkers [7,8]. These foods have been found to improve human health, lower the risk of chronic bacterial and viral diseases as well as to prevent and management of symptoms of diseases [8,9].
Among these biomolecules, β-D-glucans play a major role as biological response modifiers (BRM) in several clinical disorders such as immunomodulatory properties, antimicrobial and antioxidant activity, and hypocholesterolemic and hypolipidemic effects, HIV, cancer, diabetes, neurological, cardiovascular and immunological disorders [10,11].
Experimental planning is an important strategy for overproduction of mushroom β-D-glucans from agro-industrial wastes as it provides useful information about significant factors and the interactions between the variables [12]. There are a vast majority of agro-industrial wastes that can be used for cultivation of basidiomycete strains such as rice husks, coffee residues, fruit peels and pomaces, agricultural straw and dairy wastes. Several basidiomycete mushrooms have been used in these bioconversion processes such as Pleurotus sp. Agaricus sp., Cordyseps sp. and Hericium sp. [1,2,3,4].
Basidiomycete mushroom strains produce a heterogeneous mixture of several β-D-glucans of different sizes, structures and either bound to proteins or in free forms [13,14]. The purification of such β-D-glucans in homogeneous preparations is of crucial interest to investigate their structure-function relationship [15,16,17]. Therefore, it is of great interest to devise a specific chromatographic technique to purify these biological macromolecules as well as to investigate their structural properties. Immobilized metal affinity chromatography (IMAC) has been widely used to purify proteins with high degree of purity [18]. To our knowledge , this technique has not been used to isolate and purify β-D-glucans from basidiomycete strains. As far as literature is concerned, there are a few reports for purification of β-D-glucans from mushrooms strains by using gel filtration and ion-exchange chromatography [16,17] On the other hand, structural characterization of β-D-glucans is of great scientific interest in order to understand their interaction with cells in vivo [11,13,15] To our knowledge, intrinsic fluorescence techniques have not been used to characterize β-D-glucans from basidiomycete strains although the use of fluorescence probes bound to β-D-glucans has been reported in the literature [19,20] FTIR analysis of β-D-glucans is valuable to obtain structural information about these biological macromolecules and therefore several chromatographic factions of β-D-glucans were analysed by FTIR to investigate their structural properties [16].
The present work involved experimental planning for optimization of production of extracellular β-D-glucans from several basidiomycete strains by using several coffee and fruit agro-industrial wastes. Subsequently, these β-D-glucans were purified by IMAC by using different stationary phases and metal chelates and fluorescence and FTIR properties of these β-D-glucans were investigated in detail.
2. Materials and Methods
2.1. Chemicals
Imidazole, iminodiacetic acid (IDA), 1,4-butanediol diglycidyl ether (BDGE), Sepharose 6B, epichlorohydrin (EPI), Sepharose 4B and 6B, Congo red dye, Alcian blue 8GX, phenol, β-D-1,3-glucan from Euglena gracilis and barley β-D-glucan were obtained from Sigma -Aldrich (USA).
Potato dextrose agar (PDA) medium was supplied by HiMedia Laboratories (Mumbai, India). Milk permeate, orange and apple peels, pine sawdust, yellow lupine, rice husk, apple pomace, yellow lupin and waste coffee grounds were homemade. All other reagents were of analytical grade.
2.2. Mushroom Samples
Pleurotus ostreatus and Ganoderma carnosum were isolated from old growth forests of the Olympic Peninsula in Port Townsend - WA (USA). Pleurotus ostreatus mushroom stems were a generous gift of a mushroom grower in Amsterdam (Netherlands). These mushroom strains, Fomes fomentarius, Ganoderma applanatum, Ganoderma lucidum violeta, Irpex lacteus, Phlebia rufa and Piptoporus betulinus were kindly supplied by Department of Agronomy of University of Trás-os-Montes and Alto Douro (UTAD). These cultures were grown in PDA and maintained at 4 °C. On the other hand, the young fruiting body of Agaricus blazei, Inonotus obliquus, Polyporus umbellatus, Poria cocos, Ganoderma lucidum, Hericium erinaceus, Coriolus versicolor and Lentinula edodes were kindly supplied by Mycology Research Laboratory, Ltd. (United Kingdom).
In order to develop a culture medium for cultivation of mushroom strains for β-D-glucans production, several agro-industrial wastes were tested. The agro-industrial wastes in dry form were corncobs, rice husks, oat husks, sawdust, and lupin seeds which were supplied by the Department of Agronomy of University of Trás-os-Montes and Alto Douro (UTAD). The wet agro-industrial wastes were coffee grounds, pineapple, banana, mango and pear peels.
2.3. Methods
2.3.1. Growth and Maintenance Conditions of Mushroom Strains
All mushroom strains were grown in PDA for 2 weeks at 25 ºC and maintained at 4ºC in PDA.
2.3.2. Production of β-D-glucan from Basidiomycete Mushroom Strains in Culture Media Containing Agro-Industrial Wastes
All mushroom strains were grown in media containing 1 g/L KH2PO4, 1 g/L MgSO4, 1g/L (NH4)2SO4, 15 g/L of purified agar and 4 g/L either dry residues or 40 g/L of wet residues at pH 5.6. In a second phase, the basidiomycete cultures that exhibited growth in the media described above were inoculated into liquid medium which had the same composition as the solid media, except it did not include purified agar. The mycelia on the solid medium were scraped off and collected in Eppendorf tubes containing sterile saline. After resuspension of the mycelia, it was used as inoculum for the liquid media. The cultures were incubated at 25 °C in an incubator with orbital shaking at 150 rpm for 14 days.
2.3.3. Isolation of of β-D-Glucan from Basidiomycete Mushroom Strains
After fermentation, the mycelial biomass was separated from the fermentation broth by centrifugation for 30 min at 4,500 rpm at 4 °C. Extracellular β-D-glucans were isolated by precipitation with 4 volumes of 95% ethanol to the supernatant and the mixture was stored overnight at 4 °C. The precipitate was collected after centrifugation for 5 min, 10,000 rpm and 4 °C. Finally, the sediment was dissolved in 50 mM phosphate buffer pH 7.4.
The isolation of intracellular β-D-glucans (IBG) from mycelial biomass pellet from Pleurotus ostreatus, Ganoderma applanatum and Ganoderma carnosum and from young fruiting bodies powder from Agaricus blazei, Ganoderma lucidum, Hericium erinaceus, Coriolus versicolor, Lentinula edodes, Pleurotus ostreatus, Inonotus obliquus, Auricularia auricula, Grifola frondosa, Polyporus umbellatus, Cordyseps sinensis and Poria cocos was performed by multistep water extraction followed by extraction with alkali and acidic solutions as described previously [21]. Briefly, the aqueous extraction from mycelial biomass pellet from P. Briefly, FW1, FW2, FKOH, FHCl, FNaOH represent samples of extraction with cold H2O, hot H2O, KOH, HCl and NaOH, respectively [21].
2.3.4. Total Polysaccharides and Protein Assays
2.3.5. Congo Red Assay for Specific Determination of of β-D-glucan with Triple Helical Structure
Congo red dye colorimetric assay was carried out to quantify the concentration of β-D-glucans in several samples as described previously, using β-D-glucans from barley as standard [21].
2.3.6. Alcian Blue Dye Colorimetric Assay for of β-D-Glucan
Alcian blue dye colorimetric assay was carried out to quantify the concentration of β-D-glucans in several samples as described previously, using β-D-glucans from barley as standard [24].
2.3.7. Experimental Design to Optimize Extracellular β-D-Glucan Production
The experimental design was conducted using three factors and two levels with duplicates: basidiomycete species (Lentinula edodes and Ganoderma applanatum), agro-industrial waste concentration (40 g/L and 400 g/L for wet waste or 4 g/L and 40 g/L for dry waste), and cultivation time (3 and 7 days). The upper (+) and lower (-) levels were defined based on the preliminary results. The experimental design matrix was obtained with Design Expert version 10 software, and the results were fitted to the following factorial model equation:
where y represents the concentration of β-D-glucans, β0 is the mean of all responses, β is the regression coefficient and x are the prediction variable. The statistical analysis of the model was evaluated according to the ANOVA methodology.
2.3.8. Preparation of Stationary Phases for IMAC:
The activation of Sepharose 6B and 4B was carried out as described previously [25] by using either epichlorohydrin (EPI) or 1,4-butanediol diglycidyl ether (BDGE) as the spacer arm and either iminodiacetic acid (IDA) or nitrilotriacetic (NTA) as chelating agents.
2.3.9. Chromatographic Behaviour of β-D-glucans from Mushroom Strains on Immobilized Metal Chelates (IMAC):
A rapid batch method carried out in ELISA microtiter plates was used to study the adsorption of β-D-glucans on stationary phases [25]. In the present work, β-D-glucans adsorption to all these stationary phases was considered to be partial when β-D-glucans was detected both at the washing and elution steps. On the other hand, β-D-glucans did not bind to the column when total β-D-glucans applied to the column was recovered in five column volumes of buffer at the washing step, exhibiting a sharp activity peak. 96-well microtiter plates were used to study the adsorption of β-D-glucans from mushroom strains on immobilized metal chelates of Cu(II), Ni(II), Zn(II) and Co(II) at pH 8.0. The culture supernatant containing β-D-glucans (0.1 mg) was diluted in 20 mM sodium phosphate buffer containing 1M NaCl pH 8.0. Desorption of the β-D-glucans from metal chelates was carried out by using 20 mM phosphate buffer containing 1 M NaCl and 500 mM imidazole. All washing and elution fractions were assayed for β-D-glucan by congo red assay method and these experiments were performed in triplicates at room temperature.
2.3.10. Purification of β-D-Glucan from Basidiomycete Mushroom Strains by IMAC
According to the results of the chromatographic behaviour performed on 96-well microtiter plates, a set of experimental conditions were selected for purification of β-D-glucan from some mushroom strains. A column containing 5 mL of sedimented epoxy-activated Sepharose 6B-IDA-Cu(II) (S3) as the stationary phase was washed sequentially with 5 volumes of 50 mM copper sulfate, water and then finally with 20 mL of equilibration buffer at pH 8.0 [26]. The culture supernatant containing β-D-glucan was diluted 1:2 with equilibration buffer and applied to the column at a flow rate of 15 mL/h. The column was then washed with equilibration buffer and the chromatographic fractions were collected and read at A280, A510 and fluorescence intensity. Once the A280 was less than 0.05, β-D-glucan was desorbed from the column with a linear gradient of imidazole (0-500 mM) in the same buffer system at a flow rate of 30 mL/h and fractions were assayed for β-D-glucan and protein content by fluorescence and colorimetric assays.
