
Submitted:
30 June 2025
Posted:
30 June 2025
You are already at the latest version
Abstract
The soil-borne fungi, Setophoma terrestris and Fusarium spp., are often associated with pink root although the etiology of the disease remains doubtful. Although recognized as primary inoculum, studies show conflicting views on the formation of chlamydospores and microsclerotia in Setophoma. Therefore, this study aims to clarify the etiology of the pink root of garlic and onion and the for-mation of chlamydospores and microsclerotia in Setophoma. The isolates were obtained from symptomatic tissues of garlic, leeks, brachiaria, onions, chives and maize collected from seven different states in Brazil. Representative isolates were selected for pathogenicity tests. Sequence comparison of the tubulin gene showed Setophoma (n=50) and Fusarium clades (n=25). Garlic and onion plants inoculated with Setophoma showed pink root symptoms while plants inoculated with different Fusarium isolates remained asymptomatic. Multigene analysis of pathogenic isolates confirms only Setophoma terrestris causing pink root in garlic and onion. In addition, brachiaria, chives, and leeks are newly identified hosts of this pathogen in Brazil. To our knowledge, the main source of primary inoculum of the disease are chlamydospores, pycnidia, colonized roots of garlic, onion and plant debris of susceptible crops. The new information obtained in this study will be fundamental for researchers in the development of genotypes that are resistant to pink root and will help the efficient management of the disease.
Keywords:
diagnosis
; Alliaceae
; Pyrenochaeta
; Phoma
; taxonomy
1. Introduction
China and India are the world’s largest producers of garlic and onions. In 2022, Brazil was the 11th and 13th largest world producer of onion (1,65 million tons) and garlic (181,1 thousand tons), respectively. Due to the insufficiency of production to satisfy national demand, Brazil regularly imports from other countries [1]. The main obstacle to expanding garlic and onion cultivation is the high cost of production, primarily due to the occurrence of diseases [2].
Pink root disease is the main disease affecting garlic and onions during periods of drought stress and high temperatures [3,55]. It is characterized by an initial light pink discoloration of infected roots, which gradually become shortened, water-soaked, and dark purple before collapsing and dying. The continuous infection of newly formed roots compromises nutrient and water uptake, leading to stunted growth, leaf etiolation and wilting, and ultimately the formation of undersized, poor-quality bulbs with reduced storage potential [3]. Implementing control measures against the pathogen can enhance production by as much as 70% [62,65].
This disease was first documented in the state of Texas, United States, in 1917 [4]. In Brazil, the disease was first reported affecting onions in the Zona da Mata Mineira region in 1960 [5]. Subsequently, it was also found in the states of Rio Grande do Sul and São Paulo and currently occurs in all garlic and onion producing regions [2,6,7]. Despite reports of the pink root causal agent in China, it is listed as a quarantine pest by the Ministry of Agriculture and Rural Affairs of the People’s Republic of China [63,66].
The first causal agent of the disease was proposed as Fusarium malli [9]. Later, others Fusarium species were associated with pink root [8,10,11]. The immersion of roots in a solution of mercuric chloride (500:1) for 3 minutes revealed the presence of Phoma sp. [12]. Besides the isolation approach, pathogenicity tests revealed that Phoma terrestris is the causal agent of pink root, while Fusarium species were associated with rotten roots [12,13]. The simultaneous inoculation of both pathogens confirmed the opportunistic nature of Fusarium species, which failed to induce pink root symptoms [13]. Neverthless, Fusarium species are often associated with pink root [14,15,16,17,18,19].
The frequent recovery of isolates that form pycnidia containing setae indicated P. terrestris as the main etiological agent of pink root [20]. Based on the presence of setae on the pycnidium, P. terrestris was transferred to Pyrenochaeta terrestris (Hansen) Gorenz [21]. Later, based on phylogenetic analysis, this taxon was reclassified as Setophoma terrestris. This species is characterized by the presence of setose pycnidia, phialidic conidiogenic cells, and ellipsoidal to subcylindrical, aseptate, and hyaline conidia [22].
Although the taxonomic studies on Setophoma does not mention the presence of chlamydospores and microsclerotia, these resting structures are considered the primary inoculum source of pink root [3,19,23]. Since it is uncertain whether species of Setophoma and Fusarium could be associated with pink root, especially considering that most identifications are based on only a few isolates or limited to the examination of morphological data, it is crucial to characterize fungal isolates obtained from plant hosts exhibiting typical symptoms of the disease.
Furthermore, it has been shown that a molecular perspective, combined with morphological data, is required to resolve plant pathogen species complexes. This combined approach has proven effective in revealing previously uncharacterized species affecting different crops [24,25]. For example, molecular characterization of Setophoma isolates on tea plants revealed a total of four new species [26]. Although molecular characterization studies involving a large number of Setophoma isolates from garlic and onion are still lacking, various molecular diagnostic methods have been developed for the detection of this pathogen [19,63,64].
In Brazil, the frequent association of Fusarium with pink root symptoms has led growers to believe that this fungus may also be involved in the etiology of the disease. Therefore, this study aims to clarify the etiology of the pink root of garlic and onion and the formation of chlamydospores and microsclerotia in Setophoma.
2. Materials and Methods
2.1. Obtaining and Preserving Isolates
Root samples (n=75) were obtained in garlic, onion, leeks, brachiaria, chives and maize from 17 cities in the Brazilian states of Distrito Federal, Goiás, Minas Gerais, São Paulo, Santa Catarina, Paraná, and Bahia. Fungal isolation into pure culture was carried out by the direct method [27] in samples obtained from garlic, leeks, brachiaria, onions, chives and maize with typical symptoms of pink root in Petri dishes containing potato-dextrose-agar (PDA) medium (1,000 mL H2O, 4 g/L potatoes infusion, 25g/L agar, and 20g/L dextrose). After 7 days, pure cultures were established by transferring a fragment of a hyphal tip to a new Petri dish containing PDA. The isolates (Supplementary Table S1) were stored at Coleção de Culturas Fúngicas da UnB (CCUB, Universidade de Brasília, Brasília, Brazil) under 18°C in 2 mL microtubes by sterile glycerol 10% (v/v), Castellani method and glycerol 10% (v/v) in -80°C ultrafreezer.
