
Submitted:
12 April 2025
Posted:
15 April 2025
You are already at the latest version
Abstract
Absidia represents the most species-rich genus within the family Cunninghamellaceae, with its members commonly isolated from diverse substrates, particularly rhizosphere soil. In this study, four novel Absidia species (A. irregularis sp. nov., A. multiformis sp. nov., A. ovoidospora sp. nov., and A. verticilliformis sp. nov.) were discovered from south and southwestern Chinese soil samples through integrated morphological and molecular phylogenetic analyses. Phylogenetic analyses based on concatenated ITS, SSU, LSU, Act, and TEF1α sequence data reconstructed trees that strongly supported the monophyly of these four new taxa each. Key diagnostic features include A. irregularis (closely related to A. oblongispora) exhibiting irregular colony morphology, A. multiformis (sister to A. heterospora) demonstrating polymorphic sporangiospores, A. ovoidospora (forming a clade with A. panacisoli and A. abundans) producing distinctive ovoid sporangiospores, and A. verticilliformis (next to A. edaphica) displaying verticillately branched sporangiophores. Each novel species is formally described with comprehensive documentation, including morphological descriptions, illustrations, Fungal Names registration identifiers, designated type specimens, etymological explanations, maximum growth temperatures, and taxonomic comparisons. This work constitutes the sixth installment in a series investigating early-diverging fungal diversity in China, expanding the recognized Absidia species to 71. The findings enhance our understanding of mucoralean biodiversity in Asian tropical and subtropical ecosystems.
Keywords:
Mucoromycota
; soil-borne fungi
; new species
; taxonomy
; molecular phylogeny
1. Introduction
The genus Absidia Tiegh. (Mucoromycota Doweld, Mucoromycetes Doweld, Mucorales Dumort., Cunninghamellaceae Naumov ex R.K. Benj.) was established in 1878 and typified with A. reflexa [1, 2]. It is currently the most numerous genus in the family Cunninghamellaceae, with 67 recognized species (https://www.catalogueoflife.org/data/taxon/LSM, accessed on 2 December 2024). Its taxonomic position has been controversial since its establishment. Initially, this genus was classified in Absidiaceae [3]. Then, Benny et al. transferred it to Mucoraceae in 2001 [4]. Finally, Ashton et al. accommodated it in Cunninghamellaceae in 2009 [5]. With the deepening of research, Absidia s.l. was divided into Lichtheimia Vuill. (thermotolerant, optimum growth temperature 37–45 ℃), Absidia s.s. (mesophilic, optimum growth temperature 25–34 ℃), and Lentamyces Kerst. Hoffm. & K. Voigt (parasitic on other mucoralean fungi, optimum growth temperature 14–25 ℃) [6-9].
Strains of Absidia are distributed worldwide, ubiquitous in soil, dung, leaf litter, food, air, etc [10]. In the GlobalFungi database (https://globalfungi.com/, accessed on 22 December 2024), Absidia members are recorded from soil (37,828 records, 44.52 % of all records), root (15,865, 18.67 %), shoot (11,740, 13.82 %), topsoil (6,923, 8.15 %), rhizosphere soil (2,530, 2.98 %), deadwood (2,253, 2.65 %), air (2,080, 2.45 %), water (1,549, 1.82 %), litter (1,406, 1.65 %), mosses (265, 0.41 %), lichen (217, 0.25 %), coral (72, 0.08 %), dust (53, 0.06 %), fungal sporocarp (31, 0.04 %), and glacial ice debris (3, 0 %). In summary, soil and rhizosphere substrates account for approximately 74.32%.
Absidia coerulea, the most common species in this genus, plays an important role in bioengineering. It possesses the capability of transforming spirulina biomass into (-)-α-bisabolol [11]. It is able to catalyze the specific C-3 dehydrogenation for derivatives of ginsenoside-Rg₁, as well as hydroxylation at the 7β and 15α positions. And some metabolites in this species exhibit moderate reversal activity towards multidrug-resistant tumor cells [12].
Recently, eight fungal strains were isolated from soil in Hainan and Guizhou provinces, the south and southwest region of China. According to ITS-SSU-LSU-Act-TEF1α molecular phylogenetic analyses and morphological comparisons, these strains were classified into four new species of Absidia and described herein as A. irregularis sp. nov., A. multiformis sp. nov., A. ovoidospora sp. nov., and A. verticilliformis sp. nov. This is the sixth report of a serial work on diversity of Chinese early-diverging fungi [13-17].
2. Materials and Methods
2.1. Isolation and Morphology
Soil samples were collected in Hainan province in April 2023 and Guizhou province in August 2023, following the methods by Li et al.and Zou et al. [18, 19]. Soil sample collection started with removal of surface contaminants using a sterilized stainless steel shovel. About 100 g of homogenized soil was put into sample bags, labeled with collection date, administrative location, GPS coordinates, and altitude. Strains were obtained from the soil samples by serial dilution spread plate and single spore isolation.
About 1 g of soil samples was mixed with 10 mL sterile water to prepare 10-1 soil suspension. One millilitre of the 10-1 suspension was transferred to 9 mL of sterile water to obtain a 10-2 soil suspension. In the same way, 10-3 and 10-4 soil suspensions were made. Approximately 0.2 mL of the final 10-4 soil suspension were dispersed evenly with sterilised coating rods on Rose-Bengal Chloramphenicol agar (RBC: peptone 5.00 g/L, glucose 10.00 g/L, KH2PO4 1.00 g/L, MgSO4·7H2O 0.50 g/L, rose red 0.05 g/L, chloramphenicol 0.10 g/L, agar 15.00 g/L) [20], and then cultured in the dark at 25 ℃. Once visible, colonies were transferred and further cultured on Potato Dextrose Agar (PDA: glucose 20.00 g/L, potato 200.00 g/L, agar 20.00 g/L, pH 7) [14, 21].
Appropriately 0.1 g of soil samples were evenly sprinkled on the RBC medium with 0.06 mg/mL streptomycin and then incubated in darkness at 25 ℃ for 2–5 d. When sporangia formed, a sterilized inoculation needle was adopted to pick up a sporangium onto the PDA medium supplemented with 0.06 mg/mL streptomycin.
The maximum growth temperature was determined by a gradient method [13]. The strain was initially cultured at 25℃ for 2 d, and then increased by 1℃ per day until no growths.
Pure cultivation was applied for observing anamorphs, and pairing experiments were carried out for observing zygospores by adding 0.1 % lecithin to PDA and sealing Petri dishes to retain moisture. The morphological characteristics were observed with an optical microscope (Olympus BX53) and photographed with a high-definition colour digital camera (Olympus DP80). Each morphological character was statistically calculated against 30 measurements [22]. All strains were stored at 4 ℃ with 20 % sterilised glycerine. Cultures were deposited in the China General Microbiological Culture Collection Center, Beijing, China (CGMCC) and the Shandong Normal University, Jinan, Shandong, China (XG). Strains were deposited in the Herbarium Mycologicum Academiae Sinicae, Beijing, China (Fungarium, HMAS). Taxonomic information for the new taxa was registered in the Fungal Names repository (https://nmdc.cn/fungalnames/).
2.2. DNA Extraction and Amplification
Genomic DNA was extracted from mycelia using the CTAB method and GOMagTM Rapid Plant DNA Kit [21]. Information of the primers for PCR amplification is listed in Table 1. Amplification was performed in a final volume of 20 μL, containing 10 μL 2× Hieff Canace® Plus PCR Master Mix (Cat No.10154ES03; Yeasen Biotechnology, Shanghai, China), 0.5 μL of forward and reverse primers each (10 μM; TsingKe, Beijing, China), 1 μL template genomic DNA (1 μM) and 8 μL distilled deionised water. Molecular loci, PCR primers and programmes used in this study are listed in Table 1. PCR products were electrophoresed with 1 % agarose gel. DNA fragments were stained with TS-GelRed Nucleic Acid Gel Stain (10,000× in Water; TSJ002; Beijing Tsingke Biotech Co., Ltd.) and observed under ultraviolet light. Then a gel extraction kit (Cat# AE0101-C; Shandong Sparkiade Biotechnology Co., Ltd.) was used for gel recovery. Sanger sequencing was carried out by Biosune Co., Ltd. (Shanghai, China). Consensus sequences were assembled using MEGA v.7.0 [23]. Target sequences in some strains could not be obtained by conventional methods due to heterogeneous gene duplications, and thus they were extracted from genomic data [24]. The genomes were sequenced by Singke Biotech Co., Ltd. (Jinan, China). All sequences generated in this study were deposited at GenBank under the accession numbers in Table 2.
Relative sequences were obtained by BLAST search against the NCBI GenBank nucleotide database. SSU, ITS, LSU, Act and TEF1α sequences both generated herein and retrieved from GenBank (Table 2) were individually aligned using MAFFT 7 online service. The aligned matrices were manually proofread and then jointly analyzed. The optimal evolutionary model was determined for each partition and included in the analysis using MrModelTest v.2.3 [29]. Phylogenetic history was reconstructed using maximum likelihood (ML) algorithm with RaxML-HPC2 on XSEDE (8.2.12) and Bayesian inference (BI) algorithm with MrBayes [30-32]. Maximum Likelihood analysis was performed using the best model with 1,000 bootstrap replications. The BI analysis consisted of five million generations with four parallel runs under stopping rules and a sampling frequency of 100 generations. The burn-in score was set to 0.25, and the posterior probability (PP) was determined from the remaining trees. Initial adjustments to the phylogenetic tree were made using FigTree v.1.4.4 (http://tree.bio.ed.ac.uk/software/figtree/) and the finalization was performed using Adobe Illustrator CC 2019 (https://adobe.com/products/illustrator).
3. Results
3.1. Phylogenetic Analyses
The sequence matrix included 87 strains in 64 species of Absidia, with Cunninghamella blakesleeana CBS 782.68 as outgroup. A total of 4,656 characters comprised ITS rDNA (1–975), SSU rDNA (976–2,017), LSU rDNA (2,018–3,030), Act (3,031–3,689) and TEF1α (3,690–4,656). As many as 2,737 characters were constant, while 706 and 1,213 among the variable characters were parsimony-uninformative and informative, respectively (Supplementary File S1). MrModelTest suggested that the Dirichlet fundamental frequency and GTR-I-G evolution pattern for all partitions were adopted in Bayesian Inference. The topology of the Bayesian tree, consistent with that of the ML tree, was used as a representative to summarise the evolutionary history within the genus Absidia (Figure 1), exhibiting the phylogenetic placement of the four new species. A. irregularis is related to A. oblongispora with full supports (MLBV = 100, BIPP = 1.00), A. ovoidrospora is closely related to A. panacisoli and A. abundans with full supports (MLBV = 100, BIPP = 1.00). A. multiformis is most closely related to A. heterospora with high supports (BIPP = 0.99). And A. verticilliformis is closely related to A. edaphica with robust supports (MLBV = 94, BIPP = 1).