2.3.11. Intrinsic Fluorescence Measurements of β-D-Glucans
The samples containing β-D-glucans were also analyzed on a spectrofluorimeter (JASCO JP-8300) in quartz cuvettes with a 1 cm optical path length. Spectra Manager software was used for spectral acquisition and processing. The following parameters were set to obtain synchronous fluorescence spectra: excitation wavelength of 250-735 nm; emission wavelength of 260-750 nm; excitation and emission slit widths of 5 nm; variation of delta wavelength Dl of 5, 10, 20 and 30 nm , excitation wavelength increments of 5 nm; emission wavelength increments of 0.5 nm; response of 0.1 s; light source of Xe lamp and scan speed of 10,000 nm/min.
For steady-state fluorescence assays in a microtiter plate fluorescence reader, 100 µL of sample was pipetted into a well of a NUNC 96 microplate. The samples consisted of extracellular β-D-glucan from Lentinula edodes cultures and commercial barley β-D-glucan for comparative purposes. The samples were analyzed in triplicate, and blank assays consisting solely of deionized water and phosphate buffer were performed for each microplate. Intrinsic fluorescence was read in a microplate reader (FLUOstar OPTIMA-BMG Labtec), using excitation filters corresponding to wavelengths of 380, 400, 420, 430, 485, or 510 nm and emission filters corresponding to 480, 528, 542, 550, or 620 nm.
2.3.12. FTIR Analysis of β-D-Glucans
In order to exploit the structural features of β-D-glucans, FTIR analysis was performed. All samples were previously freeze-dried with a UNICRYO MC2L. Infrared spectra were recorded on a Bruker Vertex 70 (with OPUS 5.5 software) as KBr pellets, a total of 128 scans at a resolution of 2 cm-1 in a range of 400–4000 cm−1.
2.3.13. Statistical Analysis
SigmaPlot 16.0 (2011-2012 Systat Software inc.) was used to draw graphs in this research work. Experimental results are means of three parallel measurements and the results are presented as mean values ± standard deviation (SD). Correlation and regression analyses were performed with the Excel software 2024 package (Academic License, Microsoft of Portugal). Correlations were considered statistically significant at p < 0.05 according to Tukey HSD and Scheffé test.
3. Results and Discussion
3.1. Optimization of Cultivation Conditions for β-D-Glucan Overproduction
Basidiomycete mushroom strains were inoculated onto solid media supplemented with several agro-industrial wastes, according to the materials and methods. The data in Table 1 revealed the cultivation time required to obtain mycelial biomass across the entire surface of the solid media in the petri dishes with different basidiomycete species and agro-industrial wastes. According to Table 1, the four residues that resulted in the fastest culture growth were yellow lupine, coffee grounds, banana peels and oat hulls. Cultures grown on solid media containing these wastes were then inoculated into liquid medium containing the same agro-industrial residue. From these four agro-industrial residues tested, waste coffee grounds yielded the highest concentrations of β-D-glucan.
The data in Figure 1A have revealed that Lentinula edodes and Ganoderma applanatum exhibited the highest β-D-glucan levels when grown in media supplemented with waste coffee grounds.
Phebia rufa was the only basidiomycete strain that produced β-D-glucan when grown in medium supplemented with yellow lupin (Figure 1 B). However, the levels of β-D-glucan obtained in this medium were lower than those obtained from cultures grown in medium with coffee grounds.
The data in Figure 2 have revealed that the levels of β-D-glucan obtained by the Congo Red dye and phenol-sulfuric acid methods were rather different since the latter is a broad assay for carbohydrate analysis which detected all types of sacharides including proteoglycans, glycoproteins and glycolipds [22]. The highest β-D-glucan levels was observed on 11th day of culture, whereas highest polysaccharide levels were obtained on the 14th day of culture. β-D-glucan levels determined by the Alcian Blue dye method, exhibited a similar profile to Congo Red dye method, although with lower values and highest β-D-glucan levels were obtained on 7th day rather than 11th day of culture.
According to [21] the Congo Red dye method measured β-D-glucans with a triple helix structure whereas the Alcian Blue dye method was more sensitive and quantified β-D-glucans, regardless of the conformation presented in these biological macromolecules [24]. The levels of extracellular β-D-glucans obtained by using the Alcian Blue dye method for Lentinula edodes supernatant were surprisingly lower than those obtained using Congo Red (Figure 2). Regarding intracellular β-D-glucans from basidiomycetes, the levels reported using Alcian Blue dye were lower than those obtained with Congo Red for only a few samples. These results may be explained by the fact that the alcian blue method detects acidic and sulfated β-D-glucans whose levels may be low in such samples whereas Congo red assay detects neutral β-D-glucans with triple helix conformation [21,24].
The data in Table 2 revealed the comparative analysis of the levels of extracellular and intracellular β-D-glucans produced by these three basidiomycete strains in several agro-industrial wastes.
These data are difficult to compare with published reports on β-D-glucans production because the culture media, mushroom strains and growth conditions are significantly different from the present work.
In order to optimize the overproduction of extracellular β-D-glucans from submerged fermentation of basidiomycete strains, a systematic study was developed to analyze several factors that affect the production of these compounds. The experimental design was conducted using a 23-way full factorial design, with a total of eight duplicate experiments. After the experimental runs, statistical significance, the effect of each variable, and multivariate interactions on β-D-glucan concentration were evaluated (Table 3).
The data in Table 3 summarized the statistical analysis of variance (ANOVA). The F-value of 1.34x104 indicated that the model is significant and that there was only a 0.01% probability of such an F-value being due to noise. The coefficient of determination R2 = 0.9999 indicated that there was a statistical correlation between the response and the variables considered and that only 0.01% of the total variation was not explained by the model. The statistical analysis shows that the significant factors for β-D-glucan production are all those considered: the basidiomycete species (variable A), the agro-industrial waste concentration (variable B), the cultivation time (variable C), and the interactions between the variables AB, AC, BC, and ABC. However, the basidiomycete species, the waste concentration, and the interaction between the species and the agro-industrial waste exhibited the greatest effect. The following empirical equation was obtained to estimate the concentration of β-D-glucans:
y = 39,77-26,57xA+26,18xB-2,97xC-28,53xAB+1,27xAC-1,66xBC+14,21xABC
where: y is the concentration of β-D-glucans, A is the species of basidiomycete, B is the concentration of agro-industrial waste and C is the cultivation time. This equation was used to facilitate plotting the response surfaces which are represented in Figure 3A and Figure 3B.
The data in Figure 3A and Figure 3B exhibited the effect the optimized factors on β-D-glucan production during submerged fermentation.
The culture of Lentinula edodes in a culture medium with 400 g/L of coffee grounds produced the highest concentration of extracellular β-D-glucans (141.16 mg/L) on the third day of fermentation and was therefore the culture selected for subsequent studies (Figure 3B). These data on experimental planning are difficult to perform a comparative analysis with published reports as there are no work reported in the literature about mushroom β-D-glucans.
3.2. Intrinsic Fluorescence Spectroscopy
Since the literature search revealed lack of publications on the intrinsic fluorescence of β-D-glucans from medicinal basidiomycete strains without the use of extrinsic fluorochromes, we decided to investigate the intrinsic fluorescence of these β-D-glucans. Therefore, the sensitivity, or gain, of the microplate reader was initially evaluated. In these tests, a sample of extracellular β-D-glucans extracted from a culture of Lentinula edodes grown in 400 g/L of waste coffee grounds was used as described in the Materials and Methods section. The instrument gain/sensitivity was monitored in the range of 1800 and 2200, and the results were exhibited in Figure 4A. These data revealed an increase in fluorescence intensity as a function of sensitivity or gain with the highest peak at 528 nm and 2200 (Figure 4A).
The data in Figure 4B exhibited the highest fluorescence emission of the extracellular β-D-glucans from Lentinula edodes at 528 nm with excitation at 420 nm.
Additionally, these samples were analyzed by synchronous fluorescence spectroscopy by using a spectrofluorometer. The data in Figure 5 A exhibited synchronous fluorescence spectra for commercial barley β-D-glucan at 10 nm Dl at different concentrations by obtaining peaks at 492 and 540 nm.
Similarly, we also investigated synchronous fluorescence spectra of β-D-glucan from several mushroom strains as shown in Figure 5B. These data also revealed two peaks at 492 and 540 nm which are in agreement with barley β-D-glucan spectra.
Regarding these fluorescence data on mushroom β-D-glucan, it is not possible to perform a comparative analysis with published report as there are no work reported in the literature about intrinsic fluorescence of β-D-glucan.
3.3. Immobilized Metal Affinity Chromatography
Several chromatographic gels containing different immobilized transition metals, spacer arms and stationary phases were tested at pH 8 and the results are shown in Table 4. The chromatographic behavior of extracellular β-D-glucans from Lentinula edodes, grown in a medium supplemented with 400 g/L of waste coffee grounds, exhibited adsorption of these β-D-glucans to IMAC columns which was affected by several factors, such as the size of the spacer arm, chelating agent and metal ion (Table 4).
According to literature search, there are no reports on the use of IMAC for separation and purification of mushroom β-D-glucans. It has been shown that longer spacer arms, such as 1,4 butanediol diglycidyl ether (BDGE) increase the adsorption of monoclonal antibodies as well as contaminating proteins [25] . As far as β-D-glucans are concerned, Cu(II) immobilized chelates in the matrix exhibited the highest adsorption on S1 compared to S2, which contains a EPI as spacer arm and is therefore shorter. However, the opposite result was obtained when comparing S3 with S4 and S5 with S6. During this present investigation, two chelating agents IDA (tridentate) and NTA (tetradentate) were analyzed. From the structural point of view, IDA and NTA differ by the presence of an additional carboxymethyl group on NTA which makes it a stronger coordinator of metal ions because it has 4 valencies available compared to the 3 of IDA. Although both chelating agents have 2 valencies available to interact with the imidazole group of histidine residues, NTA has higher affinity for metal ions compared to IDA (Table 4).
The data in Table 4 has revealed β-D-glucans adsorption occurred in the following order : Cu2+ > Zn2+ > Ni2+ > Co2+, β-D-glucans from Lentinula edodes exhibited adsorption to the Sepharose 4B-BDGE-EPI-Cu2+ stationary phase (pH 8).