2.2. Total DNA Extraction
The fungal isolates were grown on Erlenmeyer flasks containing Potato-Broth (PB; 1,000 mL H2O, 4 g/L potatoes infusion, and 20g/L dextrose) medium, at 25°C for 7 days, with a 12 h photoperiod. The mycelium (100 µg) was collected using a sterile toothpick and deposited in 1.5 mL microtubes containing 20 µL of Tris-EDTA (TE) buffer. The total DNA extraction was performed using Wizard Genomic Purification Kit (Promega®) [28]. The presence and quality of total DNA were analyzed on 1% agarose gel electrophoresis with 1 Kb ladder, stained with GelRed (Biotium®), and visualized under UV light. The DNA samples were stored at -20°C.
2.3. Amplification and Sequencing
Partial sequences of the gene β-tubulin (tub) were amplified using the primer set T1/T22 [29]. This genomic region was used to make an initial identification of 75 isolates using BLASTN similarity scores. Twelve isolates of different clades and locations identified based on tub sequence data were selected to assign definitive species demarcations for Setophoma isolates. Then, partial nucleotide sequences of the four following genomic regions were sequenced: large subunit of ribosomal DNA (lsu-its), translation elongation factor (tef), glyceraldehyde-3-phosphate dehydrogenase (gapdh), and RNA polymerase subunit II (rpb2), using the primer pairs LR5-V9G [30,31], EF1F-EF2R [32], GPD1-GPD2 [33], and 5F2 and 7cR [34,35], respectively. The PCR amplifications were performed in a final volume of 12.5μL: 6.25 μL of MyTaq MasterMix 2x (Bioline, EUA), 0.3μL (10 pmol/µL) of each primer, 4.25 μL of nuclease-free water, and 1 μl of template DNA (25 ng/μL). The cycling conditions were: initial denaturation at 95°C for 1.5 min, followed by 35 cycles at 95°C for 20 s, 53°C (tub) for 45 s, 72°C for 1 min, and a final extension for 5 min. Different annealing temperatures were used according to the different genomic region amplified: 53°C (lsu-its), 56°C (tef), 54°C (gapdh and rpb2). PCR products were purified and bidirectionally Sanger-sequenced.
2.4. Phylogenetic Analyses
The DNA sequences and contig assembly of all isolates were checked for quality and ambiguity analysis through DNA Dragon software (https://www.dna-dragon.com). A Bayesian Inference (BI) phylogenetic tree was initially reconstructed using the tub sequences of 75 isolates of this study. So, BLAST searches were conducted on NCBI and Fusarioid-ID [36] for pairwise DNA alignments. Finally, a BI concatenate tree using tub, tef, gapdh, lsu, and its to Setophoma was reconstructed. Didymella pinodella CBS 531.66 was used as outgroup. The alignments were made using Mega v.7 software [37], which were concatenate posteriorly, and the best nucleotide substitution models were determined for each genomic region with MrModeltest 2.3 [38]. The website CIPRES [39] was used to run MrBayes v 3.2.1 [40]. The Markov Chain Monte Carlo (MCMC) analysis was made under analysis of 10 million generations, sampling every 1,000, with subsequent disposal of 25% first trees in analysis, followed by the assembly of consensus tree through the 7,500 remaining trees of analysis with annotation of the posterior probability (PP) values. The consensus tree was visualized in FigTree v.1.4 [41].
2.5. Morphological Characterization of Setophoma terrestris
The morphological characterization of a representative S. terrestris isolate was performed using monohyphal culture grown on oatmeal-agar (OA; 1,000 mL H2O, 30 g oat and 15 g agar) medium on Petri dishes during seven days at 25 °C, under 24h of near-ultraviolet light (UV-A or black light) to produce conidia and pycnidia. The morphological examination and documentation of pycnidia was observed and recorded by using a Leica model 205C stereomicroscope (Leica Microsystems, Nassloch, Germany) with a Leica DFC295 digital camera and Leica Qwin-Plus software. Micromorphological characteristics were analyzed by cutting pycnidia in a cryostat Leica CM1860 (Leica Microsystems, Nassloch, Germany) and mounting in clear lactoglycerol. Thirty-five measurements for conidia were carried out under magnification of ×1,000 using a Leica DM2500 light microscope (Leica Microsystems, Nassloch, Germany) equipped with a Leica DFC 490 digital camera, coupled to a computer containing Leica Qwin-Plus software.
To induce the formation of chlamydospores and microsclerotia, six representative fungal isolates were grown in: OA; synthetic low-nutrient agar (SNA; 1,000 mL H2O, KH2PO4 1 g, KNO3 1 g, MgSO4.7H2O 0.5 g, KCl 0.5 g, glucose 0.2 g, sucrose 0.2 g, and agar 20 g), malt-extract-agar (MEA; 20 g/L malt extract and 20g/L agar), PDA, PD, corn-meal-agar (CMA; 1,000 mL H2O, 50 g corn meal infused, and 15 g agar), millet substrate and modified yeast extract (MYE; sodium phosphate 2 g, magnesium sulfate 1 g, glucose 20 g, yeast extract 10 g). The isolates were incubated in the dark at 25 °C for 20 days. Subsequently, fungal structures were mounted on slides with clear lactoglycerol and analyzed under a Leica DM2500 light microscope (Leica Microsystems, Nassloch, Germany) equipped with a Leica DFC 490 digital camera, coupled to a computer containing Leica Qwin-Plus software.
