
Submitted:
09 February 2025
Posted:
10 February 2025
You are already at the latest version
Abstract
The effects of three mulch types (vermicompost, plastic, and straw) plus an untreated control, in factorial combination with three irrigation regimes (RFD: rainfed condition; SIF: one supplemental irrigation at the flowering stage; SIVF: two supplemental irrigations at the vegetative and flowering stages) on linseed growth, seed yield and oil composition, and biochemical status were investigated. Linseed plants under two irrigations and straw mulch showed the highest seed yield (300 g and 222.4 g m-2, respectively), biological yield (887.9 g m-2), and concentration of oleic and linoleic acids. The plastic mulch application under rainfed conditions or one irrigation increased stearic and palmitic acid concentrations, respectively. Water deficit stress encouraged the accumulation of proline (10.1 μmol g-1 fresh weight (FW)), total phenols (6.68 mg g-1 FW), and DPPH radical scavenging capacity (56.5%). Plants grown in straw mulched soil showed the highest total phenol content and DPPH radical scavenging capacity, while control plants displayed the highest proline concentration. Enzyme activities, including catalase and superoxide dismutase, were enhanced under straw and plastic mulch compared to control plants under rainfed conditions. Our findings suggest that straw mulch represents an effective, sustainable strategy to successfully manage linseed crops, mitigating the adverse effects of water deficit stress on plant performance.
Keywords:
Linum usitatissimum L.
; soil cover
; irrigation regime
; antioxidant enzymes
; phenolics
; soluble sugars
1. Introduction
Oilseed crops are cultivated primarily for their seed oil content, catering to various industries and diets. Key oilseed crops include soybean, sunflower, rapeseed, cotton, and peanut. [1]. Linseed (Linum usitatissimum L.) is one of the oldest cultivated oilseeds, prized for its versatile applications in foods, paints, varnishes, and linoleum flooring [2].
Linseed is rich in bioactive compounds like unsaturated fatty acids, fiber, proteins, antioxidants, and lignans, offering potential health benefits, including cancer prevention [3] and, in this respect, its α-linolenic acid content is the highest among plant sources [4]. The 18-carbon alpha-linolenic acid, is a plant-based source of essential omega-3 polyunsaturated fatty acids that are beneficial for maintaining a healthy diet [4], with positive effects on heart health, nervous system function, and inflammatory regulation [3]. Linseeds are also rich in protein and B-group vitamins [5].
Environmental changes, including decreased rainfall, pose challenges to crop cultivation, with water scarcity significantly impacting crop growth and productivity, especially during critical growth phases [6]. Insufficient moisture disrupts metabolic pathways crucial for water relations, photosynthesis, antioxidant function, and mineral uptake, affecting crop quality and yield [7]. Like other crops, linseed suffers from impaired gas exchange and leaf water relations under water stress [5]. Moisture deficits impede growth by inhibiting cell expansion, cell wall division, and differentiation [7] and photosynthesis by closing stomata [8] and destroying chlorophyll due to the excessive release of reactive oxygen species compounds (ROS) [9], leading to decreased productivity [10].
Mulch offers a practical solution to mitigate the metioned effects by enhancing soil water retention [11,12], reducing evapotranspiration [13], decreasing weed growth, and improving soil health and fertility [7]; notably, plastic mulch can effectively conserve soil moisture and enhance water use efficiency [12]. Moreover, vermicompost and straw mulch increase soil microorganism activity and water retention, especially in water-deficient conditions [9,14].
Understanding the interaction between different irrigation regimes and mulches on linseed morphological and physiological traits is crucial for optimizing crop productivity under varying moisture conditions. Therefore, this study aimed to evaluate the efficacy of plastic and straw mulch, and vermicompost in enhancing linseed seed yield, quality and oil content performances under two irrigation regimes and rainfed conditions.
2. Materials and Methods
Growing Conditions and Experimental Design
Research was conducted in 2022 at a farm located in Urmia city in Western Azerbaijan province, Iran (37° 3′ 24.82″ N, 44° 58′ 12.42″ E, 1,32 m a. s. l.), characterized by a semi-arid climate, with an average annual temperature of 13.4°C and annual precipitation of 261 mm, in a loam-clay soil (at 0–0.3 m depth), with 0.12% N, 395 mg kg–1 K, 10 mg kg–1 P, and pH 7.9. The meteorological data (Table 4) were obtained from the Iran Meteorological Organization, and the average annual temperature and precipitation over the last ten years were 12°C and 390 mm, respectively, at the research site.
The experimental protocol was based on the factorial combination of three mulch types (plastic, straw, and vermicompost) plus an untreated control (bare soil), and three irrigation regimes (RFD: rainfed condition; SIF: one supplemental irrigation at the flowering stage; SIVF: two supplemental irrigations at the vegetative and flowering stages). A randomized complete block design was used for the treatment distribution in the field, with three replicates.
Linseed seeds were sourced from the Agriculture and Natural Resources Research and Education Centre of Urmia, Iran. Each plot measured 3 × 4 m, with 10 cm spacing between the plants along the rows 50 cm apart. Seeds were sown manually at a 4 cm depth on 17 March 2022. Before sowing, a fertilization with 80 kg ha–1 urea and 100 kg ha–1 triple superphosphate was practiced. Each mulch type was applied in a 4–6 cm thick layer, covering the surface area of the plot and surrounding the linseed plants. Plant thinning occurred at the 3–4-leaf leaf stage, manual weeding was done throughout the growth period as required, and no pesticides were used during the growing season.

Table 1.
Weather data from October 2021 to September 2022 at the research site in Urmia.
| Oct | Nov | Dec | Jan | Feb | Mar | Apr | May | June | July | Aug | Sep | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Monthly average temperature (°C) | 3.3 | 30.6 | 48.9 | 19.6 | 13.4 | 20.9 | 18.1 | 58.4 | 1.2 | 1.4 | 0.1 | 0.1 |
| Monthly average precipitation (mm) | 16.3 | 9.1 | 5.4 | -1.7 | 0.5 | 5.3 | 10.7 | 16.4 | 22.4 | 25.6 | 28.0 | 25.3 |
Irrigation was applied during the vegetative and flowering stages. The amount of water applied (6400 m3 ha–1 per irrigation cycle) was determined based on the soil moisture deficit required to reach field capacity using Walker’s [15] method. Supplemental irrigation accounted for 20% of the average annual long-term rainfall in the area.
Representative leaf samples were collected at the full flowering stage, flash-frozen in liquid nitrogen, and stored at –80°C for subsequent analysis. Plants were harvested from 5 m2 within each plot, excluding the edges, at the end of the growing season on 29 June 2022.
Measurements
Growth Parameters
At maturity, ten harvested linseed plants per plot were assessed for growth parameters, including plant height, branch number, follicle number, seed number per follicle, and 100-seed weight. A 2 m2 area in the center of each plot was harvested to estimate linseed seed yield and biological yield. Plant samples were oven-dried at 72°C for 48 h before recording dry weights as biological yield. Seeds were separated from follicles and weighed after oven-drying at 72°C to adjust the moisture content to 14–15%.
Oil Content and Oilseed Yield
Linseed oil content was extracted from dried seeds using the American Oil Chemists’ Society [16] method. In summary, 5 g ground seed was subjected to a six-hour extraction using 300 mL n-hexane in a Soxhlet extractor before removing the solvent using a rotavapor (Heidolph, Schwabach, Germany). The resulting oil was collected in a dedicated glass container to facilitate further compound isolation and identification [17].
Seed oil content and oil yield were determined as follows [18]:
Oil content (%) = (Extracted oil content (%))/(5 g of linseed seed)×100 (1)
Oil content yield (kg ha–1) = Oil content (%) × Seed yield (g m–2) (2)
Fatty Acids
Fatty acids were converted into fatty acid methyl esters (FAMEs) to enhance their volatility for GC-FID analysis by mixing 0.1 g oil with 1.5 mL hexane and 0.2 mL of 2 N methanolic KOH. The mixture was vortexed for 5 s and then centrifuged at 2,500 rpm for 1 minute. The upper layer containing the FAMEs was carefully separated and stored at 4°C for further analysis. GC-FID analysis was conducted using an Agilent 6890 N GC instrument (Wilmington, DE, USA) with an FID detector. FAME separations were carried out on an HP-88 capillary column (88% cyanopropyl aryl-polysiloxane, 100 m length, 0.25 mm inner diameter, 0.2 μm film thickness) (Agilent) as follows: initial hold at 140°C for 5 minutes, followed by ramping of 4°C min-1 to 240°C, and a final hold at 240°C for 15 minutes. Nitrogen was the carrier gas at a 1.0 mL min–1 flow rate. The injection port and detector temperatures were set at 260°C and 280°C, respectively. The injector operated in split mode with a 1 30 split ratio. Data acquisition and processing were performed using ChemStation software. A commercially available FAME mixture (Supelco 37 Component FAME Mix, Bellefonte, PA, USA) was used as a reference standard to identify fatty acids.
Relative water content (RWC)
The RWC was determined as follows [19]:
% RWC = [(FW–DW)/(TW–DW)] × 100 (3)
After determining the fresh weight (FW), leaves were soaked in distilled water for 16–18 h. The turgid samples were then quickly blotted dry to remove excess surface water before determining the turgid weight (TW). The leaves were oven-dried at 70°C for 24 h to determine dry weight (DW).
