
Submitted:
14 April 2025
Posted:
14 April 2025
You are already at the latest version
Abstract
The fungal genus Cunninghamella, ubiquitously distributed as saprotrophic organisms with occasional endophytic and phytopathogenic manifestations, holds significant biomedical and biochemical importance. During systematic investigations of fungal biodiversity in China, seven novel taxa (C. amphispora sp. nov., C. cinerea sp. nov., C. flava sp. nov., C. hainanensis sp. nov., C. rhizoidea sp. nov., C. simplex sp. nov., and C. yunnanensis sp. nov.) were delineated through integrated taxonomic approaches incorporating morphological characterization, multilocus phylogenetic analyses (ITS-LSU-TEF1α), and physiological assessments. Phylogenetic reconstructions positioned these novel species within a well-supported clade along with C. bainieri and C. verticillata. New species and their diagnostic features are: C. amphispora exhibiting dimorphic sporangiola production, C. cinerea distinguished by distinctive gray pigmentation in colony, C. flava characterized by yellow colony, C. hainanensis and C. yunnanensis bearing eponymous designations reflecting their geographic origins, and C. simplex displaying simplified sporangiophore branching. Comprehensive taxonomic descriptions accompanied by photomicrographic illustration are provided herein. This study constitutes the seventh installment in an ongoing series elucidating early-diverging fungal diversity in China, expanding the global Cunninghamella taxonomic inventory to 63 species and advancing our understanding of mucoralean phylogeny.
Keywords:
Mucorales
; basal fungi
; fungal diversity
; taxonomy
; molecular phylogeny
1. Introduction
The genus Cunninghamella belongs to Mucoromycota, Mucoromycetes, Mucorales and Cunninghamellaceae [1]. It was established in 1930 by Matruchot and typified with C. echinulata (Thaxt.) Thaxt. ex Blakeslee [2,3]. Members of Cunninghamella are usually saprotrophs and sometimes endophytes, and they are ubiquitous globally in various environments, including soil, air, feces and humus [4,5,6,7]. Meantime, some species of Cunninghamella are opportunistic pathogens causing infections in immunocompromised populations [6,8,9,10]. Additionally, Cunninghamella represents a pivotal contribution to the field of biomedicine and biochemistry, producing a rich array of metabolites such as fatty acids, terpenes, nickel-iron carrier and sugars [11,12,13].
Cunninghamella species are characterised by the branching pattern of sporangiophores, the shape and size of terminal and lateral vesicles, and the shape, size and surface of sporangiola [2,4,14,15,16,17,18]. In a monographic study of 2001, Zheng and Chen [2] delimitated Cunninghamella species on the basis of morphological characteristics and maximum growth temperatures. Nevertheless, identification of certain Cunninghamella remains challenging due to considerable phenotypic variabilities [2]. Later, Yu et al. [10], Liu et al. [19] and Walther et al. [3] reached a consensus among morphological observations and molecular phylogenies of ITS rDNA (internal transcribed spacer of ribosomal DNA) and TEF1α (translation elongation factor 1 alpha), overcoming the limitations of traditional classification methods and provided a more accurate and comprehensive way.
Currently, 56 Cunninghamella species are recorded in the Index Fungorum (http://www.indexfungorum.org/, accessed on 19 December 2024), and they are distributed in Antarctica, Australia, Belgium, Brazil, China, France, India, Japan, Korea, Papua New Guinea, Spain, Switzerland, Tanzania, Thailand and the United State [2,7,10,14,16,17,20,21,22,23,24,25].
Seven novel species of Cunninghamella were discovered based on molecular, morphological and physiological characteristics during the process of isolating fungi from soil samples collected from southern China. Phylogenetic trees, descriptions and illustrations of these novel species are provided herein. This is the seventh report of a serial work on diversity of Chinese early-diverging fungi [26,27,28,29,30,31].
2. Materials and Methods
2.1. Sample Collection and Strain Isolation
In 2024, soil samples were collected in Yunnan province, China. Soil sample collection was initiated by removal of surface contaminants (including leaf litter, debris, and particulates) using a sterilized stainless steel shovel. Following surface preparation, intact soil spanning the 5–10 cm subsurface horizon were extracted. Approximately 500 g of homogenized soil material was then transferred into sterial sample bags. All collected samples were immediately labeled with waterproof markers indicating collection date, GPS coordinates (± 3 m accuracy), and altitude, followed by a temporary storage at 4°C prior to laboratory processing [32,33]. Pure strains were isolated by a combination of plate dilution coating method and single spore separation method [34,35]. About 1 g of soil sample was added into a centrifugal tube containing 10 mL of sterile water. Then, 10-1 soil suspension was obtained by shaking. One milliliter of the 10-1 soil suspension was transferred into a centrifuge tube containing 9 mL of sterile water to prepare 10-2 soil suspension. In the same way, 10-3 and 10-4 soil suspensions were successfully obtained. A portion (200 µL) of 10-3 and 10-4 soil suspensions were poured on the surface of Rose Bengal Chloramphenicol agar (RBC: peptone 5.00 g/L, glucose 10.00 g/L, MgSO4·7H2O 0.50 g/L, KH2PO4 1.00 g/L rose bengal 0.05 g/L, chloramphenicol 0.10 g/L, agar 15.00 g/L) [36] containing 0.03% streptomycin sulfate, coated evenly with a glass coating rod, and then incubated in darkness at 25℃ for 3–5 d. When colony grew, it was transferred onto Potato Dextrose Agar (PDA: 200 g potato, 20 g dextrose, 20 g agar, 1000 mL distilled water, pH 7.0) for further cultivation. After sporangium formation, sporangiospores were dispersed by suspension with sterile water. The single spores were picked up by a sterile inoculation ring and transferred to a PDA plate. The plate was placed in an incubator at 25℃ for dark cultivation. All strains were stored at 4℃ in 10% glycerol. The holotype materials were deposited in the China General Microbiological Culture Collection Center, Beijing, China (CGMCC) and Shandong Normal University, Jinan, China (XG). The holotype specimens were deposited in the Herbarium Mycologicum Academiae Sinicae, Beijing, China (Fungarium, HMAS). Taxonomic information of these new strains was uploaded to the Fungal Names repository (https://nmdc.cn/fungalnames/).
2.2. Morphology and Maximum Growth Temperature
After dark culture on PDA plate for 3–5 d, the phenotypic characteristics of fungi (obverse and reverse) were photographed using a high-definition color digital camera (DP80, Olympus, Tokyo, Japan). Micromorphological features of fungi were observed by a stereomicroscope (Olympus SZX10, OLYMPUS, Tokyo, Japan) and an optical microscope (BX53, Olympus, Tokyo, Japan). For the determination of the maximum growth temperature, all strains were initially cultured in the dark at 26℃ for 3 d, and then the temperature was gradually increased by 1℃ every day until the colony stopped growing [37,38,39,40].
2.3. DNA extraction, PCR Amplification, and Sequencing
Genomic DNA was extracted from mycelia by BeaverBeads Plant DNA Kit (Cat. No.: 70409–20; BEAVER Biomedical Engineering Co., Ltd.) [41,42]. Molecular markers ITS rDNA (internal transcribed spacer of ribosomal RNA gene), LSU rDNA (large subunit of ribosomal RNA gene), and TEF1α (translation elongation factor 1 alpha gene) were amplified by polymerase chain reaction (PCR), respectively, using the following primer pairs ITS5 (5’-GGA AGT AAA AGT CGT AAC AAG G-3’) / ITS4 (5’-TCC TCC GCT TAT TGA TAT GC-3’) [43], LR0R (5’- GTA CCC GCT GAA CTT AAG C-3’) / LR5 (5’-TCC TGA GGG AAA CTT CG-3’) [44], and EF1-983F (5’- ATG ACA CCR ACR GCR ACR GTY TG-3’) / EF1-2218R (5’-AACTTGCAGGCAATGTGG-3’) [45]. Amplification procedure consisted of a predenaturation at 95°C for 5 min, 35 cycles of denaturation at 95℃ for 30 s, annealing (ITS at 55°C for 30 s, LSU at 52°C for 30s; TEF1α at 56°C for 1 min) and extension at 72°C for 1 min, and an extra extension at 72°C for 10 min. The amplification reaction was performed in a volume of 25 µL, comprising 12 µL 2 × Hieff Canace® Plus PCR Master Mix (Yeasen Biotechnology, Cat No. 10154ES03), 10 µL ddH2O, 1 µL forward and reverse primers (10 µM) (TsingKe, Qingdao, China) and 1 µL fungal genomic DNA (about 1 µM). Completeness and concentration of extracted genomic DNA were evaluated by 1% agarose gel electrophoresis [46]. Gel recovery was performed using a Gel Extraction Kit (Cat# AE0101-C; Shandong Sparkiade Biotechnology Co., Ltd.). PCR products were sent to the Biosune Company Limited (Shanghai, China) for Sanger sequencing. Sequences were assembled and proofread using MEGA v.7.0 [47]. ITS, LSU and TEF1α sequences were submitted to GenBank for BLAST similarity searches. All sequences newly acquired in this article were deposited at GenBank under the accession number in Table S1.
2.4. Phylogenetic Analyses
Phylogenetic analyses were performed based on the combined sequences of ITS, LSU and TEF1α. Sequences both generated herein and retrieved from NCBI were presented in Table S1. Phylogenetic trees were reconstructed using maximum likelihood (ML) and Bayesian inference (BI) methods. ML analysis was performed using RAxML-HPC2 on XSEDE v.8.2.12 on the CIPRES Science Gateway website (https://www.phylo.org/) with 1,000 bootstrap replicates under the GTRGAMMA model [48]. For BI analysis, a quick start configured with an automatic stop option was executed on a Linux system server [49,50]. Random initial trees were used to run six simultaneously running Markov chains for 5,000,000 generations, with samples being taken every 1,000 generations. The first 25% of trees were discarded as burn-in, and the remaining trees were employed to measure the posterior probability (PP). The layout and adjustments of the phylogenetic tree were conducted on iTOL website (https://itol.embl.de), and then the final beautification was performed through Adobe Illustrator CC 2019.
3. Results
3.1. Phylogeny
The molecular dataset included 56 strains in Cunninghamella, with Mucor janssenii CBS 205.68 as an outgroup. The dataset consisted of 3,989 characters, covering ITS rDNA (1–1,692), LSU rDNA (1,693–2,912), and TEF1α (2,913–3,989). There were 2,098 constant, 522 variable but parsimony-uninformative, and 1,369 parsimony-informative characters. MrModelTest indicated that Dirichlet fundamental frequency along with the GTR + I + G evolution pattern was suitable for both partitions in Bayesian Inference. The topology of the Maximum Likelihood (ML) evolutionary tree was highly congruent with that of the Bayesian Inference (BI) evolutionary tree. Therefore, the ML evolutionary tree was selected as a representative for detailed illustration (Figure 1). Fourteen strains of Cunninghamella isolated in this study were grouped into seven independent clades. The new species C. simplex was closely related to C. bainieri with full supports (MLBV = 100, BIPP = 1.00), and then next to another new species C. cinerea (MLBV = 100, BIPP = 0.98). The new species C. flava had a close relationship with C. verticillata. (MLBV = 80, BIPP = 0.95). The other four species each formed their own branches, that is C. amphispora (MLBV = 100, BIPP = 1), C. hainanensis (MLBV = 100, BIPP = 1), C. rhizoidea (MLBV = 100, BIPP = 0.99), and C. yunnanensis (MLBV = 100, BIPP = 1).
3.2. Taxonomy
The descriptions and illustrations of seven novel Cunninghamella species are presented as follows.
3.2.1. Cunninghamella amphispora Z.Y. Ding & X.Y. Liu, sp. nov., Figure 2.
Type—China, Yunnan Province, Puer City, Simao District, Yixiang Town, Dakai River section (22°60'26''N, 101°09'54''E, altitude 1582.73 m), from soil, 5 July 2024, M.F. Tao and X.Y. Ji, holotype HMAS 353444, ex-holotype living culture CGMCC 3.28652 (= XG09634-9-1).
