Submitted:
12 October 2025
Posted:
13 October 2025
You are already at the latest version
Abstract
Organic agriculture is at a crossroads, facing the dual challenge of meeting rising consumer demand while enhancing productivity and resilience in the absence of synthetic inputs. For decades, vegetable breeding for organic systems has focused on the plant genome, selecting for traits like pest resistance and nutrient efficiency. While successful, this plant-centric approach overlooks a critical component of agroecological fitness: the phytobiome. Plants are not sterile entities; they are complex holobionts, hosting a vast community of microbes that profoundly influence their health, nutrition, and stress tolerance. This perspective argues for a paradigm shift in organic vegetable breeding—from a focus on the plant alone to a holistic, holobiont-aware approach. We posit that the next leap in organic productivity will come from breeding vegetable varieties that actively recruit and sustain beneficial microbial partnerships. This article deconstructs the unique microbial landscape of organic farms, outlines the key functions the phytobiome provides—from nutrient mobilization to induced systemic resistance—and proposes an integrated framework for breeding "smarter," microbially-attuned vegetables. By merging cutting-edge 'omics' technologies with farmer-centric participatory research, we can design future-proof organic vegetable systems that are not just high-yielding, but also self-regulating, resource-efficient, and truly sustainable. This requires a deep understanding of the holobiont, plant-microbe interactions, root exudates, and the integration of participatory breeding strategies for sustainable intensification.
Keywords:
plant breeding
; genomics
; holobiont breeding
; phytobiome
; participatory plant breeding
1. Introduction: A Paradigm Shift for Organic Breeding
The global appetite for organic produce has surged, driven by consumer demand for healthy food and environmental stewardship, transforming organic farming into a significant agricultural sector with worldwide organic farmland reaching 74.9 million hectares (Willer et al., 2022). However, this growth has exposed a fundamental challenge: organic systems often exhibit a yield gap compared to their conventional counterparts (Seufert et al., 2012; Ponisio et al., 2015), a gap that requires innovative solutions to close. The need for crop varieties specifically adapted to organic principles of health, ecology, fairness, and care is therefore paramount (IFOAM, 2005; Wolfe et al., 2008). For years, the response has been to breed vegetable varieties that can thrive without synthetic inputs, focusing on traits like robust root systems and innate genetic resistance (Murphy et al., 2007).
This conventional breeding approach, however, treats the plant as an autonomous organism. This perspective is fundamentally incomplete, as plants in all environments are complex superorganisms or 'holobionts'—a cohesive unit of the host plant and its associated microbiome (Vandenkoornhuyse et al., 2015). The hologenome concept posits that this collective entity, not the plant alone, is the true unit of selection in evolution (Rosenberg and Zilber-Rosenberg, 2018; Simon et al., 2019). This phytobiome, encompassing a vast community of microorganisms, acts as the plant's "second genome," performing functions the plant cannot (Turner et al., 2013; Bulgarelli et al., 2013). In conventional agriculture, many of these microbial functions are suppressed or replaced by chemical inputs (Hacquard et al., 2015), but in organic farming, they are essential for success (Berendsen et al., 2012). The very foundation of an organic system depends on this hidden world of plant-microbe partnerships (van der Heijden and Hartmann, 2016). We argue, therefore, that the current paradigm of organic vegetable breeding is reaching its limit. To unlock the full potential of organic agriculture, we must learn to breed plants that are not just genetically superior, but are also master orchestrators of their microbial allies.
2. The Organic Agroecosystem as a Unique Selective Environment
Organic farming principles inadvertently create the ideal conditions for a thriving and functional phytobiome. The prohibition of synthetic inputs fundamentally alters the soil environment, while practices like compost application and cover crops build soil organic matter, serving as a rich energy source for microbial life (Lori et al., 2017). Meta-analyses have confirmed that organic farming enhances soil microbial abundance and activity (Mäder et al., 2002) and often supports greater biodiversity compared to conventional systems (Tuck et al., 2014; Pimentel et al., 2005). This biologically-driven environment exerts strong selective pressure on plants. A variety bred for high-input systems may fail under organic conditions because it lacks the ability to engage with the soil microbiome for its needs (Richardson et al., 2009). Nutrient scarcity forces plants to "forage" for resources, a task greatly aided by phosphate-solubilizing bacteria and arbuscular mycorrhizal fungi (AMF). Without chemical pesticides, plants are more exposed to pathogens, making a robust, defensive microbiome and suppressive soils essential for survival (Weller et al., 2002). Furthermore, healthy soils rich in microbial life possess better structure, improving water retention and buffering plants against drought stress—a key climate resilience trait (Wright et al., 2005; Compant et al., 2010). The fitness of a plant in an organic system is thus inextricably linked to its ability to form beneficial symbioses (Lammerts van Bueren et al., 2011). The "ideal" organic vegetable is a systems-level phenotype that includes the plant's capacity to recruit and benefit from its microbial partners.