Among all stationary phases tested, S3 stationary phase of Sepharose 4B-BDGE-IDA-Cu(II) was the best chromatographic gel for purification of β-D-glucans with high recovery although it exhibited a partial adsorption of β-D-glucans (Figure 6 A).
The data shown in Figure 6A exhibited two β-D-glucans peaks at washing and desorption steps assayed by congo red method which correlated well with fluorescence data at 528 nm.
Since we were interested to identify protein-bound β-D-glucans in Lentinula edodes, a chromatographic profile was obtained for protein (A280) and β-D-glucan assays as well as imidazole gradient (Figure 6 B).
The data in Figure 6 B have revealed two protein and β-D-glucans peaks at the washing and desorption steps suggesting a partial adsorption of these biological macromolecules. Therefore, these β-D-glucans were either bound to proteins or some free contaminating proteins were present in these column fractions. IMAC has been widely used for purification of native and recombinant proteins containing available histidine residues on their surfaces [27,28]. As far as adsorption of β-D-glucans to immobilized metal chelates is concerned, the molecular mechanism may be due to hydrophobic interactions between the solute and long spacer arm of the stationary phase [26].
3.4. FTIR Spectroscopy
The purified fractions of β-D-glucans from IMAC of Lentinula edodes were analysed by FTIR spectroscopy. The data in Figure 7 have revealed some absorption bands characteristic of β-D-glucans.
Figure 6.
- FTIR spectra of selected IMAC fractions of extracellular Beta-D- glucan from L. edodes. .
.
.
Figure 6.
- FTIR spectra of selected IMAC fractions of extracellular Beta-D- glucan from L. edodes..
.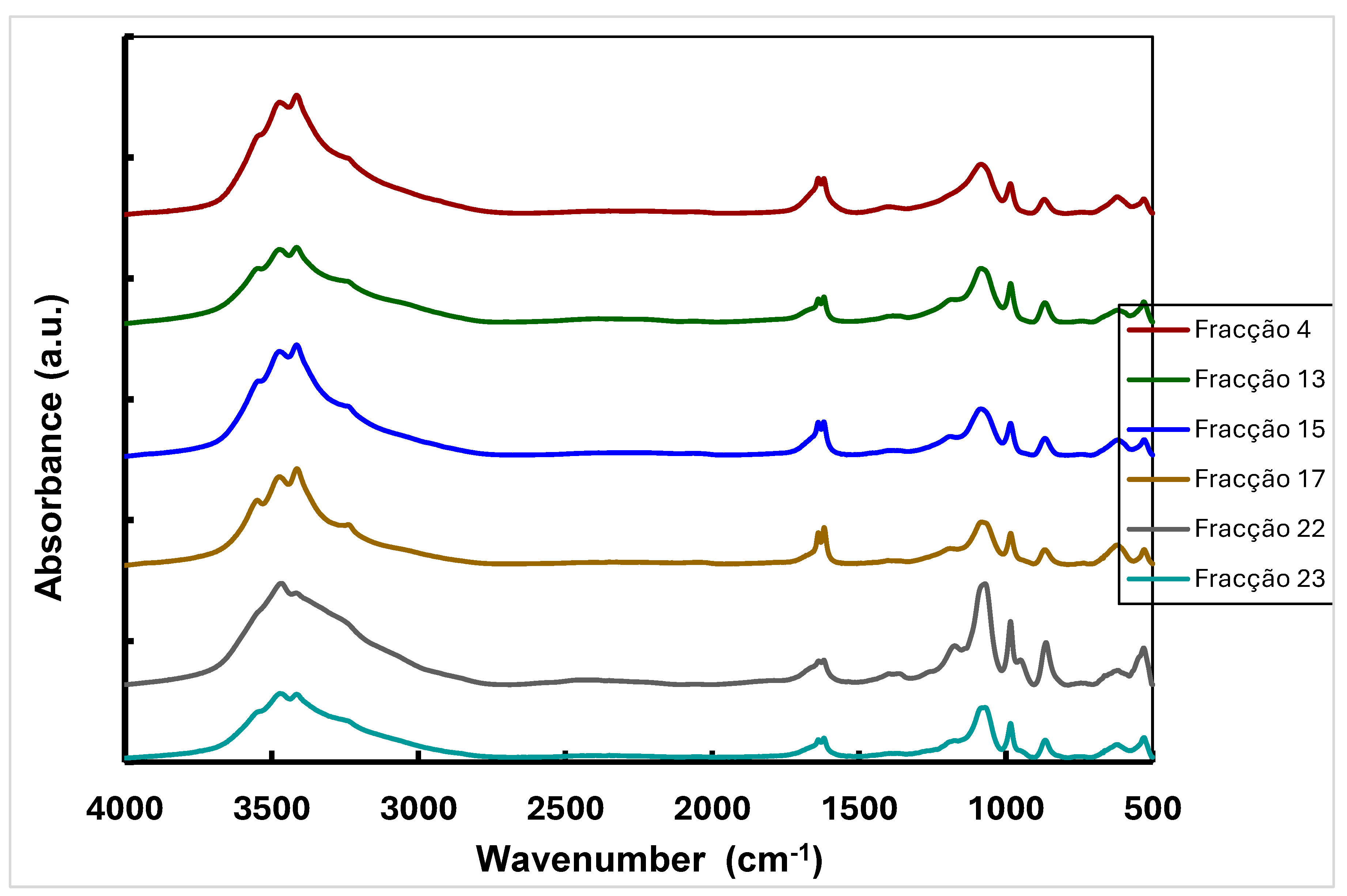
The strong band at 3474 cm-1 is due to the extension vibration of the O-H bond whereas this broad band centered near 3300 cm-1 is specific for carbohydrates [16,29]. It has been described a broad band at 3000-3500 cm-1 in the spectrum of Ganoderma apllanatum, corresponding to the O-H extension vibration in hydrogen bonds and the N-H vibration [29]. Moreover, some bands were also observed in the spectra at 1638 cm-1, corresponding to the stretching vibration of the C=O group of amide I, and at 1619 cm-1 due to deformation of the NH bond and the C-N extension of amide II, which suggest the presence of proteins. There are also absorption bands characteristic of the C-O-C deformation (1180 cm-1), the anomeric C vibration of carbohydrates (1080 cm-1) and the β conformation of carbohydrates (865 cm-1), which demonstrate the existence of β-D-glucans in all fractions [16,29].
4. Conclusions and Future Work
To author’s knowledge, this is the first report about the use of experimental planning for β-D-glucan production from mushroom strains. This work involved the optimization of culture media containing agro-industrial wastes to maximize the productivity of intra- and extracellular β-D-glucans from basidiomycete strains. The highest β-D-glucans concentration (141.16 mg/L), determined by the Congo red dye method, was obtained after extracting β-D-glucans from the fermentation broth of Lentinula edodes cultures in a medium containing 400 g/L of coffee grounds. To author’s knowledge, this is the first report about the use of IMAC for purification of β-D-glucans from L. edodes which was influenced by the size of the spacer arm, chelating agent and metal ion. To author’s knowledge, this is the first report about the intrinsic fluorescence analysis of β-D-glucans from mushroom strains. The purified fractions were analysed by fluorescence as well as by FTIR spectroscopy in order to obtain additional structural information about these biological macromolecules. However, the characterization of fractionated β-D-glucans should be complemented with future studies by using ELISA, HPLC, GC-MS, FTIR, Fluorescence spectroscopy and NMR.
Author’s contribution
AK designed the research plan and performed experiments on fluorescence properties of β-D-glucans, LM performed all the experiments and data treatment and AK wrote the MS.
Financial support
Project UID/AGR/04033/2019.
Acknowledgments
“This work was supported by: European Investment Funds by FEDER/COMPETE/POCI– Operacional Competitiveness and Internacionalization Programme, under Project POCI-01-0145-FEDER-006958 and National Funds by FCT - Portuguese Foundation for Science and Technology, under the project UID/AGR/04033/2019.”.
Conflicts of Interest
The authors declare that there are no conflicts of interest.
Abbreviations:
| BDGE | 1,4-Butanediol diglycidyl ether |
| EPI | Epiclorohydrin |
| IDA | Iminodiacetic acid |
| IMAC | Immobilized metal affinity chromatography |
| NTA | Nitrilotriacetic acid |
| PBS | Phosphate Buffered Saline |
| PDA | Potato Dextrose Agar |
References
- Luna, K.A.; Aguilar, C.N.;Ramírez-Guzmán, N.; Ruiz, H.A.;Martínez, J.L.; Chávez-González, M.L. Bioprocessing of Spent Coffee Grounds as a Sustainable Alternative for the Production of Bioactive Compounds. Fermentation 2025, 11,366. [CrossRef]
- Otieno, O.D., Mulaa, F.J., Obiero, G., Midiwo, J., Utilization of fruit waste substrates in mushroom production and manipulation of chemical composition. Biocatalysis and Agricultural Biotechnology 39, (2022), 102250 . [CrossRef]
- Kowalski, S.; Gumul, D. The Use of Waste Products from the Food Industry to Obtain High Value-Added Products. Foods 2024, 13, 847. [CrossRef]
- Khatam , K., Qazanfarzadeh, Z.,Jim’enez-Quero, A., Fungal fermentation: The blueprint for transforming industrial side streams and residues. Bioresource Technology 440 (2026) 133426 . [CrossRef]
- (5) Lysakova, V.; Streletskiy,A.; Sineva, O.; Isakova, E.;Krasnopolskaya, L. Screening of Basidiomycete Strains Capable of Synthesizing Antibacterial and Antifungal Metabolites. Int. J. Mol. Sci. 2025, 26, 9802. [CrossRef]
- Bhambri A, Srivastava M, Mahale VG, Mahale S and Karn SK (2022) Mushrooms as Potential Sources of Active Metabolites and Medicines. Front. Microbiol. 13:837266. [CrossRef]
- Fekete, M.; Lehoczki, A.; Kryczyk-Poprawa, A.; Zábó, V.; Varga, J.T.; Bálint, M.; Fazekas-Pongor, V.; Csípo˝, T.; Rza˛sa-Duran, E.; Varga, P. Functional Foods in Modern Nutrition Science: Mechanisms, Evidence, and Public Health Implications. Nutrients 2025, 17, 2153. [CrossRef]
- Rauf, A., Joshi, P. B., Ahmad, Z.,Hemeg, H. A., Olatunde, A., Naz, S., Hafeez, N., &Simal-Gandara, J. (2023). Edible mushrooms as potential functional foods in amelioration of hypertension. PhytotherapyResearch, 37(6), 2644–2660. [CrossRef]
- Dimopoulou, M.; Chinou, I.; Gortzi, O. A Systematic Review of the Seven Most Cultivated Mushrooms: Production Processes, Nutritional Value, Bioactive Properties and Impact on Non-Communicable Diseases. Agriculture 2025, 15, 1329. [CrossRef]
- Case S, O’Brien T, Ledwith AE, Chen S, Horneck Johnston CJH, Hackett EE, O’Sullivan M, Charles-Messance H, Dempsey E, Yadav S, Wilson J, Corr SC, Nagar S and Sheedy FJ (2024) b-glucans from Agaricus bisporus mushroom products drive Trained Immunity. Front. Nutr. 11:1346706. [CrossRef]
- Patel, D.K.; Dutta, S.D.; Ganguly, K.; Cho, S.-J.; Lim, K.-T. Mushroom-Derived Bioactive Molecules as Immunotherapeutic Agents: A Review. Molecules 2021, 26, 1359. [CrossRef]
- Feng, Y-L., Li, W.Q., Wu X-Q., Cheng, J-W., Ma S-Y., Statistical optimization of media for mycelial growth and exo-polysaccharide production by Lentinus edodes and a kinetic model study of two growth morphologies Biochemical Engineering Journal, 49, 2010,104-112 . [CrossRef]
- Singla, A.; Gupta, O.P.; Sagwal, V.; Kumar, A.; Patwa, N.; Mohan, N.; Ankush; Kumar, D.; Vir, O.; Singh, J.; et al. Beta-Glucan as a Soluble Dietary Fiber Source: Origins, Biosynthesis, Extraction, Purification, Structural Characteristics, Bioavailability, Biofunctional Attributes, Industrial Utilization, and Global Trade. Nutrients 2024, 16, 900. [CrossRef]
- Pérez-Bassart, Z., Fabra, M.J., Martínez-Abad, A., López-Rubio, A., Compositional differences of β-glucan-rich extracts from three relevant mushrooms obtained through a sequential extraction protocol Food Chemistry 402, 2023, 134207.