Etymology—The epithet amphispora (Lat.) refers to the species producing two types of sporangiola.
Description—Colonies on PDA at 26°C for 4 d, reaching 85 mm in diameter, fast growing with a growth rate of 21.25 mm/d, initially white, gradually becoming light gray, floccose. Hyphae branched, hyaline, smooth-walled, aseptate when juvenile, septate when old. Rhizoids not found. Stolons present. Sporangiophores arising from stolons or aerial hyphae, erect or few slightly bent, unbranced or simply branched, hyaline, single, never verticillate, always expanding upwards. 2.3–19.1 µm wide. Septa if present, single, usually just below the vesicles. Vesicles hemispherical, spherical, oval or pillar-shaped, hyaline, smooth-walled, terminally 6.4–56.6 µm long and 3.2–46.7 µm wide, laterally 8–18.9 µm long and 5.1–14.8 µm wide. Pedicels over the entire surface of vesicles, 1.4–3.4 µm long. Sporangiola on pedicels, monosporous, globose, hyaline when young, dusky brown when old, thick-walled, 6.3–16.7 µm long, 6.3–15.8 µm wide, with or without spines. Spines 0.5–2.6 µm long. Chlamydospores absent. Zygospores not found.
Maximum growth temperature—29℃..
Additional strains examined—China, Yunnan Province, Puer City, Simao District, Yixiang Town, Dakai River section (22°60'26''N, 101°09'54''E, altitude 1582.73 m), from a soil sample, 5 July 2024, M.F. Tao and X.Y. Ji, living culture XG09634-9-2.
GenBank accession numbers—CGMCC 3.28652 (ITS, PV089203; LSU, PV123104; TEF1α, PV200769), XG09634-9-2 (ITS, PV089204; LSU, PV123105; TEF1α, PV200770).
Notes—In the phylogenetic tree of ITS-LSU-TEF1α, two strains of the Cunninghamella amphispora sp. nov. formed a fully supported independent clade (MLBV = 100, BIPP = 1.00; Fig. 1).
3.2.2. Cunninghamella cinerea Z.Y. Ding & X.Y. Liu, sp. nov., Figure 3.
Type—China, Yunnan Province, Puer City, Mojiang Hani Autonomous County, Tongguan Town (23°29'93''N, 101°39'17''E, altitude 1471.21 m), from soil, 4 July 2024, M.F. Tao and X.Y. Ji, holotype HMAS 353442, ex-holotype living culture CGMCC 3.28650 (= XG09556-9-1).
Etymology—The epithet cinerea (Lat.) refers to gray colonies on PDA.
Description—Colonies on PDA at 26°C for 4 d, reaching 85 mm in diameter, fast growing with a growth rate of 21.25 mm/d, initially white, gradually turning to smoky gray with age, floccose. Hyphae branched, hyaline, smooth-walled, aseptate when juvenile, septate when mature. Rhizoids abundant, root-like, hyaline, often branched. Stolons present. Sporangiophores often arising from aerial hyphae, sometimes arising from stolons, erect or slightly bent, mainly unbranched or simply branched, hyaline, mainly single or recumbent, never verticillate, usually expanding upwards, 3.1–11.7 µm wide. Septa rarely present, usually one or two below the vesicles in the main sporangiophores. Vesicles spherical to elliptic, usually hyaline, smooth, terminally 9.6–32.3 µm long and 6.9–28.3 µm wide, laterally 11.9–20.3 µm long and 9.9–17.3 µm wide. Pedicels covering the entire surface of vesicles, 1.2–2.2 µm long. Sporangiola on pedicles on vesicles, monosporous, globose to ovoid, hyaline when young, gradually dark brown when mature, thick-walled, 5.5–16.7 µm long, 4.9–15.5 µm wide, with or without spines. Spines 0.8–1.7 µm long. Chlamydospores absent. Zygospores not found.
Maximum growth temperature—33℃ .
Additional strains examined—China, Yunnan Province, Puer City, Mojiang Hani Autonomous County, Tongguan Town (23°29'93''N, 101°39'17''E, altitude 1471.21 m), from soil, 4 July 2024, M.F. Tao and X.Y. Ji, living culture XG09556-9-2.
GenBank accession numbers—CGMCC 3.28650 (ITS, PV089197; LSU, PV123098; TEF1α, PV172612), XG09556-9-2 (ITS, PV089198; LSU, PV123099; TEF1α, PV172613).
Notes—Based on the ITS-LSU-TEF1α phylogenetic tree, two strains of the Cunninghamella cinerea sp. nov. formed a well-supported clade (MLBV = 100, BIPP = 1.00; Fig. 1), exhibiting a close relationship to C. simplex and C. bainieri (MLBV = 100, BIPP = 0.98; Fig. 1). Morphologically, the new species was distinguished from C. simplex by larger terminal vesicles and lateral vesicles (9.6–32.3 × 6.9–28.3 μm vs 5.0–28.4 ×4.1–25.8 μm; 11.9–20.3 × 9.9–17.3 μm vs 6.7–17 ×3.8–10.8 μm), shorter pedicles (1.2–2.2 μm vs 1.1–2.9), root-like rhizoids. The new species produced both spinose and smooth sporangiola, while C. simplex produced only spinos sporangiola. The new species differed from C. bainieri by non-verticillate sporangiophores, shorter pedicles (1.2–2.2 μm vs 2.5–3.5 (–6)). Besides, the new species had two shapes of sporangiola (globose and ovoid), but C. bainieri had four shapes (ovoid, ellipsoid, globose and lacrymoid). Although both the new species and C. bainieri had rhizoids, the former was root-like and the latter was finger-like. Physiologically, the maximum growth temperature of the new species was 1℃ lower than that of C. bainieri (33℃ vs 34℃ ) [2].
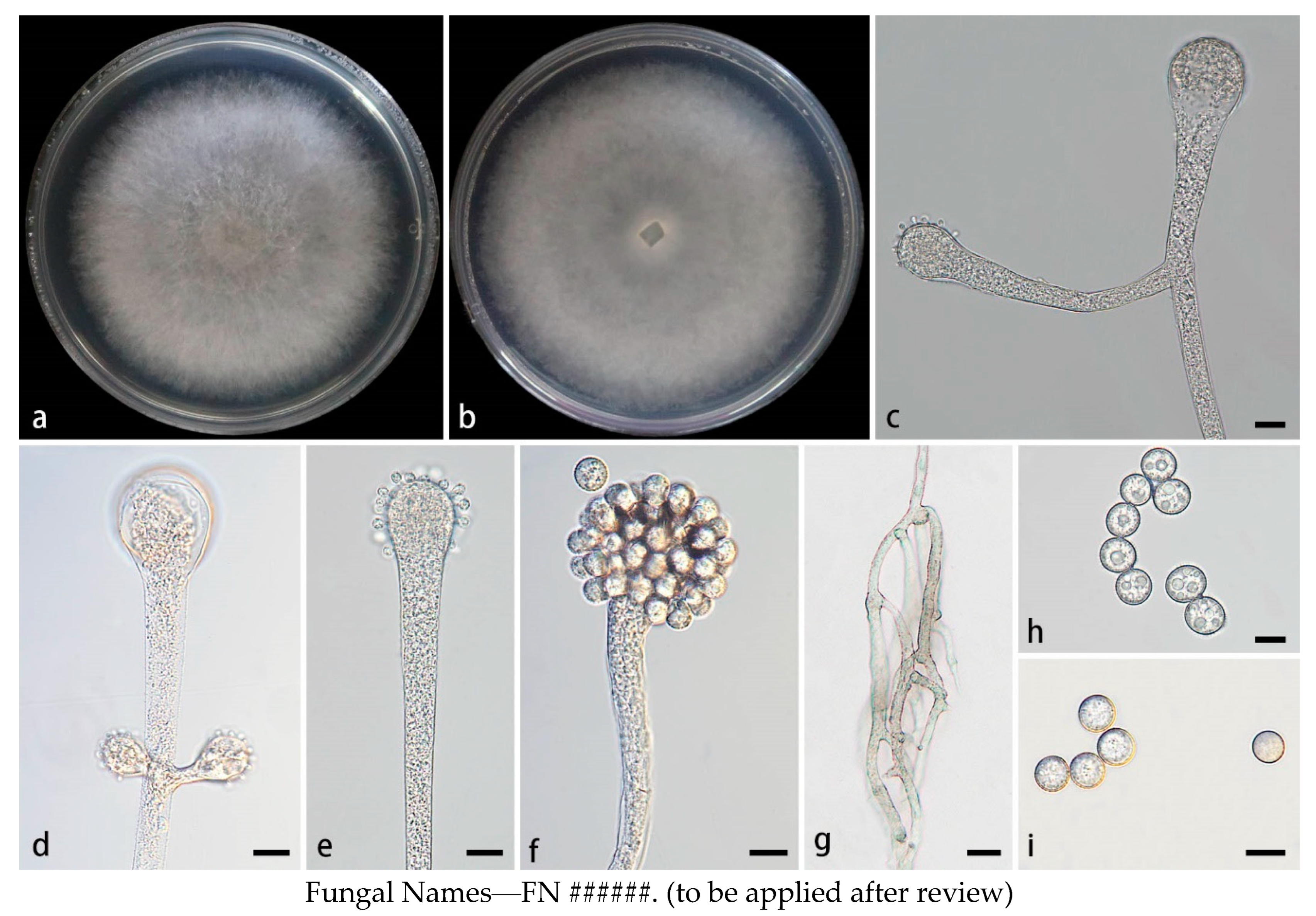
3.2.3. Cunninghamella flava Z.Y. Ding & X.Y. Liu, sp. nov., Figure 4.
Fungal Names—FN ######. (to be applied after review)
Figure 4.
Morphologies of Cunninghamella flava ex-holotype CGMCC 3.28651. (a, b) Colonies on PDA, (a) obverse, (b) reverse; (c) Sporangiophores; (d-e) Sporangiophores with branching vesicles; (f-h) Vesicles bearing sporangiola; (i) Spinose sporangiola; (j) Smooth sporangiola. Scale bars: (c-j) 10 μm.
Figure 4.
Morphologies of Cunninghamella flava ex-holotype CGMCC 3.28651. (a, b) Colonies on PDA, (a) obverse, (b) reverse; (c) Sporangiophores; (d-e) Sporangiophores with branching vesicles; (f-h) Vesicles bearing sporangiola; (i) Spinose sporangiola; (j) Smooth sporangiola. Scale bars: (c-j) 10 μm.
