
Submitted:
24 April 2025
Posted:
25 April 2025
You are already at the latest version
Abstract
The fungal family Mortierellaceae represents ubiquitous and ecologically significant components of soil ecosystems across terrestrial habitats. Through an integrative taxonomic approach combining multi-locus phylogenetic analyses (ITS, LSU, SSU rDNA, RPB1, and Act) with detailed morpho-logical examinations of rhizosphere soil isolates, four novel species within this family were pro-posed. This study describes and illustrates these taxa, elucidating their morphological distinctions from allied species and phylogenetic relationships within the family. Linnemannia rotunda sp. nov. (closely related to L. longigemmata) is distinguished by its globose sporangia and sporangiospores. Mortierella acuta sp. nov. is characterized by spiky collarettes. Mortierella oedema sp. nov. (a sister to M. macrocystis) exhibits distinctive ampulliform swellings. Mortierella tibetensis sp. nov. (clustering with M. parvispora), named for its geographic origin in Tibet. As the eighth installment in a sys-tematic investigation of early-diverging fungal diversity in China, this work expands the global taxonomic inventory of Mortierellaceae to 148 species, underscoring the ongoing discovery of cryptic biodiversity within this ecologically pivotal group.
Keywords:
Linnemannia
; Mortierella
; Mortierellales
; multi-gene phylogeny
; taxonomy
1. Introduction
Mortierellaceae, belonging to Mortierellomycota, Mortierellomycotina, Mortierellomycetes and Mortierellales (http://www.indexfungorum.org/, accessed on 14 February 2025) [1,2], typically form white, cottony zonate or rosette-like colonies, with a distinctive odor reminiscent of garlic or a freshly bathed dog [3,4,5] A swelling is usually produced at the base of sporangiophores. Mortierellaceae species are considered to be an important saprophyte [6], usually detected and isolated from soil, plant remains, insect guts, mosses, and living plant roots [3] This family is ubiquitous and widely distributed. GBIF database documents Mortierellaceae from Estonia (92,697 records), Australia (35,514), Czechia (27,899), Russian Federation (15,180), Colombia (13,733), United States of America (13,424), Italy (13,417), Lithuania (11,343), Sweden (11,074) and Norway (422), (https://www.gbif.org/, accessed on 24 February 2025). Recent studies on soil microbial communities across the globe have shown that species of Mortierellaceae are important members of the soil core microbiome. [7,8] Owing to their broad habitats, members of the genus Mortierella are able to grow at a wide range of temperatures. They live in the winter-active soil microbial community, forming substantial fungal biomass in the soil during both the snow-covered and the vegetative periods. [9] They produce polyunsaturated fatty acids, such as arachidonic acid, which are crucial for several biological functions in mammals. [10,11,12] These biological functions are widely used in commercial production, for example, in the production of biofuels. [13,14] Many Mortierellaceae species have the ability to promote plant growth, to decompose plant litter, and to remodel rhizosphere microbial communities. [15,16] Some species are also biological control agents, producing active antimicrobial metabolites. [17].
Over the past few years, Mortierellaceae has experienced an influx of a large number of new species. [18] It currently accommodates 17 genera and 144 species. Among them, the Mortierella is the most species-rich genus, with 80 species. It is followed by Linnemannia, with 24 species recorded. (https://www.catalogueoflife.org/, accessed on 15 February 2025).
In this paper, four new species, Linnemannia rotunda sp. nov., Mortierella acuta sp. nov., M. oedema sp. nov. and M. tibetensis sp. nov., were described from soil samples in China (Yunnan, Shandong and Tibet) based on evidence of molecular phylogeny, morphological characteristic and growth temperature. This is the eighth report of a serial work on diversity of Chinese early-diverging fungi [19,20,21,22,23,24,25]
2. Materials and Methods
2.1. Isolation and Morphology
In 2024, soil samples were collected in Yunnan, Tibet, and Shandong, following the methods by Zou et al. [26] and Liu et al. [27] Each sample (approximately 100 g) was placed into a sterile bag and labeled with date, vegetation type, altitude, latitude, and longitude. All samples were stored at 4°C after being transported to the laboratory. Pure strains were isolated from the soil samples using a combination of soil dilution plating and moist-chamber cultivation methods. [28]. Approximately 1 g of soil sample was placed into a 10 mL centrifuge tube containing 10 mL of sterile water and agitated on a shaker for 25 min to prepare a soil suspension. One milliliter of the initial suspension was added to nine milliliters of sterile water to obtain a 10−2 soil suspension. The process was repeated to achieve 10−3 and 10−4 soil suspensions. Approximately 200 μL of the 10−3 and 10−4 soil suspensions were pipette to the rose bengal chloramphenicol agar (RBC: peptone 5.00 g/L, KH2PO4 1.00 g/L, MgSO4·7H2O 0.50 g/L, rose bengal 0.05 g/L, glucose 10.00 g/L, chloramphenicol 0.10 g/L, agar 15.00 g/L), and evenly dispersed using a sterile triangular glass spreader. [29] The plates were cultivated at 26 ℃ in the dark for 2–5 d. Subsequently, the agar containing mycelia at the edge of the colonies was transferred to fresh potato dextrose agar (PDA: glucose 20 g/L, potato 200 g/L, agar 20 g/L). Macroscopic images were captured using a digital camera (Canon PowerShot G7X, Canon, Tokyo, Japan). For the moist-chamber method, 1 g of soil was evenly spread on the surface of PDA plates, sealed with a parafilm and incubated invertedly at 26 ℃ in the dark. After 2–3 d, target strains were purified by streaking with an inoculation loop. Two days later, the agar containing mycelia at the colony edge was transferred to fresh PDA and cultured as described above. A drop of lactic acid phenol cotton blue staining solution was mounted on the glass slide. Then, a small piece of tape was touched to the surface of the mycelia, making part of the hyphae adhere to it. It was then soaked in the lactic acid phenol cotton blue staining solution. The microscopic morphological characteristics of the fungi were observed using a stereoscope (Olympus SZX10, OLYMPUS, Tokyo, Japan) and a light microscope (Olympus BX53, OLYMPUS, Tokyo, Japan), and images were captured with a high-definition color digital camera (Olympus DP80 OLYMPU, Tokyo, Japan). [20,21,22,23,24,25,30] Structural measurements were conducted using Digimizer software (v5.6.0), with at least 25 individuals measured for each trait. The minimum and maximum growth temperature was determined using a gradient method. The culture was initially incubated at 10°C for two days, and then the temperature was reduced by 1°C each day until no further growth. This temperature was defined as the minimum growth temperature. The culture was initially incubated at 25°C for two days, and then the temperature was increased by 1°C each day until no further growth. This temperature was defined as the maximum growth temperature. All strains were kept in 10% sterilized glycerin at -20℃. The living cultures were stored in the China Microbiological Culture Collection Center, Beijing, China (CGMCC). Equivalent strains were preserved in the Shandong Normal University Culture Collection (XG). Dry culture of types was submitted to the Herbarium Mycologicum Academiae Sinicae, Beijing, China (Fungarium; HMAS). The taxonomic information was deposited to the Fungal Names repository (https://nmdc.cn/fungalnames/).