3. The Phytobiome: An Untapped Resource for Trait Improvement
Breeding for an optimized holobiont requires understanding which microbial functions are most critical. These microbial communities provide a suite of benefits that can be targeted as indirect breeding goals (Figure 1). A primary area is enhanced nutrient acquisition. The phytobiome directly addresses nutrient limitation, with breeding targeting plants that excel at partnerships with key microbial groups, including free-living nitrogen-fixing bacteria that can supply nitrogen to non-legumes (Kuan et al., 2016), and arbuscular mycorrhizal fungi, whose vast hyphal networks are extensions of the root system for phosphorus and water uptake (Smith and Read, 2008; Bonfante and Genre, 2010). This is complemented by phosphate-solubilizing microbes that unlock bound phosphorus in the soil (Alori et al., 2017).
Beyond nutrition, the microbiome provides multi-layered biotic stress resistance. Holobiont-aware breeding can enhance this "living shield" by selecting for plants that efficiently partner with competitive antagonists (Lugtenberg and Kamilova, 2009; Vinale et al., 2008). Many beneficial partners, such as Pseudomonas, Bacillus, and Trichoderma species, produce a wide array of antimicrobial compounds (Raaijmakers et al., 2002). Perhaps most elegantly, the presence of certain microbes can trigger Induced Systemic Resistance (ISR), priming the plant's immune system for a faster, stronger response to attack (Pieterse et al., 2014; Zamioudis and Pieterse, 2012). The phytobiome also confers abiotic stress tolerance. As climate change intensifies, microbes that produce stress-mitigating compounds become invaluable (Vurukonda et al., 2016). Bacteria containing the enzyme ACC deaminase can lower plant stress ethylene levels, promoting growth under drought and salinity (Glick, 2014). Finally, these interactions can improve crop quality, influencing nutritional content, flavor, and shelf-life (Bitas et al., 2013), analogous to the microbial "terroir" that defines wine quality (Gilbert et al., 2014).
4. Root Exudates: The Language of Plant-Microbe Communication
The primary mechanism through which plants actively shape their rhizosphere microbiome is the secretion of root exudates. This complex cocktail of sugars, amino acids, and secondary metabolites is a deliberate strategy to attract and nurture specific microbial partners (Sasse et al., 2018). It is now understood that different plant genotypes release distinct exudate profiles, creating a chemical fingerprint that sends a "call for partners" into the soil (Zhalnina et al., 2018). Beneficial microbes have evolved to recognize and utilize these specific compounds as food sources, leading to their proliferation near the root (Jacoby et al., 2017). This chemical dialogue is the foundation of the plant-microbe symbiosis and represents a powerful, yet largely untapped, target for plant breeding. By selecting for vegetable varieties that produce exudate profiles optimized to recruit elite nutrient-solubilizing or disease-suppressing microbes, breeders can indirectly select for a superior holobiont. This moves beyond breeding for the plant in isolation and begins to breed for the partnership itself, targeting the very language that establishes these crucial alliances.
5. A New Breeding Paradigm: Integrating 'Omics' with On-Farm Expertise
Breeding for a plant and its microbiome requires a strategy that represents a significant evolution from previous approaches (Figure 2). This requires an integrated, cyclical framework that merges high-throughput technology with farmer-centric validation (Figure 3). The most feasible approach is indirect selection: breeding a better host. By using tools like metabolomics to analyze exudate profiles, breeders can select for plants that secrete compounds favored by beneficial partners. This process is accelerated by modern 'omics' technologies (Knight et al., 2018). Metagenomics allows for the rapid profiling of the microbial community (Mueller and Sachs, 2015), while Genome-Wide Association Studies (GWAS) can pinpoint the plant genes that control the recruitment of key microbes (Wallace et al., 2018).
While powerful, these lab-based tools cannot replace real-world validation. Participatory Plant Breeding (PPB), where farmers and researchers collaborate, is a scientifically robust approach to selecting for complex, system-level traits (Ceccarelli et al., 2007; Dawson et al., 2008). Farmers' holistic selection implicitly accounts for the performance of the entire holobiont, capturing the complex Genotype x Environment x Microbiome (G x E x M) interactions that lab experiments often miss (Busby et al., 2017). Integrating farmer knowledge with 'omics' data creates a powerful feedback loop: on-farm performance guides lab-based discovery, which then informs the next cycle of on-farm breeding and selection.
