
Submitted:
03 September 2025
Posted:
08 September 2025
You are already at the latest version
Abstract
The transition to organic farming is one of the most desirable achievements of our time. Rational use of modern organic farming approaches not only enables the reduction of costs and increased yields but also limits the risks associated with the use of pesticides and chemicals. Despite the widest practical application of numerous biocontrol agents based on Bacillus strains, their metabolome, including the main active substances, often remains unknown. In order to understand the basic principles of the functioning of the Bacillus velezensis K-3618 strain, widely used in organic farming, we studied its spectrum of antimicrobial metabolites in detail. It was shown that the main antimicrobial agents of the B. velezensis K-3618 are representatives of the macrolactin family. Macrolactins are a group of 24-membered lactones, including macrolactin A (MLN A) and its acylated analogs 7-O-malonyl macrolactin A (mal-MLN A) and 7-O-succinyl macrolactin A (suc-MLN A), mostly active against Gram-positive bacterial pathogens, including multidrug-resistant strains. Suc-MLN A was the most potent antimicrobial, highly active (MIC = 0.1 μg/mL) against the common human pathogen methicillin-resistant Staphylococcus aureus (MRSA). It was shown that the primary mechanism of action of MLN A-based macrolactins is protein translation inhibition. Acylated macrolactins outperform MLN A in the prokaryotic cell-free system, displaying high efficiency in low micromolar concentrations. We observed that acylated MLN A analogs undergo pathogen-mediated biotransformation to MLN F analogs, having the antimicrobial activity reduced by two orders of magnitude. Hence, both acylation of MLNs and stabilization of the MLN A core are essential for the creation of new synthetic MLNs with improved antimicrobial activity and stability. While the counteraction against the observed biotransformations is essential to create new macrolactin-based antimicrobials for clinical practice, we speculate that these degradability modes are essential for bacterial ecology, and they are highly conserved in many Bacillus species from various ecological niches.
Keywords:
biocontrol agents
; Bacillus velezensis
; macrolactins
; biotransformation
; antibiotic resistance
; activity-guided metabolomics
Introduction
Phytopathogens pose a threat to the global economy and food industry, necessitating the search for effective counteraction measures. Application of organic farming practices and biocontrol agents enables the reduction of pesticide usage that not only manages health risks of pesticide exposure but also provides the opportunity to gain higher yields [1]. Species of the genus Bacillus are widely used as biological control agents acting through a synergy of mechanisms including competition, antagonism, induction of systemic resistance, and stimulation of plant growth [2,3]. Bacillus species are potential producers of antibiotics that protect plants by suppressing soil phytopathogens. While a high diversity of bactericidal secondary metabolites of various chemical classes are known for Bacillus [4], the exact determining of the most principle-acting antimicrobials in biocontrol agents is of high interest since it (i) enables a more comprehensive and targeted practical application of these strains and (ii) provides clues for understanding their impacts on microbiomes [5,6].
However, for the overwhelming majority of industrial biocontrol strains, the exact composition of principle-acting secondary metabolites was not characterized in detail. Bacillus velezensis strains are well known for their biocontrol activity [7,8] that shield plants from a wide range of bacterial and fungal diseases and provide growth benefits to tomato, pepper, pumpkin, and cucumber [3].
To uncover the landscape of antimicrobial metabolites of a highly efficient growth-stimulating endophytic isolate, Bacillus velezensis K-3618 (All-Russian Collection of Agricultural Microorganisms), which is widely applied as a biocontrol agent in organic farming, we used the antimicrobial activity-guided metabolomic approach. A family of macrolactins was isolated as the main antibacterial metabolites produced by B. velezensis K-3618.
Macrolactins (MLN) are a large group of natural compounds produced by various marine and soil microorganisms [9,10]. They belong to the class of macrolides, having a 24-membered lactone ring in their structure. Since 1989, when the first representatives of this group were described (MLN A-F) [11], their number has reached 33 by 2021 and continues to increase [12]. MLNs exhibit a wide range of pharmacological properties, with characteristic antibacterial activity for most of them. The main structural differences of MLNs include the position and number of olefinic bonds in the lactone ring and different types of post-modified groups.
7-O-malonyl macrolactin A (mal-MLN A) and 7-O-succinyl macrolactin A (suc-MLN A), which carry the 7-OH position of MLN A acylated by the respective dicarboxylic acids, are produced by Bacillus strains, including both soil and marine species [10]. Suc-MLN was originally isolated from the fermentation broth of the marine strain Bacillus sp. sc026 [13], mal-MLN A was isolated later from the soil strain B. subtilis ICBB 1582 along with MLN A and suc-MLN A [14]. The same three compounds were isolated from B. amyloliquefaciens strain NJN-6, which was used for the biological control of soil-borne plant pathogens [6].
The MLN backbone is assembled by the trans-acyltransferase PKS to give MLN A [15]. The reaction of acylation of MLN A in biosynthesis of mal-MLN A and suc-MLN A had remained largely enigmatic until recently, when a β-lactamase homologue, BmmI, was identified as responsible for this key step of biosynthesis [16]. BmmI can specifically attach C3−C5 alkyl acid thioesters to the 7-OH of MLN A and is substrate promiscuous toward acyl acceptors with different backbones. Pharmacological properties of these macrolactins are diverse [9]. Preclinical studies of MLN A and suc-MLN A as antimacular and antineoplastic agents have been reported, examining their potential metabolic interactions with other drugs by assessing cytochrome P450 inhibition and induction and UDP-glucuronosyltransferase inhibition in vitro [17]. The pharmacokinetics of MLN A and suc-MLN A were assessed as well [18,19]. MLN A and suc-MLN A also demonstrated antifungal activity against some important plant pathogens, and the presence of the succinyl moiety at C-7 gave the compound a higher activity against all the probed fungi [20].
Macrolactins turned out to be bacteriostatic antibiotics that inhibit a number of multidrug-resistant Gram-positive bacterial pathogens, with suc-MLN A generally exhibiting more pronounced antimicrobial activity than MLN A and mal-MLN A [14,21]. MIC of suc-MLN A against methicillin-resistant Staphylococcus aureus (MRSA) has been reported as < 0.25 μg/mL, and 2 μg/mL against vancomycin-resistant enterococci (VRE), while for MLN A MIC values vary between 1 μg/ml (MRSA) [21] and 10 μg/ml (MSSA) [22] to MBC 150 μg/ml (MSSA) [14,23].
Despite the obvious interest that macrolactins represent as antibiotics, the mechanisms of action of the numerous compounds are not fully understood. A significant contribution has been made by the recent study of the mechanism of action of MLN A [23]. The compound was shown to inhibit the first step of the elongation in protein synthesis, and the bacteriostatic and/or bactericidal action was found to be species and strain dependent. It was demonstrated that MLN A acts as an elfamycin-type antibiotic. Various types of additional activities were also reported for different representatives of the macrolactin family. Macrolactin N inhibits bacterial peptide deformylase with IC50 value of 7.5 uM [24]. MLN S and MLN B inhibit NADPH-dependent β-ketoacyl-ACP reductase (FabG) with IC50 value of 0.1 mM [25]. Transmission electron microscopy imaging showed impaired integrity and permeability of the cell membrane as result of antibacterial activity of new MLN XY against E. faecalis [4]. The activity of 7-O-succinyl macrolactin F as an inhibitor of iron uptake was also described [26].
In our research, a family of macrolactins was isolated from B. velezensis K-3618, and the mechanism of their antibacterial activity was determined. Biotransformation of the 7-O-malonyl macrolactin A (mal-MLN A) and 7-O-succinyl macrolactin A (suc-MLN A) macrolactins was discovered, indicating the principal role of acylation and double bond architecture for their antimicrobial activity. We speculate that antimicrobial activity is strongly conserved in B. velezensis strains of different origins, making them promising candidates for the development of biological control agents. Further advances in semisynthetic analogs with improved stability and selectivity will provide new avenues for the development of macrolactin-based therapeutics as well.
Results
Cultivation of the Strain, Isolation and Identification of the Metabolites with Antibiotic Activity
B. velezensis K-3618 exhibited prominent antibacterial activity against S. aureus and E. coli ΔlptD when cultivated in various liquid media (Table S1). The major fraction of antimicrobial activity was associated with macrolactins (Figure 1A): acylated 7-O-succinyl-macrolactin A (suc-MLN), 7-O-malonyl-macrolactin A (mal-MLN A), and a minor fraction of acyl-free macrolactin A (MLN A), which was monitored using a highly sensitive GFP-producing S. aureus reporter strain [27]. Isolation was performed by a two-step activity-guided fractionation: a solid-phase extraction and HPLC purification. The active components have two UV maxima, the first at 227 nm and the second at 258-262 nm (Figure 1B). Mass spectrometric analysis in positive ion mode revealed three major ions at m/z 441.20, 527.20, and 541.22, corresponding to the [M+K]+ adducts of macrolactin A (C24H35O5), 7-O-malonyl-macrolactin (CHO), and 7-O-succinyl-macrolactin (C28H38O8), respectively (Figure 1C–E, Figure S1-S3). All the compounds exhibited a characteristic pattern of fragment ions at m/z 367.22, corresponding to the known macrolactin A core structure following the loss of two water molecules [M - 2H₂O+H]+) and a secondary fragment at m/z 349.21, representing a further dehydration ([M - 3H₂O+H]+). This fragmentation pattern has been previously described and is considered characteristic for the macrolactin scaffold [6].
To unambiguously confirm that the identified macrolactins belong to the macrolactin A subfamily, the structures of the identified MLN A, mal-MLN A, and suc-MLN A were validated with NMR. The chemical shifts for the macrolactin scaffold of the investigated molecules (Table S2) are in line with previously published data for malonyl-macrolactin A [14]. The variability among the molecules is substantiated by the differing number of signals arising from the acyl substitutions, as well as the distinctive chemical shift values observed in the HSQC spectra (Table S2) [28]. Based on these findings, along with the characteristic HMBC correlations (Figure S4), it was possible to unambiguously assign three distinct forms.