2.2. DNA Extraction, PCR Amplification, and Sequencing
The DNA extraction kit (Cat. No.: 70409-20; BEAVER Biomedical Engineering Co., Ltd.) was employed for genomic DNA extraction, following the manufacturer’s instructions. The ITS, LSU, SSU, RPB1 and Act regions were amplified using the primer pairs and programs specified in Table 1. The final volume of the PCR reaction mixture is 25 μL, comprising 12.5 uL of 2 × Hieff Canace Plus PCR Master Mix with dye (Yeasen Biotechnology, Cat No. 10154ES03), 9.5 μL of ddH2O, 1 μL of forward primer (10 μM), 1 μL of reverse primer (10 μM) and 1 µL of template genomic DNA (1 ng/μL). PCR products were visualized at 254 nm on a 2% agarose electrophoresis gel. [31], and purified using a gel extraction kit (Cat# AE0101-C, Shandong Sparkjade Biotechnology Co., Ltd.). DNA sequencing was performed by Tsingke Biotechnology (Beijing, China). All sequences generated in this study were deposited in GenBank.
2.3. Phylogenetic Analyses
Newly acquired sequence data were processed using MEGA v.7.0 to ensure consistency. [36,37] Reference sequences were downloaded from GenBank according to a study on Mortierellaceae by Telagathoti et al. [18] Phylogenetic analyses were conducted for each marker, as well as a concatenation of ITS-LSU-SSU-RPB1-Act. The phylogeny of Mortierellaceae was inferred using both maximum likelihood (ML) and Bayesian inference (BI) algorithms. [38,39] These algorithms were integrated with the CIPRES Science Portal (https://www.phylo.org/, accessed February 15, 2025). ML analysis was carried out with 1,000 bootstrap replicates using RaxML 8.2.4 (https://www.phylo.org/) in CIPRES Science Gateway V. 3.3. [40,41] BI analysis was performed using the GTR + I + G model and sampling frequence of once per 1,000 generations. Eight cold Markov chains were run simultaneously for two million generations. [42,43] The phylogenetic trees resulted were optimized with the iTOL (https://itol.embl.de, accessed February 15, 2024), and refined using Adobe Illustrator CC 2019.[20]
3. Results
3.1. Molecular Phylogeny
For Linnemannia, phylogenetic analyses were performed on a dataset containing 31 strains, representing 25 species, with Mortierella cogitans (CBS 879.97) as an outgroup. The sequence matrix comprises a total of 4,527 concatenated characters: 1–634 (ITS), 635–1,622 (LSU), 1,623–2,498 (SSU), 2,499–3,700 (RPB1), and 3,701–4,527 (Act). Among these characters, 829 are parsimony-informative, along with 3,530 constant and 168 parsimony-uninformative. Bayesian tree topology is congruent with that of the ML tree (Figure 1).
For Mortierella, phylogenetic analyses were performed on a dataset containing 87 strains, representing 74 species, with Umbelopsis isabelline (NRLL 1757) and U. actotrophica (CBS 31093) employed as outgroups. The sequence matrix comprises a total of 4,527 concatenated characters: 1–990 (ITS), 991–1,959 (LSU), 1,960–3,042 (SSU), 3,043–4,401 (RPB1), and 4,402–5,273 (Act). Among these, 1,828 are parsimony-informative along with 2,542 constant and 903 parsimony-uninformative. Bayesian tree topology is consistent with the ML tree (Figure 2).
3.2. Taxonomy
3.2.1. Linnemannia rotunda X.Y. Ji, Y. Jiang & X.Y. Liu, sp. nov. Figure 3
Fungal Names—FN##### (to be applied after review)
Figure 3.
Linnemannia rotunda ex-holotype CGMCC 3.28764. (a, b) Colonies on PDA (a obverse, b reverse); (c, d) Chlamydospores; (e-i) Sporangia; (k) Typical swollen hyphae; (j, l-n) Sporangiospores; Scale bars: (c-n) 10 µm.
Figure 3.
Linnemannia rotunda ex-holotype CGMCC 3.28764. (a, b) Colonies on PDA (a obverse, b reverse); (c, d) Chlamydospores; (e-i) Sporangia; (k) Typical swollen hyphae; (j, l-n) Sporangiospores; Scale bars: (c-n) 10 µm.
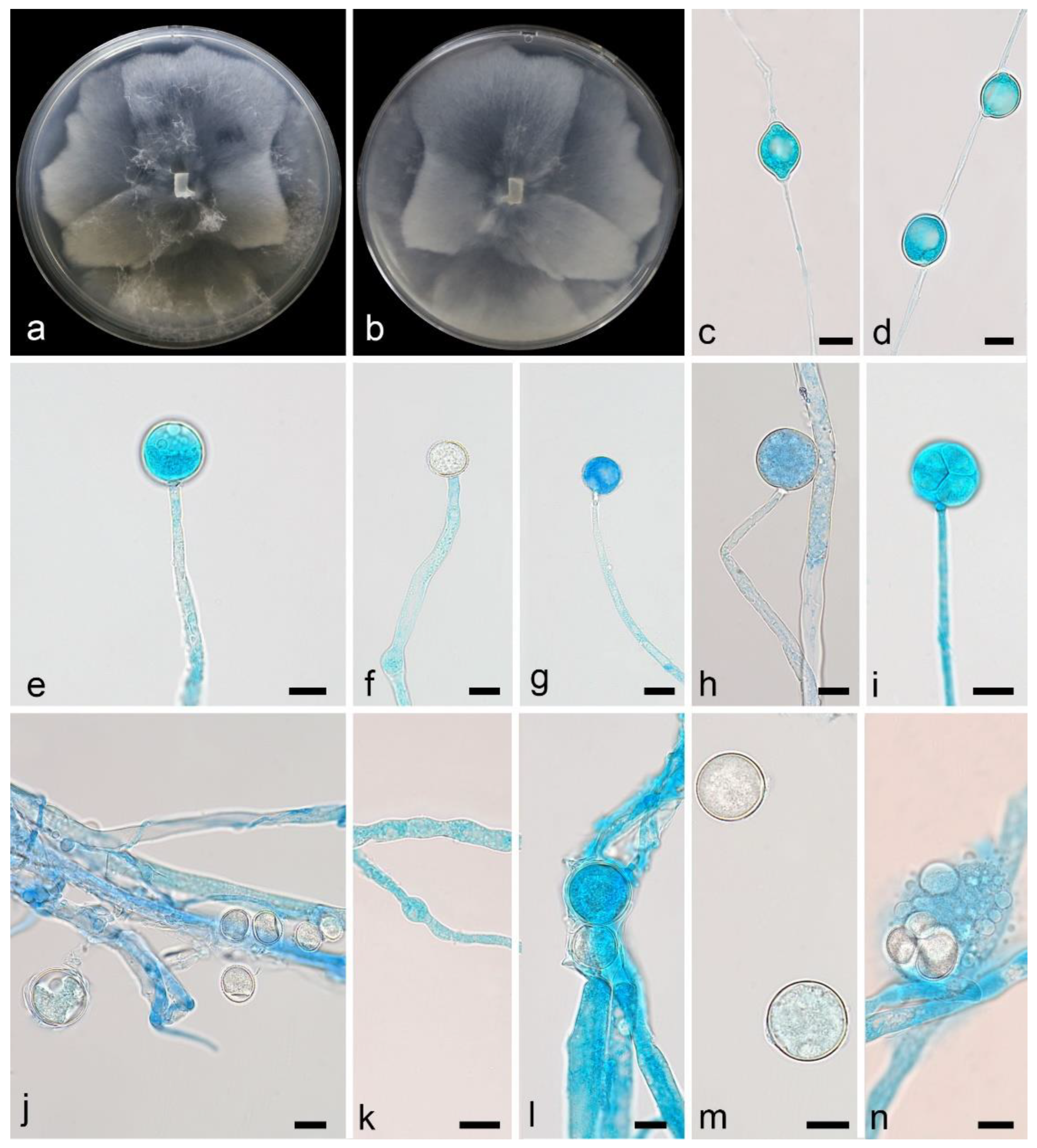
Type—China, Yunnan Province, Yuxi City, Xinping Dai Autonomous Country (23°56'39''N, 101°30'1''E, altitude 2397.53 m), from soil, 14 May 2024, X.Y. Ji, holotype HMAS 353518, ex-holotype living culture CGMCC 3.28764 (=XG08755-7-1).
