Submitted:
07 May 2025
Posted:
08 May 2025
You are already at the latest version
Abstract
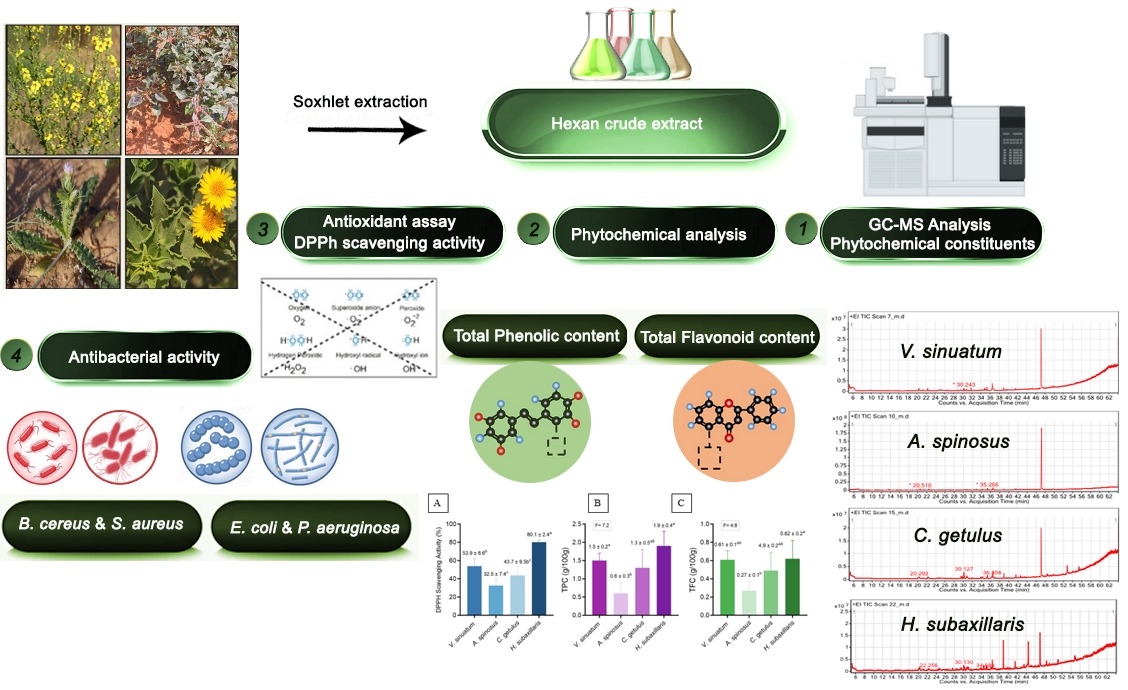
Underexplored medicinal plants serve as valuable reservoirs of bioactive compounds that exhibit complex synergistic interactions whose therapeutic potential is increasingly rec-ognized. In the current study, the antimicrobial activity, antioxidant potential, and Gas Chromatography - Mass Spectrometry (GC-MS) phytochemical screening of four medici-nal plants (Verbascum sinuatum, Amaranthus spinosus, Carduus getulus, and Heterotheca sub-axillaris) from the Gaza Strip in Palestine were investigated. For a comprehensive phyto-chemical characteristic, an analysis using GC-MS was conducted using the hexane ex-tracts of each species. For the purpose of evaluating the antioxidant active ingredients, the total phenolic content (TPC) and total flavonoid content (TFC) were measured. The 2,2-diphenyl-1-picrylhydrazyl (DPPH) test was utilized to assess antioxidant activity, while the method of disc diffusion was applied to evaluate antibacterial activity. GC-MS analysis revealed various lipophilic compounds in all species, including fatty acid esters, terpenoids, phenols, and polyacetylenes. V. sinuatum was dominated by three major con-stituents: saturated fatty acid esters, phenolic antioxidants, and a putative alkaloid. A. spinosus contained six compounds representing oxygenated monoterpenes, fatty acid de-rivatives, and aromatic compounds. Ten compounds in C. getulus represented predomi-nantly monoterpenes, diterpenes, fatty acid derivatives, and phenolic antioxidants. H. subaxillaris had eight primary metabolites dominated by terpenoids, specialized fatty acid esters, and phenolic compounds. V. sinuatum and C. getulus showed moderate bioactivity, possibly due to the synergism of fatty acid esters and diterpenoid–phenol combinations, while the monoterpenoid-rich extract of A. spinosus possessed relatively weak effects. These results make H. subaxillaris a promising candidate for developing natural products, especially to combat resistant pathogens. These plants' chemical diversity and overall bi-ological activity emphasize their potential for pharmaceutical applications and warrant further mechanistic exploration.
Keywords:
Verbascum sinuatum
; Amaranthus spinosus
; Carduus getulus
; Heterotheca subaxillaris
; GC-Ms
; antioxidant
; antimicrobial
; phenols
; flavonoids
1. Introduction
The emergence of antimicrobial resistance and the prevalence of oxidative stress-related diseases underscore the need to explore novel bioactive compounds from medicinal plants. Synergistic metabolite interactions may provide superior therapeutic effects over single-compound drugs [1]. Assessing the phytochemical content and bioactivity of underutilized species is essential for identifying their medicinal and industrial potential [2]. Recently, the use of natural substances derived from traditional plants to develop pharmaceuticals, functional foods, and cosmetic products has increased significantly [3]. Verbascum sinuatum L. (Scrophulariaceae) is a Mediterranean herbaceous plant, traditionally valued for its medicinal properties. Its aerial parts are traditionally used as an infusion and comprising bioactive constituents, such as iridoids and polyphenols [3]. Amaranthus spinosus L is a globally distributed herb that belongs to the Amaranthaceae / Chenopodiaceae family [4]. This species naturally contains bioactive metabolites including flavonoids, phenols, terpenoids, tannins, and glycosides, which are involved in the human body's important physiological and metabolic functions [5]. Studies report that the plant species has diuretic, emollient, antipyretic, and diaphoretic properties and is traditionally used to treat internal bleeding, ulcers, menstrual disorders, snake bites, gastrointestinal problems, and various skin and mucous membrane wounds [6]. Additionally, the plant exhibits anti-leprotic, anti-diabetic, anti-inflammatory and antiandrogenic effects and serves as a valuable feed source for livestock [7]. A. spinosus, often regarded as an invasive weed in cattle pastures, shows potential as a resource for methane reduction in cow-calf production systems [8]. Carduus (Asteraceae / Compositae) is a Mediterranean genus used to treat colds, gastrointestinal disorders, and rheumatism. Ethnopharmacological and phytochemical studies have highlighted its diverse bioactivities [9]. Various species have provided flavonoids, lignans, alkaloids, sterols, and triterpenes [9]. Notably, Carduus getulus Pomel exhibits a high lipid content, which has been associated with enhanced biochemical and antioxidant defenses, justifying their use in the regulation of liver disease and microbial infections [7,10]. Heterotheca subaxillaris (Lam.) Britt. & Rusby (Asteraceae/Compositae) is a wide-ranging species that occurs in the USA particularly in sandy habitats [11]. It has become a dominant invasive species in Palestine along the coastal dunes. It has a strong camphoraceous odor derived from glandular trichomes covering its aerial parts due to high concentrations of monoterpenoids (camphor, bornyl acetate, borneol) and sesquiterpenoids, which serve as herbivore deterrents [12]. H. subaxillaris produces methylated flavonoids and phenolic compounds, which enhance resistance to abiotic stresses and contribute to its invasive success in arid regions. These secondary metabolites also exhibit allelopathic properties, explaining their ecological dominance in disturbed habitats like coastal dunes [12]. Notably, terpenoid production is influenced by soil nitrogen content, demonstrating phenotypic plasticity in different environments [11,13]. The current study aims to systematically evaluate the antibacterial and antioxidant potential of four medicinal plants (V. sinuatum, A. spinosus, C. getulus, and H. subaxillaris) through integrated phytochemical profiling and bioactivity assays. By elucidating metabolite-activity relationships, we seek to advance their therapeutic development as potential sources of novel bioactive compounds.
2. Results
2.1. GC-MS and Phytochemical Profiling of V. sinuatum
The GC-MS chromatogram of V. sinuatum revealed three dominant peaks, with the most abundant component eluting at 30.243 min, accounting for 65.90% of the total peak area (Table 1, Figure 1). The main compound was recognized methyl palmitate (MW: 270.45 g/mol; Figure 2A). Its electron ionization (EI) mass spectrum exhibited characteristic fragment ions at m/z 74.02 (McLafferty rearrangement) and m/z 87.00 (acyl ion), matching NIST library entries with a match score (total score minus threshold) of 71.0. The second peak (RT: 30.840 min, 13.34% relative abundance) was identified as 3,5-ditert-butylphenol (Figure 2B) (MW: 206.32 g/mol; match score: 77.2). The spectrum displayed key fragments at m/z 190.10 [M–CH₃]⁺ and m/z 57.01 (tert-butyl ion). A third peak eluted at 34.752 min, accounting for 20.76% of the extract composition. This compound showed spectral similarity to phenethylamine derivatives, specifically a hordenine (C₁₀H₁₅NO; MW: 165.23 g/mol; Figure 2C), although the match was tentative with a lower confidence score (match score: 54.9). Fragmentation produced characteristic ions at m/z 57, supporting the tentative classification as a phenethylamine-type alkaloid. Due to its low spectral match, further confirmation via orthogonal techniques would be necessary to verify this assignment (Table 1).
2.2. GC-MS and Phytochemical Profiling of A. spinosus
The GC-MS analysis of the hexane extract of A. spinosus showed six prominent peaks representing key lipophilic phytochemicals (Table 2). The extract was dominated by monoterpenoid alcohols and fatty acid esters, constituting most of the chemical profile. The most abundant compound, eluting at 35.266 min (Peak 6) and accounting for 30.78% of the total area, was identified as isocitronellol (C₁₀H₂₀O; MW: 156.27 g/mol). The mass spectrum exhibited a dominant base peak at m/z 82.9, corresponding to [C6H10]+, formed via α-cleavage of the C–O bond in isocitronellol. Despite its moderate match score of 61.4, the compound's identification is supported by its retention time and co-occurrence with other structurally related terpenes, such as citronellol and geraniol. Further confirmation with reference standards is advised. Citronellol was identified at 20.510 min (Peak 2; 17.59%), with a match score 76.7. The mass spectrum displayed a characteristic base peak at m/z 69.01, derived from α-cleavage and other ions consistent with the hydroxylated side chain of this acyclic monoterpene. Geraniol, a structural isomer of citronellol, appeared at 22.395 min (Peak 3; 17.34%), with a high match score of 81.4. The spectrum is dominated by m/z 69.00 [C5H9]+, consistent with the α-cleavage of geraniol. Peak 5 (34.160 min; 21.75%) was identified as 11,14-Octadecadienoic acid, methyl ester (C₁₉H₃₄O₂; MW: 294.47 g/mol), a polyunsaturated fatty acid ester. Its mass spectrum showed diagnostic ions at m/z 67.02 (due to diene cleavage) and m/z 81.96, typical of linoleic acid derivatives. The high match score of 87.6 supports its confident identification. Methyl elaidate was identified at 32.923 min (Peak 4; 9.51%), with a match score of 86.9. The compound displayed intense fragment ions at m/z 74.03 (McLafferty rearrangement) and m/z 55.03 (allylic cleavage), considered hallmark ions for mono-unsaturated fatty acid esters. The earliest eluting compound, Peak 1 (15.526 min; 3.03%), was identified as linalyl acetate (C₁₂H₂₀O₂; MW: 196.29 g/mol). The mass spectrum exhibited a base peak at m/z 71.01, corresponding to the [C₅H₁₁]⁺ fragment (common for terpenes), along with other diagnostic ions at m/z 92.92 (likely the tropylium ion, C₇H₇⁺) and m/z 120.74 (indicative of acetyl cleavage).
Figure 3.
Total ion chromatogram (TIC) of A, spinosus extract with labeled peaks. (1) Linalyl acetate (2) 3,7-Dimethyloct-6-en-1-ol, (3) Geraniol (4) 9-Octadecenoic acid, methyl ester, (5) 11,14-Octadecadienoic acid, (6) Isocitronellol.
Figure 3.
Total ion chromatogram (TIC) of A, spinosus extract with labeled peaks. (1) Linalyl acetate (2) 3,7-Dimethyloct-6-en-1-ol, (3) Geraniol (4) 9-Octadecenoic acid, methyl ester, (5) 11,14-Octadecadienoic acid, (6) Isocitronellol.