6. Challenges and Future Directions
The path to breeding for the phytobiome is not without significant hurdles. The interactions within the microbial community are dizzyingly complex and highly sensitive to environmental factors (Toju et al., 2018). Developing the sophisticated bioinformatics and statistical models required to analyze G x E x M interactions will demand significant interdisciplinary collaboration. A further challenge is ensuring that the technologies and elite varieties developed are accessible to small- and medium-scale organic farmers, aligning with principles of food sovereignty (Kloppenburg, 2010). Another area requiring intense research is the development of microbial inoculants or synthetic communities (SynComs) (Liu et al., 2019). While promising, ensuring the survival and efficacy of these inoculants in competitive native soil environments remains a major bottleneck (Kaminsky et al., 2019). Future work must focus on developing robust, locally-adapted microbial solutions and the plant genotypes best equipped to host them (Finkel et al., 2017).
7. Conclusion: Co-Evolving Crops for a Sustainable Future
The principles of organic agriculture have always implicitly relied on the health of the soil ecosystem. It is time for our breeding strategies to explicitly embrace this reality. The shift from a plant-centric to a holobiont-centric breeding paradigm represents a convergence of ecology, microbiology, and genetics (O'Malley, 2015), moving us towards a systems-based understanding of crop performance essential for sustainable intensification (Pretty, 2018). Despite the challenges, the potential rewards are transformative (Bakshi et al., 2022; Lozano et al., 2020). This "back-to-nature" breeding approach is not science fiction (Palmgren et al., 2015); it is the logical next step in our co-evolutionary journey with the plants that feed us. By breeding for the holobiont, we can create organic vegetable systems that are more productive, resilient, and better aligned with the ecological principles on which they were founded, contributing to a more sustainable and food-secure future.
References
- Alori, E. T., Glick, B. R., & Babalola, O. O. (2017). Microbial phosphorus solubilization and its potential for use in sustainable agriculture. Frontiers in Microbiology, 8, 971. [CrossRef]
- Bakshi, A., Trakunjae, C., & Tisa, L. S. (2022). The plant holobiont: the next-generation of agricultural biotechnology. Biotechnology Advances, 54, 107871.
- Berendsen, R. L., Pieterse, C. M., & Bakker, P. A. (2012). The rhizosphere microbiome and plant health. Trends in Plant Science, 17(8), 478-486. [CrossRef]
- Bitas, V., Kim, H. S., Chelius, M. K., & Vivanco, J. M. (2013). The rhizosphere microbiome and its potential to enhance plant resistance to pathogens. Plant Disease, 97(3), 292-303.
- Bonfante, P., & Genre, A. (2010). Mechanisms underlying beneficial plant–fungus interactions in mycorrhizal symbiosis. Nature Communications, 1(1), 48. [CrossRef]
- Bulgarelli, D., Schlaeppi, K., Spaepen, S., van Themaat, E. V. L., & Schulze-Lefert, P. (2013). Structure and functions of the bacterial root microbiota. Annual Review of Plant Biology, 64, 325-353.
- Busby, P. E., Svedin, E., Bradford, M. A., Craine, J. M., & Peay, K. G. (2017). Tackling the G x E x M: the importance of the microbiome in breeding for crop resilience. Plant Science, 262, 117-123.
- Ceccarelli, S., Grando, S., & Baum, M. (2007). Participatory plant breeding in water-limited environments. Experimental Agriculture, 43(4), 411-435. [CrossRef]
- Compant, S., van der Heijden, M. G., & Sessitsch, A. (2010). Climate change effects on beneficial plant–microorganism interactions. FEMS Microbiology Ecology, 73(2), 197-214. [CrossRef]
- Dawson, J. C., Murphy, K. M., & Jones, S. S. (2008). Decentralized selection and participatory approaches in plant breeding for low-input systems. Euphytica, 160(2), 143-154. [CrossRef]
- Finkel, O. M., Castrillo, G., Paredes, S. H., Salas González, I., & Dangl, J. L. (2017). Understanding and exploiting plant beneficial microbes. Current Opinion in Plant Biology, 38, 155-163. [CrossRef]
- Gilbert, J. A., van der Lelie, D., & Zarraonaindia, I. (2014). Microbial terroir for wine grapes. Proceedings of the National Academy of Sciences, 111(1), 5-6. [CrossRef]
- Glick, B. R. (2014). Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiological Research, 169(1), 30-39. [CrossRef]
- Hacquard, S., Garrido-Oter, R., González, A., Spaepen, S., Ackermann, G., Lebeis, S., ... & Schulze-Lefert, P. (2015). Microbiota and host nutrition across plant and animal kingdoms. Cell Host & Microbe, 17(5), 603-616. [CrossRef]
- IFOAM. (2005). The IFOAM Norms for Organic Production and Processing. International Federation of Organic Agriculture Movements.