Etymology—The ‘’rotunda’’ (Lat.) refers to the round shape of sporangia and sporangiospores.
Description—Colonies on PDA at 16°C for 5 d, reaching 88 mm diameter, fast growing with a rate of 17.6 mm/d, garlic smell, with sparse aerial mycelia. Hyphae hyaline, 1.7–9.4 µm in diameter, sometimes swollen. Sporangiophores erect or slightly bent, unbranched, 25.5–146.0 µm long, 1.9–5.0 µm wide, sometimes with a swelling beneath sporangia. Sporangia oval to round, smooth, multi-spored, 10.4–22.3 µm long, 10.7–22.6 µm wide. Columellae present but usually tiny. Sporangiospores smooth, hyaline, mostly round, 9.6–19.0 µm in diameter. Chlamydospores present, mostly oval, 10.1–22.0 µm long, 6.6–15.6 µm wide. Zygospores not found.
Temperature requirements—Minimum growth temperature 4°C, and maximum growth temperature 28°C.
Additional specimen examined—China, Yunnan Province, Yuxi City, Xinping Dai Autonomous Country (23°56'39''N, 101°30'1''E, altitude 2397.53m), from soil, 14 May 2024, X.Y. Ji and X.Y. Liu, living culture XG08755-7-2.
Notes—The ITS rDNA phylogenetic analysis showed that the new species Linnemannia rotunda was closely related to L. longigemmata (MLBV=78, BIPP=0.96, Figure 1) [18] The new species is distinguished from L. longigemmata by 61/634 characters. Morphologically, compared to L. longigemmata, the new species has a shorter sporangiophores (25.5–146.0 µm vs 50–150.0 µm).
3.2.2. Mortierella acuta X.Y. Ji, Y. Jiang & X.Y. Liu, sp. nov. Figure 4
Fungal Names—FN##### (to be applied after review)
Type—China, Shandong Province, Tai'an City, Mount Tai (36°11'49''N, 117°7'16''E, altitude 155.8m), from soil, 12 March 2024, X.Y. Ji, holotype HMAS 353516, ex-holotype living culture CGMCC 3.28761 (=XG08182-4-1).
Etymology—The epithet ‘’acuta’’ (Lat.) refers to the spiky collarette.
Description—Colonies on PDA at 16°C for 6 d, attaining 64 mm diameter, moderately fast growing with a rate of 10.6 mm/d, garlic smell, white cottony, with a rose pattern, luxuriant and velvety after 20 d of cultivation. Hyphae hyaline, upright or bent. Sporangiophores arising from aerial mycelia, erect or slightly bent, unbranched, 22.4–72.4 µm in height, tapering from 2.8–4.3 µm to at the base to 1.0–1.4 µm at the apex. Sporangia almost spherical in shape, smooth, deliquescent, multi-spored, 9.5–12.6 µm in diameter. Columellae absent. Collarettes present. Sporangiospores transparent, mostly oval, 2.6–3.4 µm long, 1.4–1.7 µm wide. Chlamydospores present. Zygospores absent.
Temperature requirements—Minimum growth temperature 4°C, and maximum growth temperature 29°C.
Additional specimen examined—China, Shandong Province, Tai'an City, Mount Tai (36°11'49''N, 117°7'16''E, altitude 155.8m), from soil, 12 March 2024, X.Y. Ji and X.Y. Liu, living culture XG08182-4-2.
Notes—Phylogenetic analysis of the two combined genes of ITS and LSU showed that the new species M. acuta forms an independent and fully supported clade (MLBV = 100, BIPP = 1.00).
Figure 4.
Mortierella acuta ex-holotype CGMCC 3.28761. (a, b) Colonies on PDA (a obverse, b reverse); (c) Chlamydospores; (d) Sporangia; (e, f) Deliquescent sporagnia releasing sporangiospores and leaving obvious collarettes; (g) Sporangiospores; Scale bars:(c-i) 10 µm.
Figure 4.
Mortierella acuta ex-holotype CGMCC 3.28761. (a, b) Colonies on PDA (a obverse, b reverse); (c) Chlamydospores; (d) Sporangia; (e, f) Deliquescent sporagnia releasing sporangiospores and leaving obvious collarettes; (g) Sporangiospores; Scale bars:(c-i) 10 µm.
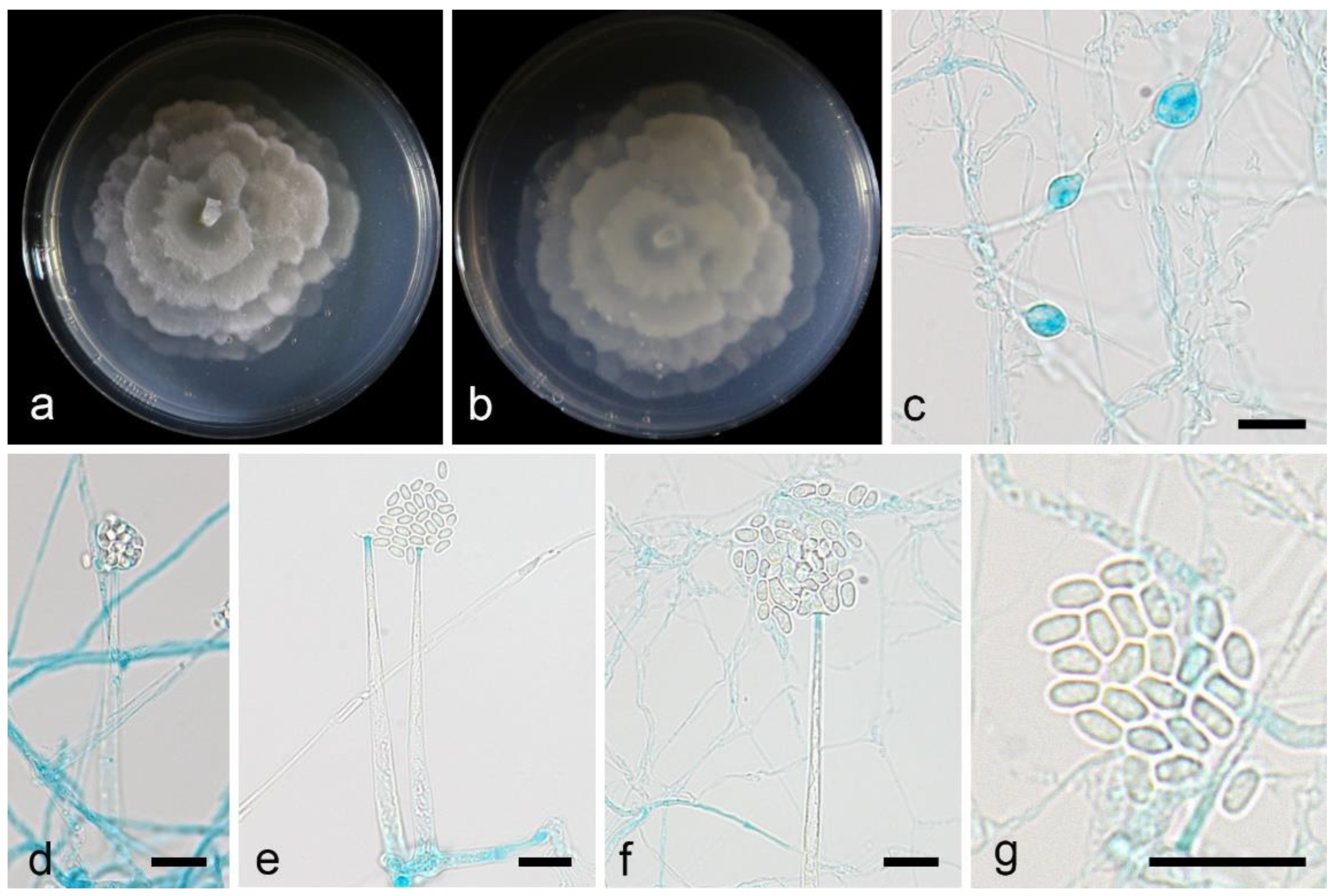
3.2.3. Mortierella oedema X.Y. Ji, Y. Jiang & X.Y. Liu, sp. nov. Figure 5
Fungal Names—FN##### (to be applied after review)
Type—China, Tibet, Shigatse City, Yadong County (27°24'37''N, 88°54'23''E, altitude 3535m), from soil, 25 June 2024, X.Y. Ji, holotype HMAS 353517, ex-holotype living culture CGMCC 3.28762 (=XG00420-1-1).
