
Submitted:
13 August 2025
Posted:
14 August 2025
You are already at the latest version
Abstract
Coat color in cats is determined by the interaction of eumelanin and pheomelanin pig-ments, regulated by multiple genes. Traditional Thai Suphalak cat, historically de-scribed as having a solid chocolate coat with yellow, amber to golden eyes, is now often seen with darker points on the face and paws. This study conducted genetic testing on five coat color genes (ASIP, TYRP1, TYR, MLPH, and MC1R) across 14 sequence vari-ants in 127 cats selected for Suphalak breed development. Cats were sourced from the Thai Cat Conservation Center (n = 61) and Thai breeders (n = 66). All cats from the Center carried color point alleles (97.54% cb, 2.46% cs), indicating previous breeding without genetic knowledge. After applying genetic selection, the frequency of color point alleles declined to 43.18%. Among the selected cats, 55 displayed a solid chocolate coat, though some carried color point or dilute alleles. Genotype of aabbCCDDEE was identified as ideal for maintaining the traditional Suphalak phenotype. While carrier cats may still be used, they should be selected with caution. Genetic testing, supported by breeding trials, enables informed selection of breeding pairs to reduce undesirable alleles and restore the authentic Suphalak appearance while preserving genetic diver-sity.
Keywords:
cat
; Supraluk
; traditional Thai cat
; coat color gene
; color point
1. Introduction
Thailand is known to be the origin of the Siamese cat, one of the oldest and most recognizable cat breeds in the world, with a fascinating history that dates back to ancient times in Southeast Asia. The Siamese cat has deep historical roots in Thailand, and ancient Thai manuscripts document the significance of cats. Dating back to ancient times, traditional Thai cats were regarded as symbols of prosperity, protection, and good fortune, and were also believed to bring luck and ward off evil spirits. The well-known Thai cat breed, the Siamese cat (or Wichien Maat in Thai), provides a clue to Thailand’s former name, Siam. The distribution of Siamese cats to Europe occurred in the 19th century through diplomatic exchanges by the Thai monarchy [1]. The ancient manuscript (Samut Khoi) includes an article on cats titled ‘Tamra Maew’, which is kept in the National Library of Thailand and dates back to the Ayutthaya period (1350-1767 AD) [2]. According to the literature, ancient traditional Thai cats consisted of 17 types of ‘good luck’ cats and 6 types of ‘unfortunate’ cats [3]. Today, only 5 traditional Thai cat breeds (Korat, Siamese, Khao Manee, Suphalak, and Konja) remain to be seen in Thailand. For more than five decades, the Suphalak cat has been extremely rare, and confusion has persisted among Thai breeders regarding its distinction from the Burmese cat. This is discussed in the book Siamese Cats: Legends and Reality, written by the foreign author Martin R. Clutterbuck in 2004. In the book, the Suphalak cat is referred to as Suphalak (Burmese or Tonkinese). The book describes the Suphalak cat in Thailand, featuring images of cats with point markings that resemble Burmese cats. These images were sourced from a well-known Thai breeder, who is widely recognized for his efforts in preserving Thai cats. The Suphalak cat is a natural breed that originated during the 14th–18th centuries in the Ayutthaya Kingdom, the same era as the Siamese cat. This breed is closely linked to the traditions and cultural significance of Thailand. Its described characteristics include Thai-style short hair, a heart-shaped face, muscular build, long tail, slim body, and eyes ranging in color from yellow to amber or gold. The ancient Thai manuscript describes the color of Suphalak cats as solid brown/chocolate across the entire body, or Thong Daeng (translated as ‘copper’ in Thai). Therefore, the Suphalak cat observed today may differ from the one described in the Thai manuscript, as it displays dark points, particularly on the face and paws.
The Thai Cat Conservation Center in Samut Songkhram Province is one of the well-known places in Thailand dedicated to preserving traditional Thai cat breeds. All remaining traditional Thai cats can be seen at this center. The center focuses on maintaining and developing the Suphalak breed according to the ancient breed standard. While the overall phenotype of the selectively bred cats is acceptable, the appearance of point markings on the extremities remains a major concern. The facial mask and darkened paws make these cats different from the characteristics described in the ancient Thai manuscripts. Their phenotype resembled that of the breed known as the Burmese cat (Figure 1). Although the Burmese cat is not mentioned in the ancient manuscript, sable-point cats were widespread in Thailand, including among feral cats. This resemblance sparked strong debates among Thai cat breeders. In early 2016, a group of Thai cat breeders decided to set up a breeding program for the Suphalak cat. To follow the traditional breed characteristics outlined in the ancient Thai manuscript, many cats that matched the description were selected for the breeding program.
In mammals, more than 100–150 genes have been identified as being associated with coat color [4,5]. The proportion of the major pigments, eumelanin and pheomelanin, affects the phenotypic expression of coat color traits [6,7,8]. Studies of coat color genes have been conducted in humans and various animal species, including the Agouti Signaling Protein (ASIP) gene [9,10,11,12,13], the Tyrosinase-Related Protein 1 (TYRP1) gene [9,14,15,16,17], the Tyrosinase (TYR) gene [17,18,19,20,21], the Melanophilin (MLPH) gene [22,23,24], and the Melanocortin 1 Receptor (MC1R) gene [9,10,15,25,26,27,28,29,30,31,32,33,34,35]. Coat color plays an important role in phenotypic authenticity and survival by contributing to camouflage, predator avoidance, and positioning within social groups, all of which are subject to natural selection. Additionally, coat color can influence mating behavior through mate preference, thereby affecting reproductive success [36,37,38,39,40].
The characteristics of coat color and patterns in cats are inherited from parents to offspring. The genes that control these phenotypes are currently being identified and studied, which could help explain the genetic inheritance of coat color. The study of these traits began in the early twentieth century [41], leading to the development of various genetic tests for coat color in domestic cats. Cat breed establishment was strongly focused on coat color and pattern traits more than utilitarian traits [42,43]. The point coloration pattern characteristic of the Siamese cat is one of the most prominent coat traits observed across various cat breeds. The Siamese allele has been identified in multiple breeds, including the Birman, Himalayan, Ragdoll, Siberian, and Tonkinese [18]. The ASIP gene, which influences whether a cat’s fur has a banded or solid appearance, maps to chromosome A3 [10]. TYRP1 gene on chromosome D4 controls black (B), chocolate (b), and cinnamon (light brown; bl) coat color [17]. It is completely dominant with 3 allelic series (B > b > bl). Chocolate (bb) alleles of TYRP1 were fixed in Havana Brown and Singapura, the same as light brown (cinnamon) (blbl) alleles were fixed in 6 breeds (Abyssinian, Devon Rex, Ocicat, Oriental, Somali, and Sphynx) [16]. The TYR gene on chromosome D1, involved in melanin synthesis, is a temperature-sensitive gene that controls point coloration on the extremities, such as the face, pinnae, paws, and tail [18]. The TYR variants cause an allelic series of C (full color) > cb (Burmese) ≥ cs (Siamese) > c, c2 (albinos) [17,18,19,21]. The C allele is the wild type; the cs allele makes the color point as found in Siamese, or known as Wichien Maat in Thailand. The Burmese allele is considered semi-dominant [44,45] or incomplete dominant to cs, resulting in a coat color phenotype that is intermediate between that of the Burmese and Siamese cats, as observed in the Tonkinese breed. The albino allele results in a cat with white fur, pink skin, and pinkish eyes. Mocha, the novel color variant (cm) recently found in cats in Thailand. The allele known as mocha is likely co-dominant to the Burmese allele and dominant to the Siamese allele [21]. However, full allelic combinations have been observed more. Nowadays, there are reports of the fixed Siamese allele (cscs) in 4 cat breeds (Siamese, Birman, Balinese, and Javanese), fixed Burmese alleles (cbcb) in 3 breeds (Burmese, European Burmese, and Singapura cats), and heterozygous (cscb) of these 2 alleles in Tonkinese cats [18]. The MLPH gene on chromosome C1 dilutes the coat color that affects both eumelanin and phaeomelanin pigment [22]. A one-base-pair deletion in exon 2 of the MLPH gene results in a premature stop codon, leading to the deletion of 11 amino acids and a shortened protein chain. The expression of the gene shows complete dominance, with D (full color) being dominant over d (dilute color). The MC1R gene is mapped to chromosome E2 in the domestic cat [10,11]. This gene is associated with felid melanism [12]. The variant in the MC1R gene linked to the amber color was identified in the Norwegian Forest Cat [24], while the variant associated with the russet color was found in the Burmese cat [35]. The Copal allele, a newly identified variant, was reported in the Kurilian Bobtail cat [46].
Based on current understanding of coat color genetics, genetic testing can be used to assist in selecting desired coat color traits. This is useful in breeding plans to determine the expected coat colors resulting from specific matings. Owing to the point markings that resemble the color points found in Siamese or Burmese cats. The hypothesis regarding the genetic inheritance that the gene may have descended from the same allelic series requires further investigation. The facial mask and darkened paws observed in the referred cat from the Thai Cat Conservation Center indicate the possible presence of the cb or cs alleles within the selectively bred population. Coat color gene identification has attracted the attention of Thai cat breeders and has been a key point of interest, serving as the first concern for action. If these alleles could be identified, the possibility of excluding cb or cs alleles from the selective breeding plan should be easier. Furthermore, screening for solid coat color and other recessive genes, such as the dilute color gene, was considered to be done simultaneously. Because the TYRP1 gene possesses various variant alleles, it was investigated as a candidate gene responsible for the chocolate coat color phenotype in the Suphalak cat. The existence of the dilute gene influences the shade of the chocolate coat color, leading to variation that may confound proper phenotypic classification. To identify desirable characteristics and avoid further distribution of the undesirable coat color alleles within the population, the objective of this study was to apply genetic testing focusing on specific traits like coat color genes for selective breeding in the Suphalak cat. Five coat color genes involving agouti (ASIP), brown (TYRP1), color point (Tyrosinase: TYR), dilute color (MLPH), and extension color (MC1R) were selected to investigate.
