
Submitted:
26 September 2025
Posted:
29 September 2025
You are already at the latest version
Abstract
Lycium barbarum L. is an important economic crop in Qinghai province, China. Howev-er, root rot seriously reduces the economic results of L. barbarum. Here, we collected the diseased L. barbarum roots from Nuomuhong Farm of Haixi Mongolian and Tibetan Autonomous Prefecture, Qinghai Province, China, to clarify the diversity, pathogenic-ity, and biological characteristics of its root rot pathogens. A total of 125 isolates were collected, and based on morphological characteristics and rDNA-ITS, TEF-1α, and RPB2 gene sequence analysis, they were identified as Fusarium equiseti, F. avenaceum, F. solani, F. citri, F. acuminatum, F. culmorum, F. sambucinum, F. incarnatum, F. oxysporum, F. tricinctum, Microdochium bolleyi, and Clonostachys rosea. These fungi were used to inocu-late the roots of 1-year-old L. barbarum seedlings using scratching and root-irrigation inoculation methods, and all isolates caused root rot. The lethal temperatures were 50°C for F. acuminatum, F. sambucinum, and F. culmorum; 55°C for F. equiseti, C. rosea, M. bolleyi, F. avenaceum, F. citri, F. incarnatum, and F. tricinctum; and 65°C for F. solani, and F. oxysporum. The best media for the 12 pathogen species were potato dextrose agar, mung bean soup culture, and carnation leaf piece agar. The optimum carbon sources were sucrose, dextrin, and soluble starch, and the optimum nitrogen sources were beef extract, peptone, and yeast extract. This is the first report that M. bolleyi, F. avenaceum, and F. citri cause root rot in L. barbarum. Moreover, our findings provide a theoretical foundation for root rot management in the future.
Keywords:
Gouqi
; root rot
; Fusarium fungi
; diversity
; pathogenicity
; biological characteristics
1. Introduction
“Gouqi” is the common name for Lycium barbarum L., which belongs to the Solanaceae family is a medicinal plant with a long history of use in traditional Chinese medicine [1]. The Lycium genus comprises approximately 70 species that vegetate in separate and distinct regions distributed from the temperate to the subtropical regions of Eurasia, North America, South America, southern Africa, and Australia [2]. In China, there are seven species and two varieties of the genus Lycium, four of which have been used by different ethnic groups. Only L. barbarum and L. chinense have been transformed into globally traded commodities. Lycium barbarum berries are red or orange-yellow and oblong or ovoid [3], and the morphological characteristics of L. chinense are similar to those of L. barbarum. The term Chinese-cultivated Gougi generally refers to L. barbarum, which has a long history of traditional use. In Northern and Central China, it has been used in traditional medicine for 2000 years to promote longevity, vision, and wellness. Since the Tang Dynasty, L. barbarum has been noted for its traditional health benefits, such as nourishing the yin, strengthening the liver and kidney, and sustaining the blood [4,5]. Modern studies have found that L. barbarum also has anti-aging, immuno-modulation, anti-cancer, and anti-fatigue. Traditional Chinese medicine makes use of both the root bark (Digupi) and fruit (Gouqizi), moreover, fruit is well known in traditional Chinese herbal medicine and has been widely used as a popular functional food [6]. The utilization of its leaves and seeds has also been mentioned in a few medicinal books [7]. The nutritional effects of L. barbarum are mainly due to its rich content of compounds such as polysaccharides, flavonoid polyphenols, carotenoids, and alkaloids [8,9]. Therefore, the demand for Gouqi and its price are increasing in China.
Fusarium species, which are part of the Nectriaceae family in the Hypocreales order within the Ascomycota phylum, are widely distributed across environments such as air, water, soil, plants, insects, and organic substrates. This genus encompasses over 200 species grouped into 22 species complexes [10]. The name Fusarium was coined by Fries in 1821, and its species are recognized as one of the most important pathogens in agriculture and horticulture, infecting a variety of crops [11]. In addition, this genus of pathogenic fungi is known in the food industry [12]. Fusarium species are major soil-borne pathogens. In China, they are common in the northwest, southwest, central, and southeast regions. They cause economic losses by limiting crop growth and yield and pose a threat to food safety and human health by producing fungal toxins, such as trichothecene, fumonisins, and zearalenone [13,14,15,16]. Of the top 10 pathogenic fungi in molecular plant pathology in 2012, F. graminearum and F. oxysporum ranked fourth and fifth, respectively, indicating that Fusarium fungi have a strong destructive effect as plant pathogens [17]. Fusarium species cause many diseases. For example, F. graminearum and F. verticillioides cause ear rot in Zea mays; Fusarium solani causes collar rot in Pisum sativum [18]. Fusarium oxysporum causes Fusarium wilt in banana, having a serious impact on banana production worldwide [19]. Fusarium graminearum causes Fusarium head blight on Triticum aestivum [20]. Fusarium fujikuroi causes rice bakanae disease [21], and F. graminearum causes Z. mays stalk rot [22]. Moreover, Fusarium species interact with other plant pathogens, and mycotoxin accumulation plays an important role in disease outbreaks [23].
Microdochium bolleyi typically resides endophytically in plant roots, especially in herbaceous plants, and is abundant in the roots of coastal herbaceous plant species along the Pacific Northwest coast of the United States [24]. Microdochium bolleyi primarily colonizing the roots of cereals and grasses, including T. aestivum and Hordeum vulgare, suppresses some plant pathogens affecting cereals. The formation of typical chlamydospore clusters, which are visible under light microscopy, in colonized plant cells makes this endophytic fungus an excellent model organism for studying plant–endophyte interactions [25]. However, some studies have shown that M. bolleyi causes crown and root rot in grasses, such as T. aestivum and H. vulgare [26], and is associated with basal rot of Agrostis stolonifera [27] and root rot in Linum usitatissimum [28]. Therefore, M. bolleyi is generally considered a commensal or weak pathogen [24].
Clonostachys rosea is a filamentous fungus with a widespread distribution across a range of habitats [29]. Clonostachys rosea is a potent biocontrol agent against F. oxysporum, using mycoparasitism and its bioactive metabolites [30], and reduces the severity of diseases caused by soil- and seed-borne pathogens [31]. However, C. rosea can also cause root rot and was first reported on Glycine max in the United States [32]. It can also cause stem end rot of Actinidia chinensis [32] and root rot of naked H. vulgare var. nudum, Xanthoceras sorbifolium [34], G. max [35], Allium sativum [36], and Astragalus membranaceus [37].
Root rot is one of the most common diseases affecting L. barbarum, seriously affecting its quality and posing a huge threat to local economic development. In the early stages of the disease, the roots swell, intensifying in the middle stage with a small amount of defoliation. In the later stage, the roots rot and show reddish brown and the plants shed many leaves until they wither and die. In the main L. barbarum cultivation areas, the root rot incidence can reach 50%, resulting in serious yield and income reduction, hindering the development of the L. barbarum industry [38]. Fusarium species have been reported as the main pathogens causing root rot in L. barbarum [39]. Our investigation into root rot of L. barbarum in Qinghai Province from 2022 to 2023 revealed that root rot caused by Fusarium commonly occurred. The objectives of this study were to determine the phylogenetic diversity, morphological characteristics, and pathogenicity of isolates to L. barbarum and to clarify their biological characteristics to provide a foundation for comprehensive disease control.
2. Materials and Methods
2.1. Collection Site
Gouqi root rot samples were collected from Nuomuhong Farm (96° 14′ 59.95″–96° 30′ 37.46″ E, 36° 24′ 20.21″–36° 26′ 45.66″ N), Dulan County, Haixi Mongolian and Tibetan Autonomous Prefecture, Qinghai Province, China, at an altitude of 2800 m. The area covers 130,000 acres, with a low annual rainfall of 43.5 mm, strong solar radiation, and more than 3100 h of sunshine per year. The average annual temperature is 4.9°C, with distinct differences between day and night, and the annual maximum and minimum temperatures are 35.8 and −31°C, respectively. The frost-free period is 70 days [40]. The soil type is gray-brown desert soil and sandy soil, which is highly suitable for Gouqi growth. On the farm, the Gouqi planting area is approximately 80,000 acres, which is the largest Gouqi planting area in Qinghai Province. In this study, 36 samples showing root rot were collected in July and August 2022 and 2023 from 12 sites on the farm.
2.2. Media Used in the Study
Potato dextrose agar (PDA) [24] (200 g of potato, 20 g of glucose, 18 g of agar, and distilled water to a volume of 1000 mL) was used to preserve, activate, and observe the morphological characteristics of fungi. For isolations, 2% water agar (2% WA) medium (20 g of agar and distilled water to a volume of 1000 mL) was used. The following media were used to evaluate the biological characteristics: carrot agar (CA) medium (200 g of carrot, 20 g of glucose, 20 g of agar, and distilled water to a volume of 1000 mL), carnation leaf-piece agar (CLA) medium (14.4 g of agar, distilled water to a volume of 1000 mL, and carnation leaves), complete (CM) medium (6 g of yeast extract, 3 g of enzymatic casein, 3 g of acid-hydrolyzed casein, 10 g of sucrose, 900 μL of dd H2O, and distilled water to a volume of 1000 mL), mung bean soup culture (MB) medium (20 g of mung beans and distilled water to a volume of 1000 mL), and Czapek (Czapek–Dox) medium (3 g of sodium nitrate, 1 g of dipotassium hydrogen phosphate, 0.5 g of magnesium sulfate, 0.5 g of potassium chloride, 0.01 g of ferrous sulfate, 30 g of sucrose, 20 g of agar, and distilled water to 1000 mL).
2.3. Isolation, Purification, and Preservation of Pathogenic Fungi
To isolate the fungi, root pieces (approximately 0.5 cm) were obtained with sterile scissors from the junction of infected and healthy tissues. They were surface-disinfected with 3% NaClO for 1.5 min and washed 3 times with sterile water. The disinfected pieces were dried and placed on 2% WA plates in an incubator at 25°C for 2 days. After the root surface became moldy, abundant conidiophores and conidia formed. Conidia were placed into sterile water with a pipette to prepare a spore suspension. Then, 200 μL of the spore suspension was spread on the surface of a 2% WA plate and placed in an incubator at 25°C for 12 h. Single spores were selected under a stereomicroscope and cultured on PDA. A total of 125 isolates were collected from rotted roots. Fungal isolates were cultured on PDA at 25°C, maintained on filter paper, and stored at −20°C for long-term conservation [41].