Bacterial Susceptibility and Cytotoxicity of the Macrolactins
To characterize the antimicrobial activity of the identified macrolactins in more detail, the susceptibility of a model panel of bacteria toward the macrolactins was estimated along with their cytotoxicity (Figure 2A). Macrolactins were highly active against Gram-positive bacteria, including the common human pathogen Staphylococcus aureus (MRSA) and opportunistic pathogens like Enterococcus faecalis and Micrococcus luteus. Oppositely, macrolactins were inactive against Gram-negative bacteria, including hypersensitive Escherichia coli BW25113 ΔtolC and ΔlptD strains. That indicates that Gram-positive bacteria are primary native bacterial targets for macrolactins. Acylated macrolactins were more active than MLN A; suc-MLN A was also more active than mal-MLN A. The estimated MICs against S. aureus were 1 μg/ml, 0.25 μg/ml, and 0.1 μg/ml for MLN A, mal-MLN A, and suc-MLN A, respectively.
The direct practical application of macrolactins is complicated by their considerable cytotoxicity. The estimated IC50 values for human cell line HEK293T are 15±2 μg/mL, 2±0.3 μg/mL, and 0.3±0.05 μg/mL for MLN A, mal-MLN A, and suc-MLN A, respectively (Figure 2A). Hence, the cytotoxicity of macrolactins generally correlates with their antimicrobial activity. However, to create efficient semisynthetic agents for antimicrobial therapy, further structure-activity relationship analysis should be provided to obtain macrolactin-based analogs with improved selectivity toward bacteria.
Inhibition of Bacterial Protein Synthesis In Vitro by the Macrolactins
It was recently proposed that the MLN A mechanism of action is translation inhibition [23]. To elucidate this observation and estimate the efficacy of acylated macrolactins as translation inhibitors, they were assessed in a prokaryotic cell-free translation system. The tested compounds (MLN A, mal-MLN A, suc-MLN) exhibit concentration-dependent inhibition of protein synthesis with estimated IC50 of 1.5±0.2, 0.8±0.2, and 0.7±0.1 μg/mL for MLN A, mal-MLN A, and suc-MLN A, respectively (Figure 2B). Hence, acylated macrolactins demonstrate a more potent inhibitory activity on protein translation in the prokaryotic cell-free system that is in line with their improved antimicrobial activity. Consequently, we speculate that, similarly to MLN A, macrolactin analogs primarily act by suppression of protein synthesis since (i) macrolactins demonstrate similar concentration ranges for protein translation inhibition and antimicrobial activity, and (ii) the antimicrobial activity of macrolactins and the efficacy of protein translation inhibition of macrolactin analogs correlate with each other.
Transformation of Acylated Macrolactins in S. aureus Cells
We hypothesized that macrolactins may undergo biotransformation in living bacterial cells, which could influence their antimicrobial activity. To test this hypothesis experimentally, we incubated mal-MLN A and suc-MLN A in the presence of S. aureus cells and analyzed culture media using HPLC-UV and LC-MS/MS (Figure 3A–D). In control experiments, only slow spontaneous hydrolysis was observed, with approximate half-lives of 35 ± 8 h and 70 ± 20 h for mal-MLN A and suc-MLN A, respectively. In contrast, living S. aureus cells degraded both mal-MLN A and suc-MLN A much more rapidly. The biodegradation kinetics were 15 ± 4 and 13 ± 5 times higher than spontaneous hydrolysis for mal-MLN A and suc-MLN A, respectively (Figure 3C,D). Moreover, S. aureus cells showed a different pattern of biodegradation products compared to the spontaneous hydrolysis of acylated macrolactins (Figure 3A,B). Detailed analysis of chromatograms revealed that S. aureus cells produced additional degradation products distinct from those resulting from a spontaneous hydrolysis. For both mal-MLN A (m/z 527.21 [M+K]+) and suc-MLN A (m/z 541.22 [M+K]+), incubation with S. aureus yielded an additional peaks at the same m/z but with shifted retention time (Figure S5) and different UV spectral pattern (Figure S6A,C). Analogously, in both cases we observed a peak at m/z 441.20 ([M+K]+), corresponding to the hydrolyzed malonyl- or succinyl-MLN F (Figure S6A,B). All these ions showed MS/MS spectra identical to the respective initial macrolactins, consistent with rearranged isomers (Figures S7–S9).
Biotransformation products were isolated, and chemical structures of compounds were defined using NMR. The structure of the most abundant product of suc-MLN A transformation was determined as 7-O-succinyl-macrolactin F (suc-MLN F) (Figure S10, Table S3). Accordingly, mal-MLN A was transformed to mal-MLN F, which was validated with mass and UV spectra (Figures S5, S6, S8). A minor component with the molecular mass, UV spectrum, and retention time identical to MLN F was observed during the transformation of mal-MLN A and suc-MLN A and then validated by NMR (Figure S10, Table S3). Biotransformation of MLN A backbone to the MLN F backbone resulted in a dramatic 25- to 100-fold reduction of antimicrobial activities of all the compounds. The measured MICs against S. aureus were 25 μg/mL, 25 μg/mL, and 6 μg/mL for MLN F, mal-MLN F, and suc-MLN F, respectively.
MLN Biosynthetic Gene Cluster is Highly Abundant in B. velezensis and B. amyloliquefaciens Species
The Bacillus velezensis K-3618 strain was originally co-isolated from potato tubers of the Charoit variety in a natural biocontrol agent screening study [29] as an endophytic microorganism displaying cellulolytic, amylolytic, and weak nitrogen-fixing activity. In laboratory and field tests, B. velezensis K-3618 demonstrated high efficiency as a complex-action biofungicide, which not only provided protection of agricultural plants from diseases, but also led to an increase in their productivity.
To reveal the biosynthetic potential of B. velezensis K-3618, we sequenced its genome. The assembly of the genome resulted in 23 contigs, with a total genome length of 3,864,279 base pairs, GC-content of 46.40%, N50 value of 471,177 bp, and N90 of 163,640 bp (GenBank NZ_JBNVTP000000000.1). The corresponding L50 and L90 values were 3 and 8, respectively, indicating that a relatively small number of large contigs represent the majority of the genome sequence. CheckM [30] analysis estimated the genome completeness at 99.63%, with no detectable contamination (0%). An antiSMASH [31] analysis of the complete genome sequence of Bacillus sp. K-3618 revealed a biosynthetic gene cluster (BGC), exhibiting high similarity to the previously described macrolatin-producting clusters (MiBiG [32] entries BGC0000181 [15,33,34] and BGC0001383 [35]). The identified cluster includes a complete set of core and tailoring enzymes required for macrolactin production (Figure 4).
To estimate the potential metabolic landscape of B. velezensis K-3618 and related strains, we performed comparative genomic analysis with 30 phylogenetically closest genomes (Supplementary file, Table S4). We first reconstructed the reference ML phylogeny based on core SNPs. Intriguingly, the distribution of species attributions does not follow a taxonomy-wise phylogenetic grouping (Figure 4). More precisely, genomes attributed to B. velezensis and B. amyloliquefaciens were mixed among the phylogenetic clades. The strain K-3618 formed a two-leaf clade with another soil-dwelling isolate BS89 belonging to B. velezensis as other representatives of the wider clade, allowing us to classify K-3618 to the B. velezensis species.