- Han B, Baruah K, Cox E, Vanrompay D and Bossier P (2020) Structure-Functional Activity Relationship of b-Glucans From the Perspective of Immunomodulation: A Mini-Review. Front. Immunol. 11:658. [CrossRef]
- Wang, W.; Tan. J.; Nima, L.; Sang. Y.; Cai. X.; Xue, H.; Polysaccharides from fungi: A review on their extraction, purification, structural features, and biological activities Food Chemistry: X 15 (2022) 100414.
- Flores, G.A.,Cusumano, G., Venanzoni, R., Angelini, P.The Glucans Mushrooms: Molecules of Significant Biological and Medicinal Value. Polysaccharides 2024, 5(3), 212- 224; [CrossRef]
- Martins, S., Karmali, A., Serralheiro, M.L. Chromatographic behaviour of monoclonal antibodies against wild-type amidase from Pseudomonas aeruginosa on immobilized metal chelates. Biomedical Chromatography 2011 . [CrossRef]
- Theint, P.P., Sakkayawong, N. Buajarern, S. Singkhonrat, J. (2025) Development of an analytical fluorescence method for quantifying β-glucan content from mushroom extracts; utilizing curcumin as a green chemical fluorophore Journal of Food Composition and Analysis, 137, Part A . [CrossRef]
- Koenig, S., Rühmann, B., Sieber, V., Schmid, J. (2017) Quantitative assay of β-(1,3)–β-(1,6)–glucans from fermentation broth using aniline blue Carbohydrate Polymers, 174, 2017, 57-64 . [CrossRef]
- Semedo, M.C., A. Karmali, and L. Fonseca (2015) A high throughput colorimetric assay of β-1,3-D-glucans by Congo red dye. J. Microbiol. Meth. 109: 140–148.
- Masuko, T., A. Minami, N. Iwasaki, T. Majima, S.-I. Nishimura, and Y.C. Lee (2005) Carbohydrate analysis by a phenol–sulfuric acid method in microplate format. Anal. Biochem. 339: 69–72.
- Bradford, M.M. (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein–dye binding. Anal. Biochem. 72: 248–254.
- Semedo, M.C., A. Karmali, and L. Fonseca (2015) A novel colorimetric assay of β-D-glucans in basidiomycete strains by alcian blue dye in a 96-well microtiter plate. Biotechnol. Prog. 31: 1526-1535.
- Freixo, MR., Karmali, A., Frazão, C., Arteiro, J.M. Production of laccase and xylanase from Coriolus versicolor grown on tomato pomace and their chromatographic behaviour on immobilized metal chelates. Process Biochemistry, 43, 2008, 1265-1274.
- Martins, S., Karmali, A., Andrade, J., Serralheiro, M. (2006), “Immobilized metal affinity chromatography of monoclonal immunoglobulin M against mutant amidase from Pseudomonas aeruginosa”, Molecular Biotechnology, 33, 103-113.