2.6. Pathogenicity
Six representative isolates of S. terrestris (CCUB 2747, 2750, 2754, 2763, 2777 and 2795) selected based on region and host and 25 isolates of Fusarium-like, identified by molecular phylogeny (tub tree), were used in pathogenicity tests on onion and garlic plants. Firstly, plastic bags with millet (500 g) and distilled water (450 mL) were previously autoclaved at 121°C for 12 minutes. Twenty mycelial disks (5 mm in diameter) from colonies with 7-days-old in BDA were transferred to sterilized millet substrate. After 30 days, when the millet has been completely colonized, 30 g were transferred to 3-liter pots containing sterilized soil and then onion and garlic seedlings (30 days old) were transplanted near the inoculum. The inoculated genotypes were ‘Ito’ (garlic) and ‘BRS 367’ (onion). The control treatment consisted in 30 g of sterile millet. Finally, plants were removed after approximately 100 days and checked for pink-root symptoms and, subsequently, taken to fungi reisolating. The isolate exhibiting the highest aggressiveness on garlic and onion was selected, and a millet substrate was employed for pathogenicity assays on leeks, brachiaria, chives, and maize. Additionally, a second methodology was tested, where the seedlings suffered root cuts (~1 cm), and were immersed in spore suspensions calibrated at 1 × 104. The control treatment consisted in seedlings immersed in sterilized water.
3. Results
3.1. Phylogenetic Analysis
After a BLAST search, it was possible to verify the presence of Setophoma sp. (n = 50) and Fusarium spp. (n = 25). The 12 selected isolates for multigenic analysis were compared to all Setophoma species (Supplementary Table S1). The tub, tef, gapdh, its, and lsu matrices had in length, respectively: 506, 575, 564, 603 and 837 bp. Although the rpb2 sequences were not used in phylogenetic analysis, they were lodged in GenBank (Accession No. OM417590 to OM417601) for future studies and identification purposes. The concatenate alignment had 3,085 sites, which 2,105 were conserved and 679 parsimony informative. The BI tree was reconstructed considering the best nucleotide substitution model for each partition in the concatenate data, GTR+I+G (tub), GTR+G (tef), SYM+G (gapdh), HKY+G (its), and SYM+I (lsu). After multigenic analysis, it was confirmed that all isolates grouped to S. terrestris (Figure 1). These sequences were deposited in Genbank with the following codes: tub = ON159208 to ON159257; tef = ON159281 to ON159292; gapdh = ON159261 to ON159272, and lsu-its = OM397056 to OM397067.
3.2. Taxonomy
Setophoma terrestris (H.N. Hansen) Gruyter, Aveskamp and Verkley, Mycologia 102, 5, 1077 (2010) [GenBank: MB514659]
Basionym: Phoma terrestris H.N. Hansen, Phytopathology 19:699. 1929.
≡ Pyrenochaeta terrestris (H.N. Hansen) Gorenz, J.C. Walker and Larson, Phytopathology 38:838. 1948.
Conidiomata pycnidial, solitary to confluent, on upper surface or submerged in agar, globose to subglobose, setose, with papillate ostioles, medium to dark brown; pycnidial wall of pseudoparenchymatal cells (Figure 2C–E). Conidiophores reduced to conidiogenous cells lining inner cavity (Figure 2G). Conidiogenous cells hyaline, smooth, phialidic, discrete. Conidia aseptate, globose, subglobose, ellipsoidal to subcylindrical to subfusoid, 4.5 to 6.0 µm length (mean = 5.14 ± 0.45 µm) and 2.0 to 3.5 µm width (mean = 2.56 ± 0.29 µm) (Figure 2F). Terminal and intercalar chlamydospores were observed in MYE (Figure 2I).
3.3. Pathogenicity Test
The six representative isolates of S. terrestris were pathogenic to onion and garlic in millet substrate and spore suspension (Figure 2A,H). Disease visualization was more consistent when using the millet substrate, whereas the method involving root cuts immersed in spore suspensions resulted in some asymptomatic roots. Differently, plants inoculated with the 25 Fusarium isolates were asymptomatic in both methodologies. Thus, S. terrestris was confirmed as the only causal agent of pink root disease in Brazil. In addition, S. terrestris isolate CCUB2754 was pathogenic to leeks, Brachiaria, chives, and maize.
4. Discussion
Although Setophoma and Fusarium are found in leaves, soil, and the rhizosphere, the highest abundance is observed in roots [55]. In Brazil, the incidence of pink root is lower in regions with mild climates (South and Southeast) compared to regions with high temperatures (Midwest). Furthermore, the disease gradually increases in areas with successive crops and can drastically increase in plantations during the summer due to high temperatures combined with periods of drought stress [19,55,69,71].
All 25 Fusarium isolates obtained in this study were considered asymptomatic on onion and garlic roots, as no visible lesions or discoloration were observed following inoculation. Although non-pathogenic Fusarium species are frequently associated with pink root symptoms [13,18,19], some species are known to cause Fusarium basal rot [68]. Conversely, Setophoma isolates caused characteristic light to dark pink discoloration on the roots of brachiaria, chives, garlic, leeks, maize, and onion (Figure 3). In our study, only light to dark pink discoloration of infected roots was observed, which is likely attributable to the controlled experimental conditions and the timing of the assessments prior to bulb harvest. In contrast, field soils harbor an active microbiota and are frequently exposed to high temperatures combined with prolonged water stress, which may exacerbate symptom severity under natural conditions.