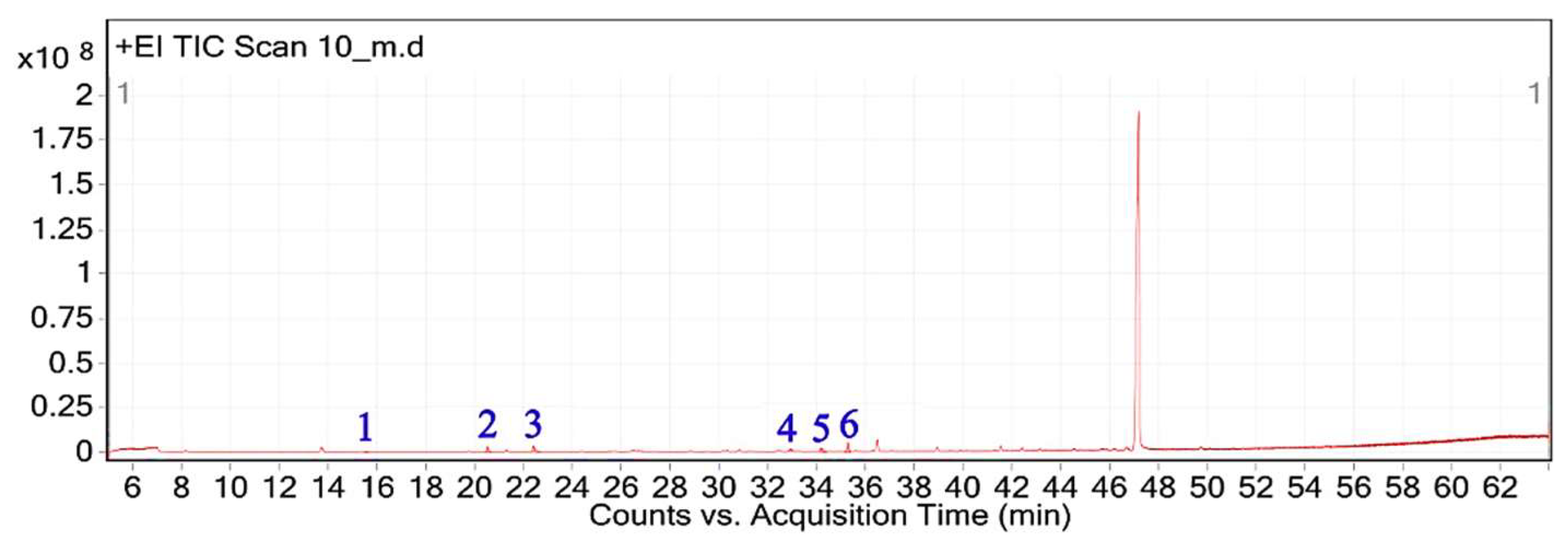
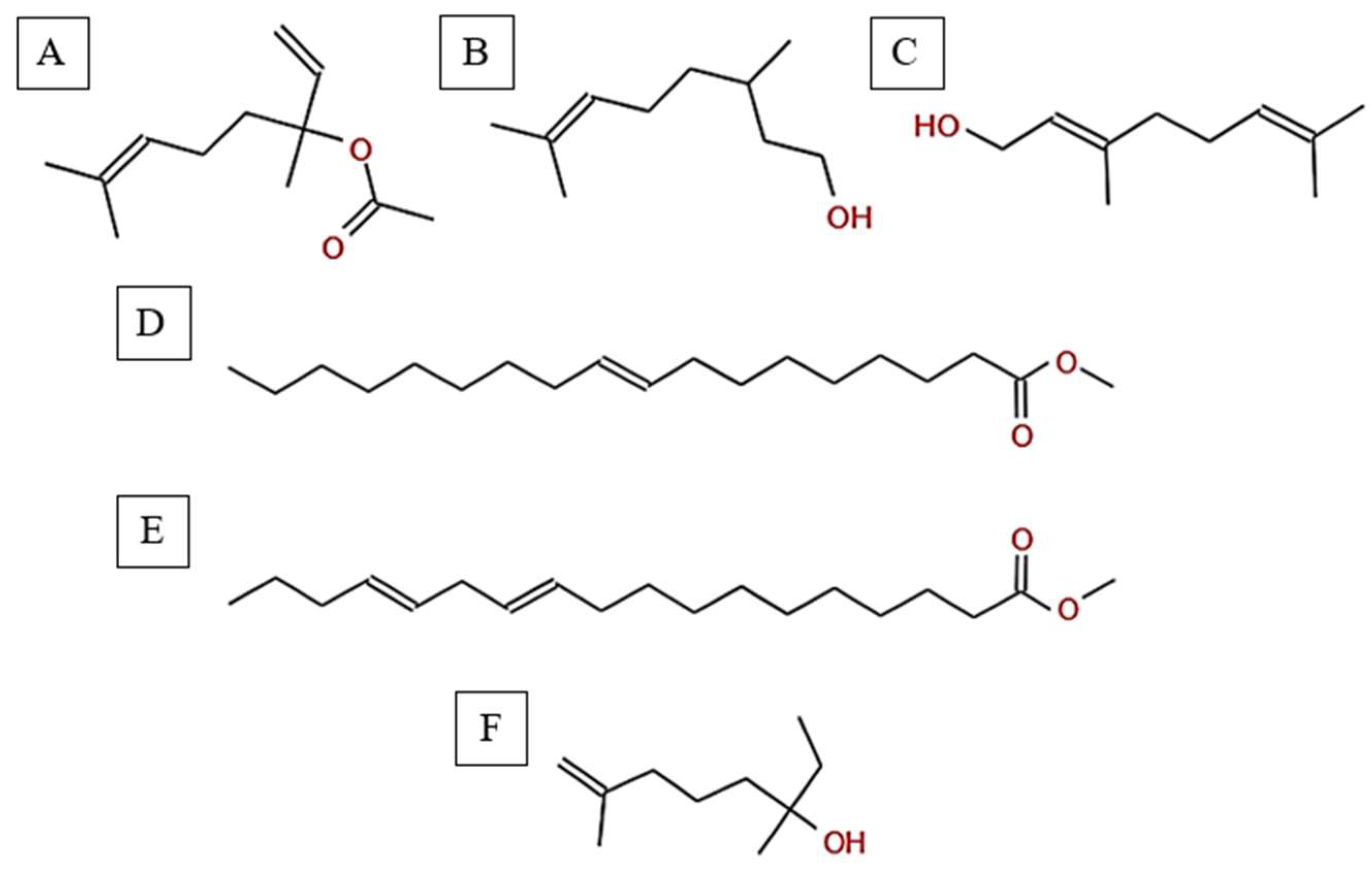
Figure 4.
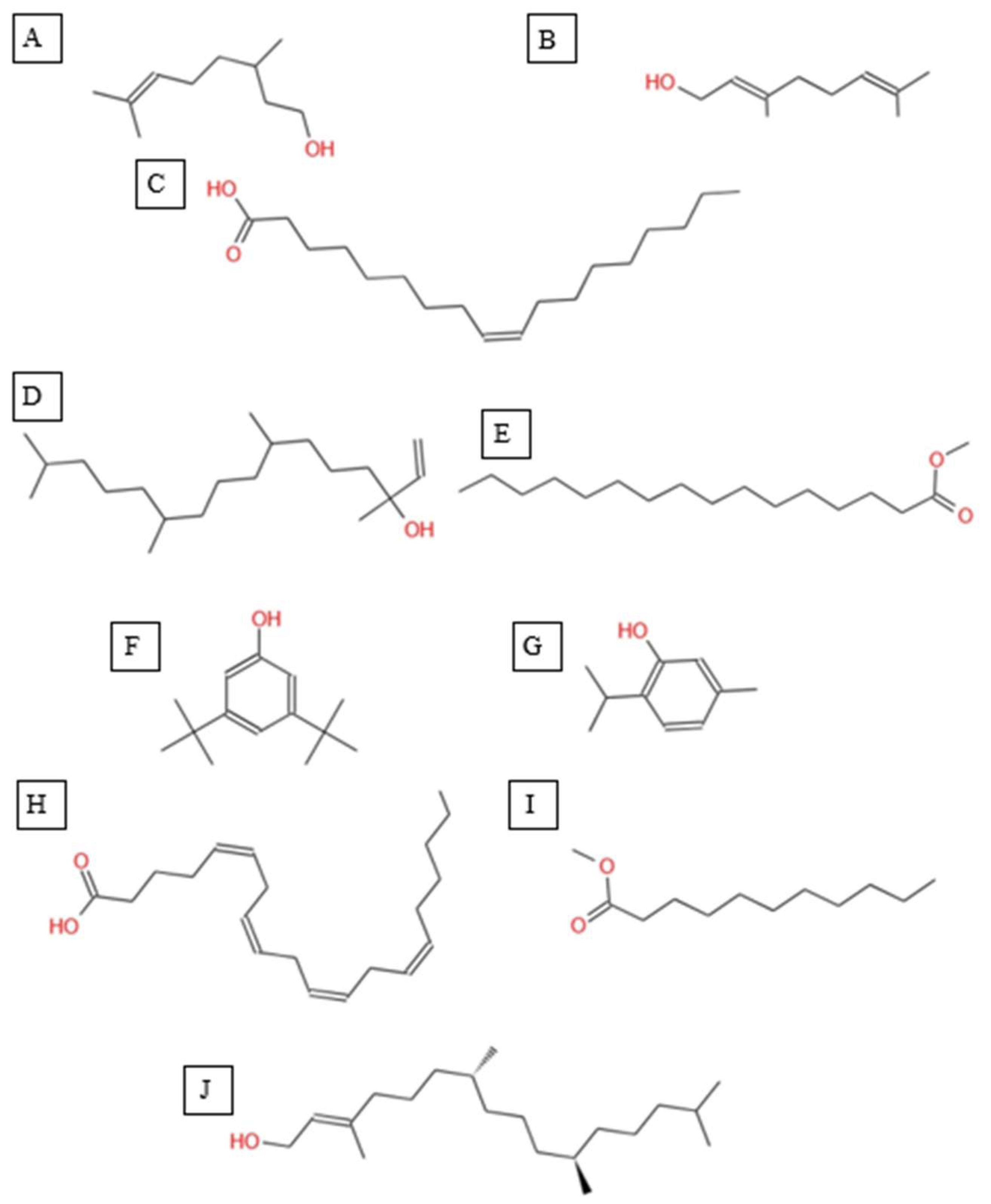
Chemical structures of key compounds identified in A. spinosus: (A) Linalyl acetate (B) 3,7-Dimethyloct-6-en-1-ol, (C) Geraniol (D) 9-Octadecenoic acid, methyl ester, (E) 11,14-Octadecadienoic acid, (F) Isocitronellol.
Figure 4.
Chemical structures of key compounds identified in A. spinosus: (A) Linalyl acetate (B) 3,7-Dimethyloct-6-en-1-ol, (C) Geraniol (D) 9-Octadecenoic acid, methyl ester, (E) 11,14-Octadecadienoic acid, (F) Isocitronellol.
2.3. GC-MS and Phytochemical Profiling of C. getulus
The GC-MS analysis of C. getulus hexane extract revealed a chemically diverse profile, with ten major peaks corresponding to different classes of lipophilic metabolites (Table 3). The extract was predominantly composed of fatty acid esters, terpenoid alcohols, and phenolic compounds, several of which are known for their pharmacological relevance. The most abundant compound, eluting at 30.127 min (Peak 5) and accounting for 66.26% of the total ion current, was identified as methyl palmitate (C₁₇H₃₄O₂; MW: 270.45 g/mol). This saturated fatty acid ester is a common component of plant cuticular waxes and has known antibacterial and anti-inflammatory properties. Its EI mass spectrum exhibited prominent fragment ions at m/z 74.02 (McLafferty rearrangement) and m/z 87.00, supporting its identification with a match score 73.8. 3,5-ditert-butylphenol, a phenolic antioxidant, was the second most abundant metabolite (12.67%; RT 30.800 min; Peak 6). The compound exhibited a high match score of 78.7 and a dominant fragment ion at m/z 190.59, reflecting the loss of a methyl group from the molecular ion. A third principal constituent was isophytol (C₂₀H₄₀O; MW: 296.5 g/mol), a diterpene alcohol, detected at RT 29.528 min (Peak 4) with 9.58% relative abundance and a base fragment at m/z 57.98. Other notable terpenoids included phytol (C₂₀H₄₀O; MW: 296.5 g/mol), eluting at 36.308 min (Peak 10; 1.05%), with a prominent fragment at m/z 70.92 and a match score of 76.1. Additionally, thymol, a monoterpenoid phenol, was identified at RT 31.776 min (Peak 7; 4.41%), characterized by m/z 134.76 and a match score of 74.1. Monoterpenoid alcohols were represented by citronellol (Peak 1; RT 20.292 min; 0.88%; m/z 68.98; match score 74.6) and Geraniol (Peak 2; RT 22.135 min; 1.29%; m/z 92.90; match score 77.6). The extract also contained unsaturated and polyunsaturated fatty acids, including oleic acid (Peak 3; RT 27.226 min; 0.69%; m/z 137.84; match score 65.3) and arachidonic acid (Peak 8; RT 33.083 min; 2.14%; m/z 78.85, match score 77.8). Undecanoic acid methyl ester (Peak 9; RT 33.798 min; 1.03%) was identified as a medium-chain fatty acid ester, with a central ion at m/z 73.94 and a match score of 64.6.
Figure 5.
Total ion chromatogram (TIC) of C. getulus extract with labeled peaks. (1) Citronellol, (2) Geraniol, (3) Oleic acid (4-9) Isophytol, Methyl palmitate, 3,5-ditert-butylphenol, Thymol, Arachidonic acid, Methyl undecanoate respectivelly, (10) Phytol.
Figure 5.
Total ion chromatogram (TIC) of C. getulus extract with labeled peaks. (1) Citronellol, (2) Geraniol, (3) Oleic acid (4-9) Isophytol, Methyl palmitate, 3,5-ditert-butylphenol, Thymol, Arachidonic acid, Methyl undecanoate respectivelly, (10) Phytol.
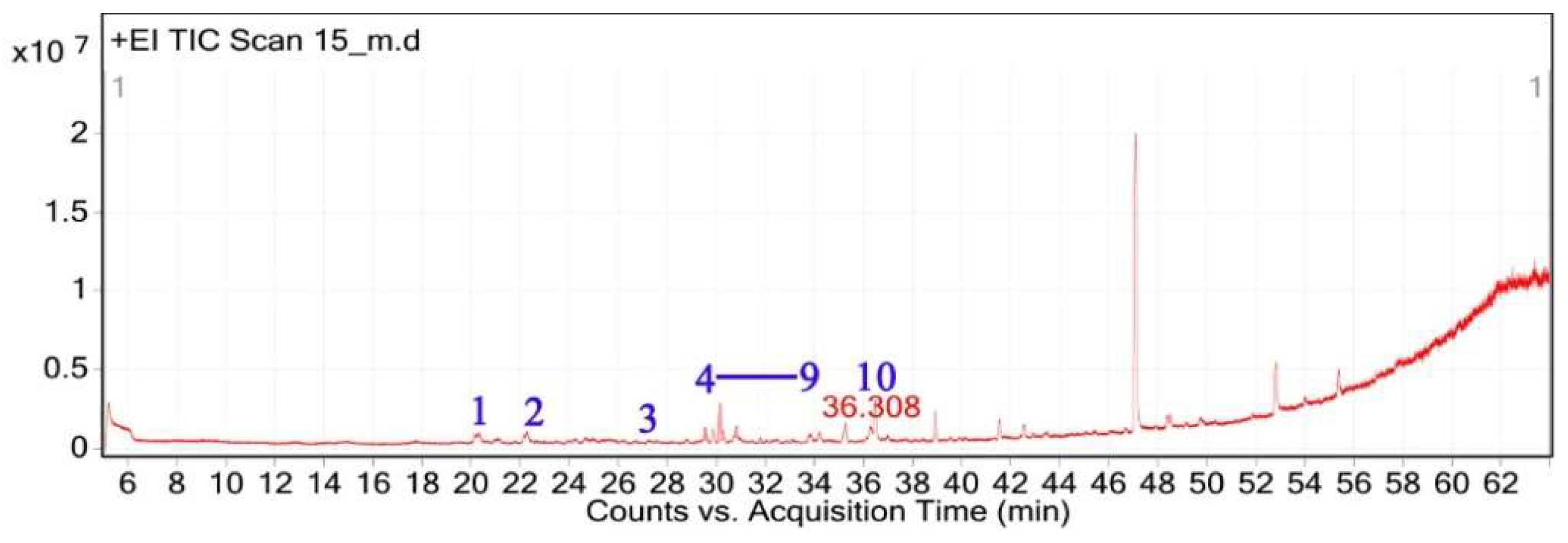
Figure 6.