- Gaberc-Porekar, V., Menart, V. (2001), “Perspectives of immobilized-metal affinity chromatography”, Journal of Biochemical and Biophysical Methods, 49, 335-360.
- Block, H. Maertens, B., Spriestersbach, A., Brinker, N., Kubicek, J.,Fabis, R., Labahn, J., Schäfer, F. Methods in Enzymology, Volume 463, 2009 Chapter 27 Immobilized-Metal Affinity Chromatography (IMAC): A Review ,439-473.
- Kozarski, M., Klaus, A., Nikšić, M., Vrvić, M., Todorović, N., Jakovljević, D., Griensven, L. (2012), “Antioxidative activities and chemical characterization of polysaccharide extracts from the widely used mushrooms Ganoderma applanatum, Ganoderma lucidum, Lentinus edodes and Trametes versicolor”, Journal of Food Composition and Analysis, 26, 144-153.
Figure 1.
Production of extracelular β-D-glucan as a function of cultivation time in culture supernatants of basidiomycete strains grown in culture medium containing waste coffee grounds (A) and yellow lupine (B). β-D-glucan was assayed by Congo red method.
Figure 1.
Production of extracelular β-D-glucan as a function of cultivation time in culture supernatants of basidiomycete strains grown in culture medium containing waste coffee grounds (A) and yellow lupine (B). β-D-glucan was assayed by Congo red method.
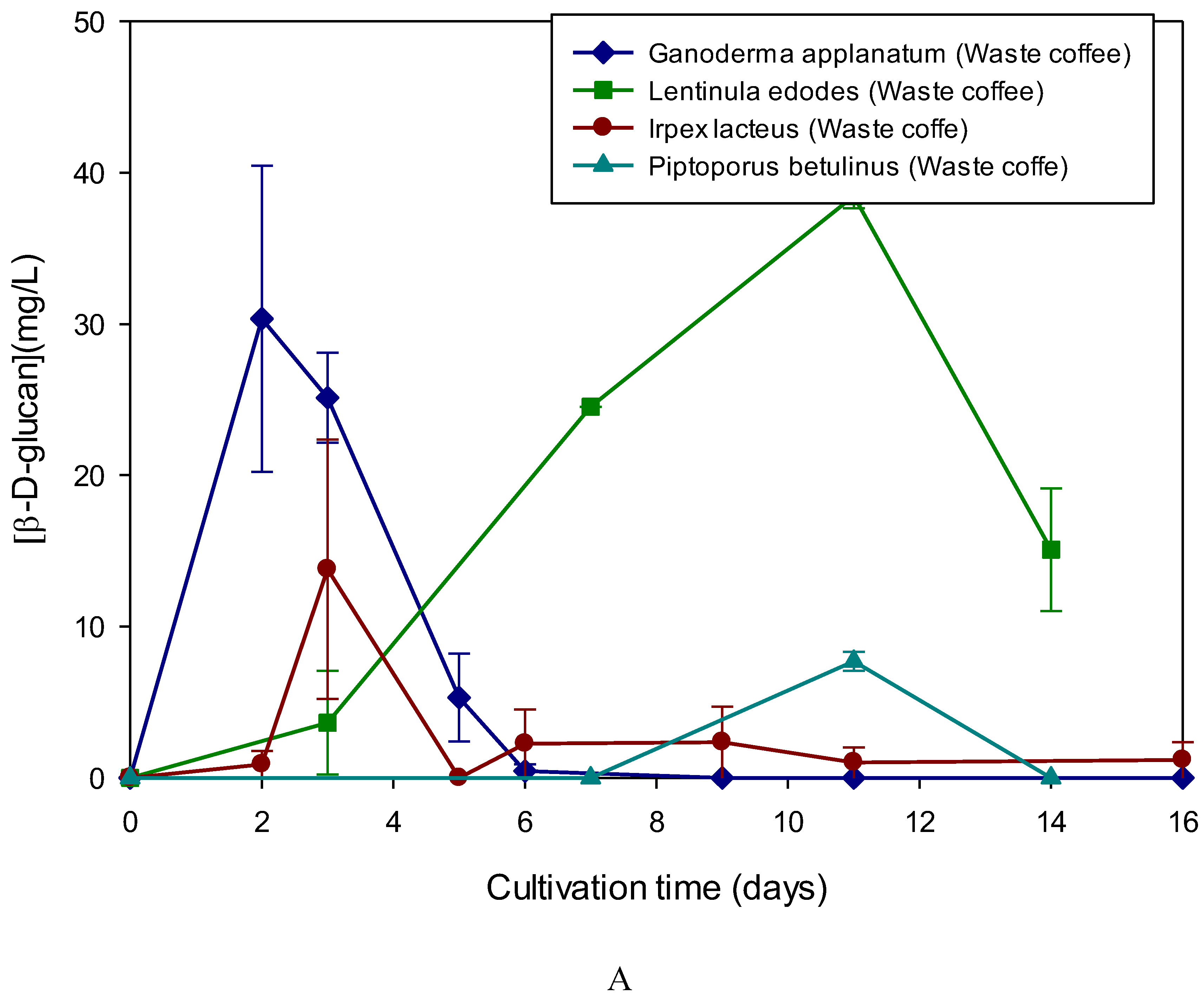
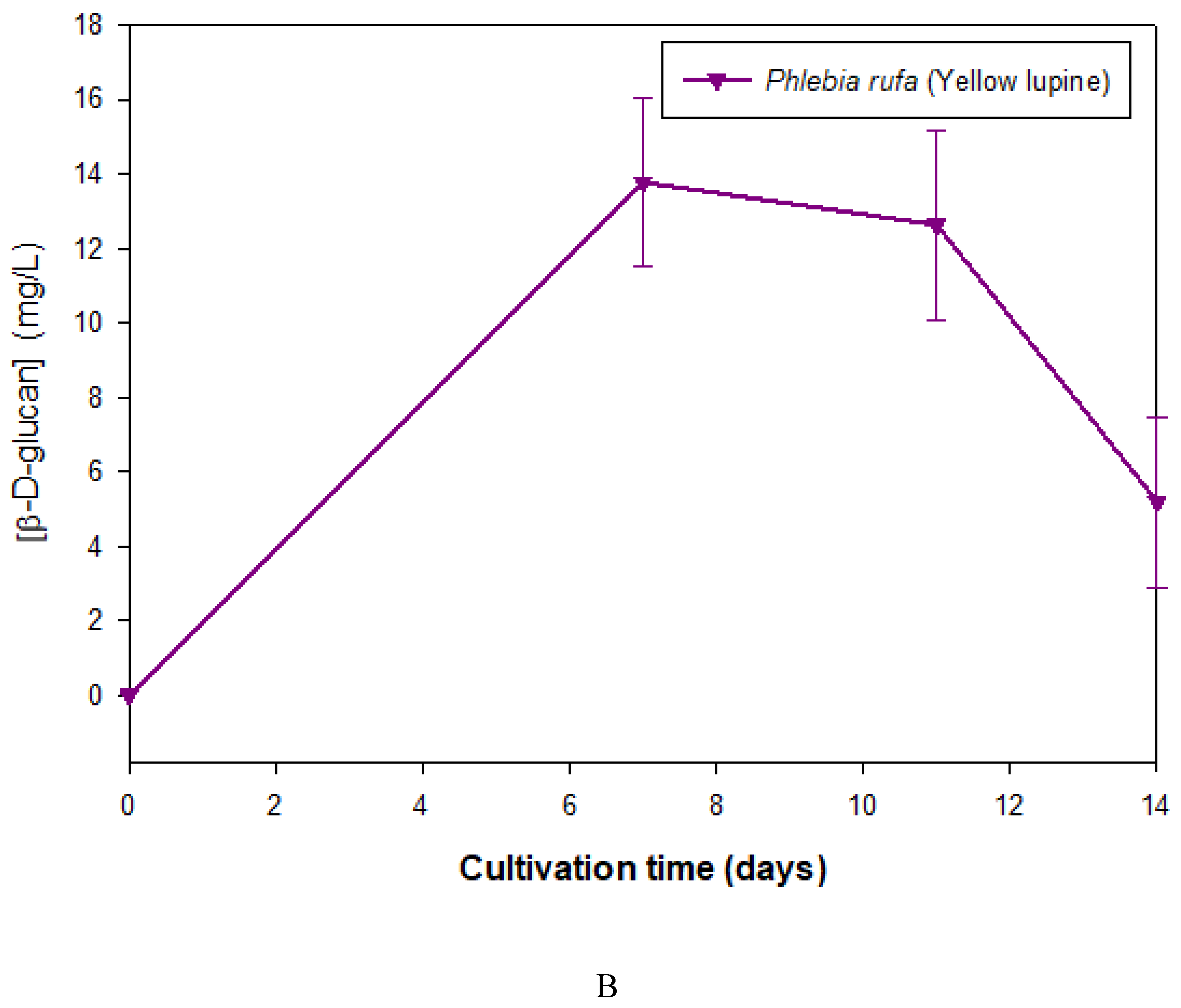
Figure 2.
Production of extracelular β-D-glucan as a function of cultivation time in culture supernatant of Lentinula edodes grown in culture medium containing waste coffee grounds . β-D-glucan was assayed by Congo red, Alcian blue and Phenol sulphuric acid methods.
Figure 2.
Production of extracelular β-D-glucan as a function of cultivation time in culture supernatant of Lentinula edodes grown in culture medium containing waste coffee grounds . β-D-glucan was assayed by Congo red, Alcian blue and Phenol sulphuric acid methods.
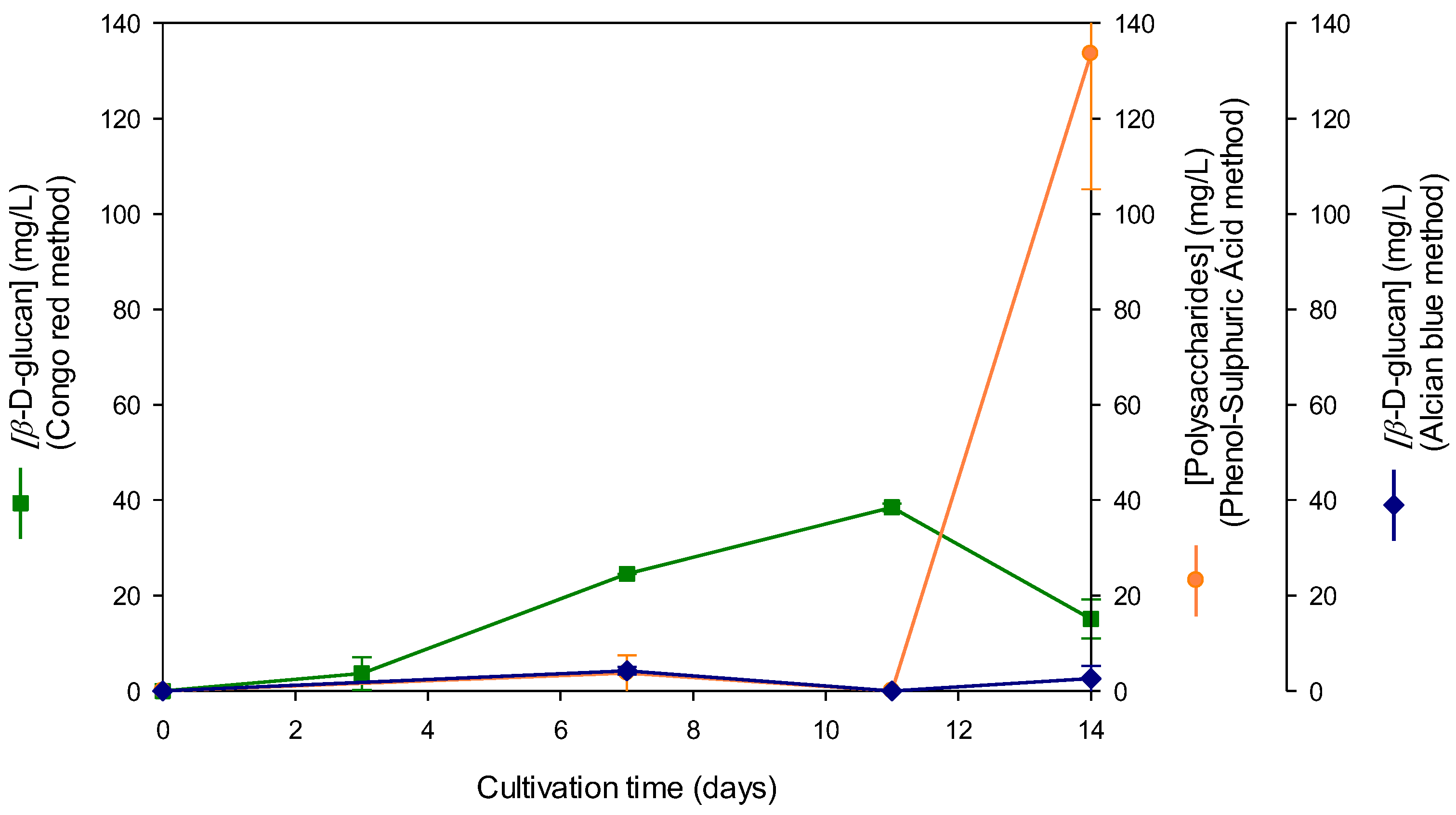
Figure 3.
A -Three-dimensional representation of the interaction of cultivation time and coffee grounds concentration on the concentration of extracellular β-D-glucans produced by Ganoderma applanatum.
Figure 3.
A -Three-dimensional representation of the interaction of cultivation time and coffee grounds concentration on the concentration of extracellular β-D-glucans produced by Ganoderma applanatum.
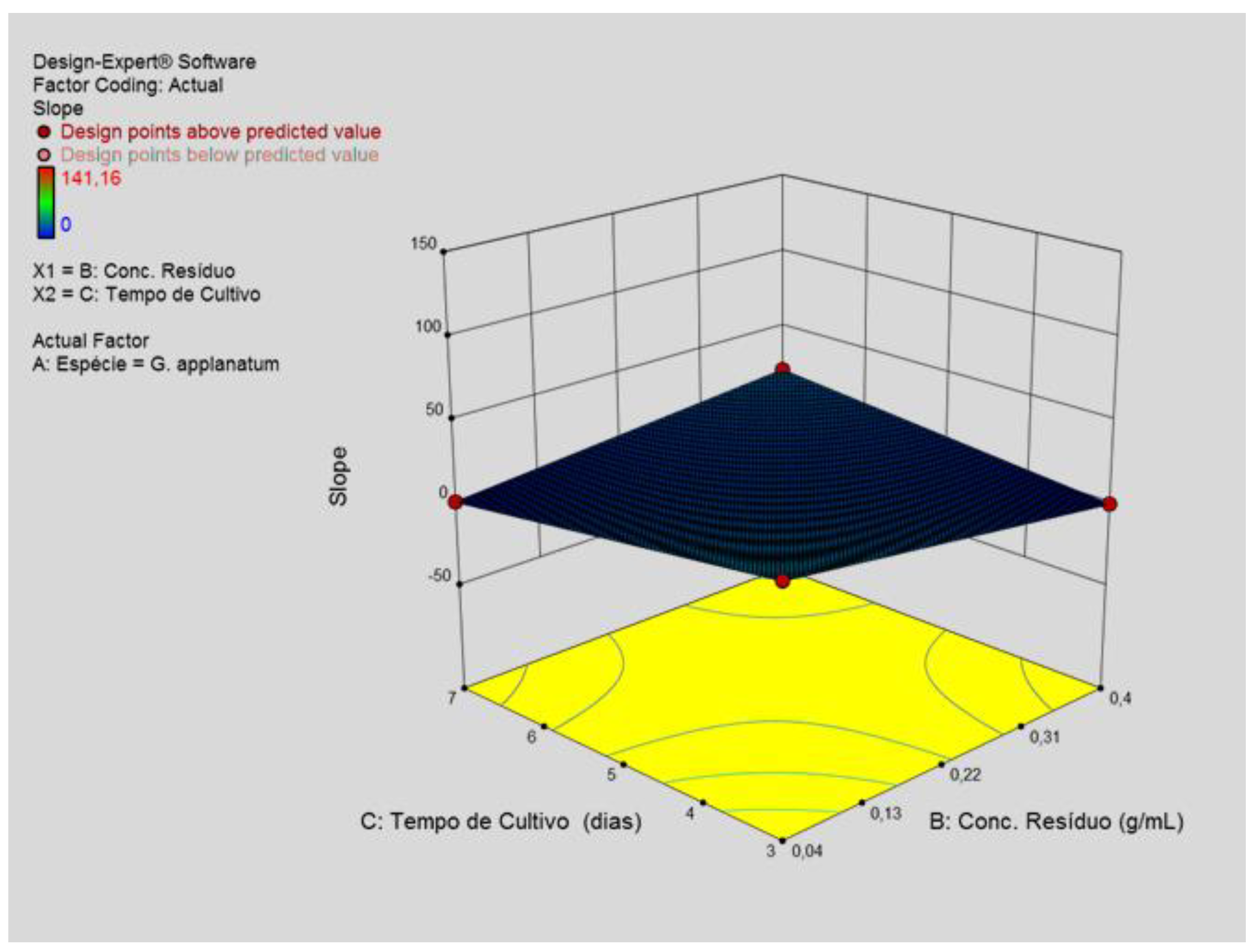
Figure 3.