Etymology—The ‘’oedema’’ (Lat.) refers to the swelling of hyphae.
Figure 5.
Mortierella oedema ex-holotype CGMCC 3.28762. (a, b) Colonies on PDA (a obverse, b reverse); (c, d) Sporangia; (e-i, k) Chlamydospores; (j) Typical swollen hyphae; (l) Sporangiospores; Scale bars:(c-l) 10 µm.
Figure 5.
Mortierella oedema ex-holotype CGMCC 3.28762. (a, b) Colonies on PDA (a obverse, b reverse); (c, d) Sporangia; (e-i, k) Chlamydospores; (j) Typical swollen hyphae; (l) Sporangiospores; Scale bars:(c-l) 10 µm.
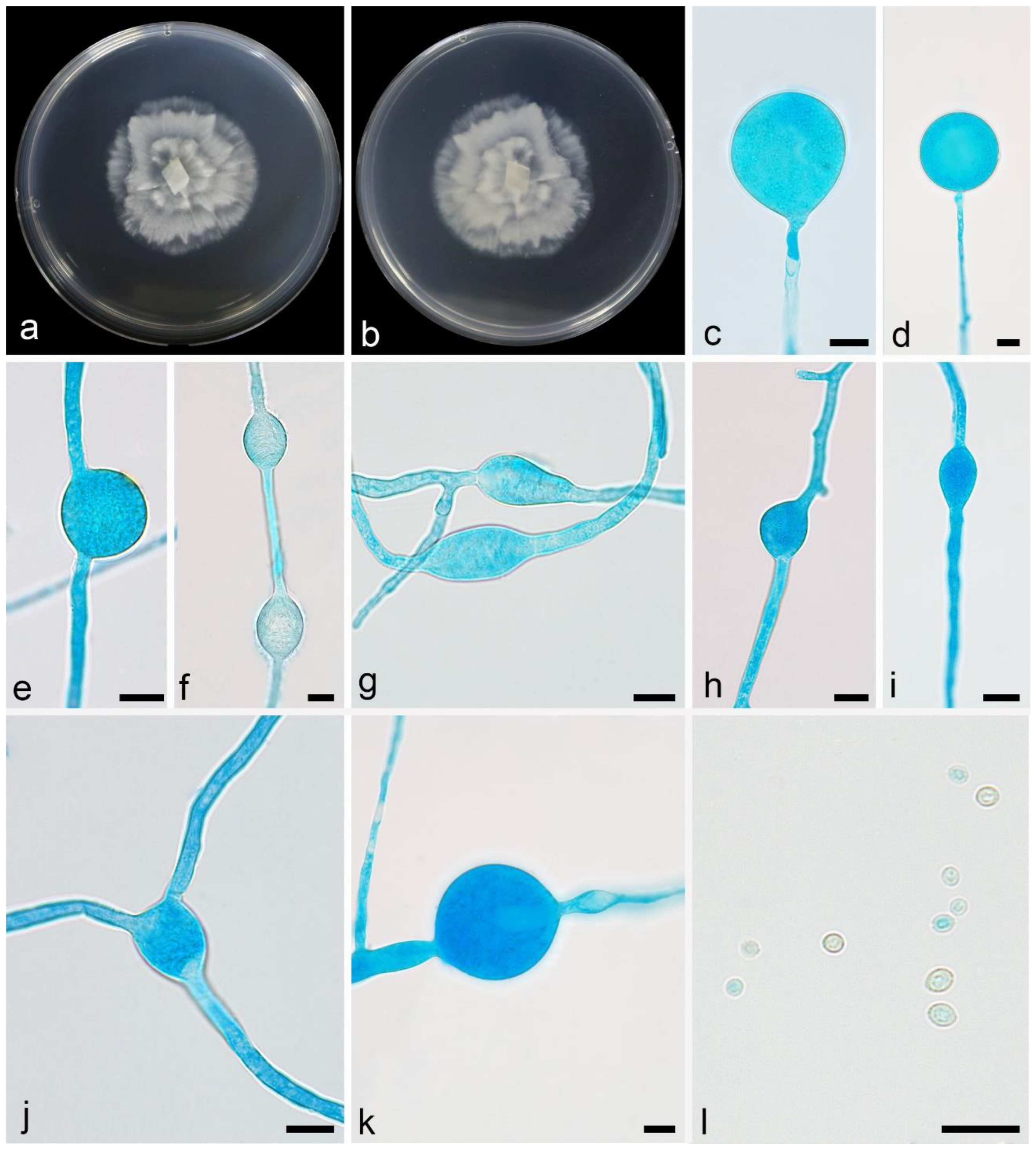
Description—Colonies on PDA at 16°C for 5 d, reaching 45 mm diameter, slow growing with a rate of 9 mm/d, garlic smell, sparse aerial mycelia, with characteristic rose pattern. Hyphae hyaline, light brown with age, 1.7–5.1 µm wide, sometimes swollen. Sporangia oval or spherical, smooth, deliquescent, hyaline, 30.7–42.5 µm in diameter. Columellae absent. Collarettes absent. Sporangiospores hyaline, smooth, oval or round, 2.2–3.1 µm long, 2.0–2.9 µm wide. Chlamydospores abundant, oval, round and irregular, 7.7–43.1 µm long and 6.9–37.7 µm wide. Zygospores not found.
Temperature requirements—Minimum growth temperature 4°C, and maximum growth temperature 28°C.
Additional specimen examined—China, Tibet, Shigatse City, Yadong County (27°24'37''N, 88°54'23''E, altitude 3535m), from soil, 25 June 2024, X.Y. Ji, living culture XG00420-1-2.
Notes—The ITS rDNA phylogenetic analysis showed that the new species M. oedema is closely related to M. macrocystis (MLBV=100, BIPP=1, Figure 2) [44] It is distinguished from M. macrocystis by 48/631 characters in ITS sequences. In sporangiospore shape, the new species is oval or round while M. macrocystis almost spherical. In chlamydospore shape, the new species is various (oval, round and irregular) while M. macrocystis globose.