In this study, the Setophoma isolates (n=50) collected in the states of Bahia, Goiás, Distrito Federal (Midwest and Northeast) and Minas Gerais, Paraná, São Paulo, Santa Catarina (South and Southeast) were obtained from symptomatic tissues on Allium cepa (n=26), A. porrum (n=4), A. sativum (n=9), A. fistulosum (n=7), Brachiaria sp. (n=2), and Zea mays (n=2). Setophoma terrestris colonizes canola, carrot, cauliflower, bunching onion, corn, cowpea, cucumber, eggplant, elephant garlic, eschalote, leeks, lima bean, melon, oats, pea, pepper, potato, pumpkin, sorghum, soybean, spinach, sugarcane, rice, tomato, wheat, and weeds. Nevertheless, only maize, garlic, onion, pea, and tomato were previously recorded as hosts of this fungus in Brazil [3,19,42,43,44,45,46,47,48,49,50]. To our knowledge, this is first report of S. terrestris associated with pink root in brachiaria, chives, and leeks in Brazil.
The clade assignments based on the concatenated-sequence tree (lsu, its, tef, tub, and gapdh) confirms fifteen Setophoma species (Figure 1). There is a proposal to split Setophoma into three genera, with Setophoma stricto sensus including S. terrestris, S. brachypodii, and S. poaceicola [61]. Large-scale studies that investigate the pink root etiology are scarce in the literature. Although several Setophoma species have been reported on Camellia in China [26], concatenate analysis (lsu, its, tef, tub, and gapdh) of isolates obtained from different geographic regions and hosts confirmed only S. terrestris causing pink root. The absence of cryptic species in Setophoma terrestris is likely, given its role as a component of the soil microbiome and its lack of host specificity. Additionally, this fungus exhibits potentially antagonistic interactions with other soil organisms and is known to produce bioactive secondary metabolites, including cytotoxic polyketides [70].
Some studies report that S. terrestris survives in the soil due to the formation of chlamydospores and microsclerotia [19,23]. The presence of chlamydospores and microsclerotia were not observed in symptomatic plants and in pathogenicity tests. After isolation in pure culture, we attempted to induce the formation of these structures on OA, SNA, MEA, CMA, PDA, PD, millet substrate, and MYE. While microsclerotia were not observed under the experimental conditions of this study, chlamydospores were observed only on MYE. Efficient use of the supplied nitrogen source during early vegetative growth is essential, while subsequent nutrient depletion induces physiological stress that promotes chlamydospore formation [56]. Several studies on S. terrestris did not mention the presence of these structures [49,51,52,67]. In addition, none of the taxonomic descriptions of the different Setophoma species reported the presence of microsclerotia and chlamydospores [22,26,53,54,57,58,59,60]. Therefore, the main source of primary inoculum of pink root disease are chlamydospores, pycnidia, colonized roots of garlic, onion and plant debris of susceptible crops.
The confirmation that only S. terrestris causes pink root in garlic and onion, and the absence of cryptic species associated with this pathogen, is essential for validating disease diagnostic protocols. Its presence in the roots of brachiaria, chives, leeks, maize, and other hosts demonstrates the ability of this fungus to survive in alternative plants during the interseason. This new information will be fundamental for breeders and plant pathologists in the development of garlic and onion genotypes resistant to pink root, and will contribute to more efficient disease management in the field.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: GenBank accession numbers of tub, lsu, its, tef, gapdh e rpb2 partial sequences of Setophoma isolates included in this study.
Author Contributions
Conceptualization, G.H.S.P., A.R., V.R.O., V.L. and D.B.P.; methodology, G.H.S.P., T.F.S. and L.F.C.; formal analysis, G.H.S.P., T.F.S. and D.B.P.; investigation, G.H.S.P., T.F.S. and D.B.P.; resources, V.R.O., V.L. and D.B.P.; data curation, G.H.S.P., L.F.C., T.F.S.; writing—original draft preparation, G.H.S.P.; writing—G.H.S.P., A.R., V.R.O., V.L., D.B.P.; supervision, V.R.O., V.L. and D.B.P. All authors have read and agreed to the published version of the manuscript.
Funding
Please add: This research was funded by Fundação de Apoio à Pesquisa do Distrito Federal, FAPDF (Grant number: 00193.00000925/2019-12).
Acknowledgments
Thanks to the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq, Brazil) for providing a scholarship to Gustavo Henrique Silva Peixoto and research fellowships to Ailton Reis and Danilo B. Pinho.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- FAO - Food and Agriculture Organization. 2024. Faostats – FAO Statistical Databases. Available at: https://www.fao.org/faostat/en/#data [Accessed 11th April 2025].
- Reis, A.; Oliveira, V.R.; Lourenço Junior, V. Identificação e Manejo Da Raiz Rosada Da Cebola. Embrapa: Brasília, DF. 6p. (Comunicado Técnico 114) 2016, 6.
- Sumner, D.R. Pink Root. In Compendium of onion and garlic diseases and pests; Schwartz, H.F., Mohan, K., Eds.; American Phytopathological Society Press: St. Paul, Minnesota, 2008; pp. 18–20. [Google Scholar]
- Taubenhaus, J.J.; Johnson, A.D. Pink Root, a New Root Disease of Onions in Texas. Phytopathology 1917, 7, 59. [Google Scholar]
- Chaves, G.M.; Erickson, H.T. Pyrenochaeta terrestris (Hansen) Gorenz, J. C. Walker y Larson, Um Novo Fungo Da Cebola (Allium cepa L.) Em Minas Gerais. Revista Ceres 1960, 63, 112–114. [Google Scholar]
- Luz, N.L. Raízes Rosadas - Uma Nova Moléstia Da Cebola Para o Rio Grande Do Sul. Revista da Faculdade de Agronomia e Veterinária da Universidade Federal do Rio Grande do Sul 1967, 9, 159–165. [Google Scholar]
- Noda, H. Reação Da Cebola (Allium Cepa L.) a Pyrenochaeta Terrestris (Hansen), Gorenz, Walker e Larson, Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo: Piracicaba, 1981.