3.2. Taxonomy
Absidia irregularisYi Xin Wang & X.Y. Liu, sp. nov. Figure 2
Fungal Names—FN 572283
Etymology—The epithet “irregularis” (Latin) pertaining to irregular colonies.
Type—China, Hainan, Changjiang Li Autonomous County, Bawangling National Forest Park, 19.0859333° N, 109.122752° E, altitude 745.3 m, from soil sample, August 9, 2023, Yi-Xin Wang (Holotype HMAS 353186, ex-holotype strain CGMCC 3.27812 = XG05674-6)
Description—Colonies growing moderately on PDA in darkness at 25 ℃ for 7 d, reaching 43.2–54.8 mm in diameter, initially white, soon becoming gray to brown, irregular and scaly at edge, cottony, reversely white to gray. Hyphae hyaline at first, brownish when mature, branched, irregular, 10.2–14.5 μm in diameter. Stolons hyaline, branched, and smooth. Rhizoids hyaline, branched, irregular or root-like. Sporangiophores on aerial mycelia, hyaline, erect or slightly curved, unbranched or slightly branched, swollen usually below sporangia, umbellately or sympodially branched, often with a septum below apophyses, 26.5–148.1 × 3.4–5.3 μm. Sporangia oval to subglobose, 27.2–32.4 × 26.5–28.9 μm, hyaline at first and then brown, deliquescent-walled, leaving a collar after releasing sporangiospores. Apophyses hyaline, smooth, bowl-shaped, 2.9–9.6 × 8.6–19.0 μm. Collars distinct. Columellae hyaline or brown, hemispherical, 10.0–19.8 × 9.8–21.0 μm, with a protruding (3.5–6.3 × 1.4–2.9 μm) at the top. Protrudings always slightly contracted in the middle. Sporangiospores hyaline, smooth, not uniform, mostly cylindrical, 3.6–4.6 × 2.3–2.9 μm. Chlamydospores present. Zygospores not observed.
Additional strain examined—China, Hainan, Changjiang Li Autonomous County, Bawangling National Forest Park, 19.0859333° N, 109.122752° E, altitude 745.3 m, from soil sample, August 9, 2023, Yi-Xin Wang (living culture XG05674-8).
GenBank accession numbers—CGMCC 3.27812 (ITS, PQ306325; LSU, PQ289020; Act, PQ807209; SSU, PQ799254; TEF1α, PV126019), XG05674-7 (ITS, PQ306326; LSU, PQ289021; Act, PQ807210; SSU, PQ799255; TEF1α, PV126020).
Maximum growth temperature—CGMCC 3.27812 (32 ℃), XG05674-7 (32 ℃).
Notes—Based on phylogenetic analyses of ITS-SSU-LSU-Act-TEF1α sequences, the two isolates of the new species Absidia irregularis formed an independent clade with full supports (MLBV = 100, BIPP = 1; Figure 1), which is closely related to A. oblongispora (BIPP = 1; Figure 1). This new species differs morphologically from A. oblongispora in apophysis and columella [33]. The new species is bigger than A. oblongispora in apophysis, 2.9–9.6 × 8.6–19.0 μm vs. 3.5–6.5 × 3.5–7.5 μm. The new species is also bigger than A. oblongispora in columellae, 10.0–19.8 × 9.8–21.0 μm vs. 7.0–15.0 × 8.5–16.5 μm. Combining the morphological and molecular phylogenetic analyses, the two isolates were classified as a new taxon, allied to A. oblongispora.
Absidia multiformisYi Xin Wang & X.Y. Liu, sp. nov. Figure 3
Fungal Names—FN 572285
Etymology—The epithet " multiformis " (Latin) pertaining to polymorphic sporangiospores.
Type—China, Hainan, Lingshui Li Autonomous County, 18.6958768° N, 109.9407998° E, altitude 151.6 m, from soil sample, April 23, 2023, Yi-Xin Wang (Holotype HMAS 353194, ex-holotype strain CGMCC 3.27807 = XG04016-2).
Description—Colonies growing moderately on PDA in darkness at 25 ℃ for 7 d, reaching 82.2–90.0 mm diameter, initially white, soon becoming gray to brown, irregular at edge, cottony, reversely white to gray. Hyphae hyaline at first, brownish when mature, branched, irregular, 4.2–12.3 μm in diameter. Stolons hyaline, branched, and smooth. Rhizoids hyaline, branched, irregular or root-like. Sporangiophores on aerial mycelia, hyaline, erect or slightly curved, unbranched or slightly branched, swollen usually below sporangia, often with a septum below apophyses, 49.9–125.4 × 2.9–3.7 μm. Sporangia oval to subglobose, 20.1–36.1 × 23.1–36.3 μm, hyaline at first and then brown, deliquescent-walled, leaving a collar after releasing sporangiospores. Apophyses hyaline, smooth, shallow mouth bowl shaped, 2.5–7.9 × 8.0–19.8 μm. Collars distinct. Columellae hyaline or brown, hemispherical, 5.3–19.4 × 8.3–22.9 μm, with long or short cylindrical protrusions at top, 1.7–6.4 × 1.1–4.0 μm. Sporangiospores hyaline, smooth, not uniform, mainly cylindrical, 4.5–6.1 × 2.5–4.4 μm, some ovoid, 4.4–5.8 × 3.9–6.1 μm, occasionally subglobose, 4.9–7.2 μm. Chlamydospores present. Zygospores present, not uniform.
Additional strain examined—China, Hainan, Lingshui Li Autonomous County, 18.6958768° N, 109.9407998° E, altitude 151.6 m, from soil sample, April 23, 2023, Yi-Xin Wang (living culture XG04016-3).
GenBank accession numbers—CGMCC 3.27807 (ITS, PQ306319; LSU, PQ803168; Act, PQ807203; SSU, PQ799260; TEF1α, PV126021), XG0 4016-3 (ITS, PQ306320; LSU, PQ803169; Act, PQ807204; SSU, PQ799261; TEF1α, PV126022).
Maximum growth temperature—CGMCC 3.27807 (32 ℃), XG0 4016-3 (32 ℃).
Notes—Based on phylogenetic analyses of ITS-SSU-LSU-Act-TEF1α sequences, the two isolates of the new species Absidia multiformis formed an independent clade with full supports (MLBV = 100, BIPP = 1; Figure 1), which is closely related to A. heterospora (BIPP = 0.99; Figure 1). This new species differs morphologically from A. heterospora in sporangiophore, sporangium, apophysis and columella [34]. It differs from A. heterospora by narrower and shorter sporangiophores, 49.9–125.4 × 2.9–3.7 μm vs. 230–1700 × 4.3–10.0 µm. It is smaller than A. heterospora in sporangia, 20.1–36.3 μm vs. 15.0–55.0 µm. In columellae, it differs from A. heterospora by more shapes and smaller size. In detail, it is hemispherical and 5.3–19.4 × 8.3–22.9 μm, while A. heterospora is regularly dorsiventrally flattened and 10.5–34 µm in diameter. Combining morphological and molecular phylogenetic analyses, the two isolates were classified as a new taxon, allied to A. heterospora.
Absidia ovoidosporaYi Xin Wang & X.Y. Liu, sp. nov. Figure 4
Fungal Names—FN 572284
Etymology—The epithet “ovoidospora” (Latin) pertaining to ovoid sporangiospores.
Type—China, Hainan, Changjiang Li Autonomous County, Bawangling National Forest Park, 25.905722° N, 107.279063° E, altitude 745.3 m, from soil sample, August 9, 2023, Yi-Xin Wang (Holotype HMAS 353185, ex-holotype strain CGMCC 3.27811 = XG05673-2).
Description—Colonies growing fast on PDA in darkness at 25 ℃ for 7 d, reaching 90 mm diameter, initially white, soon becoming gray to brown, irregular at edge, cottony, reversely white to gray. Hyphae hyaline at first, brownish when mature, branched, irregular, 5.8–13.7 μm in diameter. Stolons hyaline, branched, and smooth. Rhizoids hyaline, branched, irregular or root-like. Sporangiophores on aerial mycelia, hyaline, erect or slightly curved, unbranched or slightly branched, swollen usually present below sporangia, umbellately or sympodially branched, often with a septum below apophyses, 45.3–355.3 × 2.5–4.0 μm. Sporangia oval to subglobose, 13.6–29.0 × 13.3–28.5 μm, hyaline at first and then brown, deliquescent-walled, leaving a collar after releasing sporangiospores. Apophyses hyaline, smooth, bowl shaped and long funnel shaped, 4.1–6.9 × 8.6–10.9 μm. Columellae hyaline or brown, hemispherical with a short or long protruding at the top, 7.9–12.9 × 9.0–11.6 μm. Protrudings always slightly contracted in the middle, 1.9–4.6 × 1.7–2.6 μm. Sporangiospores hyaline, smooth, not uniform, mostly ovoid, 3.2–3.8 × 2.4–3.1 μm, some cylindrical, 3.4–4.6 × 2.2–3.0 μm μm. Chlamydospores present. Zygospores not observed.
Additional strains examined—China, Hainan, Changjiang Li Autonomous County, Bawangling National Forest Park, 25.905722° N, 107.279063° E, altitude 745.3 m, from soil sample, August 9, 2023, Yi-Xin Wang (living culture XG05673-3).
GenBank accession numbers—CGMCC 3.27811 clone1 (ITS, PQ306327; LSU, PQ803164; Act, PQ807207; SSU, PQ799256; TEF1α, PV126015), CGMCC 3.27811 clone2 (ITS, PV069753; LSU, PQ803165; Act, PV126023; SSU, PQ799257; TEF1α, PV126017), XG05673-3 clone 1 (ITS, PQ306328; LSU, PQ803166; Act, PQ807208; SSU, PQ799258; TEF1α, PV126016), XG05673-3 clone 2 (ITS, PV069754; LSU, PQ803167; Act, PV126024; SSU, PQ799259; TEF1α, PV126018).
Maximum growth temperature—CGMCC 3.27811 (30 ℃), XG05673-3 (30 ℃).