- Jacoby, R., Peukert, M., Succurro, A., Koprivova, A., & Kopriva, S. (2017). The role of soil microorganisms in plant mineral nutrition—current knowledge and future directions. Frontiers in Plant Science, 8, 1617. [CrossRef]
- Kaminsky, L. M., Trexler, R. V., Malik, A. A., Hockett, K. L., & Bell, T. H. (2019). The inherent conflicts in developing soil microbial inoculants. Trends in Biotechnology, 37(2), 140-151. [CrossRef]
- Kloppenburg, J. R. (2010). Seed sovereignty: the promise of open source biology. Wageningen University, Wageningen.
- Knight, R., Vrbanac, A., Taylor, B. C., Aksenov, A., Callewaert, C., Debelius, J., ... & Dorrestein, P. C. (2018). Best practices for analysing microbiomes. Nature Reviews Microbiology, 16(7), 410-422. [CrossRef]
- Kuan, K. B., Othman, R., Abdul Rahim, K., & Shamsuddin, Z. H. (2016). Plant growth-promoting rhizobacteria inoculation to enhance vegetative growth, nitrogen fixation and nitrogen remobilisation of maize under greenhouse conditions. PloS one, 11(3), e0152478. [CrossRef]
- Lammerts van Bueren, E. T., Jones, S. S., Tamm, L., Murphy, K. M., Myers, J. R., Leifert, C., & Messmer, M. M. (2011). The need to breed crop varieties suitable for organic farming, using wheat, tomato and broccoli as examples: a review. NJAS-Wageningen Journal of Life Sciences, 58(3-4), 193-205. [CrossRef]
- Liu, Y. X., Qin, Y., & Bai, Y. (2019). Reductionist synthetic community approaches in root microbiome research. Current Opinion in Microbiology, 49, 97-102. [CrossRef]
- Lori, M., Symnaczik, S., Mäder, P., De Deyn, G., & Gattinger, A. (2017). Organic farming enhances soil microbial abundance and activity—A meta-analysis and meta-regression. PloS One, 12(7), e0180442. [CrossRef]
- Lozano, G. L., Hamblin, M., & Leff, J. W. (2020). The gut microbiome of wild animals. Science, 367(6482), 1113-1115.
- Lugtenberg, B., & Kamilova, F. (2009). Plant-growth-promoting rhizobacteria. Annual Review of Microbiology, 63, 541-556.
- Mäder, P., Fließbach, A., Dubois, D., Gunst, L., Fried, P., & Niggli, U. (2002). Soil fertility and biodiversity in organic farming. Science, 296(5573), 1694-1697. [CrossRef]
- Mueller, U. G., & Sachs, J. L. (2015). Engineering complex mutualisms: the promise of synthetic biology. Trends in Ecology & Evolution, 30(11), 637-646.
- Murphy, K. M., Campbell, K. G., Lyon, S. R., & Jones, S. S. (2007). Evidence of varietal adaptation to organic farming systems. Field Crops Research, 102(3), 172-177. [CrossRef]
- O'Malley, M. A. (2015). The hologenome concept: a new view of evolution. BioScience, 65(3), 221-222.
- Palmgren, M. G., Edenbrandt, A. K., Vedel, S. E., Andersen, M. M., Landes, X., Østerberg, J. T., ... & Ammitzbøll, H. (2015). Are we ready for back-to-nature crop breeding? Trends in Plant Science, 20(3), 155-164. [CrossRef]
- Pieterse, C. M., Zamioudis, C., Berendsen, R. L., Weller, D. M., Van Wees, S. C., & Bakker, P. A. (2014). Induced systemic resistance by beneficial microbes. Annual Review of Phytopathology, 52, 347-375. [CrossRef]
- Pimentel, D., Hepperly, P., Hanson, J., Douds, D., & Seidel, R. (2005). Environmental, energetic, and economic comparisons of organic and conventional farming systems. BioScience, 55(7), 573-582. [CrossRef]
- Ponisio, L. C., M'Gonigle, L. K., Mace, K. C., Palomino, J., de Valpine, P., & Kremen, C. (2015). Diversification practices reduce organic to conventional yield gap. Proceedings of the Royal Society B: Biological Sciences, 282(1799), 20141396. [CrossRef]
- Pretty, J. (2018). Intensification for redesigned and sustainable food systems. Science, 362(6417), eaav0294.