- Taubenhaus, J.J. PINK ROOT OF ONIONS. Science (1979) 1919, 49, 217–218. [Google Scholar] [CrossRef]
- Taubenhaus, J.J.; Mally, F.W. Pink Root Disease of Onions and Its Control in Texas. Texas. Agricultural Experiment Station. Bulletin 273 1921, 42. [Google Scholar]
- Sideris, C.P. Species of Fusarium Isolated from Onion Roots. Phytopathology 1924, 14, 211–216. [Google Scholar]
- Sideris, C.P. Species of Fusarium Isolated from Onion Roots. Phytopathology 1929, 19, 233–268. [Google Scholar]
- Hansen, H.N. “Pink-Root” Of Onions Caused By Phoma sp. Science (1979) 1926, 64, 525–525. [Google Scholar] [CrossRef]
- Hansen, H.N. Etiology of the Pink Root Disease of Onions. Phytopathology 1929, 19, 691–704. [Google Scholar]
- Du Plessis, S.J. Pink Root and Bulbrot of Onions; Union of South Africa: Pretoria, 1933; Vol. 124. [Google Scholar]
- Tims, E.C. Pink Root of Shallots, Allium ascalonicum. Plant Disease Reports 1953, 37, 533–537. [Google Scholar]
- Watson, R.D. Rapid Identification of the Onion Pink Root Fungus. Plant Disease Reports 1961, 45, 289. [Google Scholar]
- Kodama, F.; Sugawara, Y.; Yokoyama, T. Pink Root Rot of Onion Caused by Pyrenochaeta terrestris. Phytopathological Society of Japan 1976, 42, 320–321. [Google Scholar] [CrossRef]
- Awuah, R.T.; Lorbeer, J.W. A Procedure for Isolating Pyrenochaeta terrestris from Onion Roots. Annals of Applied Biology 1989, 114, 205–208. [Google Scholar] [CrossRef]
- Yoshida, N. Seasonal Dynamics of the Pink Root Fungus (Setophoma terrestris) in Rhizosphere Soil: Effect of Crop Species and Rotation. Plant Pathol 2022, 71, 361–372. [Google Scholar] [CrossRef]
- Gorenz, A.M.; Larson, R.H.; Walker, J.C. Factors Affecting Pathogenicity of Pink Root Fungus of Onions. J Agric Res 1949, 78, 1–18. [Google Scholar]
- Gorenz, A.M.; Walker, J.C.; Lardon, R.H. Morphology and Taxonomy of the Onion Pink-Root Fungus. Phytopathology 1948, 38, 831–840. [Google Scholar]
- de Gruyter, J.; Woudenberg, J.H.C.; Aveskamp, M.M.; Verkley, G.J.M.; Groenewald, J.Z.; Crous, P.W. Systematic Reappraisal of Species in Phoma Section Paraphoma, Pyrenochaeta and Pleurophoma. Mycologia 2010, 102, 1066–1081. [Google Scholar] [CrossRef]
- Biles, C.L.; Holland, M.; Ulloa-Godinez, M.; Clason, D.; Corgan, J. Pyrenochaeta terrestris Microsclerotia Production and Pigmentation on Onion Roots. HortScience 1992, 27, 1213–1216. [Google Scholar] [CrossRef]
- Nalim, F.A.; Samuels, G.J.; Wijesundera, R.L.; Geiser, D.M. New Species from the Fusarium solani Species Complex Derived from Perithecia and Soil in the Old World Tropics. Mycologia 2011, 103, 1302–1330. [Google Scholar] [CrossRef]
- Ellis, M.L.; Paul, P.A.; Dorrance, A.E.; Broders, K.D. Two New Species of Pythium, P. Schmitthenneri and P. Selbyi Pathogens of Corn and Soybean in Ohio. Mycologia 2012, 104, 477–487. [Google Scholar] [CrossRef]
- Liu, F.; Wang, J.; Li, H.; Wang, W.; Cai, L. Setophoma spp. on Camellia sinensis. Fungal Syst Evol 2019. [Google Scholar] [CrossRef] [PubMed]
- Alfenas, A.C.; Ferreira, F.A.; Mafia, R.G.; Gonçalves, R.C. Métodos Em Fitopatologia. In Métodos em Fitopatologia; Alfenas, A.C., Mafia, R.G., Eds.; UFV: Viçosa, 2016; pp. 55–91. [Google Scholar]
- Pinho, D.B.; Firmino, A.L.; Ferreira-Junior, W.G.; Pereira, O.L. An Efficient Protocol for DNA Extraction from Meliolales and the Description of Meliola Centellae Sp. nov. Mycotaxon 2013, 122, 333–345. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E. Two Divergent Intragenomic RDNA ITS2 Types within a Monophyletic Lineage of the Fungus Fusarium Are Nonorthologous. Mol Phylogenet Evol 1997, 7, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Vilgalys, R.; Hester, M. Rapid Genetic Identification and Mapping of Enzymatically Amplified Ribosomal DNA from Several Cryptococcus Species. J Bacteriol 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed]
- de Hoog, G.S.; van den Ende, A.H.G.G. Molecular Diagnostics of Clinical Strains of Filamentous Basidiomycetes. Mycoses 1998, 41, 183–189. [Google Scholar] [CrossRef]
- Jacobs, K.; Bergdahl, D.R.; Wingfield, M.J.; Halik, S.; Seifert, K.A.; Bright, D.E.; Wingfield, B.D. Leptographium wingfieldii Introduced into North America and Found Associated with Exotic Tomicus piniperda and Native Bark Beetles. Mycol Res 2004, 108, 411–418. [Google Scholar] [CrossRef]
- Berbee, M.L.; Pirseyedi, M.; Hubbard, S. Cochliobolus Phylogenetics and the Origin of Known, Highly Virulent Pathogens, Inferred from ITS and Glyceraldehyde-3-Phosphate Dehydrogenase Gene Sequences. Mycologia 1999, 91, 964–977. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic Relationships among Ascomycetes: Evidence from an RNA Polymerse II Subunit. Mol Biol Evol 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Sung, G.-H.; Sung, J.-M.; Hywel-Jones, N.L.; Spatafora, J.W. A Multi-Gene Phylogeny of Clavicipitaceae (Ascomycota, Fungi): Identification of Localized Incongruence Using a Combinational Bootstrap Approach. Mol Phylogenet Evol 2007, 44, 1204–1223. [Google Scholar] [CrossRef]
- Crous, P.W.; Lombard, L.; Sandoval-Denis, M.; Seifert, K.A.; Schroers, H.-J.; Chaverri, P.; Gené, J.; Guarro, J.; Hirooka, Y.; Bensch, K.; et al. Fusarium: More than a Node or a Foot-Shaped Basal Cell. Stud Mycol 2021, 98, 100116. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol Biol Evol 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Nylander, J.A.A. MrModeltest Version 2. Program Distributed by the Author. Evolutionary Biology Center, Uppsala University, Uppsala, 2004. [Google Scholar]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE); IEEE, November 2010; pp. 1–8. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian Phylogenetic Inference under Mixed Models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A. FigTree. 2018. Available at: http://tree.bio.ed.ac.uk/software/figtree/. [Accessed May 14, 2022].
