
Submitted:
11 December 2024
Posted:
12 December 2024
You are already at the latest version
Abstract
This study aimed to investigate the genetic polymorphisms in Agouti signaling protein (ASIP) and melanocortin 1 receptor (MC1R) genes and their association with coat color in native Bulgarian sheep breeds. A total of 247 individuals belonging to 7 native sheep breeds from 38 herds were genotyped for the ASIP and MC1R genes. Five single nucleotide polymorphisms (SNPs) were de-tected in the MC1R gene and three haplotypes representing MC1R ED and E+ alleles were identified among the studied sheep breeds. It was found that the dominant ED allele of MC1R is absent in the breeds - Patch-faced Maritza and Dubenska. The most common haplotype (h3) of the domi-nant ED allele was associated with either black or brown coat color in native Karnobatska, Kara-kachanska and Cupper-red Shumenska breeds, respectively. We can infer that haplotype 3 alleles could be a possible result that could interpret the mechanisms of black wool colour manifestation in the native Karnobatska sheep breed, which has formed the genetic pool of this sheep breed.
Keywords:
polymorphism
; ASIP
; MC1R
; genes
; native sheep breeds
1. Introduction
The coat color (hair and fibrous cover) is an essential trait in sheep for breed identification, characterization, and morphological selection. Fiber color has critical practical applications in wool production from sheep, goats, and other farm animals [1]. The production of colored wool is gradually gaining new importance considering environmental protection, green economy, and new fashion trends in the textile industry. Conservation of color varieties within native sheep breeds is a vital breeding task in maintaining genetic diversity within the breeds.
The diversity of coat color covering the body in sheep is due to the presence, distribution, and biochemical activity in the melanocytes in which two types of melanins (eumelanin and pheomelanin) are synthesized [2]. The eumelanin in sheep is either black or brown and phaeomelanin results in a red, tan, or fawn colour [3] or non-pigmented phenotype. Many genes affect coat color in sheep, but two loci have a significant role in determining coat color - Agouti and Extension [4]. These two loci regulate the relative amounts of eumelanins and pheomelanins in the skin and fibers. In addition to Extension and Agouti loci, [5], in the literature have been reported other genes responsible for coat color in the sheep genome such as the tyrosinase-related protein 1 (TYRP1), the v-kit Hardy–Zuckerman 4 feline sarcoma viral onco-gene homolog (KIT), and the microphthalmia-associated transcription factor (MITF) and melanocyte-inducing transcription factor [5,6]. Other genes associated with the regulation of coat color development and provide a theoretical basis for selecting natural coat colors of sheep also are reported as DCT, TYR, PMEL, SLC45A2, and MLANA [7].
The Agouti locus contains a gene that codes for producing Agouti signaling protein (ASIP) [8]. It is a small paracrine signaling molecule that interacts with the products of the Extension locus. The Extension locus contains a gene that encodes the melanocortin 1 receptor (MC1R), which is present on the surface of the melanocyte membrane. The binding of MC1R to α melanocyte-stimulating hormone (αMSH) [9] induces eumelanin synthesis, while alternative binding to ASIP induces a switch in pigment synthesis from eumelanins to pheomelanins [7, 8]. The MC1R gene in sheep is located on ovine chromosome 14 (OAR14) [12]. The ASIP gene was mapped on a region of chromosome 13 [8, 10].
In sheep, classical genetic studies have identified two alleles at the Extension locus – dominant black (ED) and recessive (E+) [11, 12]. At the beginning of the 80s of the last centuries, molecular genetics was rapidly developed. In studying molecular mechanisms by which genes are expressed, new methods have begun to be applied that better reveal genetic diversity at the molecular level. Polymerase chain reaction-restriction fragment length polymorphism (RFLP-PCR) is a technique used to identify single nucleotide polymorphisms (SNPs) based on the recognition of restriction sites by restriction enzymes [16]. Using the PCR-RFLP test, two missense mutations in the MC1R gene (p.M73K and p.D121N) that determine the dominant black colour (ED allele) have been identified [17]. Initially, these mutations in the MC1R gene were found in the populations of the Norwegian Dala sheep breed [6, 11], and the Spanish rare Xalda sheep breed [18]. The second allele in the Extension locus is a wild allele (E+), widely distributed in most breeds [4]. Another MC1R missense mutation (p.R67C) was identified [1] in the Italian sheep breed Valle del Belice and the authors suggested that this is the cause of the recessive e allele at the ovine Extension series. However, it has not yet been well-documented and universally recognized. Examining MC1R mutations many authors did not find the putative recessive allele e (c.199 C>T), which was previously reported [14, 16, 17]. The first attempt to assess the genetic polymorphism of the MC1R gene in the population of 11 Greek sheep breeds also reported that the putative recessive allele e (c.199 C>T) in the Extension locus was not found, and one non-synonymous mutation that had never been reported (c.789 T> C, p.263 Leu > Pro) [21].
To characterize the genomic structure of the ASIP gene, sequence analysis of genomic DNA, BAC clones, and RT-PCR products from Romanov, Texel, Merino, and ancient Barbary sheep breeds was used [22]. The authors identified a 190-kb tandem duplication encompassing the ovine ASIP and AHCY coding regions and the ITCH promoter region as the genetic cause of the white coat color of the dominant agouti allele (Awt). Ubiquitous expression of a second copy of the ASIP coding sequence regulated by a duplicated copy of the nearby ITCH promoter causes the white sheep phenotype. The D5 deletion and the g.5172T>A SNP in exon 4 would be predicted to independently cause functional changes to the agouti protein [22]. Despite an in-depth sequence analysis by the Australian authors, they could not conclude that ASIP coding regions 5-bp deletion and a g.5172T>A SNP are the only genetic causes for the expression of recessive black color in Merino sheep.
The relationship between different alleles of ASIP and recessive black coat color in the rare Xalda breed was reported [18]. The determination in the absence of a dominant black allele (ED) cannot reject that the homozygous (Aa/Aa) genotype could cause a recessive, black-coated phenotype in sheep [18]. However, this genotype cannot be the sole cause of the recessive black colour in the Xalda breed because most black-coated individuals (109 out of 120) were not homozygous for the 5-bp deletion. Thus, they reject the Adel/Adel genotype as the sole cause of recessive black coat colour in sheep. ASIP and MC1R gene polymorphisms in two Pramenka sheep populations originated in Bosnia and Herzegovina (Dubian and Privorian sheep) were analyzed [27], in which population coat colors and color patterns are not fixed traits. Ninety-five percent of the coat color variation in Brazilian Crioula sheep was explained by epistatic interactions between specific alleles in the MC1R and ASIP genes [23]. The Brazilian author's evidence suggests an essential role of TYRP1 variants for wool color despite their low frequencies. The marker panel was efficient enough in predicting coat color in the studied animals and, therefore, can be used to implement a marker-assisted selection program in the conservation nucleus of sheep of the Crioula breed.
Mutations in ASIP and MC1R genes and their association with black coat colour have been studied in seven native Swedish sheep breeds in individuals with black, white or grey fleece [24]. Previously known mutations in ASIP recessive black allele (g.100_105del and/or g.5172T>A were associated with black coat colour in Kleovsjeo and Roslag sheep breeds and mutations in MC1R (dominant black allele): c.218T>A and/or c.361G>A) were associated with black coat colour in Swedish Finewool. Swedish sheep breeds have grey individuals in their populations, and grey is believed to result from mutations and allelic copy number variation within the ASIP duplication. Finally, a novel missense mutation in MC1R (c.452G>A) is identified in Gotland, Gute, and Vearmland sheep, and evidence of a duplication of MC1R in Gotland sheep.
The four missense mutations included two previously known mutations, c.218T>A and c.361G>A, and the two missense mutations, have been found in Norwegian Dala, Damara, Corriedale, and Spanish Merino [3, 4]. In addition, c.361G>A, c.429C>T, c.600T>G, and c.735C>T are segregated in Iranian and Moroccan sheep populations genotyped in the NEXTGEN project (http://ne xtgen.epfl.ch/;mutation information accessed from Ensembl http://www.ensembl.org/index.html).
Phenotypic description and characterization of 18 Bulgarian native sheep breeds with colored wool have been made in an extensive review[25]. Depending on the color of the fleece, the native sheep breeds in Bulgaria have been divided into three groups: 1) breeds with the fully pigmented fleece; 2) breeds with both fully pigmented and fully white fleece, and 3 breeds with spotted-colored fleece [25]. Some hypotheses about genetic loci and alleles affecting coat color in the populations of Bulgarian native sheep breeds, following the nomenclature of COGNOSAG [26] have been made [25]. No report has been made regarding the polymorphism of MC1R and ASIP genes and the potential association of its mutations with coat colors in Bulgarian native sheep breeds till now.
This study aimed to investigate the genetic polymorphisms in Agouti signaling protein (ASIP) and melanocortin 1 receptor (MC1R) genes and their association with coat color in native Bulgarian sheep breeds.
