
Submitted:
06 October 2023
Posted:
09 October 2023
You are already at the latest version
Abstract
The genus Fusarium includes several agronomically important and toxin-producing species, that are worldwide distributed and can cause a wide range of diseases. Crown and stalk rots and grain infections are among the most severe symptoms that Fusarium spp. can cause on maize. The disease development usually occurs during germination, but it may also affect the later phases of plant growth. The purpose of this study was to investigate the diversity and the patho-genicity of 41 isolates recovered from symptomatic seedlings collected in Northern Italy, and from seeds with five different geographical origins during 2019 and 2020. The pathogenicity was tested and confirmed on 23 isolates causing rotting on maize seedlings. A multi-locus phylogeny analysis, based on four genomic loci (tef1-α, rpb2, calm and tub2), was performed for 23 repre-sentative isolates. Representative isolates were identified as species belonging to three species complexes (SC). Fusarium verticillioides and F. annulatum in the F. fujikuroi SC. Fusarium commune was identified in the F. nisikadoi SC, and three different lineages were found in the Fusarium ox-ysporum SC. This study reports F. annulatum, and two lineages of the Fusarium oxysporum SC as maize pathogens for the first time in Italy.
Keywords:
Zea mays L.
; F. fujikuroi SC
; F. nisikadoi SC
; F. oxysporum SC
; multi-locus sequence typing
1. Introduction
Maize (Zea mays L.) is the first staple food in the world [1], and it represents the fifth most produced commodity in the European Union (EU), supplying food, feed and fuel [2]. Italy represents the tenth maize producer in EU, with 52,169,088 tons yielded in 2023 [3]. The Italian production is concentrated in the Northern regions, thus representing an economically relevant sector of agriculture for that area. Several pathogens can affect maize infecting seeds and seedlings and causing important plant diseases that lead to biosafety and phytosanitary problems and important yield and economic losses [4]. Stalk, crown and root rot are among the most severe diseases on maize [4]. Fungal species belonging to the Fusarium genus are one of the main causes of this disease on maize as well as on other cereals. Fusarium spp. are worldwide distributed and include a wide range of agronomically important and toxin-producing plant pathogens, causal agents of wilt, blight, tissues rot, and cankers of many horticultural, ornamental, and forest crops [5,6]. The infection occurs during seed germination, also affecting the plant in later growth phases, causing severe diseases, like root and stalk rot [7,8]. The disease can lead to a premature senescence and lodging of the plants, with different levels of severity depending on the pathogenic species involved, the phenological stage of the plant and the environmental conditions that occur during the cropping cycle. Fusarium species are also able to produce a wide range of mycotoxins, that accumulate in the plant tissues during the infection process, posing an important risk for human and animal health [6,8,9].
In Europe, the main species involved with maize diseases are F. graminearum, F. culmorum, and F. proliferatum [6]. Cases of root rot in maize are related to species of the Fusarium fujikuroi species complex (FFSC), especially to F. verticillioides [10]. Species belonging to the Fusarium oxysporum species complex (FOSC) and the Fusarium nisikadoi species complex (FNSC) were frequently recorded in maize seeds and seedlings [11]. Fusarium mycelium can survive in maize residues and seeds, and it may colonize seedlings and plants through systemic infection [12]. Previous research reported the ability of Fusarium species to infect seeds, transmit the pathogen through the plant and become a source of infection of the roots and stalk, up to the kernels [12,13,14]. The diagnosis of these diseases is often difficult due to the concurrent presence and the multiple isolation of Fusarium pathogens from the same symptomatic portion of the plant [15].
Currently more than 60 species belong to the FFSC, about 144 formae specialis are part of the FOSC, 6 species are included in FNSC, and several species are not officially assigned to a species complex [16,17,18]. Difficulties in Fusarium spp. identification lay on their morphological features, that are usually strongly influenced by environmental conditions, and on their molecular profile, because of wrong classifications of the sequences present in the public database and the nomenclature changes in the taxonomic system [19]. The molecular identification of fungi is usually obtained through sequencing of internal transcribed spacer (ITS), however, in the case of the genus Fusarium, ITS is exclusively able to discriminate the species complex, while the translation elongation factor (tef1-α) and the RNA polymerase second largest subunit (rpb2) genomic regions are highly informative [20,21]. Also, the beta-tubulin (tub2) and the calmodulin (calm) loci are used for Fusarium species identification [22]. Recently, the phylogenomic approach provided high resolution to distinguish species within the Fusarium genus [19]. Thus, the multi-locus phylogenetic analyses, combined with the traditional identification based on morphological methods can deepen the knowledge on this genus.
The purposes of this work, considering the economic importance of maize and the impact of Fusarium species on this crop, are to: (i) determine the pathogenicity of Fusarium spp. isolates obtained from maize seeds and seedlings, and (ii) combine phylogenetic analysis with morphological characterization of the isolates to identify and understand the diversity of the Fusarium species affecting maize, causing stalk and crown rot, in Northern Italy.
2. Results
2.1. Fungal isolates
The observed symptoms on maize plants consisted of browning, wilting and collapse of the seedlings, due to the decaying tissues of the stem. Disease incidence in the field was established considering the percentage of affected plants and ranged from 5 to 20%, depending on geographical location of the field. The symptoms were observed on seedlings of different maize hybrids, already at the V1 stage. Rotting kernels covered by mycelium were observed in the incubation test. The recorded percentage of seeds infected with Fusarium spp. in the incubation test ranged between 5 and 56%. Forty-one isolates, obtained from affected root, stem and crown tissue of the seedlings collected in the field and from the incubation test on seeds, were identified as belonging to Fusarium spp. (Table 1).
2.2. Pathogenicity test
A total of 36 out of 41 isolates tested caused root and crown rot like those observed in the field during spring 2019 and 2020 (Figure 1).
Different severity indexes, depending on the isolate tested, were observed. A total of 19 isolates showed disease indexes ranging from 13.3% to 46.7%, and only 17 of them showed a disease index higher than 50% (Table 2).
The identity of the re-isolated fungi was proved by sequencing the tef-1α locus, confirming the Koch’s postulates. No symptoms were observed on healthy control plants. A total of 23 out of 36 pathogenic isolates were selected as representative isolates, based on their cultural features, to proceed with molecular analyses and their characterization.
2.3. Phylogenetic analyses
The preliminary analysis conducted on the obtained sequences showed that the 23 selected isolates belong to three Fusarium species complexes, Fusarium fujikuroi SC, Fusarium nisikadoi SC and Fusarium oxysporum SC. The combined phylogeny analyses of tef-1α, rpb2, calm and tub2 performed for FFSC isolates consisted of 101 sequences, including the outgroup sequence of Fusarium foetens (CBS 120665). A total of 2210 characters (tef-1α: 1-621, rpb2: 628-1185, calm: 1192-1726, tub2: 1733-2210) were included in the analysis: 563 characters resulted as parsimony-informative, 604 as variable and parsimony uninformative, and 1025 were constant. A maximum number of 1000 equally most parsimonious trees were saved (Tree length = 2973, CI = 0.602, RI = 0.812 and RC = 0.488). Bootstrap support values obtained with the parsimony analysis are showed on the Bayesian phylogenies in Figure 2. Bayesian analyses, the dirichlet state frequency distributions were suggested by MrModeltest for analysing all the partitions. The following models, recommended by MrModeltest, were used: GTR+G for tef-1α, SYM+I+G for rpb2, SYM+G for calm, and HKY+G for tub2. In the Bayesian analysis, the tef1-α partition had 370 unique site patterns, the rpb2 partition had 191 unique site patterns, the calm partition had 233 unique site patterns, the tub2 partition had 269 unique site patterns and the analysis ran for 405000 generations, resulting in 812 trees of which 305 trees were used to calculate the posterior probabilities. In the combined analyses eight isolates clustered with seven reference isolates of F. verticillioides, while six isolates were grouped with 3 isolates known as reference of F. annulatum [19].
The combined phylogeny analysis of the three loci (tef-1α, rpb2 and calm) performed for FOSC isolates consisted of 47 sequences, including the outgroup sequence of Fusarium udum (NRRL22949). A total of 1762 characters (tef-1α: 1-589, rpb2: 596-1231, calm: 1238-1762) were included in the analysis: 77 characters resulted as parsimony-informative, 171 as variable and parsimony uninformative, and 1502 were constant. A maximum number of 1000 equally most parsimonious trees were saved (Tree length = 297, CI = 0.882, RI = 0.892 and RC = 0.787). Bootstrap support values obtained with the parsimony analysis are showed on the Bayesian phylogenies in Figure 3. For the Bayesian analyses, the dirichlet state frequency distributions were suggested by MrModeltest for analysing all the partitions. The following models, recommended by MrModeltest, were used: HKY for tef-1α, K80 for rpb2, and calm. In the Bayesian analysis, the tef1-α partition had 109 unique site patterns, the rpb2 partition had 71 unique site patterns, the calm partition had 57 unique site patterns and the analysis ran for 300000 generations, resulting in 602 trees of which 226 trees were used to calculate the posterior probabilities. In the combined analyses one isolate clustered with four reference isolates and the ex-type of F. nirenbergiae, one isolate was identified as F. cugenangense, while five isolates were identified as F. oxysporum sensu lato, cause they not cluster with anyone of reference sequences, according with the recent taxoxonomy revision of this SC reported by Lombard et al. [17].
The combined phylogeny analysis of the four loci (tef-1α, rpb2, calm and tub2) performed for FNSC isolates consisted of 15 sequences, including the outgroup sequence of Fusarium udum (NRRL22949). A total of 2024 characters (tef-1α: 1-585, rpb2: 592-1362, calm: 1369-1594, tub2: 1601-2024) were included in the analysis, 186 characters resulted as parsimony-informative, 333 as variable and parsimony uninformative, and 1487 were constant. A maximum number of 1000 equally most parsimonious trees were saved (Tree length = 616, CI = 0.959, RI = 0.922 and RC = 0.884). Bootstrap support values obtained with the parsimony analysis are showed on the Bayesian phylogenies in Figure 4. For the Bayesian analyses, the dirichlet state frequency distributions were suggested by MrModeltest for analysing all the partitions. The following models, recommended by MrModeltest, were used: HKY for tef-1α, HKY+G for rpb2, JC for calm, and SYM+G for tub2. In the Bayesian analysis, the tef1-α partition had 106 unique site patterns, the rpb2 partition had 47 unique site patterns, the calm partition had 19 unique site patterns, the tub2 partition had 57 unique site patterns and the analysis ran for 400000 generations, resulting in 802 trees of which 301 trees were used to calculate the posterior probabilities. In the combined analyses two isolates clustered with seven reference isolates of F. commune.
2.4. Morphology
Morphological features, supported by phylogenetic analysis, were assessed, and used to characterize 6 species, belonging to three species complexes, found in this study. (Figure 5, Figure 6 and Figure 7).
Seven-day-old colonies of F. verticillioides showed white abundant aerial mycelium that developed violet pigments with age. Colony radius was 55–70 mm. Monophialides were produced and appeared in V-shaped pairs, similar to “rabbit ears”. Microconidia were hyaline, oval to club shaped, aseptate, (6-)7-12(-13) x 2.5-3.5 μm (mean 7 x 3.0 μm), abundant in aerial mycelium and disposed in long chains. Macroconidia were straight and slender, with the apical cell foot-shaped, 4-6 septate, hyaline, (28-)32-49(-52) x 2.5-3 μm (mean 38.5 x 3.0 μm). Chlamydospores were absent.
F. annulatum colonies, after 7 days at 25°C on PDA reached 50-60 mm diameter. The surface was characterized by white aerial mycelium that became darker with age, while the reverse showed intense pink to purple pigments at the center of the colony. Conidiophores produce mono and polyphialides, which generates a large number of microconidia that could be grouped in long chains, on CLA. Microconidia are formed on aerial conidiophores, hyaline, oval to elliptical, aseptate, (2-)5-12(-15) x 1.5-3.5 μm (mean 8.8 x 2 μm). Macroconidia are hyaline, slender, straight to curve, with foot-shaped apical cell, 4-5 septate, (30-)35-42(-54) x 2-4 μm (mean 37 x 3 μm). Chlamydospores are absent.
F. commune colonies morphology was characterized by white to pink, abundant, floccose to fluffy mycelium on the surface and by violet pigmentation on the reverse colony. After 7 days of incubation at 25°C colony radial growth reach 45-50 mm, on PDA. F. commune produced both, mono and polyphialides. On CLA the isolates produced slightly curved 3-4 septate macroconidia (23-)28-56(-66) x 2.5-6 μm (mean 38.5 x 4 μm) and aseptate, cylindrical, and straight microconidia (3.5-)5-7(8.2) x 2-3 μm (mean 6 x 2.5 μm). Chlamydospores were produced, single or in pairs.
F. nirenbergiae colony radial growth measure 55-60 mm, after 7 days on PDA. The colony surface was characterized by an abundant pink and floccose mycelium, and by greyish pink pigments on the reverse. Conidiophores carried on the aerial mycelium produced monophialides that bears oval, aseptate microconidia (8-)9-15(-16.2) x 2-3.5 μm (mean 11.2 x 3.2 μm), and 3-4 septate, slender, straight, with a papillate apical cell and a foot shaped basal cell (26.5-)28-30(-32.2) x 2.5-4.8 μm (mean 28.5 x 3.4 μm) macroconidia. Globose chlamydospores were produced.
Figure 7.
Morphological features of the species belonging to FOSC identified in this study, (A, B, C, D, E, F) F. cugenangense, (G, H, I, J, K, L) F. nirenbergiae, and (M, N, O, P, Q, R) F. oxysporum sensu lato. a-b-g-h-m-n. Colonies on PDA above and below; c–d-i-j-o-p. conidia; e-k-q. conidiogenous cells; f-l-r. chlamydospores.— Scale bars = 10 μm.
Figure 7.
Morphological features of the species belonging to FOSC identified in this study, (A, B, C, D, E, F) F. cugenangense, (G, H, I, J, K, L) F. nirenbergiae, and (M, N, O, P, Q, R) F. oxysporum sensu lato. a-b-g-h-m-n. Colonies on PDA above and below; c–d-i-j-o-p. conidia; e-k-q. conidiogenous cells; f-l-r. chlamydospores.— Scale bars = 10 μm.