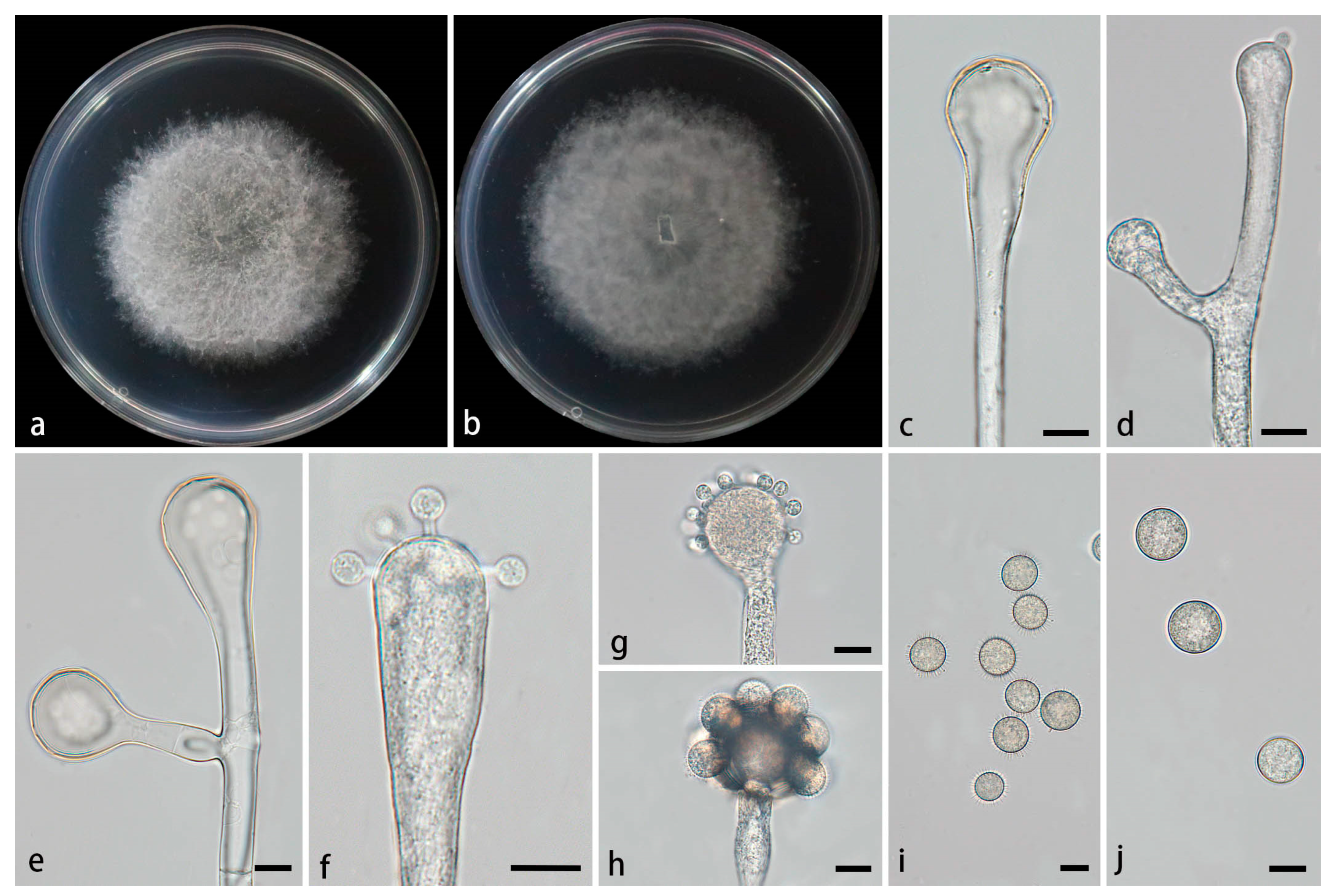
Type—China, Yunnan Province, Puer City, Mojiang Hani Autonomous County, Tongguan Town (23°29'93''N, 101°39'17''E, altitude 1471.21 m), from soil, 4 July 2024, M.F. Tao and X.Y. Ji, holotype HMAS 353443, ex-holotype living culture CGMCC 3.28651 (= XG9559-10-1).
Etymology—The epithet flava (Lat.) refers to the yellow colonies on PDA.
Description—Colonies on PDA at 26°C for 6 d, reaching 72 mm in diameter, fast growing with a growth rate of 12 mm/d, initially white, gradually turning to dry yellow with age, floccose. Hyphae branched, hyaline, smooth-walled, aseptate when juvenile, septate when mature. Rhizoids absent. Stolons present. Sporangiophores arising from aerial hyphae or stolons, erect or slightly bent, unbranched or 1–7 branched, hyaline, single, recumbent, opposite, in pairs, 1–3 verticillate, sometimes growing irregularly, swelling or bulging, then tapering upward, 3.8–25.6 µm wide. Septa if present, usually one or two below the vesicles in the main sporangiophores. Vesicles globose, pear-shaped, elliptic, usually hyaline, smooth, terminally 12.8–43.1 µm long and 11.3–44.5 µm wide, laterally 9.3–28.1 µm long and 9.4–25.3 µm wide. Pedicels on the entire surface of the vesicle, 1.6–2.2 µm long. Sporangiola on pedicles, monosporous, globose to ovoid, hyaline when young, gradually tawny when mature, thick-walled, 8.8–18.6 µm long, 8.6–18.7 µm wide, with or without spines. Spines 1.4–2.8 µm long. Chlamydospores absent. Zygospores not found.
Maximum growth temperature—33℃ .
Additional strains examined—China, Yunnan Province, Puer City, Mojiang Hani Autonomous County, Tongguan Town (23°29'93''N, 101°39'17''E, altitude 1471.21 m), from soil, 4 July 2024, M.F. Tao and X.Y. Ji, living culture XG09559-10-2.
GenBank accession numbers—CGMCC 3.28651 (ITS, PV089199; LSU, PV123100; TEF1α, PV200765), XG09559-10-2 (ITS, PV089200; LSU, PV123101; TEF1α, PV200766).
Notes—Based on the ITS-LSU-TEF1α phylogenetic tree, two strains of the Cunninghamella flava sp. nov. formed a fully supported lineage (MLBV = 100, BIPP = 1.00; Fig. 1), closely related to C. verticillata (MLBV = 80, BIPP = 0.95; Fig. 1). However, they were different in terms of morphological features such as vesicles, pedicles, rhizoids, zygosporangia and zygospores. The vesicles of the new species were characterized by various shapes including globose, pear-shaped and elliptic, while vesicles were quite regular in C. verticillata, mainly depressed-globose, subglobose to globose. The pedicles of the new species were shorter than those of C. verticillata (1.6–2.2 µm vs 2.5–4 (–6.5) µm). Rhizoids were absent in the new species, whereas present in C. verticillata. Moreover, typical zygosporangia and zygospores were generated in C. verticillata, while absent in the new species. Physiologically, the maximum growth temperature of the new species was significantly lower than those of C. verticillata (33℃ vs (37–) 39–42℃) [2].
3.2.4. Cunninghamella hainanensis Z.Y. Ding & X.Y. Liu, sp. nov., Figure 5.
Fungal Names—FN#####. (to be applied after review)
Figure 5.
Morphologies of Cunninghamella hainanensis ex-holotype CGMCC 3.28649. (a, b) Colonies on PDA, (a) obverse, (b) reverse; (c-e) Sporangiophores with branching vesicles; (f-h) Vesicles bearing sporangiola; (i) Smooth sporangiola; (j) Spinose sporangiola. Scale bars: (c-j) 10 μm.
Figure 5.
Morphologies of Cunninghamella hainanensis ex-holotype CGMCC 3.28649. (a, b) Colonies on PDA, (a) obverse, (b) reverse; (c-e) Sporangiophores with branching vesicles; (f-h) Vesicles bearing sporangiola; (i) Smooth sporangiola; (j) Spinose sporangiola. Scale bars: (c-j) 10 μm.
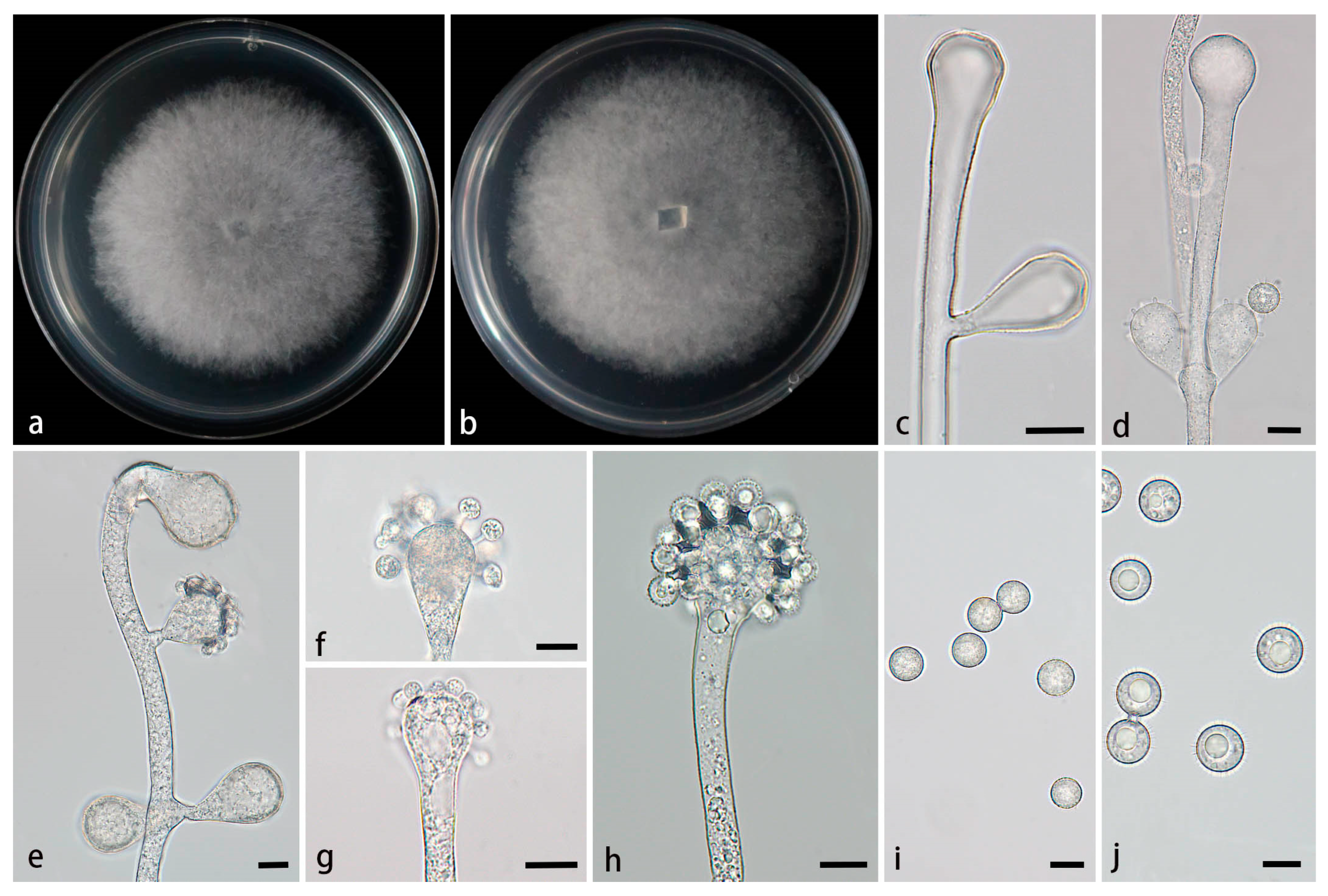
Type—China, Hainan Province, Danzhou City, near Hainan Tropical Botanical Gar den (19°51'19''N, 109°50'08''E, altitude 108 m), from a soil sample, 14 February 2024, M.F. Tao and X.Y. Ji, holotype HMAS 353441, ex-holotype living culture CGMCC 3.28649 (= XG06926-15-1).
Etymology—The epithet hainanensis (Lat.) refers to the location of the ex-holotype, Hainan Province, China.
Description—Colonies on PDA at 26°C for 4 d, reaching 85 mm in diameter, fast growing with a growth rate of 21.25 mm/d, initially white, gradually becoming light gray, floccose. Hyphae branched, hyaline, smooth-walled, aseptate when juvenile, septate with age. Rhizoids absent. Stolons present. Sporangiophores arising from stolons or aerial hyphae, mostly erect, a few slightly bent, occasionally verticillate, hyaline, single, unbranced or 1–3 branched, opposite, in pairs, 2.9-10.6 µm wide. Septa if present, usually one. Vesicles spherical, hemispherical, elliptic, smooth-walled, terminally 9.4–27 µm long and 7.9–26.5 µm wide, laterally 19.6–28.4 µm long and 12.2–22.9 µm wide. Pedicels on the entire surface of vesicles, 1.2–5.8 µm long. Sporangiola borne on pedicles, monosporous, mostly spherical, hyaline when young, dusky brown when old, thick-walled, 7.3–14 µm long, 7.8–13.4 µm wide, with or without spines. Spines 0.7–1.9 µm long. Chlamydospores absent. Zygospores not found.
Maximum growth temperature—29℃ .
Additional strains examined—China, Hainan Province, Danzhou City, near Hainan Tropical Botanical Garden (19°51'19''N, 109°50'08''E, altitude 108 m), from a soil sample, 14 February 2024, M.F. Tao and X.Y. Ji, living culture XG06926-15-2.