2. Materials and Methods
2.1. Animal Sampling
Blood samples were taken from 38 herds in Bulgaria, representing 7 native sheep breeds out of a total of 247 individuals (76 animals from Patch-faced Maritza, 26 animals from White Maritza, 38 animals from Copper-red Shumenska, 22 animals from Dubenska, 31 animals from Karakachanska, 21 animals from native Starozagorska, 33 animals from native Karnobatska (Figure 1, S1). For the White Maritza and Patch-faced Maritza sheep breeds, samples were taken from unrelated animals verified through completed pedigree. For the remaining breeds, samples were taken randomly from 3 to 5 samples per herd, and the principle of unrelatedness of the sampled animals was respected by surveying the sheep breeders.
2.2. DNA Extraction and PCR Conditions
DNA was isolated from 200 µl of baseline blood volume using the ThermoScientific™ GeneJET Genomic DNA Purification Kit, following the manufacturer's recommendations. Isolated DNA was quality checked on a 1% agarose gel stained with Green Safe DNA Gel Stain (Canvax) and stored at -20 oC. The quality and quantity of isolated genomic DNA were checked using an Epoch microplate spectrophotometer (Agilent Technologies, Santa Clara, CA, USA).
PCR-RFLP was used to genotype the animals for two single nucleotide polymorphisms (SNPs): missense mutations already identified in the ovine MC1R gene that determines the black dominant ED allele (c.361G>A or p.D121N) at the Extension locus [17], one was an SNP (c.-31G>A) located in the 5′-untranslated region (5′-UTR) of the MC1R gene [27]. Details of the PCR-RFLP protocols used for the analysis of the ASIP and MC1R polymorphisms have been already reported [28] (Table 1). PCR analyses are performed in a 20 ml volume containing 1 µL My Taq HS red Mix (Bioline). The PCR profile was as follows: 5 min at 95 0C; 35 amplification cycles from 0.3 min at 95 oC, 0.3 min at 58 oC to 59 oC, 0.3 min at 72 oC, and final extension - 5 min at 72 oC (Table 1).
Before the restriction procedure, pre-purification of the PCR products was carried out. In this step, the distinct DNA fragments were excised from 1% agarose gel and then purified, applying ISOLATE II PCR and Gel kit (BIOLINE) as described by the supplier. Restriction reaction is performed with 15 µL of the purified PCR product in a total of 20 µl reaction volume, including 3U restriction enzyme and 1X reaction buffer, 0.2 µL. BSA. The results were analyzed on a 3% agarose gel after electrophoresis.
2.3. Polymorphism in the ASIP Gene
Fragment analysis has also been used to determine whether sampled animals have a duplicate copy of DNA, including ASIP (Norris & Whan, 2008). Primers Agt16 and Agt18 spanning the duplication site yielded a unique PCR product of 242 bp, while Agt16 and Agt17 spanning the 5'-point break sequence yielded a fragment of 238 bp. Amplification was performed by asymmetric competitive PCR. If amplification results in these two fragments, the animal carries at least one copy of the duplicated chromosomal region. In contrast, when only 238 bp is amplified, the animal carries no duplicated DNA region.
2.4. Analysis of Polymorphisms in the MC1R Gene
The polymorphism (c.-31 G>A) located in the 5′-UTR of is genotyped by PCR-RFLP [19]. PCR was performed using a TGradient DLAB TC-1000-G) in a 20 μl volume containing 1 μl of DNA template, 10 pmol of each primer and PCR profile, as follows: 5 min at 95 °C; 35 amplification cycles of 30 s at 95 °C, 30 s at 58 °C, 30 s at 72 °C; and final extension - 5 min at 72 °C. The reverse primer contains a mismatched nucleotide that introduces an additional artificial restriction site for the AluI endonuclease when the c.-31 G allele is amplified. The 202 bp product was subjected to restriction digestion with 3 U AluI, and the resulting DNA fragments were separated by electrophoresis on a 3% agarose gel. The genotype of each animal will be determined as follows GG=c. 31GG (products size of 83 and 96 bp); GA=c.-31GA (products size of 83 and 96 and 119 bp); AA=c.-31AA (products size of 83 and 119 bp).
2.5. MC1R Polymorphism Analysis in Individual Samples by PCR-RFLP
PCR-RFLP genotyped a polymorphism (c.-31G>A) located in the 5′-UTR. PCR was performed using a DLab thermocycler gradient apparatus in a volume of 20 μl containing 10-50 ng DNA template, 10 pmol of each primer (Table 1). The PCR profile is as follows: 5 min at 95 °C; 35 cycles of amplification of 30 s at 95 °C, 30 s at 58 °C, 30 s at 72 °C; final extension - 5 min at 72 °C. The reverse primer contains a mismatched nucleotide that introduces an additional artificial restriction site for the AluI endonuclease when the c.-31 G allele is amplified. The resulting PCR product of 202 bp was subjected to restriction with 3 U AluI (Fermentas), and the resulting DNA fragments were separated by electrophoresis on 3% agarose gel.
2.6. SNPs Identification and Genotyping /Sequencing of MC1R Gene
SNPs were identified by sequencing amplicons of the whole coding domain sequences (CDS, 954 bp) and parts of the 5’- and 3’-untranslated regions (35 and 125 bp, resp.) of MC1R in both directions. DNA from each breed of Patch-faced Maritza, Cooper Red-Shumenska, Dubenska, Karakachanska, native Starozagorska, and native Karnobatska were used for identification mutation sites. Primers amplifying the coding region of MC1R were used for sequencing (Table 1) [20]. All amplifications were performed on TGradient DLAB TC-1000-G. The reaction was performed in 25 μL containing 50 ng DNA template, 12 μL My Taq HS red Mix (Bioline), and 10 pM of MC1R specific primers (MF and MR). The PCR condition was as described by [29]. Macrogen Europe (The Netherlands) directly sequenced the PCR products in both directions. Sequences were analyzed using Geneious software (USA) to identify polymorphisms. Highly informative SNPs were identified for genotyping.
3. Results
3.1. Relationship Between ASIP Polymorphisms and Coat Colour in Sheep
3.1.1. ASIP Genotypes
Two hundred forty-seven sheep were analyzed for the presence of a duplicated allele of the ASIP gene. ASIP alleles were determined based on their size: 242 bp for the A+ allele and 237 bp for the Adel allele. For the Patch-faced Maritza breed, seventeen of 76 animals were found to have duplicated alleles at the Agouti locus (Table 2). Duplication was found in six patch-faced animals, and duplication and deletion were found to exist simultaneously in 11 animals. In 16 animals in this breed, duplication and deletion were not detected. In 43 animals, deletion was found (Table 2). The Patch-faced Maritza sheep breed population can differentiate into three color varieties: patch-faced, fully pigmented, and piebald. "Patch-faced" is a term used to refer to a phenotype in which sheep have white body color, but on the head, ears, and legs, the animals have typical black spots. The animals appear piebald when the black spots cover most of the body. Some animals have black spots on different parts of the body. Оn the black spots on a fleeced area, it grows pigmented wool. Fully pigmented sheep can be found very rarely in this breed. Usually, in the population of Dubenska breed normally can be found pigmented and white sheep. For this study the samples were taken mostly from pigmented sheep. No duplicate copy has been found among the pigmented sheep in the Dubenska breed. There are two samples from white phenotypes for this breed in which the duplication was not detected (Table 2).
For the Karakachanska breed, in which all animals were pigmented, eleven of the sheep carry a deletion in exon 2, or as usually denoted D5 (g.100-105delAGGAA). In the samples of the Karakachanska breed, where all animals were black, as expected, there is no duplication at the Agouti locus. Only one of the animals of the Copper-red Shumenska breed was found to have duplication. The deletion D5 (g.100-105 del AGGAA) was found in the seven breeds analyzed at various frequencies. Sixteen of the 22 animals here carry the deletion D5 in the population of the Dubenska breed. In the Karakachanska sheep breed population, deletion D5 was found in only eleven animals. Interestingly, in the native Karnobatska sheep population, duplication was found in 11 animals, although all animals in the samples group are pigmented, which is an indication that the color of the animals in this sample group is encoded by the dominant ED allele in the Extension locus.
3.2. MC1R Polymorphism Analysis in Individual Samples by PCR-RFLP
3.2.1. Genotyping of the Mutation (c.361G > A; p.D121N)
Fully pigmented animals and 5 sheep from Patch-faced Maritza breed were selected for restriction analysis outside those tested by MC1R gene sequencing. Analysis of the c.361G >A mutation was obtained by digesting the 366 bp amplified region of the MC1R gene with restriction enzyme Tru1I (Fermentas; recognition sequence, TTAA) at 65°C for 5 hours (Figure S2). The resulting PCR-RFLP pattern was distinguished by one fragment of 366 bp (allele c.361G) or two fragments of 265 + 101 bp (allele c.361A). In black animals, two replacement mutations - Met→Lys at position 73 (M73K) and Asp → Asn at position 121 (D121N) - were identified, showing a complete association with black coat color. Both mutations can be easily tested by PCR-RFLP or sequencing. Black animals were found to be predominantly heterozygous at these two positions (73, 121), whereas white animals were homozygous, supporting the assumption of dominant inheritance of black coat color.