Chlorophyll and Carotenoid Content
Fresh leaf samples (0.5 g) at the full flowering stage were pulverized in liquid nitrogen, mixed with 10 mL of 80% acetone, and homogenized by centrifugation at 4,000 rpm for 15 minutes. Subsequently, the extracted pigments were quantified using a spectrophotometer (Germany, N50-Touch), enabling accurate measurement of chlorophyll a, b, and carotenoid contents [20].
Total Soluble Sugar Content
The phenol-sulfuric acid method was used to estimate leaf total soluble sugar content. Leaf tissue (0.5 g) was powdered in a mortar using liquid nitrogen, mixed with ethanol, and combined with 5% phenol. Subsequently, 5 mL of 98% sulfuric acid was added to the mixture of solutions and incubated for one hour before measuring the solution absorption at 485 nm using a spectrophotometer [21].
Proline Content
Leaf proline content was determined using the ninhydrin colorimetric method. Briefly, 0.5 g leaf tissue was ground in a mortar using liquid nitrogen, homogenized in 10 mL of 3% sulfosalicylic acid solution, and then centrifuged at 4,000 rpm for 15 minutes to obtain a clear supernatant. A glacial acetic acid solution of proline ninhydrin acid was prepared in a 1: 1: 1 ratio for the colorimetric evaluation of proline and equilibrated at 100°C for one hour to facilitate the reaction between proline and ninhydrin, forming a chromophore. The reaction was terminated by rapidly cooling the solution in an ice bath. To develop the chromophore, 4 mL toluene was added to the reaction mixture, enabling the extraction of the chromophore into the organic phase. The absorbance of the samples was measured at 515 nm using a spectrophotometer [22].
Antioxidant Enzyme Extractions and Assays
Antioxidant enzyme activities were quantified by finely pulverized 100 mg fresh material in 2 mL of 0.1 M KH2PO4 buffer, including 5% polyvinylpyrrolidone (PVP) at pH 6. Extracts from the plants were centrifuged for 30 minutes at 3°C at 15,000 rpm, with enzyme activity determined from the clear supernatant [23].
Catalase (CAT) activity was measured at 240 nm using hydrogen peroxide (H2O2) concentration change. The reaction mixture contained 1.9 mL of 50 mM K3PO4, buffered at a pH of 7, 10 mM H2O2, and 0.2 mL enzyme extract. Enzymatic activity was measured in 60 s mg–1 according to variations in protein absorption [24].
Superoxide dismutase (SOD) activity was measured at 560 nm to minimize the photochemical loss of nitroblue tetrazolium (NBT), as described by Beyer and Fridovich [25]. One unit of SOD was defined as the enzyme amount required to inhibit a 50% decrease in NBT.
Total Polyphenol Content
Total phenolic content (TPC) was evaluated using the Folin-Ciocalteau method [26]. Briefly, 1,600 µL purified water and 10 µL extracts containing methanol were combined and incubated for 5 minutes at 25°C with 200 µL Folin-Ciocalteau reagent (10% v/v) prepared in distilled water. After adding 200 µL NaCO3 (7.5%), the solution mixture was kept at 25°C in darkness for 30 minutes. The absorbance of the samples was measured at 760 nm by a UV/visible spectrophotometer (DB-20/DB-20S) for quantitative estimation of TPC. The results are presented in milligrams of equivalent gallic acid (3,4,5 trihydroxybenzoic acid) per gram of sample dry weight (mg GAE g‒1 dry weight).
DPPH (2,2-Diphenyl-1-Picrylhydrazyl-Hydrate) Radical Scavenging Activity
DPPH radical scavenging activity was measured according to Brand-Williams et al. [27]. After combining 2.0 mL DPPH solution with 15 μL methanolic extract, the mixture was left in the dark at 20°C for 30 minutes. The absorbance of the resulting solution was measured at 517 nm. Equation 4 was used to calculate DPPH inhibition.
where Abcontrol and Absample are the absorbances of the control and the sample, respectively.
Inhibition (%) = ((Abcontrol − Absample)/(Abcontrol)) × 100
Data Statistical Processing
Data were analyzed by analysis of variance, and Duncan’s multiple range test was used to assess significant differences between treatments at p < 0.05 significance level, using the SAS 9.1 software.
3. Results
Mulch type and irrigation regime mainly affected linseed plant height, branch number, follicle number, seed yield, biological yield, and oil content; their interaction was significant on follicle seed number, 100-seed weight, and oil yield (Table 1).
Biometrical and Seed Yield Parameters
Plant height under irrigation exceeded that of rainfed conditions (Figure 1), with the tallest plants (37 cm) upon two irrigations during the vegetative and flowering stages, not different from those irrigated only during the flowering stage (Figure 1), while the rainfed ones were the shortest (32.9 cm). Among the mulching treatments, straw produced the tallest plants (37.4 cm), not different from those treated with vermicompost, while the control ones were the shortest (32.4 cm) (Figure 1).
Table 1.
Analysis of variance of linseed biometrical and yield traits as influenced by mulch type and irrigation regime.
Table 1.
Analysis of variance of linseed biometrical and yield traits as influenced by mulch type and irrigation regime.
| Source of variation | Plant height | Branch number | Follicle number | Follicle seed number | 100-seed weight | Seed yield | Biological yield | Oilseed content | Oilseed yield | |
|---|---|---|---|---|---|---|---|---|---|---|
| Mulch (M) | ** | ** | ** | ** | ** | ** | ** | ** | ** | |
| Irrigation condition (IC) | ** | * | ** | * | ** | * | * | ** | ** | |
| M x IC | n.s. | n.s. | n.s. | * | ** | n.s. | n.s. | n.s. | * |
** and *: significant at p ≤ 0.01 and 0.05, respectively; n.s., not significant.
Plants irrigated twice during the vegetative and flowering stages produced the highest follicle number (94.9) (Figure 1), and the rainfed ones the lowest (53.6) not significantly different from that reached under irrigation at the flowering stage (Figure 1). As for mulching treatments, straw led to the highest number of follicles (83.7), not significantly different from that elicited by vermicompost or plastic mulch (Figure 1), and the control plants to the lowest (5.7) (Figure 1).
Plants mulched with straw and irrigated twice produced the highest number of follicle seeds (7.9), not significantly different from that recorded under one irrigation, whereas the control plants under rainfed conditions showed the fewest follicles (Figure 2).
The highest 100-seed weight was recorded with straw mulch and two irrigations (0.44 g), the lowest in control plants under rainfed conditions (0.36 g) not significantly different from that recorded upon one irrigation during the flowering stage (Figure 2).
The irrigated plants had significantly greater seed yield than those under rainfed conditions, with the highest value corresponding to two irrigations during the vegetative and flowering stages (300 g m–2) and the lowest in rainfed control plants (130.5 g m–2) (Figure 3). Regarding the mulching treatments, straw mulch elicited the highest seed yield (222.4 g m–2) and the control plants the lowest (187.3 g m–2), whereas no significant differences were recorded differ between plants treated with vermicompost and plastic mulch (Figure 3).
The highest value of biological yield was recorded under two irrigations at the vegetative and flowering stages (887.9 g m–2) and the lowest in rainfed control plants (466.3 g m–2) (Figure 3). As for the mulching treatments, straw led to the highest biological yield (703.0 g m–2) not significantly different from plastic mulch, and the control the lowest (618.6 g m–2) (Figure 3).
Oilseed Content, Yield and Fatty Acid Composition
Plants irrigated twice showed the highest oilseed content (38.5%), while the rainfed ones the lowest (33.8%) (Figure 4). Regarding the mulching treatments, straw elicited the highest oil content (37.8%) not significantly different from vermicompost or plastic mulch, while control plants had the lowest yield (33.7%) (Figure 4).
Oilseed yield was the highest in twice irrigated plants with straw mulch (129.7 g m–2) and the lowest in control under rainfed conditions (39.8 g m–2) not significantly different from that recorded upon one irrigation (Figure 4).
Physiological Traits
Relative Water Content
Mulch type and irrigation regime had a significant effect on linseed RWC (Table 3). As shown in Figure 5, two irrigations at the vegetative and flowering stages led to the highest RWC (54.8%), not significantly different from that recorded under one irrigation at the flowering stage, with the rainfed plants showing the lowest RWC (39.5%). Regarding the mulching treatments, straw mulch elicited the highest RWC (52.6%), not significantly different from plastic mulch, and the lowest RWC (44.7%) was detected in plant grown in bare soil (Figure 5).
Photosynthetic Pigments
The organic and synthetic mulches significantly increased chlorophyll a, chlorophyll b, and carotenoids compared to the untreated control (Figure 6), with the highest levels recorded in plants grown in straw mulched soil under two irrigations (1.5, 0.6 and 13.7 mg g–1 FW, respectively). No significant differences in chlorophyll b and carotenoids were recorded between the organic and synthetic mulch treatments irrigated twice. Plants grown in bare soil under rainfed conditions attained the lowest levels of chlorophyll a, b, and carotenoids (0.8, 0.3, and 7.1 mg g–1 FW, respectively) (Figure 6). Moreover, no significant differences in chlorophyll b arose between the unmulched control and organic and synthetic mulches under rainfed conditions. Similarly, no carotenoid differences were recorded between vermicompost-treated and control plants under rainfed conditions (Figure 6).