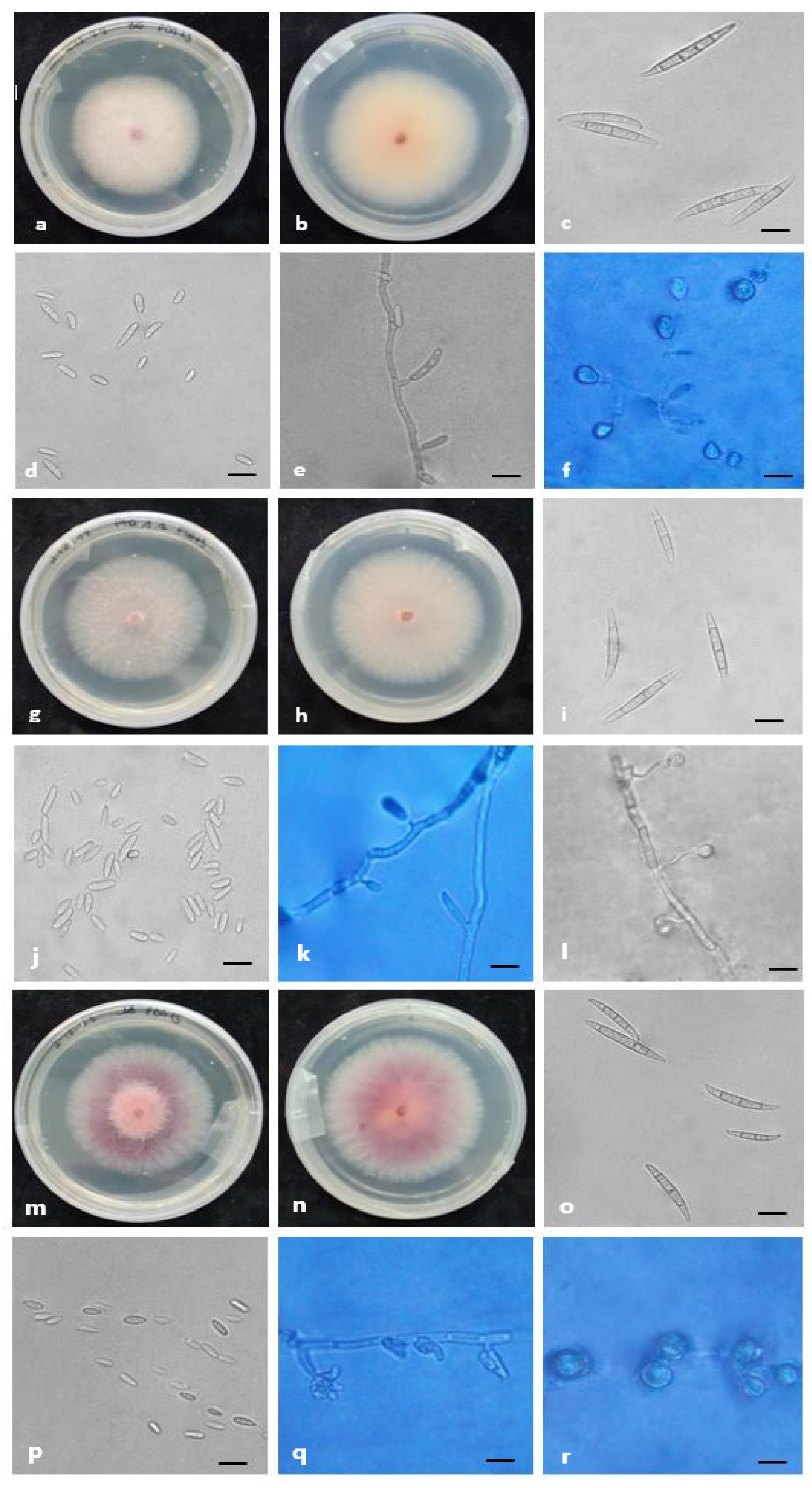
One isolate was identified as F. cugenangense, which colony morphology on PDA was characterized by white to pink, abundant, and cottony mycelium on the surface and pink at center to pale grey on colony reverse. Colony radius, after 7 days at 25°C under 12-h photoperiod on PDA, was 40–56 mm. It was characterized by monophialidic conidiogenous cells that produced 3-6 septate macroconidia (42.5-)46-55(-56.2) x 5.5-6.5 μm (mean 50.2 x 6 μm), with papillate apical cells and foot-shaped basal cells. Microconidia were abundant, oval to elliptical, 0-3 septate (7-)8.3-10.5(-13) x 4-7.5 μm (mean 9 x 5.6 μm). Chlamydospores are globose and formed single or in pairs.
The isolates classified as F. oxysporum sensu lato were characterized by an abundant pink to purple and floccose mycelium, and by purple to red pigments on the reverse. Colony radius was 50–60 mm after 7 days at 25°C under 12-h photoperiod on PDA. The isolates were characterized by conidiophores that produced monophialides, that bears slender, straight, 3-5 septate, with foot shaped basal cells and papillate apical cells macroconidia (29-)30-37(-44) x 3-4.5 μm (mean 35 x 3.8 μm). Microconidia were abundant, oval, aseptate (5.5-)6-11(-15) x 2-3 μm (mean 9.2 x 2.5 μm). Single chlamydospores were formed.
3. Discussion
Several species of Fusarium represent a severe problem for cereals cultivation and production worldwide causing relevant yield and economic losses and posing a serious threat to human and animal health due to their ability to produce mycotoxins [4].
In the present study, Fusarium spp. were isolated from maize seedlings with symptoms of root and crown rot in Northern Italy, and from rotted kernels collected in 5 different countries with the aim to investigate their diversity and pathogenicity. Isolates from seed were included, due to the ability of Fusarium species to be seed borne and seed transmitted [12,23], causing stalk, crown and root rot that can be observed in the field under favorable soil moisture and temperature conditions. A polyphasic approach was used to study the fungal isolates obtained from the affected plants including the analysis of multiple characters, since the morphological features alone, which represent the traditional identification method used for Fusarium spp. identification, are not enough to discriminate among species [16]. The combination of multi-locus sequence analysis, pathogenicity data and morphological characteristics, represent the best way to characterize fungi at species level. According to O’Donnell et al. [24], ITS region is not able to distinguish Fusarium species boundaries and for this reason was not considered in this study. The tef1-α, rpb2, tub2 and calm loci were used for Fusarium spp. identification, according to the previous phylogenetic analysis of the genus reported in literature [16,17,19]. Six different species were identified in association with infection of crown, root and seeds of maize: F. verticillioides and F. annulatum belonging to the FFSC, F. commune belonging to the FNSC, and three different lineages in the FOSC. The FFSC contains 84 described species, including a large number of cryptic species identifiable only based on phylogenetic inference [16,18,19]. The complex includes important plant pathogens and toxin-producers [16], and species belonging to FFSC can be discriminated from other complexes due to the production of macroconidia, a large amount of microconidia and sporadically of chlamydospore [19]. The results obtained in this study allowed the classification of 14 isolates in this complex, identified as F. verticillioides and F. annulatum.
Fusarium verticillioides is one of the most important species that affects maize, it is worldwide distributed and it can cause important yield and grain quality losses [25]. It is primarily reported as the causal agent of the ear rot on maize, however studies also reported the pathogen as responsible of symptoms of seedlings decay, and stalk, crown, and root rot on maize [19,26,27]. F. annulatum, firstly described by Bugnicourt et al. [28], is a species associated with symptoms of rot on different crops, such as cantaloupe melons in Spain or saffron in China [29,30]. The name F. annulatum is often confused with F. proliferatum, a well-known maize pathogen, associated with crown and root rot [15,31]. A phylogenetic analysis based on LSU, SSU, and tub2 genomic loci showed that the reference sequence of F. annulatum (CBS 258.54) introduced by Bagnicourt [28], clustered with representative strains of F. proliferatum (CBS 217.76, NRRL 25089) [32]. These results led to the wide report of F. proliferatum, instead of F. annulatum, as maize pathogen. However, a recent multi-locus phylogenetic analyses based on calm, rpb1, rpb2, and tef1-α loci, including the epitype of F. proliferatum (CBS 480.96), established that this species clustered distant from F. annulatum [19]. The same study demonstrated that several cereals pathogenic isolates, identified as F. proliferatum in previous researches [15,31,33], should be identified as F. annulatum. The present research, based on the taxonomic characterization of Yilmaz et al. [16], demonstrated the characterization of the pathogenic isolates as the species F. verticillioides and F. annulatum, which belong to the same species complex and represent the highest proportion of the pathogenic isolates infecting maize samples considered in this study. To our knowledge this is the first report of F. annulatum as causal agent of stalk, crown and root rot on maize, in Italy. Fusarium commune belongs to FNSC and it is principally known as a pathogen of rice and maize [34]. Its behavior as pathogen is similar to those of some species belonging to FOSC, causing rot and wilt of the plants [19]. Recent studies reported F. commune as causal agent of stalk, crown and root rot on maize in Italy [35], and in Liaoning province in China [36]. The phylogenetic analysis conducted by Skovgaard et al. [37] identified the species as a sister group to FOSC, a result supported by the high morphological similarity between these taxa. Species of FNSC could be distinguished from those of FOSC only because of the presence of long and thin monophialides and the occasional production of polyphialides [34,37]. To discriminate and identify the species the tef1-α genomic region was used, due to its high phylogenetic signal [34]. Fusarium oxysporum is an economically important soilborne and ubiquitous plant pathogen, that covers the fifth place in the top ten rank of the most important phytopathogens [38], and it is mainly known as causal agent of plant wilts. The challenge in identification of the species belonging to this complex is due to the inability to discriminate them on the basis of morphological features, the affected wide host range and their geographical distribution [39,40]. The tef1-α and rpb2 genomic loci provided the best resolution in distinguishing the species, as seen by Lombard et al. [17]. The calmodulin provided a little support, while the β-tubulin was excluded. Considering the current literature [17,19], the multi-locus phylogenetic analysis performed in this study allowed to identify seven isolates within three lineages of FOSC. The first lineage includes one isolate which formed a well-supported clade with the reference isolate and the ex-type of F. nirenbergiae. The second lineage includes one isolate which clustered with reference of F. cugenangense. Whilst the third lineage includes five isolates that did not cluster with any of the reference species used for the phylogenetic analyses and that were defined as F. oxysporum sensu lato. Fusarium nirenbergiae belongs to FOSC and it is reported as pathogen on saffron in China [30], and on passion fruit in Italy [41]. It was recently described as pathogen on maize in China [19], and our study represents the first finding of this species as maize pathogen in Italy. It is closely related to F. curvatum, and it can be morphologically distinguished from this species by the production of monophialidic conidiogenous cells and the production of chlamydospores, that are absent in F. curvatum [17]. For the species identification, morphological features must be supported by phylogenetic inference. The tef-1α and the rpb2 gene regions provided the best resolution to distinguish the species [17]. Fusarium cugenangense was previously included in the species F. oxysporum f. sp. cubense, the causal agent of banana wilt, however phylogenetic analyses distinguished this lineage as a new independent species [42]. This pathogen has a wide host range, such as Acer palmatum, Crocus sp., Gossypium barbadense, Hordeum vulgare, Solanum tuberosum, Smilax sp., Tulipa gesneriana, Musa nana, Musa sp., Vicia faba and Zea mays [18,19,42]. To our knowledge, this is the first report of F. cugenangense as pathogen of Zea mays in Italy. This species is closely related to F. callisthephi, F. elaeidis, and to other formae speciales, however it can be discriminated from the other species under the morphological point of view, due the septation of the macroconidia, and the only production of monophialides [17,42]. Molecular identification and discrimination were supported by the amplification of tef-1α and rpb2 loci [17]. The identification of species belonging to FOSC represents a great challenge due to the complexity and the endless evolution of the taxonomy of the genus Fusarium. During the last decades, a plethora of new species was described, increasing problems for Fusarium taxonomy users [43]. Therefore, there is an agreement on the need to stabilize the taxonomy of the complex while conducting further studies to clarify species concepts to allow the correct characterization of species within FOSC. [17,43,44]. The high species diversity, found in the present study from a molecular point of view, should be supported by analyses on the pathogenicity and host preference of these species.
The pathogenicity tests, hereby performed, confirmed that all the species were able to cause symptoms of crown and root rot in maize seedlings. This is in line with the results obtained by other scientists which contribute to increase the knowledge on the complexity of the maize microbiome and on the etiology of soilborne diseases [45,46,47]. The isolates that were confirmed as pathogenic, showed different levels of aggressiveness on maize seedlings. The F. verticillioides, F. annulatum, F. commune isolates always showed a disease index higher than 50%, except for one isolate of F. verticillioides (8.2) that showed a disease index of 20%. Regarding the isolates belonging to FOSC, one isolate of F. oxysporum sensu lato and the isolate of F. nirenbergiae showed a disease index higher than 50%, while the other isolates of F. oxysporum sensu lato and the isolate of F. cugenangense showed lower indexes, ranging from 20% to 45%. Considering the economic and agronomic relevance of maize, and the susceptibility of this crop to pathogenic Fusarium species, it is important to provide a correct diagnosis for a rapid and effective disease management. This study investigated the species involved in maize diseases associated with symptoms of stalk, crown, and root rot in Northern Italy as well as those associated with seeds from different countries. Moreover, it provides useful information on tools to analyze the target loci to identify Fusarium species laying the base for future studies on their detection to develop specific and sensitive diagnostic tools that speed up the diagnosis of these pathogens. The identification process usually requires long time and several steps, starting from the description of the symptoms, the environmental conditions in which the infection occurred, the isolation, purification and morphological and molecular identification of the causal agents of the disease observed [48]. The development of rapid, specific and accurate molecular diagnostic tools could allow the identification and quantification of multiple pathogens in symptomatic plants and seeds as well as in those not yet expressing symptoms. Further investigations should be addressed to evaluate the putative cross pathogenicity of these species and the seed-borne rate in causing the symptoms observed in the field and reproduced in this study, to provide a deeper insight on the pathogens and disease development, then to improve management sustainable control strategies.
4. Materials and Methods
4.1. Fungal isolates
During 2019 and 2020, different surveys were conducted in six maize fields in Northern Italy. The surveyed fields were in San Zenone degli Ezzelini (VI), Cigliano (VC) and Crescentino (VC). Root and crown rot symptoms were detected on seedlings of different hybrids of maize early in the season, between V1 (first leaf) and V3 (third leaf) phenological stage. Symptomatic samples were collected and washed under running tap water for 2 minutes to remove soil debris. Small sections (0.1-0.2 cm) were cut on the edge of the symptomatic portions, surface sterilized in 1% hypochlorite solution for one min, rinsed in sterile distilled water and placed on potato dextrose agar (PDA, Merck, USA) to isolate fungi. After an incubation of 72 hours at room temperature, the plates were observed and mycelial plugs from the developed fungal colonies were transferred on new PDA plates to obtain pure cultures.