PCR-RFLP with AluI restriction was performed on MC1R to identify c.-31G>A SNP in different sheep. The genotype of individual animals is indicated as follows: GG=c.-31GG; (83.96 and base pairs) GA=c.-31GA; (83,96 and 119 base pairs) AA=c.-31AA; (83 and 119 base pairs) (Figure S3). The dark pigmentation associated with genotype AA was confirmed in the analyzed animals of the Karakachanska breed, and genotype GG was absent, which was related to white coloration in sheep (Figure 2).
3.2.2. SNPs Identification and Genotyping
A sequence of about 1140 bp including the coding region and parts of the 5′- and 3′-un-translated regions (UTR) of the MC1R gene was successfully amplified. Alignment with the sheep reference (GenBank: ANH58923.1,58924-25-26-27-28-29-30, sequences showed 98% and 99% sequence homology, respectively (Figure 3). By analyzing and comparing the electropherogram of the sequences obtained from the DNA pools of 72 sheep, the results showed that five single nucleotide polymorphisms (SNPs), two nonsynonymous mutations previously associated with black coat color (c.218 T>A, p.73 Met>Lys. c.361 G>A, p.121 Asp>Asn) and three synonymous mutations (c.429 C>T, p.143 Tyr>Tyr; c.600 T>G, p.200 Leu>Leu. c.735 C>T, p.245 Ile>Ile) were identified in the CDS of the MC1R gene (GenBank accession number: KF198511) [9, 19, 27].
3.2.3. Haplotype
All alleles of the mutation (c.218A, c.361A, c.429T, c.600G, and c.735T) were found in the native Karnobatska breed, with two genotypes at each mutation site. Two nonsynonymous mutations (c.218 T>A, p.73 Met>Lys. c.361 G>A, p.121 Asp>Asn) defining the dominant black (ED) allele [17], these mutations have not been identified in Pathc-faced Maritza (white and black spotted coloration in the face) and Dubenska sheep, which have breed-specific coat pigmentation. The three synonymous mutations were transmitted with high frequency. The frequency of two nonsynonymous mutations is very low or rare in the native Starozagorska and Karakachanska breeds. The three nonsynonymous mutations in the sheep breeds, except the native Karnobatska breeds, have three genotypes. Figure 4 reports individual haplotype frequencies among the investigated breeds. Three haplotypes (haplotype1 [TGCTC], haplotype2 [TGTGT], and haplotype3 [AATGT]) were identified.
The haplotype 3 of all individual mutations was observed as predicted for all animals in the native Karnobatska sheep breed. Eleven of sixteen Cooper-red Shumenska bred animals also displayed haplotype 3 and Karakachanska breеd, where most animals are pigmented. Only one animal with black-colored fur from native Starozagorska breed, which predominately represents white coloration is present here with all five individual mutations. The white coat color breed (native Starozagorska) was only found in haplotype1 and haplotype 2. Interestingly, we also observed that the haplotype 1 and 2 frequency was significant for and very low in the Karakachanska breed.
The analysis of the results showed that five mutations were entirely associated with black (reddish-brown) coat colour in the native Karnobatska sheep population. At the same time, three haplotypes (haplotype1, haplotype2, and haplotype3) were determined from the SNP mutations in the MC1R gene. Interestingly, haplotype 3 is almost fixed in this breed (there are two missense mutations in haplotype 3 causing the ED allele). Haplotype 3 has not been detected in white animals (Figure 4). We can infer that haplotype3 alleles could be a possible result that could interpret the mechanisms of black wool colour manifestation in the Native Karnobatska sheep breed, which has formed the genetic pool of this sheep breed.
In the analysis of all pigmented sheep referring to haplotype 1, associated with light but pigmented coloration, a high level of heterozygosity was also found for the mutations tested (Figure 5).
Haplotype 3 was not found in sheep of the “Dubenska” breed. Connecting this with the results of the ASIP gene study, where there is no duplication, and in 16 out of 22 sheep, there is a deletion in exon 2, we can explain the variations in coloration and their assignment to haplotype1. As expected, the analyzed sheep are from the breed "Native Starozagorska sheep," which population is characterized by a predominantly white phenotype, and here in the pigmented representatives of the breed were found three synonymous mutations in the MC1R gene (c.429C>T, pY143Y; c.600T>G, pL200L; c.735C>T, I245I). The alleles of haplotype 3 have not been associated with patch-faced phenotype (white body color, but on the head, ears, and legs, the animals have typical black spots) in the Patch-faced Maritza breed.
4. Discussion
At the end of the 20th and the beginning of the 21st century, the Ministry of Agriculture of the Republic of Bulgaria recognized and legalized the activity of breeding organizations for native sheep breeds. Recognition of the breeding organizations nowadays usually has to be linked to the approval of breeding programs for each breed, which necessarily include a description of the breed standard containing a description of the morphological features of the breed, including the coat color [Regulation (EU) 2016/1012]
Primarily, the breed standards of the native Bulgarian breeds, such as the native Karnobatska and the Copper-red Shumenska, were aimed at pigmented sheep. For these native breeds, the standard came into conflict with biological reality and the mindset of sheep breeders. For many native breeds, the standard was influenced by the name. For example, the color should be copper-red for the Copper-red Shumenska breed. For the native Karnobatska breed, the standard defined pigmented sheep with a reddish-brown color as typical of the breed, although in the past, many authors have reported that individuals with white-coloured hair and fiber cover also occur in populations of these breeds, although rarely [25]. In the White Maritza sheep, the standard defines white as typical for the breed, although 5-10% of pigmented and less frequently piebald sheep are found in the population.
In the sheep breeding community of the country, opinions and views have been formed that the standard defines much more of what sheep should be than they are. As a result of this perception, a white lamb born into a pigmented flock or a black lamb born into a white flock was an undesirable phenotype, and often, they were excluded for breeding purposes.
In this case, the flawed thinking is that the standard reflects the reality of the breed, and, therefore, the genetic mechanism for colored lambs cannot be present in a white breed. Related to this is the wrong thinking and understanding that all sheep breeds are ancient and are ancient, isolated gene pools. In most of these breeds, none of these assumptions are true. The genetic mechanism for colored lambs persists in many breeds with white-colored coats [31].
Genetic mechanisms for white phenotypes exist to one degree or another in many sheep populations (White Maritza, native Starazagorska, Dubenska). Our study in the Copper-red Shumenska sheep breed (Figure 1i) confirmed this, where 38 fully pigmented animals were sampled (Table 2). However, in one of the blood samples, the duplication was detected, which may encode the white phenotype in the homozygous state if the Extension locus wild allele E+ is in the homozygous state. The deletion in exon 2 (g.100-105delAGGAA), denoted as D5 was present in 37 animals in the samplesed group in this breed. However, the fact that MC1R sequencing identified haplotype 3 (ED) in 11 of the animals means that pigmentation in the Copper Red Shumen breed may be encoded by both the deletion D5 if the homozygous state and if the genotype at the Extension locus is E+E+ or dominant ED allele present.
The black spots with varying sizes on the face and some body areas are crucial in selecting male and female lambs for breeding purposes in sheep breeds such as Path-faced Maritza, Elin-Pelinska [23]. Wool pigmentation is essential in selecting male and female lambs in native Bulgarian sheep breeds such as Native Karnobatska, Couper-red Shumenska, Karakachanska, and Dubenska. Thus, it would be essential to identify the genetic causes and genetic variants associated with coat colour traits in these breeds. In sheep, at least three mutations were suggested to cause the nonagouti recessive black coat colour: a 5 bp deletion in exon 2 (g.100-105delAGGAA, denoted D5), a missense mutation in exon 4 (g.5172T.A or p.C126S) and a regulatory mutation, a deletion of 5 bp in exon 2, was identified as the cause of the black recessive nonagouti (Aa) allele only indirectly deduced but not yet characterized [15, 18, 26]. Thus, it would be essential to identify the genetic mechanisms and genetic variants associated with coat colour traits in these breeds. The phenotype of an ewe that is AaAa / E+E+ and a sheep that carries the dominant allele ED in Extension locus a homozygous or heterozygous state are identical - there is no visible visual difference. In addition, a not yet characterized regulatory mutation, a deletion of 5 bp in exon 2, was identified as the cause of the black recessive nonagouti (Aa) allele [18].
For this reason, careful breeding and lambing records must be kept if the shepherd wants to trace these genetics and to preserve genetic diversity. Additionally, some studies in the past have established advantages of the recessive black color, including increased fertility, better maternal skills, lower levels of internal parasites, and fewer hoof problems [33]. These multiple survival traits are associated with codominant and recessive alleles at the Agouti locus. They are more prevalent in recessively colored ewes than those colored due to a dominant allele at the Extension locus. To investigate genes that may play essential roles in sheep skin, particularly in fiber and coat pigmentation, over 100 million transcriptome sequences were generated from the skin of white and black sheep using Illumina technology [32].