GenBank accession numbers—CGMCC 3.28649 (ITS, PV089195; LSU, PV123096; TEF1α, PV172610), XG06926-15-2 (ITS, PV089196; LSU, PV123097; TEF1α, PV172611).
Notes—Based on the ITS-LSU-TEF1α sequence, two strains of the Cunninghamella hainanensis sp. nov. formed a distinct clade with full supports (MLBV = 100, BIPP = 1.00; Fig. 1), showing a close relatedness to C. rhizoidea (BIPP = 0.99; Fig. 1). Morphological comparisons revealed notable differences between these two new species in sporangiola, spines length and rhizoids. The sporangiola of the C. hainanensis were larger than those of C. rhizoidea (7.3–14 µm × 7.79–13.4 µm vs 6.2–13.4 µm × 5.6–12.9 μm) and exhibited two varieties (smooth and spinose vs only spinose). The spines of C. hainanensis were shorter than those of C. rhizoidea (0.7–1.9 µm × 1.3–2.0 µm). Furthermore, C. hainanensis possessed numerous rhizoids, while the feature was absent in C. rhizoidea. Physiologically, C. hainanensis demonstrated a significantly lower maximum growth temperature than that of C. rhizoidea (29℃ vs 33℃).
3.2.5. Cunninghamella rhizoidea Z.Y. Ding & X.Y. Liu, sp. nov., Figure 6.
Fungal Names—FN ######. (to be applied after review)
Figure 6.
Morphologies of Cunninghamella rhizoidea ex-holotype CGMCC 3.28654. (a, b) Colonies on PDA, (a) obverse, (b) reverse; (c-d) Sporangiophores with branching vesicles; (e-f) Vesicles bearing sporangiola; (g) Rhizoids; (h) Spinose sporangiola. Scale bars: (c-f, h) 10 μm; (g) 20 μm.
Figure 6.
Morphologies of Cunninghamella rhizoidea ex-holotype CGMCC 3.28654. (a, b) Colonies on PDA, (a) obverse, (b) reverse; (c-d) Sporangiophores with branching vesicles; (e-f) Vesicles bearing sporangiola; (g) Rhizoids; (h) Spinose sporangiola. Scale bars: (c-f, h) 10 μm; (g) 20 μm.
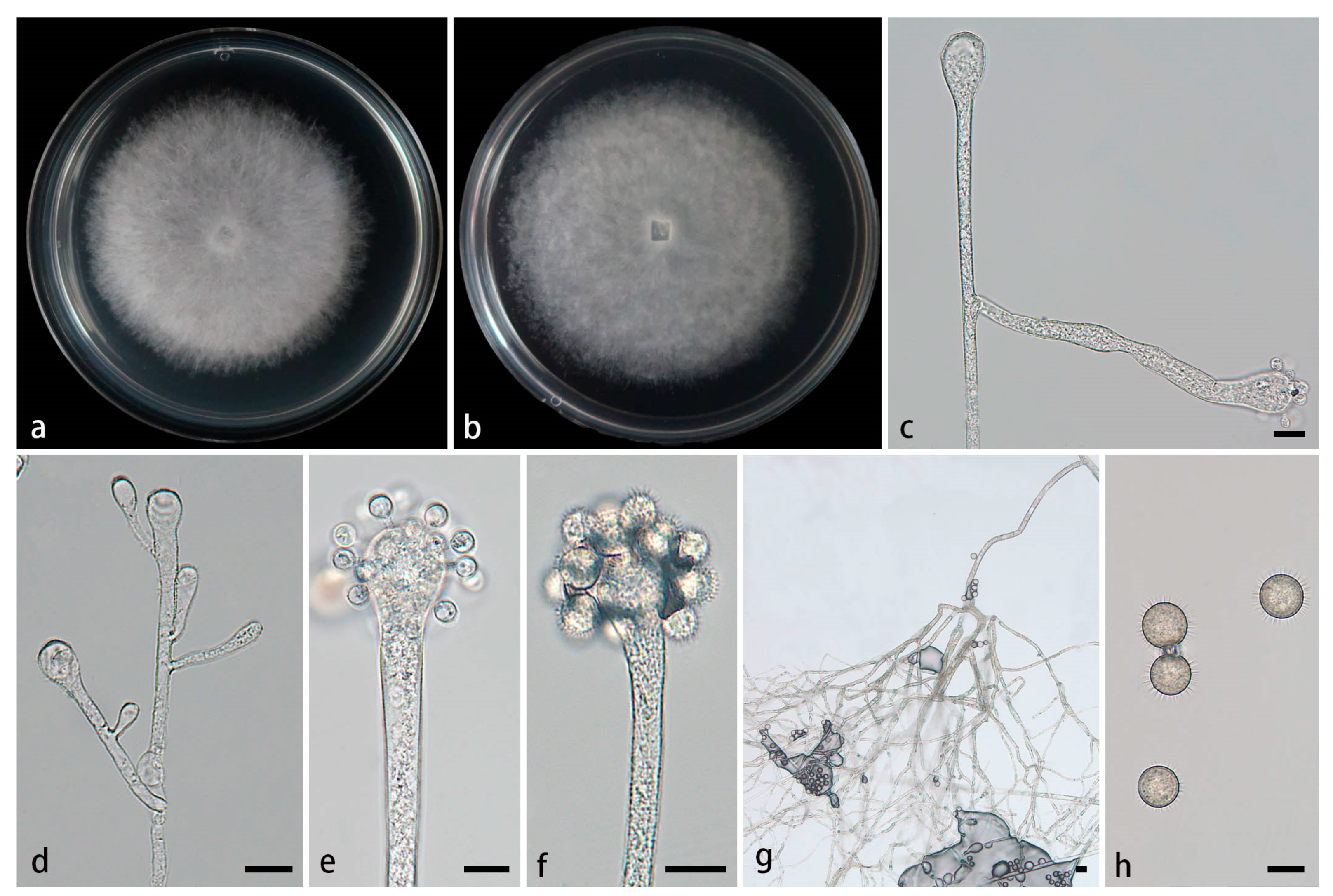
Type—China, Yunnan Province, Lincang City, Gengma Dai Autonomous County, He Pai Township (23°48'32''N, 99°41'66''E, altitude 988.59 m), from soil, 7 July 2024, M.F. Tao and X.Y. Ji, holotype HMAS 353446, ex-holotype living culture CGMCC 3.28654 (= XG09702-9-1).
Etymology—The epithet rhizoidea (Lat.) refers to the species producing abundant rhizoids.
Description—Colonies on PDA at 26°C for 4 days, reaching 85 mm in diameter, fast growing with a growth rate of 21.25 mm/d, initially white, gradually becoming gray with age, floccose. Hyphae branched, hyaline, smooth-walled, aseptate when adolescent, septate when mature. Rhizoids obviously present, root-like, complex branching, abundant. Stolons present. Sporangiophores forming from aerial hyphae or stolons, erect or slightly bent, mainly simple branches or occasionally multiple branches, hyaline, single, recumbent, opposite, occasionally verticillate, sometimes having a swollen below the sporangiophores, 2.8–11.4 µm wide. Septa if observed, usually only one in sporangiophores. Vesicles globose, club-shaped, hyaline, smooth, terminally 7.9–35.3 µm long and 4.9–32.0 µm wide, laterally 2.7–26.6 µm long and 2.4–27.1 µm wide. Pedicels forming on the surface of the vesicle, 1.5–5.4 µm long. Sporangioles formed on pedicles on vesicles, monosporous, globose to ovoid, hyaline when young, gradually tea brown when mature, thick-walled, 6.2–13.4 µm long, 5.6–12.9 µm wide, with short spines. Spinse 1.3–2.0 µm long. Chlamydospores absent. Zygospores not found.
Maximum growth temperature—33℃ .
Additional strains examined—China, Yunnan Province, Puer City, Mojiang Hani Autonomous County, Tongguan Town (23°29'93''N, 101°39'17''E, altitude 1471.21 m), from soil, 4 July 2024, M.F. Tao and X.Y. Ji, living culture XG09702-9-2.
GenBank accession numbers—CGMCC 3.28654 (ITS, PV089205; LSU, PV123106; TEF1α, PV222157), XG09702-9-2 (ITS, PV089206; LSU, PV123107; TEF1α, PV222158).
Notes—Based on the analysis of the ITS-LSU-TEF1α sequence, two strains of the Cunninghamella rhizoidea sp. nov. formed a fully supported independent lineage (MLBV = 100, BIPP = 0.99; Fig. 1), exhibiting close genetic relatedness to C. hainanensis (BIPP = 0.99; Fig. 1). Morphological comparisons further highlighted significant differences between the two species in terms of sporangiola, spine length and rhizoids (see the note to C. hainanensis above).
3.2.6. Cunninghamella simplex Z.Y. Ding & X.Y. Liu, sp. nov., Figure 7.
Fungal Names—FN ######. (to be applied after review)
Figure 7.
Morphologies of Cunninghamella simplex ex-holotype CGMCC 3.28653. (a, b) Colonies on PDA, (a) obverse, (b) reverse; (c-e) Sporangiophores with branching vesicles; (f-g) Vesicles bearing sporangiola; (h) Smooth sporangiola. Scale bars: (c-h) 10 μm.
Figure 7.
Morphologies of Cunninghamella simplex ex-holotype CGMCC 3.28653. (a, b) Colonies on PDA, (a) obverse, (b) reverse; (c-e) Sporangiophores with branching vesicles; (f-g) Vesicles bearing sporangiola; (h) Smooth sporangiola. Scale bars: (c-h) 10 μm.
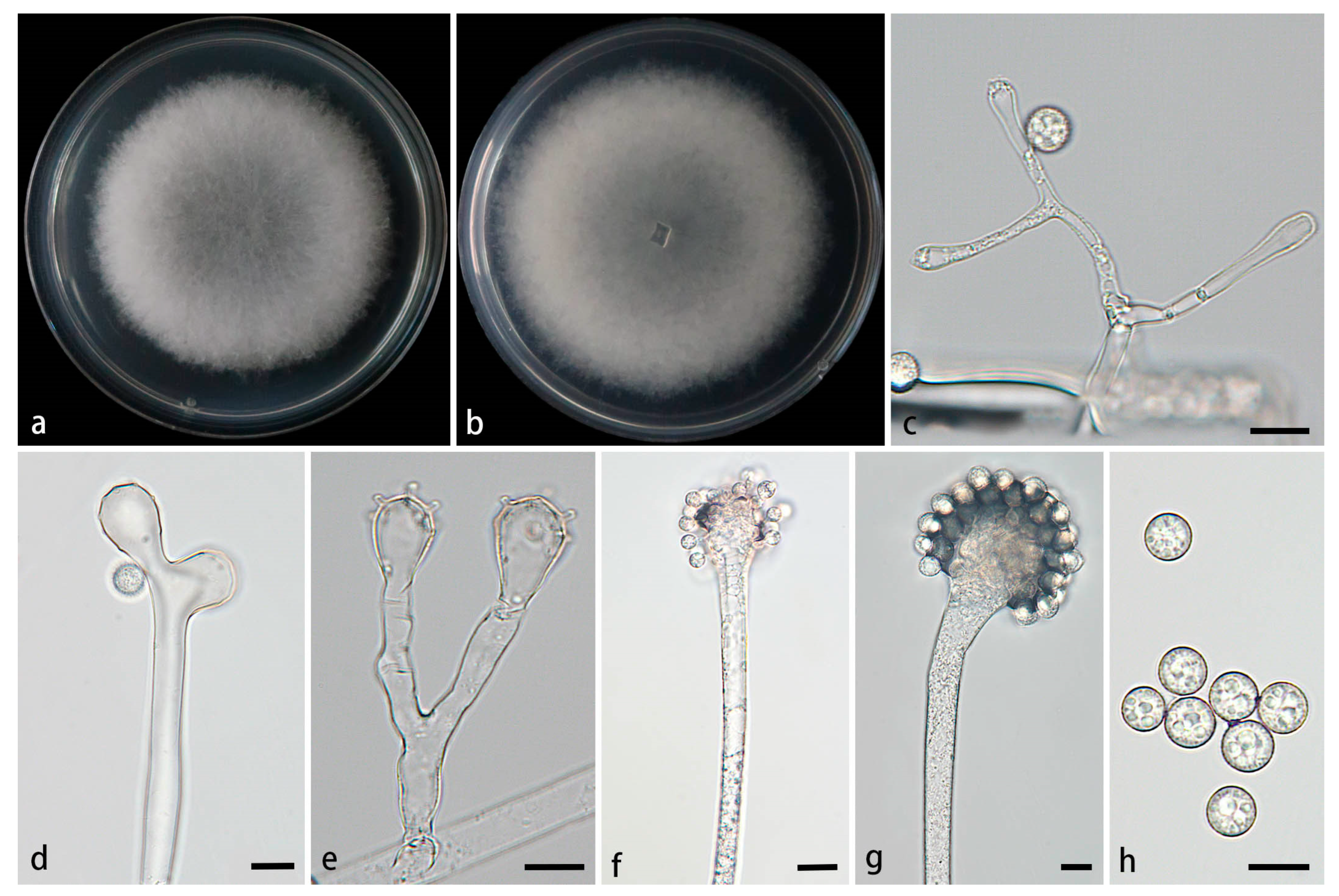
Type—China, Yinnna Province, Puer City, Simao District, Yixiang Town, Yutang Sec tion, Longlongba Jinyu Tea Estate (22°68'48''N, 101°07'32''E, altitude 1549.97 m), from soil, 5 July 2024, M.F. Tao and X.Y. Ji, holotype HMAS 353445, ex-holotype living culture CGMCC 3.28653 (= XG09611-12-1).