In 2019, from 24 commercial lots , produced in 5 different countries (France, Italy, Romania, Turkey and USA), 500 g of seeds were sampled and analysed with an incubation test to evaluate their phytosanitary conditions [49]. A total of 400 seeds of each lot was disinfected with 100 ml of a water solution containing 55.9% of commercial chlorine (5.37%), 10.4% of absolute alcohol (96%) and 10 µl of Tween 20 for 15 min and then rinsed three times with sterile distilled water and dried on sterile paper. The disinfected seeds were placed in 12 × 12 plastic boxes over three layers of sterile filter paper soaked with a 0.05% sodium hypochlorite water solution. The boxes were placed in a growth chamber at 25 °C ± 2 °C, under a 12 h near-ultraviolet light (NUV)12 h dark cycle, for 48 h, then for 24 h at -20 °C and then incubated in the growth chamber for 11 days. Colonies were isolated from seeds and placed on PDA plates to obtain pure cultures.
4.2. Pathogenicity test
The pathogenicity of the 41 isolates was assessed following the protocol described by Okello et al. [15]. Pure cultures of the isolates were grown on PDA, amended with 25 mg/L of streptomycin sulphate, for 14 days at room temperature. After two weeks, mycelium plugs (15 mm) of each isolate were transferred into conical flasks (250 ml) containing a sterile sand/corn meal substrate, prepared with 54 g of sand, 6 g of corn meal and 10 ml of deionized water per flask. Five replicate flasks were used for each isolate. The inoculated flasks were then incubated at 23±2°C for 23 day, mixing them daily. A total of 300 maize seeds (P1565, Pioneer Hi-Bred, Italy) was incubated at 23±2°C for three days in Petri dishes filled with moisturized sterile filter paper to promote their germination and to obtain seedlings for inoculation. Once germinated, six seedlings per isolate were transplanted in inoculated pots (volume 2L) filled, following the protocol described by Bilgi et al. [50], with a first layer of 40 g of perlite, followed by a second layer of 20 g of inoculum and a final layer of 20 g of perlite. A total of 123 inoculated plastic pots were used considering 2 seedling per pot and 3 pots per fungal isolate. The pots were incubated in the greenhouse at 22±2°C for 14 days. The root rot severity was assessed with a scale that ranged from 1 to 5 at 14 days post-inoculation. The adopted scale was as follow: 1 = germinated seed and healthy seedling without symptoms of root rot; 2 = germinated seeds and 1-19% of symptomatic roots; 3 = germinated seed and 20-74% of symptomatic roots; 4 = germinated seed and> 75% symptomatic roots; 5 = complete colonization of the seed and undeveloped seedling [51]. The data were expressed as disease index (DI) 0–100, calculated with the following formula: DI= [∑ (i × ni)]⁄(4 × total of plants)]×100; where i = 0–4 and ni is the number of plants with rating i. The assay was performed in triplicate and the data obtained expressed as mean value of the three replications carried out.
4.3. Data analyses
The data were subjected to analysis of variance (ANOVA), after testing that the resulting disease index data were normally distributed with a Levene's test, using SPSS Statistics v. 27.0 (IBM Corp., Armonk, NY, U.S.A.). The Duncan’s test was used to explore differences between multiple group means (p ≤ 0.05). Statistical analysis was performed with the Statistical Package for Social Science (SPSS, IBM, Chicago, IL, USA) version 27.0.
4.4. DNA extraction, PCR and sequencing
A total of 23 isolates was selected as representative based on their positive results in the pathogenicity test and used for the following analyses. Genomic DNA was extracted from each isolate, transferring 100 mg of mycelium in a 2 mL microcentrifuge tube and following the manufacturers’ instructions of the Omega E.Z.N.A.® Fungal DNA mini kit (Omega Bio-Tek, Norcross, GA, USA), after a 15-min cycle, at 25 Hz, in Tissuelyser (Qiagen®). Partial translation elongation factor-1α (tef-1α), RNA polymerase second largest subunit (rpb2), calmodulin (calm), and beta-tubulin (tub2) genomic regions were amplified using EF1 and EF2 [52], rpb2-7cr and rpb2-5f [53], CAL-228f and CAL-737r [54], CL1 and CL2A [24] and T1 [55] and Bt2b [56] primers, respectively. The PCR mixtures and the cycling conditions for the amplification of tef-1α, calm and tub2 followed the protocols described by Guarnaccia et al. [57] and Weir et al. [58]. For the rpb2 the PCR protocol by Yilmaz et al. [16] was optimized as follows: 94 °C 90 s; 40 cycles of 94 °C 30 s, 55 °C 90 s, 68 °C 2 min; 68 °C 5 min. PCR amplification was checked by electrophoresis on 1% agarose (VWR Life Science AMRESCO® biochemicals) gels stained with GelRedTM. PCR products were sequenced by BMR Genomics (Padova, Italy) and the obtained sequences were analyzed and assembled with the program Geneious v. 11.1.5 (Auckland, New Zealand).
4.5. Phylogenetic analyses
The sequences generated in this research were analysed with the NCBIs GenBank database through the BLAST-N program to determine the closest species and the species complexes to which they belong and then compared with reference sequences reported in literature [16,17,18,19,24,39,44,59,60,61,62,63,64,65,66] and downloaded from GenBank, to establish the identity of the explored isolates. All the different regions of the sequences of this study and those downloaded from GenBank were aligned with the MAFFT v. 7 online server (http://mafft.cbrc.jp/alignment/server/index.html) [67], and then manually adjusted in MEGA v. 7 [68]. A preliminary analysis was conducted on the tef1-α region (data not shown) to determine which species complex the representative isolates belonged to. Phylogeny was processed through different analyses conducted as multilocus sequence analyses using different datasets in accordance with previous studies [16,17,19]. The analysis for the FFSC and the FNSC were performed combining tef-1α, rpb2, calm and tub2 datasets, rooted with F. foetens (CBS 120665) and F. udum (NRRL 22949), respectively. The combined tef-1α, rpb2 and calm datasets were used to perform the analyses for the FOSC, rooted with F. udum (NRRL 22949). The phylogenies were based on Maximum-Parsimony (MP) and Bayesian Inference (BI) methods. The MP analysis were performed with PAUP [69], while the Bayesian analyses were carried out with MrBayes v. 3.2.5 [70], including the best evolutionary model for each partition as defined by MrModelTest v. 2.3 [71]. BI analyses were processed using four Markov Chain Monte Carlo (MCMC) chains with a sampling frequency of 1000 generations. The heating condition was set to 0.2 and the analyses end when the standard deviation of split frequencies was less than 0.01. For the MP analyses, phylogenetic relationships were estimated by heuristic searches with 100 random addition sequences. Tree bisection-reconnection was used, with the branch swapping option set on ‘best trees’ only with all characters weighted equally and alignment gaps treated as fifth state. Tree length (TL), consistency index (CI), retention index (RI) and rescaled consistence index (RC) were calculated for parsimony and the bootstrap analyses were based on 1000 replications. The clade is supported when the Bootstrap support value is ≥ 70 %, and the Bayesian PP value is ≥ 0.9. Sequences generated and used in this study were deposited in GenBank (Table 3).
4.6. Morphology
The characterization and the description of Fusarium isolates was conducted using macro and micromorphological features as described by Leslie et al. [25]. Single conidia colonies of the 23 representative isolates were grown on PDA for 10 days. Colony growth and macromorphological features were determined by placing agar plugs (5 mm), taken from the edge of actively growing cultures, on PDA plates incubated at 25±1°C, under a 12/12 h near UV light, for 7 days [66]. All the isolates were transferred on carnation leaf agar (CLA) plates [72], and incubated at 25±1°C under a 12/12h near UV light for 14 days to induce sporulation. Micromorphological features were observed and 50 random measurements of macroconidia, microconidia, conidiogenous cells and chlamydospores were done for each isolate at 40X magnification with a Leica DM2500 microscope. The observations were made by placing the plates directly under the microscope. Measurements were reported as mean value, standard deviation, minimum, and maximum values.
Author Contributions
Conceptualization, M.M and M.S.; methodology, M.S.; formal analysis, M.S. V.G. I.M.; resources, M.M.; data curation, M.S. I.M.; writing—original draft preparation, M.S.; writing—review and editing, M.S. M.M. I.M. V.G..; supervision, M.M., V.G.; funding acquisition, M.M.
Funding
This research was funded by AGROINNOVA DIAGNOSTICS Laboratory. www.agroinnova.unito.it
Data Availability Statement
Not applicable.
Acknowledgments
Authors thank CAPAC Soc. Coop. Agricola, Torino, Italy for providing the seed samples and dr. Marco Troisi, CORTEVA Agriscience, Italy, for providing the maize seedlings.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Erenstein, O.; Jaleta, M.; Sonder, K.; Mottaleb, K.; Prasanna, B. Global maize production, consumption and trade: trends and R&D implications. Food Secur. 2022, 14, 1295–1319, . [CrossRef]
- FAOSTAT. https://www.fao.org/faostat/en/%3F%23data#data/QCL/visualize (accessed 2023-07-21).
- Coltivazioni : Cereali, legumi, radici bulbi e tuberi. http://dati.istat.it/Index.aspx?QueryId=33702 (accessed 2023-07-21).
- Munkvold, G.; White, D. Compendium of Corn Diseases.; AACC International, 2016.
- Ma, L.-J.; Geiser, D. M.; Proctor, R. H.; Rooney, A. P.; O’Donnell, K.; Trail, F.; Gardiner, D. M.; Manners, J. M.; Kazan, K. Fusarium Pathogenomics. Annu. Rev. Microbiol. 2013, 67, 399–416.
- Oldenburg, E.; Höppner, F.; Ellner, F.; Weinert, J. Fusarium diseases of maize associated with mycotoxin contamination of agricultural products intended to be used for food and feed. Mycotoxin Res. 2017, 33, 167–182, . [CrossRef]
- Logrieco, A.; Bottalico, A.; Mule, G.; Moretti, A.; Perrone, G. Epidemiology of toxigenic fungi and their associated mycotoxins for some Mediterranean crops. Eur. J. Plant Pathol. 2003, 109, 645–667. [CrossRef]
- Zargaryan, N.; Kekalo, A.; Nemchenko, V. Infection of grain crops with fungi of the genus Fusarium. BIO Web Conf. 2021, 36, 04008, . [CrossRef]
- Desjardins, A. E. Fusarium Mycotoxins: Chemistry, Genetics, and Biology.; American Phytopathological Society (APS Press), 2006.
- Leyva-Madrigal, K.Y.; Larralde-Corona, C.P.; Apodaca-Sánchez, M.A.; Quiroz-Figueroa, F.R.; Mexia-Bolaños, P.A.; Portillo-Valenzuela, S.; Ordaz-Ochoa, J.; Maldonado-Mendoza, I.E. Fusarium Species from the Fusarium fujikuroi Species Complex Involved in Mixed Infections of Maize in Northern Sinaloa, Mexico. J. Phytopathol. 2014, 163, 486–497, . [CrossRef]
- Duan, C.; Qin, Z.; Yang, Z.; Li, W.; Sun, S.; Zhu, Z.; Wang, X. Identification of Pathogenic Fusarium spp. Causing Maize Ear Rot and Potential Mycotoxin Production in China. Toxins 2016, 8, 186, . [CrossRef]
- Wilke, A.L.; Bronson, C.R.; Tomas, A.; Munkvold, G.P. Seed Transmission of Fusarium verticillioides in Maize Plants Grown Under Three Different Temperature Regimes. Plant Dis. 2007, 91, 1109–1115, . [CrossRef]
- Kedera, C. J.; Leslie, J. F.; Claflin, L. E. Systemic Infection of Corn by Fusarium Moniliforme. Phytopathology 1992, 82, 1138.
- McGee, D.C.; Carlton, W.M.; Opoku, J.; Kleczewski, N.M.; Hamby, K.A.; Herbert, D.A.; Malone, S.; Mehl, H.L.; Morales, L.; Marino, T.P.; et al. Importance of Different Pathways for Maize Kernel Infection by Fusarium moniliforme. Phytopathology® 1997, 87, 209–217, . [CrossRef]
- Okello, P. N.; Petrović, K.; Kontz, B.; Mathew, F. M. Eight Species of Fusarium Cause Root Rot of Corn (Zea Mays) in South Dakota. Plant Health Prog. 2019, 20 (1), 38–43.
- Yilmaz, N.; Sandoval-Denis, M.; Lombard, L.; Visagie, C.; Wingfield, B.; Crous, P. Redefining species limits in the Fusarium fujikuroi species complex. Persoonia - Mol. Phylogeny Evol. Fungi 2021, 46, 129–162, . [CrossRef]
- Lombard, L.; Sandoval-Denis, M.; Lamprecht, S. C.; Crous, P. W. Epitypification of Fusarium Oxysporum–Clearing the Taxonomic Chaos. Persoonia-Mol. Phylogeny Evol. Fungi 2019, 43 (1), 1–47.
- Wang, M.; Crous, P.; Sandoval-Denis, M.; Han, S.; Liu, F.; Liang, J.; Duan, W.; Cai, L. Fusarium and allied genera from China: species diversity and distribution. Persoonia - Mol. Phylogeny Evol. Fungi 2022, 48, 1–53, . [CrossRef]
- Han, S.; Wang, M.; Ma, Z.; Raza, M.; Zhao, P.; Liang, J.; Gao, M.; Li, Y.; Wang, J.; Hu, D.; et al. Fusarium diversity associated with diseased cereals in China, with an updated phylogenomic assessment of the genus. Stud. Mycol. 2023, 104, 87–148, . [CrossRef]
- Geiser, D.M.; Jiménez-Gasco, M.d.M.; Kang, S.; Makalowska, I.; Veeraraghavan, N.; Ward, T.J.; Zhang, N.; Kuldau, G.A.; O'Donnell, K. FUSARIUM-ID v. 1.0: A DNA Sequence Database for Identifying Fusarium. Eur. J. Plant Pathol. 2004, 110, 473–479, . [CrossRef]
- O’donnell, K.; Ward, T.J.; Robert, V.A.R.G.; Crous, P.W.; Geiser, D.M.; Kang, S. DNA sequence-based identification of Fusarium: Current status and future directions. Phytoparasitica 2015, 43, 583–595, . [CrossRef]
- O’donnell, K.; Whitaker, B.K.; Laraba, I.; Proctor, R.H.; Brown, D.W.; Broders, K.; Kim, H.-S.; McCormick, S.P.; Busman, M.; Aoki, T.; et al. DNA Sequence-Based Identification ofFusarium: A Work in Progress. Plant Dis. 2022, 106, 1597–1609, . [CrossRef]
- Leslie, J. F. Introductory Biology of Fusarium Moniliforme. In Fumonisins in Food; Jackson, L. S., DeVries, J. W., Bullerman, L. B., Eds.; Advances in Experimental medicine and Biology; Springer US: Boston, MA, 1996; pp 153–164. [CrossRef]
- O’donnell, K.; Nirenberg, H.I.; Aoki, T.; Cigelnik, E. A Multigene phylogeny of the Gibberella fujikuroi species complex: Detection of additional phylogenetically distinct species. Mycoscience 2000, 41, 61–78, . [CrossRef]
- Leslie, J. F.; Summerell, B. A. The Fusarium Laboratory Manual; John Wiley & Sons, 2008.