B-Three-dimensional representation of the interaction of cultivation time and coffee grounds concentration on the concentration of extracellular β-D-glucans produced by Lentinula edodes.
Figure 3.
B-Three-dimensional representation of the interaction of cultivation time and coffee grounds concentration on the concentration of extracellular β-D-glucans produced by Lentinula edodes.
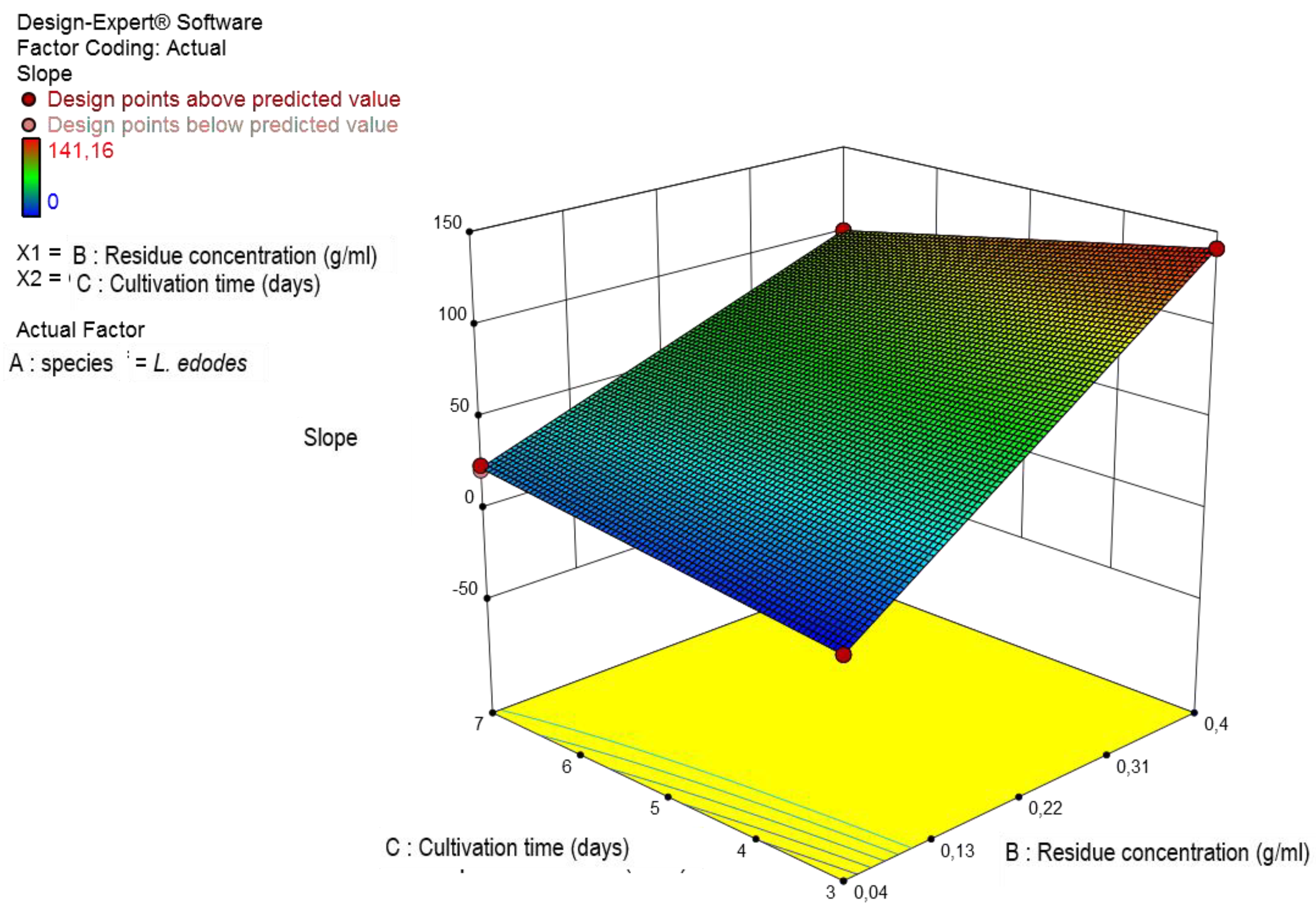
Figure 4.
A - Correlation of the fluorescence intensity of extracellular β-D-glucans from Lentinula edodes with the emission wavelength resulting from the gain change obtained in the microplate reader. The excitation wavelength was 420 nm.
Figure 4.
A - Correlation of the fluorescence intensity of extracellular β-D-glucans from Lentinula edodes with the emission wavelength resulting from the gain change obtained in the microplate reader. The excitation wavelength was 420 nm.
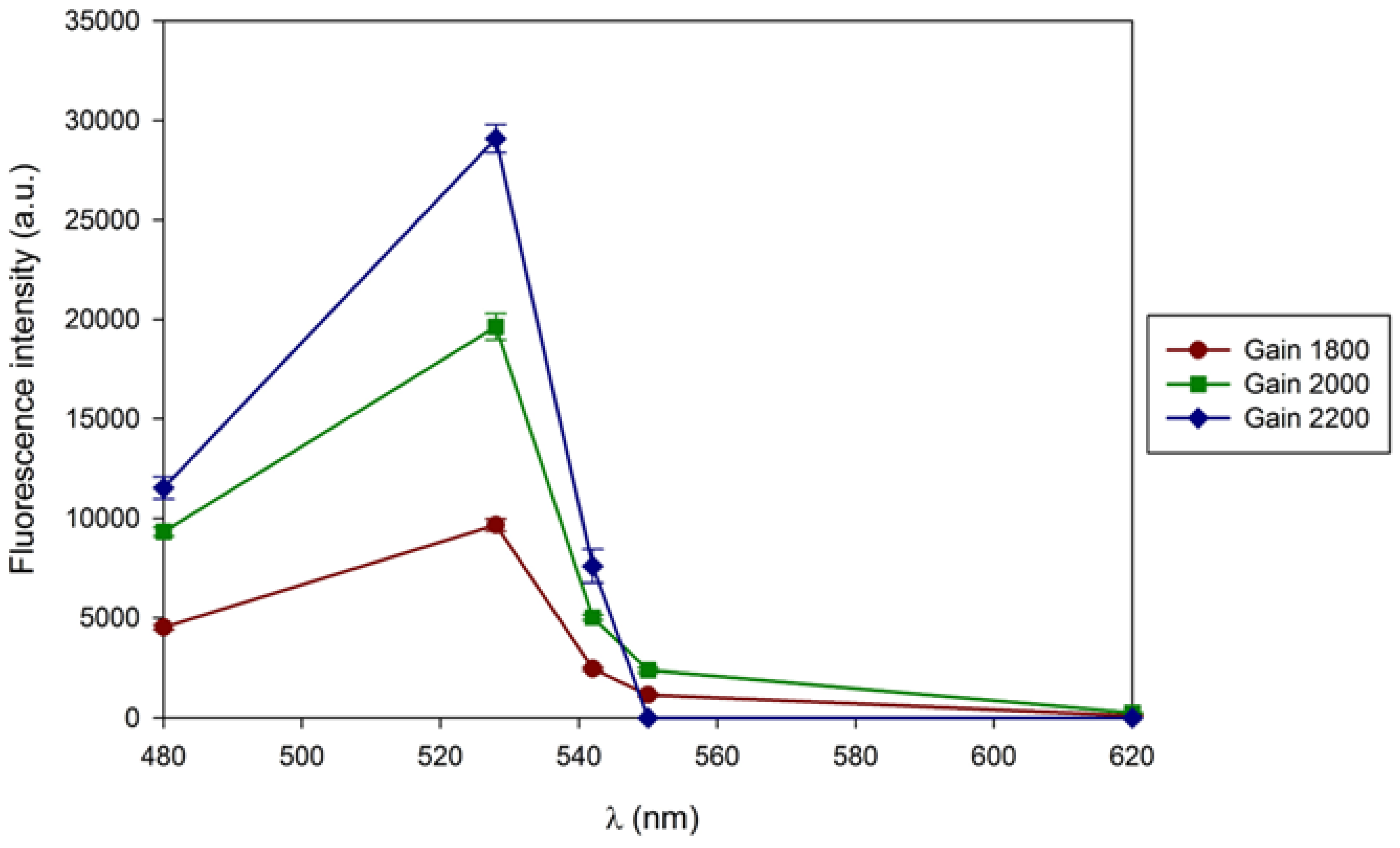
Figure 4.
B - Fluorescence intensity of extracellular β-D-glucans of Lentinula edodes. The results were obtained in a microplate reader, at different λexc and with emission filters of 480, 528, 542, 550 and 620 nm in the microplate reader.
Figure 4.
B - Fluorescence intensity of extracellular β-D-glucans of Lentinula edodes. The results were obtained in a microplate reader, at different λexc and with emission filters of 480, 528, 542, 550 and 620 nm in the microplate reader.