2. Materials and Methods
2.1. Cat Samples
Cat whole blood or buccal samples were obtained from 127 cats of the developing Suphalak cat breeds, with an emphasis on selecting cats with desirable coloring traits. Whole blood samples (0.5–1.0 mL) were collected from the cephalic vein of cats via venipuncture into EDTA tubes. Buccal cell samples were obtained using a sterile swab, which was rubbed against both inner cheek surfaces. The cotton tip of the swab was then placed into a 2 mL Eppendorf tube containing 1 mL of distilled water as a preservative medium [47]. Samples were stored at −20 °C until DNA extraction. Individual cat phenotypes were determined at the time of sample collection through visual examination or palpation, including evaluation of the tail by experienced Thai cat breeders. The cats exhibited characteristics similar to the Suphalak cat, including short hair, brown whiskers, brown/chocolate coat color with or without dark points at the extremities, and eyes ranging in color from yellow to amber or gold. The study included both male and female cats of all ages, representing all possible selective traits for developing the Suphalak cat. The sample composed of 61 cats (44 females and 17 males aged 1–4 years at the time of sample collection) from Thai cat conservation center at Samut Songkhram province date back to 2011-2018 and 66 cats from 5 Thai cat breeders from 2015-2020 (37 females and 29 males aged 4 months–3 years). Of these, 26 cats have unknown histories because they were adopted by the owners, while the remaining 101 cats were selected and bred by breeders. These cats were chosen based on their phenotype characteristics, with some having parents that resembled the Suphalak cat.
2.2. DNA Extraction
Genomic DNA was extracted from blood samples using the DNeasy® Blood Kit (Qiagen, USA). DNA from buccal swab samples was isolated using the EZNA® Tissue DNA Kit (Omega Bio-Tek, Inc., USA) according to the manufacturer’s instructions, with slight modifications [47]. The concentration and purity of the extracted DNA were assessed using a NanoDrop 2000™ spectrophotometer (Thermo Fisher Scientific, Wilmington, DE, USA). The DNA samples were subsequently used for genotyping of coat color genes.
2.3. Design Primer, Polymerase Chain Reaction, and DNA Sequencing
Fourteen sequence variants of 5 genes, including ASIP, TYRP1, TYR, MLPH, and MC1R loci, were selected to investigate (Table 1). The 2 bp deletion in exon 2 of ASIP causes the melanistic, non-agouti allele (a) [10]. Two SNPs in exon 2 (c.41G>C and c.142T>C) are associated with agouti alleles in the Asian Leopard Cat (ALC) (Prionailurus bengalensis) (APbe) [12]. The 2 SNPs of TYRP1 in exon 2 were identified in 3 allelic series (B > b > bl) [16,17]. The transversion from C to G at position 8 (c.8C>G) in exon 2 caused the amino acid change from alanine to glycine (A3G). The C to T transition at position 298 (c.298C>T) in exon 2 changed arginine to a stop codon (R100X) with light brown, chestnut, cinnamon, or fawn phenotype [16,17]. These 2 variants were correlated with the chocolate or cinnamon phenotype. The Colorpoint locus in the TYR gene was identified with 4 sequence variant targets. A sequence variant at position 679 (c.679G>T) in exon 1 leads to the substitution of glycine by tryptophan and is responsible for the Burmese coat color phenotype (cb) [17]. In exon 2, three sequence variants were investigated. The substitution of glycine with arginine at codon 301 (c.901G>A) results in the Siamese coat color (cs) [17]. A deletion of one cytosine at position 975 (975delC) is associated with the albino coat color (c) [19], and a recently identified variant, a 6-base pair deletion-insertion (6delinsAATCTC), results in the mocha coat color (cm) [21]. The Dilution locus (D, d) in the MLPH gene was identified with a sequence variance containing a T deletion at position 83 on exon 2 [22]. The extension locus (E, e) of the MC1R gene was investigated using four sequence variants: c.250G>A, associated with coat color changes from chocolate to amber; MC1R15del (Jaguar-Mel); MC1R24del (Jaguarundi-Dark); and c.439_441del, which causes melanism in jaguars, dark-brown coloration in jaguarundis, and russet coat color in Burmese cats.
Primers were designed to identify the sequence variances by using the nucleotide sequence from the domestic cat genome database with the Sequence Manipulation Suite program. The nucleotide sequences were aligned to identify the sequence variances with a program used to design primers. The major program of primer design was Primer 3 v.0.4.0, and the minor programs were GenScript Real-time PCR (TaqMan) Primer Design and PrimerQues. Subsequently, the primers were evaluated for specificity and efficiency using the BLAST software provided by the NIH NCBI (USA). The Polymerase Chain Reaction (PCR) reaction was performed with the primer in Table 1 following a protocol with pre-denaturation at 94 ºC for 3 minutes, followed by 35 cycles of denaturation at 94 ºC for 15 seconds, annealing at 57 ºC for 30 seconds, and extension at 72 ºC for 40 seconds. PCR products were run on a 1.5% agarose gel electrophoresis, and the sequence variant targets were verified through sequence analysis. In total, 127 cat DNA samples were analyzed to identify 14 sequence variants in 5 genes.
2.4. Coat Color Gene Evaluation
DNA samples from all 127cats were tested with primers for 5 genes at 14 sequence variants. PCR products were analyzed by the size of the DNA fragments on a 1.5% agarose gel. The samples were then sent for sequencing to decode the genotypes in each cat. The results were compared to reference data from two cats that had previously been tested for coat color genes from abroad. Sequence variants, allele frequencies, and genotypes were analyzed in 127 cats. Based on the results, the cats were grouped, and individuals suitable for use in breeding programs aimed at improving the Suphalak breed were selected.
2.5. The Use of Coat Color Gene for Selection and Breeding Test in Suphalak Cats
Genotype data were used to design a selective breeding program. The results from the 5 genes, including agouti, brown, colorpoint, dilute, and extension locus, were evaluated together. If consistent with the theoretical basis of coat color genetics, the ideal Suphalak cat exhibiting the desired coat color should possess the genotype aabbCCDDEE. The next step is to strategically pair these cats for a breeding test. Since the number of cats with the aabbCCDDEE genotype is limited, cats with phenotypes similar to or closely resembling the Suphalak cat, but with other genotypes, such as aabbCcsDDEE, aabbCcsDdEE, aabbCcbDDEE, aabbCcbDdEE, or aabbCCDdEE, may also be used for breeding. However, these cats carry other alleles, such as cs, cb, and d, which can be passed on to their offspring. Initially, these cats must undergo coat color gene testing, and the coat color genes in their offspring should be monitored to plan future breeding. Testing will be conducted by selecting 3 pairs of cats for breeding. The cats were paired for breeding, including both parents and offspring, and their phenotypes and genotypes were compared to ensure alignment. Additionally, the accuracy of the tested coat color genes was evaluated for their application in selection and breeding management.
2.6. Statistical Analysis
Allele frequencies, genotypes, and observed heterozygosity for the ASIP, TYRP1, TYR, MLPH, and MC1R genes in 127 cats were calculated using statistical analysis in POPGENE software version 3.2.
3. Results
3.1. Coat Color Genetic Results
All 127 cats (61 from the Thai Cat Conservation Center and 66 from Thai cat breeders) were homozygous for the 2-base pair deletion at c.123-124 in the ASIP gene. This deletion is functionally linked to the solid coat color, as it disrupts normal agouti signaling. The c.41G>C and c.142T>C mutations in exon 2, previously associated with the charcoal pattern in heterozygous (APbea), were not observed in these cats. In the TYRP1 gene, the majority of cats from the Thai Cat Conservation Center had the following genotypes: 31 cats were C/C (black), 22 were C/G (carriers of chocolate), and 8 were G/G (chocolate). In contrast, among cats from Thai cat breeders, the majority (62 cats) were G/G (chocolate), indicating selective breeding for the chocolate coat color. No cats carried the c.298C>T variant of the TYRP1 gene, indicating that the cinnamon allele is absent in both populations. The Burmese variant (c.679G>T) was found to be widespread in cats at the Thai Cat Conservation Center. Among 61 individuals, 58 were homozygous T/T, expressing the Burmese point phenotype, while 3 were compound heterozygotes (G/T). In contrast, the Siamese variant (c.901G>A) was detected at low frequency, with only three A/A (Siamese point) cats identified among cats from Thai breeders. Notably, the A/A genotype was absent in the Thai Cat Conservation Center population, where the Siamese allele appeared only in compound heterozygous form (G/A). As a result, three cats at the Conservation Center exhibited the Tonkinese point phenotype, carrying both the Burmese and Siamese alleles. No sequence variants associated with the albino or mocha alleles were found in either population. Carriers of the c.83delT variant in the MLPH gene were present in both populations, but only one homozygous dilute cat was identified among the Thai cat breeders, indicating that dilute coloration is rare, with dense pigmentation being predominant in both groups. No sequence variants were found in the MC1R gene, and no melanistic jaguar, dark-brown jaguarundi, amber, or russet coat traits were observed in either population (Table 2).
The allele frequency of the non-agouti allele was 100% in both the Thai Cat Conservation Center and among Thai cat breeders. Based on coat color genotype data from the Thai Cat Conservation Center breeding colony, allele frequencies at the TYRP1 locus were 68.85% for the black allele (B) and 31.15% for the chocolate allele (b). Among the 61 cats analyzed, 97.54% carried the cb allele, while 2.46% carried the cs allele. The wild-type C allele, which produces a full-color phenotype, was absent (0%). At the MLPH locus, which determines dilution, the allele frequency was 88.52% for the dense color allele and 11.48% for the dilute allele. In contrast, analysis of 66 samples from Thai cat breeders revealed an allele frequency of 2.27% for the black allele and 97.73% for the chocolate allele at the TYRP1 locus. For the TYR gene, allele frequencies were 56.82% for the C allele, 31.82% for the cb allele, and 11.36% for the cs allele. The frequency of the dense color allele was 90.15%, while the dilute allele occurred in 9.85% of the samples (Table 3).