Chemical structures of key compounds identified in C. getulus: (A) Geraniol, (B) Citronellol, (C) Oleic acid (D) Isophytol (E) Methyl palmitate (F) 3,5-ditert-butylphenol, (G) Thymol (H) Arachidonic acid. (I) Methyl undecanoate (J) Phytol.
Figure 6.
Chemical structures of key compounds identified in C. getulus: (A) Geraniol, (B) Citronellol, (C) Oleic acid (D) Isophytol (E) Methyl palmitate (F) 3,5-ditert-butylphenol, (G) Thymol (H) Arachidonic acid. (I) Methyl undecanoate (J) Phytol.
2.4. GC-MS and Phytochemical Profiling of H. subaxillaris
The GC-MS analysis of H. subaxillaris hexane extract revealed a chemically diverse and bioactive phytochemical profile consisting of eight major peaks, dominated by fatty acid esters, terpenoid alcohols, polyacetylenes, and phenolic antioxidants (Table 4). The most abundant compound, eluting at 30.130 min (Peak 4), was identified as methyl 8-methyl-nonanoate, constituting 36.84% of the total peak area. The EI mass spectrum exhibited a dominant McLafferty rearrangement fragment at m/z 74.03, confirming its identity as a saturated fatty acid methyl ester with a match score 74.3. Methyl palmitate (C₁₇H₃₄O₂; MW: 270.5 g/mol) was the second most abundant metabolite (18.03%, RT 30.153 min; Peak 5), showing a characteristic base ion at m/z 74.02 and a match score of 71.9. Among the early-eluting volatile terpenoids, Geraniol (Peak 1; RT 22.256 min; 3.22%) was identified with a high match score of 78.6. It exhibited key fragment ions at m/z 69.02 (from α-cleavage) and m/z 92.87 (tropylium ion). Cubebol (Peak 2; RT 27.538 min; 18.52%). The spectrum is dominated by m/z 118.76 [C8H14O]+, indicating vinyl ether cleavage in geranyl vinyl ether and a strong match score (78.0). Falcarinol (Peak 3; RT 29.072 min; 3.56%), a long-chain fatty alcohol with a polyacetylene moiety, was confirmed by ions at m/z 128.67, a typical fragment of polyyne cleavage. A unique class of polyunsaturated fatty acid esters was represented by 13,16-octadecadienoic acid methyl ester (Peak 6; RT 30.242 min; 4.31%). The spectrum exhibits a base peak at m/z 74.08 [C3H6O2]+, consistent with McLafferty rearrangement of the methyl ester group in 13,16-octadecadienoic acid methyl ester. Additional peaks at m/z 86.92 [C5H10O]+ and hydrocarbon fragments (m/z 55–69) support the assignment. 2,5-octadecadiynoic acid methyl ester (Peak 8; RT 34.889 min; 6.24%). The spectrum is dominated by m/z 83.95 [C5H7O]+, indicating cleavage near the diyne group. 3,5-ditert-butylphenol (Peak 7; RT 30.772 min; 9.28%) is a well-documented phenolic antioxidant, identified by its base peak at m/z 190.65.
Figure 7.
Total ion chromatogram (TIC) of H. subaxillaris extract with labeled peaks. (1) Geraniol, (2) Cubebol (3-7) Falcarinol, Methyl 8-methyl-nonanoate, Methyl palmitate, methyl octadeca-13,16-diynoate, and 3,5-ditert-butylphenol respectively, (8) 2,5-Octadecadiynoic acid, methyl ester.
Figure 7.
Total ion chromatogram (TIC) of H. subaxillaris extract with labeled peaks. (1) Geraniol, (2) Cubebol (3-7) Falcarinol, Methyl 8-methyl-nonanoate, Methyl palmitate, methyl octadeca-13,16-diynoate, and 3,5-ditert-butylphenol respectively, (8) 2,5-Octadecadiynoic acid, methyl ester.
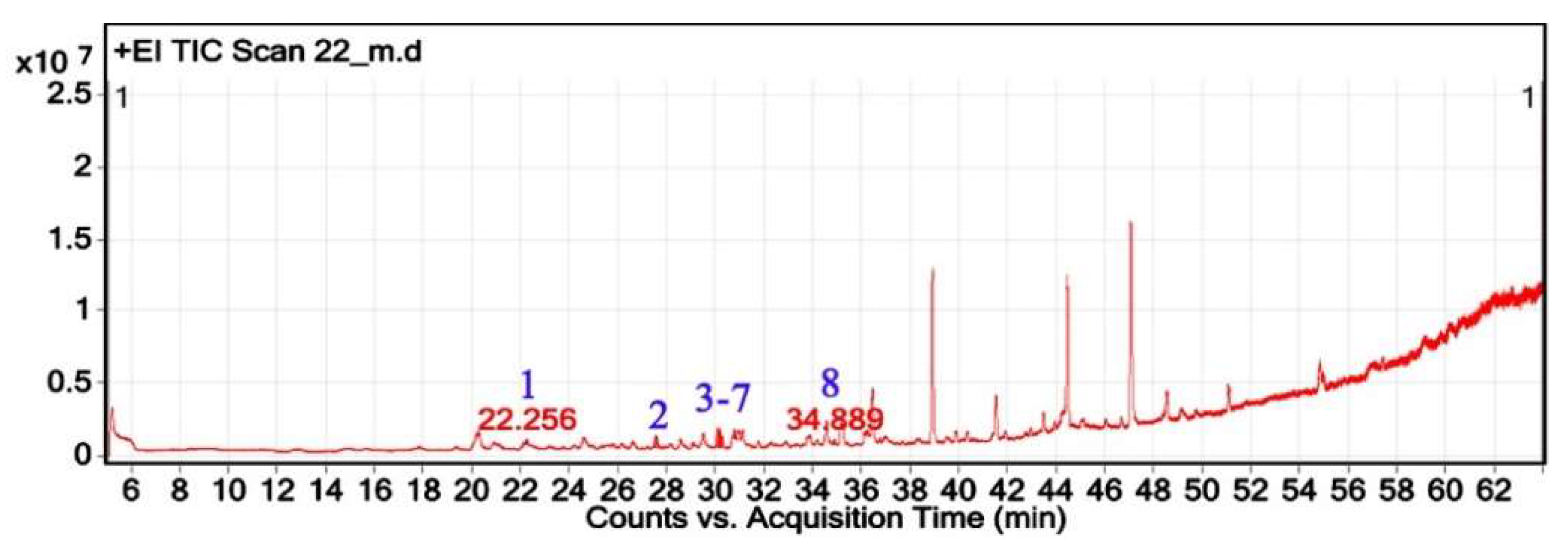
Figure 8.
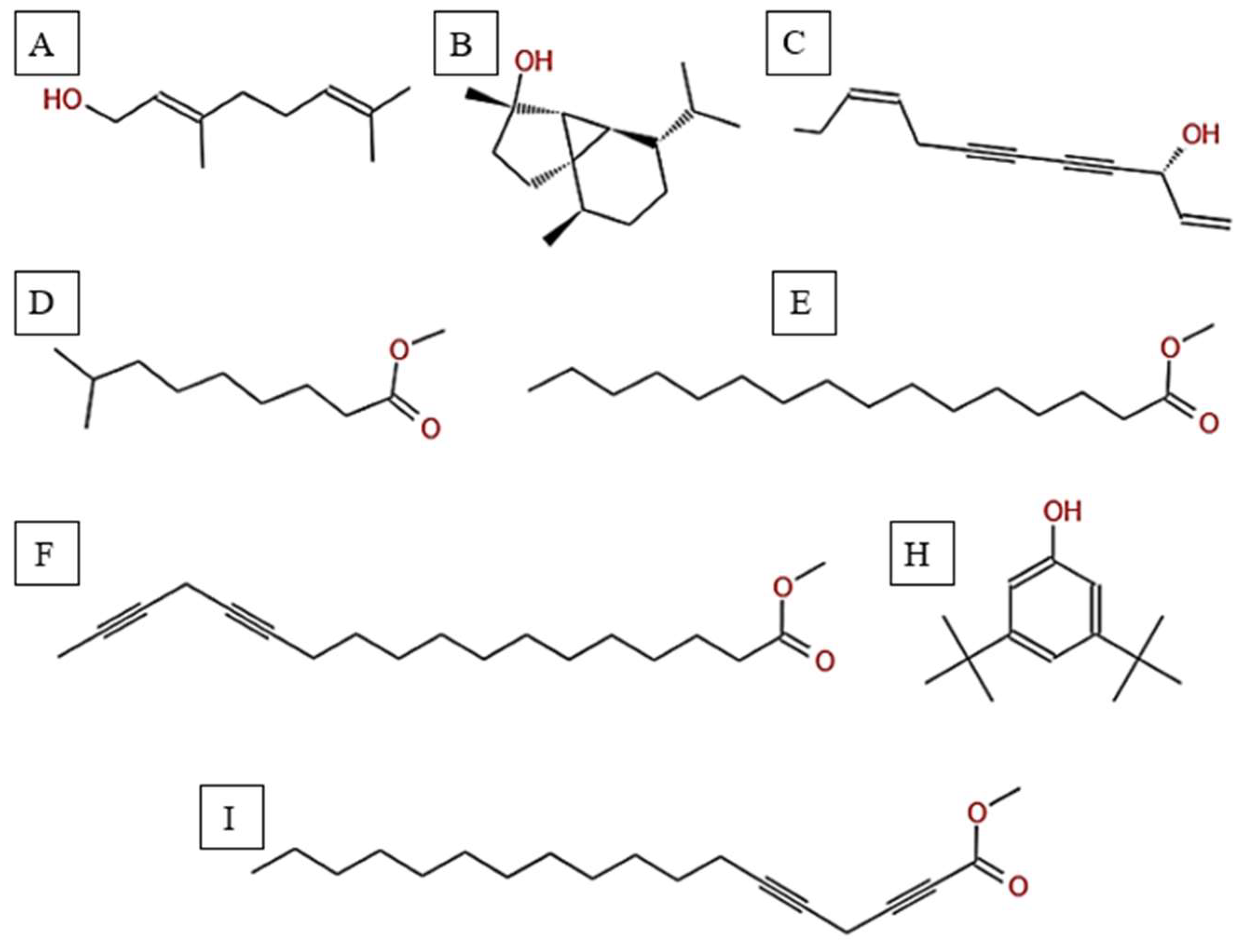
Chemical structures of key compounds identified in H. subaxillaris: (1) Geraniol, (2) Cubebol (3) Falcarinol, (4) Methyl 8-methyl-nonanoate, (5) Methyl palmitate, (6) methyl octadeca-13,16-diynoate, (7) 3,5-ditert-butylphenol, (8) 2,5-Octadecadiynoic acid, methyl ester.
Figure 8.
Chemical structures of key compounds identified in H. subaxillaris: (1) Geraniol, (2) Cubebol (3) Falcarinol, (4) Methyl 8-methyl-nonanoate, (5) Methyl palmitate, (6) methyl octadeca-13,16-diynoate, (7) 3,5-ditert-butylphenol, (8) 2,5-Octadecadiynoic acid, methyl ester.
2.5. Antibacterial Activity Analysis
The hexane crude extracts of the selected medicinal plants showed a statistically significant difference (p < 0.05) in their activity against the tested bacterial strains (Table 5). H. subaxillaris exhibited the most potent inhibition against B. cereus (19.8 ± 0.8 mm), surpassing all other species in activity. Both V. sinuatum (15.2 ± 1.5 mm) and C. getulus (13.5 ± 1.2 mm) showed moderate activity and did not differ significantly from each other, while A, spinosus exhibited the weakest activity (8.3 ± 0.6 mm). For S. aureus (MRSA), V. sinuatum (12.6 ± 0.9 mm), C. getulus (11.8 ± 0.9 mm), and H. subaxillaris (12.5 ± 0.7 mm) exhibited statistically similar and potent inhibitory effects, while A. spinosus had significantly lower activity (7.0 ± 1.1 mm). In the case of E. coli O157:H7, both V. sinuatum (9.3 ± 0.7 mm) and H. subaxillaris (10.1 ± 0.3 mm) displayed comparable and significantly higher antibacterial activity than C. getulus (6.2 ± 0.5 mm) and A. spinosus (5.5 ± 0.8 mm). Regarding P. aeruginosa, H. subaxillaris (8.4 ± 0.9 mm) and V. sinuatum (7.0 ± 0.5 mm) showed the most effective activity, while C. getulus (5.0 ± 0.4 mm) and A. spinosus (4.2 ± 0.6 mm) were significantly less active. These results highlight the stronger efficacy of H. subaxillaris and V. sinuatum against this highly resistant Gram-negative P. aeruginosa pathogen (Table 5).
2.6. Assessment of Antioxidant Capacities in Plant Extracts
Antioxidant activity measured by DPPH assay significantly varied across species (p < 0.05), with H. subaxillaris demonstrating the highest activity (80.1 ± 2.4%, p < 0.05) compared to the other plants, followed by V. sinuatum (53.9 ± 8.6%). A. spinosus and C. getulus showed moderate activity (32.5 ± 7.4% and 43.7 ± 9.5%, respectively) (Figure 9A).
2.7. Total Phenolics Content
Quantification of bioactive compounds revealed distinct compositional profiles among the four species (Figure 9). H. subaxillaris exhibited the highest total phenolic content (TPC = 1.9 ± 0.4 mg GAE/g DW), significantly surpassing A. spinosus (0.6 ± 0.3 mg GAE/g DW; p < 0.05) and showing comparable levels to V. sinuatum (1.5 ± 0.2 mg GAE/g DW) and C. getulus (1.3 ± 0.5 mg GAE/g DW; p > 0.05, F= 7.2). Notably, A. spinosus demonstrated consistently lower metabolite accumulation across both assays (Figure 9B).
2.8. Total Flavonoids Content
In the terms of total flavonoid content (TFC), H. subaxillaris (0.62 ± 0.2 mg QE/g DW) and V. sinuatum (0.61 ± 0.1 mg QE/g DW) formed a statistically homogeneous group (p = 0.87). In contrast, A. spinosus (0.27 ± 0.1 mg QE/g DW) showed significantly reduced values (p < 0.01). C. getulus displayed TFC (0.49 ± 0.2 mg QE/g DW), which did not differ significantly from either group (p > 0.05, F= 4.8) (Figure 9C).
3. Discussion
Medicinal herbs are the focus of interest as they may provide alternative pharmacological agents in the face of increasing antibiotic resistance and diseases caused by oxidative stress. In this study, the hexane crude extracts of V. sinuatum, A. spinosus, C. getulus, and H. subaxillaris were studied for their antioxidant and antimicrobial properties as well as phytochemical content.