- Murillo-Williams, A.; Munkvold, G.P. Systemic Infection by Fusarium verticillioides in Maize Plants Grown Under Three Temperature Regimes. Plant Dis. 2008, 92, 1695–1700, . [CrossRef]
- Nezhad, A. S.; Nourollahi, K. Population Genetic Structure of Fusarium Verticillioides the Causal Agent of Corn Crown and Root Rot in Ilam Province Using Microsatellite Markers. J. Crop Prot. 2020, 9 (1), 157–170.
- Bugnicourt, F. Une Espèce Fusarienne Nouvelle, Parasite Du Riz. Rev. Génerale Bot. 1952, 59, 13–18.
- Parra, M..; Gómez, J.; Aguilar, F.W.; Martinez, J.A. Fusarium annulatum causes Fusarium rot of cantaloupe melons in Spain. Phytopathol. Mediterr. 2022, 16, 269–277, . [CrossRef]
- Mirghasempour, S.A.; Studholme, D.J.; Chen, W.; Cui, D.; Mao, B. Identification and Characterization of Fusarium nirenbergiae Associated with Saffron Corm Rot Disease. Plant Dis. 2022, 106, 486–495, . [CrossRef]
- Özer, G.; Paulitz, T.C.; Imren, M.; Alkan, M.; Muminjanov, H.; Dababat, A.A. Identity and Pathogenicity of Fungi Associated with Crown and Root Rot of Dryland Winter Wheat in Azerbaijan. Plant Dis. 2020, 104, 2149–2157, . [CrossRef]
- O'Donnell, K.; Cigelnik, E.; Nirenberg, H.I. Molecular systematics and phylogeography of theGibberella fujikuroispecies complex. Mycologia 1998, 90, 465–493, . [CrossRef]
- Wulff, E.G.; Sørensen, J.L.; Lübeck, M.; Nielsen, K.F.; Thrane, U.; Torp, J. Fusarium spp. associated with rice Bakanae: ecology, genetic diversity, pathogenicity and toxigenicity. Environ. Microbiol. 2010, 12, 649–657, . [CrossRef]
- Husna, A.; Zakaria, L.; Nor, N.M.I.M. Fusarium commune associated with wilt and root rot disease in rice. Plant Pathol. 2020, 70, 123–132, . [CrossRef]
- Mezzalama, M.; Guarnaccia, V.; Martino, I.; Tabone, G.; Gullino, M.L. First Report of Fusarium commune Causing Root and Crown Rot on Maize in Italy. Plant Dis. 2021, 105, 4156–4156, . [CrossRef]
- Xi, K.; Haseeb, H.A.; Shan, L.; Guo, W.; Dai, X. First Report of Fusarium commune Causing Stalk Rot on Maize in Liaoning Province, China. Plant Dis. 2019, 103, 773–773, . [CrossRef]
- Skovgaard, K.; Rosendahl, S.; O'Donnell, K.; Nirenberg, H.I. Fusarium commune Is a New Species Identified by Morphological and Molecular Phylogenetic Data. Mycologia 2003, 95, 630–636, . [CrossRef]
- Dean, R.; Van Kan, J. A.; Pretorius, Z. A.; Hammond-Kosack, K. E.; Di Pietro, A.; Spanu, P. D.; Rudd, J. J.; Dickman, M.; Kahmann, R.; Ellis, J. The Top 10 Fungal Pathogens in Molecular Plant Pathology. Mol. Plant Pathol. 2012, 13 (4), 414–430.
- Laurence, M.H.; Walsh, J.L.; Shuttleworth, L.A.; Robinson, D.M.; Johansen, R.M.; Petrovic, T.; Vu, T.T.H.; Burgess, L.W.; Summerell, B.A.; Liew, E.C.Y. Six novel species of Fusarium from natural ecosystems in Australia. Fungal Divers. 2015, 77, 349–366, . [CrossRef]
- Maymon, M.; Sharma, G.; Hazanovsky, M.; Erlich, O.; Pessach, S.; Freeman, S.; (Lahkim), L.T. Characterization of Fusarium population associated with wilt of jojoba in Israel. Plant Pathol. 2021, 70, 793–803, . [CrossRef]
- Aiello, D.; Fiorenza, A.; Leonardi, G.R.; Vitale, A.; Polizzi, G. Fusarium nirenbergiae (Fusarium oxysporum Species Complex) Causing the Wilting of Passion Fruit in Italy. Plants 2021, 10, 2011, . [CrossRef]
- Maryani, N.; Lombard, L.; Poerba, Y.S.; Subandiyah, S.; Crous, P.W.; Kema, G.H.J. Phylogeny and genetic diversity of the banana Fusarium wilt pathogen Fusarium oxysporum f. sp. cubense in the Indonesian centre of origin. Stud. Mycol. 2018, 92, 155–194, . [CrossRef]
- Summerell, B.A. Resolving Fusarium: Current Status of the Genus. Annu. Rev. Phytopathol. 2019, 57, 323–339, . [CrossRef]
- Crous, P.; Lombard, L.; Sandoval-Denis, M.; Seifert, K.; Schroers, H.-J.; Chaverri, P.; Gené, J.; Guarro, J.; Hirooka, Y.; Bensch, K.; et al. Fusarium: more than a node or a foot-shaped basal cell. Stud. Mycol. 2021, 98, 100116, . [CrossRef]
- Moparthi, S.; Burrows, M.E.; Mgbechi-Ezeri, J.; Agindotan, B. Fusarium spp. Associated With Root Rot of Pulse Crops and Their Cross-Pathogenicity to Cereal Crops in Montana. Plant Dis. 2021, 105, 548–557, . [CrossRef]
- Gaige, A.R.; Todd, T.; Stack, J.P. Interspecific Competition for Colonization of Maize Plants Between Fusarium proliferatum and Fusarium verticillioides. Plant Dis. 2020, 104, 2102–2110, . [CrossRef]
- Xi, K.; Shan, L.; Yang, Y.; Zhang, G.; Zhang, J.; Guo, W. Species Diversity and Chemotypes of Fusarium Species Associated With Maize Stalk Rot in Yunnan Province of Southwest China. Front. Microbiol. 2021, 12, . [CrossRef]
- Summerell, B.A.; Salleh, B.; Leslie, J.F.; Felix, S.V.; Valenzuela, V.; Ortega, P.; Fierros, G.; Rojas, P.; Orona, C.A.L.; Manjarrez, J.E.R.; et al. A Utilitarian Approach to Fusarium Identification. Plant Dis. 2003, 87, 117–128, . [CrossRef]
- Warham, E. J.; Butler, L. D.; Sutton, B. C. Seed Testing of Maize and Wheat: A Laboratory Guide; CIMMYT, 1996.
- Bilgi, V.N.; Bradley, C.A.; Khot, S.D.; Grafton, K.F.; Rasmussen, J.B. Response of Dry Bean Genotypes to Fusarium Root Rot, Caused byFusarium solanif. sp.phaseoli, Under Field and Controlled Conditions. Plant Dis. 2008, 92, 1197–1200, . [CrossRef]
- Acharya, B.; Lee, S.; Mian, M.A.R.; Jun, T.-H.; McHale, L.K.; Michel, A.P.; Dorrance, A.E. Identification and mapping of quantitative trait loci (QTL) conferring resistance to Fusarium graminearum from soybean PI 567301B. Theor. Appl. Genet. 2015, 128, 827–838, . [CrossRef]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 9, 2044–2049, . [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808, . [CrossRef]
- Carbone, I.; Kohn, L. M. A Method for Designing Primer Sets for Speciation Studies in Filamentous Ascomycetes. Mycologia 1999, 91 (3), 553–556.
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330, . [CrossRef]
- O'Donnell, K.; Cigelnik, E. Two Divergent Intragenomic rDNA ITS2 Types within a Monophyletic Lineage of the FungusFusariumAre Nonorthologous. Mol. Phylogenetics Evol. 1997, 7, 103–116, . [CrossRef]
- Guarnaccia, V.; Aiello, D.; Polizzi, G.; Crous, P. W.; Sandoval-Denis, M. Soilborne Diseases Caused by Fusarium and Neocosmospora Spp. on Ornamental Plants in Italy. Phytopathol. Mediterr. 2019, 58 (1), 127–137. [CrossRef]
- Weir, B.S.; Johnston, P.R.; Damm, U. The Colletotrichum gloeosporioides species complex. Stud. Mycol. 2012, 73, 115–180, . [CrossRef]
- O’donnell, K.; Rooney, A.P.; Proctor, R.H.; Brown, D.W.; McCormick, S.P.; Ward, T.J.; Frandsen, R.J.; Lysøe, E.; Rehner, S.A.; Aoki, T.; et al. Phylogenetic analyses of RPB1 and RPB2 support a middle Cretaceous origin for a clade comprising all agriculturally and medically important fusaria. Fungal Genet. Biol. 2013, 52, 20–31, . [CrossRef]
- Zhang, Y.; Chen, C.; Mai, Z.; Lin, J.; Nie, L.; Maharachchikumbura, S.S.N.; You, C.; Xiang, M.; Hyde, K.D.; Manawasinghe, I.S. Co-infection of Fusarium aglaonematis sp. nov. and Fusarium elaeidis Causing Stem Rot in Aglaonema modestum in China. Front. Microbiol. 2022, 13, 930790, . [CrossRef]
- Costa, M.M.; Melo, M.P.; Carmo, F.S.; Moreira, G.M.; Guimarães, E.A.; Rocha, F.S.; Costa, S.S.; Abreu, L.M.; Pfenning, L.H. Fusarium species from tropical grasses in Brazil and description of two new taxa. Mycol. Prog. 2021, 20, 61–72, . [CrossRef]
- Lombard, L.; van Doorn, R.; Groenewald, J.; Tessema, T.; Kuramae, E.; Etolo, D.; Raaijmakers, J.; Crous, P. Fusarium diversity associated with the Sorghum-Striga interaction in Ethiopia. Fungal Syst. Evol. 2022, 10, 177–215, . [CrossRef]
- Vermeulen, M.; Rothmann, L.A.; Swart, W.J.; Gryzenhout, M. Fusarium casha sp. nov. and F. curculicola sp. nov. in the Fusarium fujikuroi Species Complex Isolated from Amaranthuscruentus and Three Weevil Species in South Africa. Diversity 2021, 13, 472, . [CrossRef]
- Laraba, I.; Kim, H.-S.; Proctor, R.H.; Busman, M.; O’donnell, K.; Felker, F.C.; Aime, M.C.; Koch, R.A.; Wurdack, K.J. Fusarium xyrophilum, sp. nov., a member of the Fusarium fujikuroi species complex recovered from pseudoflowers on yellow-eyed grass (Xyris spp.) from Guyana. Mycologia 2019, 112, 39–51, . [CrossRef]
- Proctor, R.H.; Van Hove, F.; Susca, A.; Stea, G.; Busman, M.; van der Lee, T.; Waalwijk, C.; Moretti, A.; Ward, T.J. Birth, death and horizontal transfer of the fumonisin biosynthetic gene cluster during the evolutionary diversification of Fusarium. Mol. Microbiol. 2013, 90, 290–306, . [CrossRef]
- Sandoval-Denis, M.; Guarnaccia, V.; Polizzi, G.; Crous, P. Symptomatic Citrus trees reveal a new pathogenic lineage in Fusarium and two new Neocosmospora species. Persoonia - Mol. Phylogeny Evol. Fungi 2018, 40, 1–25, . [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780, . [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874, . [CrossRef]
- Swofford, D.; Sullivan, J. Phylogeny Inference Based on Parsimony and Other Methods with PAUP. Phylogenetic Handb. Pract. Approach Phylogenetic Anal. Hypothesis Test. 2009, 267–312.
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice across a Large Model Space. Syst. Biol. 2012, 61, 539–542, . [CrossRef]
- Nylander, J.A.A.; Ronquist, F.; Huelsenbeck, J.P.; Nieves-Aldrey, J. Bayesian Phylogenetic Analysis of Combined Data. Syst. Biol. 2004, 53, 47–67, . [CrossRef]
- Fisher, N. L.; Burgess, L. W.; Toussoun, T. A.; Nelson, P. E. Carnation Leaves as a Substrate and for Preserving Cultures of Fusarium Species. Phytopathology 1982, 72 (1), 151–153.
Figure 1.
Symptoms caused by Fusarium spp. (A, B) observed in the field and (C, D) after pathogenicity trials on leaves, roots and crowns maize seedlings.
Figure 1.
Symptoms caused by Fusarium spp. (A, B) observed in the field and (C, D) after pathogenicity trials on leaves, roots and crowns maize seedlings.

Figure 2.
Consensus phylogram of 305 trees resulting from a Bayesian analysis of the combined tef1-α, rpb2, calm and tub2 sequences of Fusarium spp. belonging to FFSC. Bayesian posterior probability values and Bootstrap support values are indicated at the nodes. The isolates collected and species found in this study are in red. The tree was rooted to Fusarium foetens (CBS 120665).
Figure 2.
Consensus phylogram of 305 trees resulting from a Bayesian analysis of the combined tef1-α, rpb2, calm and tub2 sequences of Fusarium spp. belonging to FFSC. Bayesian posterior probability values and Bootstrap support values are indicated at the nodes. The isolates collected and species found in this study are in red. The tree was rooted to Fusarium foetens (CBS 120665).