- Crous, P.W.; Phillips, A.J.L.; Baxter, A.P. Phytopathogenic Fungi from South Africa; University of Stellenbosch, Department of Plant Pathology Press: 358, 2000. [Google Scholar]
- Wordell Filho, J.A.; Rowe, E.; Gonçalves, P.A.S.; Debarba, J.F.; Boff, P.; Thomazelli, L.F. Manejo Fitossanitário Na Cultura Da Cebola. In Doenças de origem parasitária; Wordell Filho, J.A., Boff, P., Eds.; Epagri: Florianópolis, 2006; p. 226. [Google Scholar]
- Babadoost, M. Onion Pink Root: Report on Plant Disease. 1990. University of Illinois Extension RPD No. 932. Available at: <https://ipm.illinois.edu/diseases/rpds/932.pdf> [Accessed: March 2, 2021].
- Ikeda, K.; Kuwabara, K.; Urushibara, T.; Soyai, P.; Miki, S.; Shibata, S. Pink Root Rot of Squash Caused by Setophoma Terrestris in Japan. Journal of General Plant Pathology 2012, 78, 372–375. [Google Scholar] [CrossRef]
- Levic, J.; Petrovic, T.; Stankovic, S.; Ivanovic, D. The Incidence of Pyrenochaeta terrestris in Root of Different Plant Species in Serbia. Zb Matice Srp Prir Nauk 2013, 21–30. [Google Scholar] [CrossRef]
- Punithalingram, E.; Holiday, P. CMI Descriptions of Pathogenic Fungi and Bacteria. No. 397. 1973. Available at: https://www.cabi.org/dfb/restricted?target=%2fdfb%2ffulltextpdf%2f2005%2f20056400397.pdf [Accessed: 01 March 2021].
- Yang, Y.; Zuzak, K.; Harding, M.; Neilson, E.; Feindel, D.; Feng, J. First Report of Pink Root Rot Caused by Setophoma (Pyrenochaeta) terrestris on Canola. Canadian Journal of Plant Pathology 2017, 39, 354–360. [Google Scholar] [CrossRef]
- López-López, M.; Léon-Félix, J.; Allende-Molar, R.; Lima, N.B.; Tovar-Pedraza, J.M.; García-Estrada, R.S. First Report of Setophoma terrestris Causing Corky and Pink Root of Tomato in Sinaloa, Mexico. Plant Dis 2020, 104, 1553–1553. [Google Scholar] [CrossRef]
- USDA Fungal Databases. 2024. Available at: https://fungi.ars.usda.gov/ [Accessed May 25th 2025].
- Nico, A.I.; Sánchez, M.G. Response of Different Intermediate-Day Onion Hybrids to Natural Infestation by Phoma terrestris and Fusarium oxysporum f. sp. cepae in Ciudad Real, Spain with Assessment of Different Soil Disinfestation Methods. Eur J Plant Pathol 2012, 134, 783–793. [Google Scholar] [CrossRef]
- Rivera-Méndez, W.; Brenes-Madriz, J.; Alvarado-Marchena, L. Effect of Setophoma terrestris, Sclerotium cepivorum, and Trichoderma spp. on in Vitro Onion (Allium cepa) Root Tissues and the Final Yield at the Field. Eur J Plant Pathol 2021, 160, 53–65. [Google Scholar] [CrossRef]
- Marin-Felix, Y.; Hernández-Restrepo, M.; Iturrieta-González, I.; García, D.; Gené, J.; Groenewald, J.Z.; Cai, L.; Chen, Q.; Quaedvlieg, W.; Schumacher, R.K.; et al. Genera of Phytopathogenic Fungi: GOPHY 3. Stud Mycol 2019, 94, 1–124. [Google Scholar] [CrossRef]
- Phookamsak, R.; Liu, J.-K.; Manamgoda, D.S.; Chukeatirote, E.; Mortimer, P.E.; Mckenzie, E.H.C.; Hyde, K.D. The Sexual State of Setophoma. Phytotaxa 2014, 176, 260. [Google Scholar] [CrossRef]
- Gao, C.; Montoya, L.; Xu, L.; Madera, M.; Hollingsworth, J.; Purdom, E.; Singan, V.; Vogel, J.; Hutmacher, R.B.; Dahlberg, J.A.; Coleman-Derr, D.; Lemaux, P.G.; Taylor, J.W. Fungal community assembly in drought-stressed sorghum show stochasticity, selection, and universal ecological dynamics. Nature Communications 2020, 11, 34. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Wu, B.; Zhang, S.; Li, M.; Jiang, X. Transcriptome dynamics underlying chlamydospore formation in Trichoderma virens GV29-8. Frontiers in Microbiology 2021, 12, 654855. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.-T.; Zhang, L.; Shen, H.-W.; Bao, D.-F.; Luo, Z.-L. Setophoma aseptata sp. nov. and new record of Minutisphaera aspera from Yuanjiang River Basin, China. Phytotaxa 2025, 702, 287–299. [Google Scholar] [CrossRef]
- Absalan, S.; Armand, A.; Jayawardena, R.S.; McKenzie, E.H.C.; Hyde, K.D.; Lumyong, S. Diversity of Pleosporalean Fungi Isolated from Rice (Oryza sativa L.) in Northern Thailand and Descriptions of Five New Species. J. Fungi 2024, 10, 763. [Google Scholar] [CrossRef]