Notes—Based on phylogenetic analyses of ITS-SSU-LSU-Act-TEF1α sequences, the new species Absidia ovoidospora formed two sister clades with high supports (MLBV = 96, BIPP = 1; Figure 1), which are closely related to A. panacisoli and A. abundans with full supports (MLBV = 100, BIPP = 1; Figure 1). These two clades were resulted from two clones. This new species differs morphologically from A. oblongispora in sporangiophore and sporangium [35]. The new species differs from A. panacisoli by wider sporangiophore, 2.5–4.0 μm vs. 1.7–2.8 μm. In sporangia, the new species is bigger than A. panacisoli, 13.6–29.0 × 13.3–28.5 μm vs. 13.0–21.5 × 9.4–15.5 μm. This new species differs morphologically from A. abundans in sporangiophore, sporangium and columella [9]. The new species differs from A. abundans by bigger sporangiophores, 45.3–355.3 × 2.5–4.0 μm vs. 35.0–170.0 × 2.0–3.5 μm. The new species is bigger than A. abundans in sproangia, 13.6–29.0 × 13.3–28.5 μm vs. 8.0–16.5 × 8.5–16.0 μm. In columellae, the new species is bigger than A. abundans, 7.9–12.9 × 9.0–11.6 μm vs. 4.5–10.0 × 3.5–8.0 μm. Combining morphological and molecular phylogenetic analyses, the two were classified isolates as a new taxon, allied with A. panacisoli and A. abundans.
Absidia verticilliformis, Yi Xin Wang & X.Y. Liu sp. nov. Figure 5
Fungal Names—FN 572287
Etymology—The epithet “verticilliformis” (Latin) pertaining to verticillate branches of sporangiophores.
Type—China, Hainan, Sanya City, Jiyang District, G224 Haiyu Middle Line, 18.391817° N, 109.641068° E, altitude 193.0 m, from soil sample, April 24, 2023, Yi-Xin Wang (Holotype HMAS 353183, ex-holotype strain CGMCC 3.27810 = XG04088-3)
Description—Colonies growing fast on PDA in darkness at 25 ℃ for 7 days, reaching 90 mm diameter, initially white, soon becoming gray to brown, irregular at edge, cottony, reversely white to gray, growing outward in a petal shape. Hyphae hyaline at first, brownish when mature, branched, 4.4–8.9 μm in diameter. Stolons hyaline, branched, and smooth. Rhizoids hyaline, branched, irregular or root-like. Sporangiophores on aerial mycelia, hyaline, erect or slightly curved, unbranched or slightly branched, mostly umbellately branched, swollens usually present below sporangia, often with a septum below apophyses, 22.3–368.7 × 3.3–4.6 μm. Sporangia oval to subglobose, 15.7–29.0 × 17.5–26.0 μm, hyaline at first and then brown, deliquescent-walled, mostly leaving a collar after releasing sporangiospores. Apophyses hyaline, smooth, shallow mouth bowl shaped, 4.5–14.4 × 11.1–20.4 μm. Collars distinct. Columellae hyaline or brown, hemispherical, tip with short or long cylindrical protrusions, 12.6–25.6 × 14.7–26.2 μm. Protrudings 3.1– 6.3× 1.8–2.2 μm. Sporangiospores hyaline, smooth, not uniform, mostly ovoid, 3.4–4.3 × 2.3–3.1 μm, some cylindrical, 3.9–4.4 × 2.1–3.0 μm. Chlamydospores present. Zygospores not observed.
Additional strain examined—China, Hainan, Sanya City, Jiyang District, G224 Haiyu Middle Line, 18.391817° N, 109.641068° E, altitude 193.0 m, from soil sample, April 24, 2023, Yi-Xin Wang (living culture XG04088-4)
GenBank accession numbers—CGMCC 3.27810 clone 1 (ITS, PQ306315; LSU, PQ803170; Act, PQ807205; SSU, PQ799262; TEF1α, PV126011), CGMCC 3.27810 clone 2 (ITS, PV069755; LSU, PQ803171; Act, PV126025; SSU, PQ799263; TEF1α, PV126013), XG04088-4 clone1 (ITS, PQ306316; LSU, PQ803172; Act, PQ807206; SSU, PQ799264; TEF1α, PV126012), XG04088-4 clone2 (ITS, PV069756; LSU, PQ803173; Act, PV126026; SSU, PQ799265; TEF1α, PV126014).
Maximum growth temperature—CGMCC 3.27810 (34 ℃), XG04088-4 (34 ℃).
Notes—Based on phylogenetic analyses of ITS-SSU-LSU-Act-TEF1α sequences, the two isolates of the new species Absidia verticilliformis formed two sister clades with full supports (MLBV = 100, BIPP = 1; Figure 1), which is closely related to A. edaphica (MLBV = 94, BIPP = 1; Figure 1). This new species differs morphologically from A. edaphica in sporangium, columella and sporangiospore [36]. The new species is smaller than A. edaphica in sporangia, 15.7–29.0 × 17.5–26.0 μm vs. 30.5–35.5 × 24–27 μm. In columellae, the new species is bigger than A. edaphica, 12.6–25.6 × 14.7–26.2 μm vs. 5–9.5 × 6.5–20 μm. The new species is smaller than A. edaphica in sporangiospores, 3.4–4.4 × 2.1–3.1 μm vs. 3.5–5.5 × 2–3.5 μm. Combining morphological and molecular phylogenetic analyses, the two isolates were classified as a new taxon, allied to A. edaphica.
4. Discussion
The genus Absidia was established nearly 150 years ago. Between 1878 and 2010, the taxonomic status of the genus was changed several times, and the species of the genus were divided according to the optimum growth temperature. In 2007, Kerstin Hoffmann et al. divided Absidia s. l. into Absidia (Absidiaceae s. str., mesothermal type) and Mycocladus (Mycocladiaceae, thermophilic type) [7]. In 2009, A. parricida and A. zychae were separated from the genus Absidia by Hoffmann and Voigt and reassigned to a newly described genus Lentamyces [8]. In 2010, Hoffmann et al. investigated and evaluated Absidia s. l., and finally divided it into three genera, namely Absidia s. s., Lichtheimia and Lentamyces [6]. From 2011 to 2019, the discovery of Absidia stagnated, only six new species have been described (https://www.indexfungorum.org/, accessed on 7 February 2025; [35, 37-40]). Since 2020, taxonomic studies on this genus have been extensively carried out, with a total of 49 new species described (https://www.indexfungorum.org/, accessed on 7 February 2025; [9, 10, 13, 33, 36, 41-50]). Nowadays, it is the most species-rich genus in the family Cunninghamellaceae. Together with the four new species proposed in this study, the world diversity of Absidia reaches 71 recognized species.
Absidia species predominantly inhabit soil environments across tropical, subtropical, and temperate climatic zones. In accordance with this ecological distribution pattern, the four novel species described in this study were successfully isolated from soil samples collected in tropical and subtropical geographic regions of China.
Prior to 2020, the phylogenetic tree of this genus was predominantly reconstructed utilizing the combination of ITS and LSU sequences. In 2021, Hurdeal et al. added SSU and Act sequences for reconstructing the phylogenetic [36]. Subsequently, in 2024, Tao et al. employed a more comprehensive approach, utilizing SSU, ITS, LSU, Act, and TEF1α to reconstruct the phylogenetic tree, yielding results that were largely in line with previous findings [13]. In this study, the phylogenetic history was inferred using ITS-SSU-LSU-Act-TEF1α, and newly isolated strains were grouped into four individual clades with high supports, namely Absidia irregularis sp. nov., A. multiformis sp. nov., A. ovoidospora sp. nov., and A. verticilliformis sp. nov.
Absidia irregularis is related to A. oblongispora with high supports (MLBV = 100, BIPP = 1; Figure 1), while distinguished by bigger apophyses and columellae. Absidia multiformis is most closely related to A. heterospora with high support (BIPP = 0.99; Figure 1). Morphologicaly, A. multiformis has narrower and shorter sporangiophore, smaller sporangia size, different columellae shape and size. Absidia ovoidospora is closely related to A. panacisoli and A. abundans. However, compared with A. panacisoli, A. ovoidospora had wider sporangiophore width and larger sporangium. At the same time, compared with A. abundans, the microscopical measurements of the sporangiophore, sporangium and columella of A. oviodrospora were larger. A. verticilliformis is closely related to A. edaphica. A. verticilliformis differs from A. edaphica by smaller sporangia, bigger columellae and smaller sporangiospores.
In this study, thermal tolerance thresholds of fungal strains were determined using a temperature gradient cultivation technique. Growth characterization revealed distinct maximum growth temperatures for the four Absidia species: A. ovoidospora (30℃), A. irregularis (32℃), A. multiformis (34℃), and A. verticilliformis (34℃). These thermal parameters align with the established physiological profile of Absidia s. s. (mesophilic genus with optimal growth at 25-34℃).
In summary, the molecular phylogenetic and morphological results both support the identification of the four new species. These findings further enhance our understanding of mucoralean biodiversity in Asian tropical and subtropical ecosystems.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/doi/s1, File S1: ITS-SSU-LSU- Act-TEF1a.
Author Contributions
Y.-X.W. took charge of the microscopy, DNA sequencing, data analyses and manuscript draft; Z.-Y.D. and Y.-X.W. collected samples and isolated living cultures; X.-Y.J. and Z.-Y.D. made dry cultures; Z.M. revised the manuscript; X.-Y.L. contributed to new species proposal, manuscript revision and financial support; funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the National Natural Science Foundation of China (Nos. 32170012, 32470004, 32300011), Ji’nan City’s ‘New University 20 Policies’ Initiative for Innovative Research Teams Project (202228028), Innovative Agricultural Application Technology Project of Jinan City (CX202210) and Key Technological Innovation Program of Shandong Province, China (2022CXGC020710).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The sequences were deposited in the GenBank database.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- van Tieghem, P. Troisieme memoire sur les Mucorinees. Ann Sci Nat Bot Ser 1876, 4, 312–398. [Google Scholar]
- Tedersoo, L.; Sánchez-Ramírez, S.; Kõljalg, U.; Bahram, M.; Döring, M.; Schigel, D.; May, T.; Ryberg, M.; Abarenkov, K. High-level classification of the Fungi and a tool for evolutionary ecological analyses. Fungal Diversity 2018, 90, 135–159. [Google Scholar] [CrossRef]
- Arx, J. A. v. On Mucoraceae s. str. and other families of the Mucorales. Sydowia 1982, 35, 10–26. [Google Scholar]
- Benny, G.; Humber, R.; Morton, J.: Zygomycota: Zygomycetes. 2001; 113–146.
- Kirk, P.; Cannon, P.; Minter, D.; Stalpers, J. Ainsworth and Bisby's Dictionary of the Fungi (10th edition). Reference Reviews 2009, 23, 42–42. [Google Scholar]
- Hoffmann, K. Identification of the genus Absidia (Mucorales, Zygomycetes): A comprehensive taxonomic revision. 2010; 439–460.