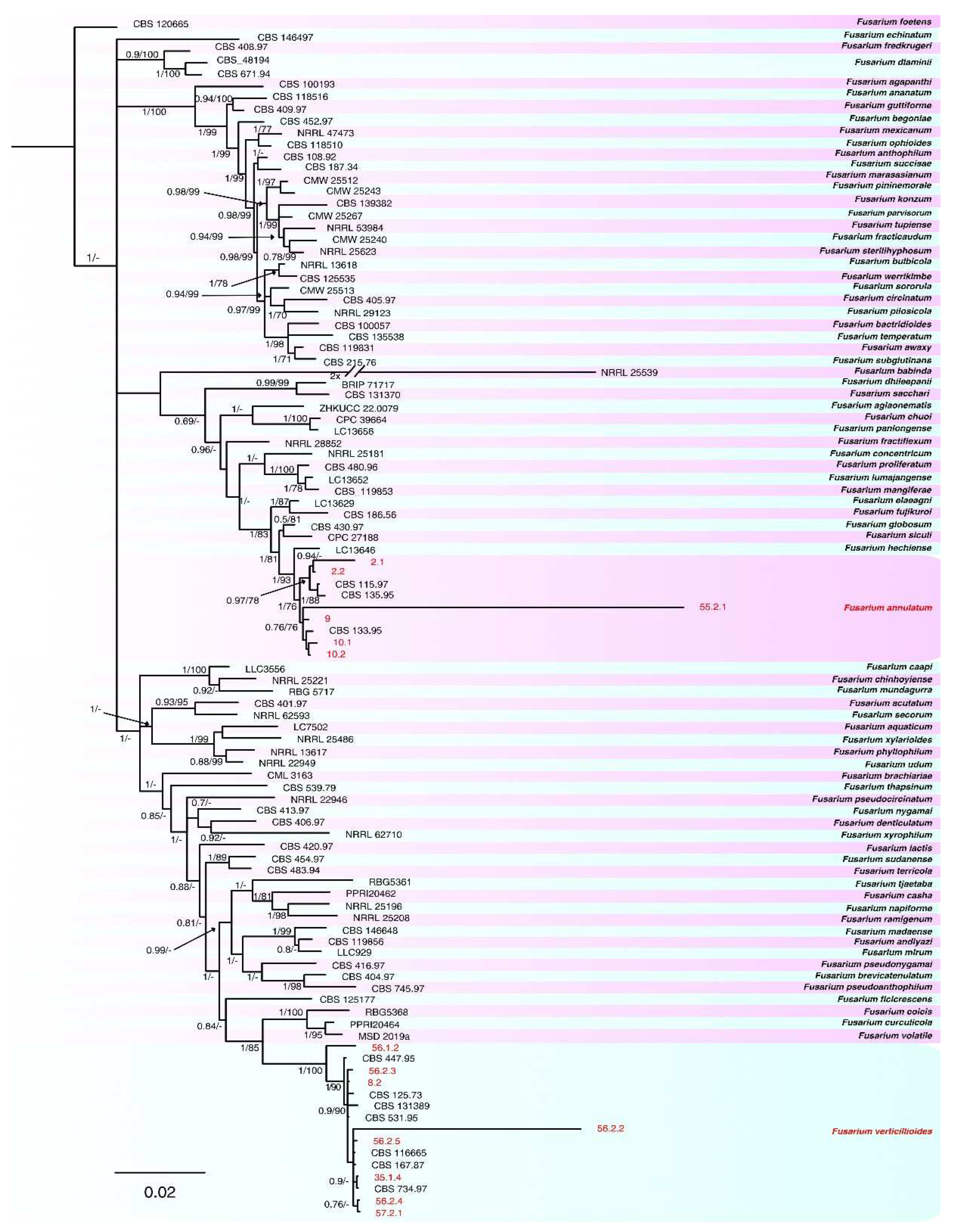
Figure 3.
Consensus phylogram of 226 trees resulting from a Bayesian analysis of the combined tef1-α, rpb2 and calm sequences of Fusarium spp. belonging to FOSC. Bayesian posterior probability values and Bootstrap support values are indicated at the nodes. The isolates collected and species found in this study are in red. The tree was rooted to Fusarium udum (NRRL22949).
Figure 3.
Consensus phylogram of 226 trees resulting from a Bayesian analysis of the combined tef1-α, rpb2 and calm sequences of Fusarium spp. belonging to FOSC. Bayesian posterior probability values and Bootstrap support values are indicated at the nodes. The isolates collected and species found in this study are in red. The tree was rooted to Fusarium udum (NRRL22949).
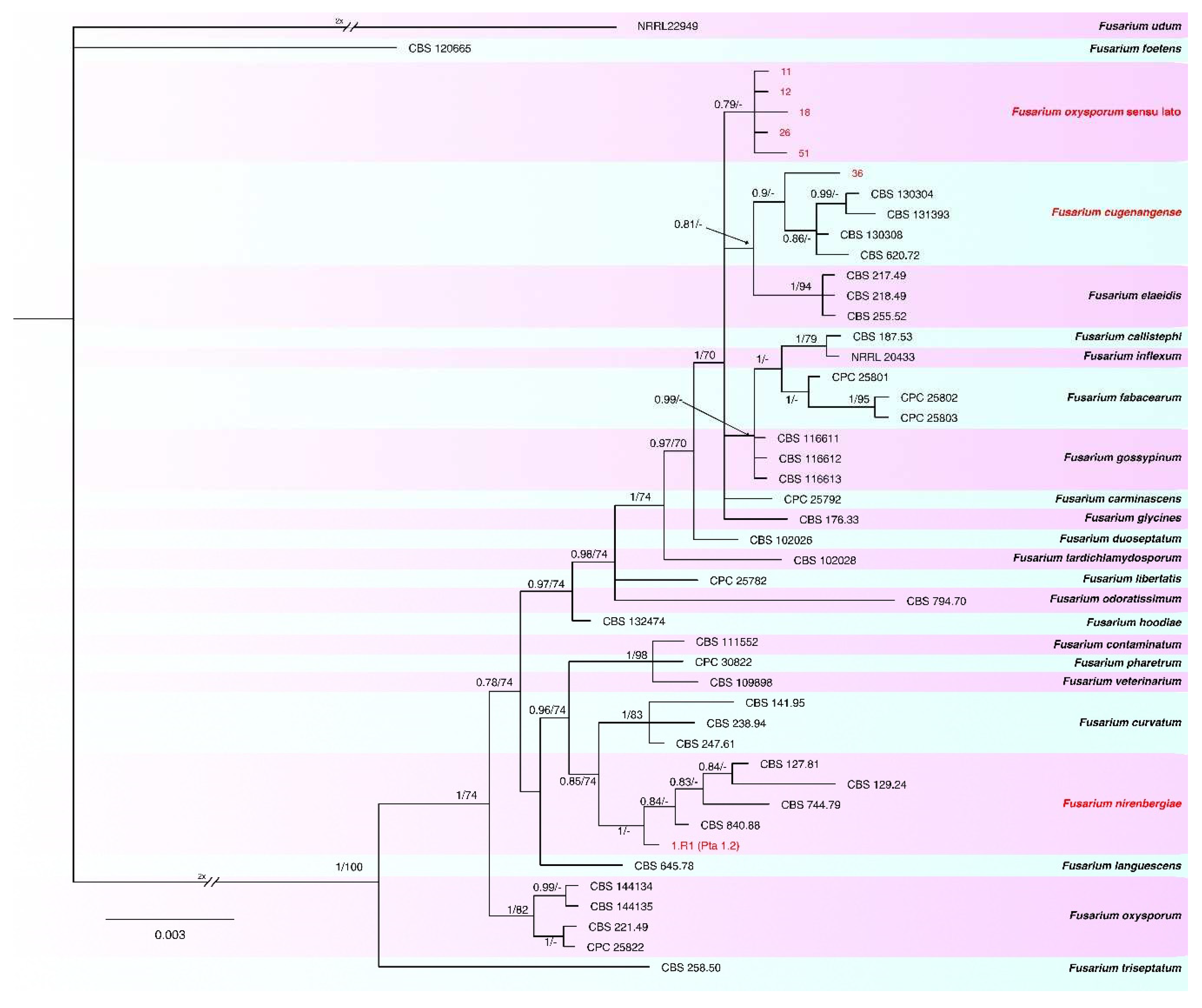
Figure 4.
Consensus phylogram of 301 trees resulting from a Bayesian analysis of the combined tef1-α, rpb2, calm and tub2 sequences of Fusarium spp. belonging to FNSC. Bayesian posterior probability values and Bootstrap support values are indicated at the nodes. The isolates collected and species found in this study are in red. The tree was rooted to Fusarium udum (NRRL22949).
Figure 4.
Consensus phylogram of 301 trees resulting from a Bayesian analysis of the combined tef1-α, rpb2, calm and tub2 sequences of Fusarium spp. belonging to FNSC. Bayesian posterior probability values and Bootstrap support values are indicated at the nodes. The isolates collected and species found in this study are in red. The tree was rooted to Fusarium udum (NRRL22949).
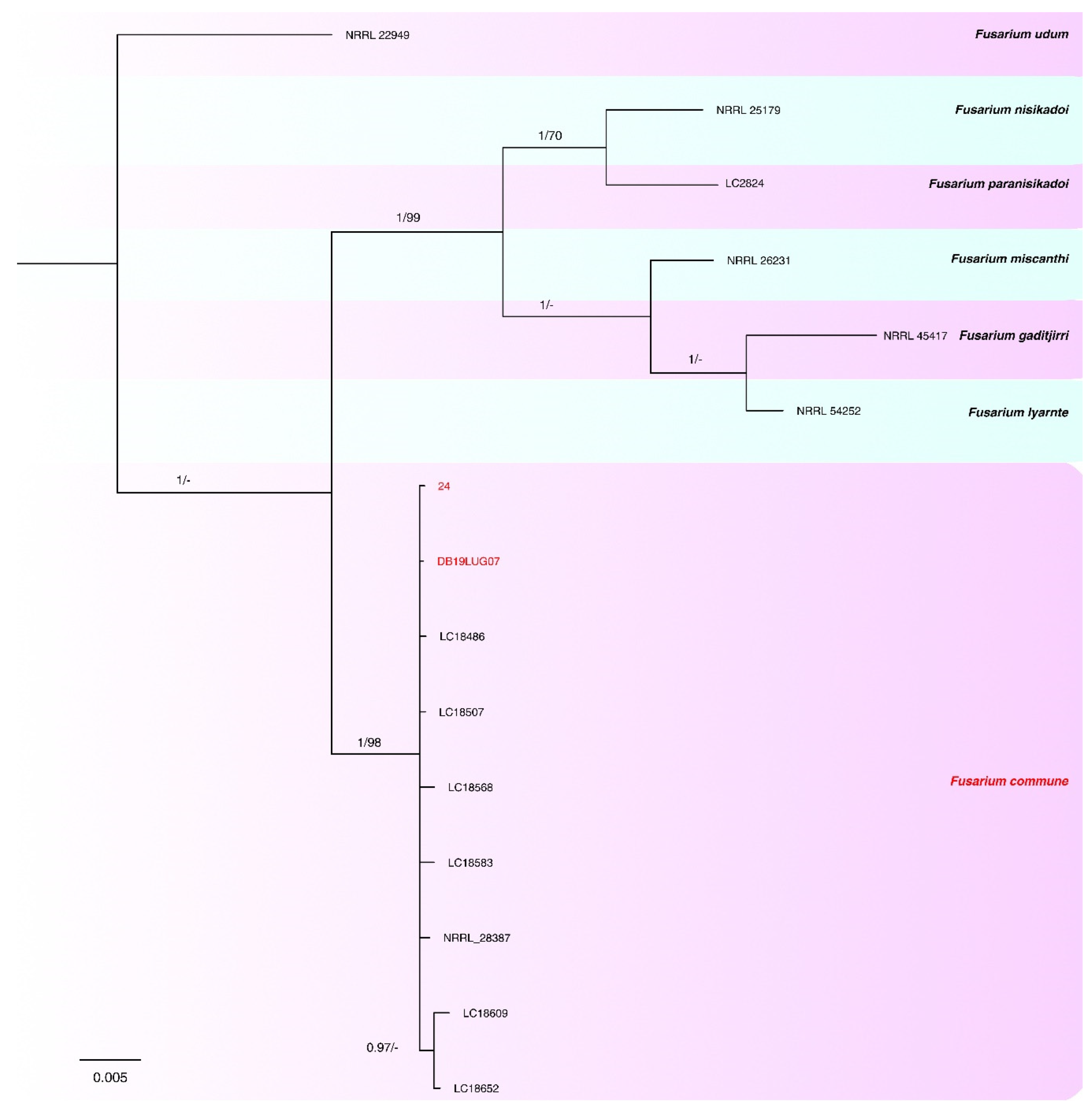
Figure 5.
Morphological features of the species belonging to FFSC identified in this study. (A, B, C, D, E, F) F. verticillioides and (G, H, I, J, K, L) F. annulatum. a-b-g-h. Colonies on PDA above and below; c–d-e-i-j-k. conidia; f-l. conidiogenous cells.— Scale bars = 10 μm.
Figure 5.
Morphological features of the species belonging to FFSC identified in this study. (A, B, C, D, E, F) F. verticillioides and (G, H, I, J, K, L) F. annulatum. a-b-g-h. Colonies on PDA above and below; c–d-e-i-j-k. conidia; f-l. conidiogenous cells.— Scale bars = 10 μm.