- Li, M.; Liu, D.; Wang, M.; Cui, K.; Chen, L.; He, L.; Zhou, L. Characterization of Setophoma henanensis sp. nov., causing root rot on peanut. BMC Microbiol 2025, 25, 329. [Google Scholar] [CrossRef]
- Liu, H.F.; Choi, H.J.; Paul, N.C.; Ariyawansa, H.A.; Sang, H.K. 2025. Discovering fungal communities in roots of Zoysia japonica and characterising novel species and their antifungal activities. IMA Fungus 2025, 16, 1–43. [Google Scholar] [CrossRef]
- Ferreira, B.W.; Guterres, D. C.; Macedo, D. M.; Barreto, R. Epityfication of Perisporiopsis struthanthi and Perisporiopsis lantanae, and the taxonomic implications for Perisporiopsidaceae, Perisporiopsis and Setophoma. ResearchSquare 2023. [Google Scholar] [CrossRef]
- Sadeghi, S.; Nasr-Esfahani, M.; Maleki, M.; Molahoseini, H.; Khankahdani, H. H.; Mohammadi, M. Fungicidal control of onion pink root caused by Setophoma terrestris and effects on soil enzyme activity. Journal of Phytopathology 2024, 172, e13349. [Google Scholar] [CrossRef]
- Zhao, P.; Feng, Z.; Cai, L.; Phurbu, D.; Duan, W.; Xie, F.; Li, X.; Liu, F. Development of an RPA-CRISPR/Cas12a Assay for Rapid and Sensitive Diagnosis of Plant Quarantine Fungus Setophoma terrestris. J. Fungi 2024, 10, 716. [Google Scholar] [CrossRef]
- Li, X.; Liu, F.; Duan, W.J. TaqMan MGB-based real-time fluorescent PCR method for the rapid detection of Setophoma terrestris. Mycosystema 2024, 43, 240100. [Google Scholar]
- Sayago, P.; Juncosa, F.; Albarracín Orio, A.G.; Luna, D. F.; Molina, G.; Lafi, J.; Ducasse, D.A. Bacillus subtilis ALBA01 alleviates onion pink root by antagonizing the pathogen Setophoma terrestris and allowing physiological status maintenance. European Journal of Plant Pathology 2020, 157, 509–519. [Google Scholar] [CrossRef]
- Zhang, F.B.; Zheng, H.L.; Cui, W.G.; Zhang, M.Q.; Yin, Y.S.; Cui, M.; Gao, M. First report of Setophoma terrestris causing pink root of garlic in China. Plant Dis. 2019, 103, 584. [Google Scholar] [CrossRef]
- López-López, A. M.; Tovar-Pedraza, J. M.; León-Félix, J.; Allende-Molar, R.; Bernardi Lima, N.; Márquez-Zequera, I.; García-Estrada, R. S. Caracterización morfológica, filogenia y patogenicidad de Setophoma terrestris causante de raíz corchosa y rosada de jitomate (Solanum lycopersicum) en Sinaloa, México. Revista Mexicana de Fitopatología 2024, 42. [Google Scholar]
- Le, D.; Audenaert, K.; Haesaert, G. Fusarium basal rot: profile of an increasingly important disease in Allium spp. Tropical Plant Pathology 2021, 46, 241-253. Tropical Plant Pathology 2021, 46, 241–253. [Google Scholar] [CrossRef]
- Abd-El-Baky, A.A.; Yousef, H.; Shalaby, S.I. Garlic pink rot disease and crop yield as affected by salinity and irrigation water deficit. Egyptian Journal of Phytopathology 2019, 47, 141–155. [Google Scholar] [CrossRef]
- El-Elimat, T.; Figueroa, M.; Raja, H. A.; Graf, T. N.; Swanson, S. M.; Falkinham III, J. O.; Oberlies, N. H. Biosynthetically distinct cytotoxic polyketides from Setophoma terrestris. European Journal of Organic Chemistry 2015, 2015, 109–121. [Google Scholar] [CrossRef]
- Ferreira, J.F.; Bosland, P.W.; Williams, P.H. The variability of Pyrenochaeta terrestris isolates based on isozyme polymorphism, cultural characteristics and virulence on differential onion breeding lines. Journal of Phytopathology 1991, 133, 289–296. [Google Scholar] [CrossRef]
Figure 1.
Bayesian phylogenetic tree based on concatenated sequences (tub, tef, its, lsu, and gapdh). Bayesian posterior probability (PP) values are indicated at the nodes, and the scale bar represents the number of expected changes per site (0.01). Branches without values indicates PP ≤ 0.95. The specimen Didymella pinodella CBS 531.66 was used as outgroup. The isolates obtained in this study were highlighted in bold. (T =Type specimen).
Figure 1.
Bayesian phylogenetic tree based on concatenated sequences (tub, tef, its, lsu, and gapdh). Bayesian posterior probability (PP) values are indicated at the nodes, and the scale bar represents the number of expected changes per site (0.01). Branches without values indicates PP ≤ 0.95. The specimen Didymella pinodella CBS 531.66 was used as outgroup. The isolates obtained in this study were highlighted in bold. (T =Type specimen).