We identified the duplication and the deletion at the Agouti locus in the Native Karnobatska breed. However, the dominant allele at the Extension locus is why all animals in the sample are pigmented. However, individuals with AaAa, EDED, or ED/E+ phenotypically are not visually distinguishable.
In the samples taken for the Karakachanska sheep breed, 31 animals were fully pigmented. The MC1R sequence results show that haplotype 3 (the dominant ED allele) was identified in 3 animals (Figure 4), and it is undoubtedly the determinant of eumelanin synthesis. In the remaining 11 pigmented sheep, the MC1R sequence showed the presence of haplotype 1 and haplotype 2, which include synonymous mutations (c.429 C>T, p.143 Tyr>Tyr; c.600 T>G, p.200 Leu>Leu. c.735 C>T, p.245 Ile>Ile) and accordance of studies of [29], but PCR performed on these identified the D9D5 deletion, which we believe is the cause of eumelanin synthesis. The analysis of the polymorphisms of the MC1R and ASIP genes in the Karakachanska breed indicates that the preferred black color in the breeding of sheep may be due to both the ED allele and the Aa allele in an individual. Selection over the years for individuals with the black coat considered typical of the breed has most likely increased the frequency of the ED and Aa alleles and reduced the frequency of the duplication in the ASIP gene, making the birth of white coat lambs very rare.
ASIP alleles were determined based on their size: 242 bp for the A+ allele and 237 bp for the Adel allele [32]. To investigate genes that may play essential roles in sheep skin, particularly in fiber and coat pigmentation, over 100 million transcriptome sequences were generated from the skin of white and black sheep using Illumina technology [30].
Identifying genomic regions, genes associated with phenotypic traits, and description of gene function are some of the applied research activities to understand the coat color genetics of livestock species [35].
The reddish-brown colour of the native Stara Zagora sheep is due to the recessive allele Aa [25]. The PCR performed on the pigmented phenotypes of the native Starozagorska sheep identified the deletion at the ASIP locus as the mutation causing the pigmented phenotype. MC1R sequencing was performed on 10 samples of pigmented native Starozagorska sheep. Haplotype 1 and haplotype 2, neutral in coat color, were detected in nine sheep, and haplotype 3 was detected in only one sample. Usually, the typical color for the native Stara Zagora sheep breed is white, which, according to classical genetics, is encoded by the dominant allele of the Agouti locus (Wt). Our studies show that the coloration of the pigmented sheep in this local population is most likely due to the recessive allele of the Agouti locus (deletion). In this breed, the study should be expanded to a more significant number of sheep since pigmented and piebald sheep are rarely found in the population.
MC1R is epistatic to Agouti alleles. White patches and white spots in mammals are due to abnormal melanoblast development or survival, reflecting the lack of mature melanocytes. White spots can be caused by defects at various stages of melanocyte development, including proliferation, survival, migration, envelope invasion, hair follicle entry, and melanocyte stem cell renewal [36]. It was reported that the presence of polymorphism among different sheep breeds and its effect on the coat color scoring confirm the 5'-UTR region of the MC1R gene as a functional candidate locus [37]. Based on PCR-RFLP analysis, these results correspond to our reporting of the nucleotide substitution of -206G>A.
Sequencing of the MC1R in the Dubenska breed (Figure 4) showed that the dominant ED allele was missing in the pigmented sheep of this breed tested and that the Extension locus contained mainly synonymous mutations encoding haplotype 1 (TGCTC) and haplotype 2 (TGTGT), which are amino acid-unchanged and colour-neutral. The pigmented phenotypes in the Duben sheep are mainly due to the deletion at the ASIP locus. Two of the animals analyzed are white and predictably belong to haplotype 1. In one of the white ewes, the mutations in the ASIP locus were not detected, whereas in the other, the deletion D5 was present. The presence of at least one non-mutated variant of, a deletion of D5, and a deletion of D9 (i.e., N5 and N9, N5 and N9 being the names of the non-mutated alleles of D5 and D9, respectively) is required for the white phenotype [22]. One possible explanation for the white phenotype in Dubenska breed is the presence of N9N5 and N9D5 genotypes.
As eumelanin results from recessive allele (Aa) in the blackish-brown phenotypic trait, the dominant AWt (white/ tan) allele in Saanen goats gives the impression of a whitish coat [38]. The absence of duplications in ASIP has previously been shown to be associated with sheep with darker fleece in Massese [1, 2], Dubian and Privorian. Modifications in one or more than two copies of the ASIP gene and the additional spontaneous mutations of D9 and/or D5 preceding the targeting modification could also involve forming coat color patterns in sheep [39]. An uncategorized mutation and removal of 5bp at exon 2 and non-sense alteration at exon 4 were considered as one of the major causes of the black recessive non-agouti (Aa) allele [40].
Genotyping the Iranian sheep populations via SSCP, led us to discover polymorphisms over the flanking region of DNA. The provided coat colour descriptions suggest that mostly the individuals with light phenotypes as white, light grey and light cream colours in the population show the heterozygous banding pattern [41].
The genotype results suggested that these mutations are not associated or not completely associated with the investigated sheep breeds. The above results indicated that the variation in the protein-coding region of ASIP did not explain the variation in the coat colour phenotypes of Bulgarian native sheep breeds.
Coat pigmentation is a crucial factor in the heat tolerance index. It determines the physiological characteristics in White Santa sheep as the individuals exhibited lower average rectal temperatures, heartbeat, and respiration rates compared to the control groups [42].
It was reported that differences between black and red coats result from the expression of different alleles of the same gene without directly affecting productive/reproductive characteristics [43].
Developing a black line targeting specific markets might also be worthwhile in maximizing production and maintaining qualities associated with black color. These are often called the BASE© of color because this abbreviation includes all four specific gene pairs: the B - black-brown locus, the A - agouti locus, the S - spotted locus, and the E - extension locus. There is no way to identify the agouti pattern alleles of a dominantly colored ewe with one copy of the ED except by carefully tracing offspring that do not carry the dominant color. Taken together, MC1R and ASIP genotypes show that the two genes interact to sinergically modulate the amount of melanin synthetised in alpaca fibre [44].
There is no way to readily identify the Agouti pattern alleles of a double dominant ewe, as all offspring will be pure black or brown, possibly with spots.
In tracing the genetics of sheep color, four specific loci determine much of what is seen as a coat colour when looking at sheep [45]. The selection against black coat color seems to hurt the genetic progress of growth traits in the Menz sheep [46]. Developing a black line targeting specific markets might also be worthwhile in maximizing production and maintaining qualities associated with black color.
The fortunes of many sheep breeds have changed in recent decades. Many native sheep breeds have remained as small population and have acquired the status of 'endangered'. On the other hand, breeders of native breeds adhere to the standard they adhered to during the breed's heyday. Adherence to the historical standard of the breed is not in itself a bad thing, but at some point, the management of the genetic population structure of these 'endangered' breeds must take precedence over any historical or economic role of the breed [47,48].
5. Conclusions
In this study, we found that the color varieties in the seven native Bulgarian sheep breeds are, in most cases, encoded by the alleles of the ASIP gene and by the recessive Aa allele. The dominant allele of the Extension locus is the leading genetic cause of the pigmentation of all individuals in the local Karnobat breed. ED can also be a genetic cause of an individual's pigmentation in the breeds Copper-red Shumenska and Karakachanska. However, in these breeds, the pigmentation can also be due to the recessive Aa allele of the ASIP gene. Within the studied samples, it was found that the dominant ED allele of MC1R is absent in the breeds - Patch-faced Maritza and Dubenska. In the native Starozagorska breed, our observations and genealogical analysis show that the pigmented variants are due to the recessive non-agouti allele (Aa), but in one of the samples, an ED allele of the Extension locus was found, and this requires additional studies of a more significant number of samples.
In our study, we found that the polymorphism of the ASIP gene cannot fully explain the "Patch-faced" phenotype in the Patch-faced Maritza sheep population, and perhaps the expression of this phenotype is influenced by other alleles in the ASIP gene and other genes as shown by the PCR-RFLP analysis of the ASIP gene. PCR-RFLP analysis in the White Maritza sheep breed showed that the white color can be expressed even without duplication, which also requires additional studies.
Our study provides novel insights into the genetic causes of different color varieties in populations of seven native Bulgarian sheep breeds. It expands our knowledge of the link between ASIP and MC1R genes and varying pigmentation patterns in sheep.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Figure S1: Sample collection, representing 7 native Bulgarian sheep breeds - map.; Figure S2: PCR-RFLP pattern after restriction with Tru1 of a product of 366 bp. M- molecular marker 50 bp; Figure S3: PCR-RFLP pattern after AluI restriction PCR-RFLP pattern after AluI restriction of MC1R.