The allele frequencies of the Burmese and Siamese alleles were high, along with a low allele frequency of the chocolate color allele, at the Thai Cat Conservation Center breeding colony. This pattern likely resulted from breeding selection that took place when the inheritance mechanisms of coat color genes were still not fully understood. In cats with the chocolate color genotype (bb), the chocolate color can reduce the visibility of the point coloration in Burmese, Tonkinese, or Siamese cats at the extremities. Cats with bbcbcb, bbcbcs, or bbcscs genotypes exhibit a fading of the point coloration, blending with the chocolate background color, compared to the sable or dark points seen in BBcbcb cats. This was the explanation of why these cats developed a Burmese appearance, despite being part of a strong selective breeding program for over five decades. The standout example was a male cat named Samsen, with the bbcbcsDD genotype. He was the only cat among those with dark-point phenotypes to exhibit a Tonkinese appearance. No cinnamon mutation (bl) was identified in any of these cats, indicating that this allele is either very rare in the Thai cat population or has not been a targeted trait in selective breeding programs. The majority of 127 cats have dense pigmentation (89.37%), meaning colors like black and chocolate are rich and not diluted into grey or lilac variants (Table 3).
Selective breeding by Thai cat breeders, based on an understanding of coat color genetic inheritance patterns, led to a marked reduction in the frequencies of the black, Burmese, and Siamese alleles compared to the high levels observed in the Thai Cat Conservation Center population. The high frequency of the b allele (97.73%) and the C allele (56.82%) among breeder cats indicates intentional selection for the chocolate coat color and full color expression (Table 3). Nevertheless, among the 127 cats examined, the cb and cs alleles remained present in heterozygous genotypes, with observed frequencies of Ccb at 23.62%, Ccs at 3.94%, cbcb at 50.39%, cbcs at 3.94%, and cscs at 2.36% (Table 4).
No genetic variation was detected in the ASIP and MC1R genes among all 127 cats. Therefore, the coat color genotypes were mainly determined based on three genes: TYRP1, TYR, and MLPH. Analysis of these genes in the 127 cats within the breeding program allowed classification into four distinct categories: 1) Burmese point that was the character of Burmese was 62 cats including 53 sable Burmese point cats (B_cbcbD_ ) and 9 chocolate Burmese cats (bbcbcbD_) 2) Tonkinese point (B_cbcsD_), this cat had the coat color between Burmese point and Siamese point was 7 cats (2 seal point Tonkinese and 5 chocolate point Tonkinese 3) Siamese point (B_cscsD_), this cat had point pattern as the Siamese cat. In this study, only two chocolate and one lilac Siamese cat were included. Additionally, 55 cats exhibited a full-body chocolate coloration and were classified as Suphalak (bbC_D_). As shown in Table 5, the ASIP and MC1R genes were excluded from the analysis and therefore are not presented in the table. Cats with Burmese point coloration were found in high numbers, especially at the Thai Cat Conservation Center. These cats exhibited dark points on the face and paws (Figure 2).
Among the 55 cats displaying a uniform chocolate coat color with no points (aabbC_D_EE), 18 cats had the genotype aabbCCDDEE, which is the desired genotype for the Suphalak cat, without any point coloration or hidden diluted fur. Of these, 7 were female and 11 were male. Among them, 7 cats (4 males and 3 females) had kinked tails. This left only 11 cats with the genotype aabbCCDDEE that conformed to the desired Suphalak cat traits and did not exhibit any other hidden characteristics. Additionally, 37 cats that exhibited coat characteristics similar to, or resembling, those of the Suphalak cat but carried hidden undesirable traits were identified with the following genotypes: 2 with aabbCCDdEE, 29 with aabbCcbDDEE, 1 with aabbCcbDdEE, 3 with aabbCcsDDEE, and 2 with aabbCcsDdEE. Although these cats displayed a solid chocolate coat color, they carried alleles that could produce point coloration at the extremities, specifically, the cb and cs alleles. The d allele, which causes coat color dilution, was also present in some individuals.
3.2. Application of the Coat Color Genes for Selective Breeding
Breeding tests were conducted using three Suphalak cat pedigrees, as illustrated in Figure 3. In Pedigree 1, a male cat with the genotype aabbCCDDEE was bred with a female cat possessing the genotype aabbCcbDDEE. The Suphalak cat parents transmitted the solid chocolate coat color phenotype, without point markings, to all four of their offspring (3 males and one female). The cb allele in the dam was passed to one of the male offspring. However, this cb allele does not show marking points in this male offspring. This suggested that the cb allele behaved as a recessive allele for markings and does not display marking points when inherited in a heterozygous form. However, this male offspring will transfer the cb allele to 50% of their offspring. Solid chocolate to solid chocolate parents were bred together. These cats will pass on the bb genotype for coat color, ensuring that the offspring all display solid chocolate coats.
Pedigree 2, the inheritance patterns between the Sire (genotype aabbCCDDEE) and the dam (genotype aabbcbcbDDEE), focusing on how their offspring will inherit the traits. Sire had a solid chocolate coat color. The dam’s appearance was chocolate color with a marking point on its extremity because she carried the cbcb genotype. All offspring will inherit one C allele from the sire and one cb allele from the dam, resulting in the solid color (Ccb genotype) for all offspring (2 males and one female). However, all of the offspring will carry the cb allele, which could be passed down to future generations. Pedigree 3, Male cat genotype aabbCCDDEE was bred with female genotype aabbCCDDEE. The male offspring was aabbCCDDEE genotype. Solid chocolate to solid chocolate Suphalak cat breeding produced all solid chocolate Suphalak cat phenotype without any carrier of the allele.
The genetic analysis of 127 cats revealed that a cat with aabbCCDDEE genotype was solid chocolate, aabbCcbDDEE was solid chocolate and a carrier of Burmese point. A cat with aabbcbcbDDEE genotype was solid chocolate with a Burmese point. Moreover, the cat with the cbcb, cbcs, cscs genotype, no matter the genotype of chocolate (BB, Bb, bb) and dilute (DD, Dd, dd) genes were expressed the dark point appearance on the extremities, especially on the face, paws, tail, ears, and genital area. Genetic inheritance of the coat color gene in Suphalak followed the genetic theory of coat color in cats. Therefore, the presence of dark points in Suphalak cats can be identified through genetic testing. Based on this pattern of genetic inheritance, mating between cats with cbcb and CC genotypes could also produce offspring exhibiting the Suphalak phenotype.
Mating chocolate cats with genotype aabbCCDDEE, aabbCcbDDEE could get male and female kittens with chocolate color that followed the hypothesis. Moreover, sire or dam with genotype aabbcbcbDDEE mating with genotype aabbCCDDEE could have Suphalak kittens as well. The results of the breeding test indicate that coat color genetic testing can be effectively applied for selective breeding in Suphalak cats. The genotypic, phenotypic, and excluding recessive phenotypes could identify the first and second categories of 18 and 37 cats with genotype aabbCCDDEE and aabbC_D_EE, respectively. Individuals possessing the aabbCCDDEE genotype exhibit the ideal Suphalak phenotype, characterized by a uniform solid chocolate coat, chocolate-colored nose leather, and brown whiskers. Ocular pigmentation ranges from amber to intense gold, and the footpads exhibit a distinct warm cinnamon coloration (Figure 4).
The important problem of breed improvement was the limited number of founding cats. In this study, 26 cats were without pedigree records, as their genetic backgrounds were unknown. These cats are at risk of carrying undesirable traits such as tabby markings, white spotting, or kinked tails. Therefore, genetic testing for coat color is essential for screening cats prior to breeding. While the five analyzed genes (ASIP, TYR, TYRP1, MLPH, and MC1R) may be sufficient for preliminary selection, the inclusion of additional coat color genes is recommended to further enhance and expedite the genetic improvement of the Suphalak cat.
Cats in the pedigrees were screened for kinked tails using radiographic examination or direct palpation. An example of the segregation pattern of the kinked tail trait within a family is shown in Figure 5. Pedigree analysis suggests that the kinked tail follows an autosomal dominant mode of inheritance, indicating that only one copy of the mutant allele is sufficient for the kinked phenotype to be expressed. However, this conclusion requires further investigation. In some cases, palpation was challenging due to minor tail abnormalities that could only be detected by experienced breeders. In Figure 6, the kinked tail is not phenotypically visible (Figure 6b and 6c) and is also difficult to detect by palpation.
4. Discussion
All five remaining traditional Thai cat breeds exhibit solid coat colors, a trait that is relatively rare in the general cat population of Thailand. The development of these solid-colored breeds likely required intensive selective breeding to eliminate the agouti allele. The observation of a 100% frequency of the non-agouti allele in the Suphalak cat further supports the existence of a strong and distinct genetic lineage. Notably, the 2-bp deletion at nucleotide positions 123–124 in the ASIP gene, which disrupts agouti signaling and leads to a solid coat color phenotype, appears to be unique to traditional Thai cats and may serve as a defining genetic marker for these breeds. The leopard cat allele was not observed segregating in traditional Thai Suphalak cat, indicating the absence of wild felid introgression in its genetic lineage. This contrasts with hybrid breeds such as the Bengal cat, which were selectively bred to incorporate Burmese and Siamese alleles influencing coat coloration. In Bengals, the compound heterozygous state of the ASIP gene (APbea) is associated with the melanistic “charcoal” phenotype, characterized by a darker facial mask and a distinct dark dorsal stripe [12]. In other species, such as mice, rats, and horses [9,48,49], loss-of-function mutations in the ASIP gene similarly result in extreme melanism, highlighting the conserved role of this gene in pigmentation across mammals. The agouti allele is dominant over the recessive non-agouti allele responsible for solid coat color. Therefore, in the absence of human selection, as in feral cat populations, most cats exhibit a tabby pattern, although some may carry the gene for solid color. When point colored cats mated with the rare solid cats, offspring with a solid coat and color-point pattern were eventually produced. However, these feral cats were not considered special by Thai people and were allowed to breed freely without selective pressure. As a result, the presence of Siamese or Burmese alleles remains evident in the point pattern phenotypes observed in the local cat population.