2.4. Pathogen Identification
2.4.1. Morphological Identification
Colony and conidia morphology were observed, and isolates were identified according to the Fusarium Laboratory Manual [42]. Representative single spore isolates were used to inoculate a PDA plate and cultured for 5 days, and a mycelial block with a diameter of 5 mm was placed on a PDA plate. After 7 days of culture in an incubator at 25°C, the diameter was measured using the cross method. Each isolate was tested three times. Conidia were used to make temporary slides, and conidial morphology was observed under a 40× optical microscope (OLYMPUS IX71, Olympus, Tokyo, Japan). To characterize conidial morphology, the length and width of at least 30 conidia per isolate were observed under a light microscope and measured with Image-Pro Plus version 6.0.0 software (Media Cybernetics, Silver Spring, MD, USA). The length of a conidium from apex to base was defined as the conidial length, and the widest part of each conidium was defined as the conidial width.
2.4.2. DNA Extraction, PCR, and Sequencing
Genomic DNA was extracted from vegetative mycelia using the cetyltrimethylammonium bromide protocol [43]. DNA amplification was performed for the DNA sequences of the nuclear ribosomal internal transcribed spacer (rDNA-ITS), the translation elongation factor 1-alpha (TEF-1α), and the RNA polymerase II gene (RPB2). The PCRs were performed in 25-μL mixtures with 1 μL of genomic DNA, 12.5 μL of 2× Taq PCR Master Mix (Biomed, Beijing, China), and 10 μM of each primer. The following PCR primers were used: rDNA-ITS amplification primers, ITS1 (5’-TCCGTAGGTGAACCTGCGG-3’) and ITS4 (5’-TCCTCCGCTTATTGATATGC-3’) [44,45]; TEF-1α gene amplification primers, EF-1 (5’-ATGGGTAAGGARGACAAGAC-3’), EF-2 (5’-GGARGTACCAGTSATCATGTT-3’) [46,47]; and RPB2 gene amplification primers, RPB2-5F (5’-GAYGAYMGWGATCAYTTYGG-3’), RPB2-7CR (5’-CCCATRGCTTGYTTRCCCAT-3’) [48]. DNA sequencing was performed at Tsingke Biological Technology Company (Beijing, China).
2.4.3. Phylogenetic Analysis
The phylogenetic tree was constructed with reference to Swofford [49]. DNA sequences were aligned with an online version of MAFFT (https://mafft.cbrc.jp/alignment/software/), and the alignment was manually adjusted using MEGA version 6.0.6 (Tokyo Metropolitan University, Tokyo, Japan). Maximum parsimony trees were constructed using PAUP* version 4.0 alpha for all datasets, and they were generated by heuristic searches with Alternaria alternata (GenBank accession number CBS 130263), as a reference taxon and 1000 bootstrap replicates. The sequences of the 21 reference strains were downloaded from the NCBI database. The NCBI accession numbers of 125 pathogens were PV592419–PV592543 for the ITS sequences, PV567778–PV567888 for TEF-1α, and PV595151–PV595275 for RPB2.
2.5. Pathogenicity Assays
All isolates were cultured on PDA plates at 25°C for 5 days, and a pathogenicity test was performed by inoculating the roots of 1-year-old L. barbarum seedlings with conidial suspensions at a concentration of 1 × 106 per mL in 0.025% Tween 20. Two inoculation methods, scratching and root-irrigation inoculation, were used to determine the pathogenicity of isolates to L. barbarum. For the scratching inoculation method, the root epidermis was evenly pierced with a sterile inoculation needle, and a 10 μL conidial suspension was used to inoculate each wound. Inoculated roots were incubated in a humid and dark chamber at 25°C for 14 days. For the root-irrigation inoculation method, the L. barbarum root epidermis was pierced, and a 15 mL spore suspension was poured into each pot to irrigate the roots. Inoculated plants were incubated in a humid and dark chamber at 25°C for 24 h and moved to a greenhouse with a 12-h light/12-h dark photoperiod for 30 days. Sterile water was used as a control. The pathogenicity test was repeated three times for each isolate.
2.6. Biological Characteristics of Pathogens
2.6.1. Effects of Different Media
PDA, CA, CLA, CM, and CB media 50 were used to investigate their effects on mycelial growth. A 5-mm diameter mycelial plug was transferred to the center of each medium and incubated at 25°C in an incubator for 7 days. Each isolate was repeated three times.
2.6.2. Effects of Temperature
To determine the effect of temperature on mycelial growth, 5-mm diameter mycelial plugs of the isolates were incubated at different temperatures (35, 40, 50, 55, 60, 65, and 70°C) for 10 min and incubated at 25°C in an incubator for 7 days. Each isolate was repeated three times.
2.6.3. Effects of pH
To determine the effect of pH on mycelial growth, PDA media were adjusted with 0.1 M HCl and 0.1 M NaOH to obtain pH values of 4.0, 5.0, 6.0, 7.0, 8.0, 9.0, 10.0, and 11.0 [50,51]. A 5-mm diameter plug was placed in the center of a PDA plate and incubated at 25°C in an incubator for 7 days. Each isolate was repeated three times.
2.6.4. Effects of Carbon and Nitrogen
Czapek–Dox medium was used as the basic medium to determine the utilization of nitrogen and carbon sources [50]. In the media, 20 g of sucrose was replaced with 20 g of soluble starch, maltose, glucose, dextrin, cellulose, mannitol, lactose, fructose, and xylose to determine the effects of different carbon sources on growth. Furthermore, 4 g of sodium nitrate was replaced with 4 g of beef extract, peptone, yeast extract, urea, potassium nitrate, ammonium nitrate, ammonium chloride, and sodium nitrate to determine the effects of different nitrogen sources on growth. A 5-mm diameter mycelial plug was transferred to the center of each carbon source medium and nitrogen source medium, and plates were incubated at 25°C in an incubator for 7 days. Each isolate was repeated three times.
2.7. Data Processing
Microsoft Excel worksheet was used for data integration, IBM Statistics SPSS 20.0 software was used for one-way ANOVA, and Duncan's multiple range test was used to test the significance of differences.
3. Results
3.1. Field Symptoms of Gouqi Root Rot and the Diversity of Pathogenic Fungi
The incidence of Gouqi root rot at Nuomuhong Farm, Dulan County, Haixi Mongolian and Tibetan Autonomous Prefecture, Qinghai Province, China, was serious, with an incidence rate of 10–50%. The leaves on the ground were yellow, and some of the Gouqi leaves fell off (Figure 1B). The epidermis near the roots fell off, and the branches and main stems turned white and dry. The main roots of the underground part were short, and the number of roots was reduced. The fibrous roots were short and thin, and main and fibrous roots were rotted (Figure 1C).
A total of 125 pathogenic fungi were isolated from 36 root samples of cultivated Gouqi and identified using morphological and molecular biology techniques. There were 34 F. equiseti, 22 C. rosea, 14 M. bolleyi, 12 F. solani, 11 F. acuminatum, 8 F. sambucinum, 6 F. avenaceum, 6 F. citri, 5 F. incarnatum, 3 F. culmorum, 2 F. oxysporum, and 2 F. tricinctum isolates, with separation frequencies of 27.2, 17.6, 11.2, 9.6, 8.8, 6.4, 4.8, 4.8, 4.0, 2.4, 1.6, and 1.6%, respectively. Among them, F. equiseti, C. rosea, and M. bolleyi were the dominant pathogens, and F. avenaceum, F. citri, F. incarnatum, F. culmorum, F. oxysporum, and F. tricinctum were rare.
3.2. Morphology and Phylogenetic Analysis of Root Rot Pathogens
The morphological characteristics of the 125 pathogens were observed (Figure 2; Table 1). Fusarium equiseti colonized the whole plate in 7 d and produced pale yellow colony. Microconidia were sparse and measured 11.56 ± 1.46 × 3.85 ± 0.56 μm. Macroconidia were slightly curved with 1–5 septa and were 26.44 ± 2.14 × 5.69 ± 1.43 μm in size. Its chlamydospores were 16.34 ± 1.58 × 14.15 ± 2.08 μm in size.
Clonostachys rosea formed a white mold, the hypha was colorless with septa, and conidiophores were broom-shaped. Conidia colorless, round or oval, no septum, 7.04 ± 0.41 × 4.36 ± 0.37 μm in size, and chlamydospores were 11.38 ± 2.25 × 12.45 ± 2.02 μm in size.
Microdochium bolleyi produced few aerial mycelia, and the middle of the colony was orange. It produced a large number of conidia that were colorless and slightly curved. Conidia were produced laterally on the conidiophore, about 9.55 ± 2.04 × 3.39 ± 0.78 μm in size, and chlamydospores were 11.28 ± 1.46 × 8.00 ± 0.59 μm in size.
The aerial mycelium of F. solani had weak growth, producing less mycelia. The mycelia were white in the early stage and produced red, purple, or blue pigments in the later growth stage. It produced many macroconidia and microconidia. Macroconidia were colorless, with 1–4 septa, and were 39.09 ± 4.63 × 7.07 ± 1.21 μm in size. Microconidia were ovate to round and single-spored, with a size of 7.63 ± 0.91 × 4.7 ± 0.68 μm. Chlamydospores were round or oval, with a size of 15.74 ± 2.02 × 14.19 ± 1.90 μm.
Fusarium acuminatum produced abundant, dense aerial hypha that was rose red and had a slow growth rate. Macroconidia were colorless and falciform, had 1–3 septa, and were 20.16 ± 3.02 × 4.79 ± 0.88 μm in size. Microconidia were hyaline, had 0–1 septa, and were 8.82 ± 1.65 × 4.32 ± 0.83 μm in size. Chlamydospores were 13.07 ± 2.15 × 12.84 ± 1.23 μm in size.
The mycelia of F. sambucinum were loose, covered the dish in 7 days, and was light orange near the center. Macroconidia were fascicled, had 1–4 septa, and were 34.75 ± 5.38 × 7.56 ± 1.20 μm in size. Microconidia were colorless, had 0–1 septa, and were 7.72 ± 1.36 × 4.22 ± 0.92 μm in size. Chlamydospores were 13.12 ± 3.80 × 9.45 ± 2.15 μm in size.
The aerial hyphae of F. avenaceum were villous and pale red, and macroconidia were thin at both ends and wide in the middle, 46.45 ± 4.96 × 5.74 ± 1.54 μm in size, and had 1–7 septa. Conidiogenous cells arose laterally, and were phialidic and slender. Chlamydospores were not found.