3.1. GC-MS Analysis and Phytochemical Characterization
GC-MS profiling of the hexane extract of V. sinuatum revealed a phytochemical profile dominated by bioactive and lipophilic metabolites, confirming its use in traditional medicine and corroborating recent evidence of its biological potential. The extract was dominated by three main constituents: Hexadecanoic acid methyl ester, Phenol, 3,5-bis(1,1-dimethyl ethyl)- and a putative hordenine, each belonging to a different phytochemical class with documented pharmacological activity. The most abundant compound, hexadecanoic acid methyl ester, exhibits antibacterial, antioxidant, hypocholesterolemic, nematicidal, pesticidal, lubricating, hemolytic, and antiandrogenic activity [14,15]. Its lipophilicity enhances penetration into Gram-positive bacteria [16], which is consistent with the observed inhibition of Bacillus cereus and MRSA. Phenol,3,5-bis(1,1-dimethylethyl)- (a sterically hindered phenolic antioxidant) suggests a possible role in the free radical scavenging activity detected in the DPPH assay. The compound belongs to the family of tert-butylphenol (TBP) antioxidants (TBP-AO), which exhibit increased stability and antioxidant activity due to their massive tert-butyl substituents that hinder oxidative degradation [17]. The extract contained lower levels of 3,5-di-tert-butylphenol, a compound known to inhibit acid production and biofilm formation in Streptococcus mutans [18], as well as certain sesquiterpenes that exhibit antioxidant, prooxidant, and other biological activities [19]. 3,5-ditert-butylphenol is a phenolic antioxidant characterized by two tert-butyl substituents [17]. Tert-butyl phenol (TBP), which consists of a phenol ring bonded to a tert-butyl group, is widely utilized as an antioxidant, UV stabilizer, and chemical precursor, owing to its high stability [20]. Its identification in V. sinuatum confirms the plant's high DPPH radical scavenging activity. Identifying a hordenine adds a neuroactive and potentially bioactive property to the phytochemical profile of V. sinuatum [21]. Hordenine and its related alkaloids are associated with central nervous system stimulation, antimicrobial activity and enzyme inhibition [22,23,24]. Although the spectral agreement was relatively low, their abundance is consistent with the documented ability of various medicinal plants to biosynthesize nitrogenous secondary metabolites, especially under stress conditions [25]. Ecologically, the presence of fat-soluble fatty acid esters and phenolic antioxidants could indicate an adaptation to abiotic stress, especially to oxidative environments or UV irradiation in arid or semi-arid habitats [26,27]. The phytochemical composition of V. sinuatum, with a high proportion of lipophilic esters and phenols, distinguishes it from the other species in this study, such as H. subaxillaris, which, on the contrary, store methylated polyacetylenes and sesquiterpenoids as diagnostic metabolites. These differences illustrate Verbascum species' interspecific metabolic specialization and ecological plasticity [28]. From a chemotaxonomic basis, phenolics, fatty acid esters, and putative alkaloids, make V. sinuatum a metabolically complex species [29]. These compounds most likely enhance protection against pathogens and oxidative stress, with bioactivity directed toward multiple targets [30]. This is consistent with natural product drug discovery trends that focus on the synergy of chemically distinct classes in crude extracts rather than single compound therapy. The hexane extract of A. spinosus contained a complex mixture of lipophilic constituents, falling into three major chemical categories: oxygenated monoterpenes, aromatic compounds, and fatty acid derivatives. Six compounds were found to be the major constituents: Linalyl acetate, 3,7-Dimethyloct-6-en-1-ol, Geraniol, 9-Octadecenoic acid, methyl ester, 11,14-Octadecadienoic acid, methyl ester, and Isocitronellol. The extract contained high levels of oxygenated monoterpenes, with citronellol and geraniol being the predominant compounds. Antimicrobial activities were attributed to these acyclic monoterpene alcohols, possibly through membrane-disrupting mechanisms [31,32]. High concentrations of isocitronellol, a structural isomer of 3,7-Dimethyloct-6-en-1-ol, and linalyl acetate, an ester of linalool [33], were also present in the extract. This is consistent with the moderate antibacterial activity previously reported for A. spinosus against MRSA and E. coli O157:H7 detected in our bioassays. The fatty acid derivatives were characterized as 9-octadecenoic acid, methyl ester, and 11,14-octadecadienoic acid, methyl ester. These molecules are responsible for the extract's lipophilic properties and can increase membrane permeability in the target microorganisms [34]. Such actions may be partially responsible for the extract's moderate activity against MRSA and its weak but measurable effect. The C. getulus hexane extract exhibited a dense array of lipophilic constituents that fall into four general categories: (1) monoterpenoids were responsible for citronellol and geraniol, (2) diterpenoids such as phytol and isophytol; (3) fatty acid derivatives such as methyl palmitate, oleic acid, arachidonic acid and methyl undecanoate; and (4) phenolic compounds such as 3,5-ditert-butylphenol and thymol. This concordance in composition suggests a potential antioxidant [35], synergistic antimicrobial effect [36,37,38,39,40] and a dual ecological role in both direct inhibition of pathogens and plant defense signaling [41]. The hexane extract of H. subaxillaris comprised a diverse array of lipophilic compounds classified into three main categories: (1) terpenoids, including the monoterpenoid geraniol and the sesquiterpenoid cubebol; (2) unique fatty acid derivatives such as methyl 8-methyl nonanoate, hexadecanoic acid methyl ester, and the polyacetylenic esters falcarinol, 13,16-octadecadienoic acid methyl ester and 2,5-octadecadienoic acid methyl ester; and (3) phenols represented by Phenol,3,5-bis(1,1-dimethyl ethyl)-. The extract was characterized by high levels of terpenoids and rare acetylenic fatty acid esters, which are rarely found in plants. In recent years, numerous novel terpenes and terpenoids have been isolated or synthesized, leading to the discovery of further new terpene-derived compounds with potential chemotherapeutic activity, some of which are already being tested in clinical trials [42]. Monoterpenes and sesquiterpenes are released from aerial parts of plants, while others are produced as part of the plant’s defense mechanisms [43,44,45]. The simultaneous occurrence of branched-chain (methyl 8-methyl-nonanoate) and polyacetylene derivatives of classical fatty acid esters (palmitate) in the species indicates a higher-order lipid metabolism. The health-promoting effects are caused by these compounds with high biological activities [46,47,48]. Scientific studies have shown that fatty acid esters of hydroxy fatty acids have anti-diabetes [49,50,51,52], anti-cancer [53,54], anti-inflammatory [51,55], cardiovascular protective [56] and hepatoprotective activities [57,58] in mammals [59]. The presence of methyl 8-methyl-nonanoate in H. subaxillaris could indicate similar roles in lipid metabolism modulation, which is worthy of investigating its ecological or pharmaceutical role. The phenolic antioxidant 3,5-di-tert-butylphenol will possess a further protective role in shielding from oxidative stress [60].
3.2. Comparative Phytochemical and Pharmacological Profiling of Four Medicinal Plant Species
The phytochemical profile of H. subaxillaris stands out among the four species for its unique combination of sesquiterpenoid alcohols, polyacetylenic fatty acid esters, and phenolic antioxidants, in contrast to the dominance of monoterpenoids in A. spinosus, the simpler phenolic ester profile of V. sinuatum and the diterpenoid-phenol synergy of C. getulus. The acetylenic compounds, which are rarely found in the other species, reflect specialized adaptations at the ecological level, e.g., survival on dry land or protection against pathogens [61], while the high content of geraniol indicates an insect-repellent effect [62]. Pharmacologically, the diverse profile is reflected in the greater antibacterial efficacy of H. subaxillaris due to synergistic membrane disruption by terpenoid-polyacetylene mixtures [63]. A. spinosus is less effective antibacterially but has a composition enriched with terpenoids and fatty acid esters, to which anti-inflammatory and hepatoprotective activities are attributed [64]. V. sinuatum is particularly niche in its high methyl palmitate content and phenolic esters, consistent with traditional respiratory use and moderate antibacterial potency [65]. In parallel, C. getulus combines diterpenoids with antimicrobial phenolics, a novel strategy that suggests anti-inflammatory potential and previously untapped synergies against resistant pathogens [66]. The chemically diverse matrix of H. subaxillaris- a balance between volatile defenses and lipid-based storage- suggests its pharmacognostic potential. At the same time, the other species offer complementary bioactivities that merit targeted investigation.
3.3. Total Phenolics Content and Total Flavonoids Content
Quantitative phytochemical profiling showed significant differences in the accumulation of total phenolic and flavonoid compounds in the four medicinal plant species. Among them, H. subaxillaris showed the highest TPC content, significantly higher than A. spinosus and with comparable values to V. sinuatum and C. getulus. Polyphenols are a large group of secondary metabolites that have an important function in the detoxification of hydrogen peroxide from plant cells and make an important contribution to the cellular antioxidant defense system [67]. Phenolic compounds are well-known as key factors in plant defense and pharmacological action and function as antioxidants, metal chelators, and enzyme modulators [68]. Their hydroxylated aromatic scaffolds facilitate the effective neutralization of free radicals, disrupt bacterial membranes, and inhibit microbial enzyme systems [69,70,71]. Their high TPC content in H. subaxillaris and V. sinuatum is likely the basis for their greater activity in free radical scavenging assays. It contributes synergistically to their antimicrobial activities, as our GC-MS evidence shows for detecting several phenolic and terpenoid metabolites. H. subaxillaris and V. sinuatum showed statistically insignificant values, which placed them in the high flavonoid category. Flavonoids, a large subgroup of phenolic compounds, exhibit various bioactivities ranging from antioxidants and antimicrobial and anti-inflammatory effects to inhibition of enzymes [72]. Various food plants have flavonoids, which are considered important nutritional components for humans [73,74]. Flavonoids are one of the most ubiquitous secondary molecules that greatly benefit humans, not only because of their function in plant coloration but also because of their various physiologically active constituents [75]. The finding of the beneficial effects of flavonoids, such as cancer prevention, has generated much research interest, including consuming foods containing flavonoids [76]. Their planar polyphenolic molecular structure enables interactions with bacterial proteins and nucleic acids and confers versatile antimicrobial activities. Polyphenols from plant parts act against bacteria via proteins, DNA, cell wall, membrane and mechanisms based on energy metabolism are involved [77]. C. getulus had a medium flavonoid concentration, and its TPC and TFC values were only moderately high and did not show a significant difference from the high and low groups. The moderately high TPC and TFC values in conjunction with the GC-MS profile dominated by lipophilic terpenoids such as phytol and sesquiterpenes suggest that non-phenolic lipophilic constituents may be primarily responsible for the reported bioactivities of C. getulus. In contrast, A. spinosus exhibited the lowest levels of phenolics and flavonoids, consistent with its relatively weak antioxidant and antibacterial activity. Such a lack of phytochemicals could be caused by lower expression of genes for phenylpropanoid biosynthesis or by ecological traits that are less reliant on polyphenolic defense mechanisms. Interestingly, despite low levels of TPC and TFC, A. spinosus was characterized by high levels of monoterpenoid alcohols and fatty acid esters, suggesting a metabolic shift towards volatile or lipid-based defense mechanisms. Overall, these results emphasize the importance of both the chemical class and the concentration of the compound for the prediction of biological activity. While the levels of phenols and flavonoids are strongly correlated with antioxidant activity, the relationship with antimicrobial activity appears to be more complex and less direct. It often depends on a synergistic relationship between phenols, terpenoids, and fatty acid derivatives. The phytochemical richness shown here demonstrates the chemotaxonomic uniqueness of each species and confirms their different uses in ethnopharmacological practice.
3.4. Antioxidant Activity
Plants are a major source of natural antioxidants, with many of their bioactive properties attributed to the presence of hydroxyl groups in their structures [78]. The strongest free radical scavenging activity, based on the DPPH assay, was observed in H. subaxillaris, followed by V. sinuatum. This superior activity correlates with their high total TPC and TFC. Phenolic compounds identified in V. sinuatum such as Phenol,2,4-bis(1,1-dimethylethyl)-, are recognized for their free radical scavenging and antitumor properties [79]. The antioxidant activity of 2,4-bis(1,1-dimethylethyl)- is attributed to its tert-butyl groups, which enhance the stability of the aromatic hydroxyl group by facilitating phenoxyl radical formation and donating hydrogen atoms to neutralize active free radicals to stop lipid peroxidation [80,81]. This observation was supported by a previous study through in silico molecular docking analysis [82]. The antioxidant activity of H. subaxillaris may also stem from its unique polyacetylenes, including monoterpenoid alcohols, sesquiterpenoid alcohols, and falcarinol, which have been reported to diminish oxidative stress[83,84,85]. Furthermore, the synergistic interaction between citronellol and geraniol emphasizes their potential role as prooxidants [86]. In contrast, the comparatively lower antioxidant activity observed in A. spinosus and C. getulus is consistent with the lower TPC and TFC levels. These results emphasize the critical role of flavonoids and related polyphenols in contributing to the free radical scavenging activity observed in medicinal plants [87,88].
3.5. Antimicrobial Activity
The hexane extracts of H. subaxillaris, A. spinosus, V. sinuatum, and C. getulus exhibited measurable inhibitory effects against four clinically significant bacterial strains, including B. cereus, MRSA, E. coli (O157:H7 serotype), and P. aeruginosa. Notably, B. cereus and MRSA were more susceptible to these lipophilic plant extracts compared to E. coli O157:H7 and P. aeruginosa, consistent with previous findings that Gram-positive bacteria are more sensitive to plant-based hydrophobic compounds [89,90,91,92]. Among all extracts, H. subaxillaris demonstrated the most significant antibacterial activity across all tested strains, followed by V. sinuatum, C. getulus, and A. spinosus. While H. subaxillaris and V. sinuatum were effective against both Gram-negative and Gram-positive bacteria, C. getulus and A. spinosus exhibited markedly weaker activity, especially against P. aeruginosa, a pathogen known for its multidrug resistance, biofilm formation, and active efflux mechanisms [93,94,95]. Phytochemicals are influenced by their chain length, degree of unsaturation, and isomerism, with cis-isomers generally exhibiting more potent activity than their trans counterparts [96]. For example, cis-6-hexadecenoic acid showed inhibitory effects at low concentrations by disrupting the proton gradient, altering membrane fluidity, and inhibiting electron transport [97]. Additionally, esterified fatty acids frequently show enhanced antibacterial potency compared to their free acid counterparts, likely contributing to the effectiveness of these extracts [98]. The vigorous antibacterial activity of H. subaxillaris is likely attributed to its unique phytochemical profile, which includes sesquiterpenoid and monoterpenoid alcohols (with known membrane-disrupting properties and anti-inflammatory properties) [99,100,101,102] , polyunsaturated and branched-chain esters (which interfere with bacterial cell wall synthesis and trigger autolysis through endogenous enzyme activation) [96], and phenolic compounds (which induce oxidative stress and cause protein denaturation) [99,100,101,103,104]. These multi-targeted effects likely synergistically enhance antimicrobial efficacy while potentially reducing resistance development. V. sinuatum also exhibited noteworthy antibacterial activity, particularly against B. cereus and MRSA. This may be due to its phytochemical composition, which includes saturated fatty acid esters, phenolic antioxidants, and alkaloids [105]. While these components share some antimicrobial properties with those in H. subaxillaris, their distinct molecular structures may offer complementary or even unique modes of action. The exceptional performance of H. subaxillaris, followed by V. sinuatum, highlights their potential for further investigation in natural product-based drug discovery targeting resistant pathogens. The synergistic combination of diverse bioactive constituents in these crude extracts may offer a strategic advantage over monotherapeutic agents in combating bacterial resistance.