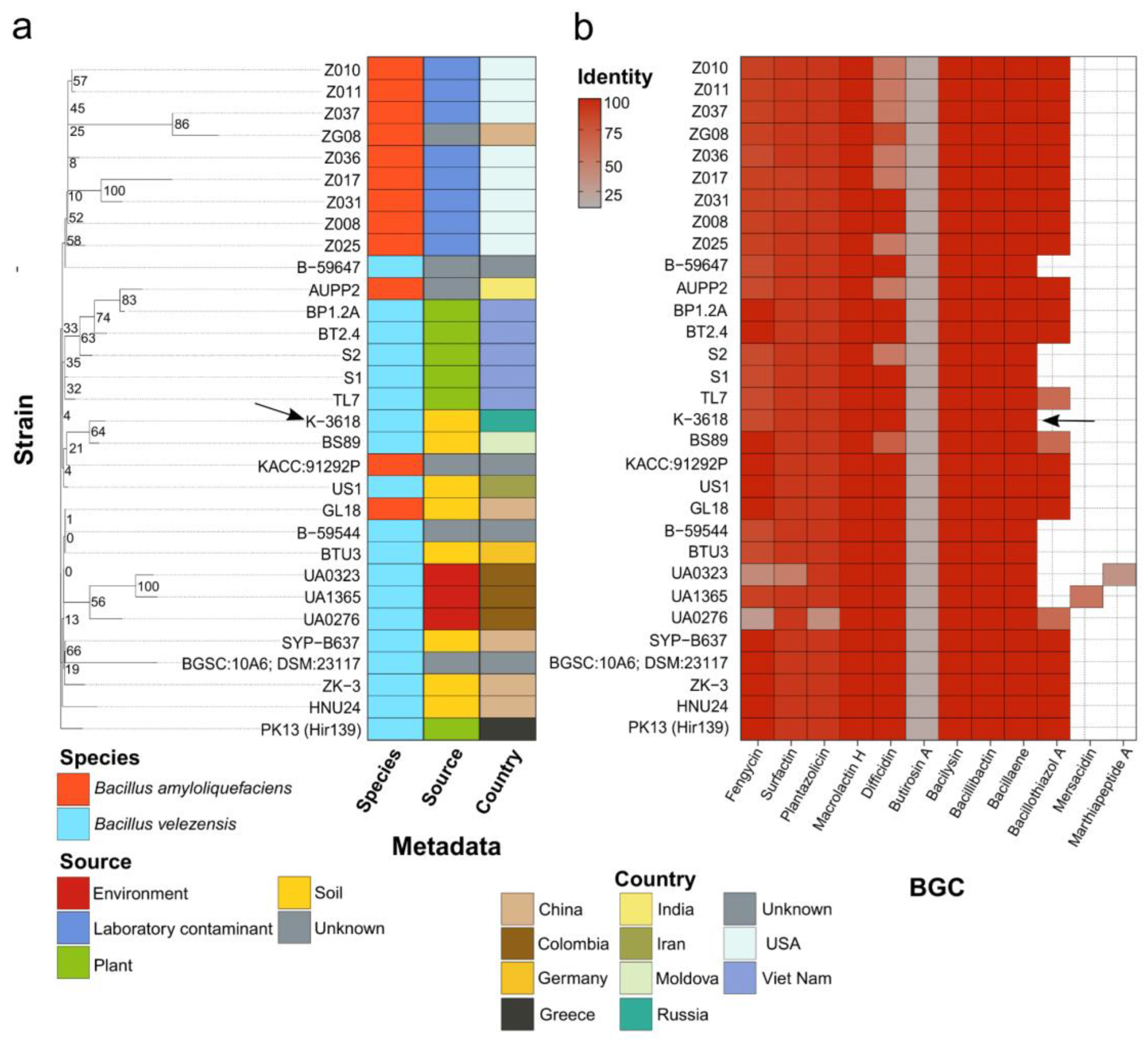
Next, we examined the distribution of known BGCs among the selected references using antiSMASH [36]. The strain K-3618 contained 9 clusters responsible for the synthesis of bacillaene, bacillibactin, bacilysin, butirosin A, difficidin, fengycin, surfactin, plantazolicin, and macrolactin H (Figure 5; Supplementary file, Table S5). These clusters were also found in all 30 reference genomes, along with bacillothiazol A detected in 22 genomes coupled with two singletons, namely, mersacidin and marthiapeptide A (Figure 5). We then mined for the core genes within the cluster of macrolactin H biosynthesis and found that all 10 core genes with 100% similarity and coverage were identified in all genomes (Supplementary file, Table S6). Hence, the observed effects of biological control of pathogenic bacteria via macrolactins potentiated with the conserved metabolic fingerprint of bacillaene, bacillibactin, and bacilysin should be strongly conserved in B. velezensis and closely related B. amyloliquefaciens strains. While B. velezensis and B. amyloliquefaciens are very close in their phylogeny, metabolic discrimination was proposed for B. velezensis based on its ability to produce macrolactin [34]. Macrolactin-producing B. amyloliquefaciens strains could be reannotated accordingly; however, most of them have another difficidin/oxydifficidin cluster. This difference may have a particular impact and impair their functionality as biocontrol and plant growth-promoting agents.
Discussion
Bacterial susceptibility to macrolactins according to literature data varies by orders of magnitude: the MIC of MLN A against S. aureus is in the range of 1-10 μg/mL [21,22], the MIC of suc-MLN A is lower than 0.25 μg/mL [21], and the value for mal-MLN A is higher than 120 μg/mL [14]. Our comparative study of three macrolactins indicates lower antibacterial activity of MLN A compared to its acylated analogs, as well as a higher activity of suc-MLN A compared with mal-MLN A that may be associated with its greater stability. We observe that the differences in the activity of these macrolactins on bacterial cells are more significant than the differences in inhibition of protein synthesis in a cell-free system. This may be due to the ability of acylated compounds to more effectively penetrate bacterial cells. A clear correlation between the minimum inhibitory concentration and toxicity was also observed, which complicates the prospects of using these compounds as antibiotic drugs.
In experiments with incubation of the antibiotics with bacterial cells, we observed two effects: dramatic acceleration of the degradation process by living S. aureus cells and formation of new metabolites that were not detected after spontaneous hydrolysis of macrolactins. We found that biodegradation of mal-MLN A and suc-MLN A occurs by rearrangement of the molecules with the formation of a keto group instead of a hydroxy group with the reduction of a double bond. Partial hydrolysis with the cleavage of the side acyl group also occurs. Both types of the modified compounds, despite having similarity to the parent compound structure, are drastically less active.
Modification of molecules during biodegradation is a common pathway for antibiotic resistance in bacteria, including staphylococci, along with such processes as enzymatic modification of the binding site, drug efflux, reduced permeability, and metabolic adaptation [37]. The mechanisms of inactivation of macrolides include a variety of modifications such as hydrolysis, reduction, acetylation, and deacylation [38]. Esterases EreA and EreB, found in S. aureus, cleave the macrocyclic ester linkage, providing resistance to the class of macrolide antibiotics. As shown in the example of tetracycline degradation, modification of an antibiotic can sometimes lead to selection against resistance due to the activity of stable degradation products [39].
Here, we observed that macrolatins undergo biotransformation in S. aureus cells, resulting in MLN A backbone rearrangement that could be described as an oxidation of α,β-unsaturated alcohols to ketones followed by double bond reduction leading to less toxic MLN F backbone products. Although enzymatic redox reactions of unsaturated compounds are quite intensively studied, we have not found any examples of one-step biotransformation such as the rearrangement detected for macrolactins. The possible pathways for two-step biotransformation may somehow simulate steps of fatty-acid synthesis, including oppositely directed oxidation of α,β-unsaturated alcohols followed by double bond reduction. The closest examples of the oxidation reactions involving bacterial enzymes were reported for the whole-cell biocatalysis processes. Biocatalytic oxidation of racemic (hetero)aromatic sec-alcohols to the corresponding ketones was reported for alcohol dehydrogenase-modified E. coli cells [40]. Enzymatic enantioselective oxidation of allyl alcohols was also reported [41]. The biotransformation reaction of macrolactins is of particular interest, as it provides the molecular pathways for antibiotic inactivation and development of antibiotic resistance in pathogenic S. aureus strains as well as natural inactivation of macrolactins. We believe the discovered biotransformations will bring new strategies for creating new antibiotics by stabilizing the architecture of a macrolactam backbone and introducing unnatural acyl analogs.
The MLN BGC is highly conserved in B. velezensis and closely related B. amyloliquefaciens species, which stresses its high significance for the biology of these species. We speculate that production of macrolactins along with the conserved metabolic fingerprint of bacillaene, bacillibactin, and bacilysin is essential for their functionality as efficient biocontrol agents. However, the functionality of the difficidin/oxydifficidin cluster may be a valuable hallmark of B. velezensis-based biocontrol agents. The abundance of the MLN BGC allows us to expand the obtained results to the numerous B. velezensis that are currently applied in agriculture, and we believe that high activity of the MLN A backbone analogs combined with their natural biodegradation pathway is of prime importance.
Materials and Methods
Bacterial Strains and Cell Lines
The macrolactin-producing strain Bacillus velezensis K-3618 was obtained from the collection of the All-Russian Research Institute of Agricultural Microbiology, ID RCAM 07246 [https://en.arriam.ru/kollekciya-kul-tur1]. The strain K 3618 was deposited as isolated from potato tubers of the Charoit variety endophytic microorganism with cellulolytic, amylolytic and weak nitrogen-fixing activity. A bacterial collection including Micrococcus luteus ATCC 4698, Bacillus subtilis 168, Enterococcus faecalis 125, Macrococcus caseolyticus 107, and Escherichia coli BL21 (DE3), Escherichia coli BW25113 ΔtolC и ΔlptD was kindly provided by Lytech Co., Ltd. (Moscow, Russia) [65]. Staphylococcus aureus GFP constitutively producing GFP was kindly provided by Andrey Shkoporov from the Department of Microbiology and Virology, Russian National Research Medical University, Moscow.
Cell line HEK293T cells were kindly provided by Dr. E. Knyazhanskaya.
Genome Sequencing, Assembly and Annotation
The Wizard Genomic DNA kit (Promega, USA) was used to isolate genomic DNA. The DNA library was prepared using the KAPA HyperPlus kit (Roche, Switzerland). Whole-genome sequencing was performed at the Lopukhin Center for Genomics, Proteomics, Metabolomics (Russia) using the NextSeq 1000 platform (Illumina, USA) and NextSeq 1000/2000 P1 reagents (300 cycles).
Raw sequencing reads were initially subjected to quality assessment using FastQC (v0.12.1) [42], and subsequent filtering of low-quality and adapter-contaminated reads was performed with fastp (v0.23.2) [43]. The genome assembly was conducted using the SPAdes toolkit (v4.0.0) [44], followed by assessment of assembly metrics via QUAST (v5.2.0) [45]. To assess taxonomic placement and evaluate genome quality, we applied CheckM (v1.2.2) [30], and the closest related genomes were identified using fastANI (v1.33) based on comparisons with RefSeq [46] assemblies of Bacillus species from the NCBI database. Genome annotation was performed using Prokka (v1.14.6) [47].
Phylogeny Reconstruction
To reconstruct the reference phylogeny, we first obtained 30 closest genomes relative to our strain according to the ANI (Average Nucleotide Identity) estimates with all available genomes of the Bacillales order deposited in the RefSeq Assembly [46] database. The pair-wise genome similarity was evaluated with the fastANI v1.33 tool [48]. The selected dataset was then used to build the pangenome applying Panaroo v1.2.8 [49] launched in the “strict” mode. The MAFFT aligner v.7 [50] was specified to align core genes defined as those present in not less than 95% of genomes. Based on the concatenated alignment of core genes, we determined the best-fit evolutionary model with ModelTest-NG v0.1.7 [51]. To reduce insignificant signals, we extracted SNPs (Single-Nucleotide Polymorphisms) with SNP-sites v2.5.1 [52]. The resulting alignment and selected evolutionary model were used for phylogeny reconstruction via the ML (Maximum Likelihood) approach implemented in the RAxML-NG v1.1.0 program [53]. A total of 1000 bootstrap replicates were specified. Visualization of the results, both the reference tree and the ordered data, such as BGCs, was performed by applying the ggplot2 v3.3.5 [54] and ape v5.6-2 [55] packages.