Etymology—The epithet simplex (Lat.) refers to the simple branching pattern of sporangiophores.
Description—Colonies on PDA at 26°C for 4 d, reaching 85 mm in diameter, fast growing with a growth rate of 21.25 mm/d, initially white, gradually turning to dusky gray, floccose. Hyphae branched, hyaline, smooth-walled, aseptate when young, septate with age. Rhizoids absent. Stolons present. Sporangiophores arising from stolons or aerial hyphae, erect or slightly bent, hyaline, unbranced or simply branched, in pairs, never verticillate, 3.1–12.2 µm wide. Septa occasionally present below vesicles in the main sporangiophores, usually only one. Vesicles spherical, oval, cylindrical, uaually hyaline, sometimes light brownish, smooth, terminally 5.0–28.4 µm long and 4.1–25.8 µm wide, laterally 6.7–17.0 µm long and 3.8–10.8 µm wide. Pedicels over the surface of vesicles, 1.1–2.9 µm long. Sporangiola borne on pedicles, monosporous, spherical to oval, hyaline when young, light grown with age, smooth, thick-walled, 5.8–10.2 µm long, 5.5–9.7 µm wide. Chlamydospores absent. Zygospores not found.
Maximum growth temperature—33℃ .
Additional strains examined—China, Yinnna Province, Puer City, Simao District, Yixiang Town, Yutang Section, Longlongba Jinyu Tea Estate (22°68'48''N, 101°07'32''E, altitude 1549.97 m), from soil, 5 July 2024, M.F. Tao and X.Y. Ji, living culture XG09611-12-2.
GenBank accession numbers—CGMCC 3.28653 (ITS, PV089201; LSU, PV123102; TEF1α, PV200767), XG09611-12-2 (ITS, PV089202; LSU, PV123103; TEF1α, PV200768).
Notes—According to the ITS-LSU-TEF1α sequence, two strains of the Cunninghamella simplex sp. nov. constituted a distinct branch with full supports (MLBV = 100, BIPP = 1.00; Fig. 1), closely related to C. bainieri (MLBV = 100, BIPP = 1.00; Fig. 1). These two species were obviously different in the morphology of sporangiophores, sporangiola, vesicles, pedicles and rhizoids. The new species was lack of pseudoverticillate and verticillate sporangiophores, while C. bainieri produced. Additionally, the new species differed from C. binariae by the simply shaped sporangiola, spherical to oval, in contrast to ovoid to ellipsoid, globose and lacrymoid of various sporangiola in C. bainieri. The difference between the new species and C. bainieri on vesicles was that the new species generates spherical, oval and cylindrical forms, whereas C. bainieri produced spherical, hemispherical and oval shapes. The pedicles of the new species were shorter than those of C. bainieri (1.1-2.9 µm vs 2.5–3.5 (–6) µm). The new species lacked rhizoids, while C. bainieri gave rise to finger-like rhizoids. Physiologically, the maximum growth temperature of the new species was one degree lower than that of C. bainieri (33℃ vs 34℃) [2].
3.2.7. Cunninghamella yunnanensis Z.Y. Ding & X.Y. Liu, sp. nov., Figure 8.
Fungal Names—FN ######. (to be applied after review)
Figure 8.
Morphologies of Cunninghamella yunnanensis ex-holotype CGMCC 3.28655. (a, b) Colonies on PDA, (a) obverse, (b) reverse; (c-d) Sporangiophores with branching vesicles; (e-f) Vesicles bearing sporangiola; (g) Rhizoids; (h) Spinose sporangiola. Scale bars: (c-h) 10 μm.
Figure 8.
Morphologies of Cunninghamella yunnanensis ex-holotype CGMCC 3.28655. (a, b) Colonies on PDA, (a) obverse, (b) reverse; (c-d) Sporangiophores with branching vesicles; (e-f) Vesicles bearing sporangiola; (g) Rhizoids; (h) Spinose sporangiola. Scale bars: (c-h) 10 μm.
Type—China, Yunnan Province, Baoshan City, Longyang District, Lujiang Dam (24°92'66''N, 98°88'15''E, altitude 703.72 m), from soil, 10 July 2024, M.F. Tao and X.Y. Ji, holotype HMAS 353447, ex-holotype living culture CGMCC 3.28655 (= XG10042-9-1).
Etymology—The epithet yunnanensis (Lat.) refers to the location, Yunnan Province, China, where the ex-holotype was collected.
Description—Colonies on PDA at 26°C for 4 d, reaching 85 mm in diameter, fast growing with a growth rate of 21.25 mm/d, initially white, gradually becoming light gray with age, floccose. Hyphae branched, hyaline, smooth-walled, exuberant, aseptate when juvenile, septate when old. Rhizoids present, root-like, hyaline, rarely branched. Stolons present. Sporangiophores producing from stolons or aerial hyphae, erect or few slightly bent, mainly unbranched, or occasionally 1–7 branched, hyaline, straight or recumbent, few verticillate, usually expanding upwards, 1.8–14.6 µm wide. Septa sometimes one or two below the vesicles in the main sporangiophores. Vesicles hemispherical, spherical, oval, elliptic, hyaline, smooth, terminally 7.1–33.8 µm long and 4.4–33.5 µm wide, laterally 5.5–13.9 µm long and 3.2–12.4 µm wide. Pedicels present throughout the vesicles, 1.1–4.1 µm long. Sporangioles borne on pedicles, monosporous, mainly globose, sometimes oval, hyaline when young, grown when mature, thick-walled, 3.3–12.4 µm long, 3.3–12.0 µm wide, with short spines. Spines 0.7–2.0 µm long. Chlamydospores absent. Zygospores not found.
Maximum growth temperature—33℃ .
Additional strains examined—China, Yunnan Province, Baoshan City, Longyang District, Lujiang Dam (24°92'66''N, 98°88'15''E, altitude 703.72 m), from soil, 10 July 2024, M.F. Tao and X.Y. Ji, living culture XG10042-9-2.
GenBank accession numbers—CGMCC 3.28655 (ITS, PV089207; LSU, PV123108; TEF1α, PV222159), XG10042-9-2 (ITS, PV089208; LSU, PV123109; TEF1α, PV222160).
Notes—Based on the ITS-LSU-TEF1α phylogenetic tree, two strains of the Cunninghamella yunnanensis sp. nov. formed a fully supported independent lineage (MLBV = 100, BIPP = 1.00; Fig. 1), basal to all the other six new species as well as C. verticillata and C. bainieri.
4. Discussion
Currently, fungal taxonomists give paramount importance to molecular data when forming new taxa or evaluating species interconnections [51,52,53]. Specifically, sequences sourced from type materials are crucial for constructing a solid phylogenetic structure, serving as the foundation for a natural classification system [54]. Nonetheless, classical morphological characteristics and physiological characteristics continued to be widely acknowledged as a vital element in this process. The morphological, physiological characteristics and molecular data together provided a comprehensive picture of the species identity and evolutionary relationships [15,20]. In this study, seven novel species of Cunninghamella from southern China were identified based on morphological, physiological and molecular data from type strains. And the morphological characteristics of these seven novel species and their relatives were systematically compared herein (Table 1).

Table 1.
morphological characteristics of Cunninghamella species.
| Species | Colonies | Sporangiophores | Vesicles | Pedicels | Sporangiola | Reference |
| C. hainanensis | PDA: 26°C 4 d, 85 mm, 21.25 mm/d, initially white, gradually becoming light gray, floccose | 2.9-10.6 µm wide, mostly erect, a few slightly bent, occasionally verticillate, unbranced or 1–3 branched, opposite, in pairs | spherical, hemispherical, elliptic, terminal vesicles 9.4–27 × 7.9–26.5µm,lateral vesicles 19.6–28.4 × 12.2–22.9 µm | 1.2–5.8 µm long | mostly spherical, 7.3–14 × 7.8–13.4µm, with or without spines, 0.7–1.9 µm long | This study |
| C.cinerea | PDA: 26°C 4 d, 85 mm, 21.25 mm/d, initially white, gradually turning to smoky gray with age, floccose | 3.1–11.7 µm wide, erect or slightly bent, mainly unbranched or simply branched, mainly single or recumbent, never verticillate | spherical to elliptic, terminal vesicles 9.6–32.3 × 6.9–28.3 µm,lateral vesicles 11.9–20.3 ×9.9–17.3 µm | 1.2–2.2 µm long | globose to ovoid, 5.5–16.7 c × 4.9–15.5 µm, with or without spines, 0.8–1.7 µm long | This study |
| C. flava | PDA: 26°C 6 d, 72 mm, 12 mm/d, initially white, gradually turning to dry yellow with age, floccose | 3.8–25.6 µm wide, erect or slightly bent, unbranched or 1–7 branched, recumbent, opposite, in pairs, 1–3 verticillate | globose, pear-shaped, elliptic, terminal vesicles 12.8–43.1 ×11.3–44.5 µm, ateral vesicles 9.3–28.1 × 9.4–25.3 µm | 1.6–2.2 µm long | globose to ovoid, 8.8–18.6 × 8.6–18.7 µm wide, with or without spines, 1.4–2.8 µm long | This study |
Table 1.