Total Soluble Sugars and Proline Content
Straw mulch and two irrigations elicited the highest total soluble sugar content (18.8 mg g–1 FW), whereas plants grown in bare soil under rainfed conditions showed the lowest level (15.0 mg g–1 FW) not significantly different from those mulched with vermicompost under rainfed conditions (Figure 7).
Under rainfed conditions, plants grown in unmulched soil had the highest proline content (10.1 μmol g–1 FW) (Figure 7), whereas no significant differences were recorded between vermicompost and plastic mulch.
Two irrigations at the vegetative and flowering stages resulted in the lowest proline content under straw mulch (5.0 μmol g–1 FW) (Figure 7), while one irrigation at the flowering stage did not lead to significant differences between vermicompost and bare soil grown plants.
Antioxidant Enzyme Activity
Under rainfed and irrigation, organic and synthetic mulches increased antioxidant enzyme activities (Figure 8). Rainfed plants treated with straw mulch had the highest CAT and SOD activities (6.9 and 16.2 μmol min–1 g–1 FW, respectively). No significant differences in CAT activity were recorded between straw and plastic mulch treatments under rainfed conditions (Figure 8). Similarly, there was no significant difference in SOD activity between vermicompost and plastic mulch treatments under rainfed conditions. Control plants irrigated twice during the vegetative and flowering stages had the lowest CAT and SOD activities (0.7 and 4.8 μmol min–1 g–1 FW, respectively) (Figure 8).
Total Phenol Content and DPPH Radical Scavenging Capacity
The organic and synthetic mulches significantly increased total phenol content and DPPH radical scavenging capacity, compared to the control (Figure 9).
Rainfed conditions and straw mulch had the highest total phenol content (6.7 mg g–1 FW) and DPPH radical scavenging capacity (56.5%), whereas no significant differences were recorded between organic and synthetic mulch in twice irrigated plants (Figure 9). Similarly, vermicompost and plastic mulch did not significantly differ in DPPH radical scavenging capacity under rainfed conditions, while no significant differences in DPPH radical scavenging capacity were recorded between organic and synthetic mulch in plants irrigated once at the flowering stage. Plants grown in bare soil irrigated twice at the vegetative and flowering stages had the lowest total phenol content (4.3 mg g–1 FW) and DPPH radical scavenging capacity (21.3%) (Figure 9).
4. Discussion
Water shortage stress impairs plant growth by reducing cell viability, development, division, and elongation, which diminishes vegetative and reproductive growth [28], and also decreases stomatal conductance due to inhibited Rubisco activity, raising crop water potential [29]. Stress-induced decline in agronomic traits and seed yield is linked to prioritized root resource allocation under low soil moisture or reduced chlorophyll content, limiting photosynthesis [30]. Stomatal closure during moisture stress reduces gas exchange, CO2 intake, and Rubisco activity, decreasing dry matter production [31]. Supplemental irrigation enhances water and nutrient availability, boosting photosynthetic capacity, growth traits, and seed yield.
In our research, organic and synthetic mulches enhanced linseed agronomic traits and seed yield under rainfed and supplemental irrigation conditions (Figure 1, Figure 2 and Figure 3), due to increased leaf surface area and durability, encouraging light use efficiency [32]. Mulch supports crop growth and production under moisture stress by promoting nutrient availability and absorption [33].
Oil yield depends on oil percentage and oilseed yield, both influenced by various factors. Reduced oil percentage under water stress in linseed was recorded by Zare et al. [34]. On the contrary, organic and synthetic mulches reportedly increase oil percentage by fostering leaf water potential, transpiration, CO₂ assimilation, hormone synthesis, root development, and water uptake under stress [35]. Hasanvand et al. [36] noted similar benefits in sunflowers, with mulches enhancing nutrient and water absorption, and photosynthetic products, thus boosting oil percentage and yield [37]. Carbohydrates are crucial for fatty acid biosynthesis and, therefore, may increase oil percentage and quality by translocation from leaves to seeds [38]. Organic and synthetic mulches enhance beneficial microorganisms, improve soil structure, water retention, and nutrient uptake, plant growth regulators, enzyme activity, and boosting growth and photosynthesis under drought stress increasing fatty acid quantity and quality, and oil percentage and seed yield in linseed [33,39].
In our research, applying mulches under rainfed and supplemental irrigation enhanced linseed oil yield by fostering soil and plant growth conditions. In this respect, straw mulch under two irrigations at vegetative and flowering stages increased linseed oil percentage as well as fatty acids like linoleic and oleic acids, compared to rainfed conditions, considered that efficient use of water, nutrients, and radiation is vital for optimizing growth and oil production. [33,39].
In the present study, mulch, particularly with straw, significantly improved RWC (Figure 5) by enhancing root development, reducing evaporation/transpiration, and increasing leaf water potential and water uptake [6]. Indeed, water stress reduces RWC by limiting root water absorption and increasing leaf transpiration, causing stomatal closure [40] and, in addition, declining turgor and RWC signal stress affect cell division and size [9].
In our research, water deficit during growth reduced chlorophyll a, b, and carotenoids (Figure 6), considered that water is essential for active chloroplasts and chlorophyll functions like energy absorption and transfer [41]. Drought-induced chlorophyll reduction is linked to water deficiency, impacting chlorophyll decomposition and oxidative damage, causing lipid, protein, and pigment breakdown [42]. Decreased chlorophyll under water stress is the consequence of reduced chloroplast membrane stability and enzyme activity, lowering assimilation and performance [7,39].
In the present investigation, supplemental irrigation in mulched soil conditions at flowering and vegetative stages increased chlorophyll content, enhancing photosynthesis (Figure 6), consistently with Gholami and Zahedi [43] reports. Straw mulch improves chlorophyll content and photosynthesis by enhancing soil properties like water content, energy, bulk density, and nutrients, supporting crop growth [7].
Soluble sugars, derived from carbon reserve hydrolysis, accumulate under water shortage which impacts growth and then photosynthesis, leading to a buildup of photosynthetic products [41,44], with sucrose acting as a signaling molecule, reducing ROS levels [42]. Soluble sugars regulate osmosis, lower water potential, retain cellular water, and maintain turgor, enhancing drought resistance [9]. Higher soluble sugar levels in plants grown in organically and synthetically mulched soils promote vegetative growth, improve osmotic adjustment, and maintain carbon balance under dehydration stress [8,43]. Sugar increase under stress also aids osmotic potential and cell membrane integrity [41].
Proline, a key organic osmolyte, accumulates under water stress, protecting cytosolic enzymes and cell structures [44] and, indeed, it is a primary osmotic adjustment mechanism, enhancing stress tolerance [9]. Its accumulation depends on factors such as ABA regulation of light-dependent proline synthesis processes and the influence of energy-rich photosynthetic compounds [45].
In the present study, plants grown in mulched soil and irrigated twice at vegetative stages and flowering managed nutrients and water more efficiently, reducing proline accumulation under stress (Figure 7) [8]. Organic and synthetic mulches improve root branching, soil organic matter, microbial activity, and cation exchange capacity, lowering proline levels [6].
In our research, CAT and SOD enzyme activity increased in linseed under water stress, reflecting an antioxidant defense response (Figure 8). In this respect, water shortage triggers antioxidant activation, which is strongly correlated with stress tolerance [40,45]. Enhanced antioxidant activity protects plants against biotic and abiotic stress by detoxifying ROS, which can damage lipids, nucleic acids, and membranes [6,8]. SOD initiates ROS defense by converting O2‒ into H2O2 and O2, while CAT neutralizes H2O2 into H2O excess, preventing cellular damage [46,47]. Organic and synthetic mulches reportedly boosts CAT and SOD activity in linseed by reducing soil moisture content, mitigating oxidative stress, maintaining ROS balance, and protecting cell membranes under severe water deficit [6,8].
In our study, organic and synthetic mulches increased total phenol content and DPPH radical scavenging capacity in both irrigation systems (Figure 9). Phenolic compounds, secondary metabolites, scavenge free radicals and protect cellular structures from lipid oxidation [17]. Mulches enhance antioxidant activity against ROS, boosting phenolic synthesis, which is beneficial under water deficit conditions [48,49]. They also improve photosynthesis, starch biosynthesis, enzyme activity, and secondary metabolite production [50]. Increased hydrocarbon concentrations support phenolic accumulation, likely due to greater carbon allocation to the shikimate pathway [51]. The mentioned compounds protect against ROS damage caused by environmental stress [52,53], under safety and health perspective [54,55].
5. Conclusions
Water deficit stress adversely affected various agronomic and physiological traits in linseed plants, including seed yield, biological yield, RWC, chlorophyll a, chlorophyll b, and carotenoid content; however, it increased proline and total phenol concentrations, antioxidant capacity, and CAT and SOD activity. Straw mulch effectively mitigated the adverse impacts of moisture shortage stress on linseed plants by increasing seed yield, biological yield, RWC, chlorophyll a, chlorophyll b, and carotenoid contents, osmotic adjustment, and antioxidant activity, and stimulating oilseed production, though also synthetic mulch had positive effects. The positive responses observed under organic and synthetic mulches indicate their potential to help linseed plants withstand water stress. The plastic and straw mulch applications under rainfed conditions or with one irrigation at the flowering stage improved linseed fixed oil quality and enzymatic and non-enzymatic activities. Within the examined mulch treatments, straw is recommended as a valuable source of beneficial substances for successful L. usitatissimum management, also reducing the harmful consequences of moisture shortage stress and providing a sustainable solution for linseed production, even under rainfed conditions.