Author Contributions
Conceptualization, D.D and M.K.; methodology, D.D., A.V., M.K; software, M.K.; validation, D.D., A.V. and G.K.; formal analysis, I.D. and M.B-S.; investigation, G.K, G.S and M.I.; resources, D.D., A.V.; data curation, D.D and M.K.; writing—original draft preparation, M.K., D.D.; writing—review and editing, M.K. and D.D; supervision, I.D and M.B-S..; project administration, D.D.; funding acquisition, D.D. All authors have read and agreed to the published version of the manuscript.
Funding
The research leading to these results received funding from the Ministry of Education and Science of the Republic of Bulgaria under the National Scientific Program INTELLIGENT ANIMAL HUSBANDRY, grant agreement no. Д01-62/18.03.2021.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data supporting the reported results can be requested from the authors.
Acknowledgments
We thank people and shepherds from several Breeding Associations: Breeding association of Maritza sheep breeds, Association of sheep breeders in Bulgaria – Balkan, Breeding Association of local sheep breed, Association for breeding of Stara Zagora sheep in Bulgaria who helped during the sampling and technical assistance of Petia Zheliazkova and Valentina Petrova from Breeding Association of Maritza sheep breeds.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Fontanesi, L.; Beretti, F.; Riggio, V.; Dall’Olio, S.; Calascibetta, D.; Russo, V.; Portolano, B. Sequence Characterization of the Melanocortin 1 Receptor (MC1R) Gene in Sheep with Different Coat Colours and Identification of the Putative e Allele at the Ovine Extension Locus. Small Ruminant Research 2010, 91, 200–207. [Google Scholar] [CrossRef]
- Barsh, G.; Gunn, T.; He, L.; Schlossman, S.; Duke-Cohan, J. Biochemical and Genetic Studies of Pigment-Type Switching. Pigment Cell Research 2000, 13, 48–53. [Google Scholar] [CrossRef]
- Lundie, R.S. The Genetics of Colour in Fat-Tailed Sheep: A Review. Trop Anim Health Prod 2011, 43, 1245–1265. [Google Scholar] [CrossRef]
- Russell, E.S. Pigmentation and Inheritance: Comparative Genetics of Coat Colour in Mammals. A. G. Searle. Logos Press, London; Academic Press, New York, 1968. Xii + 308 Pp., Illus. $17.50. Science 1968, 161, 677–678. [Google Scholar] [CrossRef]
- Koseniuk, A.; Ropka-Molik, K.; Rubiś, D.; Smołucha, G. Genetic Background of Coat Colour in Sheep. Arch. Anim. Breed. 2018, 61, 173–178. [Google Scholar] [CrossRef]
- Zhang, L.; Liang, L.; Zhang, X.; Li, W.R.; Li, H.; He, S. Understanding Key Genetic Make-up of Different Coat Colour in Bayinbuluke Sheep through a Comparative Transcriptome Profiling Analysis. Small Ruminant Research 2023, 226, 107028. [Google Scholar] [CrossRef]
- Yao, L.; Bao, A.; Hong, W.; Hou, C.; Zhang, Z.; Liang, X.; Aniwashi, J. Transcriptome Profiling Analysis Reveals Key Genes of Different Coat Color in Sheep Skin. PeerJ 2019, 7, e8077. [Google Scholar] [CrossRef]
- Bultman, S.J.; Michaud, E.J.; Woychik, R.P. Molecular Characterization of the Mouse Agouti Locus. Cell 1992, 71, 1195–1204. [Google Scholar] [CrossRef]
- Klungland, H.; Vage, D.I.; Gomez-Raya, L.; Adalsteinsson, S.; Lien, S. The Role of Melanocyte-Stimulating Hormone (MSH) Receptor in Bovine Coat Color Determination. Mammalian Genome 1995, 6, 636–639. [Google Scholar] [CrossRef] [PubMed]
- Ollmann, M.M.; Lamoreux, M.L.; Wilson, B.D.; Barsh, G.S. Interaction of Agouti Protein with the Melanocortin 1 Receptor in Vitro and in Vivo. Genes & Development 1998, 12, 316–330. [Google Scholar] [CrossRef]
- Lu, D.; Willard, D.; Patel, I.R.; Kadwell, S.; Overton, L.; Kost, T.; Luther, M.; Chen, W.; Woychik, R.P.; Wilkison, W.O.; et al. Agouti Protein Is an Antagonist of the Melanocyte-Stimulating-Hormone Receptor. Nature 1994, 371, 799–802. [Google Scholar] [CrossRef] [PubMed]
- Våge, D.I.; Fleet, M.R.; Ponz, R.; Olsen, R.T.; Monteagudo, L.V.; Tejedor, M.T.; Arruga, M.V.; Gagliardi, R.; Postiglioni, A.; Nattrass, G.S.; et al. Mapping and Characterization of the Dominant Black Colour Locus in Sheep. Pigment Cell Research 2003, 16, 693–697. [Google Scholar] [CrossRef] [PubMed]
- Parsons, Y.M.; Fleet, M.R.; Cooper, D.W. The Agouti Gene: A Positional Candidate for Recessive Self-Colour Pigmentation in Australian Merino Sheep. Aust. J. Agric. Res. 1999, 50, 1099. [Google Scholar] [CrossRef]
- Beraldi, D.; McRae, A.F.; Gratten, J.; Slate, J.; Visscher, P.M.; Pemberton, J.M. Development of a Linkage Map and Mapping of Phenotypic Polymorphisms in a Free-Living Population of Soay Sheep ( Ovis Aries ). Genetics 2006, 173, 1521–1537. [Google Scholar] [CrossRef] [PubMed]
- Lauvergne, J.-J.; Raffier, P. Génétique de La Couleur de La Toison Des Races Ovines Françaises Solognote, Bizet et Berrichonne. Genet Sel Evol 1975, 7, 263. [Google Scholar] [CrossRef] [PubMed]
- Tarach, P. Application of Polymerase Chain Reaction-Restriction Fragment Length Polymorphism (RFLP-PCR) in the Analysis of Single Nucleotide Polymorphisms (SNPs). FBOe 2021, 17, 48–53. [Google Scholar] [CrossRef]
- Våge, D.I.; Klungland, H.; Lu, D.; Cone, R.D. Molecular and Pharmacological Characterization of Dominant Black Coat Color in Sheep. Mammalian Genome 1999, 10, 39–43. [Google Scholar] [CrossRef]
- Royo, L.J.; Álvarez, I.; Arranz, J.J.; Fernández, I.; Rodríguez, A.; Pérez-Pardal, L.; Goyache, F. Differences in the Expression of the ASIP Gene Are Involved in the Recessive Black Coat Colour Pattern in Sheep: Evidence from the Rare Xalda Sheep Breed. Animal Genetics 2008, 39, 290–293. [Google Scholar] [CrossRef]
- Fontanesi, L.; Beretti, F.; Riggio, V.; Dall’Olio, S.; Calascibetta, D.; Russo, V.; Portolano, B. Sequence Characterization of the Melanocortin 1 Receptor (MC1R) Gene in Sheep with Different Coat Colours and Identification of the Putative e Allele at the Ovine Extension Locus. Small Ruminant Research 2010, 91, 200–207. [Google Scholar] [CrossRef]
- Yang, G.-L.; Fu, D.-L.; Lang, X.; Wang, Y.-T.; Cheng, S.-R.; Fang, S.-L.; Luo, Y.-Z. Mutations in MC1R Gene Determine Black Coat Color Phenotype in Chinese Sheep. The Scientific World Journal 2013, 2013, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Stamatis, C.; Giannoulis, Th.; Galliopoulou, E.; Billinis, Ch.; Mamuris, Z. Genetic Analysis of Melanocortin 1 Receptor Gene in Endangered Greek Sheep Breeds. Small Ruminant Research 2017, 157, 54–56. [Google Scholar] [CrossRef]
- Norris, B.J.; Whan, V.A. A Gene Duplication Affecting Expression of the Ovine ASIP Gene Is Responsible for White and Black Sheep. Genome Res. 2008, 18, 1282–1293. [Google Scholar] [CrossRef]
- Cavalcanti, L.C.G.; Moraes, J.C.F.; Faria, D.A.D.; McManus, C.M.; Nepomuceno, A.R.; Souza, C.J.H.D.; Caetano, A.R.; Paiva, S.R. Genetic Characterization of Coat Color Genes in Brazilian Crioula Sheep from a Conservation Nucleus. Pesq. agropec. bras. 2017, 52, 615–622. [Google Scholar] [CrossRef]
- Rochus, C.M.; Westberg Sunesson, K.; Jonas, E.; Mikko, S.; Johansson, A.M. Mutations in ASIP and MC1R : Dominant Black and Recessive Black Alleles Segregate in Native Swedish Sheep Populations. Animal Genetics 2019, 50, 712–717. [Google Scholar] [CrossRef]
- Dimov, D.; Vuchkov, A. Sheep Genetic Resources in Bulgaria with Focus on Breeds with Coloured Wool. GenResJ 2021, 2, 11–24. [Google Scholar] [CrossRef]
- The Genetics of the Dog; CABI Books; Ruvinsky, A., Sampson, J., C.A.B., International, Eds.; CABI: Wallingford, 2001; ISBN 978-0-85199-520-5. [Google Scholar]
- Fontanesi, L.; Dall’Olio, S.; Beretti, F.; Portolano, B.; Russo, V. Coat Colours in the Massese Sheep Breed Are Associated with Mutations in the Agouti Signalling Protein (ASIP) and Melanocortin 1 Receptor (MC1R) Genes. Animal 2011, 5, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Fontanesi, L.; Beretti, F.; Dall’Olio, S.; Portolano, B.; Matassino, D.; Russo, V. A Melanocortin 1 Receptor ( MC1R ) Gene Polymorphism Is Useful for Authentication of Massese Sheep Dairy Products. Journal of Dairy Research 2011, 78, 122–128. [Google Scholar] [CrossRef]
- Yang, G.-L.; Fu, D.-L.; Lang, X.; Wang, Y.-T.; Cheng, S.-R.; Fang, S.-L.; Luo, Y.-Z. Mutations in MC1R Gene Determine Black Coat Color Phenotype in Chinese Sheep. The Scientific World Journal 2013, 2013, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Fontanesi, L.; Rustempašić, A.; Brka, M.; Russo, V. Analysis of Polymorphisms in the Agouti Signalling Protein (ASIP) and Melanocortin 1 Receptor (MC1R) Genes and Association with Coat Colours in Two Pramenka Sheep Types. Small Ruminant Research 2012, 105, 89–96. [Google Scholar] [CrossRef]
- Sponenberg, P.; Reed, C. Effective Strategies for Local Breed Definition and Conservation. 2016.