Cont.
| Species | Colonies | Sporangiophores | Vesicles | Pedicels | Sporangiola | Reference |
| C.amphispora | PDA: 26°C 4 d, 85 mm, 21.25 mm/d, initially white, gradually becoming light gray, floccose | 2.3–19.1 µm wide, erect or few slightly bent, unbranced or simply branched, hyaline, single, no verticillate | hemispherical, spherical, oval, pillar-shaped, terminal vesicles 6.4–56.6 × 3.2–46.7 µm, lateral vesicles 8–18.9 × 5.1–14.8 µm | 1.4–3.4 µm long | Globose, 6.3–16.7 × 6.3–15.8 µm, with or without spines, 0.5–2.6 µm long | This study |
| C. simplex | PDA: 26°C 4 d, 85 mm, 21.25 mm/d, nitially white, gradually turning to dusky gray, floccose | 3.1–12.2 µm wide, erect or slightly bent, hyaline, unbranced or simply branched, in pairs, never verticillate | pherical, oval, cylindrical, terminal vesicles 5.0–28.4 ×4.1–25.8 µm, lateral vesicles 6.7–17.0 ×3.8–10.8 µm | 1.1–2.9 µm long | spherical to oval, 5.8–10.2 × 5.5–9.7 µm | This study |
| C. rhizoidea | PDA: 26°C 4 d, 85 mm, 21.25 mm/d, initially white, gradually becoming gray with age, floccose | .8–11.4 µm wide, erect or slightly bent, simple branches or ccasionally multiple branches, recumbent, opposite, occasionally verticillate | globose, club-shaped, terminal vesicles 7.9–35.3 × 4.9–32.0 µm; lateral vesicles 2.7–26.6 ×2.4–27.1 µm | 1.5–5.4 µm long | globose to ovoid,6.2–13.4 × 5.6–12.9 µm, with short spines, 1.3–2.0 µm long | This study |
| C. yunnanensis | PDA: 26°C 4 d, 85 mm, 21.25 mm/d, initially white, gradually becoming light gray with age, floccose | 1.8–14.6 µm wide, rect or few slightly bent, mainly unbranched, or occasionally 1–7 branched, straight or recumbent, few verticillate | emispherical, spherical, oval, elliptic, terminal vesicles 7.1–33.8 × 4.4–33.5 µm; lateral vesicles 5.5–13.9 × 3.2–12.4 µm | 1.1–4.1 µm long | mainly globose, sometimes oval, 3.3–12.4 × 3.3–12.0 µm wide, with short spines, 0.7–2.0 µm long | This study |
| C. bainieri | SMA: 27°C 4 d, 90mm, at first white, soon becoming light grey, grey, to ‘Light Mouse Grey’, reverse cream, floccose | rect, bent, or recumbent, main axes of sporangiophores (8–) 11–21µm wide; primary branches (1–) 4–10 (–18) µm wide, monopodial, pseudoverticillate or verticillate in 1–2(–3) whorls of 3–8, typically in pairs | globose, subglobose to ovoid, sometimes irregular, axial vesicles 18.5–32(–40) μm; lateral ones (8–)13.5–30 μm | 2.5–3.5 (–6) um long | ovoid to ellipsoid and 7–14.5 (–20) × 6.5-11 (–14.5) µm globose and 5.5–12.5µm, lacrymoid and 9–20 (–32.5) × 7–14.5 (–20) µm | [2] |
Table 1.
Cont.
| Species | Colonies | Sporangiophores | Vesicles | Pedicels | Sporangiola | Reference |
| C. verticillata | SMA: 28°C 5–6 d, 90mm, at first white, from the sixth day near ‘Avellaneous’ to near ‘Colonial Buff’, revcrse yellowish cream, floccose | erect, straight or recumbent, main axes of sporangiophores 7.5–17.5 (–25) µm; branches (0–) 4-15 (–25), verticillate to pseudoverticillate, rarely singly or in pairs, mostly simple, very rarely re-branched | axial ones slightly depressed-globose, subglobose to globose, 225–50 (–70) μm; lateral ones usually globose to subglobose, sometimes broadly ovoid, 8 5–27.5 (–32.5) μm | 2.5–4 (–6.5) um long | Two kinds: globose, broadly ellipsoid ovoid and bluntly pointed at one end, 6–17.5 (–20) × 5.5–13.5 (–17.5) µm; dark giant sp orangiola, globose, 11.5-17.5 (-25) µm | [2] |
The phylogenetic analysis of fourteen strains using combined ITS-LSU-TEF1α se quences revealed seven robust monophylies (Figure 1). Cunninghamella hainanensis and C. rhizoidea were sister to each other. Morphologically, the sporangiola of the former were larger than that of the latter. And the former had shorter spines and plentiful rhizoids. C. cinerea had close relationship with C. simplex and C. bainieri. Compared with C. simplex, C. cinerea had larger vesicles, shorter pedicels, rhizoids and spineless sporangiola. C. cinerea differed from C. bainieri by nonverticillate sporangiophores, shorter pedicles, as well as different shapes of rhizoids and sporangiola [2]. In contrast to C. verticillata, which had almost all globular vesicles, the vesicles were more varied in C. flava like globose, pear-shaped and elliptic. Additionally, C. flava had shorter pedicels and no rhizoids. More importantly, C. flava lacked the classical sexual stage, but C. verticillata was present [2]. In comparison to C. cinerea, C. amphispora owned wider sporangiophores, larger lateral vesicles, smaller lateral vesicles, longer pedicles and spines. In comparison to C. bainieri, C. simplex had shorter pedicles, different shapes of vesicles and sporangiola. Sporangiophores were not pseudoverticillate and verticillate in C. simplex, but they were present in C. bainieri [2].
In addition, temperature also served as crucial factor in the categorization of fungi [14]. The maximum growth temperatures of the seven newly discovered species C. amphispora, C. hainanensis, C. cinerea, C. flava, C. rhizoidea, C. simplex and C. yunnanensis were 29℃, 29℃, 33℃, 33℃, 33℃, 33℃, 33℃, respectively. Although the temperature tolerance of these new species was not very high, the maximum temperature of C. verticillata, a close relative of C. flava, was 42℃ [2]. C. amphispora and C. hainanensis are same in maximum growth temperature, but different in microscopic characteristics. In conclusion, both morphological and physiological traits are crucial for classifying fungi.
With the discovery of these seven new species, the genus Cunninghamella currently accommodates 63 species. This expansion not only broadens our taxonomic knowledge, but also paves the way for new research opportunities, promising to yield valuable insights into fungal diversity and function.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Supplementary Table S1:GenBank accession numbers of sequences used in this study;Supplementary File S1:the combined ITS-LSU- TEF1α sequence matrix used in this study.
Author Contributions
Z.Y. Ding was responsible for DNA sequencing, photo editing and paper drafting; Y. Jiang and W.X. Liu was responsible for data analyses; M.F. Tao, Y.X. Wang and X.Y. Ji collected soil samples; S. Wang was responsible for data analyses and manuscript revision; X.Y. Liu took charge of naming the new species, conceiving and revising the paper, and providing funding.
Funding
This study was supported by the National Natural Science Foundation of China (Nos. 32170012, 32470004 and 32300011), Ji’nan City’s ‘New University 20 Policies’ Initiative for Innovative Research Teams Project (202228028), Innovative Agricultural Application Technology Project of Jinan City (CX202210) and Key Technological Innovation Program of Shandong Province, China (2022CXGC020710).
Institutional Review Board Statement
Not applicable for studies involving humans or animals.
Informed Consent Statement
Not applicable.
Data Availability Statement
The sequences from the present study were submitted to the NCBI database (https://www.ncbi.nlm.nih.gov/, accessed on 12 February 2025). The sequences were deposited in the GenBank database (Table S1).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Spatafora, J.W.; Chang, Y.; Benny, G.L.; L azarus, K.; Smith, M.E.; Berbee, M.L.; Bonito, G.; Corradi, N.; Grigoriev, I.; Gryganskyi, A.; James, T.Y.; OʹDonnell, K.; Roberson, R.W.; Taylor, T.N.; Uehling, J.; Vilgalys, R.; White, M.M.; Stajich, J.E. A phylum-level phylogenetic classification of zygomycete fungi based on genome-scale data. Mycologia. 2016, 108, 1028–1046. [CrossRef]
- Zheng, R.Y.; Chen, G.Q. A monograph of Cunninghamella. Mycotaxon. 2001, 80, 1–75.
- Walther, G.; Pawłowska, J.; Alastruey-Izquierdo, A.; Wrzosek, M.; Rodriguez-Tudela, J.L.; Dolatabadi, S.; Chakrabarti, A.; de Hoog, G.S. DNA barcoding in Mucorales: an inventory of biodiversity. Persoonia. 2013, 30, 11–47. [CrossRef]
- Matruchot, L. Une Mucorinée purement conidienne, Cunninghamella africana. Annales Mycologici. 1903, 1, 45–60.
- Baijai, U.; Mehrotra, B.S. The genus Cunninghamella - a reassessment. Sydowia. 1980, 33, 1–13.
- Reed, M.D.A.; Body, P.B.; Austin, M.D.M.B.; Jr Frierson, M.D.H. Cunninghamella bertholletiae and Pneumocystis carinii pneumonia as a fatal complication of chronic lymphocytic leukemia. Human Pathology. 1988, 19, 1470–1472. [CrossRef]
- Nguyen, T.T.T.; Choi, Y.J.; Lee, H.B. Zygomycete fungi in Korea: Cunninghamella bertholletiae, Cunninghamella echinulata, and Cunninghamella elegans. Mycobiology 2017, 45, 318–326. [CrossRef]
- Gomes, M.Z.; Lewis, R.E.; Kontoyiannis, D.P. Mucormycosis caused by unusual mucormycetes, non-Rhizopus, -Mucor, and -Lichtheimia. Clin. Microbiol. Rev. 2011, 24, 411–45.
- Kwon-Chung, K.J. Taxonomy of fungi causing mucormycosis and entomophthoramycosis (zygomycosis) and nomenclature of the disease: molecular mycologic perspectives. Clin. Infect. Dis. 2012, 54 Suppl 1, S8–S15. [CrossRef]
- Yu, J.; Walther, G.; Van Diepeningen, A.D.; Gerrits Van Den Ende, A.H.G.; Li, R.Y.; Moussa, T.A.A.; Almaghrabi, O.A.; De Hoog, G.S. DNA barcoding of clinically relevant Cunninghamella species. Med. Mycol. 2015, 53, 99–106. [CrossRef]
- Fakas, S.; Papanikolaou, S.; Galiotou-Panayotou, M.; Komaitis, M.; Aggelis. G. Organic nitrogen of tomato waste hydrolysate enhances glucose uptake and lipid accumulation in Cunninghamella echinulata. J. Appl. Microbiol. 2008, 105, 1062–1070. [CrossRef]
- Alakhras, R.; Bellou, S.; Fotaki, G.; Stephanou, G.; Demopoulos, N. A.; Papanikolaou, S.; Aggelis, G. Fatty acid lithium salts from Cunninghamella echinulata have cytotoxic and genotoxic effects on HL-60 human leukemia cells. Eng. Life. Sci. 2015, 15, 243–253. [CrossRef]
- Zhao H, Lv ML, Liu Z, et al. High-yield oleaginous fungi and high-value microbial lipid resources from Mucoromycota. Bioenerg. Res. 2020, 14, 1196–1206. [CrossRef]
- Liu, C.W.; Liou, G.Y.; Chien, C.Y. New records of the genus Cunninghamella (Mucorales) in Taiwan. Fungal Science. 2005, 20, 1–9.
- Guo, J.; Wang, H.; Liu, D.; Zhang, J.N.; Zhao, Y.H.; Liu, T.X.; Xin, Z.H. Isolation of Cunninghamella bigelovii sp. nov. CGMCC 8094 as a new endophytic oleaginous fungus from Salicornia bigelovii. Mycol. Prog. 2015, 14, 1–8. 11. [CrossRef]
- Alves, A.L.; de Souza, C.A.; de Oliveira, R.J.; Cordeiro, T.R.; de Santiago, A.L. 2017 Cunninghamella clavata from Brazil: a new record for the western hemisphere. Mycotaxon. 2017, 132, 381–389.
- Bragulat, M.R.; Castella, G.; Isidoro-Ayza, M.; Domingo, M.; Cabanes, F.J. Characterization and phylogenetic analysis of a Cunninghamella bertholletiae isolate from a bottlenose dolphin (Tursiops truncatus). Revista Iberoamericana de Micología 2017, 34, 215–219. [CrossRef]
- Weitzman, I.; Crist, M.Y. Studies with clinical isolates of Cunninghamella II. Physiological and morphological studies. Mycologia 1980, 72, 661–9.
- Liu, X.Y.; Huang, H.; Zheng, R.Y. Relationships within Cunninghamella based on sequence analysis of ITS rDNA. Mycotaxon 2001, 80, 77–95.