3.2.4. Mortierella tibetensis X.Y. Ji, Y. Jiang & X.Y. Liu, sp. nov. Figure 6
Fungal Names—FN##### (to be applied after review)
Type—China, Tibet, Shigatse City, Yadong County, (27°21'53''N, 88°58'26''E, altitude 3535m), 2827m, from soil, 26 June 2024, X.Y. Ji, holotype HMAS 353519, ex-holotype living culture CGMCC 3.28763 (=XG00421-2-1).
Etymology—The ‘’tibetensis’’ (Lat.) refers to Tibet Autonomous Region of China where the type was collected.
Figure 6.
Mortierella tibetensis ex-holotype CGMCC 3.28763 (a, b) Colonies on PDA (a obverse, b reverse); (c, g) Sporangia; (h, i) Deliquescent sporangia releasing sporangiospores; (j) Curved hyphae; (k) Typical swollen hyphae; (l) Sporangiospores; Scale bars: (c-l) 10 µm.
Figure 6.
Mortierella tibetensis ex-holotype CGMCC 3.28763 (a, b) Colonies on PDA (a obverse, b reverse); (c, g) Sporangia; (h, i) Deliquescent sporangia releasing sporangiospores; (j) Curved hyphae; (k) Typical swollen hyphae; (l) Sporangiospores; Scale bars: (c-l) 10 µm.
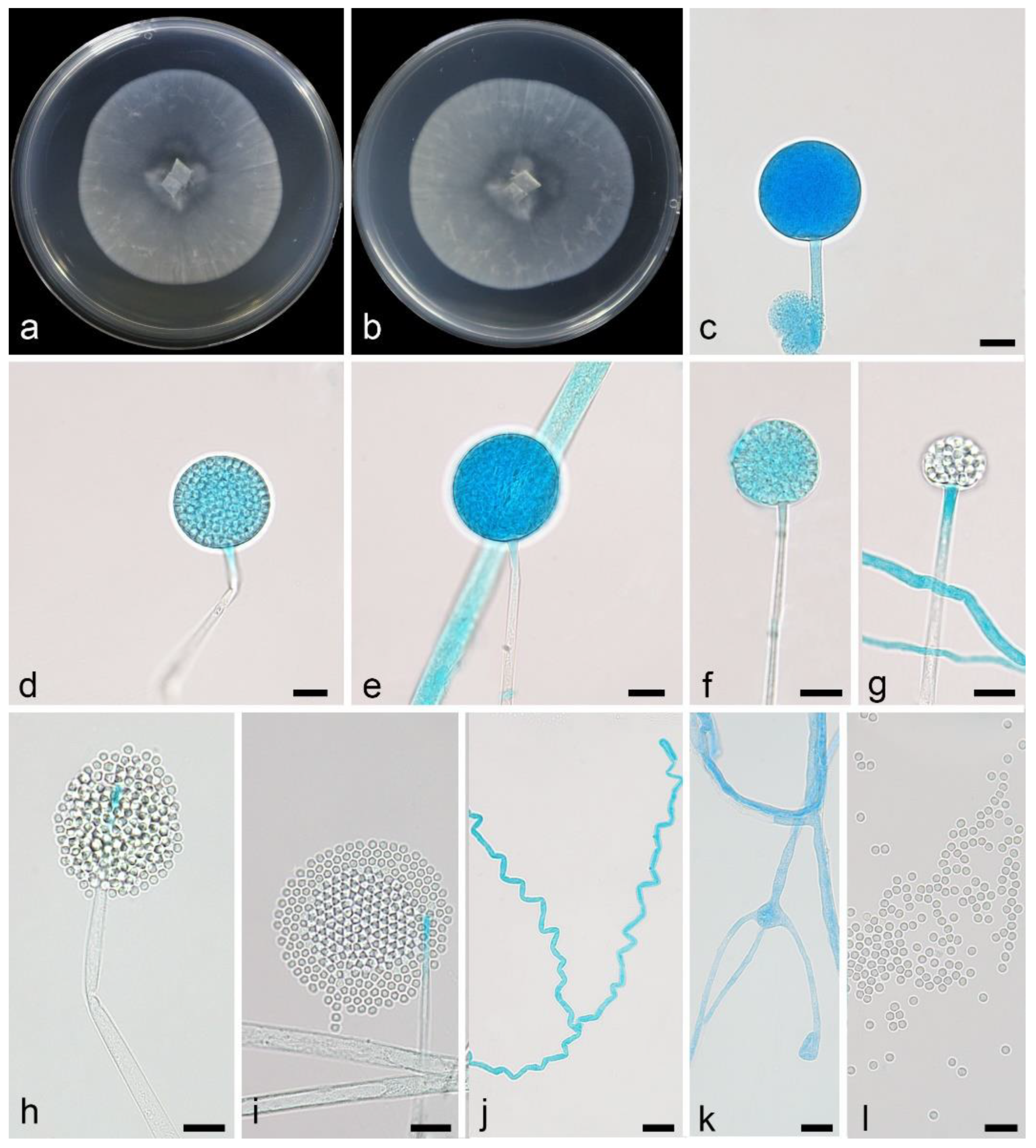
Description—Colonies on PDA at 16°C for 7 d, reaching 59 mm diameter, slow growing with a rate of 8.4 mm/d, garlic smell and a wet dog smell, with sparse aerial mycelia. Hyphae hyaline, upright or bent, sometimes swollen. Sporangiophores arising from aerial mycelia, erect or slightly bent, unbranched, 112–406 µm in height, tapering from 4.2–6.9 µm to at the base to 1.6–2.6 µm at the apex. Sporangia almost spherical in shape, smooth, deliquescent, multi-spored, 12.8–30.3 µm in diameter. Columellae absent. Collarettes absent. Sporangiospores hyaline, mostly pentagonal or hexagonal, smooth, 1.8–5.9 µm vertical heigh. Chlamydospores absent. Zygospores not observed.
Temperature requirements—Minimum growth temperature 4°C, and maximum growth temperature 29°C.
Additional specimen examined—China, Tibet, Shigatse City, Yadong County (27°21'53''N, 88°58'26''E, altitude 3535m), from soil, 26 June 2024, X.Y. Ji and X.Y. Liu, living culture XG00421-2-2.
Notes—Phylogenetic analysis of three loci (ITS, LSU, SSU) showed that the new species M. tibetensis was closely related to M. parvispora (MLBV=88, BIPP=1, Figure 2) [45]. The new species is distinguished from M. parvispora by 49/648, 18/969 and 25/1019 characters in ITS, LSU and SSU sequences, respectively. Due to lack of morphological description in protologue for M. parvispora, no comparisons are able to be made.
4. Discussion
Mortierellaceae is a fungal family with an extremely high ecological and physiological diversity that allows it to be widely distributed worldwide. [46] It covers a wide range of ecosystems from cold regions to temperate and tropical. For example, some species exhibit significant activity in alpine and polar environments, while others are widespread in temperate and tropical soils. Members of this family typically live in soil, but also have extensive associations with plant roots, insect guts, and other microorganisms. [47] For example, some bacterial species of Pseudomonas often live in symbiosis with Mortierellaceae fungi, and this symbiosis may have an impact on the fungi's volatile organic compounds (VOCs). Some soil samples in Yunnan Province, Tibet Autonomous Region and Shandong Province were investigated in this study. The city Yuxi in Yunnan Province have a subtropical monsoon climate with complex terrain, mild and humid terrain, and abundant precipitation. The climatic environment is conducive to the growth of various microorganisms. Tai'an City, Shandong Province belongs to the warm temperate continental semi-humid monsoon climate zone, with four distinct seasons, suitable cold and summer, synchronized light and temperature, and rain and heat in the same season. Yadong County of Tibet Autonomous Region, has a plateau and mountainous climate, with significant seasonal changes and extreme weather phenomena. [48,49] Four new species of the family Mortierellaceae were obtained from these regions.