4. Materials and Methods
4.1. Collection and Identification of Plant Material
Fresh and healthy leaves of various plant species, including V. sinuatum L., A. spinosus L., C. getulus Pomel and H. subaxillaris (Lam.) Britt. & Rusby were collected at various locations in the Gaza Strip, Palestine, during the flowering period between March and July 2023. The Gaza Strip (31°25′N, 34°20′E), an arid coastal region on the south-eastern Mediterranean coast, is about 365 km² in extent. With around 2.3 million inhabitants, it is one of the most densely populated regions in the world. It is bordered to the southwest by Egypt (11 km) and to the east and north by the occupied Palestinian territories (51 km). The climate is Mediterranean, with hot summers (average 25°C), mild winters (13°C) and between 200 mm in the south and 400 mm in the north per year. Plant species were taxonomically identified by the Botany Department, Al-Aqsa University, Gaza. The leaves were washed with tap water, air-dried at room temperature for ten days, then ground and stored in a cool, dry environment. Phytochemical studies were carried out at the Analytical Chemistry and Desert Soils Laboratories of Cairo University Research Park, Faculty of Agriculture, Cairo University, Egypt.
Figure 10.
Illustrative photographs of the studied plant species recorded in the Gaza Strip, Palestine: (A) V. sinuatum; (B) A. spinosus; (C) C. getulus; (D) H. subaxillaris. (https://flora.org.il/en/plants).
Figure 10.
Illustrative photographs of the studied plant species recorded in the Gaza Strip, Palestine: (A) V. sinuatum; (B) A. spinosus; (C) C. getulus; (D) H. subaxillaris. (https://flora.org.il/en/plants).

4.2. Extraction of Plant Material
Ten grams (g) of each sample of leaf powder was extracted in hexane, ethyl acetate and methanol (1:10 w/v) using a Soxhlet apparatus for 6 hours for each solvent. The resulting filtrates were concentrated under reduced pressure at 45 °C, and the remaining solvents were evaporated under nitrogen stream. The extracts were stored at −20 °C in amber vials for later analysis [106]
4.3. Gas Chromatography-Mass Spectrometry Analysis
The chemical profile was generated on an Agilent 7000 Triple Quadrupole GC–MS instrument using an Elite-5MS column. The analytical conditions were electron ionization (70 eV), helium as carrier gas (1 mL/min, 30:1 split) and a temperature ramp from 110 °C to 280 °C in 36 minutes. Mass spectra in the range of m/z 45–450 were matched to the NIST library, and compound abundance was determined from the percent peak area using Turbomass software [107].
4.4. Analysis and Characterization of Compounds of Plant Extracts
A combined approach identified the compounds: spectral matching with the mass spectral libraries of NIST (2017) and Wiley (11th edition), calculation of the retention index (RI) in comparison to n-alkanes (C8–C40). Only compounds with > 70% agreement were considered confidently identified. Quantification was based on individual peak areas (% of total ion chromatogram), processed using Mass Hunter Qualitative Analysis, which provided comprehensive results, including retention times, peak areas, and fragmentation profiles. Triplicate analysis of leaf extracts under identical conditions enabled reproducibility with a relative standard deviation (RSD) of less than 5%. Sample preparation, extract treatment, and GC-MS analysis were combined in one step for data interpretation based on spectral libraries, mass Fragmentation, and RI confirmation.
4.5. Measurement of Total Phenolic Content
Total phenolic compounds in hexane extracts were quantitated by Folin–Ciocalteu assay [108]. 0.5 g of fresh material was extracted with 10 mL hexane, filtered, and stored in refrigerator for a week. A gallic acid stock solution (100,000 ppm) was prepared, and a calibration curve (10–200 ppm) was prepared. To determine, 200 μL of the extract was mixed with 400 μL of the 10% Folin–Ciocalteu reagent, followed by 800 μL of 10% Na₂CO₃ after 3 minutes. Samples, standards, and blanks were stored in the dark for 1 hour. The absorbance at 725 nm was measured. The TPC from the standard curve was calculated and expressed as % gallic acid equivalents (%GAE) as follows: TPC (%GAE) = (standard curve concentration/sample weight) × dilution factor × 10,000
4.6. Measurement of Total Flavonoid Content
Total flavonoids were quantitated according to the aluminum chloride colorimetric assay reported by [109]). Plant material (0.5 g) was homogenized in hexane (10 mL) and filtered. A quercetin stock solution (1000 ppm) was prepared (0.1 g in 100 mL), and a working solution (100 ppm) was diluted. A standard curve (20–640 ppm) was created from the standard solutions. For the assay, 125 μL of extract was mixed with NaNO₂ (75 μL of a 5% solution) and AlCl₃ (150 μL of a 10% solution), followed by NaOH (750 μL of 1 M ) addition, and the volume adjusted to 2.5 mL. Standard solutions and blanks underwent the same treatment. After 15 min in darkness, absorbance was measured at 510 nm. Flavonoid content was calculated as quercetin equivalents (%QE) using a calibration curve and the formula: TFC (%QE) = (standard curve concentration/sample weight) × dilution factor × 10,000.
4.7. DPPH Radical Scavenging Assay
Antioxidant activity of the extracts was assessed using the DPPH (1,1-diphenyl-2-picrylhydrazyl) method on the basis of violet DPPH radical reducibility to the yellow DPPH-H form when interacting with antioxidants. All the tested extracts at various concentrations were examined. In brief, 0.5 mL of DPPH solution (50 mg/100 mL) was mixed with 4.5 mL of hexane, and 0.1 mL of extract was added. The mixtures were shaken and incubated in the dark for 45 minutes and subsequently assayed for 515 nm absorbance against a blank [110].
4.8. Measurement of Antimicrobial Activity
The antimicrobial activity of hexane crude extracts was tested against two Gram-positive bacteria (Staphylococcus aureus MRSA and Bacillus cereus) and two Gram-negative bacteria (Escherichia coli O157:H7 and Pseudomonas aeruginosa) from Cairo University bacterial culture collection using the agar well diffusion method [111]. Bacterial cultures were grown to logarithmic phase at 37°C in nutrient broth, adjusted to 1–5 × 10⁵ CFU/mL, and inoculated onto Mueller-Hinton agar plates. Wells (6 mm) were filled with 100 µL of each extract (dissolved in ≤1% DMSO), and the plates were incubated at 37°C for 24 hours. Zones of inhibition were measured with a digital caliper. Streptomycin (10 µg/mL) served as a positive control, and DMSO (1% v/v) as a negative control.
4.9. Statistical Analysis
All experimental data were analyzed using GraphPad Prism 9.0 (GraphPad Software, San Diego, CA, USA). Continuous variables were tested for normality distribution using the Shapiro-Wilk test. Parametric data were compared using a one-way ANOVA followed by Dunnett's test. Results are presented as mean ± SD (standard deviation) of at least three independent experiments. Statistical significance was set at p < 0.05.
5. Conclusions
This study provides a comprehensive phytochemical analysis of four Mediterranean medicinal plants (V. sinuatum, A. spinosus, C. getulus, and H. subaxillaris), revealing a rich diversity of lipophilic metabolites. H. subaxillaris emerged as the most potent antibacterial and antioxidant agent, linked to its unique combination of sesquiterpenoids, polyacetylenic fatty acids, and phenolic antioxidants. V. sinuatum and C. getulus also demonstrated considerable activity due to their phenolic and diterpenoid profiles. The weak performance of A. spinosus underscores the role of phytochemical diversity in bioactivity. These findings underscore the value of phytochemical profiling for bioprospecting and pave the way for future pharmacognostic exploration of structurally diverse natural compounds from underexplored medicinal flora. Future work should isolate key metabolites (e.g., falcarinol, methyl 8-methyl-nonanoate) to mechanisms of action and elucidate structure-activity relationships.
Author Contributions
Conceptualization, M.A.A; methodology, M.A.A and M.E.; software, M.E.; validation, M.A.A and M.E.; formal analysis, M.A.A and M.E.; investigation, M.A.A and M.E.; resources, M.A.A.; data curation, M.E.; writing—original draft preparation, M.A.A. and M.E.; writing—review and editing, X.X.; visualization, X.X.; supervision, X.X.; project administration, X.X.; funding acquisition, M.A.A. and M.E.. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Data are contained within the article.
Acknowledgments
The authors thank colleagues from the Faculty of Agriculture Research Park, Cairo University, for assistance with lab analyses and support in this study. Appreciation is also extended to Dr. Ahmed K. Junina, Dr. Adham M. Abu Hatab, and Dr. Sorowar Chowdhury for their help in improving the manuscript's language and academic quality.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Dubale, S.; Kebebe, D.; Zeynudin, A.; Abdissa, N.; Suleman, S. Phytochemical Screening and Antimicrobial Activity Evaluation of Selected Medicinal Plants in Ethiopia. J. Exp. Pharmacol. 2023, 15, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Selseleh, M.; Nejad Ebrahimi, S.; Aliahmadi, A.; Sonboli, A.; Mirjalili, M.H. Metabolic profiling, antioxidant, and antibacterial activity of some Iranian Verbascum L. species. Industrial Crops and Products 2020, 153, 112609. [Google Scholar] [CrossRef]
- Donn, P.; Barciela, P.; Perez-Vazquez, A.; Cassani, L.; Simal-Gandara, J.; Prieto, M.A. Bioactive Compounds of Verbascum sinuatum L.: Health Benefits and Potential as New Ingredients for Industrial Applications. Biomolecules 2023, 13, 427. [Google Scholar] [CrossRef] [PubMed]
- Ruth, O.N.; Unathi, K.; Nomali, N.; Chinsamy, M. Underutilization Versus Nutritional-Nutraceutical Potential of the Amaranthus Food Plant: A Mini-Review. Applied Sciences 2021, 11, 6879. [Google Scholar] [CrossRef]
- Adegbola, P.I.; Adetutu, A.; Olaniyi, T.D. Antioxidant activity of Amaranthus species from the Amaranthaceae family – A review. South African Journal of Botany 2020, 133, 111–117. [Google Scholar] [CrossRef]
- Mamuru, S.; Kaigamma, I.; Muluh, E. Preliminary Phytochemical Screening and GC-MS Analysis of Aqueous and Ethanolic Extracts of Amaranthus spinosus Leaves. Journal of Natural Products and Resources 2019, 5, 213–215. [Google Scholar] [CrossRef]
- Abou Auda, M. Phytochemical and proximate analysis of wild plants from the Gaza Strip, Palestine. Biodiversitas Journal of Biological Diversity 2025, 26. [Google Scholar] [CrossRef]
- Cuervo, W.; Larrauri, M.; Gomez-Lopez, C.; DiLorenzo, N. Invasive Pigweed (Amaranthus spinosus) as a Potential Source of Plant Secondary Metabolites to Mitigate Enteric Methane Emissions in Beef Cattle. Grasses 2025, 4, 14. [Google Scholar] [CrossRef]
- Al-Shammari, L.A.; Hassan, W.H.B.; Al-Youssef, H.M. Phytochemical and biological studies of Carduus pycnocephalus L. Journal of Saudi Chemical Society 2015, 19, 410–416. [Google Scholar] [CrossRef]
- Abou Auda, M. Taxonomical knowledge, biological spectra and ethnomedicinal plant inventory of Asteraceae family in various areas of Gaza strip, Palestine. Pak. J. Bot 2023, 55, 2369–2380. [Google Scholar] [CrossRef]
- Morimoto, M.; Cantrell, C.L.; Libous-Bailey, L.; Duke, S.O. Phytotoxicity of constituents of glandular trichomes and the leaf surface of camphorweed, Heterotheca subaxillaris. Phytochemistry 2009, 70, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Rajasekar, R.; Thanasamy, R.; Samuel, M.; Edison, T.N.J.I.; Raman, N. Ecofriendly synthesis of silver nanoparticles using Heterotheca subaxillaris flower and its catalytic performance on reduction of methyl orange. Biochem. Eng. J. 2022, 187, 108447. [Google Scholar] [CrossRef]
- Sternberg, M. From America to the Holy Land: disentangling plant traits of the invasive Heterotheca subaxillaris (Lam.) Britton & Rusby. Plant Ecology 2016, 217, 1307–1314. [Google Scholar] [CrossRef]
- Desbois, A.P.; Smith, V.J. Antibacterial free fatty acids: activities, mechanisms of action and biotechnological potential. Appl. Microbiol. Biotechnol. 2010, 85, 1629–1642. [Google Scholar] [CrossRef] [PubMed]
- Krishnamoorthy, K.; Subramaniam, P. Phytochemical Profiling of Leaf, Stem, and Tuber Parts of Solena amplexicaulis (Lam.) Gandhi Using GC-MS. Int Sch Res Notices 2014, 2014, 567409. [Google Scholar] [CrossRef]
- Shaaban, M.T.; Ghaly, M.F.; Fahmi, S.M. Antibacterial activities of hexadecanoic acid methyl ester and green-synthesized silver nanoparticles against multidrug-resistant bacteria. J. Basic Microbiol. 2021, 61, 557–568. [Google Scholar] [CrossRef]
- Hoang, N.M.H.; Park, K. Applications of Tert-Butyl-Phenolic Antioxidants in Consumer Products and Their Potential Toxicities in Humans. Toxics 2024, 12, 869. [Google Scholar] [CrossRef]
- Vijayakumar, K.; MuhilVannan, S. 3, 5-Di-tert-butylphenol combat against Streptococcus mutans by impeding acidogenicity, acidurance and biofilm formation. World J. Microbiol. Biotechnol. 2021, 37, 202. [Google Scholar] [CrossRef]
- Bartikova, H.; Hanusova, V.; Skalova, L.; Ambroz, M.; Bousova, I. Antioxidant, pro-oxidant and other biological activities of sesquiterpenes. Curr. Top. Med. Chem. 2014, 14, 2478–2494. [Google Scholar] [CrossRef]
- Fiege, H.; Voges, H.W.; Hamamoto, T.; Umemura, S.; Iwata, T.; Miki, H.; Fujita, Y.; Buysch, H.J.; Garbe, D.; Paulus, W. Phenol derivatives. Ullmann's encyclopedia of industrial chemistry, 2000. [Google Scholar] [CrossRef]