- A., T. Allelic and Genotypic Frequencies of ASIP and MC1R Genes in Four West African Sheep Populations. Afr. J. Biotechnol. 2012, 11. [CrossRef]
- Colour Inheritance in Icelandic Sheep and Relation between Colour, Fertility and Fertilization. 1970.
- Fan, R.; Xie, J.; Bai, J.; Wang, H.; Tian, X.; Bai, R.; Jia, X.; Yang, L.; Song, Y.; Herrid, M.; et al. Skin Transcriptome Profiles Associated with Coat Color in Sheep. BMC Genomics 2013, 14, 389. [Google Scholar] [CrossRef] [PubMed]
- Gebreselassie, G.; Berihulay, H.; Jiang, L.; Ma, Y. Review on Genomic Regions and Candidate Genes Associated with Economically Important Production and Reproduction Traits in Sheep (Ovies Aries). Animals 2019, 10, 33. [Google Scholar] [CrossRef]
- Baxter, L.L.; Hou, L.; Loftus, S.K.; Pavan, W.J. Spotlight on Spotted Mice: A Review of White Spotting Mouse Mutants and Associated Human Pigmentation Disorders. Pigment Cell Research 2004, 17, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Najafi, M. Comparative Analysis of Gene Structure in 5-Flanking Region of Mc1r Gene in Indigenous Sheep Breeds 2024.
- Martin, P.M.; Palhière, I.; Ricard, A.; Tosser-Klopp, G.; Rupp, R. Correction: Genome Wide Association Study Identifies New Loci Associated with Undesired Coat Color Phenotypes in Saanen Goats. PLoS ONE 2017, 12, e0186029. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, W.; Liu, C.; Peng, X.; Lin, J.; He, S.; Li, X.; Han, B.; Zhang, N.; Wu, Y.; et al. Alteration of Sheep Coat Color Pattern by Disruption of ASIP Gene via CRISPR Cas9. Sci Rep 2017, 7, 8149. [Google Scholar] [CrossRef]
- Gratten, J.; Pilkington, J.G.; Brown, E.A.; Beraldi, D.; Pemberton, J.M.; Slate, J. The Genetic Basis of Recessive Self-Colour Pattern in a Wild Sheep Population. Heredity 2010, 104, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Amin, M.; Masoudi, A.A.; Amirinia, C.; Emrani, H. Molecular Study of the Extension Locus in Association with Coat Colour Variation of Iranian Indigenous Sheep Breeds. Russ J Genet 2018, 54, 464–471. [Google Scholar] [CrossRef]
- Castanheira, M.; Paiva, S.R.; Louvandini, H.; Landim, A.; Fiorvanti, M.C.S.; Dallago, B.S.; Correa, P.S.; McManus, C. Use of Heat Tolerance Traits in Discriminating between Groups of Sheep in Central Brazil. Trop Anim Health Prod 2010, 42, 1821–1828. [Google Scholar] [CrossRef]
- Muniz, M.M.M.; Caetano, A.R.; McManus, C.; Cavalcanti, L.C.G.; Façanha, D.A.E.; Leite, J.H.G.M.; Facó, O.; Paiva, S.R. Application of Genomic Data to Assist a Community-Based Breeding Program: A Preliminary Study of Coat Color Genetics in Morada Nova Sheep. Livestock Science 2016, 190, 89–93. [Google Scholar] [CrossRef]
- Pallotti, S.; Chandramohan, B.; Pediconi, D.; Nocelli, C.; La Terza, A.; Renieri, C. Interaction between the Melanocortin 1 Receptor ( MC1R ) and Agouti Signalling Protein Genes ( ASIP ), and Their Association with Black and Brown Coat Colour Phenotypes in Peruvian Alpaca. Italian Journal of Animal Science 2020, 19, 1518–1522. [Google Scholar] [CrossRef]
- Lundie, R.S. The Genetics of Colour in Fat-Tailed Sheep: A Review. Trop Anim Health Prod 2011, 43, 1245–1265. [Google Scholar] [CrossRef]
- Getachew, T.; Abebe, A.; Gizaw, S.; Rischkowsky, B.; Bisrat, A.; Haile, A. Coat Color Alterations over the Years and Their Association with Growth Performances in the Menz Sheep Central Nucleus and Community-Based Breeding Programs. Trop Anim Health Prod 2020, 52, 2977–2985. [Google Scholar] [CrossRef]
- An Introduction to Heritage Breeds, D.P. Sponenberg, J. Beranger and A. Martin Edited by D. Burns & A. Larkin Hansen Storey Publishing, North Adams Published in 2014, Pp. 240 ISBN 978-1-61212-125-3 (Pbk.: Alk. Paper) ISBN 978-1-61212-130-7 (Hardcover: Alk. Paper) e-ISBN 978-1-61212-462-9. Anim. Genet. Resour. 2015, 56, 155. [Google Scholar] [CrossRef]
- Sponenberg, D.P.; Martin, A.; Couch, C.; Beranger, J. Conservation Strategies for Local Breed Biodiversity. Diversity 2019, 11, 177. [Google Scholar] [CrossRef]
Figure 1.
Pictures of Bulgarian native sheep breeds with different coat color: a – Patch-faced Maritza; b – piebald phenotype in Patch-faced Maritza breed; c – Piebald phenotype in White Maritza breed; d – native Starozagorska; e – White Maritza breed; f – Dubenska; g – native Karnobatska; h – Karakachanska; i – Copper-red Shumenska; .
Figure 1.
Pictures of Bulgarian native sheep breeds with different coat color: a – Patch-faced Maritza; b – piebald phenotype in Patch-faced Maritza breed; c – Piebald phenotype in White Maritza breed; d – native Starozagorska; e – White Maritza breed; f – Dubenska; g – native Karnobatska; h – Karakachanska; i – Copper-red Shumenska; .

Figure 2.
Genotype distribution among pigmented animals after AluI restriction.