Brown color in domestic cats has been recognized for more than 100 years [50]. The ancient cat breeds, such as Siamese and Abyssinians were carried the allelic series of the brown color variant [16]. The high cb and cs alleles could suggest the existence of the Siamese breed during the breeding selection of the Suphalak cat. In contrast, the dilute allele (d) had a low frequency (< 10%), meaning that the blue Korat cat was not the best choice for breeders to develop the Suphalak cat. The allele frequencies and genotypes of these 5 genes indicated a strong relationship between Suphalak and albino gene series, especially the Burmese allele. All cats from the Thai Cat Conservation Center had color point alleles (97.54% cb, 2.46% cs). The optical illusion of the thermosensitive Burmese allele (cb) produced a sable or brown appearance [16]. This allele seems to create the illusion of a black cat, with the point allele appearing nearly chocolate in color. Discrepancies between genetic and visual coat color. There is no mention of the Burmese cat in any known ancient Thai manuscripts. Nonetheless, the result of high Burmese (cb) allele frequency could imply the existence of this allele among Traditional Thai cats for a long period. It is reasonable to assume that the color point variant has long been in the Thai native cat lineage. The first Burmese cat imported into the United States in 1930 was notably a female cat, and this cat had a heterozygous genotype combining both Burmese and Siamese alleles [44]. The presence of the Siamese allele in the Burmese suggests a shared genetic heritage or at least a historical connection between the two breeds. In Thailand, the Burmese allele has been found in both feral cats and those in the Cat Conservation Center. It seems likely that the Burmese gene, or at least a version of it, was indeed present in the region of Thailand well before the 19th century. The high frequency of the Burmese allele in these populations suggests that the gene might have been circulating in the region for centuries. The discovery of Burmese appearance in feral cats further supports the idea that these traits were present in Thailand well before the breed’s modern development in the West.
In this study, there was no evidence of the albino and mocha coat color alleles of the TYR gene in any of the cats. A blue-eyed or odd-eyed white coat was observed in one traditional Thai cat, known as the Khao Manee. The albino allele in the TYR gene is associated with a pink-eyed white coat. Mocha is a recently identified coat color, first discovered in Thailand. Sequence variants associated with the charcoal or russet colors at the MC1R gene were also not found. The russet color was first identified in Burmese cats in New Zealand [35]. This suggests that the mutation responsible for the russet color has likely never existed in cats with a Burmese appearance in Thailand. Overall, the absence of any sequence variants related to these alleles in the 127 cats analyzed suggests that they may not exist in the Thai cat population. However, further investigation is needed to confirm this finding. Evidence suggests that the albino gene series may have ancient origins in Thailand, as indicated by descriptions of Siamese cats found in Thai manuscripts dating back approximately 600 years. Point-patterned cats, both purebred and feral, have been found in greater numbers in Thailand than in other countries. The Siamese and Burmese mutation was likely to be identical by descent from the traditional Thai cat rather than being a new mutation. These traits were present in the region long before the Siamese cat gained international recognition in Western countries in the 19th century.
The analysis of coat color genes has deepened the understanding of genetic inheritance in Suphalak cats. Due to the fixation of the ASIP and MC1R genes, Suphalak cats consistently exhibit a solid coat color. As a result, a completely solid chocolate-colored Suphalak can present the same phenotype (appearance) despite having any of the following three underlying genotypes: chocolate (bb genotype), color point (CC or Ccb, Ccs genotype), and dilution (DD or Dd genotype). A cat with the aabbCCDDEE genotype was considered the ideal choice for selective breeding. However, cats with the aabbCcbDDEE genotype should be selected with caution. Offspring should be screened to exclude the cb, cs, or d alleles. In the chocolate locus, the B allele is dominant over the b allele, so BB genotype cats would produce 100% black cats, making them the least desirable for breeding.
During the ongoing breed development, cats that closely resembled Suphalak cats in appearance were included in the breeding program due to the limited number of founder cats, some of which had an unknown genetic background. Without genetic screening, relying solely on coat color criteria could introduce new undesirable traits to the population, such as kinked tails or white spotting. Therefore, other genes, such as those affecting the kinked tail, were further considered for improving the breeding process. While palpation is a commonly used method in field conditions or by breeders, its accuracy can be limited, particularly when abnormalities are subtle. In some cases, mild kinks may not be externally visible and can be easily overlooked, especially by untrained observers. Experienced breeders may be able to detect these subtle deformities through careful palpation; however, even then, there is potential for misclassification. This highlights the importance of radiographic confirmation in cases where the presence of kinked tails is uncertain. The findings suggest that relying solely on visual inspection or palpation may underestimate the true prevalence of kinked tails in a population, especially when the phenotype is mild. These observations have implications for pedigree analysis and inheritance studies. If mild cases are missed or inaccurately scored, this could obscure the true inheritance pattern of the trait. Therefore, combining palpation with radiographic imaging provides a more reliable method for identifying kinked tail phenotypes, particularly when assessing the genetic basis of the trait in breeding programs or conservation colonies.
The kinked tail also occurred in the Burmese cat during breed development, and at that time, the kinked tail had not been bred out of the Burmese stock [44]. The tail of a traditional Thai cat is an important feature in its breed standard. A good-quality traditional Thai cat is typically characterized by a long tail that tapers to a point without any kinks, particularly in the effort to restore the breed to its historical standard as described in ancient Thai manuscripts. The tail’s appearance is part of the overall symmetry and elegance that the breed is known for. From the breeder’s perspective, the presence of kinked tails in these breeding selections resulted from the limited number of founder cats. Although the kinked tail does not affect the health or function of the cat, it has often been tolerated in local cat populations. However, in the context of breed standardization, its elimination is essential for the Suphalak to meet established breed standard criteria. Outcrossing to develop new cat breeds has also been practiced in other breeds, such as the Selkirk Rex. This strategy can enhance genetic diversity; however, it also carries the risk of introducing undesirable traits, including heritable diseases [51]. Consequently, the use of unknown or genetically unrelated cats in breed development requires careful evaluation to avoid the inadvertent incorporation of deleterious alleles. Breeders should be fully aware of the genetic implications of their mating decisions.
5. Conclusions
Over the past several decades, Thai cat breeders have encountered numerous challenges in the development of the Suphalak cat. Solid chocolate cats were very rare and difficult to find in general. Most Thai people overlooked or were unaware of the value of these cats unless they were breeders. As a result, these remaining cats were not selected for breeding programs. Due to the recessive expression of the chocolate allele, combined with the illusion effect of the point alleles, these cats were often obscured in randomly bred cats without careful observation. After gaining a deeper understanding of coat color genetic inheritance, genetic testing became essential in supporting the breeding plan to revive the Suphalak cat. In 2024, the Suphalak cat is officially recognized as a purebred breed by the World Cat Federation (WCF) [52], following its successful development through selective breeding. The breed standard defines the coat color as warm chocolate. The nose leather is chocolate brown, the whiskers are brown, and the paw pads are a warm cinnamon color. The eye color ranges from golden yellow to intense gold.
Author Contributions
Conceptualization, J.P.; methodology, J.P., S.L., G.N., K.K., S.P.; formal analysis, J.P., S.P.; investigation, J.P.; resources, J.P, K.K.; data curation, J.P., S.P.; writing—original draft preparation, J.P.; writing—review and editing, J.P., S.L., G.N., K.K., and S.P. All authors have read and agreed to the published version of the manuscript
Funding
This research was financially supported by the Faculty of Veterinary Medicine, Kasetsart University (FFK).
Institutional Review Board Statement
The animal study protocol was submitted and approved by the Kasetsart University Institutional Animal Care and Use Committee, Faculty of Veterinary Medicine, Kasetsart University (protocol no. ACKU 01660).
Informed Consent Statement
Not applicable
Data Availability Statement
Data are contained within the article. Further inquiries can be directed to the corresponding author.