- Su, Y.; Liu, Y.; He, D.; Hu, G.; Wang, H.; Ye, B.; He, Y.; Gao, X.; Liu, D. Hordenine inhibits neuroinflammation and exerts neuroprotective effects via inhibiting NF-κB and MAPK signaling pathways in vivo and in vitro. Int. Immunopharmacol. 2022, 108, 108694. [Google Scholar] [CrossRef]
- Zhang, X.; Du, L.; Zhang, J.; Li, C.; Zhang, J.; Lv, X. Hordenine protects against lipopolysaccharide-induced acute lung injury by inhibiting inflammation. Front. Pharmacol. 2021, 12, 712232. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-C.; Lee, J.-H.; Kim, M.-H.; Lee, J.-A.; Kim, Y.B.; Jung, E.; Kim, Y.-S.; Lee, J.; Park, D. Hordenine, a single compound produced during barley germination, inhibits melanogenesis in human melanocytes. Food Chem. 2013, 141, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Zhang, Q.; Ding, C.; Wen, F.; Sun, F.; Liu, Y.; Tao, C.; Yao, J. Beneficial Effects of Hordenine on a Model of Ulcerative Colitis. Molecules 2023, 28, 2834. [Google Scholar] [CrossRef] [PubMed]
- Jamwal, K.; Bhattacharya, S.; Puri, S. Plant growth regulator mediated consequences of secondary metabolites in medicinal plants. Journal of Applied Research on Medicinal and Aromatic Plants 2018, 9, 26–38. [Google Scholar] [CrossRef]
- Rao, M.J.; Zheng, B. The Role of Polyphenols in Abiotic Stress Tolerance and Their Antioxidant Properties to Scavenge Reactive Oxygen Species and Free Radicals. Antioxidants 2025, 14, 74. [Google Scholar] [CrossRef]
- Mishra, N.; Jiang, C.; Chen, L.; Paul, A.; Chatterjee, A.; Shen, G. Achieving abiotic stress tolerance in plants through antioxidative defense mechanisms. Frontiers in Plant Science 2023, 14, 1110622. [Google Scholar] [CrossRef]
- Seipel, T.; Alexander, J.M.; Daehler, C.C.; Rew, L.J.; Edwards, P.J.; Dar, P.A.; McDougall, K.; Naylor, B.; Parks, C.; Pollnac, F.W. Performance of the herb Verbascum thapsus along environmental gradients in its native and non-native ranges. J. Biogeogr. 2015, 42, 132–143. [Google Scholar] [CrossRef]
- Georgiev, M.I.; Ali, K.; Alipieva, K.; Verpoorte, R.; Choi, Y.H. Metabolic differentiations and classification of Verbascum species by NMR-based metabolomics. Phytochemistry 2011, 72, 2045–2051. [Google Scholar] [CrossRef]
- Saini, N.; Anmol, A.; Kumar, S.; Wani, A.W.; Bakshi, M.; Dhiman, Z. Exploring phenolic compounds as natural stress alleviators in plants- a comprehensive review. Physiol. Mol. Plant Pathol. 2024, 133, 102383. [Google Scholar] [CrossRef]
- Jayaraj, R.L.; Azimullah, S.; Parekh, K.A.; Ojha, S.K.; Beiram, R. Effect of citronellol on oxidative stress, neuroinflammation and autophagy pathways in an in vivo model of Parkinson's disease. Heliyon 2022, 8, e11434. [Google Scholar] [CrossRef]
- Chen, W.; Viljoen, A.M. Geraniol – A review update. South African Journal of Botany 2022, 150, 1205–1219. [Google Scholar] [CrossRef]
- Peana, A.T.; D'Aquila, P.S.; Panin, F.; Serra, G.; Pippia, P.; Moretti, M.D.L. Anti-inflammatory activity of linalool and linalyl acetate constituents of essential oils. Phytomedicine 2002, 9, 721–726. [Google Scholar] [CrossRef] [PubMed]
- Dembitsky, V.M.; Shkrob, I.; Rozentsvet, O.A. Fatty acid amides from freshwater green alga Rhizoclonium hieroglyphicum. Phytochemistry 2000, 54, 965–967. [Google Scholar] [CrossRef] [PubMed]
- Javan, A.J.; Javan, M.J. Electronic structure of some thymol derivatives correlated with the radical scavenging activity: Theoretical study. Food Chem. 2014, 165, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Tlais, A.Z.A.; Rantsiou, K.; Filannino, P.; Cocolin, L.S.; Cavoski, I.; Gobbetti, M.; Di Cagno, R. Ecological linkages between biotechnologically relevant autochthonous microorganisms and phenolic compounds in sugar apple fruit (Annona squamosa L.). Int. J. Food Microbiol. 2023, 387, 110057. [Google Scholar] [CrossRef]
- Sytar, O.; Hemmerich, I.; Zivcak, M.; Rauh, C.; Brestic, M. Comparative analysis of bioactive phenolic compounds composition from 26 medicinal plants. Saudi J. Biol. Sci. 2018, 25, 631–641. [Google Scholar] [CrossRef]
- Kasmi, S.; Hamdi, A.; Atmani-Kilani, D.; Debbache-Benaida, N.; Jaramillo-Carmona, S.; Rodríguez-Arcos, R.; Jiménez-Araujo, A.; Ayouni, K.; Atmani, D.; Guillén-Bejarano, R. Characterization of phenolic compounds isolated from the Fraxinus angustifolia plant and several associated bioactivities. Journal of Herbal Medicine 2021, 29, 100485. [Google Scholar] [CrossRef]
- Hossain, M.A.; Shah, M.D.; Gnanaraj, C.; Iqbal, M. In vitro total phenolics, flavonoids contents and antioxidant activity of essential oil, various organic extracts from the leaves of tropical medicinal plant Tetrastigma from Sabah. Asian Pac. J. Trop. Med. 2011, 4, 717–721. [Google Scholar] [CrossRef]
- Guadie, A.; Dakone, D.; Unbushe, D.; Wang, A.; Xia, S. Antibacterial activity of selected medicinal plants used by traditional healers in Genta Meyche (Southern Ethiopia) for the treatment of gastrointestinal disorders. Journal of Herbal Medicine 2020, 22, 100338. [Google Scholar] [CrossRef]
- Chauhan, K.R.; Le, T.C.; Chintakunta, P.K.; Lakshman, D.K. Phyto-fungicides: Structure activity relationships of the thymol derivatives against Rhizoctonia solani. Journal of Agricultural Chemistry and Environment 2017, 6, 175. [Google Scholar] [CrossRef]
- Zhu, W.; Liu, X.; Wang, Y.; Tong, Y.; Hu, Y. Discovery of a novel series of α-terpineol derivatives as promising anti-asthmatic agents: Their design, synthesis, and biological evaluation. Eur. J. Med. Chem. 2018, 143, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Niinemets, Ü.; Monson, R.K. Biology, controls and models of tree volatile organic compound emissions. 2013. [Google Scholar] [CrossRef]
- Lange, B.M. The evolution of plant secretory structures and emergence of terpenoid chemical diversity. Annu. Rev. Plant Biol. 2015, 66, 139–159. [Google Scholar] [CrossRef] [PubMed]
- Staudt, M.; Lhoutellier, L. Volatile organic compound emission from holm oak infested by gypsy moth larvae: evidence for distinct responses in damaged and undamaged leaves. Tree physiology 2007, 27, 1433–1440. [Google Scholar] [CrossRef]
- Yore, Mark M.; Syed, I.; Moraes-Vieira, Pedro M.; Zhang, T.; Herman, Mark A.; Homan, Edwin A.; Patel, Rajesh T.; Lee, J.; Chen, S.; Peroni, Odile D.; et al. Discovery of a Class of Endogenous Mammalian Lipids with Anti-Diabetic and Anti-inflammatory Effects. Cell 2014, 159, 318–332. [CrossRef]
- Patel, R.; Santoro, A.; Hofer, P.; Tan, D.; Oberer, M.; Nelson, A.T.; Konduri, S.; Siegel, D.; Zechner, R.; Saghatelian, A.; et al. ATGL is a biosynthetic enzyme for fatty acid esters of hydroxy fatty acids. Nature 2022, 606, 968–975. [Google Scholar] [CrossRef]
- Riecan, M.; Paluchova, V.; Lopes, M.; Brejchova, K.; Kuda, O. Branched and linear fatty acid esters of hydroxy fatty acids (FAHFA) relevant to human health. Pharmacol. Ther. 2022, 231, 107972. [Google Scholar] [CrossRef]
- Syed, I.; Lee, J.; Moraes-Vieira, P.M.; Donaldson, C.J.; Sontheimer, A.; Aryal, P.; Wellenstein, K.; Kolar, M.J.; Nelson, A.T.; Siegel, D.; et al. Palmitic Acid Hydroxystearic Acids Activate GPR40, Which Is Involved in Their Beneficial Effects on Glucose Homeostasis. Cell Metab. 2018, 27, 419–427.e414. [Google Scholar] [CrossRef] [PubMed]
- Muoio, D.M.; Newgard, C.B. The good in fat. Nature 2014, 516, 49–50. [Google Scholar] [CrossRef]
- Moraes-Vieira, P.M.; Saghatelian, A.; Kahn, B.B. GLUT4 expression in adipocytes regulates de novo lipogenesis and levels of a novel class of lipids with antidiabetic and anti-inflammatory effects. Diabetes 2016, 65, 1808–1815. [Google Scholar] [CrossRef]
- Syed, I.; de Celis, M.F.R.; Mohan, J.F.; Moraes-Vieira, P.M.; Vijayakumar, A.; Nelson, A.T.; Siegel, D.; Saghatelian, A.; Mathis, D.; Kahn, B.B. PAHSAs attenuate immune responses and promote β cell survival in autoimmune diabetic mice. The Journal of clinical investigation 2019, 129, 3717–3731. [Google Scholar] [CrossRef]
- Rodríguez, J.P.; Guijas, C.; Astudillo, A.M.; Rubio, J.M.; Balboa, M.A.; Balsinde, J. Sequestration of 9-Hydroxystearic Acid in FAHFA (Fatty Acid Esters of Hydroxy Fatty Acids) as a Protective Mechanism for Colon Carcinoma Cells to Avoid Apoptotic Cell Death. Cancers (Basel) 2019, 11, 524. [Google Scholar] [CrossRef]
- Zhu, Q.-F.; Yan, J.-W.; Gao, Y.; Zhang, J.-W.; Yuan, B.-F.; Feng, Y.-Q. Highly sensitive determination of fatty acid esters of hydroxyl fatty acids by liquid chromatography-mass spectrometry. Journal of Chromatography B 2017, 1061-1062, 34–40. [Google Scholar] [CrossRef]
- Kolar, M.J.; Konduri, S.; Chang, T.; Wang, H.; McNerlin, C.; Ohlsson, L.; Härröd, M.; Siegel, D.; Saghatelian, A. Linoleic acid esters of hydroxy linoleic acids are anti-inflammatory lipids found in plants and mammals. J. Biol. Chem. 2019, 294, 10698–10707. [Google Scholar] [CrossRef] [PubMed]
- Dongoran, R.A.; Lin, T.-J.; Byekyet, A.; Tang, S.-C.; Yang, J.-H.; Liu, C.-H. Determination of Major Endogenous FAHFAs in Healthy Human Circulation: The Correlations with Several Circulating Cardiovascular-Related Biomarkers and Anti-Inflammatory Effects on RAW 264.7 Cells. Biomolecules 2020, 10, 1689. [Google Scholar] [CrossRef] [PubMed]
- Benlebna, M.; Balas, L.; Bonafos, B.; Pessemesse, L.; Fouret, G.; Vigor, C.; Gaillet, S.; Grober, J.; Bernex, F.; Landrier, J.-F.; et al. Long-term intake of 9-PAHPA or 9-OAHPA modulates favorably the basal metabolism and exerts an insulin sensitizing effect in obesogenic diet-fed mice. Eur. J. Nutr. 2021, 60, 2013–2027. [Google Scholar] [CrossRef]
- Defour, M.; van Weeghel, M.; Hermans, J.; Kersten, S. Hepatic ADTRP overexpression does not influence lipid and glucose metabolism. American Journal of Physiology-Cell Physiology 2021, 321, C585–C595. [Google Scholar] [CrossRef]
- Minto, R.E.; Blacklock, B.J. Biosynthesis and function of polyacetylenes and allied natural products. Prog. Lipid Res. 2008, 47, 233–306. [Google Scholar] [CrossRef]
- Daniel, J.W.; Green, T.; Phillips, P.J. Metabolism of the phenolic antioxidant 3,5-Di-tert-butyl-4-hydroxyanisole (Topanol 354). II. Biotransformation in man, rat and dog. Food Cosmet. Toxicol. 1973, 11, 781–792. [Google Scholar] [CrossRef] [PubMed]
- Seigler, D.S. Acetylenic Compounds. In Plant Secondary Metabolism, Seigler, D.S., Ed.; Springer US: Boston, MA, 1998; pp. 42–50. [Google Scholar]
- Chen, W.; Viljoen, A.M. Geraniol — A review of a commercially important fragrance material. South African Journal of Botany 2010, 76, 643–651. [Google Scholar] [CrossRef]
- Avato, P.; Vitali, C.; Mongelli, P.; Tava, A. Antimicrobial activity of polyacetylenes from Bellis perennis and their synthetic derivatives. Planta Med. 1997, 63, 503–507. [Google Scholar] [CrossRef]
- Yao, P.; Liu, Y. Terpenoids: Natural Compounds for Non-Alcoholic Fatty Liver Disease (NAFLD) Therapy. Molecules 2023, 28, 272. [Google Scholar] [CrossRef]
- Yabalak, E.; Ibrahim, F.; Eliuz, E.A.E.; Everest, A.; Gizir, A.M. Evaluation of chemical composition, trace element content, antioxidant and antimicrobial activities of Verbascum pseudoholotrichum. Plant Biosystems-An International Journal Dealing with all Aspects of Plant Biology 2022, 156, 313–322. [Google Scholar] [CrossRef]
- Kozyra, M.; Komsta, Ł.; Wojtanowski, K. Analysis of phenolic compounds and antioxidant activity of methanolic extracts from inflorescences of Carduus sp. Phytochemistry Letters 2019, 31, 256–262. [Google Scholar] [CrossRef]
- Supritha, P.; Radha, K.V. Estimation of phenolic compounds present in the plant extracts using high pressure liquid chromatography, antioxidant properties and its antibacterial activity. Indian Journal of Pharmaceutical Education and Research 2018, 52, 321–326. [Google Scholar] [CrossRef]
- Sun, W.; Shahrajabian, M.H. Therapeutic Potential of Phenolic Compounds in Medicinal Plants—Natural Health Products for Human Health. Molecules 2023, 28, 1845. [Google Scholar] [CrossRef]
- Cui, Y.; Oh, Y.J.; Lim, J.; Youn, M.; Lee, I.; Pak, H.K.; Park, W.; Jo, W.; Park, S. AFM study of the differential inhibitory effects of the green tea polyphenol (−)-epigallocatechin-3-gallate (EGCG) against Gram-positive and Gram-negative bacteria. Food Microbiol. 2012, 29, 80–87. [Google Scholar] [CrossRef]
- Fathima, A.; Rao, J.R. Selective toxicity of Catechin—a natural flavonoid towards bacteria. Appl. Microbiol. Biotechnol. 2016, 100, 6395–6402. [Google Scholar] [CrossRef]
- Veiko, A.G.; Sekowski, S.; Lapshina, E.A.; Wilczewska, A.Z.; Markiewicz, K.H.; Zamaraeva, M.; Zhao, H.-c.; Zavodnik, I.B. Flavonoids modulate liposomal membrane structure, regulate mitochondrial membrane permeability and prevent erythrocyte oxidative damage. Biochimica et Biophysica Acta (BBA) - Biomembranes 2020, 1862, 183442. [Google Scholar] [CrossRef]
- Mutha, R.E.; Tatiya, A.U.; Surana, S.J. Flavonoids as natural phenolic compounds and their role in therapeutics: an overview. Future Journal of Pharmaceutical Sciences 2021, 7, 25. [Google Scholar] [CrossRef]
- Verri, W.A.; Vicentini, F.T.M.C.; Baracat, M.M.; Georgetti, S.R.; Cardoso, R.D.R.; Cunha, T.M.; Ferreira, S.H.; Cunha, F.Q.; Fonseca, M.J.V.; Casagrande, R. Chapter 9 - Flavonoids as Anti-Inflammatory and Analgesic Drugs: Mechanisms of Action and Perspectives in the Development of Pharmaceutical Forms. In Studies in Natural Products Chemistry, Atta ur, R., Ed.; Elsevier: 2012; Volume 36, pp. 297-330.