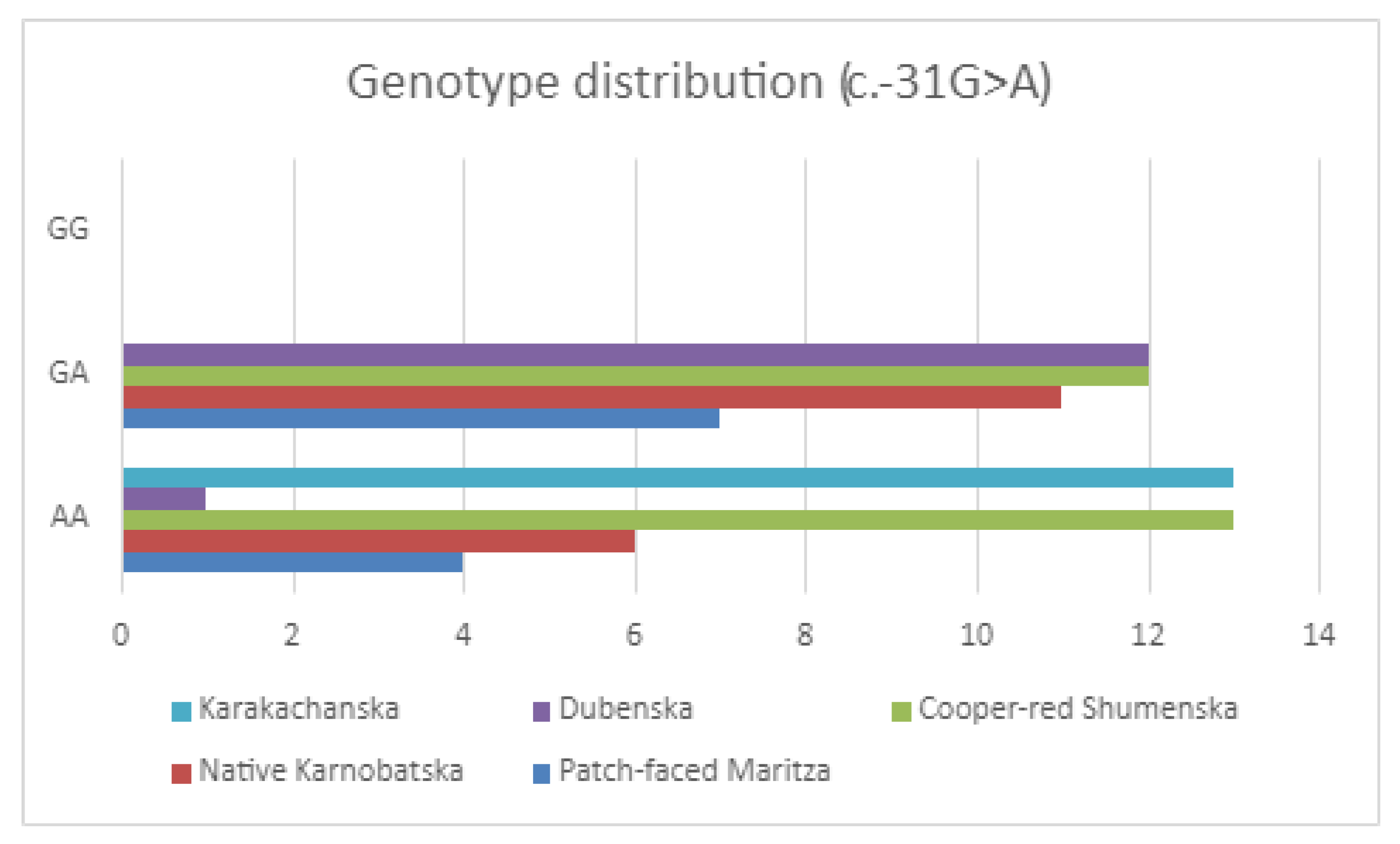
Figure 3.
Multiple alignment for detection of five SNP-s in MC1R gene.
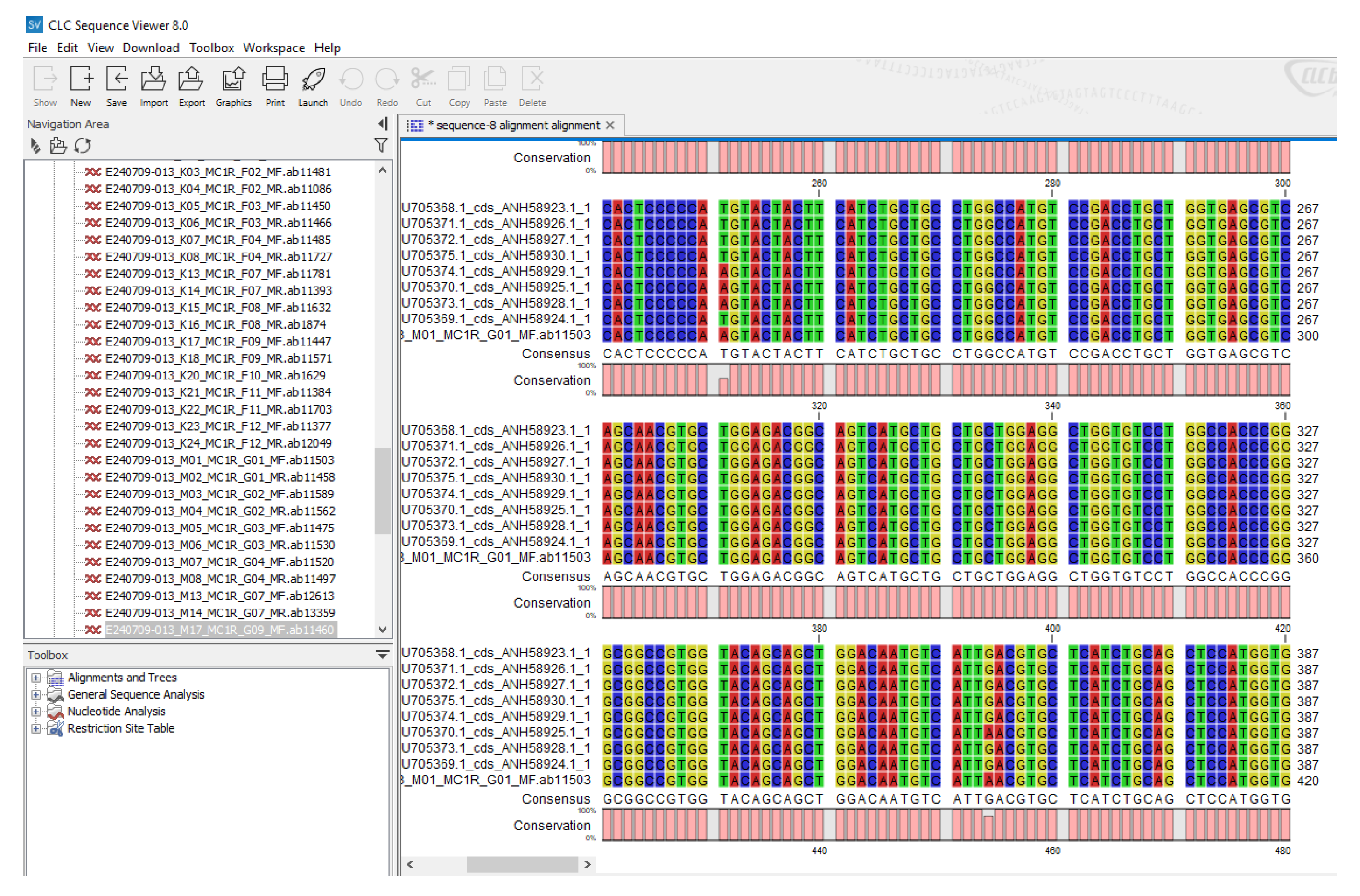
Figure 4.
Distribution of haplotypes detected after the sequencing analyzis of pigmented breeds.
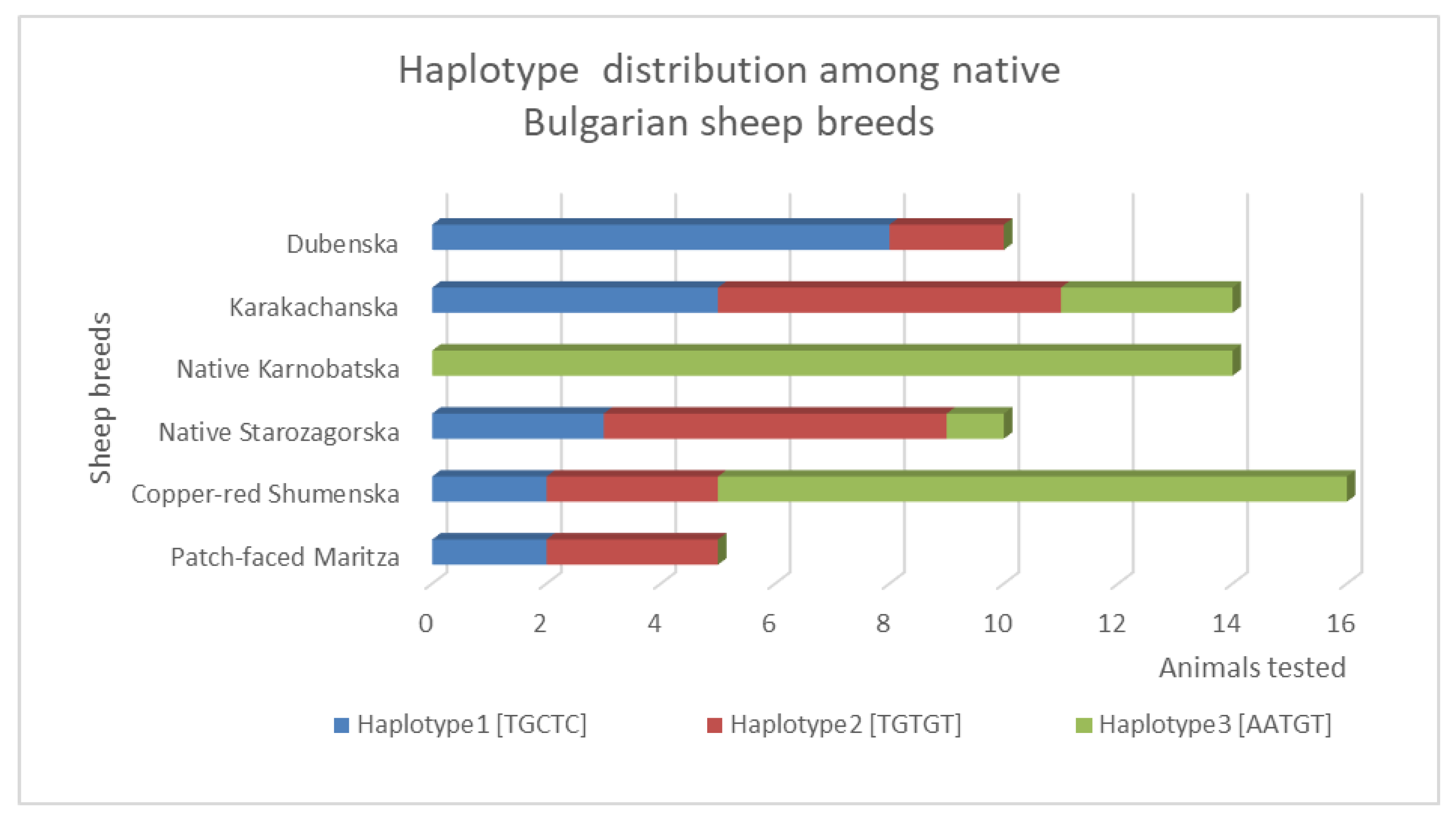
Figure 5.