Cultivation, Isolation and Purification of Macrolactins
The strain K-3818 was cultivated on the agar medium at the temperature in the range of 37 C. The strain was transferred from the surface of the agar to a 750 mL Erlenmeyer flask with 50 mL of of 2YT nutrient medium of the following composition, g/L: tryptone—16, yeast extract—10, NaCl—5, distilled water—up to 1 L, pH 7.0. Cultivation was carried out at 28C for 24 h on a thermostat shaker Innova 40 (New Brunswick Scientific, USA) at 150 rpm. Using the first culture as an inoculum for seeding at a concentration of 3% v/v , the second generation culture was grown in 150 ml on the same medium and under the same conditions.
Bacterial cells were eliminated from culture broth by centrifugation at 5000 rpm on Sigma 3-16KL centrifuge and filtration through 0.47 μm MCE membrane filter (Millipore, USA). 1 L of cleared supernatant was loaded on the 30 ml cartridge packed with 7 g of LPS-500-H polymer sorbent (copolymer divinylbenzene - hydrophilic monomer, pore size 50-1000 Å, 70 μm, Technosorbent, RF) at flow rate 15 ml/min using peristaltic pump (Masterflex L/S variable speed pump Systems, Masterflex, Germany). Extraction was performed using a syringe, passing 15 ml of a mixture of water and acetonitrile (ACN) at a rate of about 15 ml/min, successively with an ACN content of 0, 10, 20, 35, 50, 75 and 100%. Activity of the collected fractions was tested using a reporter S. aureus GFP strain. Active fractions eluted at 35, 50 and 75 % ACN were further analyzed by HPLC on RP column using Nexera X2 LC 30A instrument (Shimadzu, Japan) equipped with SPD-M20A detector. HPLC conditions for the screening analysis were as following: HC C18 150 x 4.6 mm, 5 μm, 110 Å; eluent solvents A – 0.1% AA, B – MeCN; gradient elution from 30% B to 90% B at 8 min, then 90% of solvent B; flow rate 1.5 mL/min, UV 260 nm. HPLC fractions were collected, tested for activity, and the active fractions containing pure active substances were isolated and then analyzed by LCMS. Mal-MLN A was contained in fractions 35 and 50% ACN, MLN A and suc-MLN A were in fractions of 50 and 75% ACN. All three fractions were combined for subsequent isolation of the compounds by HPLC on semipreparative column Gemini NX C18 150x20 mm, 10 μm, 110 Å (Phenomenex) using PuriFlash 5.250 instrument (Interchim). Isocratic elution at 43% of solvent B for 20 min at flow rate 16 mL/min were applied.
LC-MS Analysis
LCMS analysis was carried out on an Ultimate 3000 RSLCnano HPLC system connected to an Orbitrap Fusion Lumos mass spectrometer (ThermoFisher Scientific, Waltham, MA, USA) with the loading pump used for analytical flow gradient delivery. Samples were separated on a Gemini NX-C18 3 μm 100 Å column 100*2.1 mm at 200 μL/min flow rate in the linear gradient of acetonitrile in water with the addition of 10 mM ammonium formate and 0.1% FA. UV data were collected at 275 nm. MS1 and MS2 spectra were recorded at 30K and 15K resolution, respectively, with HCD fragmentation. Raw data were collected and processed on Thermo Xcalibur Qual ver. 4.3.73.11.
NMR
The structures of the compounds were elucidated using the conventional heteronuclear NMR approach. The compounds presumably identified according HRMS as macrolactin A, 7-O-malonyl macrolactin A, 7-O-succinyl macrolactin A were dissolved in CDCl3 (99.8%, Solvex, RF) and placed into the 5 mm NMR tube (Wilmad, USA). Other samples were dissolved in MeOD-d4 (100%, Sigma-Aldrich, USA) and placed into a 5 mm shigemi NMR tube (SHIGEMI, Japan). NMR spectra were recorded at 30°C using the Bruker Avance II 700 MHz NMR spectrometer (Bruker, Billerica, MA, USA) equipped with a TCI cryogenic probe. The 1D 1H,13C and 2D 1H,13C-HSQC; 1H, 13C-HMBC, DQF-COSY and 2D TOCSY spectra were recorded for each sample.
Antibacterial Activity Assessment
Determination of the activity of the pure compounds was carried out using the method of serial 2X-fold dilutions starting from the final concentrations of 50 μg/mL. Individual colonies of bacteria from the plate with 2YT agar medium (10 g/L yeast extract; 16 g/L tryptone; 5 g/L NaCl, 15 g/L agar) were transferred to 5 mL of 2YT nutrient medium (10 g/L yeast extract; 16 g/L tryptone; 5 g/L NaCl) and incubated at 37℃ overnight. The overnight cultures were transferred to a fresh nutrient medium in a ratio 1:100 and incubated at 37℃ for 3 h. Next, the cultures were diluted to the OD600 ~0.001 a.u. and applied to the serial 2-fold dilutions of compounds under investigation. MICs of antibiotics were determined after 16 h of incubation in a 96-well plate at a wavelength of 600 nm using a Varioskan multimodal reader (Thermo Fisher Scientific, USA).
Cytotoxicity Assessment
To prepare MTT solution, thiazolyl blue tetrazolium bromide was dissolved in phosphate-buffered saline, pH = 7.4, to a concentration of 5 mg/mL, and sterilely filtered through a 0.2 μm filter. To prepare a solubilizing solution, 40% DMF was diluted in 2% acetic acid, and a total of 16% sodium dodecyl sulfate was added to the resulting solution, then pH was adjusted to 4.7. Double dilutions of the test compounds were prepared in DMSO as a negative control in the culture medium. A final concentration series of eight 2-fold dilutions was started at 6.4 μg/mL. Incubation was carried out in a humidified atmosphere at 37◦C with 8% CO2 for 72 h. Cytotoxicity was assessed in HEK293T cells according to the method, described in [56]. Cell viability was determined by colorimetric assessment of cell metabolic activity after incubation for 72 h. All measurements were performed in triplicate biological replicates. A total of 10 μL per well of MTT solution was added to the test samples to a final concentration of 0.45 mg/mL and incubated for an hour at 37 ℃. Then, 100 μL of solubilizing solution was added to each well, the result was measured at 570 nm using a Varioskan multimodal reader (Thermo Scientific, USA).
The graph of the dependence of the average survival value on the concentration of the test substance plotted on a logarithmic scale was constructed using MS Excel 2013 software. A well with cells that did not contain the test compounds was taken as 100% viability. Relative survival was calculated as the ratio of the absorbance for a given well to the absorbance for a well without drug, in percent. The cytotoxic concentration for each compound (IC50) was defined as the concentration at which a cytotoxic effect was induced in 50% of the cells in the monolayer. Doxorubicin was used as positive control.
Inhibition Assays in Cell-Free Translation System
To examine the inhibition of translation by MLN A, mal-MLN A, suc-MLN, we used the E. coli S30 Extract System for Linear Templates (Promega, USA). The reaction mixture (total volume 5 μL) contained: 2 μL of S30 Premix, 1.5 μL of S30 extract, 0.1 μL of the RNase inhibitor Ribolock, 0.5 μL of amino acid mixture, 0.5 μL of Fluc mRNA (200 ng/μL), 0.5 μL of the tested compound (in 10% DMSO). Erythromycin (25 μg/mL) and 10% DMSO were used as controls. The mixture (without mRNA) was prepared on ice and incubated for 5 minutes at room temperature to allow the antibiotic to bind to the ribosome. After mRNA was added, translation proceeded for 20 minutes at 37°C. After incubation, 5 μL of Bright-Glo (Promega, USA) was added. Luminescence was measured using a Victor X5 2030 plate reader (Perkin Elmer, USA). The results were normalized on DMSO.
Biotransformation of with suc-MLN A and mal-MLN A by S.aureus
Macrolactins were solubilized in DMSO at a concentration of 5 mg/mL and added into 2YT medium containing 20 μg/mL chloramphenicol to achieve a final concentration of 0.25 mg/mL. S. aureus GFP was cultured in 2YT medium, supplemented with 20 μg/mL chloramphenicol up to the culture density OD600 = 3. The bacterial cells were centrifugated at 5000 g for 10 min. The cell pellet was resuspended in 2YT medium supplemented with either mal-MLN A or suc-MLN A to attain a final concentration 0.25 mg/mL and bacterial cell density of 7.5×109 CFU/ml. Two types of controls were prepared: 800 μL of the cell suspension without suc-MLN or mal-MLN A and 800 μL of each compound in 2YT media. In the small scale experiment there were only one sample of every kind (experimental suspension, control suspension and control with a compound), in the large scale experiment there were 20 experimental aliquots of 800 µL. The resulting aliquots, as well as control samples were incubated at 37°C with shaking at 200 RPM in conical 15 mL tubes.
Aliquots of 50 μl suspension samples and control samples were taken after incubation periods of 1, 2, 4, 8, 16, and 24 h. In the large scale experiment after 24 h incubation all suspension samples were combined and prepared by the same procedure.