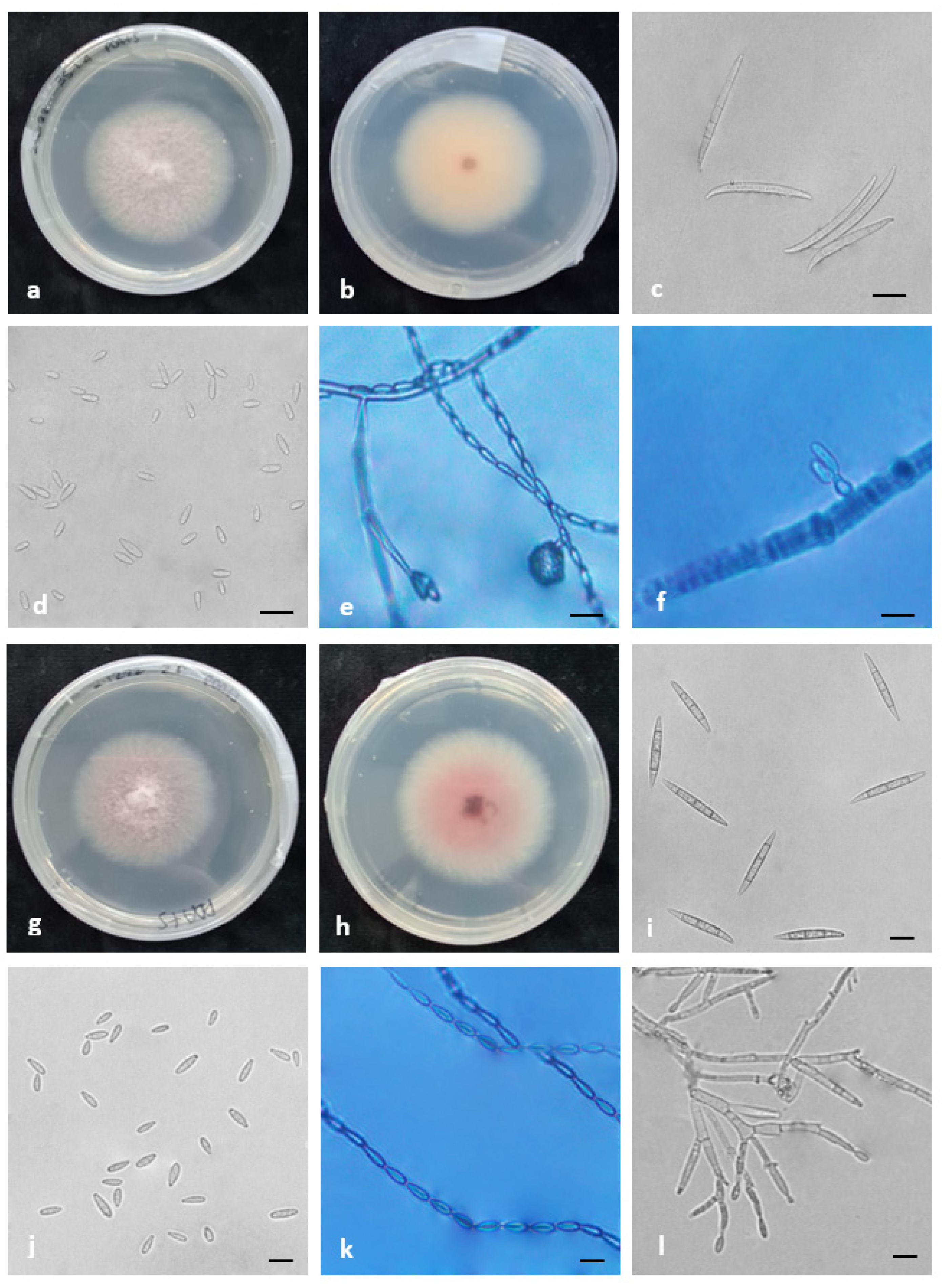
Figure 6.
Morphological features of F. commune, the species belonging to FNSC identified in this study. a-b. Colonies on PDA above and below; c–d. conidia; e. conidiogenous cells; f. chlamydospores.— Scale bars = 10 μm.
Figure 6.
Morphological features of F. commune, the species belonging to FNSC identified in this study. a-b. Colonies on PDA above and below; c–d. conidia; e. conidiogenous cells; f. chlamydospores.— Scale bars = 10 μm.
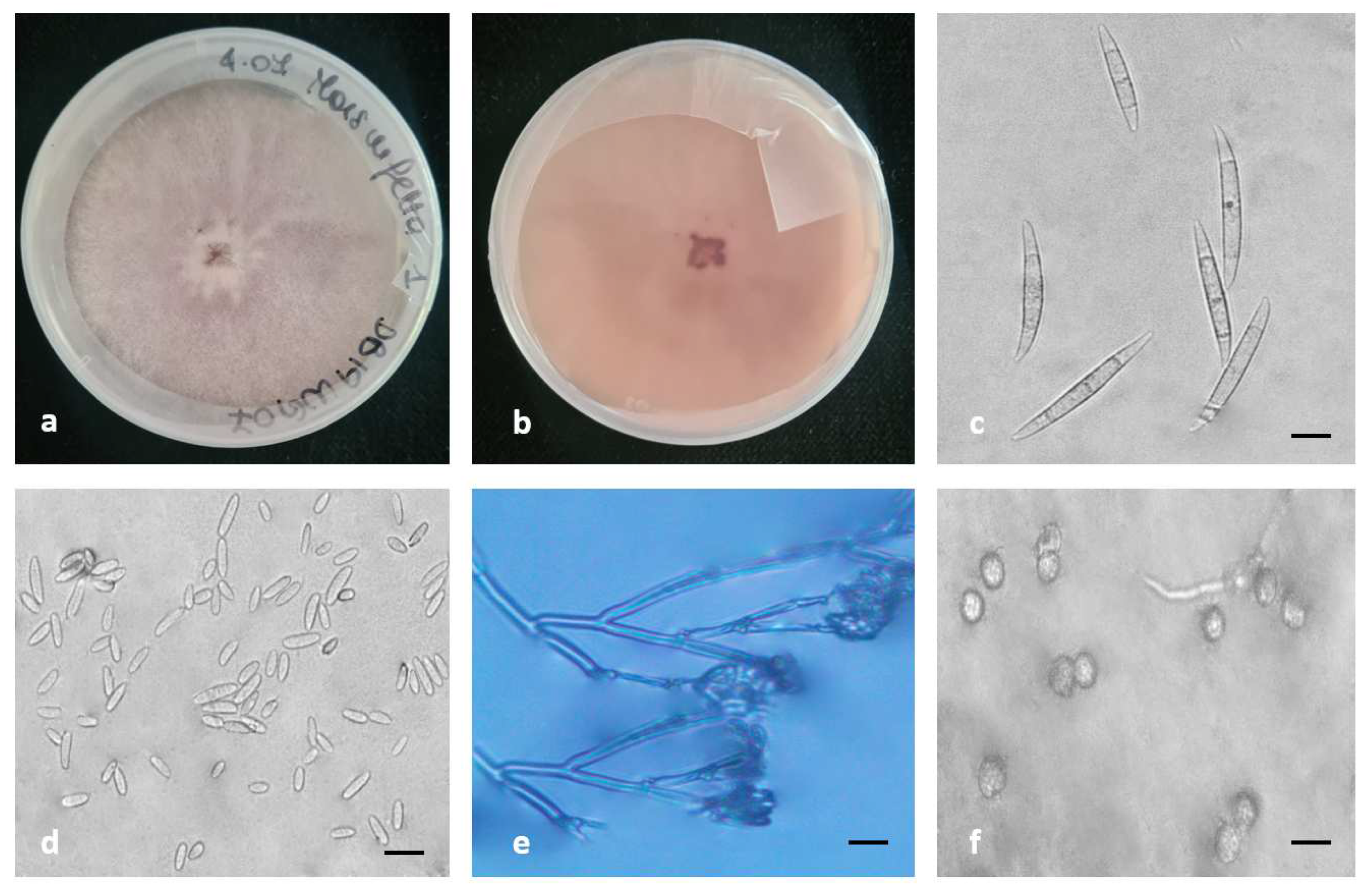

Table 1.
Fusarium spp. isolates used in this study (isolate code, origin of the sample, hybrid, FAO class, symptomatic portion isolated and year of isolation).
Table 1.
Fusarium spp. isolates used in this study (isolate code, origin of the sample, hybrid, FAO class, symptomatic portion isolated and year of isolation).
| Isolate code | Origin | Hybrid | Fao Class | Symptomatic portion | Year of isolation |
|---|---|---|---|---|---|
| DB19LUG07 | San Zenone degli Ezzelini (VI)-Italy | unknown | unknown | Root | 2019 |
| DB19LUG16 | San Zenone degli Ezzelini (VI)-Italy | unknown | unknown | Root | 2019 |
| DB19LUG20 | San Zenone degli Ezzelini (VI)-Italy | unknown | unknown | Root | 2019 |
| DB19LUG25 | San Zenone degli Ezzelini (VI)-Italy | unknown | unknown | Root | 2019 |
| 2.1 | Livorno Ferraris (VC)-Italy | P1547 | 600-130 days | Root | 2019 |
| 2.2 | Livorno Ferraris (VC)-Italy | P1547 | 600-130 days | Root | 2019 |
| 8.1 | Cigliano (VC)-Italy | - | - | Root | 2019 |
| 8.2 | Cigliano (VC)-Italy | - | - | Root | 2019 |
| 9 | USA | PR32B10 | 600-132 days | Seed | 2019 |
| 10.1 | France | P0423 | 400-116 days | Seed | 2019 |
| 10.2 | France | P0423 | 400-116 days | Seed | 2019 |
| 11 | Italy | unknown | unknown | Seed | 2019 |
| 12 | Italy | SY ANTEX | 600-130 days | Seed | 2019 |
| 18 | Turkey | DKC6752 | 600-128 days | Seed | 2019 |
| 19 | Romania | DKC5830 | 500-x days | Seed | 2019 |
| 21 | Crescentino (VC)-Italy | P1547 | 600-130 days | Stem | 2019 |
| 23 | Crescentino (VC)-Italy | P1547 | 600-130 days | Root | 2019 |
| 24 | Crescentino (VC)-Italy | P1916 | 600-130 days | Root | 2019 |
| 26 | Crescentino (VC)-Italy | P1916 | 600-130 days | Stem | 2019 |
| 28 | Crescentino (VC)-Italy | P1916 | 600-130 days | Root | 2019 |
| 29 | Cigliano (VC)-Italy | P1517W | 600-128 days | Root | 2019 |
| 30 | Cigliano (VC)-Italy | P1517W | 600-128 days | Root | 2019 |
| 31 | Cigliano (VC)-Italy | P1517W | 600-128 days | Stem | 2019 |
| 32 | Cigliano (VC)-Italy | P1517W | 600-128 days | Stem | 2019 |
| 35.1.4 | Cigliano (VC)-Italy | P1517W | 600-128 days | Root | 2019 |
| 36 | Cigliano (VC)-Italy | P1517W | 600-128 days | Stem | 2019 |
| 40 | Cigliano (VC)-Italy | P1517W | 600-128 days | Root | 2019 |
| 41 | Cigliano (VC)-Italy | P1547 | 600-130 days | Root | 2019 |
| 44 | Cigliano (VC)-Italy | P1547 | 600-130 days | Root | 2019 |
| 50 | Cigliano (VC)-Italy | P1547 | 600-130 days | Root | 2019 |
| 51 | Cigliano (VC)-Italy | unknown | unknown | Stem | 2019 |
| 55.2.1 | Cigliano (VC)-Italy | unknown | unknown | Crown | 2019 |
| 56.1.2 | Cigliano (VC)-Italy | unknown | unknown | Root | 2019 |
| 56.2.2 | Cigliano (VC)-Italy | unknown | unknown | Root | 2019 |
| 56.2.3 | Cigliano (VC)-Italy | unknown | unknown | Root | 2019 |
| 56.2.4 | Cigliano (VC)-Italy | unknown | unknown | Root | 2019 |
| 56.2.5 | Cigliano (VC)-Italy | unknown | unknown | Root | 2019 |
| 57.2.1 | Cigliano (VC)-Italy | unknown | unknown | Root | 2019 |
| 1.RI (Pta 1.1) | San Zenone degli Ezzelini (VI)-Italy | unknown | unknown | Crown | 2020 |
| 1.RI (Pta 1.2) | San Zenone degli Ezzelini (VI)-Italy | unknown | unknown | Crown | 2020 |
| 1.RII (Pta 3.2) | San Zenone degli Ezzelini (VI)-Italy | unknown | unknown | Crown | 2020 |
Table 2.
Results of pathogenicity test performed on the 41 Fusarium isolates isolated, at 14 days. Number of plants recorded for each index and disease index (0–100) of each isolate (ANOVA and Duncan p < 0.05%).
Table 2.
Results of pathogenicity test performed on the 41 Fusarium isolates isolated, at 14 days. Number of plants recorded for each index and disease index (0–100) of each isolate (ANOVA and Duncan p < 0.05%).
| ID Sample | Severity index of root and crown rot (number of plant) | Disease index | |||||||
| 1 | 2 | 3 | 4 | 5 | (DI) 0-100 | ||||
| DB19LUG07 | 0 | 3 | 3 | 0 | 0 | 50.0 | abcde | ||
| DB19LUG16 | 0 | 6 | 0 | 0 | 0 | 40.0 | cdefg | ||
| DB19LUG20 | 4 | 2 | 0 | 0 | 0 | 13.3 | gh | ||
| DB19LUG25 | 3 | 3 | 0 | 0 | 0 | 20.0 | gh | ||
| 2.1 | 0 | 0 | 0 | 4 | 2 | 86.7 | a | ||
| 2.2 | 0 | 0 | 0 | 3 | 3 | 90.0 | a | ||
| 8.1 | 6 | 0 | 0 | 0 | 0 | 0.0 | h | ||
| 8.2 | 3 | 3 | 0 | 0 | 0 | 20.0 | fgh | ||
| 9 | 0 | 0 | 0 | 6 | 0 | 80.0 | ab | ||
| 10.1 | 0 | 0 | 2 | 0 | 4 | 86.7 | a | ||
| 10.2 | 0 | 0 | 1 | 2 | 3 | 86.7 | a | ||
| 11 | 3 | 3 | 0 | 0 | 0 | 20.0 | efgh | ||
| 12 | 0 | 3 | 0 | 0 | 3 | 70.0 | abc | ||
| 18 | 2 | 3 | 0 | 0 | 1 | 36.7 | efgh | ||
| 19 | 6 | 0 | 0 | 0 | 0 | 0.0 | h | ||
| 21 | 2 | 4 | 0 | 0 | 0 | 26.7 | efgh | ||
| 23 | 2 | 4 | 0 | 0 | 0 | 26.7 | efgh | ||
| 24 | 0 | 3 | 0 | 0 | 3 | 70.0 | abc | ||
| 26 | 0 | 4 | 2 | 0 | 0 | 46.7 | bcdef | ||
| 28 | 3 | 3 | 0 | 0 | 0 | 20.0 | efgh | ||
| 29 | 0 | 6 | 0 | 0 | 0 | 40.0 | cdefg | ||
| 30 | 3 | 3 | 0 | 0 | 0 | 20.0 | efgh | ||
| 31 | 2 | 4 | 0 | 0 | 0 | 26.7 | efgh | ||
| 32 | 4 | 2 | 0 | 0 | 0 | 13.3 | gh | ||
| 35.1.4 | 0 | 1 | 1 | 2 | 2 | 76.7 | abc | ||
| 36 | 3 | 3 | 0 | 0 | 0 | 20.0 | efgh | ||
| 40 | 0 | 4 | 2 | 0 | 0 | 46.7 | bcdef | ||
| 41 | 6 | 0 | 0 | 0 | 0 | 0.0 | h | ||
| 44 | 6 | 0 | 0 | 0 | 0 | 0.0 | h | ||
| 50 | 6 | 0 | 0 | 0 | 0 | 0.0 | h | ||
| 51 | 2 | 2 | 2 | 0 | 0 | 33.3 | defgh | ||
| 55.2.1 | 0 | 1 | 1 | 2 | 2 | 76.7 | abc | ||
| 56.1.2 | 0 | 0 | 0 | 4 | 2 | 86.7 | a | ||
| 56.2.2 | 0 | 0 | 2 | 2 | 2 | 80.0 | ab | ||
| 56.2.3 | 0 | 0 | 0 | 3 | 3 | 90.0 | a | ||
| 56.2.4 | 0 | 0 | 2 | 2 | 2 | 80.0 | ab | ||
| 56.2.5 | 0 | 0 | 2 | 4 | 0 | 73.3 | abc | ||
| 57.2.1 | 0 | 0 | 0 | 4 | 2 | 86.7 | a | ||
| 1.RI (Pta 1.1) | 2 | 2 | 0 | 0 | 2 | 46.7 | cdefg | ||
| 1.RI (Pta 1.2) | 0 | 2 | 2 | 0 | 2 | 66.7 | abcd | ||
| 1.RII (Pta 3.2) | 3 | 3 | 0 | 0 | 0 | 20.0 | efgh | ||
| Healthy control | 6 | 0 | 0 | 0 | 0 | 0.0 | h | ||
Table 3.