An electropherogram representing the positions (arrow) of three of the mutations with heterozygous peaks of pigmented animals assigned to haplotype 1.
Figure 5.
An electropherogram representing the positions (arrow) of three of the mutations with heterozygous peaks of pigmented animals assigned to haplotype 1.
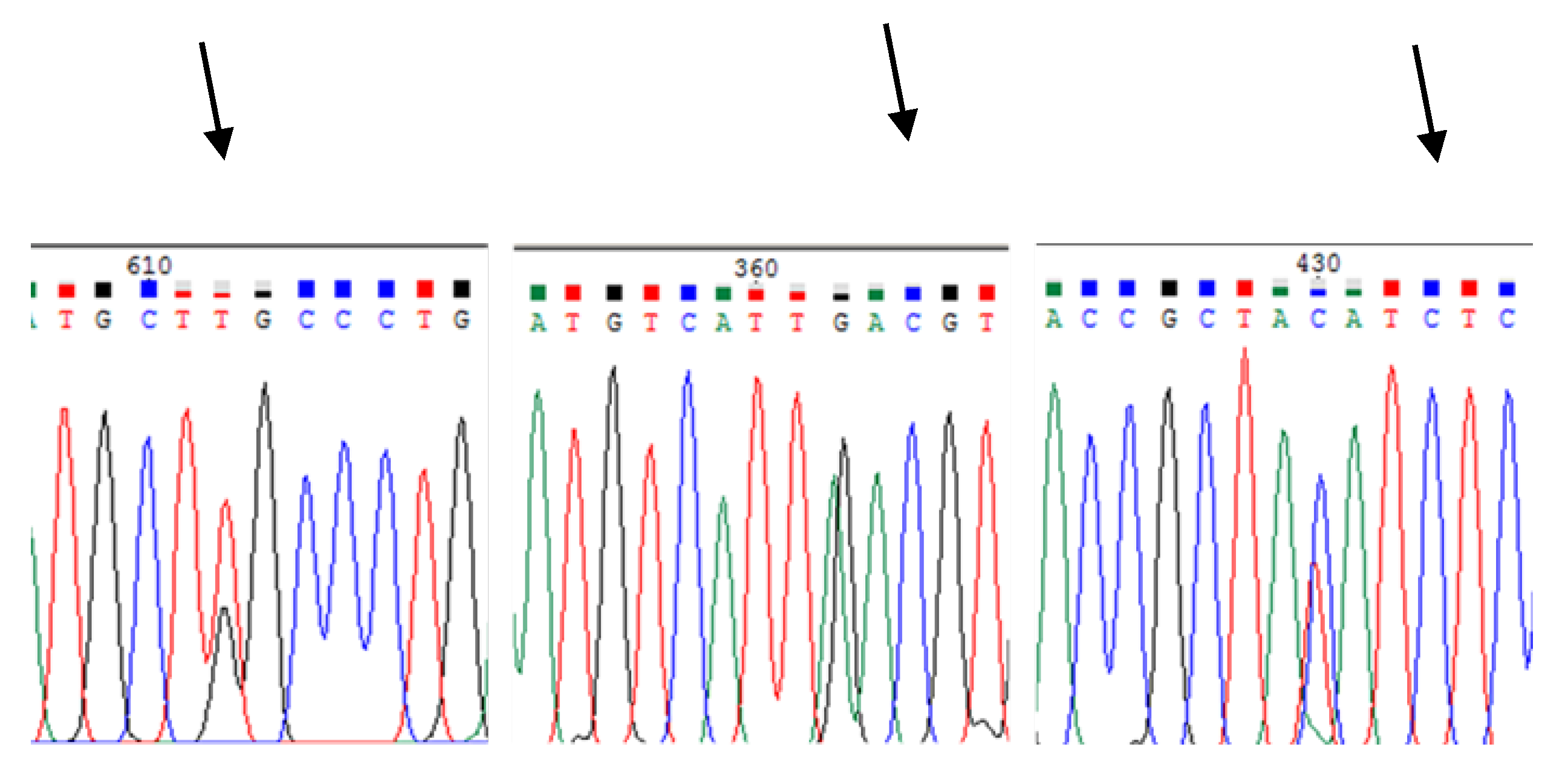

Table 1.
Primers, PCR conditions, and genotyping analyses for ASIP and MC1R polymorphisms. .
| Gene | Primer pair name | Forward and reverse primers (5′ –3′ ) | PCRa | Use |
|---|---|---|---|---|
| ASIP | ex2_del | CAGCAGGTGGGGTTGAGCACGCTGC CTACCTGACTGCCTTCTCTG |
59/25/174 | Fragment analysis (genotyping of the exon 2 deletions) |
| ASIP | Agt17 Agt18 Agt16 |
GTTTCTGCTGGACCTCTTGTTC GTGCCTTGTGAGGTAGAGATGGTGTT CAGCAATGAGGACGTGAGTTT |
58/23/238–242 | Fragment analysis (duplication breakpoint analysis, asymmetric competitive PCR) |
| MC1R | MC1R-5’UTR | GCAACTGCACATCCAGAGAA CGCAGGGAGCAGGAAAGGGTGCTAG |
58/35/202 | PCR-RFLP analysis with AluI (c.-31G>A) |
| MC1R | 3 MC1R | GTGAGCGTCAGCAACGTG ACATAGAGGACGGCCATCAG |
61/35/366 | PCR-RFLP analysis with Tru1I (c.361G>A; p.D121N) |
| MF: MR: |
GAGAGCAAGCACCCTTTCCT GAGAGTCCTGTGATTCCCCT |
60/35/1170 | Complete coding region of MC1R |
Annealing temperature (0C)/no. of cycles/amplified fragment size (bp). .
Table 2.
Distribution of the analyzed mutations (duplication, deletion, and the presence simultaneously), in analyzed sheep breeds. .
Table 2.
Distribution of the analyzed mutations (duplication, deletion, and the presence simultaneously), in analyzed sheep breeds. .
| Breeds | Phenotype | n | duplication | deletion | dupl+del | -/- |
|---|---|---|---|---|---|---|
| Patch-faced Maritza | 76 | 6 | 43 | 11 | 16 | |
| Patch-faced* | 59 | 4 | 35 | 6 | 14 | |
| Fully pigmented** | 10 | 1 | 3 | 4 | 2 | |
| Piebald | 7 | 1 | 5 | 1 | - | |
| White Maritza | 26 | 1 | 12 | 10 | 3 | |
| White | 17 | - | 8 | 6 | 1 | |
| Fully pigmented | 6 | 1 | 4 | 2 | 1 | |
| Piebald | 3 | - | - | 2 | 1 | |
| Dubenska | 22 | - | 16 | - | 6 | |
| White | 2 | - | 1 | - | 1 | |
| Fully pigmented | 20 | - | 15 | - | 5 | |
| native Starozagorska | 21 | 4 | 17 | - | - | |
| White | 4 | 4 | - | - | - | |
| Fully pigmented | 17 | - | 17 | - | - | |
|
Copper-red Shumenska |
38 | 1 | 37 | - | - | |
| Fully pigmented | 38 | - | - | - | - | |
| native Karnobatska | 33 | 10 | 23 | - | - | |
| Fully pigmented | 33 | - | - | - | - | |
| Karakachanska | 31 | |||||
| Fully pigmented | 31 | - | 11 | - | 20 |
*Patch-faced - term used to refer to a phenotype in which sheep have white color of body but, on the head, ears and legs, the animals have a typical black spots. **Fully pigmented – term used to summarize different nuances of reddish-brown to black.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