Author Contributions
Conceptualization, EM, ERC, AD, and KHMS; methodology, EM, ERC, OCM, MS, and GC; software, ERC, AD, and MS; validation, EM, ERC, AD, OCM, GC, and KHMS; formal analysis, EM and AD; investigation, EM and ERC; resources, ERC and AD; data curation, EM and AD; writing—original draft preparation, EM and AD; writing—review and editing, ERC, AD, OCM, GC and KHMS; supervision, ERC; project administration, ERC and KHMS.
Funding
This research received no specific grants from public, commercial, or not-for-profit funding agencies.
Ethical Approval
This article contains no studies with human participants or animals performed by any author.
Data Availability Statement
The data supporting this study’s findings are available on request from the corresponding author.
Conflict of Interest
The authors declare that they have no conflict of interest.
References
- Chugh, V.; Kaur, D.; Purwar, S.; Kaushik, P.; Sharma, V.; Kumar, H.; Rai, A.; Singh, C.M.; Kamaluddin Dubey, R.B. Applications of molecular markers for developing abiotic-stress-resilient oilseed crops. Life 2023, 13, 8. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Zou, Y.; Tian, R.; Huang, X.; Liu, L.; Wang, B.; Peng, D. Transcriptome analysis of fiber development under high-temperature stress in flax (Linum usitatissimum L.). Ind. Crops Prod. 2023, 195, 116019. [Google Scholar] [CrossRef]
- Paliwal, S.; Tripathi, M.K.; Tiwari, S.; Tripathi, N.; Payasi, D.K.; Tiwari, P.N.; Singh, K.; Yadav, R.K.; Asati, R.; Chauhan, S. Molecular advances to combat different biotic and abiotic stresses in linseed (Linum usitatissimum L.): A comprehensive review. Genes 2023, 14, 1461. [Google Scholar] [CrossRef]
- Hajibabaie, F.; Abedpoor, N.; Safavi, K.; Taghian, F. Natural remedies medicine derived from flaxseed (secoisolariciresinol diglucoside, lignans, and α-linolenic acid) improve network targeting efficiency of diabetic heart conditions based on computational chemistry techniques and pharmacophore modeling. J. Food Biochem. 2022, 46, 14480. [Google Scholar] [CrossRef]
- Wang, N.; Qi, F.; Wang, F.; Lin, Y.; Xiaoyang, C.; Peng, Z.; Zhang, B.; Qi, X.; Deyholos, M.K.; Zhang, J. Evaluation of differentially expressed genes in leaves vs. roots subjected to drought stress in flax (Linum usitatissimum L.). Int. J. Mol. Sci. 2023, 24, 12019. [Google Scholar] [CrossRef] [PubMed]
- Behzadnejad, J.; Tahmasebi-Sarvestani, Z.; Aein, A.; Mokhtassi-Bidgoli, A. Wheat straw mulching helps improve yield in sesame (Sesamum indicum L.) under drought stress. Int. J. Plant Prod. 2020, 14, 389–400. [Google Scholar] [CrossRef]
- Lahmod, N.R.; Alkooranee, J.T.; Alshammary, A.A.G.; Rodrigo-Comino, J. Effect of wheat straw as a cover crop on the chlorophyll, seed, and oilseed yield of Trigonella foeunm graecum L under water deficiency and weed competition. Plants 2019, 8, 503. [Google Scholar] [CrossRef]
- Ahmad, S.; Zaheer, M.S.; Ali, H.H.; Erinle, K.O.; Wani, S.H.; Iqbal, R.; Okone, O.G.; Raza, A.; Waqas, M.M.; Nawaz, M. Physiological and biochemical properties of wheat (Triticum aestivum L.) under different mulching and water management systems in the semi-arid region of Punjab, Pakistan. Arid. L. Res. Manag. 2022, 36, 181–196. [Google Scholar] [CrossRef]
- Gholami, R.; Fahadi Hoveizeh, N.; Zahedi, S.M.; Arji, I. Effect of organic and synthetic mulches on some morpho-physiological and yield parameters of ‘Zard’ olive cultivar subjected to three irrigation levels in field conditions. South African J. Bot. 2023, 162, 749–760. [Google Scholar] [CrossRef]
- Rezaei-Chiyaneh, E.; Seyyedi, S.M.; Ebrahimian, E.; Moghaddam, S.S.; Damalas, C.A. Exogenous application of gamma-aminobutyric acid (GABA) alleviates the effect of water deficit stress in black cumin (Nigella sativa L.). Ind. Crops Prod. 2018, 112, 741–748. [Google Scholar] [CrossRef]
- de Aquino, G.S.; de Conti Medina, C.; Shahab, M.; Santiago, A.D.; Cunha, A.C.B.; Kussaba, D.A.O.; Carvalho, J.B.; Moreira, A. Does straw mulch partial-removal from soil interfere in yield and industrial quality sugarcane? A long-term study. Ind. Crops Prod. 2018, 111, 573–578. [Google Scholar] [CrossRef]
- Zheng, J.; Fan, J.; Zhou, M.; Zhang, F.; Liao, Z.; Lai, Z.; Yan, S.; Guo, J.; Li, Z.; Xiang, Y. Ridge-furrow plastic film mulching enhances grain yield and yield stability of rainfed maize by improving resources capture and use efficiency in a semi-humid drought-prone region. Agric. Water Manag. 2022, 269, 107654. [Google Scholar] [CrossRef]
- Chai, Y.; Chai, Q.; Li, R.; Li, Y.; Yang, C.; Cheng, H.; Chang, L.; Chai, S. Straw strip mulching in a semiarid rainfed agroecosystem achieves winter wheat yields similar to those of full plastic mulching by optimizing the soil hydrothermal regime. Crop J. 2022, 10, 879–892. [Google Scholar] [CrossRef]
- Rezaei-Chiyaneh, E.; Mahdavikia, H.; Hadi, H.; Muhittin, K.; Caruso, G.; Siddique, K.H.M. The effect of exogenously applied plant growth regulators and zinc on some physiological characteristics and essential oil constituents of Moldavian balm (Dracocephalum moldavica L.) under water stress. Physiol. Mol. Biol. Plants 2021, 27, 2201–2214. [Google Scholar] [CrossRef] [PubMed]
- Walker, W.R. Irrigation engineering — Sprinkler, trickle, surface irrigation principles, design and agricultural practices. Agric. Water Manag. 1984, 9, 263–264. [Google Scholar] [CrossRef]
- AOCS. Official methods and recommended practices of the American oil. 4th Ed. Soc Champaign, IL, 1993; 6–38. [Google Scholar]
- Zamani, F.; Amirnia, R.; Rezaei-Chiyaneh, E.; Gheshlaghi, M.; von Cossel, M.; Siddique, K.H.M. Optimizing essential oil, fatty acid profiles, and phenolic compounds of dragon’s head (Lallemantia iberica) intercropped with chickpea (Cicer arietinum L.) with biofertilizer inoculation under rainfed conditions in a semi-arid region. Arch. Agron. Soil Sci. 2022, 69, 1687–1704. [Google Scholar] [CrossRef]
- Taghizadeh, Y.; Amirnia, R.; Rezaei-Chiyaneh, E.; Ghiyasi, M.; Razavi, B.S.; Siddique, K.H.M. Co-Inoculation of mycorrhizal fungi with bacterial fertilizer along with intercropping scenarios improves seed yield and oil constituents of sesame. J. Soil Sci. Plant Nutr. 2023, 23, 2258–2272. [Google Scholar] [CrossRef]
- Saadat, B.; Pirzad, A.; Jalilian, J. Yield-related biochemical response of understory mycorrhizal yellow sweet clover (Melilotus officinalis L.) to drought in agrisilviculture. Arch. Agron. Soil Sci. 2020, 67, 1603–1620. [Google Scholar] [CrossRef]
- Lichtenthaler, H. Determination of total carotenoids and chlorophylls a and b of leaf in different solvents. Biochem. Soc. Trans. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Tejera, N.A.; Campos, R.; Sanjuan, J.; Lluch, C. Nitrogenase and antioxidant enzyme activities in Phaseolus vulgaris nodules formed by Rhizobium tropid isogenic strains with varying tolerance to salt stress. J. Plant Physiol. 2004, 161, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Beyer, W.F.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.; Liu, R.H.; Nock, J.F.; Holliday, D.; Watkins, C.B. Temperature and relative humidity effects on quality, total ascorbic acid, phenolics and flavonoid concentrations, and antioxidant activity of strawberry. Postharvest Biol. Technol. 2007, 45, 349–357. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT - Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Rezaei-Chiyaneh, E.; Mahdavikia, H.; Alipour, H.; Dolatabadian, A.; Battaglia, L.M.; Harrison, T.M. Biostimulants alleviate water deficit stress and enhance essential oil productivity: a case study with savory. Sci. Rep. 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- Javaid, M.M.; Algwaiz, H.I.M.; Waheed, H.; Ashraf, M.; Mahmood, A.; Li, F.M.; Attia, K.A.; Nadeem, M.A.; AlKahtani, M.D.; Fiaz, S.; Nadeem, M. Ridge-furrow mulching enhances capture and utilization of rainfall for improved maize production under rain-fed conditions. Agronomy 2022, 12, 1187. [Google Scholar] [CrossRef]