- Hoffmann, K.; Discher, S.; Voigt, K. Revision of the genus Absidia (Mucorales, Zygomycetes) based on physiological, phylogenetic, and morphological characters; thermotolerant Absidia spp. form a coherent group, Mycocladiaceae fam. nov. Mycological Research 2007, 111, 1169–1183. [Google Scholar] [CrossRef]
- Hoffmann, K.; Voigt, K. Absidia parricida plays a dominant role in biotrophic fusion parasitism among mucoralean fungi (Zygomycetes): Lentamyces, a new genus for A. parricida and A. zychae. Plant Biology 2009, 11, 537–554. [Google Scholar] [CrossRef]
- Zhao, H.; Nie, Y.; Tongkai, Z.; Wang, Y.-J.; Wang, M.; Dai, Y.-C.; Liu, X.-Y. Species diversity and ecological habitat of Absidia (Cunninghamellaceae, Mucorales) with emphasis on five new species from forest and grassland soil in China. Journal of Fungi 2022, 8, 471. [Google Scholar] [CrossRef]
- Zhao, H.; Nie, Y.; Zong, T.; Dai, Y.; Liu, X. Three new species of Absidia (Mucoromycota) from China based on phylogeny, morphology and physiology. Diversity 2022, 14, 132. [Google Scholar] [CrossRef]
- Braga, A. R.; Nunes, M. C.; Raymundo, A. The experimental development of emulsions enriched and stabilized by recovering matter from spirulina biomass: Valorization of residue into a sustainable protein source. Molecules. [CrossRef] [PubMed]
- Liu, X.; Qiao, L.; Xie, D.; Zhang, Y.; Zou, J.; Chen, X.; Dai, J. Microbial transformation of ginsenoside-Rg1 by Absidia coerulea and the reversal activity of the metabolites towards multi-drug resistant tumor cells. Fitoterapia 2011, 82, 1313–1317. [Google Scholar] [CrossRef]
- Tao, M.-F.; Ding, Z.-Y.; Wang, Y.-X.; Zhang, Z.-X.; Zhao, H.; Meng, Z.; Liu, X.-Y. Unveiling species diversity within early-diverging fungi from China II: Three new species of Absidia (Cunninghamellaceae, Mucoromycota) from Hainan Province. MycoKeys 2024, 110, 255–272. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-X.; Zhao, H.; Jiang, Y.; Liu, X.-Y.; Tao, M.-F.; Liu, X.-Y. Unveiling species diversity within early-diverging fungi from China III: Six new species and a new record of Gongronella (Cunninghamellaceae, Mucoromycota). MycoKeys 2024, 110, 287–317. [Google Scholar] [CrossRef]
- Zhao, H.; Nie, Y.; Huang, B.; Liu, X.-Y. Unveiling species diversity within early-diverging fungi from China I: three new species of Backusella (Backusellaceae, Mucoromycota). MycoKeys 2024, 109, 285–304. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.-Y.; Ji, X.-Y.; Tao, M.-F.; Jiang, Y.; Liu, W.-X.; Wang, Y.-X.; Meng, Z.; Liu, X.-Y. Unveiling species diversity within early-diverging fungi from China IV: Four new species of Absidia (Cunninghamellaceae, Mucoromycota). MycoKeys (under review).
- Ji, X.-Y.; Ding, Z.-Y.; Nie, Y.; Zhao, H.; Wang, S.; Huang, B.; Liu, X.-Y. Unveiling species diversity within early-diverging fungi from China V: Five new species of Absidia (Cunninghamellaceae, Mucoromycota). MycoKeys (under review).
- Li, W.; Wei, Y.; Zou, Y.; Liu, P.; Li, Z.; Gontcharov, A.; Stephenson, S.; Wang, Q.; Zhang, S.; li, y. Dictyostelid cellular slime molds from the russian far east. Protist 2020, 171, 125756. [Google Scholar] [CrossRef]
- Zou, Y.; Hou, J.; Guo, S.; Li, C.; Li, Z.; Stephenson, S. L.; Pavlov, I. N.; Liu, P.; Li, Y. Diversity of dictyostelid cellular slime molds, including two species new to science, in forest soils of Changbai Mountain, China. Microbiology Spectrum 2022, 10, e0240222. [Google Scholar] [CrossRef]
- Corry, J. E. L., Curtis, G. D. W., Baird, R. M., Eds.; Elsevier, Rose Bengal Chloramphenicol (RBC) agar. In Progress in Industrial Microbiology 1995, 34, 431–433.
- Wang, Y.-X.; Zhao, H.; Ding, Z.-Y.; Ji, X.-Y.; Zhang, Z.; Wang, S.; Zhang, X.; Liu, X.-Y. Three new species of Gongronella (Cunninghamellaceae, Mucorales) from soil in Hainan, China based on morphology and molecular phylogeny. Journal of Fungi 2023, 9, 1182. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, R.; Liu, S.; Mu, T.; Zhang, X.; Xia, J. Morphological and phylogenetic analyses reveal two new species of Sporocadaceae from Hainan, China. MycoKeys 2022, 88, 171–192. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Molecular Biology and Evolution 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Voigt, K.; Wostemeyer, J. Reliable amplification of actin genes facilitates deep-level phylogeny. Microbiological research 2000, 155, 179–195. [Google Scholar] [CrossRef]
- White, T.; Bruns, T.; Lee, S.; Taylor, J.; Innis, M.; Gelfand, D.; Sninsky, J.: Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. 1990; Vol. 31; pp 315–322.
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. Journal of Bacteriology 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L. M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- O'Donnell, K.; Lutzoni, F.; Ward, T.; Benny, G. Evolutionary relationships among mucoralean fungi (Zygomycota): Evidence for family polyphyly on a large scale. Mycologia 2001, 93, 286–296. [Google Scholar] [CrossRef]
- Nylander, J. MrModeltest V2. Program distributed by the author. Bioinformatics 2004, 24, 581–583. [Google Scholar] [CrossRef]
- Huelsenbeck, J. P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Mark, P.; Ayres, D.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.; Huelsenbeck, J. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Systematic biology 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Nie, Y.; Zong, T.-K.; Wang, K.; Lv, M.-L.; Cui, Y.-J.; Tohtirjap, A.; Chen, J.-J.; Zhao, C.-L.; Wu, F.; Cui, B.-K.; Yuan, Y.; Dai, Y.-C.; Liu, X.-Y. Species diversity, updated classification and divergence times of the phylum Mucoromycota. Fungal Diversity 2023, 123, 49–157. [Google Scholar] [CrossRef]
- Hesseltine, C. W. J. J. E. The genus Absidia: Gongronella and cylindrical-spored species of Absidia. Mycologia 1964, 4, 568–601. [Google Scholar] [CrossRef]
- Zhang, T.-Y.; Yu, Y.; Zhu, H.; Yang, S.-Z.; Yang, T.-M.; Zhang, M.-y.; Zhang, Y.-X. Absidia panacisoli sp. nov., isolated from rhizosphere of Panax notoginseng. International Journal of Systematic and Evolutionary Microbiology 2018, 68, 2468–2472. [Google Scholar] [CrossRef]
- Hurdeal, V. G.; Gentekaki, E.; Lee, H. B.; Jeewon, R.; Hyde, K. D.; Tibpromma, S.; Mortimer, P. E.; Xu, J.-c. Mucoralean fungi in Thailand: Novel species of Absidia from tropical forest soil. Cryptogamie, Mycologie 2021, 42, 39–61. [Google Scholar] [CrossRef]
- Ariyawansa, H.; Hyde, K.; Jayasiri, S.; Buyck, B.; Kandawatte, T.; Dai, D.-Q.; Dai, Y.; Daranagama, D.; Jayawardena, R.; Lücking, R.; Ghobad-Nejhad, M.; Niskanen, T.; Thambugala, K.; Voigt, K.; Zhao, R.-L.; Li, G.-J.; Doilom, M.; Boonmee, S.; Yang, Z.; Chen, X.-H. Fungal Diversity Notes 111–252—taxonomic and phylogenetic contributions to fungal taxa. Fungal diversity 2015, 75, 27–274. [Google Scholar] [CrossRef]