Fusarium citri grew abundant, dense pale-yellow mycelia. Its macroconidia were colorless and laterally on conidiophores, with 1–5 septa. The tip of macroconidia was curved, and they were 30.84 ± 5.41 × 5.72 ± 1.02 μm in size. Microconidia were rare. Chlamydospores were round or oval and were 11.32 ± 2.10 × 10.57 ± 2.93 μm in size.
Fusarium incarnatum grew fast and had dense hyphae, and the surface was dark yellow. Macroconidia were colorless and 30.02 ± 1.51 × 6.17 ± 1.07 μm in size and had 1–5 septa. Microconidia were rare. Chlamydospores were 13.06 ± 3.48 × 16.73 ± 2.54 μm in size.
The aerial hypha of F. culmorum was sparse, and the colony was red. The top and bottom sides of the colony were initially dark red in the middle, surrounded by light red, and the dish was covered after 7 days. Macroconidia were pale yellow or colorless, with 1–4 septa, and were 37.99 ± 1.96 × 8.97 ± 0.46 μm in size. Microconidia were colorless, ovoid, and 8.24 ± 1.56 × 4.43 ± 1.14 μm in size and had 0–2 septa. Chlamydospores were round or ovoid and 13.77 ± 2.71 × 11.91 ± 1.79 μm in size.
The aerial hyphae of F. oxysporum were panniform and grew fast. The colony was light purple in the middle. Microconidia were rich, colorless, 7.22 ± 1.12 × 3.26 ± 0.70 μm in size, and had 0–1 septum. Macroconidia were colorless, 33.68 ± 2.21 × 11.45 ± 0.42 μm in size, and had 1–4 septa. Chlamydospores were 13.91 ± 1.36 × 12.77 ± 1.68 μm in size.
Fusarium tricinctum had white flocculent mycelia that covered the dish in 7 d. Its macroconidia were colorless, falciform, and slightly curved, with 1–5 septa, and were 33.97 ± 4.95 × 7.17 ± 1.34 μm in size. Microconidia were limoniform, oval, or pyriform, with 0–1 septa, and were 13.20 ± 2.16 × 4.52 ± 1.44 μm in size. Chlamydospores were 14.76 ± 6.45 × 13.56 ± 2.25 μm in size.
Values are presented as the mean ± standard deviatioThe single most parsimonious trees were generated from the combined sequences of rDNA-ITS and RPB2 genes (Figure 3), as the TEF-1α gene of M. bolleyi was not amplified using the primers used in this study, and individual rDNA-ITS, TEF-1α, and RPB2 genes from 125 isolates (Figures S1–S3). A total of 22 strains published in the NCBI database were used as reference strains (gray characters in Figures 3 and S1–S3). Overall, the trees in each phylogeny had the same topology, and 11 or 12 clades with high bootstrap values were resolved in the phylogenies inferred for F. equiseti, C. rosea, M. Bolleyi, F. solani, F. acuminatum, F. sambucinum, F. avenaceum, F. citri, F. incarnatum, F. culmorum, F. oxysporum, and F. tricinctum. In total, 22 isolates and 2 control strains (SJ123 and SJ62) clustered in the C. rosea clade, and 11 isolates and 2 control strains (NRRL5 4212 and NRRL 54214) clustered in the F. acuminatum clade. P612A, P415, P612B, and P111 clustered in the F. culmorum clade; N131 and GQ421 in the F. oxysporum clade; and GQ42 with control isolates CBS 110262, CBS133023, and CBS 25350 in the F. tricinctum clade. Five isolates and two control strains (CBS 132194 and CBS 133024) clustered in the F. incarnatum clade, and six isolates and two control strains (CBS 12173 and NRRL 54939) clustered in the F. avenaceum clade. Five isolates and two control strains (CBS 130905 and NRRL 25084) clustered in the F. citri clade, and nine isolates and two control strains (CBS 14695 and QJ 20911) clustered in the F. sambucinum clade. Thirty-four isolates and two control strain (CBS 18534 and CBS 30794) clustered in the F. equiseti clade. Twelve isolates and two control strains (CBS 102429 and CBS 131775) clustered in the F. solani clade. Fourteen isolates and two control strains (CBS 54092 and KAS 1350) clustered in the M. bolleyi clade.
3.3. Pathogenicity Analysis
The pathogenicity test was conducted using scratching and root-irrigation inoculation methods. The results of scratching inoculation showed that all isolates caused root rot in L. barbarum (Figure 4). Notably, distinct black lesions were observed after inoculation with F. solani, F. avenaceum, and F. citri. In contrast, the control group displayed lighter coloration with no symptoms. Robust mycelial growth was visible, with discoloration and decay around the wound, following inoculation with F. tricinctum, F. oxysporum, and C. rosea. Inoculation with F. oxysporum, F. equiseti, and F. sambucinum resulted in significant root decay and a color change at the inoculation site, confirming the disease. Inoculation with M. bolleyi and F. culmorum led to color changes and rot at the inoculation site.
The results of root-irrigation inoculation showed that all isolates could cause root rot. Thirty days after inoculation, the plants exhibited stunted growth and wilting and drying symptoms (Figure 5A and B). Root rot was evident, with internal discoloration visible upon cutting, and black lesions appeared centrally. In contrast, the blank control roots showed no decay or discoloration, maintaining a milky white appearance. Inoculation with F. solani, F. oxysporum, F. sambucinum, and F. avenaceum resulted in pronounced internal root discoloration, with the blackened areas larger than those observed after inoculation with other pathogens (Figure 5C and D). Pathogens were re-isolated from the diseased roots, and their morphology matched that of the field pathogens (Figure 2A and B).
3.4. Biological Characteristics of Root Rot Pathogens
3.4.1. Effect of Different Media on Mycelial Growth
The optimum culture medium for the 12 pathogen species was determined using PDA, CM, MB, CLA, and CA media. The results showed that the best media were PDA, MB, and CLA. The optimum culture medium for F. solani, F. sambucinum, F. avenaceum, F. citri, F. incarnatum, F. culmorum, and F. oxysporum was PDA, with growth rates ranging from 0.78 to 0.80 mm/d. The optimum culture medium for F. equiseti, C. rosea, F. culmorum, F. oxysporum, and F. citri was MB, with growth rates ranging from 0.61 to 0.80 mm/d. The optimum culture medium for F. culmorum, F. oxysporum, F. tricinctum, M. bolleyi, F. acuminatum, and F. citri was CLA, with growth rates ranging from 0.69 to 0.80 mm/d. Fusarium equiseti, C. rosea, and Fusarium acuminatum had the worst growth on PDA, with growth rates of 0.68 ± 0.06, 0.44 ± 0.04, and 0.52±0.01 mm/d, respectively. Microdochium bolleyi showed the worst growth on CA, with growth rates of 0.28 ± 0.02. MB resulted in the worst growth for F. avenaceum, with growth rates of 0.56±0.06 mm/d. (Table 2, Figure S4).
3.4.2. Effect of Temperature on Mycelial Growth
The lethal temperatures of the 12 pathogen species were between 50 and 65°C. The lethal temperatures were 50°C for F. acuminatum, F. sambucinum, and F. culmorum; 55°C for F. equiseti, C. rosea, M. bolleyi, F. avenaceum, F. citri, F. incarnatum, and F. tricinctum; and 65°C for F. solani, and F. oxysporum (Table 3, Figure S5).
3.4.3. Effect of pH on Mycelial Growth
The optimum pH of the 12 pathogen species ranged from 4 to 11. The optimum pH of C. rosea, F. citri, F. oxysporum, F. sambucinum, F. equiseti, and F. tricinctum was 4–7, with growth rates ranging from 0.52 ± 0.02 to 0.80 ± 0.01 mm/d. The optimum pH of F. incarnatum, F. culmorum, C. rosea, F. solani, F. citri, F. oxysporum, F. sambucinum, F. equiseti, F. tricinctum, and F. avenaceum was 7–11, with growth rates from 0.53 ± 0.03 to 0.80 ± 0.01 mm/d. The optimum pH of M. bolleyi was 9, showing growth rates of 0.59 ± 0.02 mm/d. The optimum pH of F. acuminatum was 8, with a growth rate of 0.73 ± 0.04 mm/d (Table 4, Figure S6).
3.4.4. Effect of Carbon and Nitrogen Sources on Mycelial Growth
The optimum carbon sources for the 12 pathogen species were screened using 10 carbon sources. The optimum carbon source for F. equiseti, F. citri, F. incarnatum, F. culmorum, and F. oxysporum was sucrose, with growth rates of all were 0.80 mm/d. The optimum carbon source for C. rosea was fructose, with growth rates of 0.57 ± 0.03 mm/d. Dextrin was the optimum carbon source for F. incarnatum, F. culmorum, F. oxysporum, and F. tricinctum with growth rates of all were 0.80 mm/d. The optimum carbon source for F. acuminatum was cellulose, showing a growth rate of 0.72 ± 0.03 mm/d. Soluble starch was the optimum carbon source for F. avenaceum, F. incarnatum, F. culmorum, and F. oxysporum, resulting in growth rates of 0.76 ± 0.0, 0.80±0.01, 0.80±0.01, and 0.80±0.01 mm/d, respectively. The growth rates of CK for F. equiseti, F. incarnatum, F. culmorum, and F. oxysporum were also fast, but the mycelia were only a thin layer and grew poorly. Glucose, lactose, cellulose, and xylose were not good carbon sources for any of the 12 pathogen species. For example, lactose as the carbon source resulted in the worst growth for F. equiseti, F. acuminatum, F. citri and F. tricinctum, with growth rates ranging from 0.39 ± 0.03 to 0.65 ± 0.03 mm/d (Table 5, Figure S7).
The optimum nitrogen sources for the 12 pathogen species were determined using 8 nitrogen sources, and we found that beef extract, peptone, and yeast powder resulted in optimum growth. The optimum nitrogen source for F. oxysporum, F. tricinctum, F. equiseti, M. bolleyi, F. sambucinum, F. incarnatum, and F. culmorum was beef extract, showing growth rates of 0.80 mm/d. Peptone was the optimum nitrogen source for F. equiseti, C. rosea, M. bolleyi, F. incarnatum, F. culmorum, F. oxysporum, F. tricinctum. Yeast powder was the optimum nitrogen source for F. equiseti, M. bolleyi, F. sambucinum, F. avenaceum, F. citri, F. incarnatum, F. culmorum, F. oxysporum, F. tricinctum, and with growth rates ranging from 0.78 to 0.80 mm/d. Saltpeter was the optimum nitrogen source for F. equiseti, and C. rosea, with a growth rate of 0.80±0.01, and 0.51 ± 0.06 mm/d, respectively. The optimum nitrogen source for F. solani was sodium nitrate, with a growth rate of 0.79 ± 0.01 mm/d. The growth rates of CK for F. equiseti, F. acuminatum, F. culmorum, and F. oxysporum were fast, however, the mycelia were sparse and only a thin layer. The worst growth was observed on the media with ammonium chloride, and ammonium nitrate, respectively, with growth rates ranging from 0.28 ± 0.04 to 0.79 ± 0.00 mm/d (Table 6, Figure S8).