- Qu, Y.; Feng, B.L. Straw mulching improved yield of field buckwheat (Fagopyrum) by increasing water-temperature use and soil carbon in rain-fed farmland. Acta Ecol. Sin. 2022, 42, 11–16. [Google Scholar] [CrossRef]
- Chai, Y.; Chai, Q.; Yang, C.; Chen, Y.; Li, R.; Li, Y.; Chang, L.; Lan, X.; Cheng, H.; Chai, S. Plastic film mulching increases yield, water productivity, and net income of rain-fed winter wheat compared with no mulching in semiarid Northwest China. Agric. Water Manag. 2022, 262, 107420. [Google Scholar] [CrossRef]
- Zhang, M.; Song, D.; Pu, X.; Dang, P.; Qin, X.; Siddique, K.H. Effect of different straw returning measures on resource use efficiency and spring maize yield under a plastic film mulch system. Eur. J. Agron. 2022, 134, 126461. [Google Scholar] [CrossRef]
- Wang, N.; Chen, H.; Ding, D.; Zhang, T.; Li, C.; Luo, X.; Chu, X.; Feng, H.; Wei, Y.; Siddique, K.H. Plastic film mulching affects field water balance components, grain yield, and water productivity of rainfed maize in the Loess Plateau, China: A synthetic analysis of multi-site observations. Agric. Water Manag. 2022, 266, 107570. [Google Scholar] [CrossRef]
- Zare, S.; Mirlohi, A.; Sabzalian, M.R.; Saeidi, G.; Koçak, M.Z.; Hano, C. Water stress and seed color interacting to impact seed and oil yield, protein, mucilage, and secoisolariciresinol diglucoside content in cultivated flax (Linum usitatissimum L.). Plants 2023, 12, 1632. [Google Scholar] [CrossRef]
- Cui, Z.; Yan, B.; Gao, Y.; Wu, B.; Wang, Y.; Wang, H.; Xu, P.; Zhao, B.; Cao, Z.; Zhang, Y.; Xie, Y. Agronomic cultivation measures on productivity of oilseed flax: A review. Oil Crop Sci. 2022, 7, 53–62. [Google Scholar] [CrossRef]
- Hasanvand, R.K.; Jalal-Abadi, A.L.; Abdali Mashhadi, A.; Kochekzadeh, A.; Siyahpoosh, A. The effect of different types of mulch and different cultivation methods on the quantitative and qualitative traits of sunflower (Helianthus annuus L.) in Ahvaz climate. J. Cent. Eur. Agric. 2023, 24, 115–125. [Google Scholar] [CrossRef]
- Ye, H.; Wen, Y.; Chen, Z.; Zhang, T.; Li, S.; Guan, M.; Zhang, Y.; Su, S. Relationship of soil microbiota to seed kernel metabolism in camellia oleifera under mulched. Front. Plant Sci. 2022, 13, 920604. [Google Scholar] [CrossRef]
- Rahangdale, N.; Kumawat, N.; Jadav, M.L.; Bhagat, D.V.; Singh, M.; Yadav, R.K. Symbiotic efficiency, productivity and profitability of soybean as influenced by liquid bio-inoculants and straw mulch. Int. J. Bio-resource Stress Manag. 2022, 13, 9–16. [Google Scholar] [CrossRef]
- Kikhazaleh, M.; Ramroudi, M.; Galavi, M.; Ghanbari, S.A.; Fanaei, H.R. Effect of potassium on yield and some qualitative and physiological traits of safflower (Carthamus tinctorius L.) under drought stress conditions. J. Plant Nutr. 2023, 46, 2380–2392. [Google Scholar] [CrossRef]
- Heydarzadeh, S.; Arena, C.; Vitale, E.; Rahimi, A.; Mirzapour, M.; Nasar, J.; Kisaka, O.; Sow, S.; Ranjan, S.; Gitari, H. Impact of different fertilizer sources under supplemental irrigation and rainfed conditions on eco-physiological responses and yield characteristics of dragon’s head (Lallemantia iberica). Plants 2023, 12, 1693. [Google Scholar] [CrossRef] [PubMed]
- Maroof, T.; Tahir, M.; Zahid, N.; Khan, M.N.; Younas, M.T.; Shahzad, J.; Zafar, T.; Zeeshan, M.; Pervaiz, A.Y.A. Effect of different mulching materials on weed emergence and growth characteristics of strawberry under rainfed condition at Rawalakot Azad Jammu and Kashmir. J. App. Res. Plant Sci. 2024, 5, 46–57. [Google Scholar]
- Liao, Z.; Zeng, H.; Fan, J.; Lai, Z.; Zhang, C.; Zhang, F.; Wang, H.; Cheng, M.; Guo, J.; Li, Z.; Wu, P. Effects of plant density, nitrogen rate and supplemental irrigation on photosynthesis, root growth, seed yield and water-nitrogen use efficiency of soybean under ridge-furrow plastic mulching. Agric. Water Manag. 2022, 268, 107688. [Google Scholar] [CrossRef]
- Gholami, R.; Zahedi, S.M. Effects of deficit irrigation and mulching on morpho-physiological and biochemical characteristics of konservolia olives. Gesunde Pflanz 2020, 72, 49–55. [Google Scholar] [CrossRef]
- Zamani, F.; Amirnia, R.; Rezaei-Chiyaneh, E.; Gheshlaghi, M.; von Cossel, M.; Siddique, K.H.M. Alleviating plant water stress with biofertilizers: A case study for dragon’s head (Lallemantia iberica) and chickpea (Cicer arietinum L.) in a rainfed intercropping system. Int. J. Plant Prod. 2023, 17, 251–264. [Google Scholar] [CrossRef]
- Hussain, S.; Wang, J.J.; Asad Naseer, M.; Saqib, M.; Siddiqui, M.H.; Ihsan, F.; Xiaoli, C.; Xiaolong, R.; Hussain, S.; Ramzan, H.N. Water stress memory in wheat/maize intercropping regulated photosynthetic and antioxidative responses under rainfed conditions. Sci. Rep. 2023, 13, 13688. [Google Scholar] [CrossRef] [PubMed]
- Perveen, S.; Parvaiz, M.; Shahbaz, M.; Saeed, M.; Zafar, S. Triacontanol positively influences growth, yield, biochemical attributes and antioxidant enzymes of two linseed (Linum usitatissimum l.) accessions differing in drought tolerance. Pakistan J. Bot. 2022, 54. [Google Scholar] [CrossRef]
- Heydarzadeh, S.; Jalilian, J.; Pirzad, A.; Jamei, R.; Petrussa, E. Fodder value and physiological aspects of rainfed smooth vetch affected by biofertilizers and supplementary irrigation in an agri-silviculture system. Agrofor. Syst. 2022, 96, 221–232. [Google Scholar] [CrossRef]
- Sałata, A.; Nurzyńska-Wierdak, R.; Kalisz, A.; Moreno-Ramón, H. Impacts of alexandrian clover living mulch on the yield, phenolic content, and antioxidant capacity of leek and shallot. Agronomy 2022, 12, 2602. [Google Scholar] [CrossRef]
- Zheng, Z.; Zhou, W.; Yang, Q.; Jiang, J.; Miao, M. Effect of furrow straw mulching and straw decomposer application on celery (Apium graveolens L.) production and soil improvement. Agronomy 2023, 13, 2774. [Google Scholar] [CrossRef]
- Paunović, S.M.; Mašković, P. Phenolic compounds, antioxidant and cytotoxic activity in berry and leaf extracts of black currant (Ribes nigrum L.) as affected by soil management systems. Erwerbs-Obstbau 2020, 62, 293–300. [Google Scholar] [CrossRef]
- Sałata, A.; Sękara, A.; Pandino, G.; Mauromicale, G.; Lombardo, S. Living mulch as sustainable tool to improve leaf biomass and phytochemical yield of Cynara cardunculus var. altilis. Agronomy 2023, 13, 1274. [Google Scholar] [CrossRef]
- Murariu, O.C.; Lipsa, F.D.; Cârlescu, P.M.; Frunză, G.; Ciobanu, M.M.; Cara, I.G.; Murariu, F.; Stoica, F.; Albu, A.; Tallarita, A.V.; et al. The Effect of Including Sea Buckthorn Berry By-Products on White Chocolate Quality and Bioactive Characteristics under a Circular Economy Context. Plants 2024, 13, 2799. [Google Scholar] [CrossRef]
- Irimia, L.M.; Patriche, C.V.; Murariu, O.C. The imparct of climate change on viticultural potential and wine grape varieties of a temperate wine growing region. Appl. Ecol. Environ. Res. 2018, 16(3), 2663–2680. [Google Scholar] [CrossRef]
- Murariu, F.; Voda, A.D.; Murariu, O.C. Researches on food safety assessment—Supporting a healthy lifestyle for the population from NE of Romania. J. Biotechnol. 2019, 305S, S68. [Google Scholar] [CrossRef]
- Mineată, I.; Murariu, O.C.; Sîrbu, S.; Tallarita, A.V.; Caruso, G.; Jităreanu, C.D. Effects of Ripening Phase and Cultivar under Sustainable Management on Fruit Quality and Antioxidants of Sweet Cherry. Horticulturae 2024, 10, 720. [Google Scholar] [CrossRef]
Figure 1.