In recent years, the phylogenetic analysis of the Mortierellaceae family has primarily relied on morphological characteristics and the ITS + LSU + SSU sequences. In this study, the phylogenetic tree was reconstructed with the additional inclusion of Act and RPB1 protein-coding sequences. The results were largely consistent with those from previous studies based on ITS + LSU. Based on morphology and molecular phylogenetic analyses, four novel species were identified in the family Mortierellaceae, namely L. rotunda sp. nov., M. acuta sp. nov., M. oedema sp. nov. and M. tibetensis sp. nov. By analyzing these data, strong supports were obtained for the clades of these species (Fig 1: L. rotunda 100 MLBV and 1.00 BIPP; Figure 2: M. acuta 100 MLBV and 1.00 BIPP, M. oedema 100 MLBV and 0.99 BIPP, M. tibetensis 100 MLBV and 1.00 BIPP). At the same time, in terms of morphological structure and physiology, we also found some differences between these four newly discovered species and their relatives. They have great differences in sporangiospores size, stolon width and sporangium size; However, other features are not described in the protologues of those rlatives. [18,44,45,50,51]
Some species of the Mortierellaceae family play important roles in ecosystems, such as promoting plant growth, breaking down plant debris, and remodeling rhizosphere microbial communities. In addition, fungi in this family have important applications in the field of biotechnology, for example as industrial producers of polyunsaturated fatty acids such as arachidonic acid; Metabolites of some species act as antibacterial and insecticides. [52,53] In recent years, significant progress has been made in taxonomic research in the family Mortierellaceae. Through multi-locus phylogenetic analyses, the Mortierella s.l. was reclassified into 13 monophyletic genera. These taxonomic adjustments provide a clearer framework for future research and contribute to a better understanding of the diversity and evolution. However, there are still many unsampled areas, especially in the geographical and ecological distribution on a global scale, which needs to be further explored.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. The GenBank accession number of the sequence used in this study (Table S2, S3).
Funding
This study was supported by the National Natural Science Foundation of China (Nos. 32170012, 32470004, 32300011). The Key Technological Innovation Program of Shandong Province, China (No. 2022CXGC020710), the Jinan City’s ‘New University 20 Policies’ Initiative for Innovative Research Teams Project (No. 202228028) and the Innovative Agricultural Application Technology Project of Jinan City (no. CX202210).
Conflict of interest
The authors declare no conflicts of interest.
Institutional Review Board Statement
Not applicable.
Author contributions
Conceptualization, X.-Y.J.; software, X.-Y.J. and Z.-Y.D.; Data curation, X.-Y.J.; methodology, X.-Y.J.; writing, review and editing, X.-Y.J. and X.Y.L.; formal analysis, F.L.; resources, Y.J. and Z.M.; supervision, X.-Y.L. and Z.M.; funding acquisition, X.-Y.L. All authors have read and agreed to the published version of the manuscript.
Data availability
The sequences of this study have been submitted to the NCBI database (https://www.ncbi.nlm.nih.gov/, accessed February 15, 2025) with accession numbers were shown in Tables S2 and S3.
References
- Wijayawardene, N.N.; Hyde, K.D.; Mikhailov, K.V.; Péter, G.; Aptroot, A.; Pires-Zottarelli, C.L.A.; Goto, B.T.; Tokarev, Y.S.; Haelewaters, D.; Karunarathna, S.C.; et al. Classes and phyla of the kingdom Fungi. Fungal Diversity 2024, 128, 1–165. [Google Scholar] [CrossRef]
- Smith, M.E.; Gryganskyi, A.; Bonito, G.; Nouhra, E.; Moreno-Arroyo, B.; Benny, G. Phylogenetic analysis of the genus Modicella reveals an independent evolutionary origin of sporocarp-forming fungi in the Mortierellales. Fungal Genet Biol. 2013, 61, 61–68. [Google Scholar] [CrossRef]
- Gams, W. A key to the species of Mortierella. Persoonia 1977, 9, 381–391. [Google Scholar]
- Petkovits, T.; Nagy, L.G.; Hoffmann, K.; Wagner, L.; Nyilasi, I.; Griebel, T.; Schnabelrauch, D.; Vogel, H.; Voigt, K.; Vágvölgyi, C.; et al. Data partitions, Bayesian analysis and phylogeny of the zygomycetous fungal family Mortierellaceae, inferred from nuclear ribosomal DNA sequences. PLoS One 2011, 6, e27507. [Google Scholar] [CrossRef] [PubMed]
- Wagner, L.; Stielow, B.; Hoffmann, K.; Petkovits, T.; Papp, T.; Vágvölgyi, C.; de Hoog, G.S.; Verkley, G.; Voigt, K. A comprehensive molecular phylogeny of the Mortierellales (Mortierellomycotina) based on nuclear ribosomal DNA. Persoonia 2013, 30, 77–93. [Google Scholar] [CrossRef]
- Linnemann, G. Die Mucorineen-Gattung Mortierella Coemans. Pflanzenforschung 1941, 23. [Google Scholar]
- Tedersoo, L.; Bahram, M.; Põlme, S.; Kõljalg, U.; Yorou, N.S.; Wijesundera, R.; Ruiz, L.V.; Vasco-Palacios, A.V.; Thu, P.Q.; Suija, A.; et al. Global diversity and geography of soil fungi. Science 2014, 346, 6213. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, Z.; Lv, X.; Li, Y.; Li, Z. High-throughput sequencing reveals the diversity and community structure of rhizosphere fungi of Ferula sinkiangensis at different soil depths. Scientific Reports 2019, 9, 6558. [Google Scholar] [CrossRef]
- Kuhnert, R.; Oberkofler, I.; Peintner, U. Fungal growth and biomass development is boosted by plants in snow-covered soil. Microbial Ecology 2012, 64, 79–90. [Google Scholar] [CrossRef]
- Holland, H.L. Biotransformation of organic sulfides. Natural Product Reports 2001, 18, 171–181. [Google Scholar] [CrossRef]
- Higashiyama, K.; Fujikawa, S.; Park, E.Y.; Shimizu, S. Production of arachidonic acid by Mortierella fungi. Biotechnology and Bioprocess Engineering 2002, 7, 252–262. [Google Scholar] [CrossRef]
- Yadav, D.R.; Kim, S.W.; Babu, A.G.; Adhikari, M.; Kim, C.; Lee, H.B.; Lee, L.S. First report of Mortierella alpina (Mortierellaceae, Zygomycota) isolated from crop field soil in Korea. Mycobiology 2014, 42, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Ward, O.P. Production of high yields of arachidonic acid in a fed-batch system by Mortierella alpina ATCC 32222. Applied Microbiology and Biotechnology 1997, 48, 1–5. [Google Scholar] [CrossRef]
- Du, Z.Y.; Alvaro, J.; Hyden, B.; Zienkiewicz, K.; Benning, N.; Zienkiewicz, A.; Bonito, G.; Benning, C. Enhancing oil production and harvest by combining the marine alga Nannochloropsis oceanica and the oleaginous fungus Mortierella elongata. Biotechnology for Biofuels 2018, 11, 174. [Google Scholar] [CrossRef]