4. Discussion
Root rot is one of the main reasons for the decrease in L. barbarum yield, and Fusarium species are the main causative pathogens [38]. The root rot pathogens of L. barbarum mainly damage its rhizome, and diseased plants exhibit symptoms including root rot, leaf etiolation, wilting, and stunted growth, resulting in reduced water and nutrient transport and sometimes plant death [52]. The yield and quality of Gouqi berry decrease, causing serious economic losses. Nuomuhong Farm in Qinghai Province is one of the main L. barbarum–producing areas. The large-scale cultivation of L. barbarum has brought huge economic benefits. However, root rot is becoming increasingly serious in L. barbarum. To effectively control root rot in L. barbarum, it is necessary to identify the pathogens causing this disease.
Fusarium species are the most widely distributed fungi, and they are connected to the environment, plants, and insects [10]. Compound infection and toxin secretion by Fusarium cause many diseases, reducing yield and quality and economic benefits. In this study, 36 root samples were collected from 12 locations on Nuomuhong Farm, Qinghai Province, China, and 125 isolates were confirmed as pathogens based on Koch’s postulates. The pathogenic isolates showed differences in colony color and morphological characteristics of conidia. Molecular biology analyses revealed that these pathogens belonged to 12 species. Among the isolated pathogens, F. equiseti, C. rosea, and M. bolleyi were dominant. Fusarium avenaceum, F. citri, F. incarnatum, F. culmorum, F. oxysporum, and F. tricinctum were isolated less frequently. Additionally, F. sambucinum, F. acuminatum, and F. solani were common species. To our knowledge, this is the first report of M. bolleyi, F. avenaceum, and F. citri as pathogens causing root rot in L. barbarum in China.
Uwaremwe et al. confirmed the presence of Fusarium species, including F. oxysporum, F. solani, F. tricinctum, and F. chlamydosporum, and A. alternata, which are responsible for root rot disease, in Gouqi plants in Gansu and Ningxia provinces, China [53]. Bai et al. first reported root rot caused by F. culmorum and F. equiseti on L. barbarum in Qinghai Province, China [52]. Feng et al. found that fungi isolated from Gouqi roots in the Qaidam Basin, China, were mainly Fusarium species, including F. lichenicola, F. oxysporum, F. redolens, and F. solani; Plectosphaerella plurivora; and Ple. cucumerina. Mortierella alpina, Mucor hiemalis, Penicillium janthinellum, Verticillium nonalfalfae, C. rosea, and Pen. simplicissimum were also identified [54]. Compared with these studies, in Qinghai, the composition of pathogenic fungi causing root rot on Gouqi differed. In this study, we found 12 species of pathogenic fungi, but F. chlamydosporum, A. alternata, Plectosphaerella plurivora, Mortierella alpina, and P. cucumerina were not found. We also identified F. oxysporum, F. solani, F. tricinctum, F. culmorum, F. equiseti, and C. rosea. This could have occurred because of regional differences and different planting years.
Pathogenic fungi infect the roots and rhizomes of L. barbarum, and root rot occurs from the seedling stage to the mature plant stage. In this study, the pathogenicity of 125 isolates was tested, and they could infect 1-year-old L. barbarum seedlings, as shown using scratching and root-irrigation inoculation techniques. Fusarium also affects other medicinal plants, such as Angelica [55] and A. membranaceus var. mongholicus [56], and leads to root rot and wilt in several important economic crops, including Cucumis melo [57], C. sativus [58] and Solanum lycopersicum [59]. Other plants, such as Codonopsis pilosula and Zea mays, also suffer from root and stem rot due to this pathogen. Furthermore, Fusarium causes needle blight in coniferous trees, such as Pinus thunbergii [60], and damping-off disease in P. pinea [61]. It also secretes toxins that contaminate fruit, reducing their shelf life. Common toxins produced by Fusarium include trichothecenes, zearalenones, fumonisins, beauvericin, and enniatins, all of which pose health risks to plants, livestock, and humans [62,63]. Efforts to control Fusarium in China primarily focus on selecting biocontrol microorganisms, including Metarhizium robertsii [64], fluorescent Pseudomonas [65], and Trichoderma harzianum [66], which have shown a control effect against root rot.
Morphological characteristics of pathogenic fungi from different hosts or different ecological regions vary to some extent. Research on the biological characteristics of plant pathogens lays the foundation for determining the occurrence patterns of diseases and formulating prevention and control measures. Therefore, the biological characteristics, including the optimal pH, temperature, and carbon and nitrogen sources, were also determined. Li et al. found that optimum growth conditions for F. oxysporum, causing black rot in Gastrodia elata, were as follows: pH 9, 28°C, fructose as a carbon source, and beef extract as a nitrogen source. The strain died after 40 min of treatment at 60°C [67]. Here, the optimum carbon source for F. oxysporum were sucrose, dextrin, soluble starch, and fructose, etc. The optimum nitrogen sources were beef extract, peptone, and yeast powder. The lethal temperature was 65°C, and the best pH was 10. The lethal temperature, and best pH differed from those reported by Li et al. [67]. According to Li et al., the optimum growth conditions for F. solani, causing black rot in Gastrodia elata, were as follows: pH 7, 30°C, mannitol as a carbon source, sodium nitrate as a nitrogen source, and PDA medium as a base medium. The strain died after 40 min of treatment at 65°C [67]. In this study, the optimum culture medium for F. solani was PDA, and the lethal temperature was 65°C. A pH of 10, a carbon source of mannitol, and a nitrogen source of sodium nitrate were optimal. Therefore, the results for F. solani are similar to those found by Li et al. [67]. Nikitin et al. found that F. acuminatum and F. culmorum were mostly identified in temperate climates and did not develop at temperatures above 25°C [68]. Li et al. found that F. sambucinum and F. culmorum, causing potato dry rot, were sensitive to temperature and pH. Fusarium sambucinum showed favorable growth on the glucose-based culture, while F. culmorum preferred the sucrose-based culture [69]. Here, the lethal temperature of F. acuminatum was 50°C, with the lethal temperature for F. culmorum was 50°C, and the best carbon source were sucrose, glucose, dextrin, and soluble starch. The biological characteristics of F. acuminatum and F. culmorum differed from those observed by Li et al. [69] and Nikitin et al. [68]. These differences in the biological characteristics might be related to the source, hosts, and environment of the isolates.
5. Conclusions
This study has provided preliminary insights into the fungal species responsible for root rot in L. barbarum at Nuomuhong Farm in Qinghai Province, China. Following Koch's postulates, 12 pathogen species were identified to cause root rot in L. barbarum. The dominant pathogens included F. equiseti, C. rosea, and M. bolleyi. This research marks the first report of M. bolleyi, F. avenaceum, and F. citri as pathogens causing L. barbarum root rot in China. The lethal temperatures for these 12 pathogen species generally exceeded 50°C. The best media for the 12 pathogens were PDA, CM, and CLA. The optimum carbon sources were sucrose, dextrin and soluble starch, and the optimum nitrogen sources were beef extract, peptone, and yeast extract. These results establish a foundation for the comprehensive prevention and control of Gouqi root rot. Future studies should focus on the screening and development of control agents for these 12 pathogenic fungal species.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1. Maximum parsimony tree of root rot pathogens from L. barbarum inferred from rDNA-ITS sequences. The tree length is 3561. The CI, RI, and RC index were 0.537, 0.950, and 0.510, respectively. Figure S2. Maximum parsimony tree of root rot pathogens from L. barbarum inferred from TEF-1α gene sequences. The tree length is 3871. The CI, RI, and RC index were 0.582, 0.952, and 0.554, respectively. Figure S3. Maximum parsimony tree of root rot pathogens from L. barbarum inferred from RPB2 gene sequences. The tree length is 7092. The CI, RI, and RC index were 0.594, 0.935, and 0.555, respectively. Figure S4. Effects of different media on the pathogen growth rate. Figure S5. Effects of different temperatures on the pathogen growth rate. Figure S6. Effects of different pH values on the pathogen growth rate. Figure S7. Effects of different carbon sources on the pathogen growth rate. Figure S8. Effects of different nitrogen sources on the pathogen growth rate.
Author Contributions
Writing this paper, Yongbao Zhao; Designed the experiments, Hexing Qi; Performed the experiments, Yongbao Zhao and Linshan Wang and Lijie Liu, Hexing Qi; Analyzed the data, Yongbao Zhao and Kaifu Zheng and Chengwen Zheng and Hexing Qi. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Zhejiang Province Key Research and Development Program, grant number 2023C04023.
Institutional Review Board Statement
This study not involving humans or animals.
Data Availability Statement
The original contributions presented in this study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Conflicts of Interest
We declare that no conflict of interest exists.
References
- Wetters, S.; Horn, T.; Nick, P. Goji Who? Morphological and DNA Based Authentication of a "Superfood". Front Plant Sci 2018, 9, 1859. [Google Scholar] [PubMed]
- Mocan, A.; Vlase, L.; Vodnar, D.C.; Bischin, C.; Hanganu, D.; Gheldiu, A.-M.; Oprean, R.; Silaghi-Dumitrescu, R.; Crișan, G. Polyphenolic Content, Antioxidant and Antimicrobial Activities of Lycium barbarum L. and Lycium chinense Mill. Leaves. Molecules 2014, 19, 10056–10073. [Google Scholar] [CrossRef]
- Yao, R.; Heinrich, M.; Weckerle, C.S. The genus Lycium as food and medicine: A botanical, ethnobotanical and historical review. J Ethnopharmacol 2018, 212, 50–66. [Google Scholar] [CrossRef]
- Burke, D.S.; Smidt, C.R.; Vuong, L.T. Momordica cochinchinensis, Rosa, roxburghii, wolfberry, and sea buckthorn-highly nutritional fruits supported by tradition and science. Curr Top Nutraceut R 2005, 3(4), 259.
- Han, Y.; Zhou, Y.; Shan, T.; Li, W.; Liu, H. Immunomodulatory Effect of Lycium barbarum Polysaccharides against Liver Fibrosis Based on the Intelligent Medical Internet of Things. J Healthc Eng 2022, 2022, 6280265. [Google Scholar] [CrossRef]
- Luo, Q.; Cai, Y.; Yan, J.; Sun, M.; Corke, H. Hypoglycemic and hypolipidemic effects and antioxidant activity of fruit extracts from Lycium barbarum. Life Sci 2004, 76(2), 137-149.