- Li, G. J.; Hyde, K. D.; Zhao, R. L.; Hongsanan, S.; Abdel-Aziz, F. A.; Abdel-Wahab, M. A.; Alvarado, P.; Alves-Silva, G.; Ammirati, J. F.; Ariyawansa, H. A.; Baghela, A.; Bahkali, A. H.; Beug, M.; Bhat, D. J.; Bojantchev, D.; Boonpratuang, T.; Bulgakov, T. S.; Camporesi, E.; Boro, M. C.; Ceska, O.; Chakraborty, D.; Chen, J. J.; Chethana, K. W. T.; Chomnunti, P.; Consiglio, G.; Cui, B. K.; Dai, D. Q.; Dai, Y. C.; Daranagama, D. A.; Das, K.; Dayarathne, M. C.; De Crop, E.; De Oliveira, R. J. V.; de Souza, C. A. F.; de Souza, J. I.; Dentinger, B. T. M.; Dissanayake, A. J.; Doilom, M.; Drechsler-Santos, E. R.; Ghobad-Nejhad, M.; Gilmore, S. P.; Góes-Neto, A.; Gorczak, M.; Haitjema, C. H.; Hapuarachchi, K. K.; Hashimoto, A.; He, M. Q.; Henske, J. K.; Hirayama, K.; Iribarren, M. J.; Jayasiri, S. C.; Jayawardena, R. S.; Jeon, S. J.; Jerônimo, G. H.; Jesus, A. L.; Jones, E. B. G.; Kang, J. C.; Karunarathna, S. C.; Kirk, P. M.; Konta, S.; Kuhnert, E.; Langer, E.; Lee, H. S.; Lee, H. B.; Li, W. J.; Li, X. H.; Liimatainen, K.; Lima, D. X.; Lin, C. G.; Liu, J. K.; Liu, X. Z.; Liu, Z. Y.; Luangsa-ard, J. J.; Lücking, R.; Lumbsch, H. T.; Lumyong, S.; Leaño, E. M.; Marano, A. V.; Matsumura, M.; McKenzie, E. H. C.; Mongkolsamrit, S.; Mortimer, P. E.; Nguyen, T. T. T.; Niskanen, T.; Norphanphoun, C.; O’Malley, M. A.; Parnmen, S.; Pawłowska, J.; Perera, R. H.; Phookamsak, R.; Phukhamsakda, C.; Pires-Zottarelli, C. L. A.; Raspé, O.; Reck, M. A.; Rocha, S. C. O.; de Santiago, A. L. C. M. A.; Senanayake, I. C.; Setti, L.; Shang, Q. J.; Singh, S. K. Fungal diversity notes 253–366: taxonomic and phylogenetic contributions to fungal taxa. Fungal Diversity 2016, 78, 1–237. [Google Scholar] [CrossRef]
- Crous, P.; Luangsa-Ard, J.; Wingfield, M. J.; Carnegie, A.; Hernández-Restrepo, M.; Lombard, L.; Roux, J.; Barreto, R.; Baseia, I.; Cano, J.; Martín, M.; Morozova, O.; Stchigel, A. M.; Summerell, B.; Brandrud, T. E.; Dima, B.; García, D.; Giraldo López, A.; Guarro, J.; Groenewald, J. Z. Fungal Planet description sheets- 785–867. Persoonia 2018, 41, 238–417. [Google Scholar] [CrossRef]
- Wanasinghe, D. N.; Phukhamsakda, C.; Hyde, K. D.; Jeewon, R.; Lee, H. B.; Gareth Jones, E. B.; Tibpromma, S.; Tennakoon, D. S.; Dissanayake, A. J.; Jayasiri, S. C.; Gafforov, Y.; Camporesi, E.; Bulgakov, T. S.; Ekanayake, A. H.; Perera, R. H.; Samarakoon, M. C.; Goonasekara, I. D.; Mapook, A.; Li, W.-J.; Senanayake, I. C.; Li, J.; Norphanphoun, C.; Doilom, M.; Bahkali, A. H.; Xu, J.; Mortimer, P. E.; Tibell, L.; Tibell, S.; Karunarathna, S. C. Fungal diversity notes 709–839: taxonomic and phylogenetic contributions to fungal taxa with an emphasis on fungi on Rosaceae. Fungal Diversity 2018, 89, 1–236. [Google Scholar] [CrossRef]
- Lima, D.; Cordeiro, T.; Fragoso de Souza, C.; Oliveira, R.; Lee, H.; Souza-Motta, C.; Santiago, A. Morphological and molecular evidence for two new species of Absidia from Neotropic soil. Phytotaxa 2020, 446, 61–71. [Google Scholar] [CrossRef]
- Rafhaella, T.; Cordeiro, L.; Thuong, T.; Nguyen, T.; Lima, D.; Silva, S.; Lima, C.; D'Arc, J.; Leitão, A.; Melo, L.; Gurgel, S.; Lee, H.; Luiz, A.; Monteiro, C.; Santiago, A. Two new species of the industrially relevant genus Absidia (Mucorales) from soil of the Brazilian Atlantic Forest. Acta Botanica Brasilica 2020, 34. [Google Scholar]
- Arina, C.; Lima, L.; Diogo, X.; Lima, T.; Rafhaella, L.; Cordeiro, T.; Lee, H.; Thuong, T.; Nguyen, T.; Melo, L.; Gurgel, S.; Santiago, A. Absidia bonitoensis (Mucorales, Mucoromycota),a new species isolated from the soil of an uplandAtlantic forest in Northeastern Brazil. Nova Hedwigia 2021, 112, 241–251. [Google Scholar]
- LeitÃO, J.; Cordeiro, T.; Nguyen, T.; Lee, H.; Gurgel, L.; Santiago, A. Absidia aguabelensis sp. nov.: A new mucoralean fungi isolated from a semiarid region in Brazil. Phytotaxa 2021, 516, 83–91. [Google Scholar] [CrossRef]
- Urquhart, A.; Idnurm, A. Absidia healeyae: a new species of Absidia (Mucorales) isolated from Victoria, Australia. Mycoscience 2021, 62, 331–335. [Google Scholar] [CrossRef]
- Zhao, H.; Zhu, J.; Tongkai, Z.; Liu, X.-L.; Ren, L.-Y.; Lin, Q.; Qiao, M.; Nie, Y.; Zhang, Z.-D.; Liu, X.-Y. Two New Species in the Family Cunninghamellaceae from China. Mycobiology 2021, 49, 1–9. [Google Scholar] [CrossRef]
- Luo, C.; Chen, F.; Phookamsak, R.; Sun, F.; Xu, J.; Jiang, H. Polyphasic taxonomic study of Absidia menglianensis sp. nov. (Cunninghamellaceae, Mucorales) isolated from an avocado plantation in Yunnan, China. Studies in Fungi 2024, 9, e018. [Google Scholar] [CrossRef]
- Nguyen, T. T. T.; Santiago, A.; Hallsworth, J. E.; Cordeiro, T.; Voigt, K.; Kirk, P.; Crous, P. W.; Júnior, M. A. M.; Elsztein, C.; Lee, H. New Mucorales from opposite ends of the world. Studies in Mycology 2024, 109, 273–321. [Google Scholar] [CrossRef] [PubMed]
- Htet, Y. M.; Yasanthika, W. A. E.; Hurdeal, V. G.; Gomes De Farias, A. R. Absidia thailandica sp. nov., an addition to the diversity of soil fungi from Thailand. Phytotaxa 2024, 646, 157–168. [Google Scholar] [CrossRef]
- Hurdeal, V. G.; Jones, E. B. G.; Gentekaki, E. Absidia zygospora (Mucoromycetes), a new species from Nan Province, Thailand.
Figure 1.
The Beyesian phylogenetic tree of Absidia based on ITS-SSU-LSU-Act-TEF1α sequences, with Cunninghamella blakesleeana CBS 782.68 as outgroup. The maximum likelihood bootstrap value (MLBV) ≥ 75, and the Bayesian inference posterior probability (BIPP) ≥ 0.85 are shown at the first and second positions and separated by a slash "/" on relevant nodes. The ex-types or ex-holotypes are in bold and marked with an asterisk "*", and strains involved in this study are in red. Branches shortened to fit the page are represented by double slashes “//” and folds “×”. The scale at the bottom centre indicates 0.2 substitutions per site.
Figure 1.
The Beyesian phylogenetic tree of Absidia based on ITS-SSU-LSU-Act-TEF1α sequences, with Cunninghamella blakesleeana CBS 782.68 as outgroup. The maximum likelihood bootstrap value (MLBV) ≥ 75, and the Bayesian inference posterior probability (BIPP) ≥ 0.85 are shown at the first and second positions and separated by a slash "/" on relevant nodes. The ex-types or ex-holotypes are in bold and marked with an asterisk "*", and strains involved in this study are in red. Branches shortened to fit the page are represented by double slashes “//” and folds “×”. The scale at the bottom centre indicates 0.2 substitutions per site.
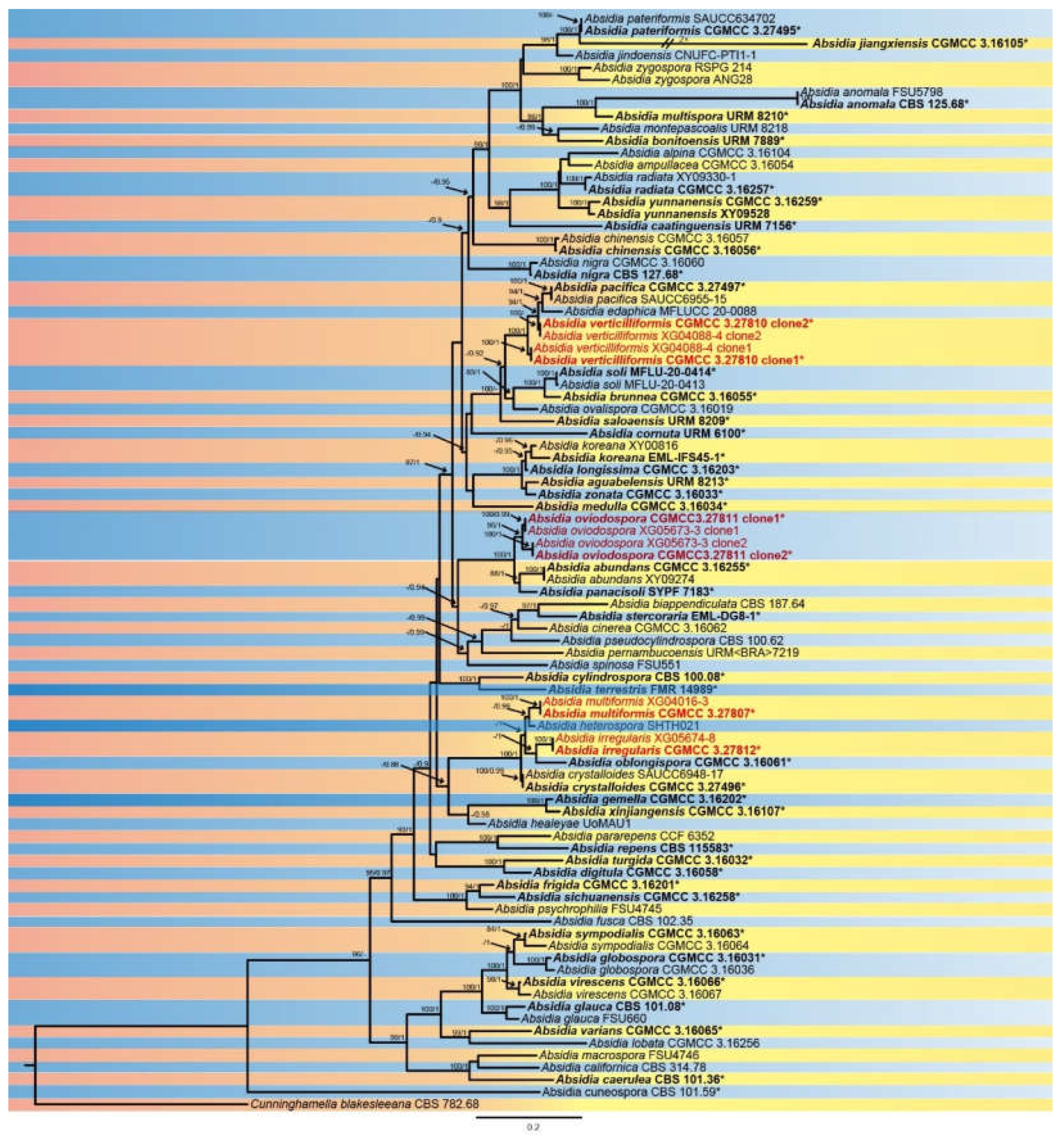
Figure 2.
Absidia irregularis ex-holotype CGMCC 3.27812. a, b, Colonies on PDA (a, obverse; b, reverse); c, d, An unbranched sporangiophore with a sporangium; e, f, Columellae, collars, sporangiospores and apophyses; g, Branched sporangiophores with sporangia; h, Branched sporangiophores with columellae, collars, sporangia and apophyses; i, j, Rhizoids; k, Sporangiospores; Bars: c–k 10 μm.