- Patil, V.M.; Masand, N. Chapter 12 - Anticancer Potential of Flavonoids: Chemistry, Biological Activities, and Future Perspectives. In Studies in Natural Products Chemistry, Atta ur, R., Ed.; Elsevier: 2018; Volume 59, pp. 401-430.
- Muhaisen, H.M. Introduction and interpretation of flavonoids. Advanced Science, Engineering and Medicine 2014, 6, 1235–1250. [Google Scholar] [CrossRef]
- Khajuria, R.; Singh, S.; Bahl, A. General introduction and sources of flavonoids. Current aspects of flavonoids: Their role in cancer treatment, -7. [CrossRef]
- Lobiuc, A.; Pavăl, N.-E.; Mangalagiu, I.I.; Gheorghiță, R.; Teliban, G.-C.; Amăriucăi-Mantu, D.; Stoleru, V. Future Antimicrobials: Natural and Functionalized Phenolics. Molecules 2023, 28, 1114. [Google Scholar] [CrossRef] [PubMed]
- Abbas, Z.K.; Saggu, S.; Sakeran, M.I.; Zidan, N.; Rehman, H.; Ansari, A.A. Phytochemical, antioxidant and mineral composition of hydroalcoholic extract of chicory (Cichorium intybus L.) leaves. Saudi J. Biol. Sci. 2015, 22, 322–326. [Google Scholar] [CrossRef]
- Rahman, M.M.; Hossain, A.S.M.S.; Mostofa, M.G.; Khan, M.A.; Ali, R.; Mosaddik, A.; Sadik, M.G.; Alam, A.H.M.K. Evaluation of anti-ROS and anticancer properties of Tabebuia pallida L. Leaves. Clinical Phytoscience 2019, 5, 17. [Google Scholar] [CrossRef]
- Ali, H.M.; Abo-Shady, A.; Sharaf Eldeen, H.A.; Soror, H.A.; Shousha, W.G.; Abdel-Barry, O.A.; Saleh, A.M. Structural features, kinetics and SAR study of radical scavenging and antioxidant activities of phenolic and anilinic compounds. Chem. Cent. J. 2013, 7, 53. [Google Scholar] [CrossRef]
- Zhao, F.; Wang, P.; Lucardi, R.D.; Su, Z.; Li, S. Natural Sources and Bioactivities of 2,4-Di-Tert-Butylphenol and Its Analogs. Toxins (Basel) 2020, 12, 35. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Reza, A.; Khan, Z.; Munira, M.S.; Khatoon, M.M.; Kabir, S.R.; Sadik, M.G.; Ağagündüz, D.; Capasso, R.; Kazi, M.; et al. Apoptosis-inducing anti-proliferative and quantitative phytochemical profiling with in silico study of antioxidant-rich Leea aequata L. leaves. Heliyon 2024, 10, e23400. [Google Scholar] [CrossRef]
- Yu, L.; Cheng, W.; Tian, M.; Wu, Z.; Wei, X.; Cheng, X.; Yang, M.; Ma, X. Antioxidant Activity and Volatile Oil Analysis of Ethanol Extract of Phoebe zhennan S. Lee et F. N. Wei Leaves. Forests 2024, 15, 236. [Google Scholar] [CrossRef]
- Medbouhi, A.; Merad, N.; Khadir, A.; Bendahou, M.; Djabou, N.; Costa, J.; Muselli, A. Chemical composition and biological investigations of Eryngium triquetrum essential oil from Algeria. Chem. Biodivers. 2018, 15, e1700343. [Google Scholar] [CrossRef]
- Tel-Çayan, G.; Duru, M.E. Chemical characterization and antioxidant activity of Eryngium pseudothoriifolium and E. thorifolium essential oils. 2019. [Google Scholar] [CrossRef]
- Viuda-Martos, M.; López-Marcos, M.; Fernández-López, J.; Sendra, E.; López-Vargas, J.; Pérez-Álvarez, J. Role of fiber in cardiovascular diseases: a review. Compr Rev Food Sci Food Saf 9: 240–258. 2010. [Google Scholar] [CrossRef]
- Sharififar, F.; Dehghn-Nudeh, G.; Mirtajaldini, M. Major flavonoids with antioxidant activity from Teucrium polium L. Food Chem. 2009, 112, 885–888. [Google Scholar] [CrossRef]
- Khan, R.A.; Khan, M.R.; Sahreen, S.; Ahmed, M. Assessment of flavonoids contents and in vitro antioxidant activity of Launaea procumbens. Chem. Cent. J. 2012, 6, 43. [Google Scholar] [CrossRef]
- Benramdane, E.; Chougui, N.; Ramos, P.A.B.; Makhloufi, N.; Tamendjari, A.; Silvestre, A.J.D.; Santos, S.A.O. Lipophilic Compounds and Antibacterial Activity of Opuntia ficus-indica Root Extracts from Algeria. Int. J. Mol. Sci. 2022, 23, 11161. [Google Scholar] [CrossRef] [PubMed]
- Aydın Kurç, M.; Orak, H.H.; Gülen, D.; Caliskan, H.; Argon, M.; Sabudak, T. Antimicrobial and Antioxidant Efficacy of the Lipophilic Extract of Cirsium vulgare. Molecules 2023, 28, 7177. [Google Scholar] [CrossRef]
- Palmeri, R.; Parafati, L.; Arena, E.; Grassenio, E.; Restuccia, C.; Fallico, B. Antioxidant and Antimicrobial Properties of Semi-Processed Frozen Prickly Pear Juice as Affected by Cultivar and Harvest Time. Foods 2020, 9, 235. [Google Scholar] [CrossRef] [PubMed]
- Jubair, N.; Rajagopal, M.; Chinnappan, S.; Abdullah, N.B.; Fatima, A. Review on the antibacterial mechanism of plant-derived compounds against multidrug-resistant bacteria (MDR). Evidence-Based Complementary and Alternative Medicine 2021, 2021, 3663315. [Google Scholar] [CrossRef] [PubMed]
- Lorusso, A.B.; Carrara, J.A.; Barroso, C.D.N.; Tuon, F.F.; Faoro, H. Role of Efflux Pumps on Antimicrobial Resistance in Pseudomonas aeruginosa. Int. J. Mol. Sci. 2022, 23. [Google Scholar] [CrossRef]
- Tuon, F.F.; Dantas, L.R.; Suss, P.H.; Tasca Ribeiro, V.S. Pathogenesis of the Pseudomonas aeruginosa Biofilm: A Review. Pathogens 2022, 11, 300. [Google Scholar] [CrossRef]
- Mirani, Z.A.; Naz, S.; Khan, F.; Aziz, M.; Asadullah; Khan, M.N.; Khan, S.I. Antibacterial fatty acids destabilize hydrophobic and multicellular aggregates of biofilm in S. aureus. The Journal of Antibiotics 2017, 70, 115–121. [CrossRef]
- Alves, E.; Dias, M.; Lopes, D.; Almeida, A.; Domingues, M.d.R.; Rey, F. Antimicrobial Lipids from Plants and Marine Organisms: An Overview of the Current State-of-the-Art and Future Prospects. Antibiotics 2020, 9, 441. [Google Scholar] [CrossRef]
- Cartron, M.L.; England, S.R.; Chiriac, A.I.; Josten, M.; Turner, R.; Rauter, Y.; Hurd, A.; Sahl, H.G.; Jones, S.; Foster, S.J. Bactericidal activity of the human skin fatty acid cis-6-hexadecanoic acid on Staphylococcus aureus. Antimicrob. Agents Chemother. 2014, 58, 3599–3609. [Google Scholar] [CrossRef]
- Marshall, D.L.B. , Lloyd B. Antimicrobial activity of sucrose fatty acid ester emulsifiers. J. Food Sci. 1986, 51, 468–470. [Google Scholar] [CrossRef]
- Sikkema, J.; de Bont, J.A.; Poolman, B. Interactions of cyclic hydrocarbons with biological membranes. J. Biol. Chem. 1994, 269, 8022–8028. [Google Scholar] [CrossRef] [PubMed]
- Sikkema, J.; de Bont, J.A.; Poolman, B. Mechanisms of membrane toxicity of hydrocarbons. Microbiol. Rev. 1995, 59, 201–222. [Google Scholar] [CrossRef] [PubMed]
- Helander, I.M.; Alakomi, H.-L.; Latva-Kala, K.; Mattila-Sandholm, T.; Pol, I.; Smid, E.J.; Gorris, L.G.; von Wright, A. Characterization of the action of selected essential oil components on Gram-negative bacteria. J. Agric. Food Chem. 1998, 46, 3590–3595. [Google Scholar] [CrossRef]
- Li, H.; Song, X.; Li, H.; Zhu, L.; Cao, S.; Liu, J. Sesquiterpenes and Monoterpenes from the Leaves and Stems of Illicium simonsii and Their Antibacterial Activity. Molecules 2022, 27, 1115. [Google Scholar] [CrossRef]
- De Rossi, L.; Rocchetti, G.; Lucini, L.; Rebecchi, A. Antimicrobial Potential of Polyphenols: Mechanisms of Action and Microbial Responses—A Narrative Review. Antioxidants 2025, 14, 200. [Google Scholar] [CrossRef]
- Trombetta, D.; Castelli, F.; Sarpietro, M.G.; Venuti, V.; Cristani, M.; Daniele, C.; Saija, A.; Mazzanti, G.; Bisignano, G. Mechanisms of antibacterial action of three monoterpenes. Antimicrob. Agents Chemother. 2005, 49, 2474–2478. [Google Scholar] [CrossRef]
- Senatore, F.; Rigano, D.; Formisano, C.; Grassia, A.; Basile, A.; Sorbo, S. Phytogrowth-inhibitory and antibacterial activity of Verbascum sinuatum. Fitoterapia 2007, 78, 244–247. [Google Scholar] [CrossRef]
- Lee, N.Y.; Yunus, M.A.C.; Idham, Z.; Ruslan, M.S.H.; Aziz, A.H.A.; Irwansyah, N. Extraction and identification of bioactive compounds from agarwood leaves. IOP Conference Series: Materials Science and Engineering 2016, 162, 012028. [Google Scholar] [CrossRef]
- Ezhilan, B.P.; Neelamegam, R. GC-MS analysis of phytocomponents in the ethanol extract of Polygonum chinense L. Pharmacognosy Res. 2012, 4, 11–14. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. American journal of Enology and Viticulture 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Hossain, T.J.; Mozumder, H.A.; Ali, F.; Akther, K. Inhibition of pathogenic microbes by the lactic acid bacteria Limosilactobacillus fermentum strain LAB-1 and Levilactobacillus brevis strain LAB-5 isolated from the dairy beverage borhani. Current Research in Nutrition and Food Science 2022, 10, 928–939. [Google Scholar] [CrossRef]
Figure 1.
Total ion chromatogram (TIC) of V. sinuatum extract with labeled peaks. (1) Methyl palmitate (2) 3,5-ditert-butylphenol, (3) Hordenine.
Figure 1.
Total ion chromatogram (TIC) of V. sinuatum extract with labeled peaks. (1) Methyl palmitate (2) 3,5-ditert-butylphenol, (3) Hordenine.