- Potterat, O. Goji (Lycium barbarum and L. chinense), Phytochemistry, pharmacology and safety in the perspective of traditional uses and recent popularity. Planta Med 2010, 76(1), 7-19.
- Liang, X.; An, W.; Li, Y.; Qin, X.; Zhao, J.; Su, S. Effects of different nitrogen application rates and picking batches on the nutritional components of Lycium barbarum L. fruits. Front Plant Sci 2024, 15, 355832.
- Zhao, J.; Li, H.; Yin, Y.; An, W.; Qin, X.; Wang, Y.; Fan, Y. Li, Y.; Cao, Y. Fruit ripening in Lycium barbarum and Lycium ruthenicum is associated with distinct gene expression patterns. Febs Open Bi 2020, 10(8), 1550-1567.
- Gnaim, R.; Ledesma-Amaro, R. Synthetic biology of Fusarium for the sustainable production of valuable bioproducts. Blotechnol Adv 2025, 108579.
- Summerell, B.A. Resolving Fusarium, Current status of the genus. Annual review of phytopathol 2019, 57(1), 323-339.
- Doshi, P.; Šerá, B. Role of Non-Thermal Plasma in Fusarium Inactivation and Mycotoxin Decontamination. Plants 2023, 12, 627. [Google Scholar] [CrossRef]
- Zheng, J.; Wang, L.; Hou, W.; Han, Y. Fusarium oxysporum Associated with Fusarium Wilt on Pennisetum sinese in China. Pathogens 2022,11(9), 999-999.
- Khairullina, A.; Micic, N.; Jørgensen, H.J.L.; Bjarnholt, N.; Bülow, L.; Collinge, D.B.; Jensen, B. Biocontrol Effect of Clonostachys rosea on Fusarium graminearum Infection and Mycotoxin Detoxification in Oat (Avena sativa). Plants 2023, 12, 500. [Google Scholar] [CrossRef]
- Thenuwara, G.; Akhtar, P.; Javed, B.; Singh, B.; Byrne, H.J.; Tian, F. Recent Advancements in Lateral Flow Assays for Food Mycotoxin Detection: A Review of Nanoparticle-Based Methods and Innovations. Toxins 2025, 17, 348. [Google Scholar] [CrossRef]
- Ren, X.; Fan, L.; Li, G.; Lyagin, I.V.; Zhang, B.; Ning, M.; Yan, M.; Gao, J.; Wang, F.; Guo, C.; et al. Interaction of Trichoderma Species with Fusarium graminearum Growth and Its Trichothecene Biosynthesis as Further Contribution in Selection of Potential Biocontrol Agents. J. Fungi 2025, 11, 521. [Google Scholar] [CrossRef]
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; Foster, G.D. The Top 10 fungal pathogens in molecular plant pathology. Mol Plant Pathol 2012, 13(4), 414-30.
- Lal, D.; Dev, D.; Kumari, S.; Pandey, S. ; Aparna; Sharma, N.; Nandni, S.; Jha, R.K.; Singh, A. Fusarium wilt pandemic, current understanding and molecular perspectives. Funct Integr Genomic 2024, 24(2), 41.
- Munhoz, T.; Vargas, J.; Teixeira, L.; Staver, C.; Dita, M. Fusarium Tropical Race 4 in Latin America and the Caribbean, status and global research advances towards disease management. Front Plant Sci 2024, 15, 1397617. [Google Scholar] [CrossRef] [PubMed]
- Szabo-Hever, A.; Sharma, J.S.; Faris, J.D.; Zhong, S.; Friesen, T.L.; Fiedler, J.D.; Green, A.J.; Bai, G.; Elias, E.M.; Xu, S.S. Identification and mapping of quantitative trait loci for Fusarium head blight resistance in a synthetic hexaploid × hard red spring wheat population. Plant Genome 2025, 18(3), e70073.
- Chen, S.Y.; Lai, M.H.; Tung, C.W.; Wu, D.H.; Chang, F.Y.; Lin, T.C.; Chung, C.L. Genome-wide association mapping of gene loci affecting disease resistance in the rice-Fusarium fujikuroi pathosystem. Rice (N Y) 2019, 12(1), 85.
- Xie, F.; Sun, Y.; Zhang, H.; Cui, J.; Wang, Q.; Gao, X. ZmBAK1 confers maize resistance to Gibberella stalk rot caused by Fusarium graminearum via activating PAMP-triggered immunity. Plant Signal Behav. 2025, 20(1), 2502739.
- Karlsson, I.; Persson, P.; Friberg, H. Fusarium Head Blight From a Microbiome Perspective. Front Microblol 2021, 12, 628373–628373. [Google Scholar] [CrossRef] [PubMed]
- Pašakinskienė, I.; Stakelienė, V.; Matijošiūtė, S.; Martūnas, J. Diversity of Endophytic Fungi and Bacteria Inhabiting the Roots of the Woodland Grass, Festuca gigantea (Poaceae). Diversity 2024, 16, 453. [Google Scholar] [CrossRef]
- Matušinsky, P.; Florová, V.; Sedláková, B.; Mlčoch, P.; Bleša, D. Colonization dynamic and distribution of the endophytic fungus Microdochium bolleyi in plants measured by qPCR. PLoS One 2024, 19(1), e0297633.
- Ünal, F. Phylogenetic analysis of Microdochium spp. associated with turfgrass and their pathogenicity in cereals. Peer J 2024, 12, e16837–e16837. [Google Scholar] [CrossRef] [PubMed]
- Sung, K.H.; Wan, G.K.; Hyo, W.C.; Lee, S.Y. Identification of Microdochium bolleyi associated with basal rot of creeping bent grass in Korea. Mycobiology 2008, 36(2), 77-80.
- Black, D.L.R.; Brown, A. E. Microdochium bolleyi associated with root rot of flax. Plant Pathol 1986, 35(4), 592-659.
- Liu, J.; Sun, S.; Chen, K.; Gao, S.; Chen, D.; Zhang, Y.; Hu, C. Discerning promotion mechanisms of fungi Clonostachys rosea on growth of freshwater microalga Chlorella sp. by non-contact culture. Algal Research 2025, 86103967-103967.
- Shravani, V.; Iruthayasamy, J.; Nallusamy, S.; Govindasamy, J.; Eswaran, K.; Annaiyan, S. Unveiling Clonostachys rosea bioactives, Investigating antifungal potential against Fusarium oxysporum through in vitro and in silico studies. Research Square 2024.
- Petrucci, A.; Cesarini, M.; Vicente, I.; Merani, L.; Jensen, B.; Collinge, D.B.; Sarrocco, S. Exploring the syner1gistic potential of Trichoderma gamsii T6085 and Clonostachys rosea IK726 for biological control of Fusarium head blight in wheat. Microbiol Res 2025, 296128152.
- Bienapfl, J. C.; Floyd, C. M.; Percich, J. A.; Malvick, D. K. (2012). First report of Clonostachys rosea causing root rot of soybean in the United States. Plant Dis 2012, 96(11), 1700-1700.
- Zhao, J.; Xu, X.; Xu, X.; Guo, L.; Wang, X.; Xiang, W.; Zhao, J. Identification and pathogenicity of Clonostachys spp. and its co-inoculation with Fusarium species on soybean root. Plant Pathol 2024, 73(7), 1801-1811.
- Li, Y.L.; Sun, Z.M.; Li, S.K.; Zhao, X.S.; Abiodun, O. ; Han; Z. M. First report of root rot caused by Clonostachys rosea on Xanthoceras sorbifolium in China. Plant Dis 2024, 108(3), 798.
- Bienapfl, J.C.; Floyd, C.M.; Percich, J.A.; Malvick, D.K. First report of Clonostachys rosea Causing Root Rot of Soybean in the United States. Plant Dis 2012, 96(11), 1700.
- Díaz, R.; Chávez, E.C.; Delgado-Ortiz, J. C.; Velazquez Guerrero, J. J.; Roque, A.; Ochoa, Y.M. First report of Clonostachys rosea causing root rot on garlic in Mexico. Plant Dis 2022, 106(11), 3000.
- Qi, H.; Duan, X.; Xu, W.; Zhou, Y.; Ma, H.; Ma, W.; Ma, G. First report disease of Clonostachys rosea causing root rot on Astragalus membranaceus in China. Plant Dis 2022, 106(6), 1752.
- Li, N.; Chen, W.; Wang, B.; Zhang, C.; Wang, Y.; Li, R.; Yan, Y.; He, J. Arbuscular mycorrhizal fungi improve the disease resistance of Lycium barbarum to root rot by activating phenylpropane metabolism. Front Plant Sci 2024, 15, 1459651. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.Y.; Gao, H.H.; Han, Q.; Jia, C.B.; Swingle, B.; Gao, M.; Su, J.Y. First report of Fusarium redolens causing root rot of Goji berry cv. ‘Ningqi-7’ in China. J Phytopathol 2024, 172(2), e13278.
- Zhang, T.; Zhang, Z.; Li, Y.; He, K. The Effects of Saline Stress on the Growth of Two Shrub Species in the Qaidam Basin of Northwestern China. Sustainability-basel 2019, 11, 828. [Google Scholar] [CrossRef]
- Chen, X. L.; Yang, J.; Peng, Y. L. Large-scale insertional mutagenesis in Magnaporthe oryzae by Agrobacterium tumefaciens-mediated transformation. Methods in molecular biology 2011, 722, 213–224. [Google Scholar]
- Leslie, J.F.; Summerell, B.A. The Fusarium laboratory manual. John Wiley & Sons 2008.
- Xu, J. R.; Hamer, J. E. MAP kinase and cAMP signaling regulate infection structure formation and pathogenic growth in the rice blast fungus Magnaporthe grisea. Genes development 1996, 10(21), 2696–2706.
- Utama, G.L.; Lestari, W.D.; Kayaputri, I.L.; Balia, R.L. Balia. Indigenous yeast with cellulose-degrading activity in napa cabbage (Brassica pekinensis L.) waste, Characterisation and species identification. Foods and Raw Materials 2019, 321-328.
- White, T. J.; Bruns, T.; Lee, S. J. W. T.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR protocols, a guide to methods and applications 1990, 18(1), 315-322.
- Wong, J.Y.; Jeffries, P. Diversity of pathogenic Fusarium populations associated with asparagus roots in decline soils in Spain and the UK. Plant Pathol 2006, 55(3), 331-342.