Acknowledgments
The authors gratefully acknowledge Kasetsart University for their support and data contribution to this study. Financial support was provided by the Faculty of Veterinary Medicine, Kasetsart University. We also sincerely appreciate the valuable data and assistance provided by the Thai Cat Conservation Center, the International Maew Boran Association (TIMBA), the Vadhana Cattery, the Nong Khaem Thai Cat House, and Thai cat breeders.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ASIP | Agouti Signaling Protein |
| DNA | Deoxyribonucleic Acid |
| EDTA | Ethylenediaminetetraacetic Acid |
| TYRP1 | Tyrosinase-Related Protein 1 |
| TYR | Tyrosinase |
| MLPH | Melanophilin |
| MC1R | Melanocortin 1 Receptor |
| PCR | Polymerase Chain Reaction |
| ºC | Degrees Celsius |
References
- Clutterbuck, M.R. Siamese Cats: Legends and Reality; White Lotus Co., Ltd: Bangkok, Thailand, 2004; pp. 1–244. [Google Scholar]
- Vella, C.M.; Shelton, L.M.; McGonagle, J.J.; Stanglein, T.W. Robinson’s Genetics for Cat Breeders and Veterinarians, 4th ed.; Butterworth-Heinemann: Oxford, UK, 2005; pp. 1–496. [Google Scholar]
- Ubolrat, K.; Sudtisa, L.; Kavil, N.; Janjira, P. Genetic diversity and inbreeding situation of Korat and Siamese cats based on microsatellite markers. Vet. Integr. Sci. 2019, 17, 51–64. [Google Scholar]
- Barsh, G. S. The genetics of pigmentation: from fancy genes to complex traits. Trends Genet. 1996, 12, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Cieslak, M.; Reissmann, M.; Hofreiter, M.; Ludwig, A. Colours of domestication. Biol. Rev. Camb. Philos. Soc. 2011, 86, 885–899. [Google Scholar] [CrossRef] [PubMed]
- Hearing, V.J.; Tsukamoto, K. Enzymatic control of pigmentation in mammals. FASEB J. 1991, 5, 2902–2909. [Google Scholar] [CrossRef] [PubMed]
- Jackson, I.J. Colour-coded switches. Nature. 1993, 362, 587–588. [Google Scholar] [CrossRef]
- Liu, X.; Yongdong, P.; Xinhao, Z.; Xinrui, W.; Wenting, C.; Xiyan, K.; Huili, L.; Wei, R.; Muhammad, Z.K.; Changfa, W. Coloration in Equine: Overview of Candidate Genes Associated with Coat Color Phenotypes. Animals 2024, 14, 1802. [Google Scholar] [CrossRef]
- Rieder, S.; Taourit, S.; Mariat, D.; Langlois, B.; Guerin, G. Mutations in the agouti (ASIP), the extension (MC1R), and the brown (TYRP1) loci and their association to coat color phenotypes in horses (Equus caballus). Mamm. Genome 2001, 12, 450–455. [Google Scholar] [CrossRef]
- Eizirik, E.; Yuhki, N.; Johnson, W.E.; Menotti-Raymond, M.; Hannah, S.S.; O’Brien, S.J. Molecular genetics and evolution of melanism in the cat family. Curr. Biol. 2003, 13, 448–453. [Google Scholar] [CrossRef]
- Schneider, A.; David, V.A.; Johnson, W.E.; O’Brien, S.J.; Barsh, G.S.; Menotti-Raymond, M.; Eizirik, E. How the leopard hides its spots: ASIP mutations and melanism in wild cats. PLoS ONE 2012, 7, e50386. [Google Scholar] [CrossRef]
- Gershony, L.C.; Penedo, M.C.; Davis, B.W.; Murphy, W.J.; Helps, C.R.; Lyons, L.A. Who’s behind that mask and cape? The Asian leopard cat’s Agouti (ASIP) allele likely affects coat colour phenotype in the Bengal cat breed. Anim. Genet. 2014, 45, 893–897. [Google Scholar] [CrossRef]
- Sun, T.; Li, S.; Xia, X.; Ji, C.; Zhang, G.; Yu, J.; Jiang, G.; Dang, R.; Lei, C. ASIP gene variation in Chinese donkeys. Anim. Genet. 2017, 48, 372–373. [Google Scholar] [CrossRef]
- Javerzat, S. , Jackson, I.J. White-based brown (Tyrp1B-w) is a dominant mutation causing reduced hair pigmentation owing to a chromosomal inversion. Mamm. Genome. 1998, 9, 469–71. [Google Scholar] [CrossRef] [PubMed]
- Schmutz, S.M.; Berryere, T.G.; Goldfinch, A.D. TYRP1 and MC1R genotypes and their effects on coat color in dogs. Mamm. Genome. 2002, 13, 380–387. [Google Scholar] [CrossRef]
- Lyons, L.A.; Foe, I.T.; Rah, H.C.; Grahn, R.A. Chocolate-coated cats: TYRP1 mutations for brown color in domestic cats. Mamm. Genome. 2005, 16, 356–366. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Kuntzel, A.; Eizirik, E.; O’Brien, S.J.; Menotti-Raymond, M. Tyrosinase and tyrosinase related protein 1 alleles specify domestic cat coat color phenotypes of the albino and brown loci. J. Hered. 2005, 96, 289–301. [Google Scholar] [CrossRef] [PubMed]
- Lyons, L.A.; Imes, D.L.; Rah, H.C.; Grahn, R.A. Tyrosinase mutations associated with Siamese and Burmese patterns in the domestic cat (Felis catus). Anim. Genet. 2005, 36, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Imes, D.L.; Geary, L.A.; Grahn, R.A.; Lyons, L.A. Albinism in the domestic cat (Felis catus) is associated with a tyrosinase (TYR) mutation. Anim. Genet. 2006, 37, 175–178. [Google Scholar] [CrossRef]
- Utzeri, V.J.; Bertolini, F.; Ribani, A.; Schiavo, G.; Dall’Olio, S.; Fontanesi, L. The albinism of the feral Asinara white donkeys (Equus asinus) is determined by a missense mutation in a highly conserved position of the tyrosinase (TYR) gene deduced protein. Anim. Genet. 2016, 47, 120–124. [Google Scholar] [CrossRef]
- Yu, Y.; Grahn, R.A.; Lyons, L.A. Mocha tyrosinase variant: a new flavour of cat coat coloration. Anim. Genet. 2019, 50, 182–186. [Google Scholar] [CrossRef]
- Ishida, Y.; David, V.A.; Eizirik, E.; Schaffer, A.A.; Neelam, B.A.; Roelke, M.E.; Hannah, S.S.; O’Brien, S.J.; Menotti-Raymond, M. A homozygous single-base deletion in MLPH causes the dilute coat color phenotype in the domestic cat. Genomics 2006, 88, 698–705. [Google Scholar] [CrossRef]
- Drögemüller, C.; Ute, P.; Bianca, H.; Anne-Rose, G.-A.; Tosso, L. A Noncoding Melanophilin Gene (MLPH) SNP at the Splice Donor of Exon 1 Represents a Candidate Causal Mutation for Coat Color Dilution in Dogs. J. Hered. 2007, 98, 468–473. [Google Scholar] [CrossRef] [PubMed]
- Peterschmitt, M.; Grain, F.; Arnaud, B.; Deleage, G.; Lambert, V. Mutation in the melanocortin 1 receptor is associated with amber colour in the Norwegian Forest Cat. Anim. Genet. 2009, 40, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Robbins, L.; Nadeau, J.; Johnson, K.R.; Kelly, M.; Roselli-Rehfuss, L.; Baack, E. Pigmentation phenotypes of variant extension locus alleles result from point mutations that alter MSH receptor function. Cell 1993, 72, 827–834. [Google Scholar] [CrossRef]
- Valverde, P.; Healy, E.; Jackson, I.; Rees, J.; Thody, A. Variants of the melanocyte-stimulating hormone receptor gene are associated with red hair and fair skin in humans. Nat. Genet. 1995, 11, 328–330. [Google Scholar] [CrossRef]
- Joerg, H.; Fries, H.; Meijerink, E.; Stranzinger, G. Red coat color in Holstein cattle is associated with a deletion in the MSHR gene. Mamm. Genome 1996, 7, 317–318. [Google Scholar] [CrossRef]
- Marklund, L.; Moller, M.; Sandberg, K.; Andersson, L. A missense mutation in the gene for melanocyte-stimulating hormone receptor (MC1R) is associated with the chestnut coat color in horses. Mamm. Genome 1996, 7, 895–899. [Google Scholar] [CrossRef]
- Våge, D.I.; Lu, D.; Klungland, H.; Lien, S.; Adalsteinsson, S.; Cone, R.D. A non-epistatic interaction of agouti and extension in the fox, Vulpes vulpes. Nat. Genet. 1997, 15, 311–315. [Google Scholar] [CrossRef]
- Kijas, J.M.H.; Wales, R.; Törnsten, A.; Chardon, P.; Moller, M.; Andersson, L. Melanocortin receptor 1 (MC1R) mutations and coat color in pigs. Genetics. 1998, 150, 1177–1185. [Google Scholar] [CrossRef]
- Våge, D.I.; Klungland, H.; Dongsi, L.; Cone, R.D. Molecular and pharmacological characterization of dominant black coat color in sheep. Mamm. Genome 1999, 10, 39–43. [Google Scholar] [CrossRef]
- Everts, R.E.; Rothuizen, J.; Van Oost, B.A. Identification of a premature stop codon in the melanocyte-stimulating hormone receptor gene (MC1R) in Labrador and Golden retrievers with yellow coat colour. Anim. Genet. 2000, 31, 194–199. [Google Scholar] [CrossRef]