Analysis of Reaction Broth, Isolation and Purification of Target Compounds After Incubation of the Macrolactins with S.aureus Cells
Sample preparation. The samples of 50 μL taken at fixed time points were centrifuged at 10,000 g for 10 min; thereafter the supernatant and cell pellets were separately frozen in liquid nitrogen and stored at -80°C. Prior to analysis, the medium samples were thawed and treated with addition of 100 µL ethyl acetate then vortexed for 15 s before incubation on a tabletop shaker for 5 min. They were then vortexed again for 15 s and centrifuged at 10,000g for 5 min. The resulting ethyl acetate phase was separated, vacuum-dried. For further analysis, every dry residue sample was dissolved in 100 μL of a mixture of 0.1% acetic acid and acetonitrile 1:1 and analyzed by HPLC-UV or HPLC-MS/MS.
HPLC analysis and isolation of compounds. HPLC analysis were performed on RP column using Nexera X2 LC 30A instrument (Shimadzu, Japan) equipped with SPD-M20A detector. HPLC conditions were as following: Luna C18(2) 150 x 4 mm, 3 μm, 110 Å (Phenomenex) eluent solvent A - 10 mM NH4OAc, pH 5, solvent B – MeCN; isocratic elution at 40% of solvent B for 15 min, then gradient to 90 % ACN for 17 min and cleaning the column for 3 min; flow rate 0.75 mL/min, UV 260 nm. In the large-scale experiment isolation and purification of the compounds of interest were performed on semipreparative column Gemini NX C18 150 x 10 mm, 5 μm, 110 Å (Phenomenex).
Conclusions
This study elucidates the molecular basis for the antimicrobial potential of the widely used biocontrol strain Bacillus velezensis K-3618 and establishes macrolactins as its principal effectors. Mechanistic assays indicate that MLN A–based macrolactins primarily act by inhibiting protein translation. Acylation at the 7-O position substantially enhances activity, with acylated analogs outperforming MLN A in prokaryotic cell-free systems at low micromolar concentrations. Oppositely, we uncovered pathogen-mediated biotransformation of MLN A to MLN F analogs, which reduces activity by approximately two orders of magnitude. These findings define two complementary design approaches for next-generation macrolactins—preserving and stabilizing the MLN A core while optimizing acylation to resist degradative conversion. Our data suggest an ecological rationale for the observed degradability: such biotransformations may function as built-in attenuation, balancing efficacy with environmental persistence across Bacillus species in diverse niches. Together, these results provide a mechanistic and chemical blueprint for developing macrolactin-based antibacterials and improving the sustainability and reliability of Bacillus-driven organic farming.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Data Availability Statement
The raw genome sequencing data were submitted to the NCBI with BioSample number SAMN48451324, under BioProject PRJNA1262192. The assembled genome is available in the NCBI Assembly database under the accession JBNVTP000000000.
Acknowledgments
This work was supported by the Ministry of Science and Higher Education of the Russian Federation (Agreement No. 075-15-2024-536).
References
- Baker, B.P.; Green, T.A.; Loker, A.J. Biological Control and Integrated Pest Management in Organic and Conventional Systems. Biol. Control 2020, 140, 104095. [CrossRef]
- Zhong, X.; Jin, Y.; Ren, H.; Hong, T.; Zheng, J.; Fan, W.; Hong, J.; Chen, Z.; Wang, A.; Lu, H.; et al. Research Progress of Bacillus Velezensis in Plant Disease Resistance and Growth Promotion. Front. Ind. Microbiol. 2024, 2, 1442980. [CrossRef]
- Rabbee, M.F.; Hwang, B.-S.; Baek, K.-H. Bacillus Velezensis: A Beneficial Biocontrol Agent or Facultative Phytopathogen for Sustainable Agriculture. Agronomy 2023, 13, 840. [CrossRef]
- Tran, C.; Cock, I.E.; Chen, X.; Feng, Y. Antimicrobial Bacillus: Metabolites and Their Mode of Action. Antibiotics 2022, 11, 88. [CrossRef]
- Chen, L.; Wang, X.; Liu, Y. Contribution of Macrolactin in Bacillus Velezensis CLA178 to the Antagonistic Activities against Agrobacterium Tumefaciens C58. Arch. Microbiol. 2021, 203, 1743–1752. [CrossRef]
- Yuan, J.; Li, B.; Zhang, N.; Waseem, R.; Shen, Q.; Huang, Q. Production of Bacillomycin- and Macrolactin-Type Antibiotics by Bacillus Amyloliquefaciens NJN-6 for Suppressing Soilborne Plant Pathogens. J. Agric. Food Chem. 2012, 60, 2976–2981. [CrossRef]
- Wang, J.; Peng, Y.; Xie, S.; Yu, X.; Bian, C.; Wu, H.; Wang, Y.; Ding, T. Biocontrol and Molecular Characterization of Bacillus Velezensis D against Tobacco Bacterial Wilt. Phytopathol. Res. 2023, 5, 50. [CrossRef]
- Yousfi, S.; Krier, F.; Deracinois, B.; Steels, S.; Coutte, F.; Frikha-Gargouri, O. Characterization of Bacillus Velezensis 32a Metabolites and Their Synergistic Bioactivity against Crown Gall Disease. Microbiol. Res. 2024, 280, 127569. [CrossRef]
- Lu, X.; Xu, Q.; Liu, X.; Cao, X.; Ni, K.; Jiao, B. Marine Drugs – Macrolactins. Chem. Biodivers. 2008, 5, 1669–1674. [CrossRef]
- Wu, T.; Xiao, F.; Li, W. Macrolactins: Biological Activity and Biosynthesis. Mar. Life Sci. Technol. 2021, 3, 62–68. [CrossRef]
- Gustafson, K.; Roman, M.; Fenical, W. The Macrolactins, a Novel Class of Antiviral and Cytotoxic Macrolides from a Deep-Sea Marine Bacterium. J. Am. Chem. Soc. 1989, 111, 7519–7524. [CrossRef]
- Xu, Y.; Song, Y.; Ning, Y.; Li, S.; Qu, Y.; Jiao, B.; Lu, X. Macrolactin XY, a Macrolactin Antibiotic from Marine-Derived Bacillus Subtilis Sp. 18. Mar. Drugs 2024, 22, 331. [CrossRef]
- Jaruchoktaweechai, C.; Suwanborirux, K.; Tanasupawatt, S.; Kittakoop, P.; Menasveta, P. New Macrolactins from a Marine Bacillus Sp. Sc026. J. Nat. Prod. 2000, 63, 984–986. [CrossRef]
- Romero-Tabarez, M.; Jansen, R.; Sylla, M.; Lünsdorf, H.; Häußler, S.; Santosa, D.A.; Timmis, K.N.; Molinari, G. 7- O -Malonyl Macrolactin A, a New Macrolactin Antibiotic from Bacillus Subtilis Active against Methicillin-Resistant Staphylococcus Aureus , Vancomycin-Resistant Enterococci, and a Small-Colony Variant of Burkholderia Cepacia. Antimicrob. Agents Chemother. 2006, 50, 1701–1709. [CrossRef]
- Schneider, K.; Chen, X.-H.; Vater, J.; Franke, P.; Nicholson, G.; Borriss, R.; Süssmuth, R.D. Macrolactin Is the Polyketide Biosynthesis Product of the Pks2 Cluster of Bacillus Amyloliquefaciens FZB42. J. Nat. Prod. 2007, 70, 1417–1423. [CrossRef]
- Xiao, F.; Dong, S.; Liu, Y.; Feng, Y.; Li, H.; Yun, C.-H.; Cui, Q.; Li, W. Structural Basis of Specificity for Carboxyl-Terminated Acyl Donors in a Bacterial Acyltransferase. J. Am. Chem. Soc. 2020, 142, 16031–16038. [CrossRef]
- Bae, S.H.; Kwon, M.J.; Park, J.B.; Kim, D.; Kim, D.-H.; Kang, J.-S.; Kim, C.-G.; Oh, E.; Bae, S.K. Metabolic Drug-Drug Interaction Potential of Macrolactin A and 7- O -Succinyl Macrolactin A Assessed by Evaluating Cytochrome P450 Inhibition and Induction and UDP-Glucuronosyltransferase Inhibition In Vitro. Antimicrob. Agents Chemother. 2014, 58, 5036–5046. [CrossRef]
- Kim, J.M.; Jung, J.W.; Kim, D.H.; Kang, J.S.; Kim, C.G.; Kang, H.E. A Simple and Sensitive HPLC-UV Determination of 7- O -succinyl Macrolactin A in Rat Plasma and Urine and Its Application to a Pharmacokinetic Study. Biomed. Chromatogr. 2013, 27, 273–279. [CrossRef]
- Jung, J.W.; Kim, J.M.; Kwon, M.H.; Kim, D.H.; Kang, H.E. Pharmacokinetics of Macrolactin A and 7- O -Succinyl Macrolactin A in Mice. Xenobiotica 2014, 44, 547–554. [CrossRef]
- Salazar, F.; Ortiz, A.; Sansinenea, E. A Strong Antifungal Activity of 7-O-Succinyl Macrolactin A vs Macrolactin A from Bacillus Amyloliquefaciens ELI149. Curr. Microbiol. 2020, 77, 3409–3413. [CrossRef]
- Kim, D.H.; Kim, H.K.; Kim, K.M.; Kim, C.K.; Jeong, M.H.; Ko, C.Y.; Moon, K.H.; Kang, J.S. Antibacterial Activities of Macrolactin a and 7-O-Succinyl Macrolactin a from Bacillus Polyfermenticus KJS-2 against Vancomycin-Resistant Enterococci and Methicillin-Resistant Staphylococcus Aureus. Arch. Pharm. Res. 2011, 34, 147–152. [CrossRef]
- Nagao, T.; Adachi, K.; Sakai, M.; Nishijima, M.; Sano, H. Novel Macrolactins as Antibiotic Lactones from a Marine Bacterium. J. Antibiot. (Tokyo) 2001, 54, 333–339. [CrossRef]
- Vasilchenko, A.S.; Lukyanov, D.A.; Dilbaryan, D.S.; Usachev, K.S.; Poshvina, D.V.; Taldaev, A.K.; Nikandrova, A.A.; Imamutdinova, A.N.; Garaeva, N.S.; Bikmullin, A.G.; et al. Macrolactin a Is an Inhibitor of Protein Biosynthesis in Bacteria. Biochimie 2025, 232, 25–34. [CrossRef]
- Yoo, J.-S.; Zheng, C.-J.; Lee, S.; Kwak, J.-H.; Kim, W.-G. Macrolactin N, a New Peptide Deformylase Inhibitor Produced by Bacillus Subtilis. Bioorg. Med. Chem. Lett. 2006, 16, 4889–4892. [CrossRef]
- Sohn, M.-J.; Zheng, C.-J.; Kim, W.-G. Macrolactin S, a New Antibacterial Agent with Fab G-Inhibitory Activity from Bacillus Sp. AT28. J. Antibiot. (Tokyo) 2008, 61, 687–691. [CrossRef]
- Kalai, K.; Rufus, A.C.; Manz, A.M.; Elangovan, E. Unveiling the Antimicrobial Potential of 7-O-Succinyl Macrolactin F from Bacillus Subtilis Group against HtsA Siderophore Receptor of Staphylococcus Aureus: A Computational Exploration. Biomed. Biotechnol. Res. J. 2024, 8, 92–99. [CrossRef]
- Terekhov, S.S.; Smirnov, I.V.; Malakhova, M.V.; Samoilov, A.E.; Manolov, A.I.; Nazarov, A.S.; Danilov, D.V.; Dubiley, S.A.; Osterman, I.A.; Rubtsova, M.P.; et al. Ultrahigh-Throughput Functional Profiling of Microbiota Communities. Proc. Natl. Acad. Sci. 2018, 115, 9551–9556. [CrossRef]
- Duddeck, H. E. Pretsch, P. Bühlmann, C. Affolter. Structure Determination of Organic Compounds—Tables of Spectra Data. Springer, Berlin, 2000. 421 Pp. plus CD-ROM. Price £ 40.39, DM 79.00. ISBN 3 540 67815 8. Magn. Reson. Chem. 2002, 40, 247–247. [CrossRef]
- Bacillus Subtilis CH13: A Highly Effective Biocontrol Agent for the Integrated Management of Plant Diseases. IOBC-WPRS.