- Bueno, C.J.; Fischer, I.H.; Rosa D.D.; Firmino, A.C.; Harakava, R.; Oliveira C.M.G.; Furtado, E.L. Fusarium solani F. sp. Passiflorae: a new forma specialis causing collar rot in yellow passion fruit. Plant Pathol 2014, 63(2), 382-389.
- Zhao, J.; Huang, M. Characterization and In Vitro Fungicide Sensitivity of Two Fusarium spp. Associated with Stem Rot of Dragon Fruit in Guizhou; China. J Fungi 2023, 9(12), 1178.
- Swofford, D.L. PAUP* [Phylogenetic Analysis Using Parsimony (*and Other Methods)], Version 4. Sinauer Associates 2003, Sunderland, MA.
- Zhao, X.; Gao, C.; Zhang, X.; Du, M.; Wang, J.; Wang, X.; Lu, B.; Chen, C.; Yang, L.; Zhang, Y.; et al. Identification and biological characteristics of Fusarium tobaccum sp. nov.; a novel species causing tobacco root rot in Jilin Province; China. Microbiol Spectr 2024, 12(12), e00925-24.
- Yang, E.; Fan, L.; Yan, J.; Jiang, Y.; Doucette, C.; Fillmore, S. ; Walker; B. Influence of culture media; pH and temperature on growth and bacteriocin production of bacteriocinogenic lactic acid bacteria. AMB express 2018, 8, 1–14. [Google Scholar]
- Bai, L.; Li, X.; Cao, Y.; Song, Z.; Ma, K.; Fan, Y.; Ma, M. Fusarium culmorum and Fusarium equiseti causing root rot disease on Lycium barbarum (goji berry) in China. Plant Dis 2020, 104(11), 3066.
- Uwaremwe, C.; Yue, L.; Liu, Y.; Tian, Y.; Zhao, X.; Wang, Y.; Xie, Z.; Zhang, Y.; Cui, Z.; Wang, R. Molecular identification and pathogenicity of Fusarium and Alternaria species associated with root rot disease of wolfberry in Gansu and Ningxia provinces; China. Plant Pathol 2021, 70(2), 397-406.
- Feng, Zhilin; Xiao, Y. ; Li, N.; Gao, Q.; Wang J.; Chen, S.; Xing, R. Effects of root rot on microbial communities associated with goji berry (Lycium barbarum) in the Qaidam Basin; China European Journal of Plant Pathol 2023, 167, 853–866.
- Zhang, Z.; Zhang, W.; Wang, X.; Kou, Z.; Wang, Y.; Islam, R.; Zhang, J.Q.; Liu, L.; Xie, T.; Tian, Y.Q. Isolation and identification of antagonistic bacteria of Angelica root rot and their mechanism as biological control. Biological Control 2023, 177, 105120. [Google Scholar] [CrossRef]
- Qi, H.; Qi, X.; Xu, W.; An, Z.; Fan, T. First Report of Root Rot Caused by Fusarium avenaceum and Fusarium flocciferum on Astragalus membranaceus var. mongholicus in China. J Phytopathol 2024, 172(5), e13397.
- Sabahi, F.; Banihashemi, Z.; Mirtalebi, M.; Rep, M.; Cacciola, S.O. Molecular Variability of the Fusarium solani Species Complex Associated with Fusarium Wilt of Melon in Iran. J. Fungi 2023, 9, 486. [Google Scholar] [CrossRef]
- Williams, H. Root and stem rot of cucumber caused by Fusarium oxysporum f. sp. radicis-cucumerinum f. sp. nov. Plant Dis 1996, 80(3), 313.
- Ye, Q.; Wang, R.; Ruan, M.; Yao, Z.; Cheng, Y.; Wan, H.; Li, Z.; Yang, Y.; Zhou, G. Genetic diversity and identification of wilt and root rot pathogens of tomato in China. Plant Dis 2020, 104(6), 1715-1724.
- Ding, X.L.; Lin, S.X.; Zhao, R.W.; Ye, J.R. First report of needle blight on Pinus thunbergii Caused by Fusarium proliferatum in China. Plant Dis 2022, 106(11), 2989.
- Tahat, M.M.; Aldakil, H.A.; Alananbeh, K.M. First report of damping off disease caused by Fusarium oxysporum on Pinus pinea in Jordan. Plant Dis 2021, 105(12), 4153.
- Desjardins, A.E.; Proctor, R.H. Molecular biology of Fusarium mycotoxins. International journal of food microbiol 2007, 119(1-2), 47-50.
- Wang, Q.; Xu, L. Beauvericin, a Bioactive Compound Produced by Fungi: A Short Review. Molecules 2012, 17, 2367–2377. [Google Scholar] [CrossRef]
- He, J.; Zhang, X.; Wang, Q.; Li, N.; Ding, D.; Wang, B. Optimization of the fermentation conditions of Metarhizium robertsii and its biological control of wolfberry root rot disease. Microorganisms 2023, 11(10), 2380.
- Lemanceau, P.; Alabouvette, C. Biological control of fusarium diseases by fluorescent Pseudomonas and non-pathogenic Fusarium. Crop protection 1991, 10(4), 279–286. [Google Scholar] [CrossRef]
- Sivan, A; Chet, I. Biological control of Fusarium spp. in cotton; wheat and muskmelon by Trichoderma harzianum. J. Phytopathol 1986, 116(1), 39-47.
- Li, J.; He, K.; Zhang, Q.; Wu, X.; Li, Z.; Pan, X.; Wang, Y.; Li, C.; Zhang, M. Draft Genome and Biological Characteristics of Fusarium solani and Fusarium oxysporum Causing Black Rot in Gastrodia elata. Int. J. Mol. Sci 2023, 24, 4545. [Google Scholar] [CrossRef] [PubMed]
- Nikitin, D.A.; Ivanova, E.A.; Semenov, M.V.; Zhelezova, A.D.; Ksenofontova, N.A.; Tkhakakhova, A.K.; Kholodov, V.A. Diversity, Ecological Characteristics and Identification of Some Problematic Phytopathogenic Fusarium in Soil: A Review. Diversity 2023, 15, 49. [Google Scholar] [CrossRef]
- Li, F.; Jiang, X.; Sun, M.; Xu, H.; Shi, L.; Shan, W. Screening of culture conditions for pathogens of potato dry rot. Acta Agr Scand B-S P 2014, 64(8), 694-699.
Figure 1.
Symptoms of root rot disease on L. barbarum in the field: A, healthy Gouqi; B, diseased Gouqi; and C, rotted roots of Gouqi.
Figure 1.
Symptoms of root rot disease on L. barbarum in the field: A, healthy Gouqi; B, diseased Gouqi; and C, rotted roots of Gouqi.

Figure 2.
Morphological characteristics of root rot pathogens of Gouqi: A, colony morphology of original isolates; B, colony morphology of re-isolated isolates from infected roots after artificial inoculation; C, conidiophores; D, conidia; and E, chlamydospores.
Figure 2.
Morphological characteristics of root rot pathogens of Gouqi: A, colony morphology of original isolates; B, colony morphology of re-isolated isolates from infected roots after artificial inoculation; C, conidiophores; D, conidia; and E, chlamydospores.
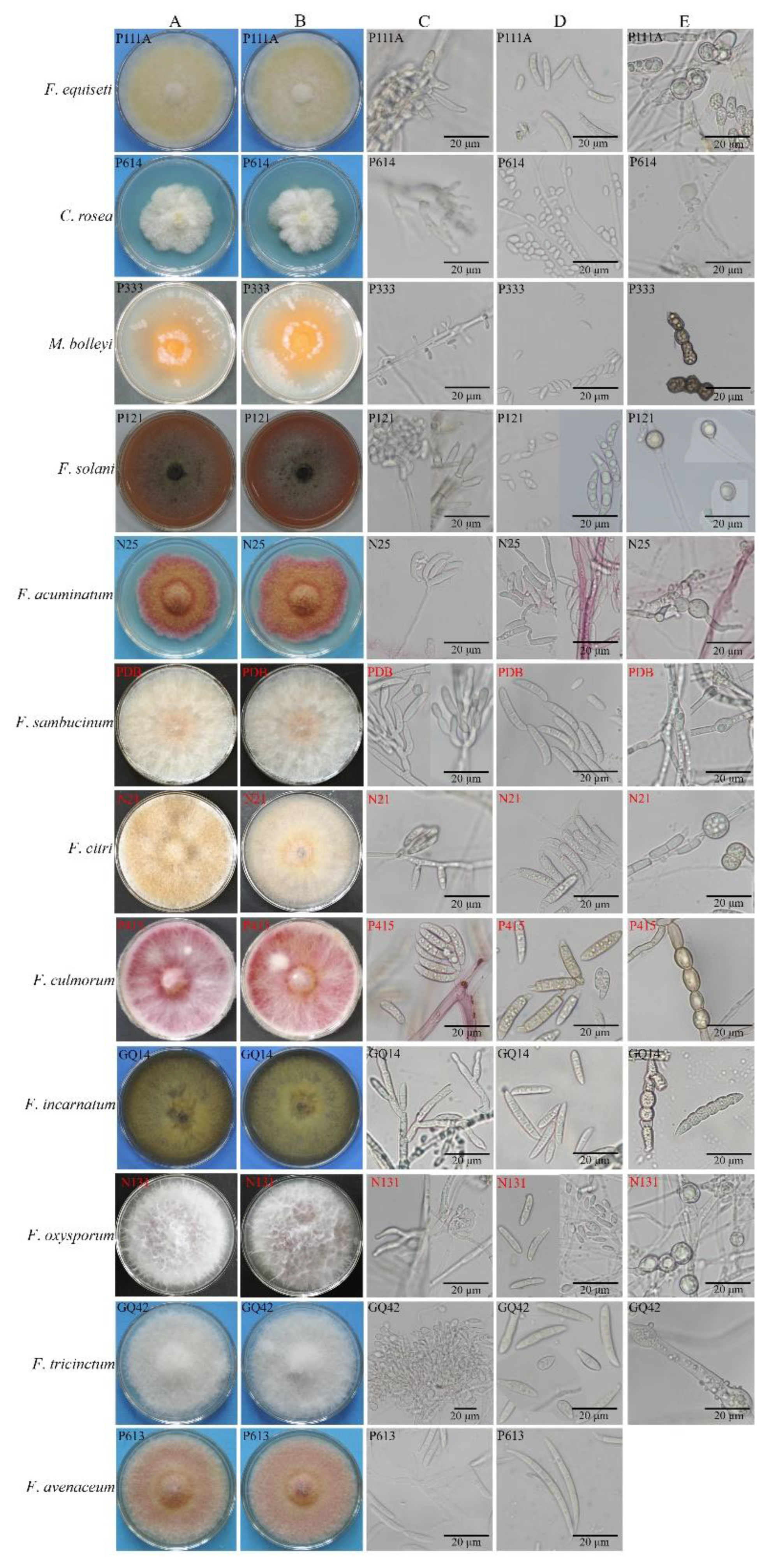
Figure 3.