- Rouzaud, F.; Martin, J.; Gallet, P.F.; Delourme, D.; Goulemont-Leger, V.; Amigues, Y.; Ménissier, F.; Levéziel, H.; Julien, R.; Oulmouden, A. A first genotyping assay of French cattle breeds based on a new allele of the extension gene encoding the melanocortin-1 receptor (MC1R). Genet. Sel. Evol. 2000, 32, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Abitbol, M.; Legrand, R.; Tiret, L.A. Missense mutation in melanocortin 1 receptor is associated with the red coat color in donkeys. Anim. Genet. 2014, 45, 878–880. [Google Scholar] [CrossRef] [PubMed]
- Gustafson, N. A.; Gandolfi, B.; Lyons, L. A. Not another type of potato: MC1R and the russet coloration of Burmese cats. Anim. Genet. 2016, 47, 704–705. [Google Scholar]
- Becerril, C.M.; Wilcox, C.J.; Lawlor, T.J.; Wiggans, G.R.; Webb, D.W. Effects of percentage of white coat color on Holstein production and reproduction in a subtropical climate. J. Dairy Sci. 1993, 76, 2286–2291. [Google Scholar] [CrossRef]
- Johansson, A.; Pielberg, G.; Andersson, L.; Edfors – Lilja, I. Polymorphism at the porcine dominant white/KIT locus influences coat color and peripheral blood cell measures. Anim. Genet. 2005, 36, 288–296. [Google Scholar] [CrossRef]
- Charon, K.M.; Lipka, K.R. The effect of a coat colour-associated gene polymorphism on animal health review. Ann. Anim. Sci. 2015, 15, 3–17. [Google Scholar] [CrossRef]
- Kennah, J.L.; Peers, M.J.L.; Wal, E.V.; Majchrzak, Y.N.; Menzies, A.K.; Studd, E.K.; Boonstra, R.; Humphries, M.M.; Jung, T.S.; Kenney, A.J.; Krebs, C.J.; Boutin, S. Coat color mismatch improves survival of a keystone boreal herbivore: Energetic advantages exceed lost camouflage. Ecology 2023, 104, e3882. [Google Scholar] [CrossRef]
- Oli, M.K.; Kenney, A.J.; Boonstra, R.; Boutin, S.; Murray, D.L.; Peers, M.J.L.; Gilbert, B.S.; Jung, T.S.; Chaudhary, V.; Hines, J.E.; Krebs, C.J. Does coat colour influence survival? A test in a cyclic population of snowshoe hares. Proc. R. Soc. B Biol. Sci. 2023, 290, 20221421. [Google Scholar] [CrossRef]
- Little, C.C. Color inheritance in cats, with special reference to the colors black, yellow, and tortoise-shell. Genetics. 1919, 8, 279–290. [Google Scholar]
- Kurushima, J.D.; Lipinski, M.J.; Gandolfi, B.; Froenicke, L.; Grahn, J.C.; Grahn, R.A.; Lyons, L.A. Variation of cats under domestication: genetic assignment of domestic cats to breeds and worldwide random-bred populations. Anim Genet. 2012, 44, 311–24. [Google Scholar] [CrossRef]
- Montague, M.J.; Li, G.; Gandolfi, B.; Khan, R.; Aken, B.L.; Searle, S.M.J.; Warren, W.C. Comparative analysis of the domestic cat genome reveals genetic signatures underlying feline biology and domestication. Proc. Natl. Acad. Sci. 2014, 111, 17230–17235. [Google Scholar] [CrossRef]
- Thompson, J.C.; Virginia, C.C.; Clyde, E.K.; Madeleine, D. Genetics of the Burmese cat. J. Hered. 1943, 34, 119–123. [Google Scholar] [CrossRef]
- Weir, H. Siamese cats. In: Our cats. In Our cats and all about them, Weir, H. ed.; Boston and New York: Houghton, Mifflin, 1889; pp. 73–79. [Google Scholar]
- Abitbol, M.; Gache, V. Copal, a new MC1R allele in the domestic cat. Anim. Genet. 2019, 50, 550–554. [Google Scholar] [CrossRef]
- Phavaphutanon, J.; Laopium, S.; Nanklang, K.; Sirinarumitr, K.; Kornkaewrat, K.; Pinyopummin, A.; Viriyarumpa, J.; Suthanmapinunt, P.; Vorawattanatham, N. Buccal Swab as a Source of Noninvasive Technique for Genomic DNA Collection in Felidae. Thai J. Vet. Med. 2013, 43, 417–422. [Google Scholar] [CrossRef]
- Hustad, C.M.; Perry, W.L.; Siracusa, L.D.; Rasberry, C.; Cobb, L.; Cattanach, B.M.; Kovatch, R.; Copeland, N.G.; Jenkins, N.A. Molecular genetic characterization of six recessive viable alleles of the mouse agouti locus. Genetics. 1995, 140, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Kuramoto, T.; Nomoto, T.; Sugimura, T.; Ushijima, T. Cloning of the rat agouti gene and identification of the rat nonagouti mutation. Mamm. Genome 2001, 12, 469–471. [Google Scholar] [CrossRef] [PubMed]
- CFA. The Cat Fanciers’ Association Cat Encyclopedia; Simon & Schuster, USA: New York, 1993; pp. 128–136. [Google Scholar]
- Serina, F.; Hasan, A.; Barbara, G.; Jennifer, D.K.; Alejandro, C.; Christine, V.; Leslie, A. L.; Gottfried, B. Selkirk Rex: Morphological and genetic characterization of a new cat breed. J. Hered. 2012, 103, 727–733. [Google Scholar] [CrossRef]
- WCF. World Cat Federation. Available online: http://wcf.de/en (accessed on 18 July 2025).
Figure 1.
A Suphalak cat from the Thai Cat Conservation Center. The text in the image indicates the name in Thai as “Suphalak/Thongdang” (meaning copper or red gold), exhibiting a chocolate or copper-colored coat with a facial mask and darkened coloration on the paws, which deviates from the traditional standard of a uniformly solid coat described in historical records.
Figure 1.
A Suphalak cat from the Thai Cat Conservation Center. The text in the image indicates the name in Thai as “Suphalak/Thongdang” (meaning copper or red gold), exhibiting a chocolate or copper-colored coat with a facial mask and darkened coloration on the paws, which deviates from the traditional standard of a uniformly solid coat described in historical records.

Figure 2.
Coat color in cats with color point patterns. (a) and (b) depict cats with a Burmese coat color pattern, showing dark pigmentation concentrated at the extremities (ears, face, and paws); (c) shows a cat with a Burmese point pattern that carries the dilute allele. Genotypes associated with coat color are displayed below each image.
Figure 2.
Coat color in cats with color point patterns. (a) and (b) depict cats with a Burmese coat color pattern, showing dark pigmentation concentrated at the extremities (ears, face, and paws); (c) shows a cat with a Burmese point pattern that carries the dilute allele. Genotypes associated with coat color are displayed below each image.

Figure 3.
Three independent pedigrees demonstrating coat color genetic inheritance involving the ASIP, TYRP1, TYR, MLPH, and MC1R genes, showing segregation of phenotypes across three different genotypes. (a) pedigree 1; (b) pedigree 2; (c) pedigree 3. Square: male, circle: female, open symbol: chocolate coat color, open symbol with thick margin: chocolate coat color with dark point at the extremity. Coded genotypes are shown below the diagram.
Figure 3.
Three independent pedigrees demonstrating coat color genetic inheritance involving the ASIP, TYRP1, TYR, MLPH, and MC1R genes, showing segregation of phenotypes across three different genotypes. (a) pedigree 1; (b) pedigree 2; (c) pedigree 3. Square: male, circle: female, open symbol: chocolate coat color, open symbol with thick margin: chocolate coat color with dark point at the extremity. Coded genotypes are shown below the diagram.
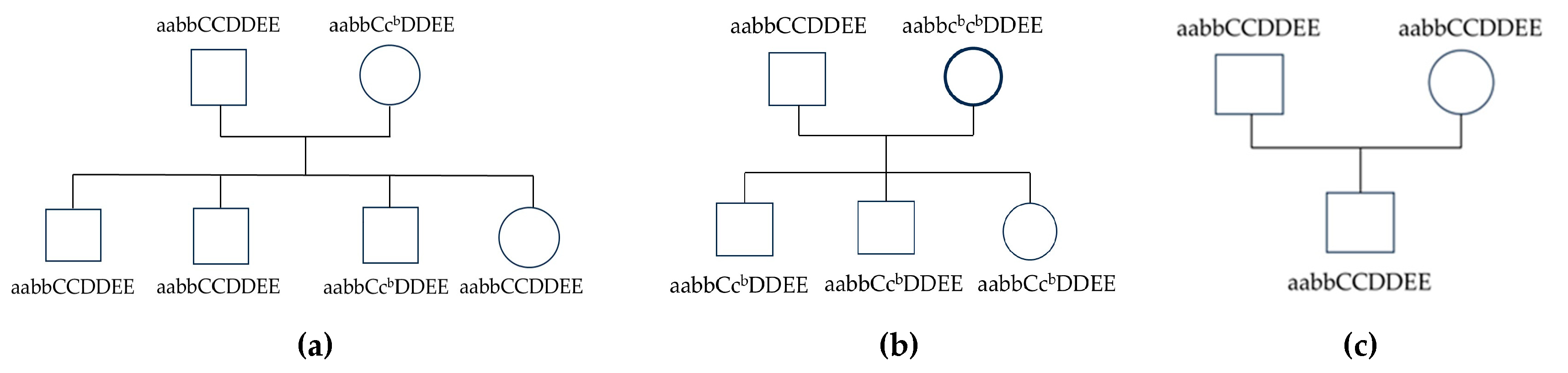
Figure 4.
The ideal Suphalak cat phenotype: (a) Solid chocolate coat color; nose leather is chocolate, and whiskers are brown; (b) Amber to intense gold eye color; (c) the paw pad exhibits a warm cinnamon color.
Figure 4.
The ideal Suphalak cat phenotype: (a) Solid chocolate coat color; nose leather is chocolate, and whiskers are brown; (b) Amber to intense gold eye color; (c) the paw pad exhibits a warm cinnamon color.

Figure 5.
Segregation of kinked tail phenotype in the Suphalak pedigree. Squares represent males; circles represent females. Open symbols indicate cats with a chocolate coat color, while open symbols with thick borders represent chocolate coat color with dark points at the extremities. Genotypes are shown below each symbol; however, ASIP and MC1R genotypes have been omitted. Asterisks (*) denote cats with kinked tails.
Figure 5.
Segregation of kinked tail phenotype in the Suphalak pedigree. Squares represent males; circles represent females. Open symbols indicate cats with a chocolate coat color, while open symbols with thick borders represent chocolate coat color with dark points at the extremities. Genotypes are shown below each symbol; however, ASIP and MC1R genotypes have been omitted. Asterisks (*) denote cats with kinked tails.