- Raaijmakers, J. M., Vlami, M., & de Souza, J. T. (2002). Antibiotic production by bacterial biocontrol agents. Antonie van Leeuwenhoek, 81(1-4), 537-547. [CrossRef]
- Richardson, A. E., Barea, J. M., McNeill, A. M., & Prigent-Combaret, C. (2009). Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant and Soil, 321(1-2), 305-339. [CrossRef]
- Rosenberg, E., & Zilber-Rosenberg, I. (2018). The hologenome concept of evolution, 25 years later. Microbiome, 6(1), 78.
- Sasse, J., Martinoia, E., & Northen, T. (2018). Feed your friends: do plant exudates shape the root microbiome? Trends in Plant Science, 23(1), 25-41. [CrossRef]
- Seufert, V., Ramankutty, N., & Foley, J. A. (2012). Comparing the yields of organic and conventional agriculture. Nature, 485(7397), 229-232.
- Simon, J. C., Marchesi, J. R., Mougel, C., & Selosse, M. A. (2019). Host-microbiota interactions: from holobiont theory to analysis. Microbiome, 7(1), 5. [CrossRef]
- Smith, S. E., & Read, D. J. (2008). Mycorrhizal symbiosis. Academic press.
- Toju, H., Peay, K. G., Yamamichi, M., Narisawa, K., Hiruma, K., Naito, K., ... & Kiers, E. T. (2018). 'Core microbiomes' for sustainable agroecosystems. Nature Plants, 4(5), 247-257. [CrossRef]
- Tuck, S. L., Winqvist, C., Mota, F., Ahnström, J., Turnbull, L. A., & Bengtsson, J. (2014). Land-use intensity and the effects of organic farming on biodiversity: a hierarchical meta-analysis. Journal of Applied Ecology, 51(3), 746-755. [CrossRef]
- Turner, T. R., James, E. K., & Poole, P. S. (2013). The plant microbiome. Genome Biology, 14(6), 209.
- van der Heijden, M. G., & Hartmann, M. (2016). Networking in the plant microbiome. PLoS Biology, 14(2), e1002378.
- Vandenkoornhuyse, P., Quaiser, A., Duhamel, M., Le Van, A., & Dufresne, A. (2015). The importance of the microbiome of the plant holobiont. New Phytologist, 206(4), 1196-1206. [CrossRef]
- Vinale, F., Sivasithamparam, K., Ghisalberti, E. L., Marra, R., Woo, S. L., & Lorito, M. (2008). Trichoderma–plant–pathogen interactions. Soil Biology and Biochemistry, 40(1), 1-10.
- Vurukonda, S. S. K. P., Vardharajula, S., Shrivastava, M., & SkZ, A. (2016). Enhancement of drought stress tolerance in crops by plant growth promoting rhizobacteria. Microbiological Research, 184, 13-24. [CrossRef]
- Wallace, J. G., Zhang, X., Beyene, Y., Semagn, K., Olsen, M., Prasanna, B. M., & Buckler, E. S. (2018). The future of agriculture: breeding for resilience. Trends in Genetics, 34(11), 849-852.
- Weller, D. M., Raaijmakers, J. M., Gardener, B. B. M., & Thomashow, L. S. (2002). Microbial populations responsible for specific soil suppressiveness to plant pathogens. Annual Review of Phytopathology, 40(1), 309-348.
- Willer, H., Trávníček, J., Meier, C., & Schlatter, B. (2022). The World of Organic Agriculture. Statistics and Emerging Trends 2022. Research Institute of Organic Agriculture FiBL, Frick, and IFOAM – Organics International, Bonn.
- Wolfe, M. S., Baresel, J. P., Desclaux, D., Goldringer, I., Hoad, S., Kovacs, G., ... & Lammerts van Bueren, E. T. (2008). Developments in breeding cereals for organic agriculture. Euphytica, 163(3), 323-346. [CrossRef]
- Wright, D., Scholes, J., & Read, D. (2005). The role of arbuscular mycorrhiza in the growth of a T-DNA mutant of Arabidopsis thaliana deficient in P absorption. Plant, Cell & Environment, 28(7), 879-888.
- Zamioudis, C., & Pieterse, C. M. (2012). Modulation of host immunity by beneficial microbes. Molecular Plant-Microbe Interactions, 25(2), 139-150. [CrossRef]
- Zhalnina, K., Louie, K. B., Hao, Z., Mansoori, N., da Rocha, U. N., Shi, S., ... & Northen, T. R. (2018). Dynamic root exudate chemistry and microbial substrate preferences drive patterns in rhizosphere microbial community assembly. Nature Microbiology, 3(4), 470-480. [CrossRef]
Figure 1.