Effect of mulch type and irrigation regime on linseed plant height, branch number, and follicle number. RFD: rainfed condition; SIF: one irrigation at the flowering stage; SIVF: two irrigations at the vegetative and flowering stages. Lowercase letters above the bars indicate significant differences between treatments at p ≤ 0.05.Plants irrigated during the vegetative and flowering stages showed the highest number of branches (5.9), those under rainfed conditions the lowest one (4.9) not significantly different from that recorded under one irrigation during the flowering stage (Figure 1). Among the mulching treatments, straw elicited 5.5 branches issue not significantly different from vermicompost but higher than the control (4.8) (Figure 1).
Figure 1.
Effect of mulch type and irrigation regime on linseed plant height, branch number, and follicle number. RFD: rainfed condition; SIF: one irrigation at the flowering stage; SIVF: two irrigations at the vegetative and flowering stages. Lowercase letters above the bars indicate significant differences between treatments at p ≤ 0.05.Plants irrigated during the vegetative and flowering stages showed the highest number of branches (5.9), those under rainfed conditions the lowest one (4.9) not significantly different from that recorded under one irrigation during the flowering stage (Figure 1). Among the mulching treatments, straw elicited 5.5 branches issue not significantly different from vermicompost but higher than the control (4.8) (Figure 1).
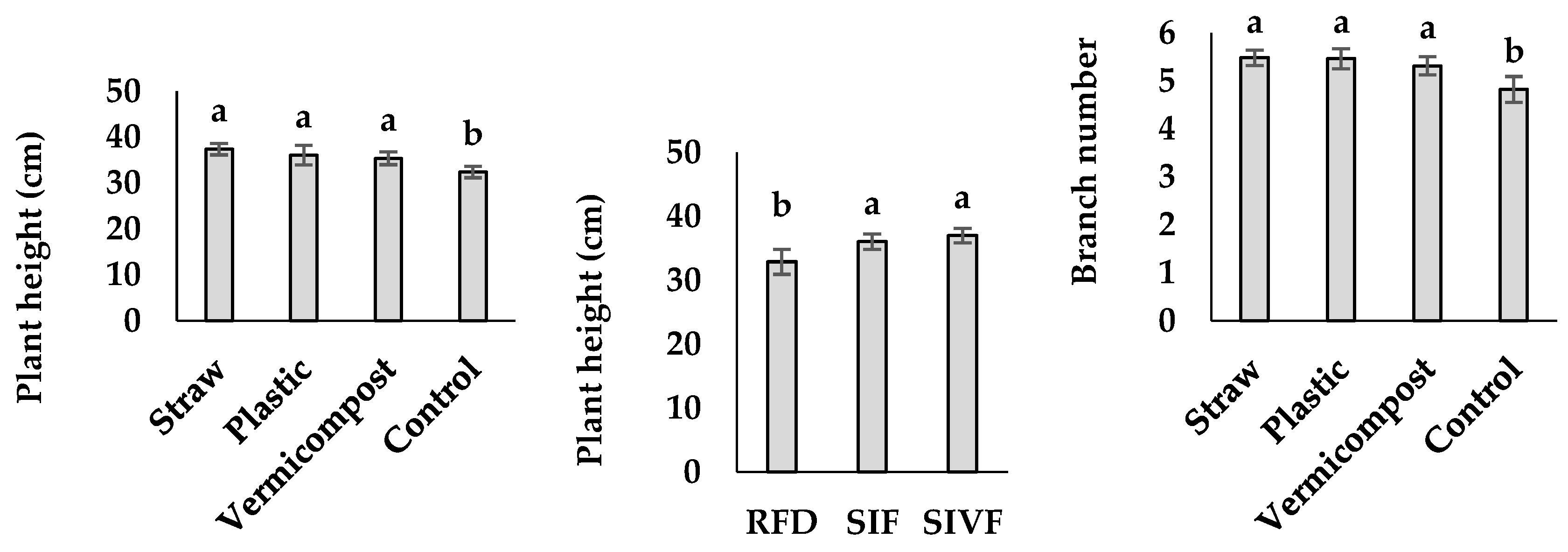
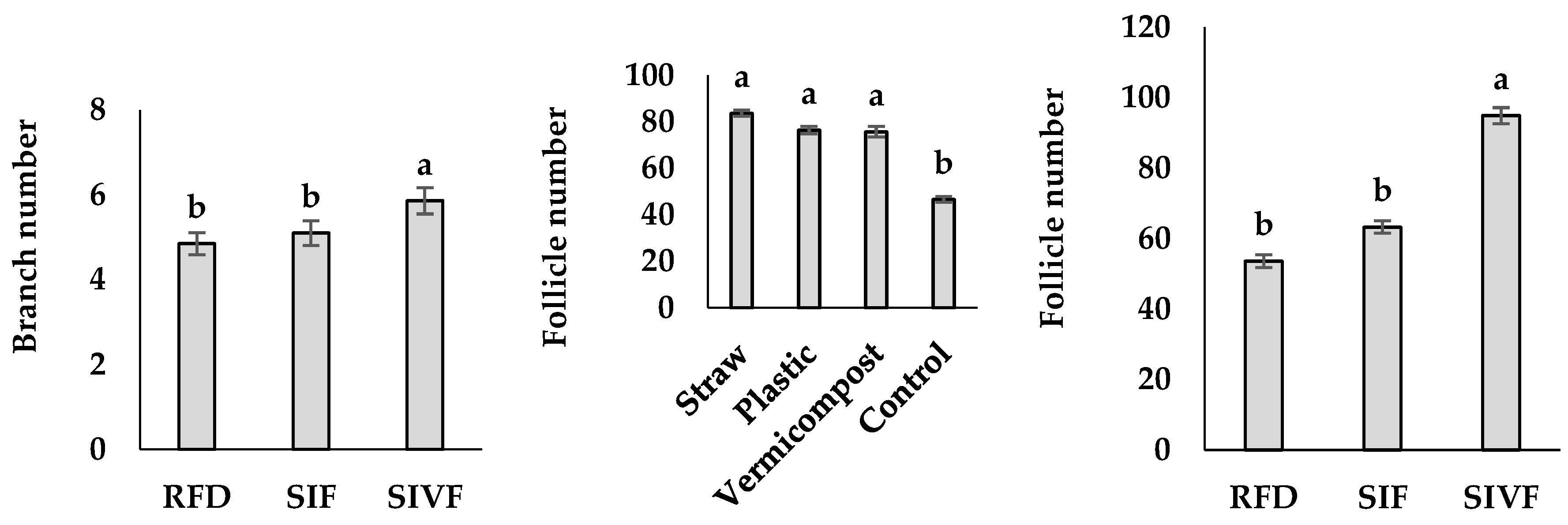
Figure 2.
Interaction between mulch type and irrigation regime on follicle seed number and 100-seed weight in linseed. RFD: rainfed condition; SIF: one supplemental irrigation in the flowering stage; SIVF: two supplemental irrigations in the vegetative and flowering stages. Lowercase letters above the bars indicate significant differences between treatments at p ≤ 0.05.
Figure 2.
Interaction between mulch type and irrigation regime on follicle seed number and 100-seed weight in linseed. RFD: rainfed condition; SIF: one supplemental irrigation in the flowering stage; SIVF: two supplemental irrigations in the vegetative and flowering stages. Lowercase letters above the bars indicate significant differences between treatments at p ≤ 0.05.