Maximum parsimony tree of root rot pathogens from Gouqi inferred from combined rDNA-ITS, EF-1α, and RPB2 sequences. The tree length is 14355. The consistency index (CI), retention index (RI), and rescaled consistency (RC) index were 0.531, 0.939, and 0.498, respectively.
Figure 3.
Maximum parsimony tree of root rot pathogens from Gouqi inferred from combined rDNA-ITS, EF-1α, and RPB2 sequences. The tree length is 14355. The consistency index (CI), retention index (RI), and rescaled consistency (RC) index were 0.531, 0.939, and 0.498, respectively.
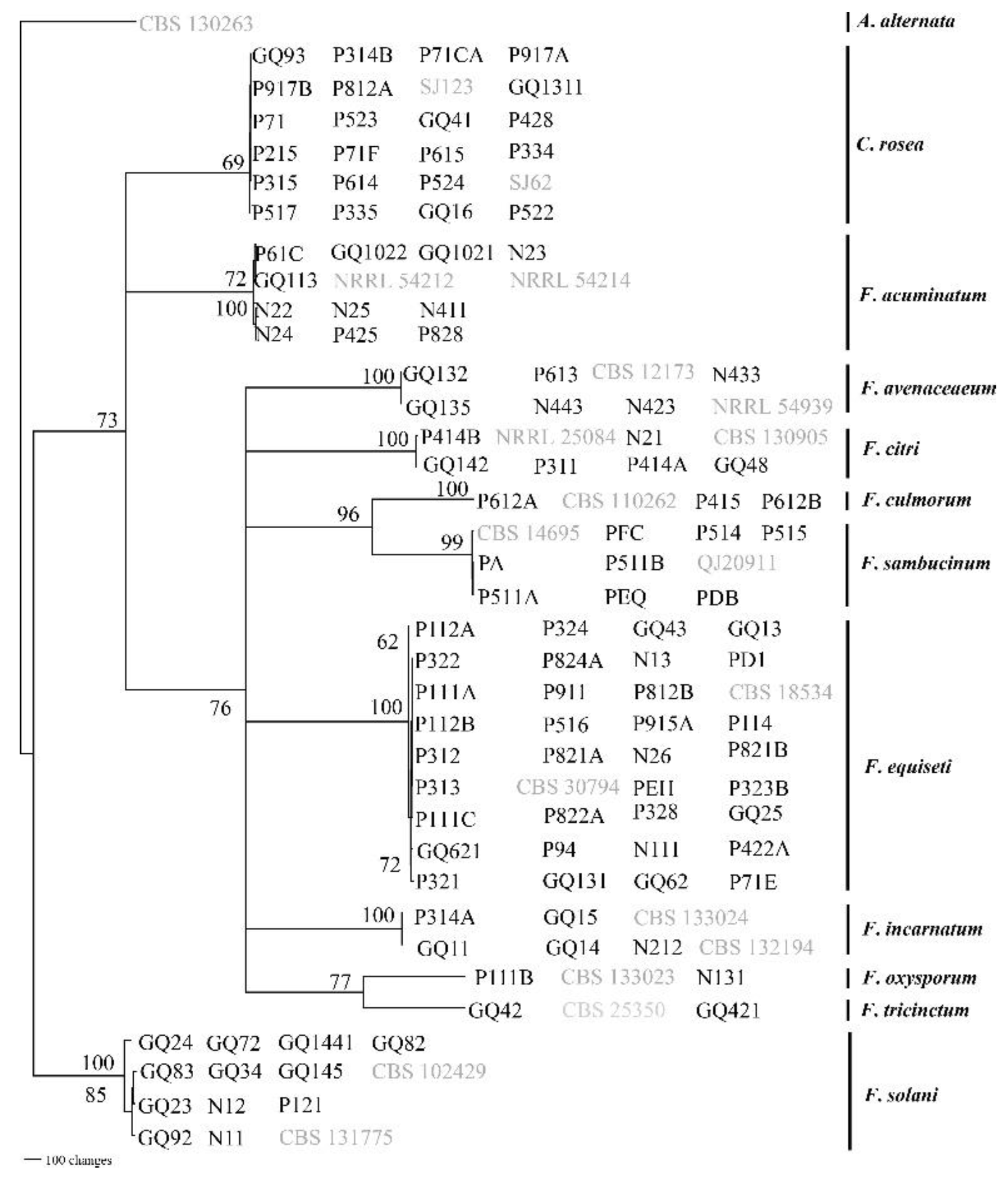
Figure 4.
Root rot symptoms of Gouqi using the scratching inoculation method.

Figure 5.
Root rot symptoms of Gouqi using the root-irrigation inoculation method. A, aboveground part; B, inoculated seedlings; C, underground part; and D, root cross-section.
Figure 5.
Root rot symptoms of Gouqi using the root-irrigation inoculation method. A, aboveground part; B, inoculated seedlings; C, underground part; and D, root cross-section.


Table 1.
Conidia length and width of twelve species of root rot pathogens from L. barbarum (μm).
| Types of spores | F. equiseti | C. rosea | M. bolleyi | F. solani | F. acuminatum | F. sambucinum | F. avenaceum | F. citri | F. incarnatum | F. culmorum | F. oxysporum | F. tricinctum | |
| conidia | microconidium | 11.56±1.46×3.85±0.56 | 7.04±0.41×4.36±0.37 | 9.55±2.04×3.39±0.78 | 7.63±0.91×4.7±0.68 | 8.82±1.65×4.32±0.83 | 7.72±1.36×4.22±0.92 | rare | rare | rare | 8.24±1.56×4.43 ±1.14 | 7.22±1.12×3.26±0.70 | 13.20±2.16×4.52±1.44 |
| macroconidium | 26.44±2.14×5.69±1.43 | 39.09±4.63 ×7.07±1.21 | 20.16±3.02×4.79±0.88 | 34.75±5.38 ×7.56±1.20 | 46.45±4.96×5.74 ±1.54 | 30.84±5.41×5.72 ±1.02 | 30.02±1.51×6.17±1.07 | 37.99±1.96×8.97±0.46 | 33.68±2.21×11.45±0.42 | 33.97±4.95×7.17±1.34 | |||
| chlamydospore | 16.34±1.58×14.15±2.08 | 11.38±2.25×12.45±2.02 | 11.28±1.46×8.00±0.59 | 15.74±2.02×14.19±1.90 | 13.07±2.15×12.84±1.23 | 13.12±3.80× 9.45±2.15 | not found | 11.32±2.10×10.57±2.93 | 13.06±3.48 ×16.73±2.54 | 13.77±2.71×11.91±1.79 | 13.91±1.36×12.77±1.68 | 14.76±4.45×13.56±2.25 | |
Table 2.
Effects of different media on the growth rate of root rot pathogens from L. barbarum (mm).
| Media | F. equiseti | C. rosea | M. bolleyi | F. solani | F. acuminatum | F. sambucinum | F. avenaceum | F. citri | F. incarnatum | F. culmorum | F. oxysporum | F. tricinctum |
| PDA | 0.68±0.06b | 0.44±0.04b | 0.47±0.19bc | 0.78±0.01a | 0.52±0.01c | 0.80±0.00a | 0.80±0.01a | 0.80±0.00ab | 0.80±0.00a | 0.80±0.00a | 0.80±0.00a | 0.72±0.08ab |
| CM | 0.79±0.00a | 0.50±0.01b | 0.34±0.02c | 0.77±0.01b | 0.52±0.06c | 0.80±0.01a | 0.62±0.07b | 0.80±0.01a | 0.80±0.01a | 0.80±0.00a | 0.78±0.01c | 0.64±0.09bc |
| CA | 0.79±0.00a | 0.50±0.07b | 0.28±0.02c | 0.72±0.01d | 0.62±0.07b | 0.80±0.00a | 0.77±0.03a | 0.79±0.01b | 0.80±0.00a | 0.80±0.00a | 0.79±0.01b | 0.68±0.07bc |
| MB | 0.80±0.01a | 0.61±0.04a | 0.60±0.16ab | 0.76±0.01b | 0.53±0.01c | 0.76±0.04a | 0.56±0.06c | 0.80±0.00ab | 0.81±0.01a | 0.80±0.00a | 0.80±0.01a | 0.61±0.05c |
| CLA | 0.79±0.01a | 0.48±0.03b | 0.69±0.03a | 0.73±0.00c | 0.74±0.03a | 0.78±0.03a | 0.79±0.01a | 0.80±0.01a | 0.79±0.02b | 0.80±0.00a | 0.80±0.00a | 0.80±0.01a |
Values are presented as the mean ± standard deviation. Letters represent the significance of differences.
Table 3.
Effects of different temperatures on the growth rate of pathogens from L. barbarum (mm).
| Temperature | F. equiseti | C. rosea | M. bolleyi | F. solani | F. acuminatum | F. sambucinum | F. avenaceum | F. citri | F. incarnatum | F. culmorum | F. oxysporum | F. tricinctum |
| 35℃ | 0.76±0.03a | 0.51±0.05a | 0.36±0.03a | 0.77±0.01a | 0.61±0.01a | 0.79±0.01a | 0.80±0.01a | 0.79±0.01a | 0.79±0.01a | 0.80±0.01a | 0.79±0a | 0.79±0.01a |
| 40℃ | 0.78±0.02a | 0.51±0.01a | 0.34±0.01a | 0.78±0.01a | 0.61±0.03a | 0.79±0.01a | 0.78±0.01a | 0.80±0.01a | 0.79±0.01a | 0.79±0.01a | 0.80±0.01a | 0.76±0.04a |
| 50℃ | 0.77±0.03a | 0.40±0.03b | 0.25±0.01b | 0.77±0.01a | 0 | 0 | 0.17±0b | 0.58±0.12b | 0.79±0.01a | 0 | 0.80±0.01a | 0.63±0.05b |
| 55℃ | 0 | 0 | 0 | 0.47±0b | 0 | 0 | 0 | 0 | 0 | 0 | 0.79±0.01a | 0 |
| 60℃ | 0 | 0 | 0 | 0.35±0c | 0 | 0 | 0 | 0 | 0 | 0 | 0.80±0.01a | 0 |
| 65℃ | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 70℃ | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
Values are presented as the mean ± standard deviation. Letters represent the significance of differences.