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the Quality of Microbial Genomes Recovered from Isolates, Single Cells, and Metagenomes. Genome Res. 2015, 25, 1043–1055. [CrossRef]
- Gilchrist, C.L.M.; Chooi, Y.-H. Clinker & Clustermap.Js: Automatic Generation of Gene Cluster Comparison Figures. Bioinformatics 2021, 37, 2473–2475. [CrossRef]
- Zdouc, M.M.; Blin, K.; Louwen, N.L.L.; Navarro, J.; Loureiro, C.; Bader, C.D.; Bailey, C.B.; Barra, L.; Booth, T.J.; Bozhüyük, K.A.J.; et al. MIBiG 4.0: Advancing Biosynthetic Gene Cluster Curation through Global Collaboration. Nucleic Acids Res. 2025, 53, D678–D690. [CrossRef]
- Chen, X.-H.; Vater, J.; Piel, J.; Franke, P.; Scholz, R.; Schneider, K.; Koumoutsi, A.; Hitzeroth, G.; Grammel, N.; Strittmatter, A.W.; et al. Structural and Functional Characterization of Three Polyketide Synthase Gene Clusters in Bacillus Amyloliquefaciens FZB 42. J. Bacteriol. 2006, 188, 4024–4036. [CrossRef]
- Chen, X.H.; Koumoutsi, A.; Scholz, R.; Eisenreich, A.; Schneider, K.; Heinemeyer, I.; Morgenstern, B.; Voss, B.; Hess, W.R.; Reva, O.; et al. Comparative Analysis of the Complete Genome Sequence of the Plant Growth–Promoting Bacterium Bacillus Amyloliquefaciens FZB42. Nat. Biotechnol. 2007, 25, 1007–1014. [CrossRef]
- Liu, Y.; Qin, W.; Liu, Q.; Zhang, J.; Li, H.; Xu, S.; Ren, P.; Tian, L.; Li, W. Genome-wide Identification and Characterization of Macrolide Glycosyltransferases from a Marine-derived Bacillus Strain and Their Phylogenetic Distribution. Environ. Microbiol. 2016, 18, 4770–4781. [CrossRef]
- Blin, K.; Shaw, S.; Augustijn, H.E.; Reitz, Z.L.; Biermann, F.; Alanjary, M.; Fetter, A.; Terlouw, B.R.; Metcalf, W.W.; Helfrich, E.J.N.; et al. antiSMASH 7.0: New and Improved Predictions for Detection, Regulation, Chemical Structures and Visualisation. Nucleic Acids Res. 2023, 51, W46–W50. [CrossRef]
- Foster, T.J. Antibiotic Resistance in Staphylococcus Aureus. Current Status and Future Prospects. FEMS Microbiol. Rev. 2017, 41, 430–449. [CrossRef]
- Matsumoto, K.; Kawabata, Y.; Okada, S.; Takahashi, J.; Hashimoto, K.; Nagai, Y.; Tatsuta, J.; Hatanaka, M. Enantioselective Microbial Oxidation of Allyl Alcohols. Chem. Lett. 2007, 36, 1428–1429. [CrossRef]
- Wilson, D.N. Ribosome-Targeting Antibiotics and Mechanisms of Bacterial Resistance. Nat. Rev. Microbiol. 2014, 12, 35–48. [CrossRef]
- ADH-Catalyzed Biooxidation of (Hetero)Aromatic sec-Alcohols to Ketones Employing Vinyl Acetate as Acetaldehyde Surrogate - Rudzka - 2024 - ChemCatChem - Wiley Online Library Available online: https://chemistry-europe.onlinelibrary.wiley.com/doi/10.1002/cctc.202400803 (accessed on 12 April 2025).
- Gandomkar, S.; Jost, E.; Loidolt, D.; Swoboda, A.; Pickl, M.; Elaily, W.; Daniel, B.; Fraaije, M.W.; Macheroux, P.; Kroutil, W. Biocatalytic Enantioselective Oxidation of Sec -Allylic Alcohols with Flavin-Dependent Oxidases. Adv. Synth. Catal. 2019, 361, 5264–5271. [CrossRef]
- Babraham Bioinformatics - FastQC A Quality Control Tool for High Throughput Sequence Data Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 16 May 2025).
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality Assessment Tool for Genome Assemblies. Bioinformatics 2013, 29, 1072–1075. [CrossRef]
- O’Leary, N.A.; Wright, M.W.; Brister, J.R.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D.; et al. Reference Sequence (RefSeq) Database at NCBI: Current Status, Taxonomic Expansion, and Functional Annotation. Nucleic Acids Res. 2016, 44, D733–D745. [CrossRef]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [CrossRef]
- Jain, C.; Rodriguez-R, L.M.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High Throughput ANI Analysis of 90K Prokaryotic Genomes Reveals Clear Species Boundaries. Nat. Commun. 2018, 9, 5114. [CrossRef]
- Tonkin-Hill, G.; MacAlasdair, N.; Ruis, C.; Weimann, A.; Horesh, G.; Lees, J.A.; Gladstone, R.A.; Lo, S.; Beaudoin, C.; Floto, R.A.; et al. Producing Polished Prokaryotic Pangenomes with the Panaroo Pipeline. Genome Biol. 2020, 21, 180. [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [CrossRef]
- Darriba, D.; Posada, D.; Kozlov, A.M.; Stamatakis, A.; Morel, B.; Flouri, T. ModelTest-NG: A New and Scalable Tool for the Selection of DNA and Protein Evolutionary Models. Mol. Biol. Evol. 2020, 37, 291–294. [CrossRef]
- Page, A.J.; Taylor, B.; Delaney, A.J.; Soares, J.; Seemann, T.; Keane, J.A.; Harris, S.R. SNP-Sites: Rapid Efficient Extraction of SNPs from Multi-FASTA Alignments. Microb. Genomics 2016, 2. [CrossRef]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A Fast, Scalable and User-Friendly Tool for Maximum Likelihood Phylogenetic Inference. Bioinformatics 2019, 35, 4453–4455. [CrossRef]
- Ggplot2: Elegant Graphics for Data Analysis | SpringerLink Available online: https://link.springer.com/book/10.1007/978-0-387-98141-3 (accessed on 16 May 2025).
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of Phylogenetics and Evolution in R Language. Bioinformatics 2004, 20, 289–290. [CrossRef]
- Riss, T.L.; Moravec, R.A.; Niles, A.L.; Duellman, S.; Benink, H.A.; Worzella, T.J.; Minor, L. Cell Viability Assays. In Assay Guidance Manual; Markossian, S., Grossman, A., Arkin, M., Auld, D., Austin, C., Baell, J., Brimacombe, K., Chung, T.D.Y., Coussens, N.P., Dahlin, J.L., Devanarayan, V., Foley, T.L., Glicksman, M., Gorshkov, K., Haas, J.V., Hall, M.D., Hoare, S., Inglese, J., Iversen, P.W., Lal-Nag, M., Li, Z., Manro, J.R., McGee, J., McManus, O., Pearson, M., Riss, T., Saradjian, P., Sittampalam, G.S., Tarselli, M., Trask, O.J., Weidner, J.R., Wildey, M.J., Wilson, K., Xia, M., Xu, X., Eds.; Eli Lilly & Company and the National Center for Advancing Translational Sciences: Bethesda (MD), 2004.
Figure 1.