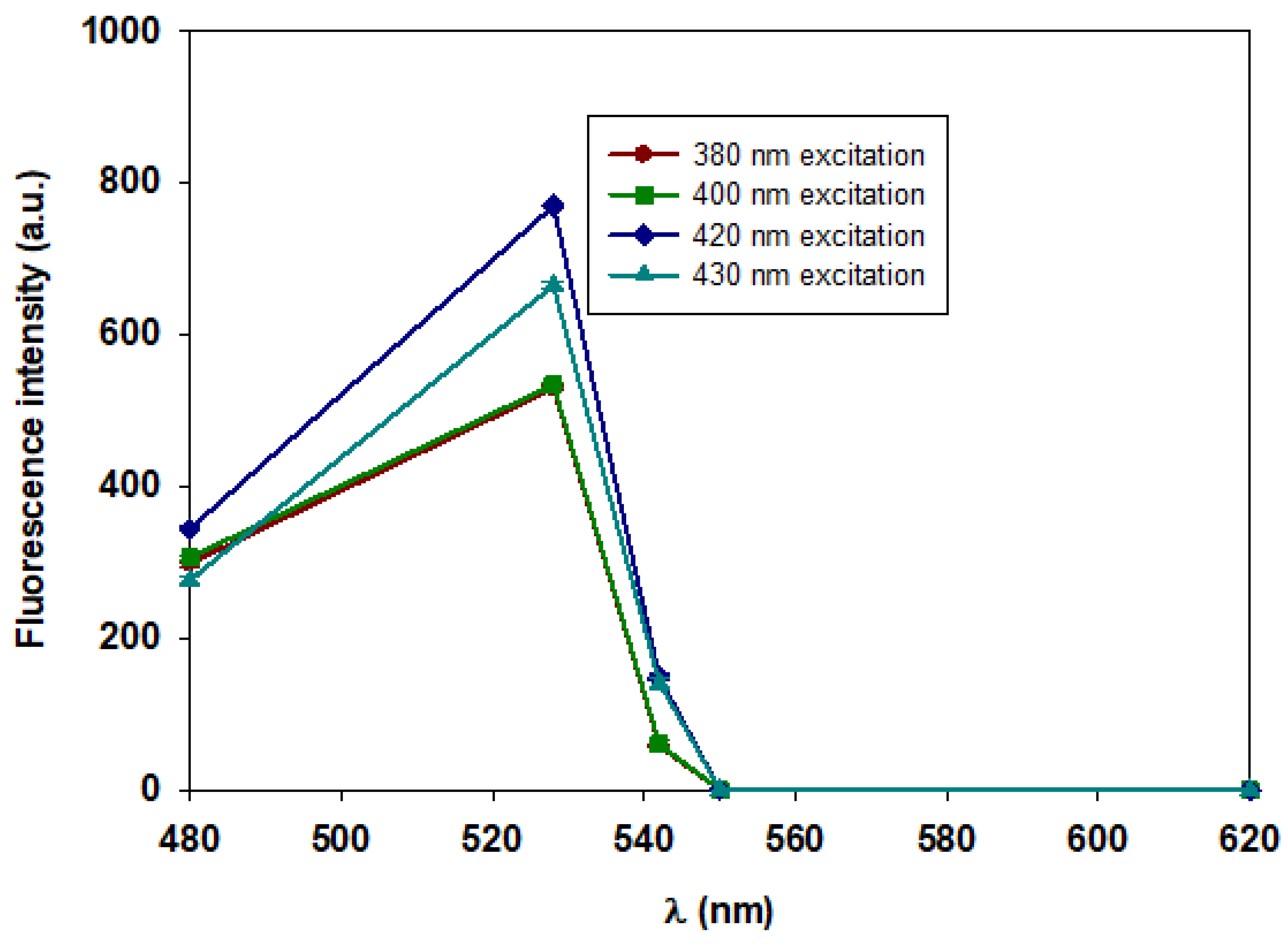
Figure 5.
A- Synchronous fluorescence spectra of commercial barley β-D-glucan with high sensitivity and Dl of 10 nm: ____ 0.25 mg ____ 0.50 mg.
Figure 5.
A- Synchronous fluorescence spectra of commercial barley β-D-glucan with high sensitivity and Dl of 10 nm: ____ 0.25 mg ____ 0.50 mg.
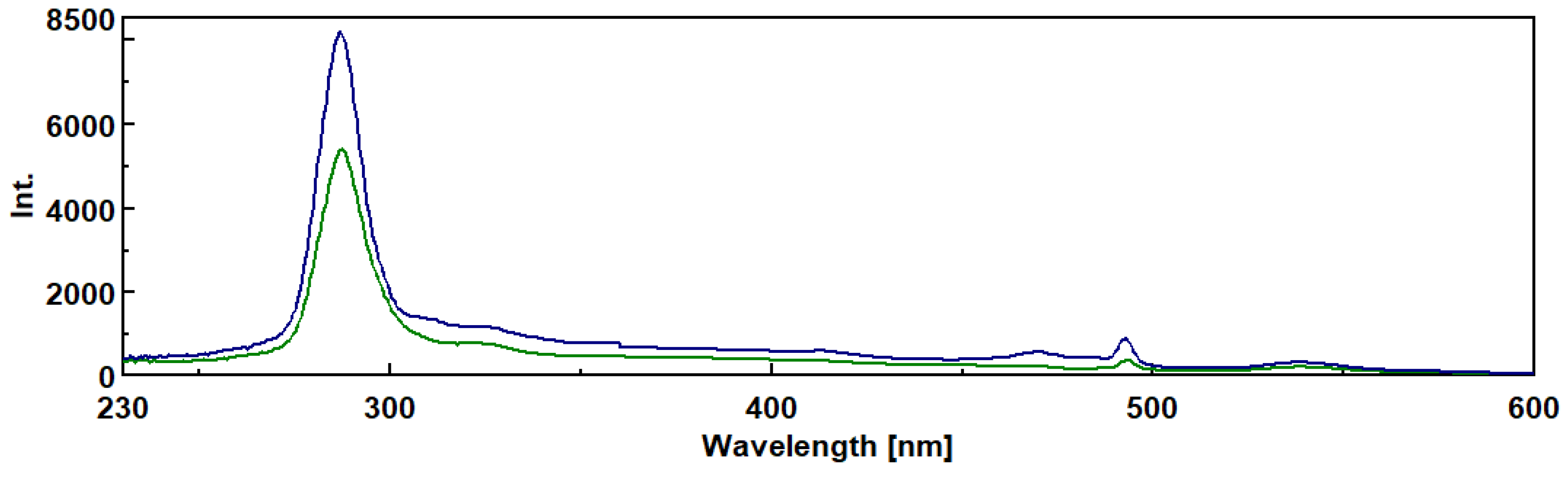
Figure 5.
B- Synchronous fluorescence spectra of β-D-glucan from medicinal mushroom fruiting bodies with high sensitivity and delta wavelength of 20 nm: ____ Auricularia auricula ; ____ Agaricus brasiliensis; ____ Cordyseps sinensis.
Figure 5.
B- Synchronous fluorescence spectra of β-D-glucan from medicinal mushroom fruiting bodies with high sensitivity and delta wavelength of 20 nm: ____ Auricularia auricula ; ____ Agaricus brasiliensis; ____ Cordyseps sinensis.
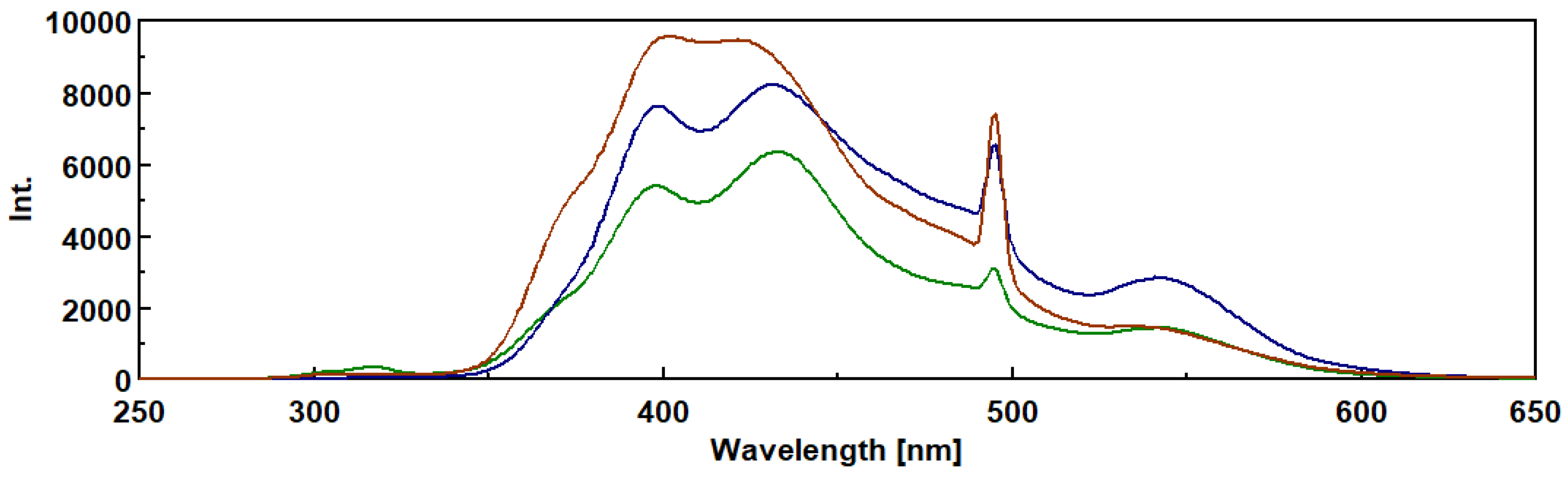
Figure 6.
A- Chromatographic behaviour of extracelular beta-D-glucans from Lentinula edodes on Sepharose 4B-BDGE-IDA-Cu(II) column at pH 8 and they were eluted with a linear gradient of imidazole in the range of 0-50 mM, followed by 50 mM and 500 mM imidazole in buffer system as described in Materials and Methods. Column fractions were analysed for fluorescence and β-D-glucans were assayed by Congo red method.
Figure 6.