Origin, host, culture and sequence GenBank accession number of Fusarium isolates used and generated in this study. Newly generated accession numbers are in bold.
Table 3.
Origin, host, culture and sequence GenBank accession number of Fusarium isolates used and generated in this study. Newly generated accession numbers are in bold.
| Species | Complex | Collection | Host | Origin | tef1-α | rpb2 | calm | tub2 | Reference |
|---|---|---|---|---|---|---|---|---|---|
| F. acutatum | FFSC | CBS 401.97 | Cajanus cajan | India | MW402124 | MW402813 | MW402458 | MW402322 | Yilmaz et al. (2021) |
| F. agapanthi | FFSC | CBS 100193 | Agapanthus praecox | New Zealand | MW401959 | MW402727 | MW402363 | MW402160 | Yilmaz et al. (2021) |
| F. aglaonematis | FFSC | ZHKUCC 22-0079 | Aglaonema modestum | China | ON330439 | ON330445 | ON330436 | ON330442 | Zhang et al. (2022) |
| F. ananatum | FFSC | CBS 118516 | Ananas comosus | South Africa | LT996091 | LT996137 | MW402376 | MN534089 | Yilmaz et al. (2021) |
| F. andiyazi | FFSC | CBS 119856 | Sorghum grain | Ethiopia | MN533989 | MN534286 | MN534174 | MN534081 | Yilmaz et al. (2021) |
| F. annulatum | FFSC | CBS 115.97 | Dianthus caryophyllus | Italy | MW401973 | MW402785 | MW402373 | MW402173 | Yilmaz et al. (2021) |
| F. annulatum | FFSC | CBS 133.95 | Dianthus caryophyllus | The Netherlands | MW402040 | MW402743 | MW402407 | MW402239 | Yilmaz et al. (2021) |
| F. annulatum | FFSC | CBS 135.95 | Dianthus caryophyllus | The Netherlands | MW402043 | MW402745 | MW402408 | MW402242 | Yilmaz et al. (2021) |
| F. annulatum | FFSC | 2.1 | Zea mays | Italy | OR565982 | OR566043 | OR566020 | OR566004 | This study |
| F. annulatum | FFSC | 2.2 | Zea mays | Italy | OR565983 | OR566044 | OR566021 | OR566005 | This study |
| F. annulatum | FFSC | 9 | Zea mays | Italy | OR565984 | OR566045 | OR566022 | OR566006 | This study |
| F. annulatum | FFSC | 10.1 | Zea mays | Italy | OR565985 | OR566046 | OR566023 | OR566007 | This study |
| F. annulatum | FFSC | 10.2 | Zea mays | Italy | OR565986 | OR566047 | OR566024 | OR566008 | This study |
| F. annulatum | FFSC | 55 | Zea mays | Italy | OR565987 | OR566048 | OR566025 | OR566009 | This study |
| F. anthophilum | FFSC | CBS 108.92 | Hippeastrum leaf | The Netherlands | MW401965 | MW402783 | MW402368 | MW402166 | Yilmaz et al. (2021) |
| F. aquaticum | FFSC | LC7502 | Water | China | MW580448 | MW474394 | MW566275 | MW533730 | Wang (2022) |
| F. awaxy | FFSC | CBS 119831 | Environmental | New Guinea | MN534056 | MN534237 | MN534167 | MN534108 | Yilmaz et al. (2021) |
| F. babinda | FFSC | NRRL 25539 | Unknown | Unknown | KU171718 | KU171698 | KU171418 | KU171778 | O' Donnell et al. (2013) |
| F. bactridioides | FFSC | CBS 100057 | Cronartium conigenum on Pinus leiophylla | USA | MN533993 | MN534235 | MN534173 | MN534112 | Yilmaz et al. (2021) |
| F. begoniae | FFSC | CBS 452.97 | Begonia elatior hybrid | Germany | MN533994 | MN534243 | MN534163 | MN534101 | Yilmaz et al. (2021) |
| F. braichiariae | FFSC | CML 3163 | Brachiaria decumbens | Brazil | MT901349 | MT901315 | - | MT901322 | Moreira Costa et al. (2021) |
| F. brevicatenulatum | FFSC | CBS 404.97 | Striga asiatica | Madagascar | MN533995 | MN534295 | MT010979 | MN534063 | Yilmaz et al. (2021) |
| F. bulbicola | FFSC | NRRL13618 | Nerine bowdenii bulb | The Netherlands | KF466415 | MW402767 | MW402450 | KF466437 | Yilmaz et al. (2021) |
| F. caapi | FFSC | LLC3556 | Sorghum | Ethiopia | OP486950 | OP486519 | OP485837 | - | Lombard et al. (2022) |
| F. callistephi | FOSC | CBS 187.53 | Callistephus chinensis | The Netherlands | MH484966 | MH484875 | MH484693 | MH485057 | Lombard et al. (2019) |
| F. carminascens | FOSC | CPC 25792 | Zea mays | South Africa | MH485025 | MH484934 | MH484752 | MH485116 | Lombard et al. (2019) |
| F. casha | FFSC | PPRI20462 | Amaranthus cruentus | South Africa | MF787262 | - | - | MF787256 | Vermeulen et al. (2021) |
| F. chinhoyiense | FFSC | NRRL 25221 | Zea mays | Zimbabwe | MN534050 | MN534262 | MN534196 | MN534082 | Yilmaz et al. (2021) |
| F. chuoi | FFSC | CPC 39664 | Unknown | Unknown | OK626308 | OK626302 | OK626304 | OK626310 | Yilmaz et al. (2021) |
| F. circinatum | FFSC | CBS 405.97 | Pinus radiata | USA | MN533997 | MN534252 | MN534199 | MN534097 | Yilmaz et al. (2021) |
| F. coicis | FFSC | RBG5368 | Coix gasteenii | Australia | KP083251 | KP083274 | LT996178 | LT996115 | Laurence et al. (2015) |
| F. commune | FNSC | NRRL 28387 | Dianthus caryophyllus | The Netherlands | HM057338 | JX171638 | KU171420 | AY329043 | Han et al. (2023) |
| F. commune | FNSC | LC18507 | Zea mays | China | OQ125095 | OQ125101 | - | - | Han et al. (2023) |
| F. commune | FNSC | LC18486 | Zea mays | China | OQ125094 | OQ125100 | - | - | Han et al. (2023) |
| F. commune | FNSC | LC18652 | Zea mays | China | OQ125093 | OQ125099 | - | - | Han et al. (2023) |
| F. commune | FNSC | LC18609 | Zea mays | China | OQ125092 | OQ125098 | - | - | Han et al. (2023) |
| F. commune | FNSC | LC18583 | Zea mays | China | OQ125097 | OQ125103 | - | - | Han et al. (2023) |
| F. commune | FNSC | LC18568 | Zea mays | China | OQ125096 | OQ125102 | - | - | Han et al. (2023) |
| F. commune | FNSC | DB19LUG07 | Zea mays | Italy | MW419921 | MW419923 | OR566042 | OR566011 | Mezzalama et al. (2021), This study |
| F. commune | FNSC | 24 | Zea mays | Italy | OR565988 | OR566049 | OR566026 | OR566010 | This study |
| F. concentricum | FFSC | CBS 450.97 | Musa sapientum | Costa Rica | AF160282 | JF741086 | MW402467 | MW402334 | Yilmaz et al. (2021) |
| F. contaminatum | FOSC | CBS 111552 | Pasteurized fruit juice | The Netherlands | MH484991 | MH484900 | MH484718 | MH485082 | Lombard et al. (2019) |
| F. cugenangense | FOSC | CBS 620.72 | Crocus sp. | Germany | MH484970 | MH484879 | MH484697 | MH485061 | Lombard et al. (2019) |
| F. cugenangense | FOSC | CBS 130308 | Human toe nail | New Zealand | MH485011 | MH484920 | MH484738 | MH485102 | Lombard et al. (2019) |
| F. cugenangense | FOSC | CBS 131393 | Vicia faba | Australia | MH485019 | MH484928 | MH484746 | MH485110 | Lombard et al. (2019) |
| F. cugenangense | FOSC | CBS 130304 | Gossypium barbadense | China | MH485012 | MH484921 | MH484739 | MH485103 | Lombard et al. (2019) |
| F. cugenangense | FOSC | 36 | Zea mays | Italy | OR565989 | OR566050 | OR566027 | - | This study |
| F. curculicola | FFSC | PPRI20464 | Amaranthus cruentus | South Africa | MF787267 | MN605063 | - | MF787259 | Vermeulen et al. (2021) |
| F. curvatum | FOSC | CBS 247.61 | Matthiola incana | Germany | MH484967 | MH484876 | MH484694 | MH485058 | Lombard et al. (2019) |
| F. curvatum | FOSC | CBS 238.94 | Beaucarnia sp. | The Netherlands | MH484984 | MH484893 | MH484711 | MH485075 | Lombard et al. (2019) |
| F. curvatum | FOSC | CBS 141.95 | Hedera helix | The Netherlands | MH484985 | MH484894 | MH484712 | MH485076 | Lombard et al. (2019) |
| F. denticulatum | FFSC | CBS 406.97 | Ipomoea batatas | Cuba | MN533999 | MN534273 | MN534185 | MN534067 | Yilmaz et al. (2021) |
| F. dhileepanii | FFSC | BRIP 71717 | Unknown | Unknown | OK509072 | OK533536 | - | - | Yilmaz et al. (2021) |
| F. dlaminii | FFSC | CBS 481.94 | Unknown | Unknown | MN534003 | MN534257 | MN534151 | MN534139 | Yilmaz et al. (2021) |
| F. dlaminii | FFSC | CBS 671.94 | Soil | South Africa | MN534004 | MN534254 | MN534152 | MN534136 | Yilmaz et al. (2021) |
| F. duoseptatum | FOSC | CBS 102026 | Musa sapientum | Malaysia | MH484987 | MH484896 | MH484714 | MH485078 | Lombard et al. (2019) |
| F. echinatum | FFSC | CBS 146497 | Unidentified tree | South Africa | MW834273 | MW834004 | MW834110 | MW834301 | Crous et al. (2021) |
| F. elaeagni | FFSC | LC13629 | Elaeagnus pungens | China | MW580468 | MW474414 | MW566295 | MW533750 | Wang (2022) |
| F. elaeidis | FOSC | CBS 217.49 | Elaeis sp. | Zaire | MH484961 | MH484870 | MH484688 | MH485052 | Lombard et al. (2019) |
| F. elaeidis | FOSC | CBS 255.52 | Elaeis guineensis | Unknown | MH484965 | MH484874 | MH484692 | MH485056 | Lombard et al. (2019) |
| F. elaeidis | FOSC | CBS 218.49 | Elaeis sp. | Zaire | MH484962 | MH484871 | MH484689 | MH485053 | Lombard et al. (2019) |
| F. fabacearum | FOSC | CPC 25801 | Zea mays | South Africa | MH485029 | MH484938 | MH484756 | MH485120 | Lombard et al. (2019) |
| F. fabacearum | FOSC | CPC 25802 | Glycine max | South Africa | MH485030 | MH484939 | MH484757 | MH485121 | Lombard et al. (2019) |
| F. fabacearum | FOSC | CPC 25803 | Glycine max | South Africa | MH485031 | MH484940 | MH484758 | MH485122 | Lombard et al. (2019) |
| F. ficicrescens | FFSC | CBS 125177 | Environmental | Iran | MN534006 | MN534281 | MN534176 | MN534071 | Yilmaz et al. (2021) |
| F. foetens | FOSC | CBS 120665 | Nicotiana tabacum | Iran | MH485009 | MH484918 | MH484736 | MH485100 | Lombard et al. (2019) |
| F. fracticaudum | FFSC | CMW:25240 | Pinus maximinoi | Colombia | MN534009 | MN534231 | MN534161 | MN534103 | Yilmaz et al. (2021) |
| F. fractiflexum | FFSC | NRRL 28852 | Cymbidium sp. | Japan | AF160288 | LT575064 | AF158341 | AF160315 | Yilmaz et al. (2021) |
| F. fredkrugeri | FFSC | CBS 408.97 | Soil | Maryland | MW402126 | MW402814 | MW402461 | MW402324 | Yilmaz et al. (2021) |
| F. fujikuroi | FFSC | CBS 186.56 | Unknown | Unknown | MW402108 | EF470116 | MW402447 | MW402306 | Yilmaz et al. (2021) |
| F. gaditjirri | FNSC | NRRL 45417 | Hetepogon triticeus | Australia | MN193881 | MN193909 | KU171424 | KU171784 | Sandoval-Denis et al. (2018) |
| F. globosum | FFSC | CBS 430.97 | Zea mays seed | South Africa | MN534013 | MN534265 | MN534219 | MN534125 | Yilmaz et al. (2021) |
| F. glycines | FOSC | CBS 176.33 | Linum usitatissium | Unknown | MH484959 | MH484868 | MH484686 | MH485050 | Lombard et al. (2019) |
| F. gossypinum | FOSC | CBS 116611 | Gossypium hirsutum | Ivory Coast | MH484998 | MH484907 | MH484725 | MH485089 | Lombard et al. (2019) |
| F. gossypinum | FOSC | CBS 116613 | Gossypium hirsutum | Ivory Coast | MH485000 | MH484909 | MH484727 | MH485091 | Lombard et al. (2019) |
| F. gossypinum | FOSC | CBS 116612 | Gossypium hirsutum | Ivory Coast | MH484999 | MH484908 | MH484726 | MH485090 | Lombard et al. (2019) |
| F. guttiforme | FFSC | CBS 409.97 | Ananas comosus | Brazil | MT010999 | MT010967 | MT010901 | MT011048 | Yilmaz et al. (2021) |
| F. hechiense | FFSC | LC13646 | Musa nana | China | MW580496 | MW474442 | MW566323 | MW533775 | Wang (2022) |
| F. hoodiae | FOSC | CBS 132474 | Hoodia gordonii | South Africa | MH485020 | MH484929 | MH484747 | MH485111 | Lombard et al. (2019) |