- Li, F.; Zhang, S.; Wang, Y.; Li, Y.; Li, P.; Chen, L.; Jie, X.; Hu, D.; Feng, B.; Yue, K.; et al. Rare fungus, Mortierella capitata, promotes crop growth by stimulating primary metabolisms related genes and reshaping rhizosphere bacterial community. Soil Biology and Biochemistry 2020, 151, 108017. [Google Scholar] [CrossRef]
- Ozimek, E.; Hanaka, A. Mortierella species as the plant growth-prmoting fungi present in the agricultural soils. Agriculture 2021, 11, 7. [Google Scholar] [CrossRef]
- Shemshura, O.N.; Shemsheyeva, Z.N.; Sadanov, A.K.; Lozovicka, B.; Kamzolova, S.V.; Morgunov, L.V. Antifungal potential of organic acids produced by Mortierella alpina. International Journal of Engineering & Technology 2018, 7, 1218–1221. [Google Scholar] [CrossRef]
- Telagathoti, A.; Probst, M.; Mandolini, E.; Peintner, U. Mortierellaceae from subalpine and alpine habitats: new species of Entomortierella, Linnemannia, Mortierella, Podila and Tyroliella gen. nov. Stud Mycol. 2022, 103, 25–58. [Google Scholar] [CrossRef]
- Zhao, H.; Nie, Y.; Huang, B.; Liu, X.Y. Unveiling species diversity within early-diverging fungi from China I: three new species of Backusella (Backusellaceae, Mucoromycota). MycoKeys 2024, 109, 285–304. [Google Scholar] [CrossRef]
- Tao, M.F.; Ding, Z.Y.; Wang, Y.X.; Zhang, Z.X.; Zhao, H.; Meng, Z.; Liu, X.Y. Unveiling species diversity within early-diverging fungi from China II: Three new species of Absidia (Cunninghamellaceae,Mucoromycota) from Hainan Province. MycoKeys 2024, 110, 255–272. [Google Scholar] [CrossRef]
- Wang, Y.X.; Zhao, H.; Jiang, Y.; Liu, X.Y.; Tao, M.F.; Liu, X.Y. Unveiling species diversity within early-diverging fungi from China III: Six new species and a new record of Gongronella (Cunninghamellaceae, Mucoromycota). MycoKeys 2024, 110, 287–317. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.Y.; Ji, X.Y.; Tao, M.F.; Liu, W.X.; Jiang, Y.; Zhao, H.; Meng, Z.; Liu, X.Y. Unveiling species diversity within early-diverging fungi from China IV: Four new species of Absidia (Cunninghamellaceae, Mucoromycota). MycoKeys 2025. (in review).
- Ji, X.Y.; Ding, Z.Y.; Nie, Y.; Zhao, H.; Wang, S.; Huang, B.; Liu, X.Y. Unveiling species diversity within early-diverging fungi from China V: Five new species of Absidia (Cunninghamellaceae, Mucoromycota). MycoKeys 2025. (in review).
- Wang, Y.X.; Ding, Z.Y.; Ji, X.Y.; Meng, Z.; Liu, X.Y. Unveiling species diversity within early-diverging fungi from China Ⅵ: Four Absidia sp. nov. (Mucorales) in Guizhou and Hainan. Microorganisms 2025. (in review).
- Ding, Z.Y.; Tao, M.F.; Ji, X.Y.; Jiang, Y.; Liu, W.X.; Wang, S.; Liu, X.Y. Unveiling species diversity within early-diverging fungi from China Ⅶ: Seven new species of Cunninghamella (Cunninghamellaceae, Mucoromycota). J. Fungi 2025. (in review).
- Zou, Y.; Hou, J.; Guo, S.; Li, Z.; Stephenson, S.L.; Pavlov, I.N.; Liu, P.; Li, Y. Diversity of dictyostelid cellular slime molds, including two species new to science, in forest soils of Changbai Mountain, China. Microbiol Spectr 2022, 10, 5. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Zou, Y.; Li, S.; Stephenson, S.L.; Wang, Q.; Li, Y. Two new species of dictyostelid cellular slime molds in high-elevation habitats on the Qinghai-Tibet Plateau, China. Sci Rep 2019, 9, 5. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Lv, M.L.; Liu, Z.; Zhang, M.Z.; Wang, Y.N.; Ju, X.; Song, Z.; Ren, L.Y.; Jia, B.S.; Qiao, M.; et al. High-yield oleaginous fungi and high-value microbial lipid resources from Mucoromycota. BioEnerg. Res. 2021, 14, 1196–1206. [Google Scholar] [CrossRef]
- Corry, J.E.L. Rose bengal chloramphenicol (RBC) agar. In Progress in Industrial Microbiology; Elsevier: Amsterdam, The Netherlands, 1995; pp. 431–433. [Google Scholar]
- Jiang, Y.; Zhang, Z.; Zhang, J.; Wang, S.; Zhang, X. Morphological and phylogenetic analyses reveal three new species of Phyllosticta (Botryosphaeriales, Phyllostictaceae) in China. J. Fungi 2024, 10, 7. [Google Scholar] [CrossRef]
- Zhang, Z.X.; Liu, R.Y.; Liu, S.B.; Mu, T.C.; Zhang, X.G.; Xia, J.W. Morphological and phylogenetic analyses reveal two new species of Sporocadaceae from Hainan, China. MycoKeys 2022, 88, 171–192. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, F.J.R.M.; Lee, S.H.; Taylor, L.; Shawe-Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protocols: A Guide to Methods and Applications 1990, 31, 315–322. [Google Scholar] [CrossRef]
- Hurdeal, V.G.; Jones, E.B.G.; Gentekaki, E. Absidia zygospore (Mucoromycetes), a new species from Nan province, Thailand. Studies in Fungi 2023, 8, 15. [Google Scholar] [CrossRef]
- Stiller, J.W.; Hall, B.D. The origin of red algae: implications for plastid evolution. Proceedings of the National Academy of Sciences of the United States of America 1997, 94, 4520–4525. [Google Scholar] [CrossRef]
- Voigt, K.; Wöstemeyer, J. Reliable amplification of actin genes facilitates deep-level phylogeny. Microbiological Research 2000, 155, 179–195. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef] [PubMed]
- Nie, Y.; Cai, Y.; Gao, Y.; Yu, D.S.; Wang, Z.M.; Liu, X.Y.; Huang, B. Three new species of Conidiobolus sensu stricto from plant debris in eastern China. MycoKeys 2020, 73, 133–149. [Google Scholar] [CrossRef] [PubMed]
- Nie, Y.; Yu, D.S.; Wang, C.F.; Liu, X.Y.; Huang, B. A taxonomic revision of the genus Conidiobolus (Ancylistaceae, Entomophthorales): Four clades including three new genera. MycoKeys 2020, 66, 55–81. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. 2010 Gateway Computing Environments Workshop (GCE), 2010, 1–8. [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Molecular Biology and Evolution 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Sys. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Wagner, L.; Stielow, B.; Hoffmann, K.; Petkovits, T.; Papp, T.; Vágvölgyi, C.; de Hoog, G.S.; Verkley, G.; Voigt, K. A comprehensive molecular phylogeny of the Mortierellales (Mortierellomycotina) based on nuclear ribosomal DNA. Persoonia 2013, 30, 77–93. [Google Scholar] [CrossRef]
- Hoffmann, K.; Telle, S.; Walther, G.; Eckhart, M.; Kirchmair, M.; Prillinger, H.; Prazenica, A.; Newcombe, G.; Dölz, F.; TamásPapp.; et al. Diversity, genotypic identification, ultrastructural and phylogenetic characterization of zygomycetes from different ecological habitats and climatic regions: Limitations and utility of nuclear ribosomal DNA barcode markers. Mycology 2008, 263–312.