Isolation and identification of the macrolactins. (A) General structure of macrolacin A scaffold with R-group substitutions. (B) HPLC-UV chromatogram of the active fraction extracted at 50 % ACN at 260 nm. The peaks corresponding to the macrolactin A (MLN A), 7-O-malonyl-macrolactin A (mal-MLN A), and 7-O-succinyl-macrolactin A (suc-MLN A) are indicated in blue. Inset: UV absorption spectrum with characteristic maxima at 227 and 262 nm for MLN A, mal-MLN A, and suc-MLN A. (C-E) Positive-ion mode ESI-MS spectra of MLN A (C), mal-MLN A (D), and suc-MLN A (E).
Figure 1.
Isolation and identification of the macrolactins. (A) General structure of macrolacin A scaffold with R-group substitutions. (B) HPLC-UV chromatogram of the active fraction extracted at 50 % ACN at 260 nm. The peaks corresponding to the macrolactin A (MLN A), 7-O-malonyl-macrolactin A (mal-MLN A), and 7-O-succinyl-macrolactin A (suc-MLN A) are indicated in blue. Inset: UV absorption spectrum with characteristic maxima at 227 and 262 nm for MLN A, mal-MLN A, and suc-MLN A. (C-E) Positive-ion mode ESI-MS spectra of MLN A (C), mal-MLN A (D), and suc-MLN A (E).
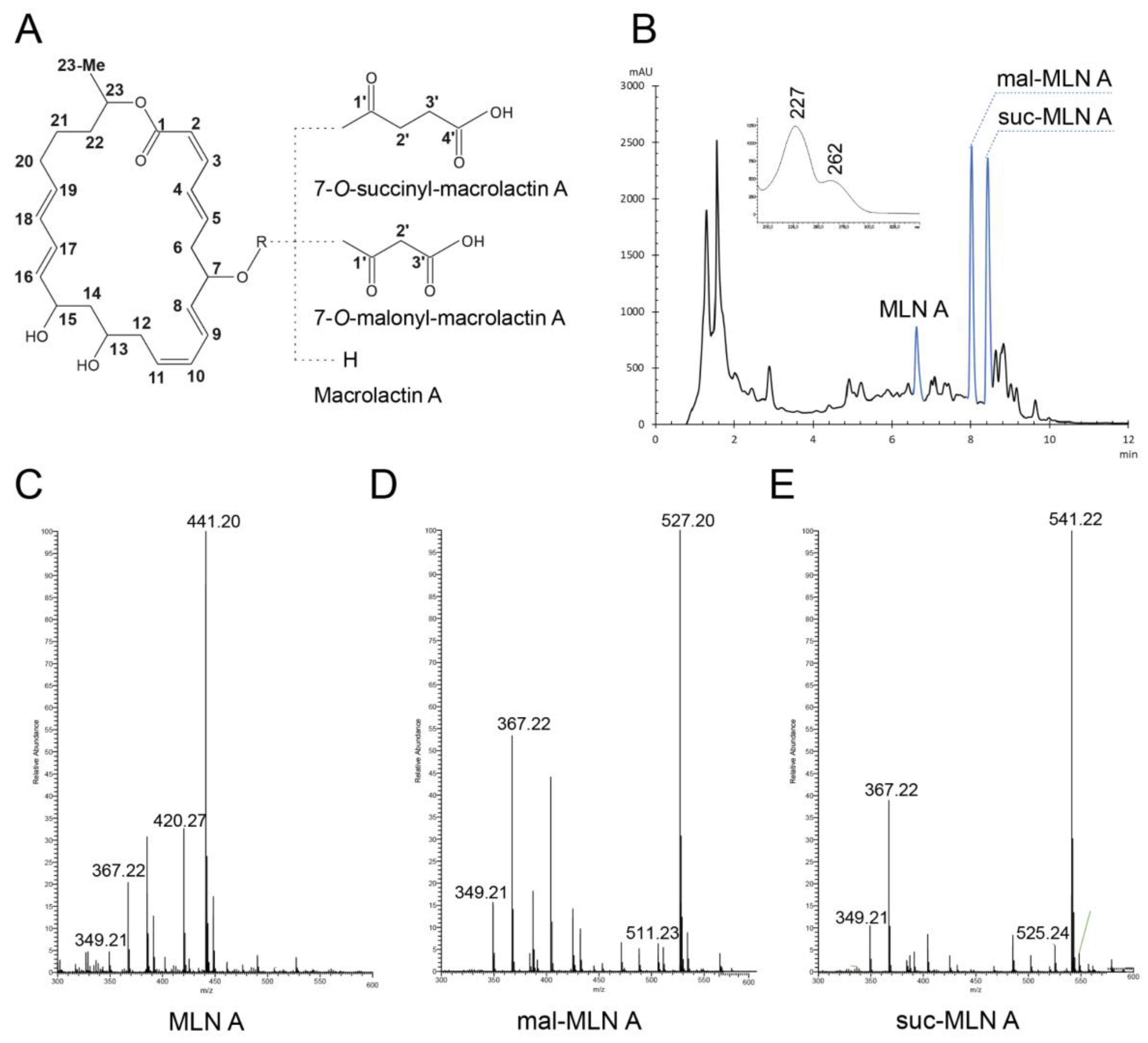
Figure 2.
Antimicrobial activity of macrolactins is mediated by inhibition of protein translation. (A) Antimicrobial activity spectra and toxicity of the macrolactins. (B) Inhibition of protein synthesis by MLN A, mal-MLN A, and suc-MLN A. Erythromycin, 25 μg/mL, and DMSO were used as positive and negative controls, respectively.
Figure 2.
Antimicrobial activity of macrolactins is mediated by inhibition of protein translation. (A) Antimicrobial activity spectra and toxicity of the macrolactins. (B) Inhibition of protein synthesis by MLN A, mal-MLN A, and suc-MLN A. Erythromycin, 25 μg/mL, and DMSO were used as positive and negative controls, respectively.
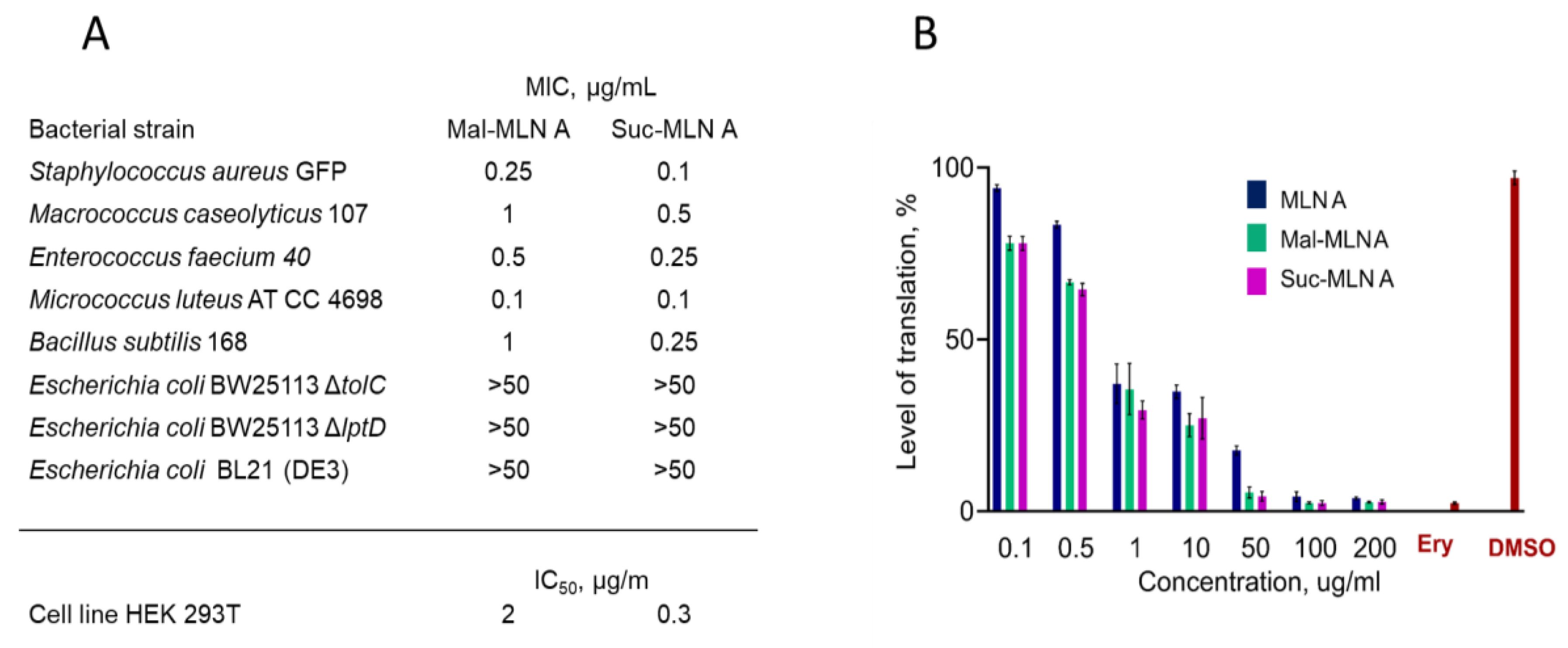
Figure 3.
Degradation of the macrolactins is accelerated when incubated in 2YT medium with S.aureus cells according HPLC. (A,B) HPLC data. (C) Mal-MLN A degradation. (D) Suc-MLN A degradation. (E) Products of mal-MLN A transformation with bacterial cells. (F) Products of suc-MLN A transformation with bacterial cells.
Figure 3.
Degradation of the macrolactins is accelerated when incubated in 2YT medium with S.aureus cells according HPLC. (A,B) HPLC data. (C) Mal-MLN A degradation. (D) Suc-MLN A degradation. (E) Products of mal-MLN A transformation with bacterial cells. (F) Products of suc-MLN A transformation with bacterial cells.