| F. inflexum | FOSC | NRRL 20433 | Vicia faba | Germany | AF008479 | JX171583 | AF158366 | – | O’Donnell et al. (2013) |
| F. konzum | FFSC | CBS 139382 | Unknown | Unknown | MW402071 | MW402804 | MW402418 | MW402270 | Yilmaz et al. (2021) |
| F. lactis | FFSC | CBS 420.97 | Ficus carica | USA | MN534015 | - | MN534181 | MN534078 | Yilmaz et al. (2021) |
| F. languescens | FOSC | CBS 645.78 | Solanum lycopersicum | Morocco | MH484971 | MH484880 | MH484698 | MH485062 | Lombard et al. (2019) |
| F. libertatis | FOSC | CPC 25782 | Asphalatus sp. | South Africa | MH485023 | MH484932 | MH484750 | MH485114 | Lombard et al. (2019) |
| F. lumajangense | FFSC | LC13652 | Arenga caudata | China | MW580503 | MW474449 | MW566330 | MW533782 | Wang (2022) |
| F. lyarnte | FNSC | NRRL 54252 | Sorghum interjectum | Australia | MN193880 | MN193908 | - | - | Sandoval-Denis et al. (2018) |
| F. madaense | FFSC | CBS 146648 | Arachis hypogaea | Nigeria | MW402095 | MW402761 | MW402436 | MW402294 | Yilmaz et al. (2021) |
| F. mangiferae | FFSC | CBS 119853 | Mangifera sp. | South Africa | MN534016 | MN534270 | MN534225 | MN534140 | Yilmaz et al. (2021) |
| F. marasasianum | FFSC | CMW:25512 | Pinus tecunumanii | Colombia | MN534018 | MN534249 | MN534208 | MN534113 | Yilmaz et al. (2021) |
| F. mexicanum | FFSC | NRRL 47473 | Mangifera indica | Mexico | GU737416 | LR792615 | GU737389 | GU737308 | Yilmaz et al. (2021) |
| F. mirum | FFSC | LLC929 | Sorghum | Ethiopia | OP487012 | OP486581 | OP485896 | - | Lombard et al. (2022) |
| F. miscanthi | FNSC | NRRL 26231 | Miscanthus sinensis | Japan | KU171725 | KU171705 | KU171425 | KU171785 | Han et al. (2023) |
| F. mundagurra | FFSC | RBG5717 | Soil | Australia | KP083256 | KP083276 | MN534214 | MN534146 | Yilmaz et al. (2021) |
| F. napiforme | FFSC | NRRL25196 | Pennisetum typhoides | South Africa | MN193863 | MN534291 | MN534192 | MN534085 | Laraba et al. (2020) |
| F. nirenbergiae | FOSC | CBS 129.24 | Secale cereale | Unknown | MH484955 | MH484864 | MH484682 | MH485046 | Lombard et al. (2019) |
| F. nirenbergiae | FOSC | CBS 127.81 | Chrysantemum sp. | USA | MH484974 | MH484883 | MH484701 | MH485065 | Lombard et al. (2019) |
| F. nirenbergiae | FOSC | CBS 840.88 | Dianthus caryophyllus | The Netherlands | MH484978 | MH484887 | MH484705 | MH485069 | Lombard et al. (2019) |
| F. nirenbergiae | FOSC | CBS 744.79 | Passiflora edulis | Brazil | MH484973 | MH484882 | MH484700 | MH485064 | Lombard et al. (2019) |
| F. nirenbergiae | FOSC | 1RI (Pta 1.2) | Zea mays | Italy | OR565990 | OR566051 | OR566028 | - | This study |
| F. nisikadoi | FNSC | NRRL 25179 | Phyllostachys nigra | Japan | MN193879 | MN193907 | - | - | Sandoval-Denis et al. (2018) |
| F. nygamai | FFSC | CBS 413.97 | Oryza sativa | Morocco | MW402127 | MW402815 | MW402462 | MW402325 | Yilmaz et al. (2021) |
| F. odoratissimum | FOSC | CBS 794.70 | Albizzia julibrissin | Iran | MH484969 | MH484878 | MH484696 | MH485060 | Lombard et al. (2019) |
| F. ophioides | FFSC | CBS 118510 | Panicum maximum | South Africa | MN534020 | MN534301 | MN534201 | MN534121 | Yilmaz et al. (2021) |
| F. oxysporum | FOSC | CBS 144134 | Solanum tuberosum | Germany | MH485044 | MH484953 | MH484771 | MH485135 | Lombard et al. (2019) |
| F. oxysporum | FOSC | CBS 144135 | Solanum tuberosum | Germany | MH485045 | MH484954 | MH484772 | MH485136 | Lombard et al. (2019) |
| F. oxysporum | FOSC | CBS 221.49 | Camellia sinensis | South East Asia | MH484963 | MH484872 | MH484690 | MH485054 | Lombard et al. (2019) |
| F. oxysporum | FOSC | CPC 25822 | Protea sp. | South Africa | MH485034 | MH484943 | MH484761 | MH485125 | Lombard et al. (2019) |
| F. oxysporum sensu lato | FOSC | 11 | Zea mays | Italy | OR565991 | OR566052 | OR566029 | - | This study |
| F. oxysporum sensu lato | FOSC | 12 | Zea mays | Italy | OR565992 | OR566053 | OR566030 | - | This study |
| F. oxysporum sensu lato | FOSC | 18 | Zea mays | Italy | OR565993 | OR566054 | OR566031 | - | This study |
| F. oxysporum sensu lato | FOSC | 26 | Zea mays | Italy | OR565994 | OR566055 | OR566032 | - | This study |
| F. oxysporum sensu lato | FOSC | 51 | Zea mays | Italy | OR565995 | OR566056 | OR566033 | - | This study |
| F. panlongense | FFSC | LC13656 | Musa nana | China | MW580510 | MW474456 | MW566337 | MW533789 | Wang (2022) |
| F. paranisikadoi | FNSC | LC2824 | Zea mays | China | MW594317 | MW474550 | - | MW533921 | Han et al. (2023) |
| F. parvisorum | FFSC | CMW:25267 | Pinus patula | Colombia | KJ541060 | - | - | KJ541055 | Yilmaz et al. (2021) |
| F. pharetrum | FOSC | CPC 30822 | Aliodendron dichotomum | South Africa | MH485042 | MH484951 | MH484769 | MH485133 | Lombard et al. (2019) |
| F. phyllophilum | FFSC | NRRL13617 | Dracaena deremensis | Italy | MN193864 | KF466410 | KF466333 | KF466443 | Laraba et al. (2020) |
| F. pilosicola | FFSC | NRRL 29123 | Bidens pilosa | USA | MN534054 | MN534247 | MN534165 | MN534098 | Yilmaz et al. (2021) |
| F. pininemorale | FFSC | CMW:25243 | Pinus tecunumanii | Colombia | MN534026 | MN534250 | MN534211 | MN534115 | Yilmaz et al. (2021) |
| F. proliferatum | FFSC | CBS 480.96 | Tropical rain forest soil | Papua New Guinea | MN534059 | MN534272 | MN534217 | MN534129 | Yilmaz et al. (2021) |
| F. pseudoanthophilum | FFSC | CBS 745.97 | Zea mays | Zimbabwe | MW402148 | MW402820 | MW402476 | MW402349 | Yilmaz et al. (2021) |
| F. pseudocircinatum | FFSC | NRRL22946 | Solanum sp. | Ghana | AF160271 | MN534277 | MN534190 | MN534069 | O' Donnell et al. (2000) |
| F. pseudonygamai | FFSC | CBS 416.97 | Pennisetum typhoides | Nigeria | MN534030 | MN534283 | MN534194 | MN534064 | Yilmaz et al. (2021) |
| F. ramigenum | FFSC | NRRL25208 | Ficus carica | USA | KF466423 | KF466412 | MN534187 | MN534145 | Proctor et al. (2013) |
| F. sacchari | FFSC | CBS 131370 | Oryzae australiensis | Australia | MW402031 | MW402793 | MW402404 | MW402230 | Yilmaz et al. (2021) |
| F. secorum | FFSC | NRRL 62593 | Beta vulgaris | USA | KJ189225 | – | KJ189235 | – | Yilmaz et al. (2021) |
| F. siculi | FFSC | CPC 27188 | Citrus sinensis | Italy | LT746214 | LT746327 | LT746189 | LT746346 | Sandoval-Denis et al. (2018) |
| F. sororula | FFSC | CMW:25513 | Pinus tecunumanii | Colombia | MN534035 | MN534246 | MN534210 | MN534114 | Yilmaz et al. (2021) |
| F. sterilihyposum | FFSC | NRRL 25623 | Mangifera sp. | South Africa | MN193869 | MN193897 | AF158353 | AF160316 | Yilmaz et al. (2021) |
| F. subglutinans | FFSC | CBS 215.76 | Zea mays | Germany | MN534061 | MN534241 | MN534171 | MN534109 | Yilmaz et al. (2021) |
| F. succisae | FFSC | CBS 187.34 | Zostera marina | UK | MW402109 | MW402810 | MW402448 | MW402307 | Yilmaz et al. (2021) |
| F. sudanense | FFSC | CBS 454.97 | Striga hermonthica | Sudan | MN534037 | MN534278 | MN534179 | MN534073 | Yilmaz et al. (2021) |
| F. tardichlamydosporum | FOSC | CBS 102028 | Musa sapientum | Malaysia | MH484988 | MH484897 | MH484715 | MH485079 | Lombard et al. (2019) |
| F. temperatum | FFSC | CBS 135538 | Pulmonary infection (human) | Mexico | MN534039 | MN534239 | MN534168 | MN534111 | Yilmaz et al. (2021) |
| F. terricola | FFSC | CBS 483.94 | Soil | Australia | MN534042 | LT996156 | MN534189 | MN534076 | Yilmaz et al. (2021) |
| F. thapsinum | FFSC | CBS 539.79 | Man, white grained mycetoma | Italy | MW402140 | MW402818 | MW402472 | MW402340 | Yilmaz et al. (2021) |
| F. tjaetaba | FFSC | RBG5361 | Sorghum interjectum | Australia | KP083263 | KP083275 | LT996187 | GU737296 | Laurence et al. (2015) |
| F. triseptatum | FOSC | CBS 258.50 | Ipomoea batatas | USA | MH484964 | MH484873 | MH484691 | MH485055 | Lombard et al. (2019) |
| F. tupiense | FFSC | NRRL 53984 | Mangifera indica | Brazil | GU737404 | LR792619 | GU737377 | GU737350 | Yilmaz et al. (2021) |
| F. udum | FFSC | NRRL22949 | Unknown | Unknown | AF160275 | LT996172 | MW402442 | U34433 | O' Donnell et al. (2000) |
| F. verticillioides | FFSC | CBS 116665 | Solanum lycopersicum | Unknown | MW401976 | MW402825 | MW402375 | MW402176 | Yilmaz et al. (2021) |
| F. verticillioides | FFSC | CBS 125.73 | Trichosanthes dioica | India | MW402012 | MW402791 | MW402392 | MW402212 | Yilmaz et al. (2021) |
| F. verticillioides | FFSC | CBS 167.87 | Pinus seed | USA | MW402101 | - | MW402441 | MW402300 | Yilmaz et al. (2021) |
| F. verticillioides | FFSC | CBS 447.95 | Asparagus | Unknown | MW402133 | MW402770 | MW402466 | MW402332 | Yilmaz et al. (2021) |
| F. verticillioides | FFSC | CBS 531.95 | Zea mays | Unknown | MW402136 | MW402771 | MW402468 | MW402336 | Yilmaz et al. (2021) |
| F. verticillioides | FFSC | CBS 131389 | Environmental | Australia | KU711695 | KU604226 | MN534193 | KU603857 | Yilmaz et al. (2021) |
| F. verticillioides | FFSC | CBS 734.97 | Zea mays | Germany | MW402146 | EF470122 | AF158315 | MW402346 | Yilmaz et al. (2021) |
| F. verticillioides | FFSC | 8.2 | Zea mays | Italy | OR565996 | OR566057 | OR566034 | OR566012 | This study |
| F. verticillioides | FFSC | 35.1.4 | Zea mays | Italy | OR565997 | OR566058 | OR566035 | OR566013 | This study |
| F. verticillioides | FFSC | 56.1.2 | Zea mays | Italy | OR565998 | OR566059 | OR566036 | OR566014 | This study |
| F. verticillioides | FFSC | 56.2.2 | Zea mays | Italy | OR565999 | OR566060 | OR566037 | OR566015 | This study |
| F. verticillioides | FFSC | 56.2.3 | Zea mays | Italy | OR566000 | OR566061 | OR566038 | OR566016 | This study |
| F. verticillioides | FFSC | 56.2.4 | Zea mays | Italy | OR566001 | OR566062 | OR566039 | OR566017 | This study |
| F. verticillioides | FFSC | 56.2.5 | Zea mays | Italy | OR566002 | OR566063 | OR566040 | OR566018 | This study |
| F. verticillioides | FFSC | 57.2.1 | Zea mays | Italy | OR566003 | OR566064 | OR566041 | OR566019 | This study |
| F. veterinarium | FOSC | CBS 109898 | Shark peritoneum | The Netherlands | MH484990 | MH484899 | MH484717 | MH485081 | Lombard et al. (2019) |
| F. volatile | FFSC | CBS 143874 | Human bronchoalveolar lavage fluid | French Guiana | LR596007 | LR596006 | MK984595 | LR596008 | Yilmaz et al. (2021) |
| F. werrikimbe | FFSC | CBS 125535 | Sorghum leiocladum | Australia | MW928846 | MN534304 | MN534203 | MN534104 | Yilmaz et al. (2021) |
| F. xylarioides | FFSC | NRRL25486 | Coffea trunk | Ivory Coast | MN193874 | HM068355 | MW402455 | AY707118 | Laraba et al. (2020) |
| F. xyrophilum | FFSC | NRRL 62710 | Xyris spp. | Guyana | MN193875 | MN193903 | – | – | Yilmaz et al. (2021) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