Table 4.
Effects of different Ph on the growth rate of pathogens from L. barbarum (mm).
| pH value |
F. equiseti |
C. rosea |
M. bolleyi |
F. solani |
F. acuminatum |
F. sambucinum |
F. avenaceum |
F. citri |
F. incarnatum |
F. culmorum |
F. oxysporum |
F. tricinctum |
| 4 | 0.59±0.02f | 0.50±0.03c | 0.43±0.01b | 0.68±0.01e | 0.45±0.03d | 0.79±0.01ab | 0.52±0.01e | 0.80±0.01a | 0.78±0.01e | 0.79±0.01ab | 0.70±0.02c | 0.67±0.04d |
| 5 | 0.70±0.02cd | 0.52±0.02abc | 0.36±0.01cde | 0.71±0.00c | 0.57±0.01c | 0.79±0.01ab | 0.70±0.02cd | 0.79±0.01ab | 0.79±0.01cd | 0.79±0.00b | 0.79±0.00a | 0.77±0.02abc |
| 6 | 0.73±0.01abc | 0.54±0.03ab | 0.35±0.01e | 0.73±0.00ab | 0.63±0.03b | 0.79±0.01ab | 0.76±0.03b | 0.79±0.01ab | 0.79±0.01bc | 0.79±0.00b | 0.79±0.00a | 0.78±0.01ab |
| 7 | 0.75±0.01a | 0.53±0.00ab | 0.35±0.01de | 0.70±0.01d | 0.64±0.03b | 0.79±0.00b | 0.79±0.01ab | 0.79±0.01ab | 0.78±0.01de | 0.79±0.00b | 0.79±0.02a | 0.79±0.01a |
| 8 | 0.73±0.04ab | 0.53±0.03ab | 0.38±0.01c | 0.72±0.01b | 0.73±0.04a | 0.80±0.01a | 0.79±0.01ab | 0.79±0.01ab | 0.80±0.00a | 0.79±0.00b | 0.75±0.01b | 0.78±0.01ab |
| 9 | 0.73±0.02abc | 0.54±0.02a | 0.59±0.02a | 0.73±0.01ab | 0.58±0.02c | 0.80±0.01a | 0.76±0.03b | 0.80±0.01a | 0.80±0.00a | 0.79±0.01ab | 0.79±0.01a | 0.78±0.01ab |
| 10 | 0.66±0.03e | 0.51±0.01bc | 0.36±0.04cd | 0.74±0.01a | 0.59±0.03c | 0.79±0.01ab | 0.72±0.02c | 0.79±0.00b | 0.80±0.00a | 0.80±0.01a | 0.80±0.01a | 0.74±0.05c |
| 11 | 0.70±0.02bcd | 0.52±0.01abc | 0.36±0.02cde | 0.73±0.00ab | 0.58±0.02c | 0.79±0.01ab | 0.68±0.03d | 0.79±0.00b | 0.80±0.01ab | 0.79±0.01ab | 0.79±0.01a | 0.75±0.02bc |
Values are presented as the mean ± standard deviation. Letters represent the significance of differences.
Table 5.
Effects of different carbon sources on the growth rate of pathogens from L. barbarum (mm).
| Carbon source | F. equiseti | C. rosea | M. bolleyi | F. solani | F. acuminatum | F. sambucinum | F. avenaceum | F. citri | F. incarnatum | F. culmorum | F. oxysporum | F. tricinctum |
| CK | 0.80±0.01ab | 0.44±0.03cd | 0.30±0.01c | 0.78±0.01a | 0.64±0.01b | 0.75±0.02b | 0.62±0.03cd | 0.73±0.03bc | 0.79±0.01ab | 0.79±0.00a | 0.79±0.01ab | 0.65±0.02d |
| glucose | 0.76±0.05b | 0.43±0.02d | 0.35±0.02b | 0.79±0.01a | 0.48±0.02d | 0.72±0.06bc | 0.59±0.03de | 0.73±0.04bc | 0.77±0.03b | 0.80±0.00a | 0.80±0.01a | 0.74±0.03b |
| sucrose | 0.80±0.00ab | 0.43±0.01cd | 0.63±0.02a | 0.79±0.00a | 0.46±0.02d | 0.79±0.01a | 0.64±0.05cd | 0.80±0.00a | 0.80±0.00a | 0.80±0.00a | 0.80±0.01a | 0.79±0.01a |
| mannitol | 0.80±0.01ab | 0.45±0.03cd | 0.37±0.20c | 0.80±0.01a | 0.39±0.02e | 0.75±0.01b | 0.46±0.03g | 0.79±0.00a | 0.79±0.01ab | 0.79±0.01a | 0.80±0.01a | 0.71±0.06c |
| dextrin | 0.79±0.01ab | 0.48±0.07bc | 0.30±0.03c | 0.74±0.01c | 0.52±0.01c | 0.80±0.01a | 0.75±0.04a | 0.79±0.01a | 0.80±0.01a | 0.80±0.01a | 0.80±0.01a | 0.80±0.00a |
| maltose | 0.70±0.05c | 0.50±0.06b | 0.29±0.02c | 0.73±0.02c | 0.66±0.05a | 0.61±0.04d | 0.69±0.02b | 0.75±0.02bc | 0.80±0.01a | 0.79±0.01a | 0.80±0.01a | 0.72±0.02bc |
| soluble starch | 0.79±0.01ab | 0.43±0.03d | 0.30±0.03c | 0.76±0.02b | 0.69±0.03a | 0.80±0.01a | 0.76±0.04a | 0.79±0.01a | 0.80±0.01a | 0.80±0.01a | 0.80±0.01a | 0.79±0.01a |
| lactose | 0.65±0.03d | 0.44±0.08bcd | 0.24±0.03de | 0.73±0.01c | 0.39±0.03e | 0.71±0.01c | 0.56±0.03ef | 0.63±0.04d | 0.64±0.02d | 0.69±0.17b | 0.78±0.01b | 0.47±0.02f |
| cellulose | 0.79±0.01ab | 0.51±0.04b | 0.37±0.02b | 0.79±0.01a | 0.72±0.03a | 0.79±0.01a | 0.70±0.03b | 0.79±0.00a | 0.17±0.02e | 0.79±0.00a | 0.79±0.01ab | 0.78±0.01a |
| fructose | 0.79±0.01ab | 0.57±0.03a | 0.25±0.02de | 0.79±0.00a | 0.53±0.02c | 0.79±0.00a | 0.58±0.02def | 0.79±0.00a | 0.78±0.01ab | 0.79±0.00a | 0.80±0.01a | 0.70±0.02c |
| xylose | 0.72±0.04c | 0.44±0.02cd | 0.21±0.03e | 0.76±0.01b | 0.46±0.04d | 0.74±0.02bc | 0.54±0.02f | 0.72±0.01c | 0.75±0.02c | 0.79±0.01a | 0.79±0.01ab | 0.59±0.02e |
Values are presented as the mean ± standard deviation. Letters represent the significance of differences.
Table 6.
Effects of different nitrogen sources on the growth rate of pathogens from L. barbarum (mm).
Table 6.
Effects of different nitrogen sources on the growth rate of pathogens from L. barbarum (mm).
| Nitrogen source | F. equiseti | C. rosea | M. bolleyi | F. solani | F. acuminatum | F. sambucinum | F. avenaceum | F. citri | F. incarnatum | F. culmorum | F. oxysporum | F. tricinctum |
| CK | 0.80±0.00a | 0.44±0.04bc | 0.63±0.02c | 0.71±0.02b | 0.71±0.05a | 0.69±0.03d | 0.54±0.02c | 0.62±0.08c | 0.61±0.04e | 0.80±0.01ab | 0.74±0.02c | 0.73±0.01b |
| beef extract | 0.80±0.00a | 0.43±0.01bc | 0.80±0.00a | 0.76±0.02a | 0.57±0.10cd | 0.80±0.00a | 0.75±0.03a | 0.71±0.00b | 0.80±0.00a | 0.80±0.00a | 0.80±0.00a | 0.80±0.01a |
| urea | 0.80±0.01a | 0.38±0.01cd | 0.69±0.01b | 0.64±0.01c | 0.37±0.01f | 0.71±0.02cd | 0.49±0.08d | 0.68±0.03b | 0.72±0.04c | 0.80±0.00a | 0.79±0.01ab | 0.49±0.02d |
| peptone | 0.80±0.00a | 0.51±0.10a | 0.80±0.00a | 0.72±0.02b | 0.64±0.05bc | 0.74±0.04bc | 0.76±0.02a | 0.78±0.04a | 0.80±0.00a | 0.80±0.00a | 0.80±0.01ab | 0.80±0.01a |
| yeast powder | 0.80±0.01a | 0.45±0.01ab | 0.80±0.00a | 0.71±0.01b | 0.59±0.06bc | 0.80±0.01a | 0.78±0.02a | 0.80±0.00a | 0.80±0.00a | 0.80±0.01ab | 0.80±0.00a | 0.80±0.00a |
| sodium nitrate | 0.79±0.01a | 0.42±0.02bc | 0.25±0.01f | 0.79±0.01a | 0.51±0.02d | 0.77±0.02ab | 0.69±0.04b | 0.79±0.01a | 0.78±0.01ab | 0.80±0.00a | 0.79±0.01ab | 0.73±0.04b |
| ammonium chloride | 0.64±0.05b | 0.28±0.04e | 0.59±0.05d | 0.49±0.05e | 0.35±0.05f | 0.53±0.07e | 0.34±0.04e | 0.42±0.03d | 0.50±0.04f | 0.78±0.03c | 0.67±0.03d | 0.45±0.05e |
| ammonium nitrate | 0.79±0.00a | 0.35±0.03d | 0.71±0.03b | 0.60±0.02d | 0.44±0.01e | 0.78±0.01ab | 0.48±0.04d | 0.63±0.06c | 0.65±0.02d | 0.79±0.01abc | 0.78±0.01b | 0.65±0.03c |
| saltpeter | 0.80±0.01a | 0.51±0.06a | 0.34±0.01e | 0.78±0.02a | 0.52±0.01d | 0.78±0.01ab | 0.66±0.06b | 0.73±0.03b | 0.76±0.03b | 0.79±0.00abc | 0.78±0.01b | 0.79±0.01a |
Values are presented as the mean ± standard deviation. Letters represent the significance of differences.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