- Hyde, K.D.; Hongsanan, S.; Jeewon, R.; Bhat, D.J.; McKenzie, E.H.C.; Jones, E.B.G.; Phookamsak, R.; Ariyawansa, H.A.; Boonmee, S.; Zhao, Q.; Abdel-Aziz, F.A.; Abdel-Wahab, M.; Banmai, S.; Chomnunti, P.; Cui, B.K.; Daranagama, D.A.; Das, K.; Dayarathne, M.C.; De Silva, N.I.; Dissanayake, A.J.; Doilom, M.; Ekanayaka, A.H.; Gibertoni, T.B.; Góes-Neto, A.; Huang, S.K.; Jayasiri, S.C.; Jayawardena, R.S.; Konta, S.; Lee, H.B.; Li, W.J.; Lin, C.G.; Liu, J.K.; Lu, Y.Z.; Luo, Z.L.; Manawasinghe, I.S.; Manimohan, P.; Mapook, A.; Niskanen, T.; Norphanphoun, C.; Papizadeh, M.; Perera, R.H.; Phukhamsakda, C.; Richter, C.; De Santiago, A.L.C.M.; Drechsler-Santos, E.R.; Senanayake, I.C.; Tanaka, K. Fungal diversity notes 367-490: taxonomic and phylogenetic contributions to fungal taxa. Fungal Diversity 2016, 80, 1–270. [CrossRef]
- Suwannarach, N.; Kumla, J.; Supo, C.; Honda, Y.; Nakazawa, T.; Khanongnuch, C.; Wongputtisin, P. Cunninghamella saisamornae (Cunninghamellaceae, Mucorales), a new soil fungus from northern Thailand. Phytotaxa 2021,509, 291–300. [CrossRef]
- Zhang, Z.Y.; Zhao, Y.X.; Shen, X.; Chen, W.H.; Han, Y.F.; Huang, J.Z.; Liang, Z.Q. Molecular phylogeny and morphology of Cunninghamella guizhouensis sp. nov. (Cunninghamellaceae, Mucorales), from soil in Guizhou, China. Phytotaxa 2020, 455, 31–39. [CrossRef]
- Wang, Y.J.; Zhao, T.; Wu, W.Y.; Wang, M.; Liu, X.Y. Cunninghamella verrucosa sp. nov. (Mucorales, Mucoromycota) from Guangdong province in China. Phytotaxa 2022, 560: 274–284.
- Zhao, H.; Zhu, J.; Zong, T.K.; Liu, X.L.; Ren, L.Y.; Lin, Q.; Qiao, M.; Nie, Y.; Zhang, Z.D.; Liu, X.Y. Two new species in the family Cunninghamellaceae from China. Mycobiology 2021, 49, 142–150. [CrossRef]
- Zhao, H.; Nie, Y.; Zong, T.; Wang, K.; Lv, M.; Cui, Y.; Tohtirjap, A.; Chen, J.; Zhao, C.; Wu, F.; Cui, B.; Yuan, Y.; Dai, Y.; Liu, X.Y. Species diversity, updated classification and divergence times of the phylum Mucoromycota. Fungal Diversity 2023, 123, 49–157. [CrossRef]
- Zhao, H.; Nie, Y.; Huang, B.; Liu, X.Y. Unveiling species diversity within early-diverging fungi from China I: three new species of Backusella (Backusellaceae, Mucoromycota). MycoKeys 2024, 109, 285–304. [CrossRef]
- Tao, M.F.; Ding, Z.Y.; Wang, Y.X.; Zhang, Z.X.; Zhao, H.; Meng, Z.; Liu, X.Y. Unveiling species diversity within early-diverging fungi from China II: Three new species of Absidia (Cunninghamellaceae, Mucoromycota) from Hainan Province. MycoKeys 2024, 110, 255–272. [CrossRef]
- Wang, Y.X.; Zhao, H.; Jiang, Y.; Liu, X.Y.; Tao, M.F.; Liu, X.Y. Unveiling species diversity within early-diverging fungi from China III: Six new species and a new record of Gongronella. (Cunninghamellaceae, Mucoromycota). MycoKeys 2024, 110, 287–317. [CrossRef]
- Ding, Z.Y.; Ji, X.Y.; Tao, M.F.; Jiang, Y.; Liu, W.X.; Wang, Y.X.; Meng, Z.; Liu, X.Y. Unveiling species diversity within early-diverging fungi from China IV: Four new species of Absidia (Cunninghamellaceae, Mucoromycota). MycoKeys 2025. (under review).
- Ji, X.Y.; Ding, Z.Y.; Nie, Y.; Zhao, H.; Wang, S.; Huang, B.; Liu, X.Y. Unveiling species diversity: within early-diverging fungi. From China V: Five new species of Absidia (Cunninghamellaceae, Mucoromycota) MycoKeys 2025. (under review).
- Wang, Y.X.; Ding, Z.Y.; Ji, X.Y.; Meng, Z.; Liu, X.Y. Unveiling species diversity within early-diverging fungi from China VI: Four Absidia sp. nov. (Mucorales) in Guizhou and Hainan. Microorganisms 2025. (under review).
- Li, W. X.; Wei, Y. H.; Zou, Y.; Liu, P.; Li, Z.; Gontcharov, A. A.; Stephenson, S. L.; Wang, Q.; Zhang, S. H.; Li, Y. Dictyostelid cellular slime molds from the Russian Far East. Protist 2020, 171, 125756. [CrossRef]
- Zou, Y.; Hou, J. G.; Guo, S. N.; Li, C. T.; Li, Z.; Stephenson, S. L.; Pavlov, I. N.; Liu, P.; Li, Y. Diversity of dictyostelid cellular slime molds, including two species new to science, in forest soils of Changbai Mountain, China. Microbiol. Spectr. 2022, 10, e0240222. [CrossRef]
- Zhao, H.; Nie, Y.; Zong, T.; Dai, Y.; Liu, X.Y. Three new species of Absidia (Mucoromycota) from China based on phylogeny, morphology and physiology. Diversity 2022, 14, 132. [CrossRef]
- Zhao, H.; Nie, Y.; Zong, T.; Wang, Y.; Wang, M.; Dai, Y.; Liu, X.Y. Species diversity and ecological habitat of Absidia (Cunninghamellaceae, Mucorales) with emphasis on five new species from forest and grassland soil in China. Journal of Fungi 2022, 8, 471. [CrossRef]
- Corry, J.E.L. Rose bengal chloramphenicol (RBC) agar. In: Progress in Industrial Microbiology; Elsevier: Amsterdam, The Netherlands, 1995, pp. 431–433.
- Zheng, R.Y.; Chen, G.Q.; Huang, H.; Lt g t S, Liu, X.Y. A monograph of Rhizopus. Sydowia 2007, 59, 273–372.
- Zheng, R.Y.; Liu, X.Y.; Li, R.Y. More Rhizomucor causing human mucormycosis from China: R. chlamydosporus sp. nov. Sydowia 2009, 61, 135–147.
- Zheng, R.Y.; Liu, X.Y. Taxa of Pilaira (Mucorales, Zygomycota) from China. Nova Hedwigia 2009, 88, 255–267. [CrossRef]
- Zong, T.K.; Zha, H.; Liu, X.L.; Ren, L.Y.; Zhao, C.L.; Liu, X.Y. Taxonomy and phylogeny of four new species in Absidia (Cunninghamellaceae, Mucorales) from China. Frontiers in Microbiology 2021, 12, 677836. [CrossRef]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [CrossRef]
- Guo, L.D.; Hyde, K.D.; Liew, E.C.Y. Identification of endophytic fungi from Livistona chinensis based on morphology and rDNA sequences. New Phytol. 2000, 147, 617–630. http://doi.org/10.1046/j.1469-8137.2000.00716.x.
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press: New York, NY, USA, 1990; pp. 315–322.
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [CrossRef]
- O’Donnell, K.; Lutzoni, F.M.; Ward, T.J.; Benny, G.L. Evolutionary relationships among mucoralean fungi Zygomycota: Evidence for family polyphyly on a large scale. Mycologia 2001, 93, 286–297. [CrossRef]
- Zhang, Z.; Liu, R.; Liu, S.; Mu, T.; Zhang, X.; Xia, J. Morphological and phylogenetic analyses reveal two new species of Sporocadaceae from Hainan, China. MycoKeys 2022, 88, 171–192. [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [CrossRef]
- Nylander, J. MrModeltest V2: Program distributed by the author. Bioinformatics 2004, 24, 581–583. [CrossRef]
- Bauer, R.; Garnica, S.; Oberwinkler, F.; Riess, K.; Weiß, M.; Begerow, D. Entorrhizomycota: a new fungal phylum reveals new perspectives on the evolution of fungi. PLoS One 2015, 10, e0128183. [CrossRef]
- Jeewon, R.; Hyde, K.D. Establishing species boundaries and new taxa among fungi: recommendations to resolve taxonomic ambiguities. Mycosphere 2016, 7, 1669–1677. [CrossRef]
- Maharachchikumbura, S.S.N.; Hyde, K.D.; Jones, E.B.G.; McKenzie, E.H.C.; Bhat, J.D.; Dayarathne, M.C.; Huang, S.K.; Norphanphoun, C.; Senanayake, I.C.; Perera, R.H.; Shang, Q.J.; Xiao, Y.P.; D’souza, M.J.; Hongsanan, S.; Jayawardena, R.S.; Daranagama, D.A.; Konta, S.; Goonasekara, I.D.; Zhuang, W.Y.; Jeewon, R.; Phillips, A.J.L.; Abdel-Wahab, M.A.; Al-Sadi, A.M.; Bahkali, A.H.; Boonmee, S.; Boonyuen, N.; Cheewangkoon, R.; Dissanayake, A.J.; Kang, J.C.; Li, Q.R.; Liu, J.K.; Liu, X.Z.; Liu, Z.Y.; Luangsaard, J.J.; Pang, K.L.; Phookamsak, R.; Promputtha, I.; Suetrong, S.; Stadler, M.; Wen, T.C.; Wijayawardene, N.N. Families of Sordariomycetes. Fungal Diversity 2016, 79, 1–317. [CrossRef]
- Hu, D.M.; Wang, M.; Cai, L. Phylogenetic assessment and taxonomic revision of Mariannaea. Mycological Progress 2016, 16, 271–283. [CrossRef]
Figure 1.
The Maximum Likelihood (ML) phylogenetic tree of Cunninghamella based on sequences of ITS, LSU rDNA and TEF1α, with Mucor jansseni as an outgroup. Nodes are annotated with ML bootstrap values (MLBV≥70%) and BI posterior probabilities (BIPP≥0.9), separated by slashes "/". The bold black represents ex-type or ex-holotype strains with an asterisk "*". Strains isolated in this study are displayed in red. The scale at the bottom left indicates 0.1 substitutions per site.
Figure 1.
The Maximum Likelihood (ML) phylogenetic tree of Cunninghamella based on sequences of ITS, LSU rDNA and TEF1α, with Mucor jansseni as an outgroup. Nodes are annotated with ML bootstrap values (MLBV≥70%) and BI posterior probabilities (BIPP≥0.9), separated by slashes "/". The bold black represents ex-type or ex-holotype strains with an asterisk "*". Strains isolated in this study are displayed in red. The scale at the bottom left indicates 0.1 substitutions per site.