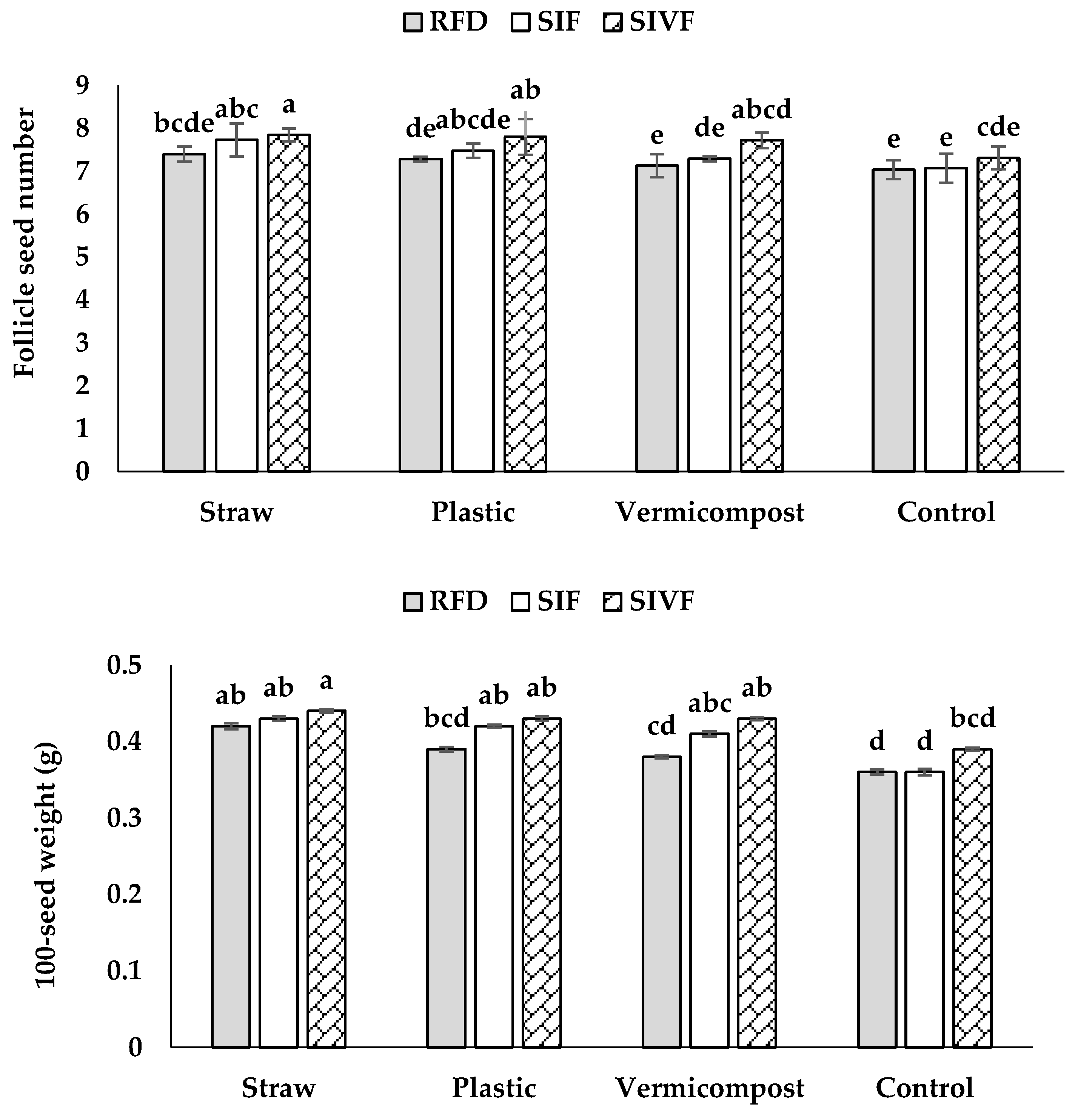
Figure 3.
Main effects of mulch application and irrigation conditions on linseed seed and biological yields. RFD: rainfed condition; SIF: one supplemental irrigation in the flowering stage; SIVF: two supplemental irrigations in the vegetative and flowering stages. Lowercase letters above the bars indicate significant treatment differences (p ≤ 0.05).
Figure 3.
Main effects of mulch application and irrigation conditions on linseed seed and biological yields. RFD: rainfed condition; SIF: one supplemental irrigation in the flowering stage; SIVF: two supplemental irrigations in the vegetative and flowering stages. Lowercase letters above the bars indicate significant treatment differences (p ≤ 0.05).
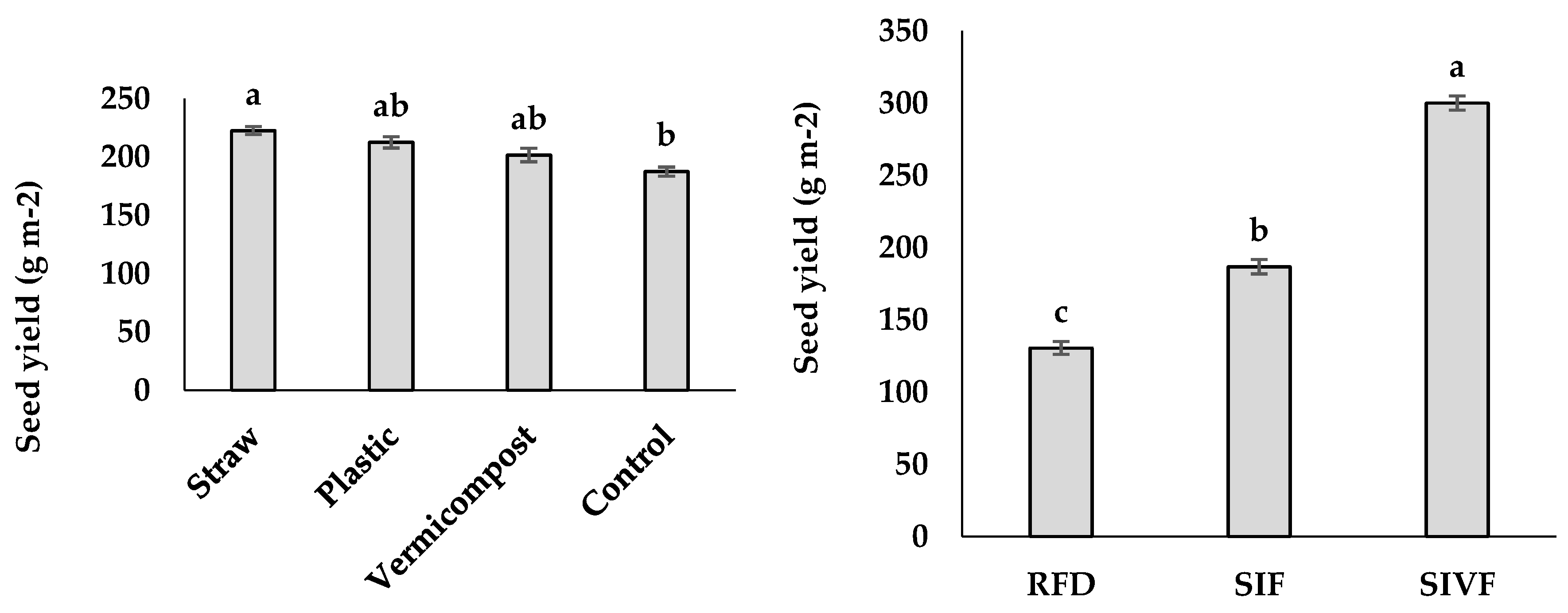
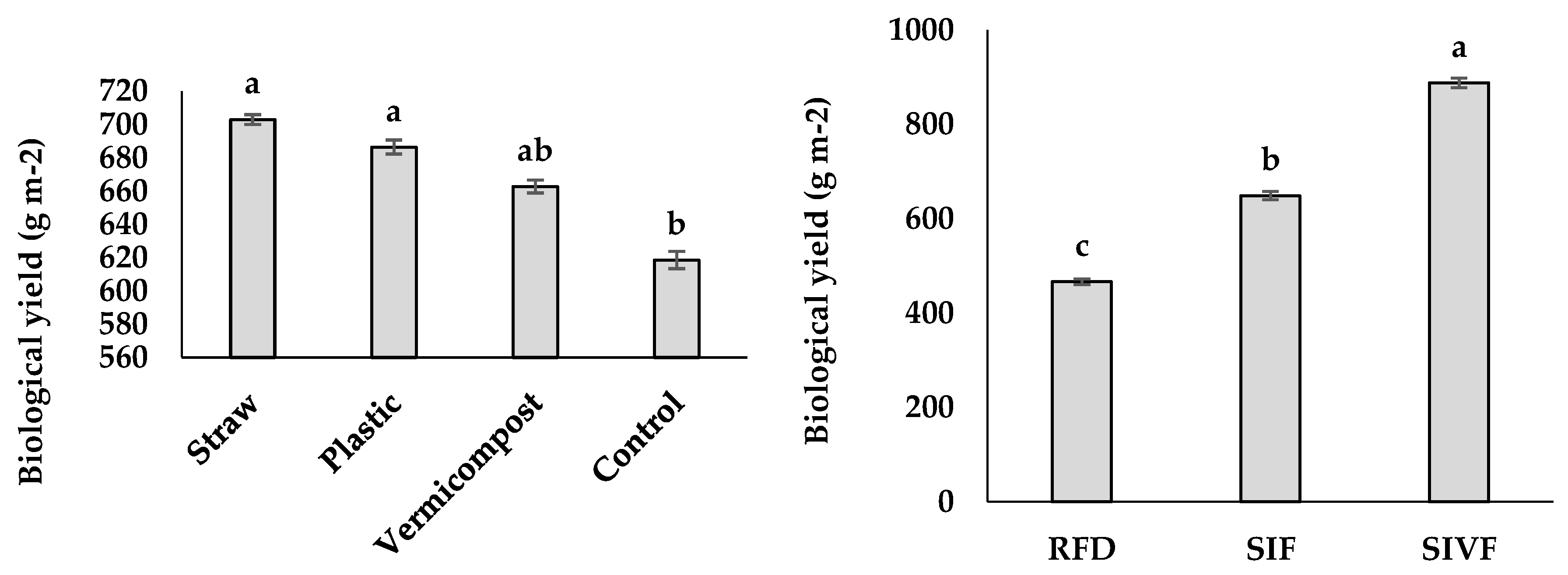
Figure 4.
Main effects of mulch application and irrigation regime on oilseed content and yield in linseed. RFD: rainfed condition; SIF: one supplemental irrigation in the flowering stage; SIVF: two supplemental irrigations in the vegetative and flowering stages. Lowercase letters above the bars indicate significant differences between treatments at p ≤ 0.05.The GC-MS and GC-FID analysis allowed to identify the most significant fatty acid components in linseed oil: oleic acid (23.7–21.5%), linoleic acid (12.9–10.7%), palmitic acid (7.4–4.7%), and stearic acid (5.8–5.0%) (Table 2). Control plants showed the highest oleic acid content (23.7%) when irrigated only at the flowering stage and the lowest (21.5%) under rainfed conditions (Table 2). Plants irrigated twice at the vegetative and flowering stages had the highest linoleic acid content (12.9%) when grown in straw mulched soil and the lowest (10.7%) in bare soil (Table 2). Plastic mulch and one irrigation at the flowering stage elicited the highest palmitic acid content (7.4%), while the lowest value was recorded with straw mulch and two irrigations (4.7%) (Table 2). Plants grown under plastic mulch and rainfed conditions had the highest stearic acid content (5.8%) and those undergone straw mulch and two irrigations the lowest (5.0%) (Table 2).