Figure 2.
Absidia irregularis ex-holotype CGMCC 3.27812. a, b, Colonies on PDA (a, obverse; b, reverse); c, d, An unbranched sporangiophore with a sporangium; e, f, Columellae, collars, sporangiospores and apophyses; g, Branched sporangiophores with sporangia; h, Branched sporangiophores with columellae, collars, sporangia and apophyses; i, j, Rhizoids; k, Sporangiospores; Bars: c–k 10 μm.
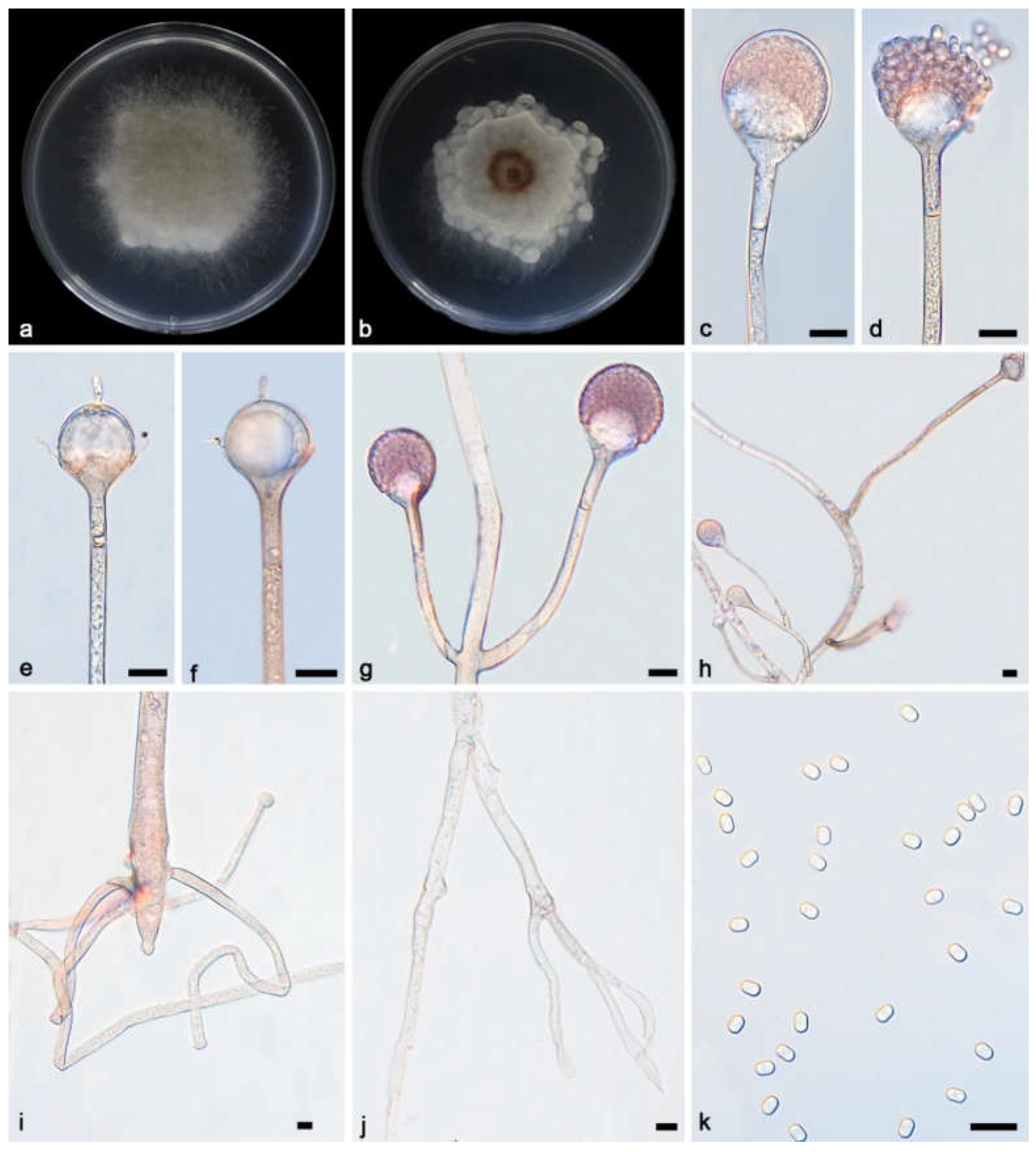
Figure 3.
Absidia multiformis ex-holotype CGMCC 3.27807. a, b, Colonies on PDA (a, obverse; b, reverse); c, d, An unbranched sporangiophore with a sporangium; e, f, Columellae, collars, sporangiospores and apophyses; g, Branched sporangiophores with columellae, collars, sporangium and apophyses; h, i, Rhizoids; j, k, Sporangiospores; k, m, Zygospores; Bars: c–m 10 μm.
Figure 3.
Absidia multiformis ex-holotype CGMCC 3.27807. a, b, Colonies on PDA (a, obverse; b, reverse); c, d, An unbranched sporangiophore with a sporangium; e, f, Columellae, collars, sporangiospores and apophyses; g, Branched sporangiophores with columellae, collars, sporangium and apophyses; h, i, Rhizoids; j, k, Sporangiospores; k, m, Zygospores; Bars: c–m 10 μm.
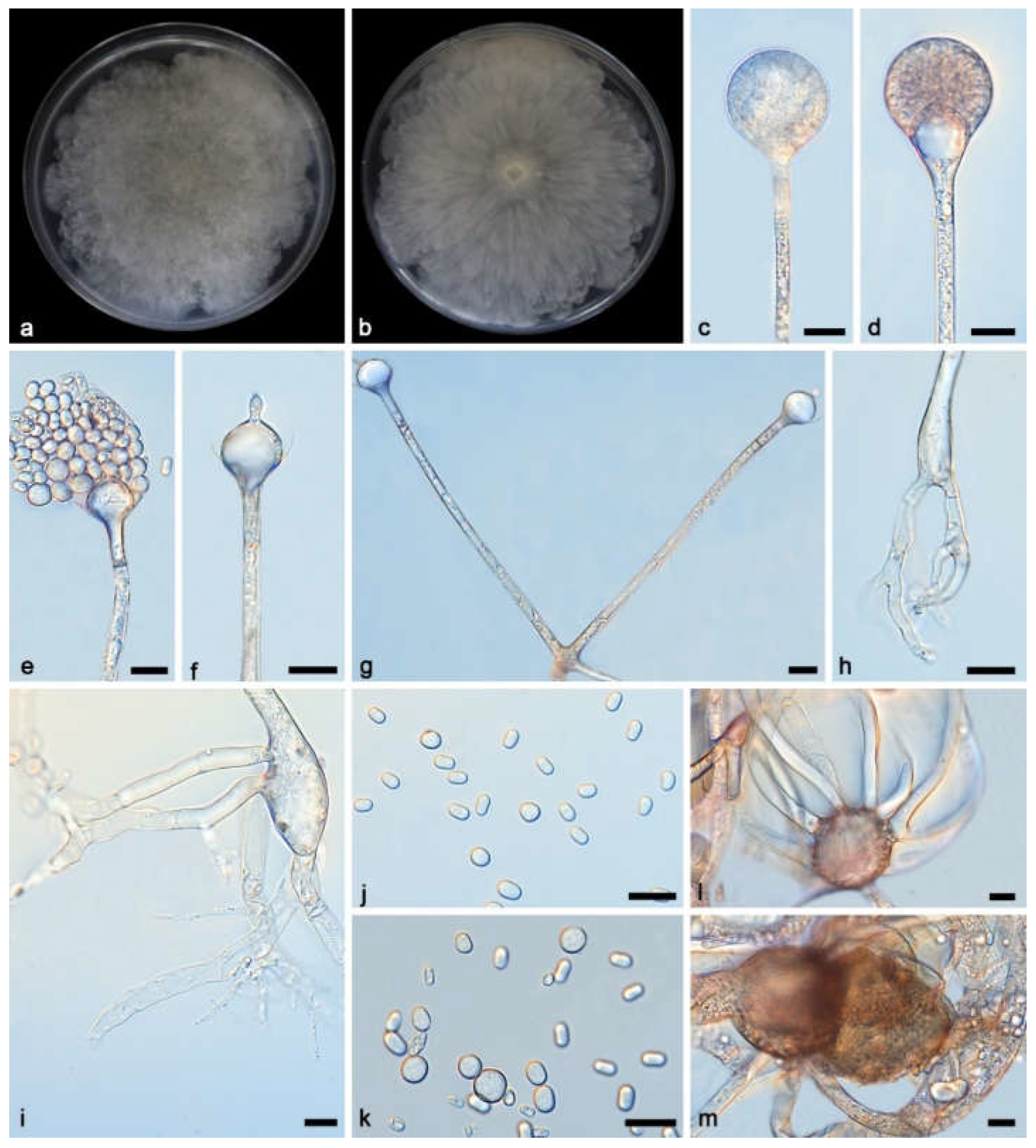
Figure 4.
Absidia oviodospora ex-holotype CGMCC 3.27811. a, b, Colonies on PDA (a, obverse; b, reverse); c–e, An unbranched sporangiophore with a sporangium; f, Columellae, collars, sporangiospores and apophyses; g, Unbranched sporangiophores with swelling and sporangium; h, Branched sporangiophores with columellae, collars and apophyses; i, Branched sporangiophores with sporangia; j, Rhizoids; k–m, Giant cells; n, Sporangiospores; Bars: c–n 10 μm.
Figure 4.
Absidia oviodospora ex-holotype CGMCC 3.27811. a, b, Colonies on PDA (a, obverse; b, reverse); c–e, An unbranched sporangiophore with a sporangium; f, Columellae, collars, sporangiospores and apophyses; g, Unbranched sporangiophores with swelling and sporangium; h, Branched sporangiophores with columellae, collars and apophyses; i, Branched sporangiophores with sporangia; j, Rhizoids; k–m, Giant cells; n, Sporangiospores; Bars: c–n 10 μm.