A- Chromatographic behaviour of extracelular beta-D-glucans from Lentinula edodes on Sepharose 4B-BDGE-IDA-Cu(II) column at pH 8 and they were eluted with a linear gradient of imidazole in the range of 0-50 mM, followed by 50 mM and 500 mM imidazole in buffer system as described in Materials and Methods. Column fractions were analysed for fluorescence and β-D-glucans were assayed by Congo red method.
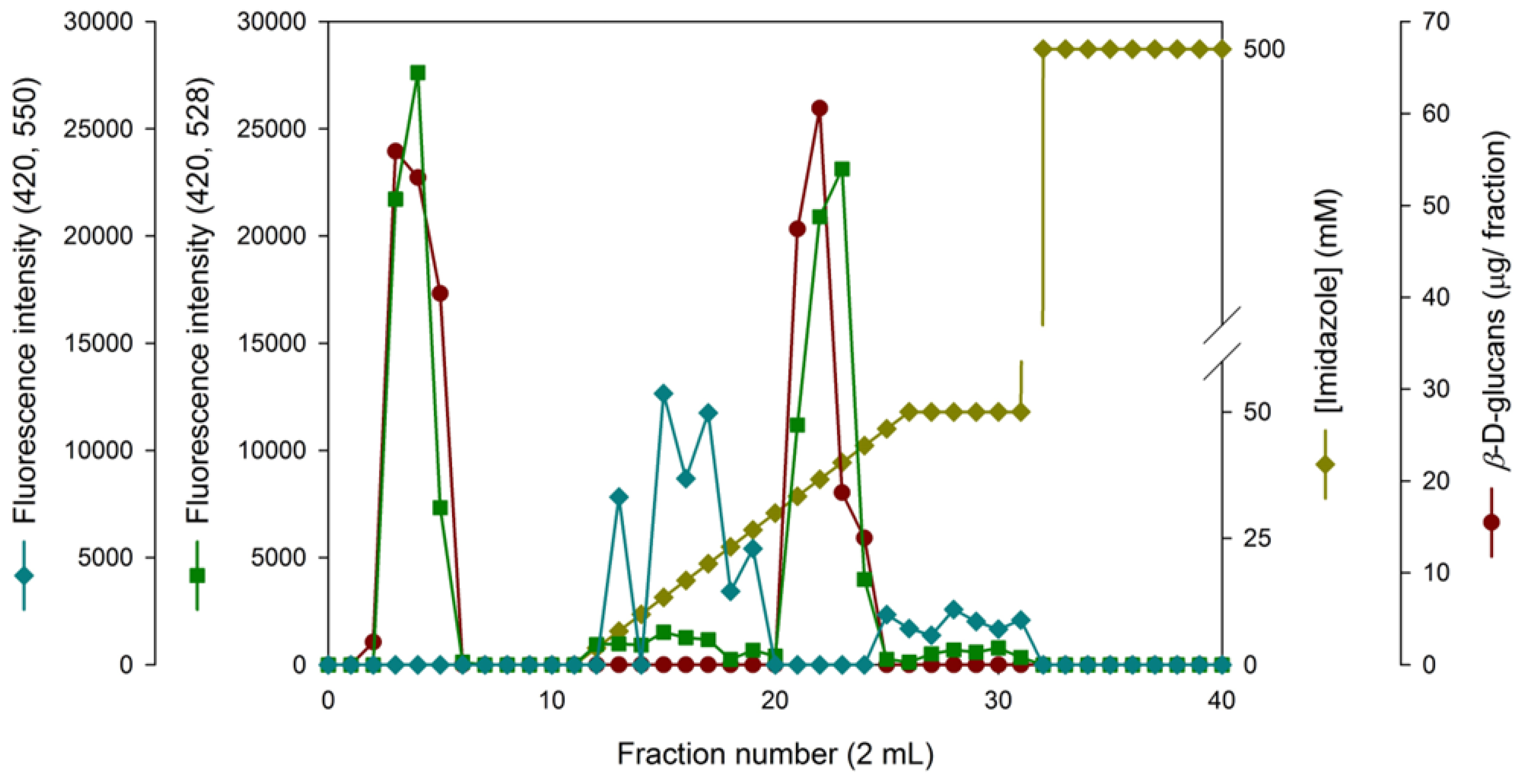
Figure 6.
B - Chromatographic behaviour of extracellular beta-D. glucans from Lentinula edodes. The sample was applied to a column packed with Sepharose 4B-BDGE-IDA-Cu(II) at pH 8 and eluted with a linear gradient of 0-50 mM imidazole, followed by 50 mM and 500 mM imidazole, in the buffer system described in Materials and Methods. The chromatographic fractions were analyzed for beta-D- glucan by Congo red dye method and proteins were monitored at A280.
Figure 6.
B - Chromatographic behaviour of extracellular beta-D. glucans from Lentinula edodes. The sample was applied to a column packed with Sepharose 4B-BDGE-IDA-Cu(II) at pH 8 and eluted with a linear gradient of 0-50 mM imidazole, followed by 50 mM and 500 mM imidazole, in the buffer system described in Materials and Methods. The chromatographic fractions were analyzed for beta-D- glucan by Congo red dye method and proteins were monitored at A280.

Table 1.
Growth of basidiomycete strains in solid culture media and their time of growth.
| Cultivation Time (days) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Agro-Industrial wastes | Oatmeal | Yellow lupine | Waste coffee ground | Banana peel | Pear peel | Pineapple peel | Mango peel | |
| Basidiomycete strains | ||||||||
| F. fomentarius | 11 | 11 | 11 | +60 | 11 | 28 | 28 | |
| G. applanatum | +60 | 56 | 9 | 12 | 12 | 12 | 12 | |
| G. carnosum | 37 | 11 | 11 | 11 | 11 | 14 | 14 | |
| G. lucidum violeta | 37 | 16 | 9 | 13 | 14 | 14 | 14 | |
| I. lacteus | 5 | 5 | 13 | 13 | 13 | 9 | 13 | |
| L. edodes | 18 | 16 | 18 | 18 | 11 | 14 | 11 | |
| P. rufa | 37 | 30 | 15 | 30 | 15 | 14 | 30 | |
| P. betulinus | 50 | 18 | 18 | 18 | +60 | 45 | 45 | |
| P. ostreatus | 37 | 31 | 23 | 18 | 18 | 28 | 28 | |
Table 2.
Assay of intracelular and extracelular β-glucans from several basidiomycete strains grown in agro-industrial residues by Congo Red method.
Table 2.
Assay of intracelular and extracelular β-glucans from several basidiomycete strains grown in agro-industrial residues by Congo Red method.
| Basidiomycete strains | Agro-Industrial Residue | β-Glucan Concentration (mg/L) | ||||
|---|---|---|---|---|---|---|
| Culture Media Filtrate | Cold Water Fraction | Hot Water Fraction | KOH Fraction | HCl Fraction |
||
| G. applanatum | Banana Peel | 0 | 0 | 0 | 0 | 0 |
| Waste Coffee Ground | 2,96x101± 5,79x10-1 | 0 | 8,04x10-1± 2,57x10-1 | 1,38± 2,82x10-1 | 3,16± 5,19x10-1 | |
| G. carnosum | Banana Peel | 0 | 0 | 1,83x10-1± 1.24x10-2 | 4,16± 1,69x10-1 | 9,76± 1,88 |
| Waste Coffee Ground | 4,26± 1,91 | 0 | 1,57± 1,31x10-2 | 1,03x101± 8,64 | 1,83x101± 1,27 | |
| L. edodes | Banana Peel | 0 | 0 | 0 | 3,46± 4,02x10-2 | 7,56± 5,19x10-1 |
| Waste Coffee Ground | 3,85x101± 4,41 | 0 | 9,14x10-1± 2,99x10-3 | 1,48x101± 2,97 | 1,56± 1,96x10-1 | |
Table 3.
ANOVA of the Factorial Design Model Proposed.
| Source | Sum of Squares | Degree of Freedom | Mean Square | F-Value | P-Value, Prob>F |
|---|---|---|---|---|---|
| Model | 3,87x104 | 7 | 5,53x103 | 1,34x104 | <0,0001 |
| A- Species | 1,13x104 | 1 | 1,13x104 | 2,74x104 | <0,0001 |
| B- Residue Concentration | 1,10x104 | 1 | 1,10x104 | 2,66x104 | <0,0001 |
| C- Time of Cultivation | 1,41x102 | 1 | 1,41x102 | 3,41x102 | <0,0001 |
| AB | 1,30x104 | 1 | 1,30x104 | 3,16x104 | <0,0001 |
| AC | 2,56x101 | 1 | 2,56x101 | 6,21x101 | <0,0001 |
| BC | 4,40x101 | 1 | 4,40x101 | 1,07x102 | <0,0001 |
| ABC | 3,23x103 | 1 | 3,23x103 | 7,83x103 | <0,0001 |
| Residual | 3,30 | 8 | 4,10x10-1 | ||
| Corrected Total | 3,87x104 | 15 |
R2= 0,9999; CV= 1,62 %; Adequate Precisio n= 310,74; Contribution of A= 29,17 %; Contribution of B= 28,31 %; and Contribution of C= 0,36 %.
Table 4.
Chromatographic behavior of extracelular -glucans from Lentinula edodes on immobilized metal chelates.
Table 4.
Chromatographic behavior of extracelular -glucans from Lentinula edodes on immobilized metal chelates.
| Stationary phase | pH | Cu(II)-IDA | Ni(II)-IDA | Co(II)-IDA | Zn(II)-IDA |
|---|---|---|---|---|---|
| Sepharose 6B- BDGE (S1) | 8 | ± | - | - | ± |
| Sepharose 6B- EPI (S2) | 8 | - | - | - | ± |
| Sepharose 4B- BDGE (S3) | 8 | ±/- | - | - | ± |
| Sepharose 4B- EPI (S4) | 8 | + | ± | - | ± |
| Stationary phase | pH | Cu(II)-NTA | Ni(II)-NTA | Zn(II)-NTA | Co(II)-NTA |
| Sepharose 6B- BDGE (S5) | 8 | - | - | - | - |
| Sepharose 6B- EPI (S6) | 8 | ± | - | - | - |
(+) represents adsorption of β-D-glucans in the stationary phase. (±) represents partial adsorption of β-D-glucans in the stationary phase. (-) represents a non-adsorption of β-D-glucans in the stationary phase since the total amount of β-D-glucans was recovered in four fractions at washing step. Lentinula. edodes was grown in a culture medium containing waste coffee ground and β-D-glucans were assayed by Congo red method and chromatographic runs were carried out as described in Materials and Methods.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