Figure 2.
Morphological characteristics of S. terrestris CCUB 2754. A: Culture grown on oatmeal-agar (left, above; right, reverse); B: Setose pycnidia. Bar = 0.1mm; C-D: Setose pycnidium and conidia. Bar = 10µm; E: conidia. Bar = 10µm; F: Conidiogenous cell. Bar = 10µm. G: Intercalar chlamydospores. Bar = 10µm; Setose pycnidium on garlic root. Bar = 0.1mm.
Figure 2.
Morphological characteristics of S. terrestris CCUB 2754. A: Culture grown on oatmeal-agar (left, above; right, reverse); B: Setose pycnidia. Bar = 0.1mm; C-D: Setose pycnidium and conidia. Bar = 10µm; E: conidia. Bar = 10µm; F: Conidiogenous cell. Bar = 10µm. G: Intercalar chlamydospores. Bar = 10µm; Setose pycnidium on garlic root. Bar = 0.1mm.
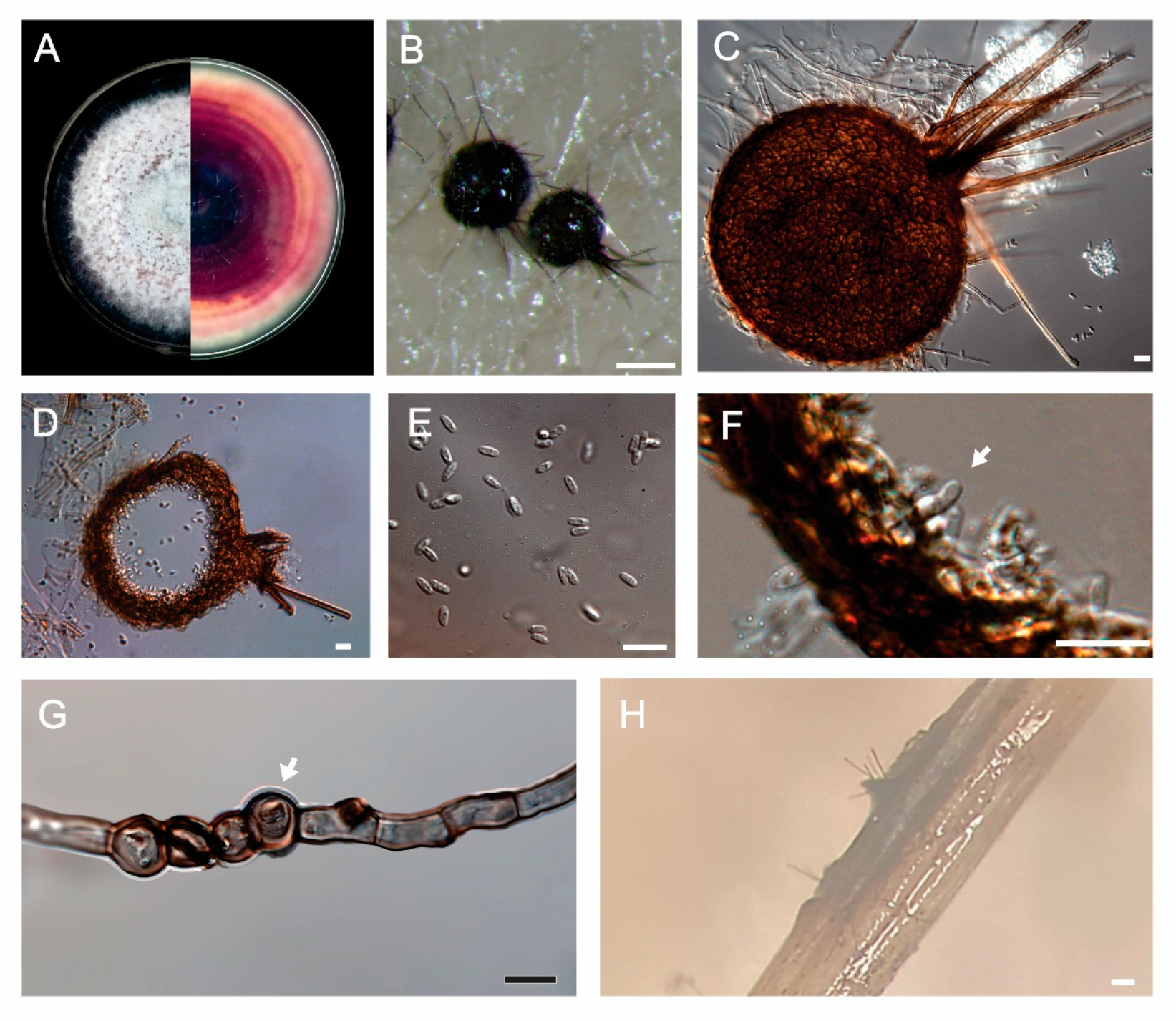
Figure 3.
Pink root symptoms under field and greenhouse conditions. (a) garlic and (b) onion showing symptoms in field; (c) garlic, (e) onion, (k) chives, (l)leeks and (n) brachiaria inoculated with S. terrestris isolate CCUB 2754 under greenhouse conditions; (d) garlic, (f) onion, (i) chives, (j) leeks and (m) brachiaria mock-inoculated controls; (g) garlic and (h) onion asyntomatic after inoculation with Fusarium sp.
Figure 3.
Pink root symptoms under field and greenhouse conditions. (a) garlic and (b) onion showing symptoms in field; (c) garlic, (e) onion, (k) chives, (l)leeks and (n) brachiaria inoculated with S. terrestris isolate CCUB 2754 under greenhouse conditions; (d) garlic, (f) onion, (i) chives, (j) leeks and (m) brachiaria mock-inoculated controls; (g) garlic and (h) onion asyntomatic after inoculation with Fusarium sp.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