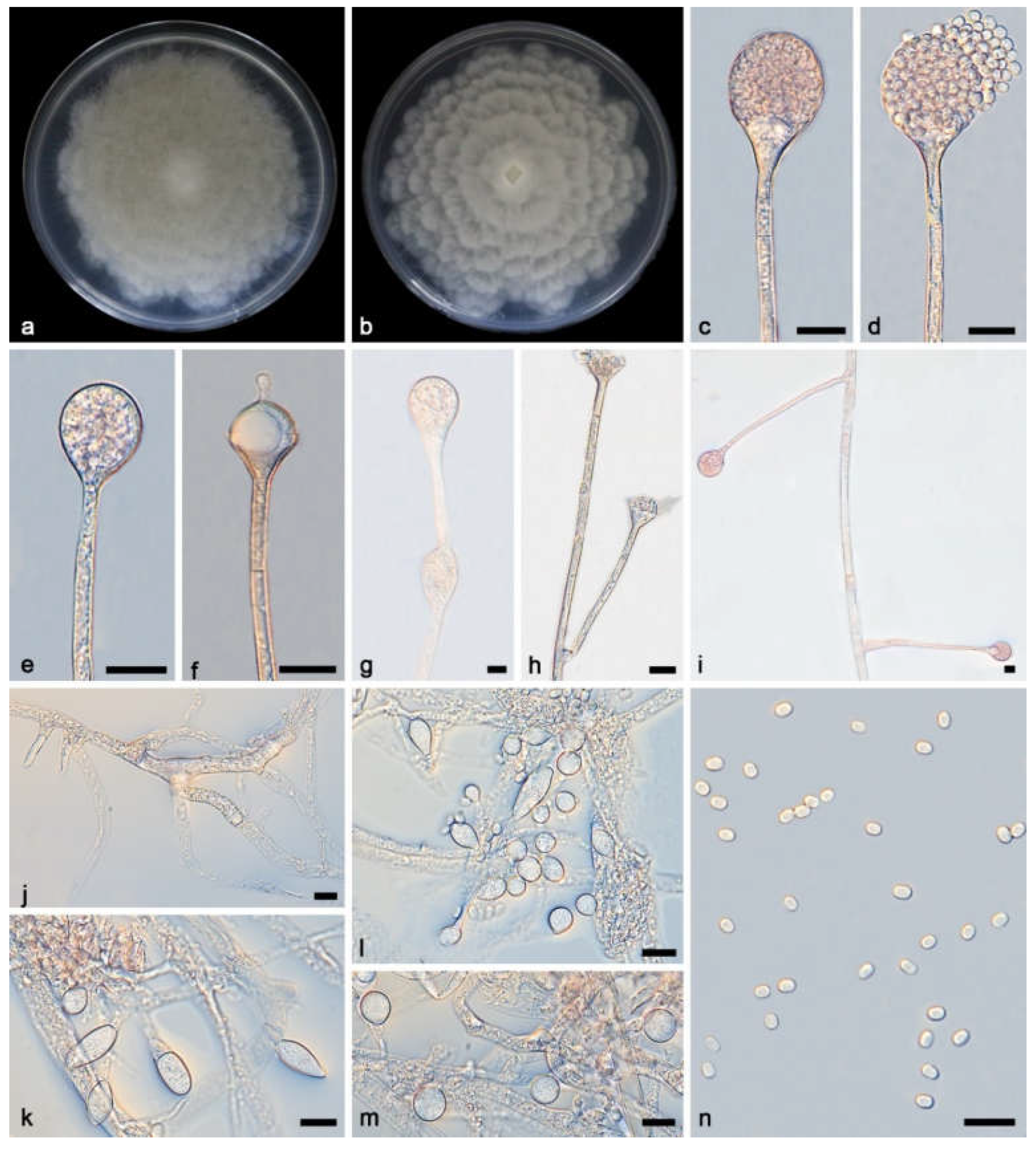
Figure 5.
Absidia verticilliformis ex-holotype CGMCC 3.27810. a, b, Colonies on PDA (a, obverse; b, reverse); c–e, An unbranched sporangiophore with a sporangium; f, Columellae, collars, sporangiospores and apophyses; g, h, Branched sporangiophores with columellae, collars, sporangiospores and apophyses; i, j, Rhizoids; k, Sporangiospores; Bars: c–k 10 μm.
Figure 5.
Absidia verticilliformis ex-holotype CGMCC 3.27810. a, b, Colonies on PDA (a, obverse; b, reverse); c–e, An unbranched sporangiophore with a sporangium; f, Columellae, collars, sporangiospores and apophyses; g, h, Branched sporangiophores with columellae, collars, sporangiospores and apophyses; i, j, Rhizoids; k, Sporangiospores; Bars: c–k 10 μm.
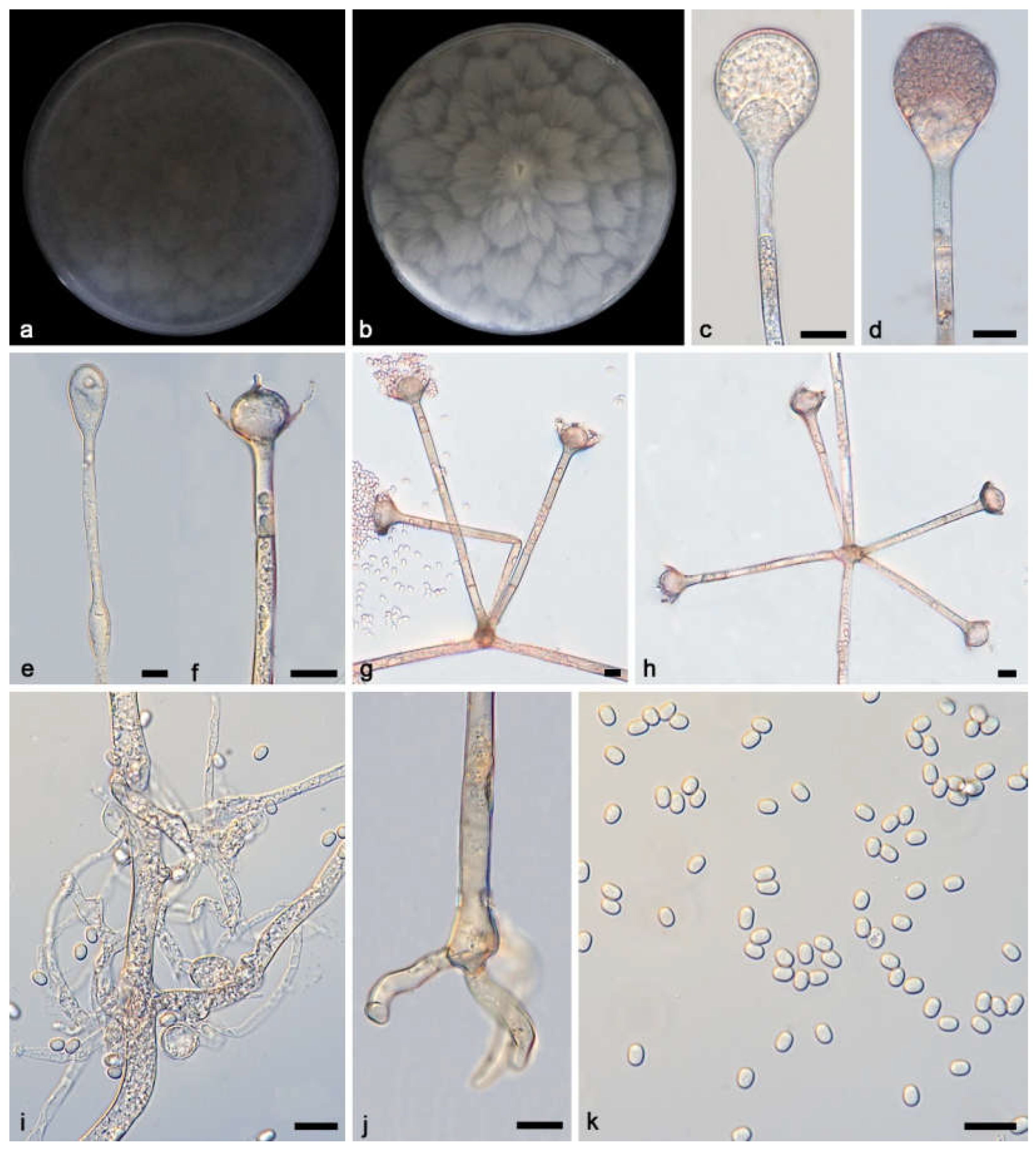

Table 1.
Molecular loci, PCR primers and programmes used in this study.
| Loci | PCR primers | Sequence (5’ – 3’) | PCR cycles | References |
|---|---|---|---|---|
| Act | ACT-512F | ATG TGC AAG GCC GGT TTC GC | 95℃ 3 min; (95℃ 1 min, 55℃ 1 min, 72℃ 1 min) × 30 cycles; 72℃ 10 min | [25] |
| ACT-783R | TAC GAG TCC TTC TGG CCC AT | |||
| ITS | ITS5 | GGA AGT AAA AGT CGT AAC AAG G | 95℃ 5 min; (95℃ 30 s, 55℃ 30 s, 72℃ 1 min) × 35 cycles; 72℃ 10 min | [26] |
| ITS4 | TCC TCC GCT TAT TGA TAT GC | |||
| LSU | LR0R | GTA CCC GCT GAA CTT AAG C | 95℃ 5 min; (95℃ 50 s, 47℃ 30 s, 72℃ 1.5 min) × 35 cycles; 72℃ 10 min | [27] |
| LR7 | TAC TAC CAC CAA GAT CT | |||
| SSU | NS1 | GTA GTC ATA TGC TTG TCT C C | 95 °C 5 min; (94 °C 60 s, 54 °C 50 s, 72 °C 60 s) × 37 cycles; 72 °C 10 min | [26] |
| NS4 | CTT CCG TCA ATT CCT TTA AG | |||
| TEF1α | EF1-728F | CAT CGA GAA GTT CGA GAA GG | 95℃ 5 min; (95℃ 30 s, 55℃ 60 s, 72℃ 1 min) × 30 cycles; 72℃ 10 min | [28] |
| EF2 | GGA RGT ACC AGT SAT CAT GTT |
Table 2.