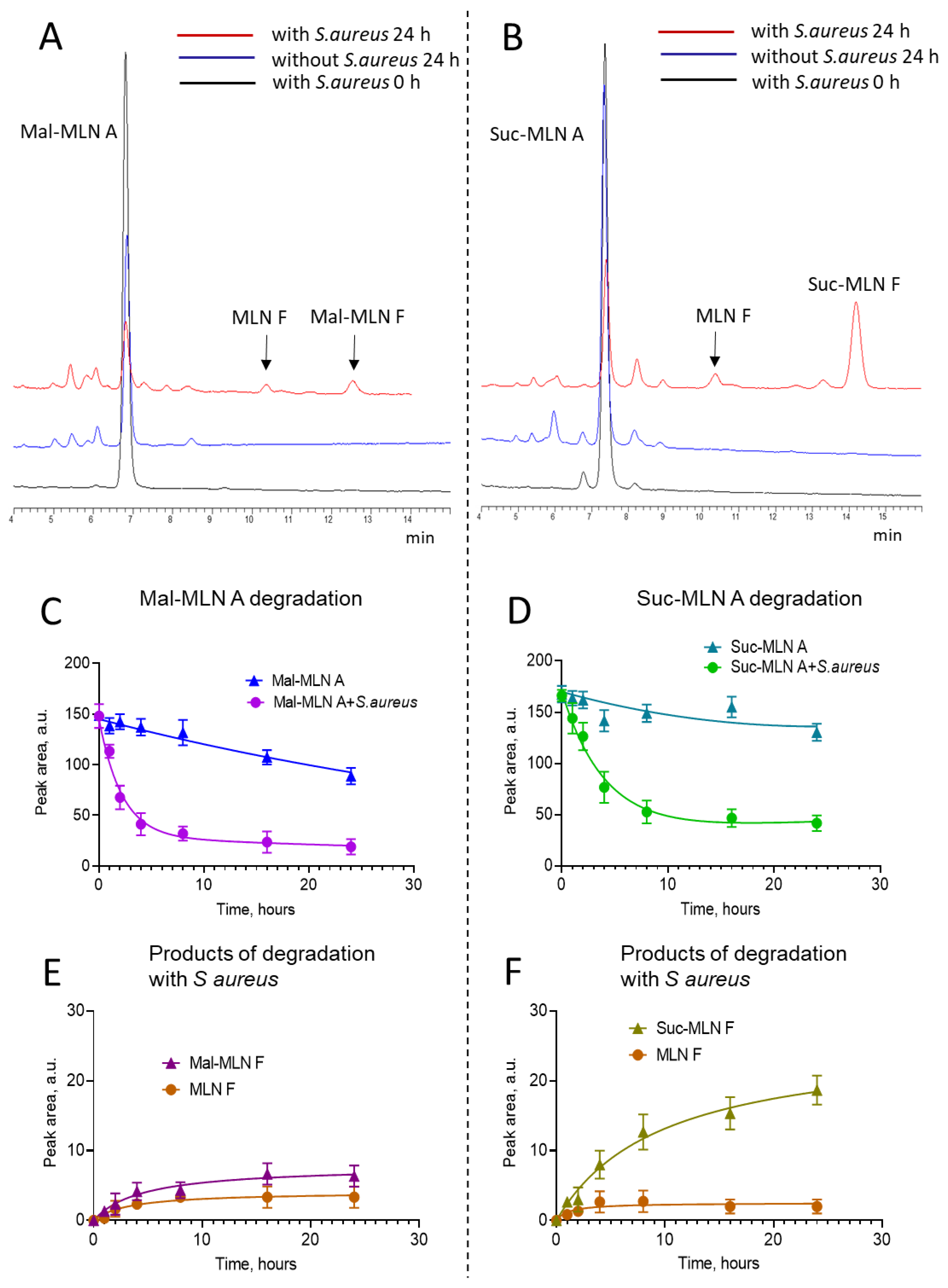
Figure 4.
Comparison of the macrolactin BGCs (BGC0000181) with region 8.1 in the complete genome sequence of Bacillus sp. K-3618, and the selected homologues clusters in closely related species, generated using clinker tool [31]. Homologous genes are color-coded, and labels indicate the corresponding protein identities.
Figure 4.
Comparison of the macrolactin BGCs (BGC0000181) with region 8.1 in the complete genome sequence of Bacillus sp. K-3618, and the selected homologues clusters in closely related species, generated using clinker tool [31]. Homologous genes are color-coded, and labels indicate the corresponding protein identities.
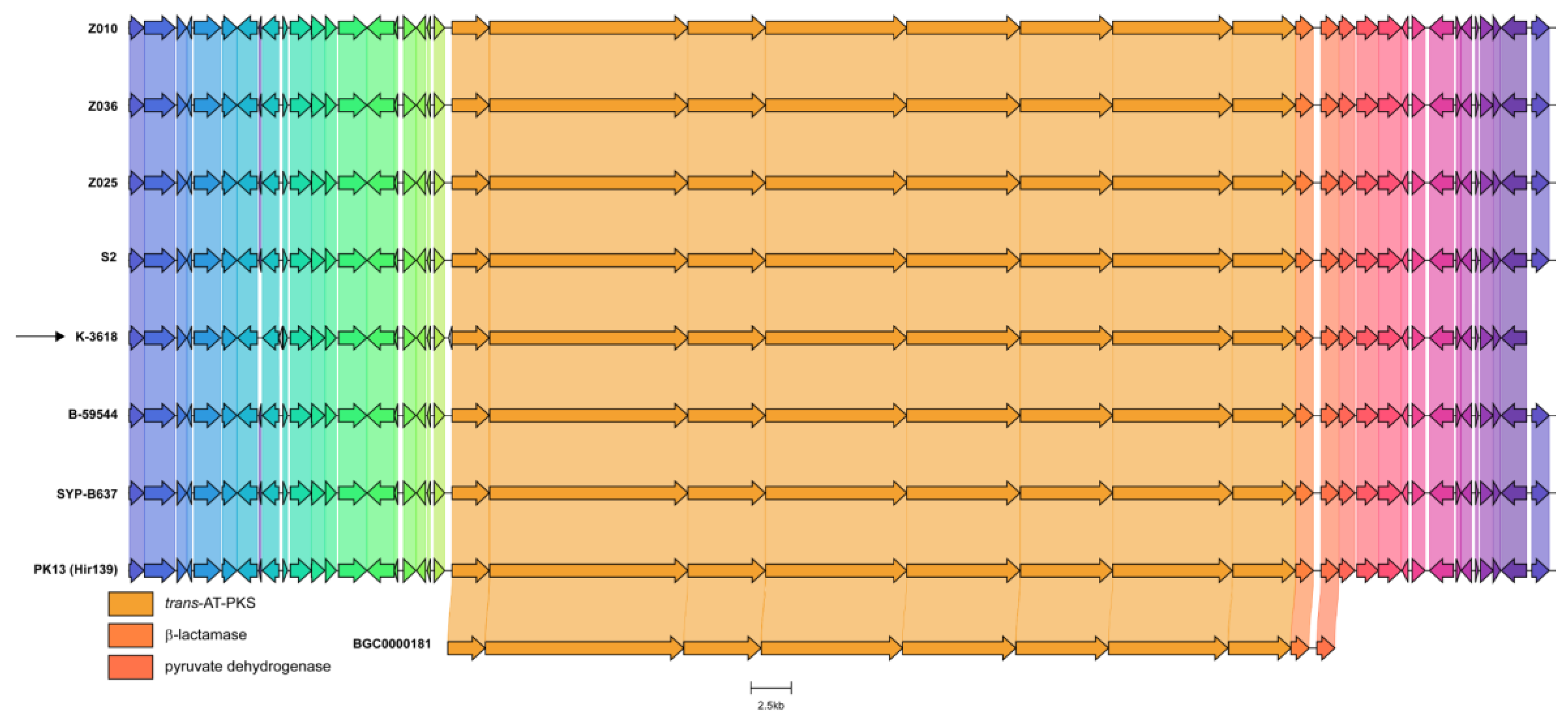
Figure 5.
Phylogeny-wise distribution of BGCs (biosynthetic gene clusters) in closest reference genomes relative to the B. velezensis K-3618 strain. (a) Reference ML (Maximum likelihood) phylogeny reconstructed on core SNPs of 30 closest strains to the K-3618 strain based on the ANI (Average Nucleotide Identity) metrics. The tree was obtained using the RAxML-NG v1.1.0 [7] software. The numbers near the branches represent bootstrap support values with 1000 replicates. The adjacent left most colored bars illustrate the strains’ metadata, namely, taxonomic attribution, isolation source, and country of origin for the studied strains colorized according to the respective features. The black arrow points at the K-3618 strain. (b) The distribution of BGCs identified in the analyzed genomes using the antiSMASH v7.1 utility [10]. The rows correspond to strains ordered according to the reference phylogeny. The color of the tiles is proportional to the overall similarity with known BGCs calculated by the mentioned software.
Figure 5.
Phylogeny-wise distribution of BGCs (biosynthetic gene clusters) in closest reference genomes relative to the B. velezensis K-3618 strain. (a) Reference ML (Maximum likelihood) phylogeny reconstructed on core SNPs of 30 closest strains to the K-3618 strain based on the ANI (Average Nucleotide Identity) metrics. The tree was obtained using the RAxML-NG v1.1.0 [7] software. The numbers near the branches represent bootstrap support values with 1000 replicates. The adjacent left most colored bars illustrate the strains’ metadata, namely, taxonomic attribution, isolation source, and country of origin for the studied strains colorized according to the respective features. The black arrow points at the K-3618 strain. (b) The distribution of BGCs identified in the analyzed genomes using the antiSMASH v7.1 utility [10]. The rows correspond to strains ordered according to the reference phylogeny. The color of the tiles is proportional to the overall similarity with known BGCs calculated by the mentioned software.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