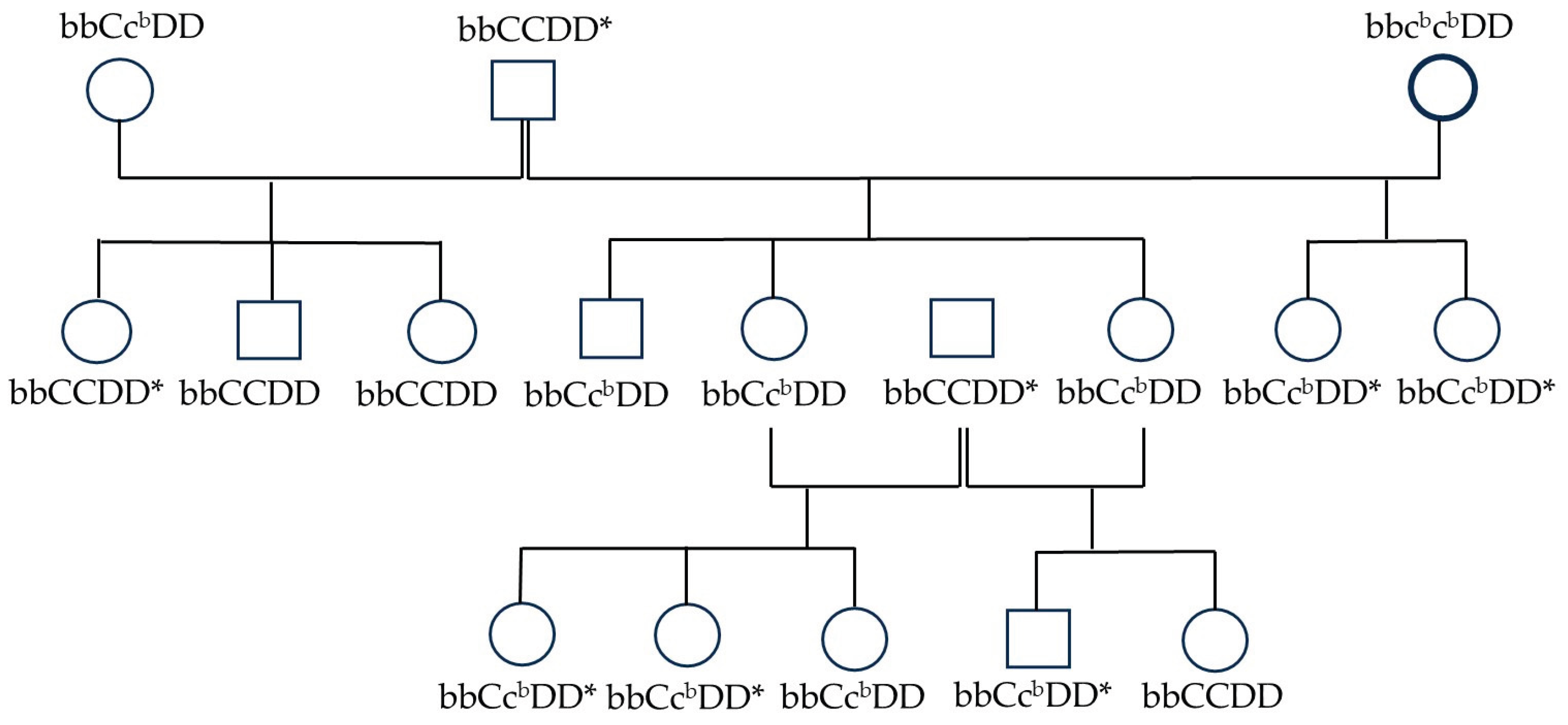
Figure 6.
Radiograph of the Suphalak cat tail (a) normal tail; (b) kinked tail; (c) kinked tail at the tip of the caudal vertebrae.
Figure 6.
Radiograph of the Suphalak cat tail (a) normal tail; (b) kinked tail; (c) kinked tail at the tip of the caudal vertebrae.
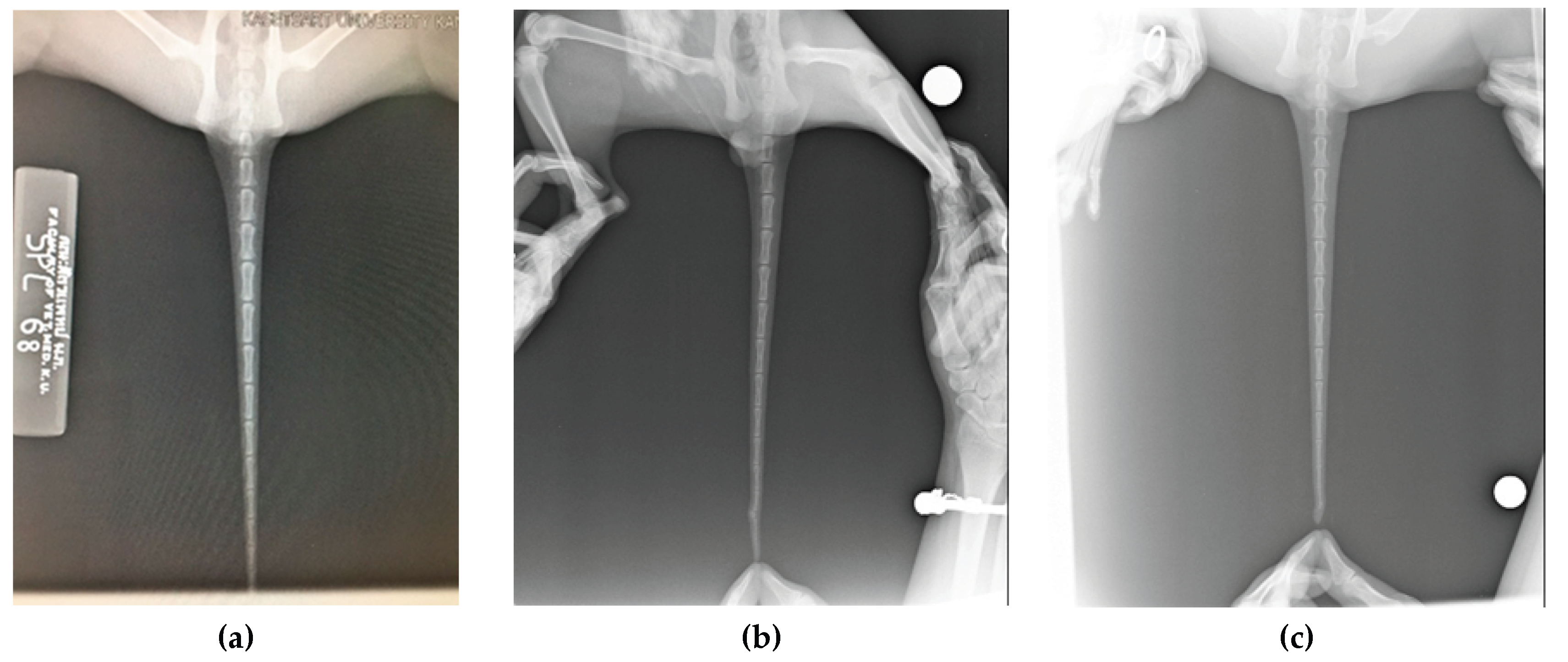

Table 1.
Genes, primer sequences, and sequence variants used for coat color gene testing in the Suphalak cat.
Table 1.
Genes, primer sequences, and sequence variants used for coat color gene testing in the Suphalak cat.
| Gene |
5’-3’ Forward and reverse primers |
Product size (bp) | Sequence variants | Identify color |
| ASIP | F- CTTTCTGGTCCTCTGGCTCCAC R- GGTGGTATGCAGAGCTTTTCCAA |
500 | 41G>C | Wild type (A; agouti) Non-agouti (a; solid) ALC agouti (APbe), |
| 123-124 2bp del | Non-agouti (a; solid) | |||
| 142T>C | ALC agouti (APbe) | |||
| TYRP1 | F- AACCAGAGATCTGTTCTTCACTCTT R- CTATGAGAACTCTCTGGTCACAAGC |
428 | 8C>G 298C>T |
Wild type (B; black) Chocolate (b) Cinnamon (bl) |
| TYR | F- TTTATAATCGGACCTGCCAGTG R- TCTGATATTGTATGTCCAGGATGTCTT |
598 | 679G>T | Wild type (C; full color) Burmese (cb) |
| F- CGAAGCCACAAACTGCGAGA R- CCTGAGGCTGCCAACCATCT |
700 | 901G>A 975delC 6delinsAATCTC |
Siamese (cs) Albino (c) Mocha (cm) |
|
| MLPH | F- GTGTGATCCTGACAGGCAGAG R- GGCTCGGCCTCATACACTC |
152 | 83delT | Wild type (D; full color) Dilute (d) |
| MC1R | F- TGCTGGGCTCCCTCAACTC R- GTACCGCAGCGCGTAGAAGA |
500 | 250G>A 15del (Jaguar-Mel) 24del (Jaguarundi-Dark) 439-441del |
Wild type (E; extension) Amber (e) Melanistic jaguar (e*) Dark-brown jaguarundi (e**) Russet (er) |
Table 2.
Sequence variants of coat color genes (ASIP, TYRP1, TYR, MLPH, and MC1R) observed in cats from the Thai Cat Conservation Center (n = 61) and Thai cat breeders (n = 66).
Table 2.
Sequence variants of coat color genes (ASIP, TYRP1, TYR, MLPH, and MC1R) observed in cats from the Thai Cat Conservation Center (n = 61) and Thai cat breeders (n = 66).