Information of strains used in this study.
| Species | Strains | GenBank accession numbers | ||||
| ITS | LSU | TEF1α | Act | SSU | ||
| Absidia abundans | CGMCC 3.16255* | NR_182590 | ON074683 | n.a. | n.a. | n.a. |
| A. abundans | XY09274 | ON074696 | ON074682 | n.a. | n.a. | n.a. |
| A. aguabelensis | URM 8213* | NR_189383 | NG_241934 | n.a. | n.a. | n.a. |
| A. alpina | CGMCC 3.16104 | OL678133 | n.a. | n.a. | n.a. | n.a. |
| A. ampullacea | CGMCC 3.16054 | MZ354138 | MZ350132 | n.a. | n.a. | n.a. |
| A. anomala | CBS 125.68* | MH859085 | MH870799 | n.a. | n.a. | n.a. |
| A. anomala | FSU5798 | EF030523 | n.a. | n.a. | EF030535 | n.a. |
| A. biappendiculata | CBS 187.64 | MZ354153 | MZ350147 | MZ357420 | MZ357438 | |
| A. bonitoensis | URM 7889* | MN977786 | MN977805 | n.a. | n.a. | n.a. |
| A. brunnea | CGMCC 3.16055* | MZ354139 | MZ350133 | MZ357403 | MZ357421 | n.a. |
| A. caatinguensis | URM 7156* | NR_154704 | NG_058582 | n.a. | n.a. | n.a. |
| A. caerulea | CBS101.36* | MH855718 | MH867230 | n.a. | n.a. | n.a. |
| A. californica | CBS 314.78 | JN205816 | MH872902 | n.a. | n.a. | n.a. |
| A. chinensis | CGMCC 3.16057 | MZ354141 | MZ350135 | n.a. | MZ357422 | n.a. |
| A. chinensis | CGMCC 3.16056* | MZ354140 | MZ350134 | n.a. | n.a. | n.a. |
| A. cinerea | CGMCC 3.16062 | MZ354146 | MZ350140 | MZ357407 | MZ357427 | n.a. |
| A. cornuta | URM 6100* | NR_172976 | MN625255 | n.a. | n.a. | n.a. |
| A. crystalloides | CGMCC3.27496* | PP377803 | PP373736 | PP790574 | PP790582 | PP779723 |
| A. crystalloides | SAUCC693201 | PP377804 | PP373737 | PP790573 | PP790581 | PP779722 |
| A. cuneospora | CBS 101.59* | MH857828 | MH869361 | n.a. | n.a. | n.a. |
| A. cuneospora | FSU5890 | EF030524 | n.a. | n.a. | EF030533 | n.a. |
| A. cylindrospora | CBS 100.08 | JN205822 | JN206588 | n.a. | n.a. | n.a. |
| A. digitula | CGMCC 3.16058* | MZ354142 | MZ350136 | MZ357404 | MZ357423 | n.a. |
| A. edaphica | MFLUCC 20-0088 | NR_172305 | NG_075367 | n.a. | MT410739 | NG_074951 |
| A. frigida | CGMCC 3.16201* | NR_182565 | OM030223 | n.a. | n.a. | n.a. |
| A. fusca | CBS 102.35* | NR_103625 | NG_058552 | n.a. | n.a. | n.a. |
| A. gemella | CGMCC 3.16202* | OM108488 | OM030224 | n.a. | n.a. | n.a. |
| A. glauca | CBS 101.08* | MH854573 | MH866105 | n.a. | n.a. | n.a. |
| A. glauca | FSU660 | AY944879 | EU736302 | EU736248 | EU736225 | EU736275 |
| A. globospora | CGMCC 3.16031* | NR_189829 | MW671544 | MZ357412 | MZ357431 | n.a. |
| A. globospora | CGMCC 3.16036 | MW671539 | MW671546 | MZ357414 | MZ357433 | n.a. |
| A. healeyae | UoMAU1 | n.a. | MT436027 | n.a. | n.a. | n.a. |
| A. heterospora | SHTH021 | JN942683 | JN982936 | n.a. | n.a. | JQ004928 |
| A.irregularis | CGMCC 3.27812* | PQ306325 | PQ289020 | PV126019 | PQ807209 | PQ799254 |
| A.irregularis | XG05674-7 | PQ306326 | PQ289021 | PV126020 | PQ807210 | PQ799255 |
| A. jiangxiensis | CGMCC 3.16105* | OL678134 | PP780377 | n.a. | n.a. | n.a. |
| A. jindoensis | CNUFC-PTI1-1 | MF926622 | MF926616 | MF926513 | MF926510 | MF926626 |
| A. koreana | EML-IFS45-1* | KR030062 | KR030056 | KR030060 | KR030058 | KT321298 |
| A. koreana | XY00816 | OL620083 | ON123771 | n.a. | n.a. | n.a. |
| A. lobata | CGMCC 3.16256 | ON074690 | ON074679 | n.a. | n.a. | n.a. |
| A. longissima | CGMCC 3.16203* | NR_182566 | OM030225 | n.a. | n.a. | n.a. |
| A. macrospora | FSU4746 | AY944882 | EU736303 | EU736249 | AY944760 | EU736276 |
| A. medulla | CGMCC 3.16034 | NR_189832 | MW671549 | MZ357417 | MZ357436 | n.a. |
| A. montepascoalis | URM 8218 | NR_172995 | n.a. | n.a. | n.a. | n.a. |
| A.multiformis | CGMCC 3.27807* | PQ306319 | PQ803168 | PV126021 | PQ807203 | PQ799260 |
| A.multiformis | XG04016-3 | PQ306320 | PQ803169 | PV126022 | PQ807204 | PQ799261 |
| A. multispora | URM 8210* | MN953780 | MN953782 | n.a. | n.a. | n.a. |
| A. nigra | CBS 127.68* | NR_173068 | MZ350146 | MZ357419 | MZ357437 | n.a. |
| A. nigra | CGMCC 3.16060 | MZ354144 | MZ350138 | MZ357406 | MZ357425 | n.a. |
| A. oblongispora | CGMCC 3.16061 | MZ354145 | MZ350139 | n.a. | MZ357426 | n.a. |
| A. ovalispora | CGMCC 3.16019 | NR_176748 | MW264131 | n.a. | n.a. | n.a. |
| A.ovoidospora | CGMCC 3.27811 clone1* | PQ306327 | PQ803164 | PV126015 | PQ807207 | PQ799256 |
| A. ovoidospora | CGMCC 3.27811 clone2* | PV069753 | PQ803165 | PV126017 | PV126023 | PQ799257 |
| A. ovoidospora | XG05673-3 clone1 | PQ306328 | PQ803166 | PV126016 | PQ807208 | PQ799258 |
| A. ovoidospora | XG05673-3 clone2 | PV069754 | PQ803167 | PV126018 | PV126024 | PQ799259 |
| A. pacifica | CGMCC3.27497* | PP377802 | PP373735 | PP839793 | PP790579 | PP779720 |
| A. pacifica | SAUCC413601 | PP377801 | PP373734 | PP839794 | PP790580 | PP779721 |
| A. panacisoli | SYPF 7183* | MF522181 | MF522180 | MF624251 | n.a. | MF522179 |
| A. pararepens | CCF 6352 | MT193669 | MT192308 | n.a. | n.a. | n.a. |
| A. pateriformis | CGMCC3.27495* | PP377805 | PP373738 | PP790575 | PP790583 | PP779724 |
| A. pateriformis | SAUCC634702 | PP377806 | PP373739 | PP790576 | PP790584 | PP779725 |
| A. pernambucoensis | URM<BRA>7219 | MN635568 | MN635569 | n.a. | n.a. | n.a. |
| A. pseudocylindrospora | EML-FSDY6-2 | KU923817 | KU923814 | n.a. | KU923815 | KU923819 |
| A. pseudocylindrospora | CBS 100.62* | NR_145276 | MH869688 | n.a. | n.a. | n.a. |
| A. psychrophilia | FSU4745 | AY944874 | EU736306 | EU736252 | AY944762 | EU736279 |
| A. radiata | CGMCC 3.16257 | ON074698 | ON074684 | n.a. | n.a. | n.a. |
| A. radiata | XY09330-1 | ON074699 | ON074685 | n.a. | n.a. | n.a. |
| A. repens | CBS 115583* | NR_103624 | NG_058551 | n.a. | n.a. | n.a. |
| A. saloaensis | URM 8209* | MN953781 | MN953783 | n.a. | n.a. | n.a. |
| A. sichuanensis | CGMCC 3.16258* | NR_182589 | ON074688 | n.a. | n.a. | n.a. |
| A. soli | MFLU-20-0414* | MT396373 | MT393988 | n.a. | n.a. | MT394049 |
| A. soli | MFLU 20-0413 | MT396371 | MT393985 | n.a. | n.a. | MT394046 |
| A. spinosa | FSU551 | AY944887 | EU736307 | EU736253 | EU736227 | EU736280 |
| A. stercoraria | EML-DG8-1* | KU168828 | KT921998 | KT922002 | KT922000 | NG_065640 |
| A. sympodialis | CGMCC 3.16063* | MZ354147 | MZ350141 | n.a. | n.a. | n.a. |
| A. sympodialis | CGMCC 3.16064 | MZ354148 | MZ350142 | MZ357408 | n.a. | n.a. |
| A. terrestris | FMR 14989* | LT795003 | LT795005 | n.a. | n.a. | n.a. |
| A. turgida | CGMCC 3.16032* | NR_189830 | NG_241931 | MZ357415 | MZ357434 | n.a. |
| A. varians | CGMCC 3.16065* | MZ354149 | MZ350143 | MZ357409 | MZ357428 | n.a. |
| A.verticilliformis | CGMCC 3.27810 clone1* | PQ306315 | PQ803170 | PV126011 | PQ807205 | PQ799262 |
| A.verticilliformis | CGMCC 3.27810 clone2* | PV069755 | PQ803171 | PV126013 | PV126025 | PQ799263 |
| A.verticilliformis | XG04088-4 clone1 | PQ306316 | PQ803172 | PV126012 | PQ807206 | PQ799264 |
| A.verticilliformis | XG04088-4 clone2 | PV069756 | PQ803173 | PV126014 | PV126026 | PQ799265 |
| A. virescens | CGMCC 3.16066* | MZ354150 | MZ350144 | MZ357410 | MZ357429 | n.a. |
| A. virescens | CGMCC 3.16067 | MZ354151 | MZ350145 | MZ357411 | MZ357430 | n.a. |
| A. xinjiangensis | CGMCC 3.16107* | OL678136 | n.a. | n.a. | n.a. | n.a. |
| A. yunnanensis | XY09528 | ON074701 | ON074686 | n.a. | n.a. | n.a. |
| A. yunnanensis | CGMCC 3.16259* | NR_182591 | NG_149054 | n.a. | n.a. | n.a. |
| A. zonata | CGMCC 3.16033* | NR_189831 | MW671548 | MZ357416 | MZ357435 | n.a. |
| A. zygospora | RSPG 214 | KC478527 | n.a. | n.a. | n.a. | n.a. |
| A. zygospora | ANG28 | DQ914420 | n.a. | n.a. | n.a. | n.a. |
| Cunninghamella blakesleeana | CBS 782.68 | JN205869 | MH870950 | n.a. | n.a. | n.a. |
Notes: New species proposed in this study are in bold. Ex-type or ex-holotype strains are labelled with an asterisk “*”. The abbreviation “n.a.” stands for “not available”.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