The Organic Vegetable Holobiont and its Key Functions. A schematic of the vegetable phytobiome, highlighting the ecosystem services provided by microbial partners in an organic context. The plant actively shapes its rhizosphere microbiome via root exudates. In return, microbial communities provide benefits including nutrient acquisition, pathogen suppression, and stress tolerance. Holobiont-aware breeding aims to optimize these beneficial interactions.
Figure 1.
The Organic Vegetable Holobiont and its Key Functions. A schematic of the vegetable phytobiome, highlighting the ecosystem services provided by microbial partners in an organic context. The plant actively shapes its rhizosphere microbiome via root exudates. In return, microbial communities provide benefits including nutrient acquisition, pathogen suppression, and stress tolerance. Holobiont-aware breeding aims to optimize these beneficial interactions.
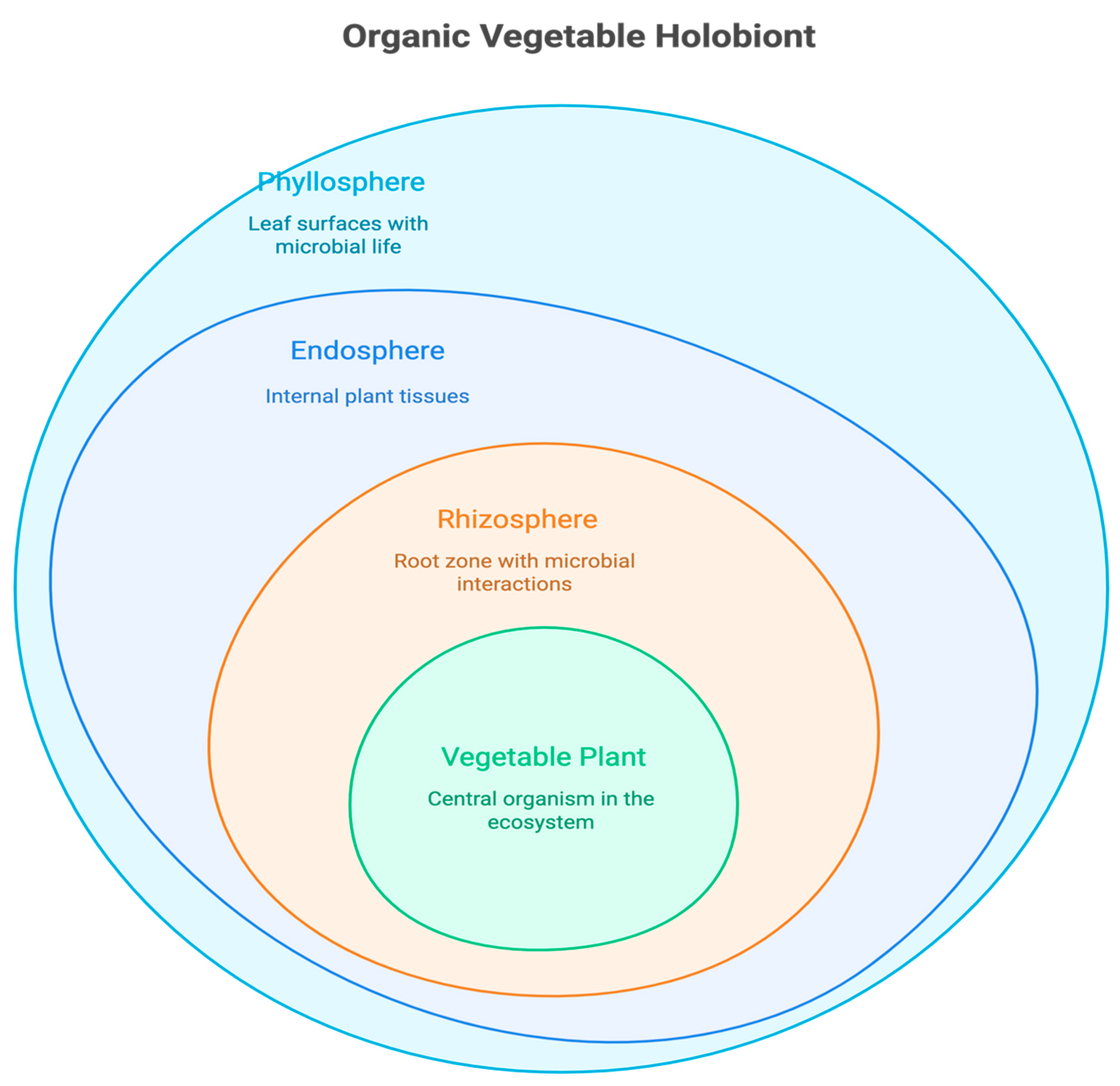
Figure 2.