| Gene | Sequence variants | Thai cat conservation center | Thai cat breeders | |||||
| ASIP | c.41G>C | Agouti | Agouti | ALC agouti | Agouti | Agouti | ALC agouti | |
| G/G | G/C | C/C | G/G | G/C | C/C | |||
| 61 | 0 | 0 | 66 | 0 | 0 | |||
| Agouti | Agouti | Solid | Agouti | Agouti | Solid | |||
| c.123-124 2bp del | CA/CA | CA/- | -/- | CA/CA | CA/- | -/- | ||
| 0 | 0 | 61 | 0 | 0 | 66 | |||
| Agouti | Agouti | ALC agouti | Agouti | Agouti | ALC agouti | |||
| c.142T>C | T/T | T/C | C/C | T/T | T/C | C/C | ||
| 61 | 0 | 0 | 66 | 0 | 0 | |||
| TYRP1 | c.8C>G | Black | Black | Chocolate | Black | Black | Chocolate | |
| C/C | C/G | G/G | C/C | C/G | G/G | |||
| 31 | 22 | 8 | 2 | 2 | 62 | |||
| c.298C>T | Black | Black | Cinnamon | Black | Black | Cinnamon | ||
| C/C | C/T | T/T | C/C | C/T | T/T | |||
| 61 | 0 | 0 | 66 | 0 | 0 | |||
| TYR | c.679G>T | Full color | Full color | Burmese | Full color | Full color | Burmese | |
| G/G | G/T | T/T | G/G | G/T | T/T | |||
| 0 | 3 | 58 | 28 | 34 | 4 | |||
| c.901G>A | Full color | Full color | Siamese | Full color | Full color | Siamese | ||
| G/G | G/A | A/A | G/G | G/A | A/A | |||
| 58 | 3 | 0 | 54 | 9 | 3 | |||
| c.975delC | Full color | Full color | Albino | Full color | Full color | Albino | ||
| 61 | 0 | 0 | 66 | 0 | 0 | |||
| 6delinsAATCTC | Full color | Full color | Mocha | Full color | Full color | Mocha | ||
| 61 | 0 | 0 | 66 | 0 | 0 | |||
| MLPH | c.83delT | Dense color | Dense color | Dilute | Dense color | Dense color | Dilute | |
| T/T | T/- | -/- | T/T | T/- | -/- | |||
| 47 | 14 | 0 | 54 | 11 | 1 | |||
| MC1R | c.250G>A | Extension | Extension | Amber | Extension | Extension | Amber | |
| G/G | G/A | A/A | G/G | G/A | A/A | |||
| 61 | 0 | 0 | 66 | 0 | 0 | |||
| 15del | Extension | Extension | Jaguar-Mel | Extension | Extension | Jaguar-Mel | ||
| (Jaguar-Mel) | WT/WT | WT/15del | 15del/15del | WT/WT | WT/15del | 15del/15del | ||
| 61 | 0 | 0 | 66 | 0 | 0 | |||
| 24del | Extension | Extension | Jaguarundi-Dark | Extension | Extension | Jaguarundi-Dark | ||
| (Jaguarundi-Dark) | WT/WT | WT/24del | 24del/24del | WT/WT | WT/24del | 24del/24del | ||
| 61 | 0 | 0 | 66 | 0 | 0 | |||
| c.439-441del | Extension | Extension | Russet | Extension | Extension | Russet | ||
| WT/WT | WT/- | -/- | WT/WT | WT/- | -/- | |||
| 61 | 0 | 0 | 66 | 0 | 0 | |||
Note: MC1R-15del (Jaguar-Mel) = melanistic jaguar allele; MC1R-24del (Jaguarundi-Dark) = dark-brown jaguarundi allele.
Table 3.
Allele frequencies of ASIP, TYRP1, TYR, MLPH, and MC1R genes in cats from the Thai Cat Conservation Center (n = 61) and Thai cat breeders (n = 66).
Table 3.
Allele frequencies of ASIP, TYRP1, TYR, MLPH, and MC1R genes in cats from the Thai Cat Conservation Center (n = 61) and Thai cat breeders (n = 66).
| Gene | Allele | Predicted Phenotype |
Thai Cat Conservation Center (n = 61) |
Thai Cat breeders (n = 66) |
Total (n = 127) |
| ASIP | A | Agouti | 0 (0.00) | 0 (0.00) | 0 (0.00) |
| a | Solid | 122 (100) | 132 (100) | 254 (100) | |
| APbe | AAPbe(Agouti), APbea(Charcoal) | 0 (0.00) | 0 (0.00) | 0 (0.00) | |
| TYRP1 | B | Black | 84 (68.85) | 3 (2.27) | 87 (34.25) |
| b | Chocolate | 38 (31.15) | 129 (97.73) | 167 (65.75) | |
| bl | Cinnamon | 0 (0.00) | 0 (0.00) | 0 (0.00) | |
| TYR | C | Full color | 0 (0.00) | 75 (56.82) | 75 (29.53) |
| cb | Burmese colorpoint | 119 (97.54) | 42 (31.82) | 161 (63.39) | |
| cs | Siamese colorpoint | 3 (2.46) | 15 (11.36) | 18 (7.09) | |
| c | Albino | 0 (0.00) | 0 (0.00) | 0 (0.00) | |
| cm | Mocha | 0 (0.00) | 0 (0.00) | 0 (0.00) | |
| MLPH | D | Dense pigment | 108 (88.52) | 119 (90.15) | 227 (89.37) |
| d | Dilute | 14 (11.48) | 13 (9.85) | 27 (10.63) | |
| MC1R | E | Extension | 122 (100) | 132 (100) | 254 (100) |
| e | Amber | 0 (0.00) | 0 (0.00) | 0 (0.00) | |
| e* | Melanistic jaguar | 0 (0.00) | 0 (0.00) | 0 (0.00) | |
| e** | Dark-brown jaguarundi | 0 (0.00) | 0 (0.00) | 0 (0.00) | |
| er | Russet | 0 (0.00) | 0 (0.00) | 0 (0.00) |
Note: Percentages are shown in parentheses.
Table 4.
Genotype distribution at TYRP1, TYR, and MLPH loci in cats from the Thai Cat Conservation Center (n = 61) and Thai cat breeders (n = 66).
Table 4.
Genotype distribution at TYRP1, TYR, and MLPH loci in cats from the Thai Cat Conservation Center (n = 61) and Thai cat breeders (n = 66).
| Gene | Genotype | Predicted Phenotype |
Thai Cat Conservation Center (n = 61) |
Thai Cat breeders (n = 66) |
Total (n = 127) |
| ASIP | AA | Agouti | 0 (0.00) | 0 (0.00) | 0 (0.00) |
| Aa | Agouti | 0 (0.00) | 0 (0.00) | 0 (0.00) | |
| AAPbe | Agouti | 0 (0.00) | 0 (0.00) | 0 (0.00) | |
| APbea | Charcoal | 0 (0.00) | 0 (0.00) | 0 (0.00) | |
| aa | Solid | 61 (100) | 66 (100) | 127 (100) | |
| TYRP1 | BB | black | 31 (50.82) | 1 (1.52) | 32 (25.20) |
| Bb | black (carrier of chocolate) | 22 (36.07) | 1 (1.52) | 23 (18.11) | |
| bb | chocolate | 8 (13.11) | 64 (96.97) | 72 (56.69) | |
| TYR | CC | full color | 0 (0.00) | 20 (30.30) | 20 (15.75) |
| Ccb | full color (carrier of Burmese point | 0 (0.00) | 30 (45.45) | 30 (23.62) | |
| Ccs | full color (carrier of Siamese point | 0 (0.00) | 5 (7.58) | 5 (3.94) | |
| cbcb | Burmese (colorpoint) | 58 (95.08) | 6 (9.10) | 64 (50.39) | |
| cbcs | Tonkinese (colorpoint) | 3 (4.92) | 2 (3.03) | 5 (3.94) | |
| cscs | Siamese (colorpoint) | 0 (0.00) | 3 (4.54) | 3 (2.36) | |
| MLPH | DD | dense | 47 (77.05) | 54 (81.82) | 101 (79.53) |
| Dd | dense (carrier of dilute) | 14 (22.95) | 11 (16.67) | 25 (19.69) | |
| dd | dilute | 0 (0.00) | 1 (1.52) | 1 (0.79) | |
| MC1R | EE | extension | 61 (100) | 66 (100) | 127 (100) |
| E_ | extension | 0 (0.00) | 0 (0.00) | 0 (0.00) | |
| e* e* | Melanistic jaguar | 0 (0.00) | 0 (0.00) | 0 (0.00) | |
| e** e** | Dark-brown jaguarundi | 0 (0.00) | 0 (0.00) | 0 (0.00) | |
| er er | russet | 0 (0.00) | 0 (0.00) | 0 (0.00) |
Note: Percentages are shown in parentheses.
Table 5.
Genotypes of the TYRP1, TYR, and MLPH genes in 61 cats from the Thai Cat Conservation Centre and 66 cats from Thai cat breeders.
Table 5.
Genotypes of the TYRP1, TYR, and MLPH genes in 61 cats from the Thai Cat Conservation Centre and 66 cats from Thai cat breeders.
| Gene | Genotype | Predicted coat color phenotype |
Thai Cat Conservation Center (n = 61) |
Thai Cat breeders (n = 66) |
Total (n = 127) |
|
Solid black (B_C_D_) |
BBCCDD | Solid black | 0 (0%) |
0 (0%) |
0(0%) |
| Sable Burmese | BBcbcbDD | Sable Burmese | 26 | 26 | |
| (B_cbcbD_) | BBcbcbDd | Sable Burmese (carrier of dilute) | 5 | 1 | 6 |
| BbcbcbDD | Sable Burmese (carrier of chocolate) | 14 | 14 | ||
| BbcbcbDd | Sable Burmese (carrier of chocolate and dilute) | 6 | 1 | 7 | |
| Total | 53 | ||||
| Chocolate Burmese | bbcbcbDD | Chocolate Burmese | 6 | 2 | 8 |
| (bbcbcbD_) | bbcbcbDd | Chocolate Burmese (carrier of dilute) | 1 | - | 1 |
| Total | 9 | ||||
| Seal point Tonkinese | BbcbcsDd | Tonkinese point (carrier of chocolate and dilute) | 2 | - | 2 |
| (B_cbcsD_) | - | - | - | ||
| Total | 2 | ||||
| Chocolate point Tonkinese | bbcbcsDD | chocolate mink | 1 | 1 | 2 |
| (bbcbcsD_) | bbcbcsDd | chocolate mink (carrier of dilute) | - | 3 | 3 |
| Total | 5 | ||||
| Chocolate point | bbcscsDD | Siamese chocolate point | - | 1 | 1 |
| Siamese | bbcscsDd | Siamese chocolate point (carrier of dilute) | - | 1 | 1 |
| (bbcscsD_) | bbcscsdd | Siamese liac point | - | 1 | 1 |
| Total | 3 | ||||
| Solid chocolate | bbCCDD | Solid chocolate | - | 18 | 18 |
| Suphalak | bbCCDd | Solid chocolate (carrier of dilute) | - | 2 | 2 |
| (bbC_D_) | bbCcbDD | Solid chocolate (carrier of Burmese point) | - | 29 | 29 |
| bbCcbDd | Solid chocolate (carrier of Burmese point and dilute) | - | 1 | 1 | |
| bbCcsDD | Solid chocolate (carrier of Siamese point) | - | 3 | 3 | |
| bbCcsDd | Solid chocolate (carrier of Siamese point and dilute) | - | 2 | 2 | |
| Total | 55 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