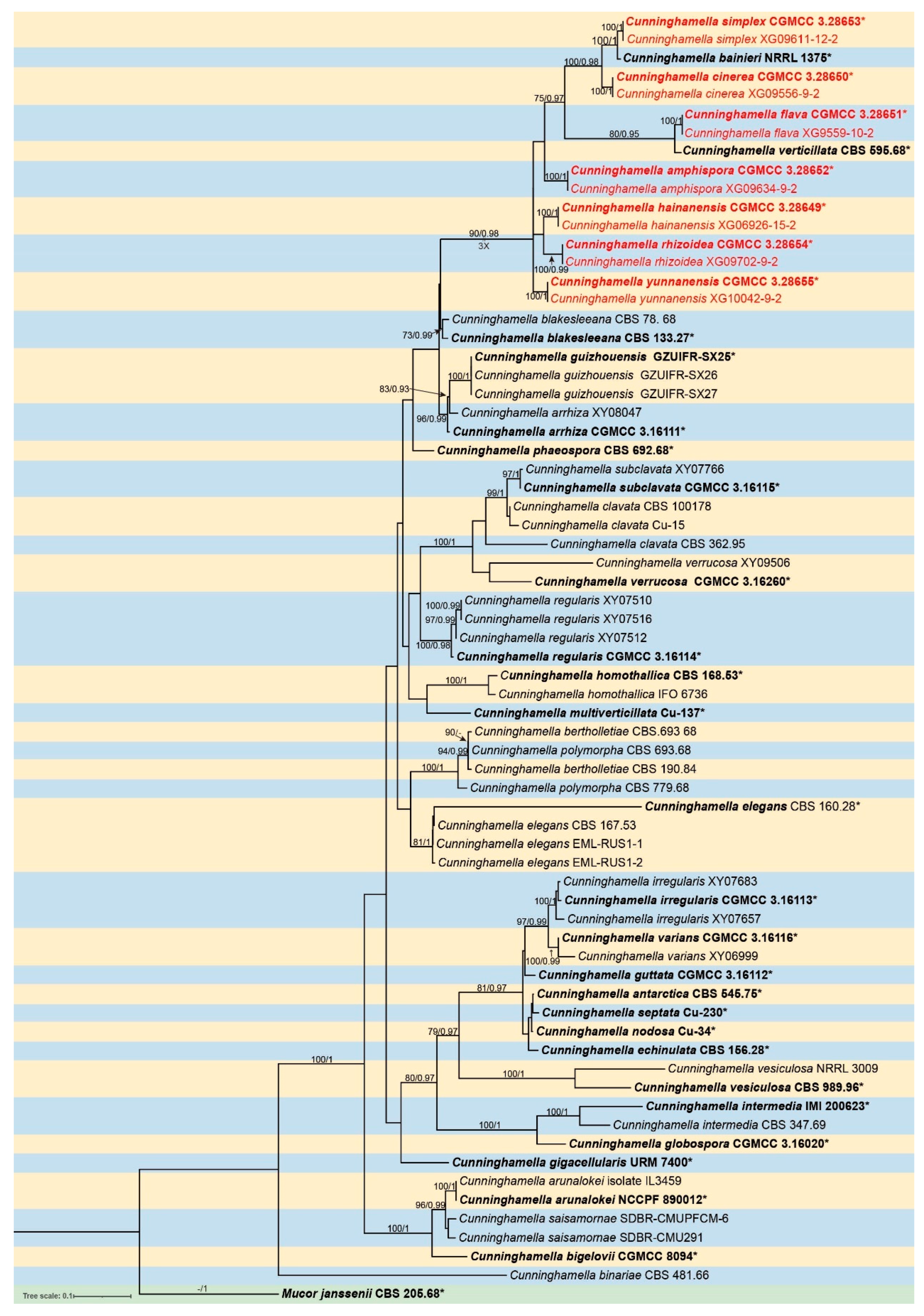
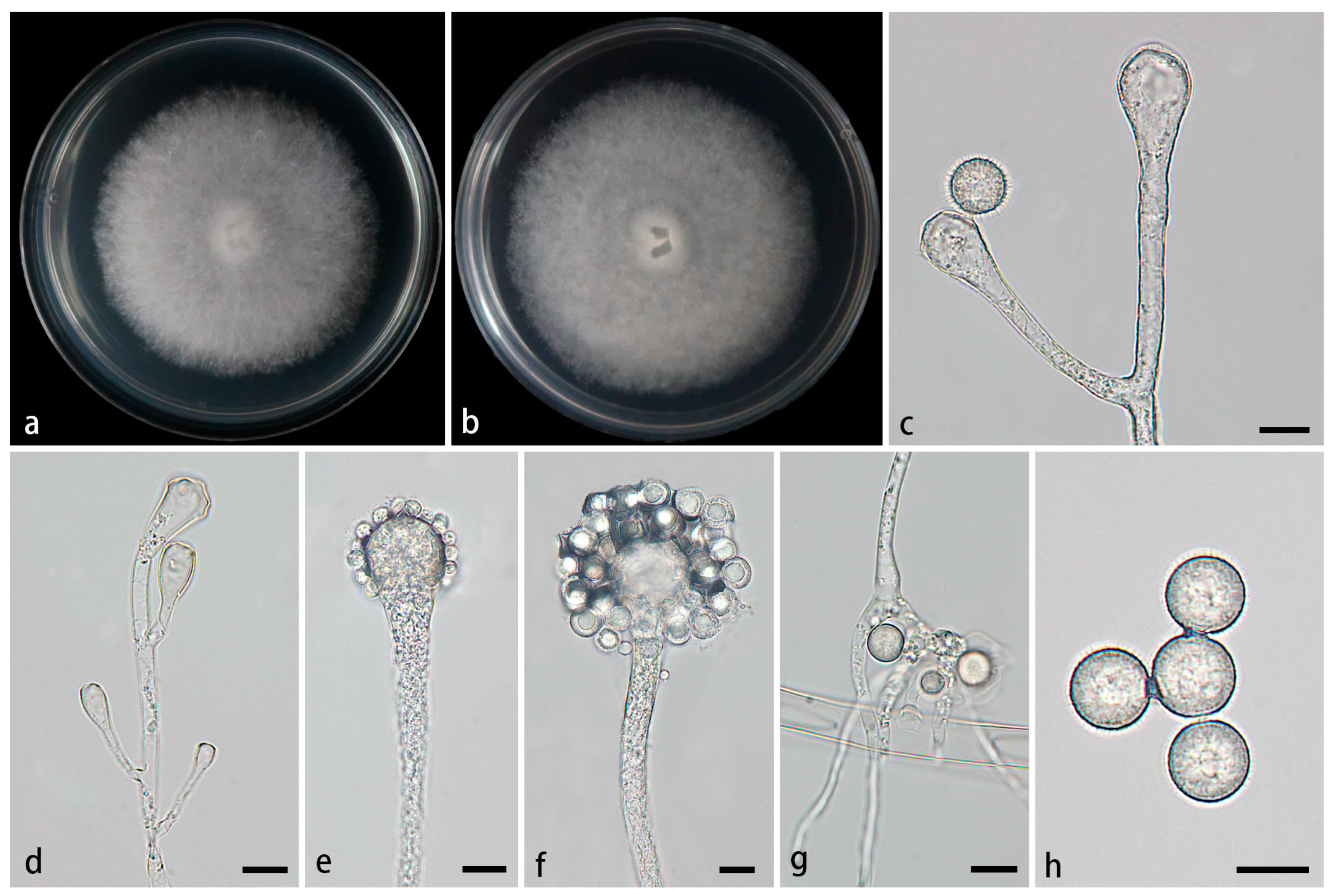
Figure 2.
Morphologies of Cunninghamella amphispora ex-holotype CGMCC 3.28652. (a, b) Colonies on PDA, (a) obverse, (b) reverse; (c-e) Sporangiophores with branching vesicles; (f-g) Vesicles bearing sporangiola; (h) Spinose sporangiola; (i) Smooth sporangiola. Scale bars: (c-i) 10 μm.
Figure 2.
Morphologies of Cunninghamella amphispora ex-holotype CGMCC 3.28652. (a, b) Colonies on PDA, (a) obverse, (b) reverse; (c-e) Sporangiophores with branching vesicles; (f-g) Vesicles bearing sporangiola; (h) Spinose sporangiola; (i) Smooth sporangiola. Scale bars: (c-i) 10 μm.
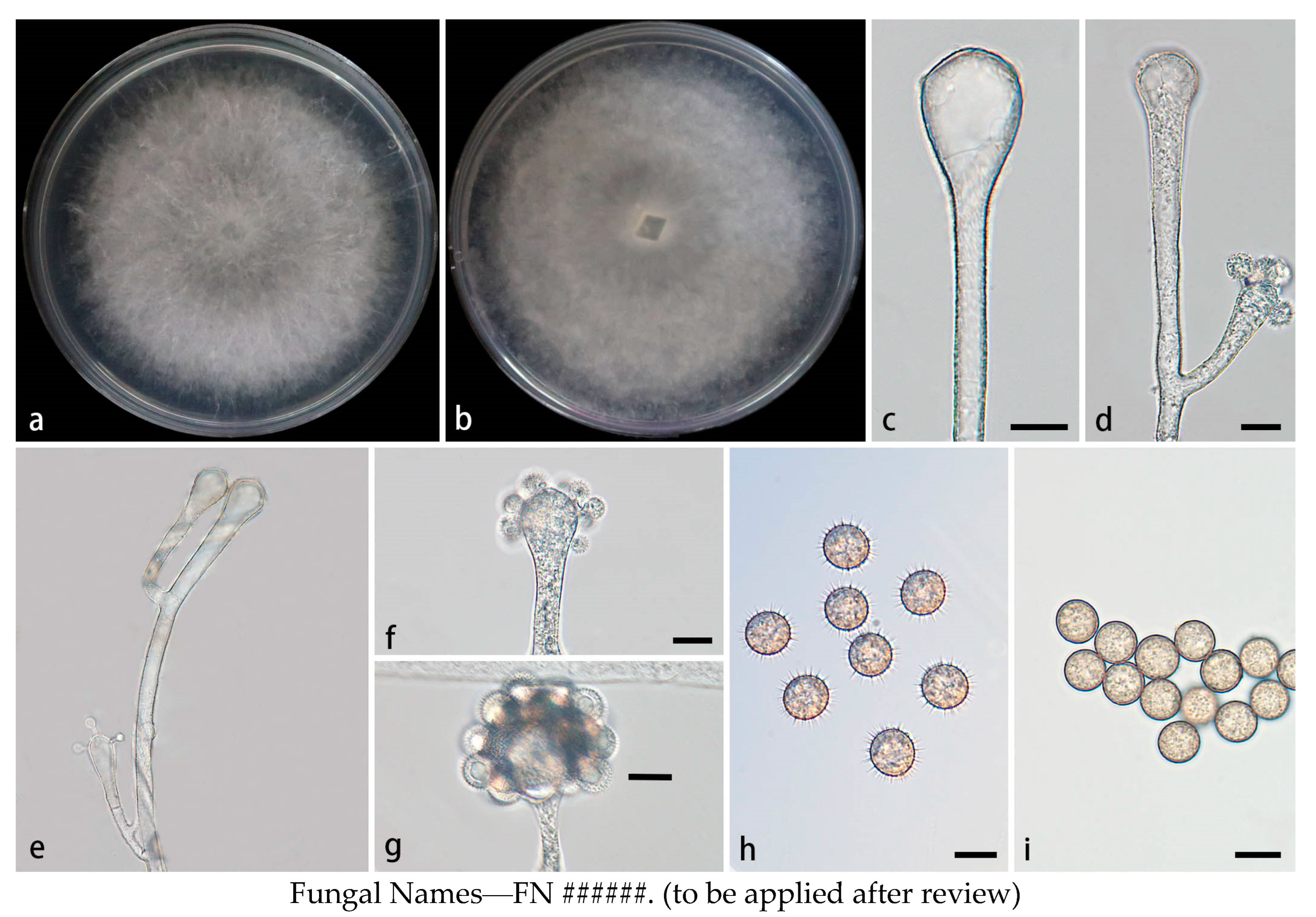
Figure 3.
Morphologies of Cunninghamella cinerea ex-holotype CGMCC 3.28650. (a, b) Colonies on PDA, (a) obverse, (b) reverse; (c-d) Sporangiophores with branching vesicles; (e-f) Vesicles bearing sporangiola; (g) Rhizoids; (h) Spinose sporangiola; (i) Smooth sporangiola. Scale bars: (c-f, h-i) 10 μm; (g) 20 μm.
Figure 3.
Morphologies of Cunninghamella cinerea ex-holotype CGMCC 3.28650. (a, b) Colonies on PDA, (a) obverse, (b) reverse; (c-d) Sporangiophores with branching vesicles; (e-f) Vesicles bearing sporangiola; (g) Rhizoids; (h) Spinose sporangiola; (i) Smooth sporangiola. Scale bars: (c-f, h-i) 10 μm; (g) 20 μm.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