The Evolution of Plant Breeding Paradigms for Organic Systems. A conceptual model illustrating the evolution of breeding strategies. (A) Classical breeding relied on phenotypic observation. (B) The molecular era integrated knowledge of the plant's genotype. (C) The proposed new paradigm, holobiont-aware breeding, treats the plant and its microbiome as a single, co-adapted unit of selection, essential for optimizing performance in complex organic agroecosystems.
Figure 2.
The Evolution of Plant Breeding Paradigms for Organic Systems. A conceptual model illustrating the evolution of breeding strategies. (A) Classical breeding relied on phenotypic observation. (B) The molecular era integrated knowledge of the plant's genotype. (C) The proposed new paradigm, holobiont-aware breeding, treats the plant and its microbiome as a single, co-adapted unit of selection, essential for optimizing performance in complex organic agroecosystems.
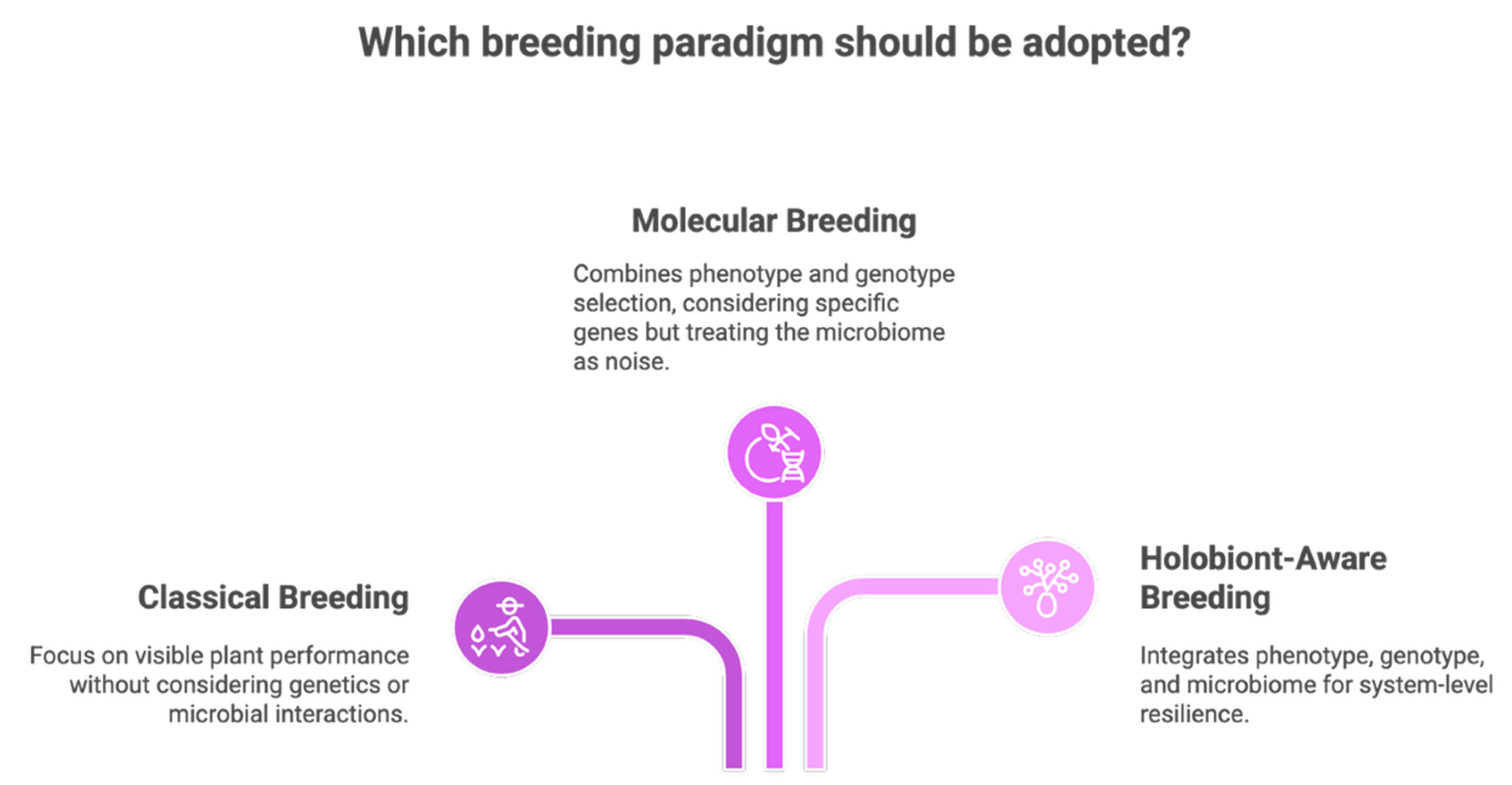
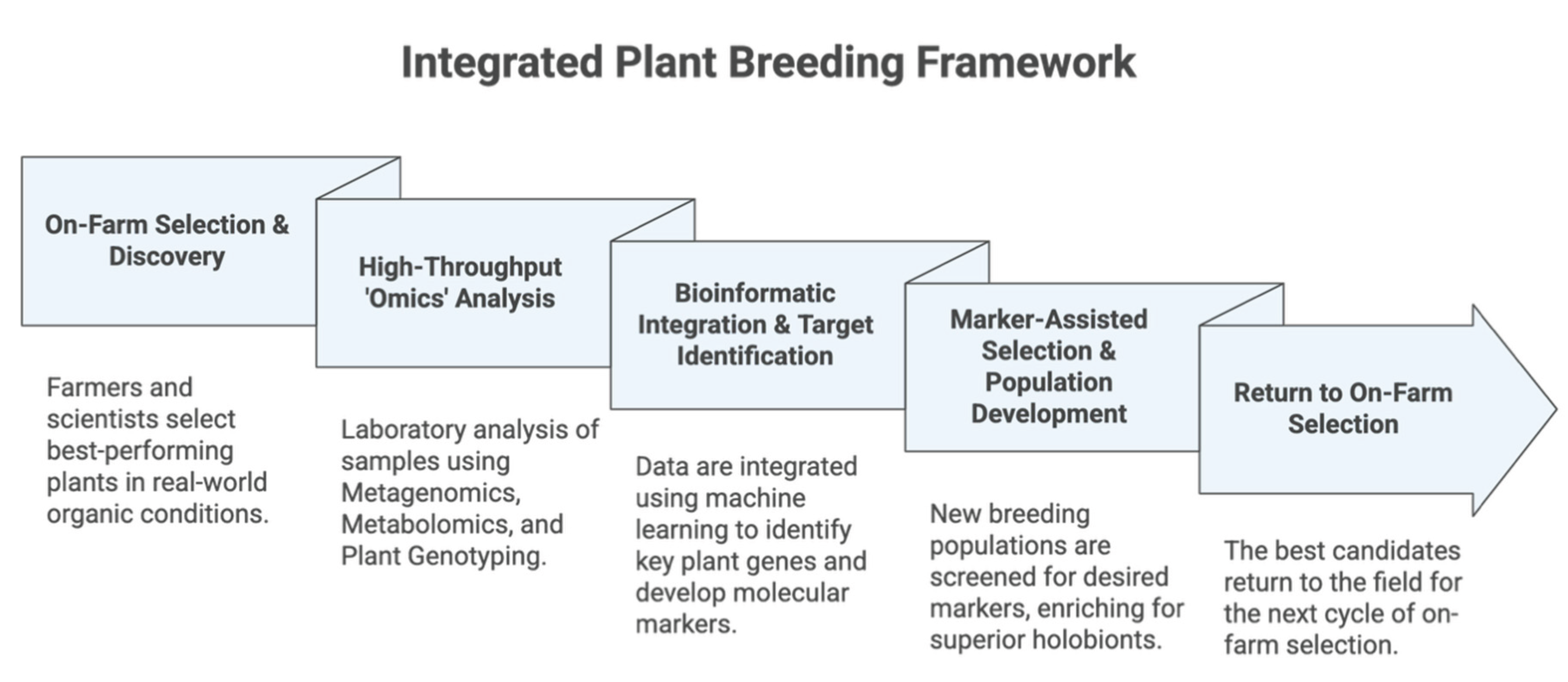
Figure 3.
An Integrated Framework for Holobiont-Aware Vegetable Breeding. A proposed framework integrating participatory plant breeding with 'omics' technologies. This cyclical process begins with on-farm selection, which grounds the program in real-world performance. High-throughput 'omics' and bioinformatics dissect the basis of successful plant-microbe partnerships, creating molecular tools to accelerate the development of organically-adapted varieties optimized at the holobiont level.
Figure 3.
An Integrated Framework for Holobiont-Aware Vegetable Breeding. A proposed framework integrating participatory plant breeding with 'omics' technologies. This cyclical process begins with on-farm selection, which grounds the program in real-world performance. High-throughput 'omics' and bioinformatics dissect the basis of successful plant-microbe partnerships, creating molecular tools to accelerate the development of organically-adapted varieties optimized at the holobiont level.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