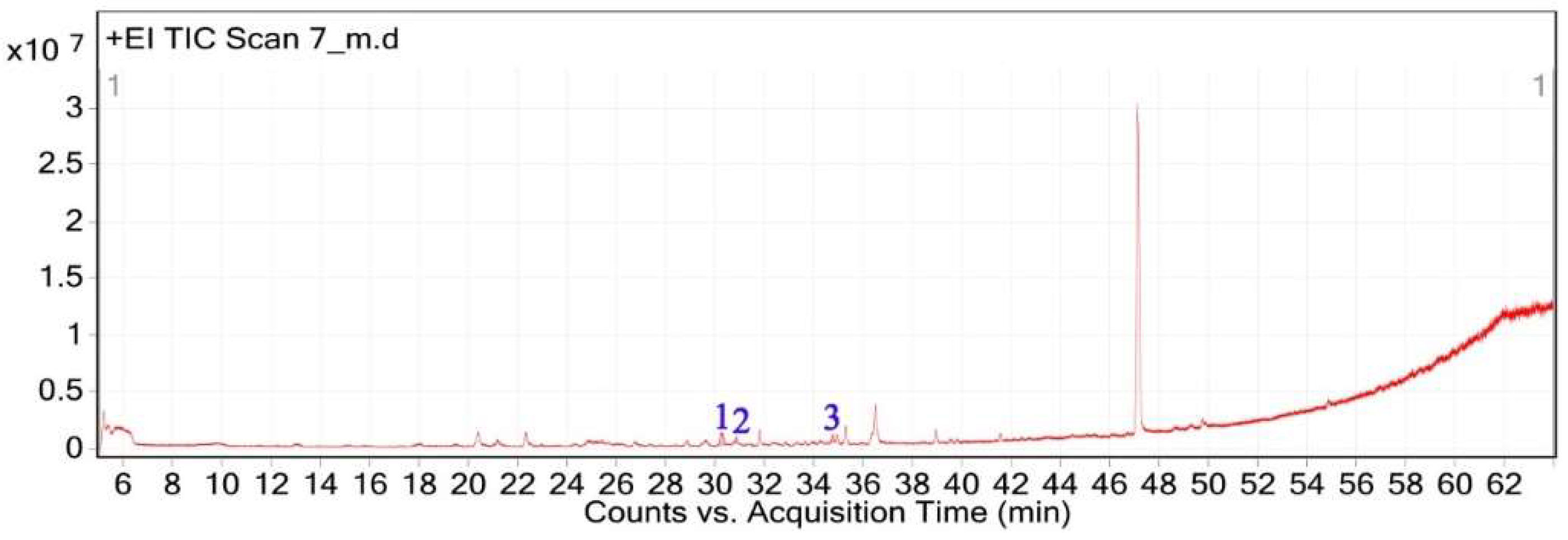
Figure 2.
Chemical structures of key compounds identified in V. sinuatum: (A) Hexadecanoic acid methyl ester (B), (C) Hordenine.
Figure 2.
Chemical structures of key compounds identified in V. sinuatum: (A) Hexadecanoic acid methyl ester (B), (C) Hordenine.
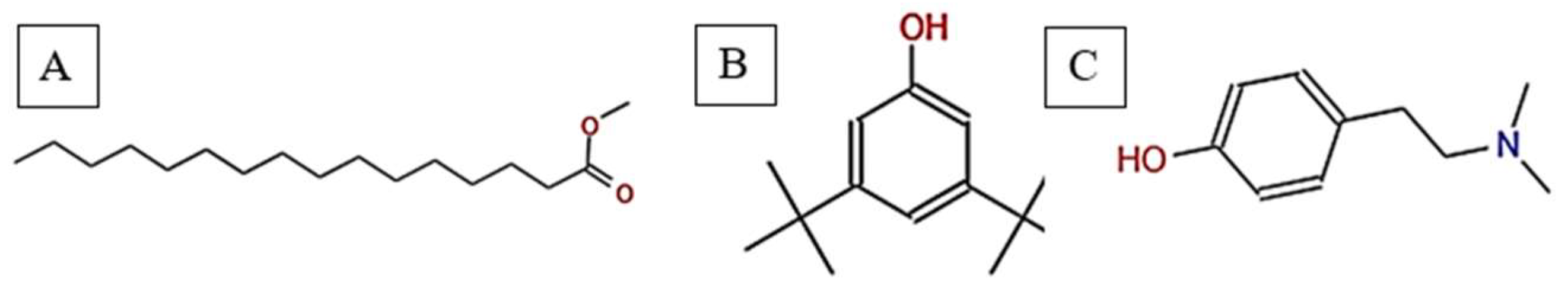
Figure 9.
(A) Antioxidant activity; (B) TPC; (C) TFC of V. sinuatum, A. spinosus, C. getulus, and H. subaxillaris hexane extracts. Values are expressed as mean ± standard deviation (n = 3, from independent experiments). Different lowercase letters designate significant differences among plants based on one-way ANOVA followed by Tukey’s HSD post hoc test (p < 0.05).
Figure 9.
(A) Antioxidant activity; (B) TPC; (C) TFC of V. sinuatum, A. spinosus, C. getulus, and H. subaxillaris hexane extracts. Values are expressed as mean ± standard deviation (n = 3, from independent experiments). Different lowercase letters designate significant differences among plants based on one-way ANOVA followed by Tukey’s HSD post hoc test (p < 0.05).
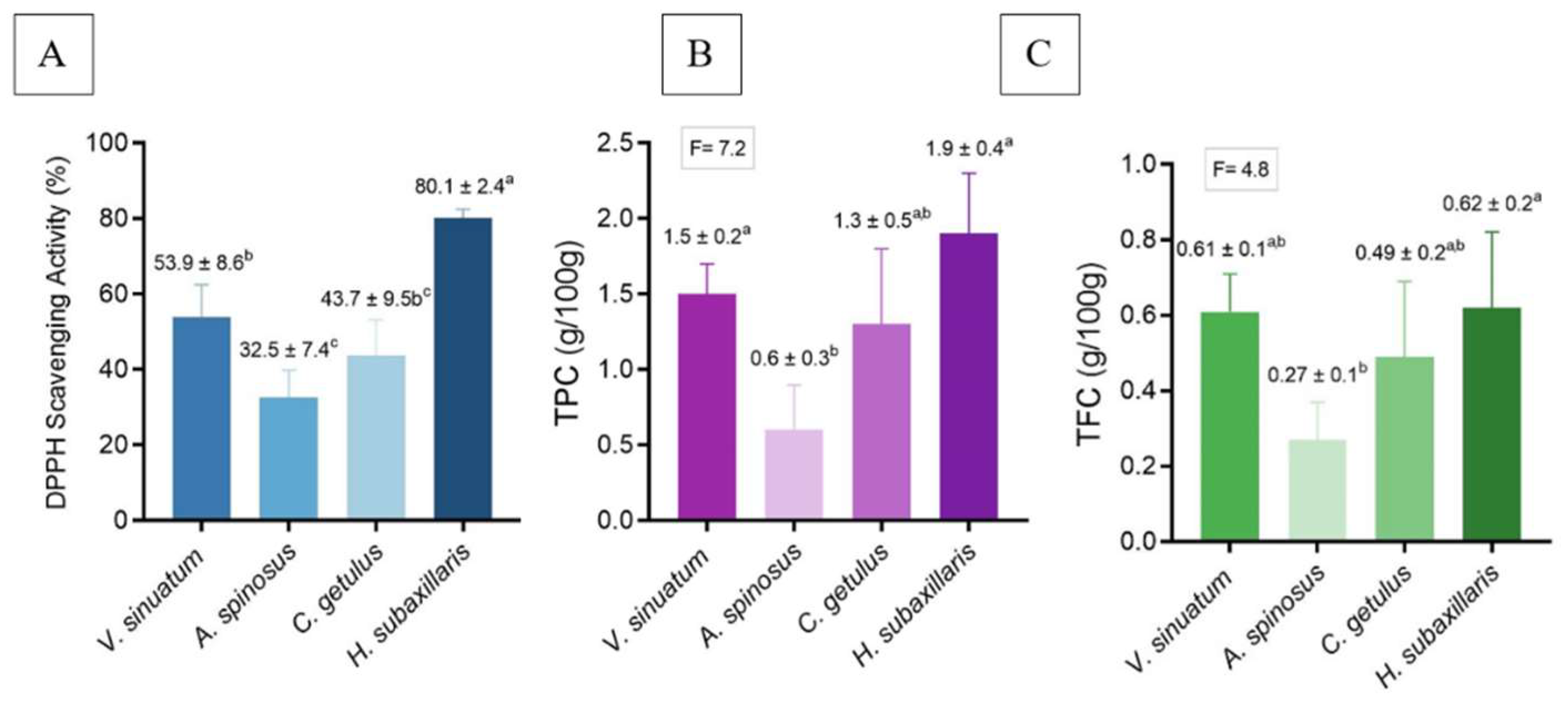

Table 1.
Phytochemical constituents of V. sinuatum extract characterized by GC-MS analysis.
| Peak No. | RT (min) | Area (%) | Chemical Compounds | MF | MW (g/mol) | Key m/z | Match score | Phytochemical Class |
|---|---|---|---|---|---|---|---|---|
| 1 | 30.243 | 65.90 | Methyl palmitate | C₁₇H₃₄O₂ | 270.45 | 74.02 | 71.0 | Saturated fatty acid ester |
| 2 | 30.840 | 13.34 | 3,5-ditert-butylphenol | C₁₄H₂₂O | 206.32 | 190.57 | 75.6 | Phenolic antioxidant |
| 3 | 34.752 | 20.76 | Hordenine | C₁₀H₁₅NO | 165.23 | 190.57 | 75.6 | Alkaloid (phenethylamine) |
MF: Molecular Formula; MW: Molecular Weight.
Table 2.
Phytochemical constituents of A. spinosus extract characterized by GC-MS analysis.
| Peak No. | RT (min) | Area (%) |
Chemical Compounds | MF | MW (g/mol) | Key m/z | Match score | Phytochemical Class |
|---|---|---|---|---|---|---|---|---|
| 1 | 15.526 | 3.03 | Linalyl acetate | C₁₂H₂₀O₂ | 196.29 | 71.01 | 72.8 | Monoterpene ester |
| 2 | 20.51 | 17.59 | Citronellol | C₁₀H₂₀O | 156.27 | 69.01 | 76.7 | Monoterpenoid alcohol |
| 3 | 22.395 | 17.34 | Geraniol | C₁₀H₁₈O | 154.25 | 92.88 | 81.4 | Monoterpenoid alcohol |
| 4 | 32.923 | 9.51 | Methyl elaidate | C₁₉H₃₆O₂ | 296.49 | 55.03 | 86.9 | Fatty acid ester |
| 5 | 34.16 | 21.75 | 11,14-Octadecadienoic acid, methyl ester | C₁₉H₃₄O₂ | 294.47 | 67.02 | 87.6 | Polyunsaturated fatty acid ester |
| 6 | 35.266 | 30.78 | Isocitronellol | C₁₀H₂₀O | 156.27 | 152.77 | 61.4 | Monoterpenoid alcohol |
MF: Molecular Formula; MW: Molecular Weight.
Table 3.
Phytochemical constituents of C. getulus extract characterized by GC-MS analysis.
| Peak No. | RT (min) |
Area (%) |
Chemical Compounds | MF | MW (g/mol) | Key m/z | Match score | Phytochemical Class |
|---|---|---|---|---|---|---|---|---|
| 1 | 20.292 | 0.88 | Citronellol | C₁₀H₂₀O | 156.26 | 69.01 | 74.6 | Monoterpenoid alcohol |
| 2 | 22.135 | 1.29 | Geraniol | C₁₀H₁₈O | 154.25 | 92.90 | 77.6 | Monoterpenoid alcohol |
| 3 | 27.226 | 0.69 | Oleic acid | C₁₈H₃₄O₂ | 282.5 | 137.84 | 65.3 | Unsaturated fatty acid |
| 4 | 29.528 | 9.58 | Isophytol | C₂₀H₄₀O | 296.5 | 57.98 | 70.3 | Diterpene alcohol |
| 5 | 30.127 | 66.26 | Methyl palmitate | C₁₇H₃₄O₂ | 270.45 | 74.02 | 73.8 | Saturated fatty acid ester |
| 6 | 30.800 | 12.67 | 3,5-ditert-butylphenol | C₁₄H₂₂O | 206.32 | 190.59 | 78.7 | Phenolic antioxidant |
| 7 | 31.776 | 4.41 | Thymol | C₁₀H₁₄O | 150.22 | 134.76 | 74.1 | Monoterpenoid phenol |
| 8 | 33.083 | 2.14 | Arachidonic acid | C₂₀H₃₂O₂ | 304.5 | 78.85 | 77.8 | PUFA |
| 9 | 33.798 | 1.03 | Methyl undecanoate | C₁₂H₂₄O₂ | 200.32 | 74.98 | 64.6 | Fatty acid ester |
| 10 | 36.308 | 1.05 | Phytol | C₂₀H₄₀O | 296.5 | 70.92 | 76.1 | Diterpene alcohol |
MF: Molecular Formula; MW: Molecular Weight.
Table 4.
Phytochemical constituents of H. subaxillaris extract characterized by GC-MS analysis.
| Peak No. | RT (min) |
Area (%) |
Chemical Compounds | Molecular Formula | MW (g/mol) | Key m/z | Match score | Phytochemical Class |
|---|---|---|---|---|---|---|---|---|
| 1 | 22.256 | 3.22 | Geraniol | C₁₀H₁₈O | 154.25 | 69.03 | 78.6 | Monoterpenoid alcohol |
| 2 | 27.538 | 18.52 | Cubebol | C₁₅H₂₆O | 222.37 | 81.87 | 78.0 | Sesquiterpenoid alcohol |
| 3 | 29.072 | 3.56 | Falcarinol | C₁₇H₂₄O | 244.38 | 114.75 | 80.1 | Long-chain fatty alcohol |
| 4 | 30.153 | 36.84 | Methyl 8-methyl-nonanoate | C₁₁H₂₂O₂ | 186.29 | 74.02 | 74.3 | Branched fatty acid ester |
| 5 | 30.130 | 18.03 | Methyl palmitate | C₁₇H₃₄O₂ | 270.5 | 114.75 | 71.9 | fatty acid methyl ester |
| 6 | 30.242 | 4.31 | methyl octadeca-13,16-diynoate | C₁₉H₃₀O₂ | 290.45 | 190.59 | 78.7 | Polyunsaturated fatty acid ester |
| 7 | 30.772 | 9.28 | 3,5-ditert-butylphenol | C₁₄H₂₂O | 206.32 | 55.04 | 72.9 | Phenolic antioxidant |
| 8 | 34.889 | 6.24 | 2,5-Octadecadiynoic acid, methyl ester | C₁₉H₃₀O₂ | 290.45 | 190.59 | 78.7 | Polyunsaturated fatty acid ester |
MF: Molecular Formula; MW: Molecular Weight.
Table 5.
Comparative inhibition zones (mm) of various plant species, hexane crude extract against pathogenic bacteria.
Table 5.
Comparative inhibition zones (mm) of various plant species, hexane crude extract against pathogenic bacteria.
| V. sinuatum | A. spinosus | C. getulus | H. subaxillaris | |
|---|---|---|---|---|
| B. cereus | 15.2 ± 1.5b | 8.3 ± 0.6c | 13.5 ± 1.2b | 19.8 ± 0.8a |
| S. aureus | 12.6 ± 0.9a | 7.0 ± 1.1b | 11.8 ± 0.9a | 12.5 ± 0.7a |
| E. coli | 9.3 ± 0.7a | 5.5 ± 0.8b | 6.2 ± 0.5b | 10.1 ± 0.3a |
| P. aeruginosa | 7.0 ± 0.5a | 4.2 ± 0.6b | 5.0 ± 0.4b | 8.4 ± 0.9a |
Mean values labeled with different superscript letters (a–c) are significantly different (p < 0.05), according to Tukey’s HSD test. SD: Standard deviation of triplicate experiments.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