- Kuhnert, R.; Oberkofler, I.; Peintner, U. Fungal growth and biomass development is boosted by plants in snow-covered soil. Microbial Ecology 2012, 64, 79–90. [Google Scholar] [CrossRef]
- Dixon-Stewart, D. Species of Mortierella isolated from soil. Trans Br Mycol Soc. 1932, 17, 208–220. [Google Scholar] [CrossRef]
- Wu, F.; Yuan, H.S.; Zhou, L.W.; Yuan, Y.; Cui, B.K.; Dai, Y.C. Polypore diversity in south China. Mycosystema 2020, 39, 653–682. [Google Scholar] [CrossRef]
- Wang, K.; Chen, S.L.; Dai, Y.C.; Jia, Z.F.; Li, T.H.; Liu, T.Z.; Phurbu, D.; Mamut, R.; Sun, G.Y.; Bau, T.; et al. Overview of China’s nomenclature novelties of fungi in the new century (2000-2020). Mycosystema 2021, 40, 822–833. [Google Scholar] [CrossRef]
- Karunarathna, S.C.; Dong, Y.; Karasaki, S.; Tibpromma, S.; Hyde, K.D.; Lumyong, S.; Xu, J.; Sheng, J.; Mortimer, P.E. Discovery of novel fungal species and pathogens on bat carcasses in a cave in Yunnan Province, China. Emerg Microbes Infect. 2020, 9, 1554–1566. [Google Scholar] [CrossRef]
- Vandepol, N.; Liber, J.; Desirò, A.; Na, H.; Kennedy, M.; Barry, K.; Grigoriev, I.V.; Miller, A.N.; O'Donnell, K.; Stajich, J.E.; Bonito, G. Resolving the Mortierellaceae phylogeny through synthesis of multi-gene phylogenetics and phylogenomics. Fungal Divers. 2020, 104, 267–289. [Google Scholar] [CrossRef]
- DiLegge, M.J.; Manter, D.K.; Vivanco, J.M. A novel approach to determine generalist nematophagous microbes reveals Mortierella globalpina as a new biocontrol agent against Meloidogyne spp. nematodes. Scientific Reports 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Edgington, S.; Thompson, E.; Moore, D.; Hughes, K.A.; Bridge, P. Investigating the insecticidal potential of Geomyces (Myxotrichaceae: Helotiales) and Mortierella (Mortierellacea: Mortierellales) isolated from Antarctica. SpringerPlus 2014, 3, 289. [Google Scholar] [CrossRef]
Figure 1.
The ML phylogenetic tree of the genus Linnemannia based on the concatenated alignment of ITS, LSU, SSU, RPB1 and Act sequences, with Mortierella cogitans serving as outgroup. Branches are labeled with Maximum Likelihood Bootstrap Value (left, MLBV≥70) and Bayesian Inference Posterior Probability (right, BIPP≥0.90), which are separated by a slash "/". New species are highlighted in red. Branches shortened due to space constrains are indicated by double slashes "//" and the number of folds. Strains marked with an asterisk "*" and in bold represent ex-type or ex-holotypes. The bottom-right scale bar indicates 0.1 substitutions per site.
Figure 1.
The ML phylogenetic tree of the genus Linnemannia based on the concatenated alignment of ITS, LSU, SSU, RPB1 and Act sequences, with Mortierella cogitans serving as outgroup. Branches are labeled with Maximum Likelihood Bootstrap Value (left, MLBV≥70) and Bayesian Inference Posterior Probability (right, BIPP≥0.90), which are separated by a slash "/". New species are highlighted in red. Branches shortened due to space constrains are indicated by double slashes "//" and the number of folds. Strains marked with an asterisk "*" and in bold represent ex-type or ex-holotypes. The bottom-right scale bar indicates 0.1 substitutions per site.

Figure 2.
The ML phylogenetic tree of the genus Mortierella based on the concatenated alignment of ITS, LSU, SSU, RPB1 and Act sequences, with Umbelopsis isabellina and U. actotrophica serving as outgroups. Branches are labeled with Maximum Likelihood Bootstrap Value (left, MLBV≥70) and Bayesian Inference Posterior Probability (right, BIPP≥0.90), which are separated by a slash "/". New species are highlighted in red. Branches shortened due to space constrains are indicated by double slashes "//" and the number of folds. Strains marked with an asterisk"*" and in bold represent ex-types or ex-holotypes. The bottom-right scale bar indicates 0.1 substitutions per site.
Figure 2.
The ML phylogenetic tree of the genus Mortierella based on the concatenated alignment of ITS, LSU, SSU, RPB1 and Act sequences, with Umbelopsis isabellina and U. actotrophica serving as outgroups. Branches are labeled with Maximum Likelihood Bootstrap Value (left, MLBV≥70) and Bayesian Inference Posterior Probability (right, BIPP≥0.90), which are separated by a slash "/". New species are highlighted in red. Branches shortened due to space constrains are indicated by double slashes "//" and the number of folds. Strains marked with an asterisk"*" and in bold represent ex-types or ex-holotypes. The bottom-right scale bar indicates 0.1 substitutions per site.


Table 1.
PCR information used in this study.
| Loci | PCR primers | Primer sequence (5’ – 3’) | PCR cycles | References |
|---|---|---|---|---|
| ITS | ITS5 | GGA AGT AAA AGT CGT AAC AAG G | 95 °C 5 min; (95 °C: 30 s, 55 °C: 30 s, 72 °C: 1 min) × 35 cycles; 72 °C 10 min | [32] |
| ITS4 | TCC TCC GCT TAT TGA TAT GC | |||
| LSU | LR0R | GTA CCC GCT GAA CTT AAG C | 95 °C 5 min; (94 °C: 30 s, 52 °C: 45 s, 72 °C: 1.5 min) × 30 cycles; 72 °C 10 min | [33] |
| LR5 | TCC TGA GGG AAA CTT CG | |||
| RPB1 |
RPB1-Af |
GAR TGY CCD GGD CAY TTY GG |
95 °C 3 min; (94 °C: 40 s, 60 °C: 40 s, 72 °C: 2 min) × 9 (94 °C: 45 s, 55 °C: 1.5 min, 72 °C: 2 min) × 37 cycles; 72 °C 10 min | [34] |
| RPB1-Cr | CCN GCD ATN TCR TTR TCC ATR TA | |||
| Act | ACT-1 | TGG GAC GAT ATG GAI AAI ATC TGG CA | 95 °C 3 min; (95 °C: 60 s, 55 °C: 60 s, 72 °C: 1 min) × 30 cycles; 72 °C 10 min | [35] |
| ACT-4R | TC ITC GTA TIC TIG CTI IGA IAT CCA CA T | |||
| SSU | NS1 | GTA GTC ATA TGC TTG TCT CC | 95 °C 5 min; (94 ℃: 60 s, 54 ℃: 50 s, 72 ℃: 1 min) × 37 cycles; 72 °C 10 min | [32] |
| NS4 | CTT CCG TCA ATT CCT TTA AG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