Figure 4.
Main effects of mulch application and irrigation regime on oilseed content and yield in linseed. RFD: rainfed condition; SIF: one supplemental irrigation in the flowering stage; SIVF: two supplemental irrigations in the vegetative and flowering stages. Lowercase letters above the bars indicate significant differences between treatments at p ≤ 0.05.The GC-MS and GC-FID analysis allowed to identify the most significant fatty acid components in linseed oil: oleic acid (23.7–21.5%), linoleic acid (12.9–10.7%), palmitic acid (7.4–4.7%), and stearic acid (5.8–5.0%) (Table 2). Control plants showed the highest oleic acid content (23.7%) when irrigated only at the flowering stage and the lowest (21.5%) under rainfed conditions (Table 2). Plants irrigated twice at the vegetative and flowering stages had the highest linoleic acid content (12.9%) when grown in straw mulched soil and the lowest (10.7%) in bare soil (Table 2). Plastic mulch and one irrigation at the flowering stage elicited the highest palmitic acid content (7.4%), while the lowest value was recorded with straw mulch and two irrigations (4.7%) (Table 2). Plants grown under plastic mulch and rainfed conditions had the highest stearic acid content (5.8%) and those undergone straw mulch and two irrigations the lowest (5.0%) (Table 2).
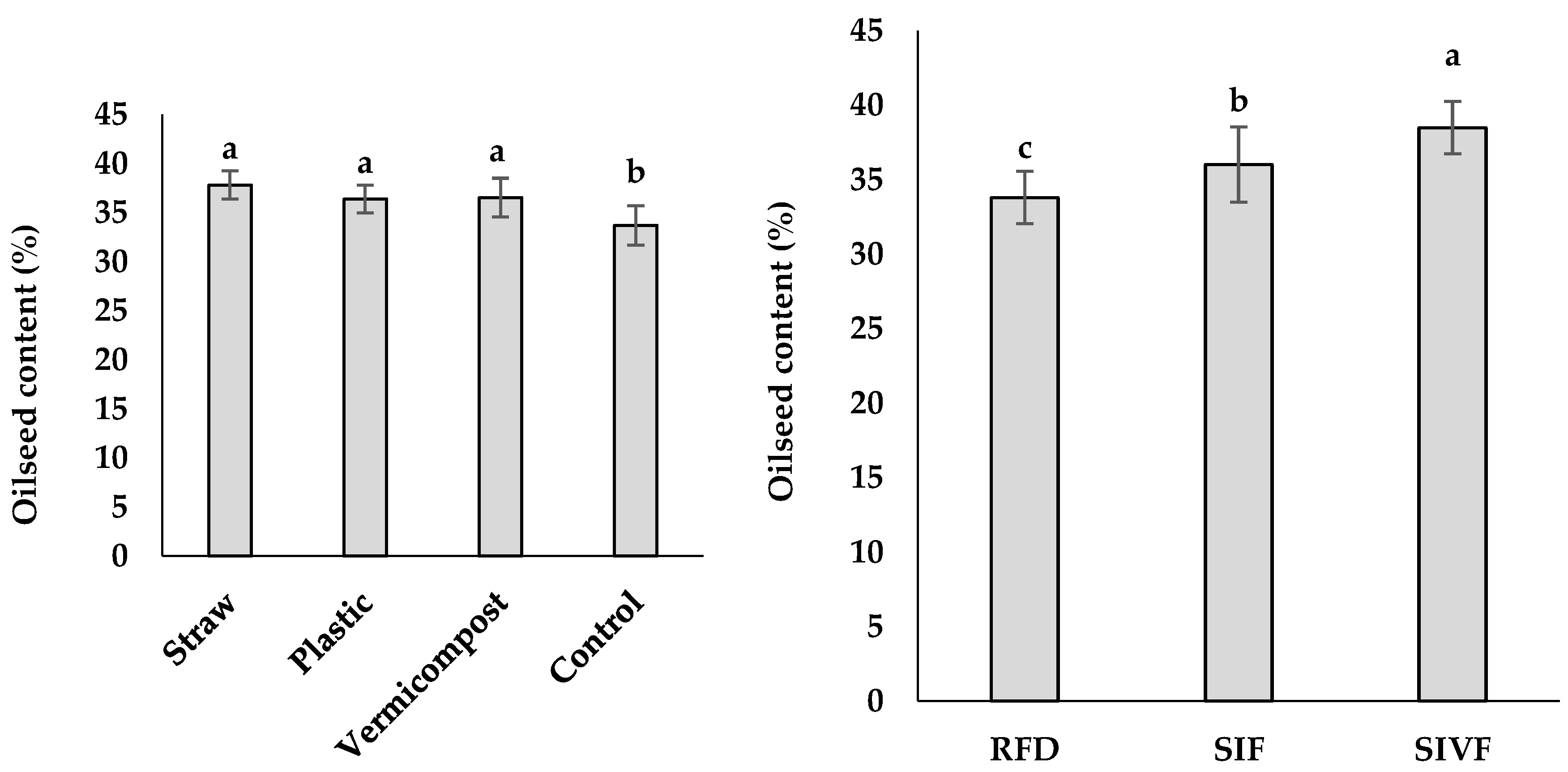
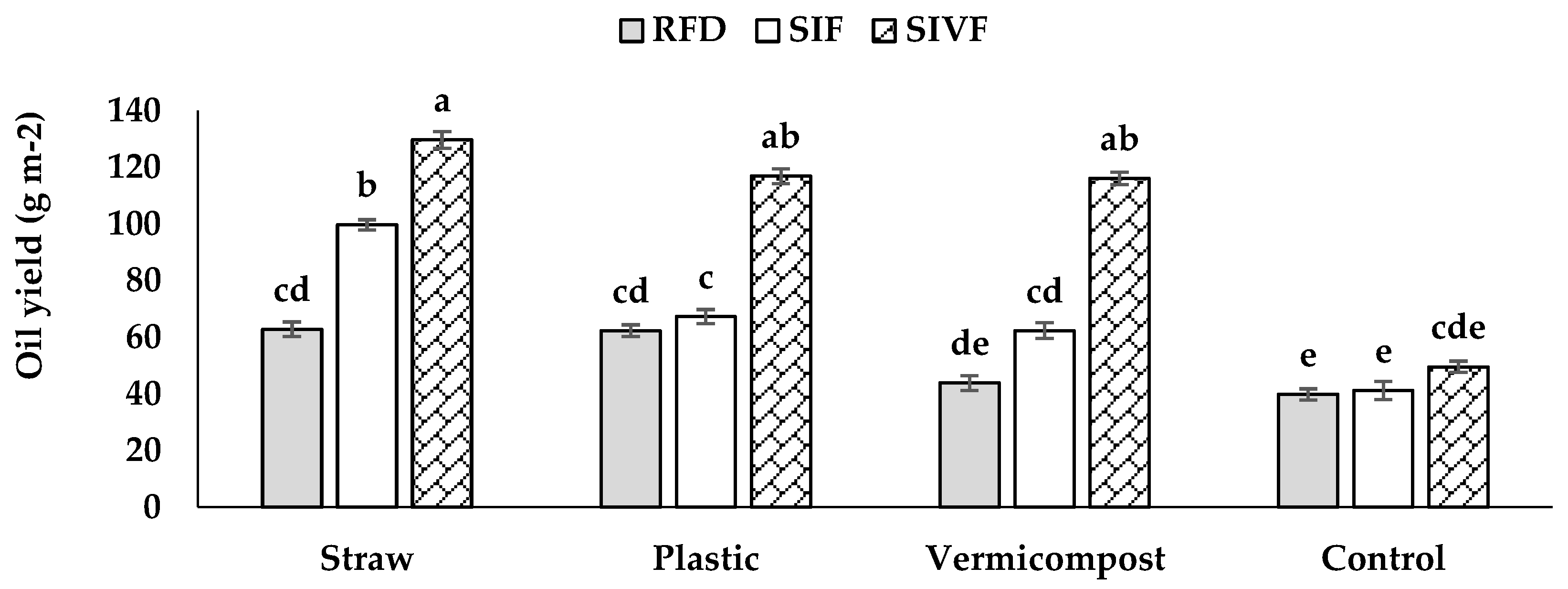
Figure 5.
Effect of mulch type and irrigation regime on relative water content in linseed. RFD: rainfed condition; SIF: one supplemental irrigation in the flowering stage; SIVF: two supplemental irrigations in the vegetative and flowering stages. Lowercase letters above the bars indicate significant differences between treatments at p ≤ 0.05.
Figure 5.
Effect of mulch type and irrigation regime on relative water content in linseed. RFD: rainfed condition; SIF: one supplemental irrigation in the flowering stage; SIVF: two supplemental irrigations in the vegetative and flowering stages. Lowercase letters above the bars indicate significant differences between treatments at p ≤ 0.05.
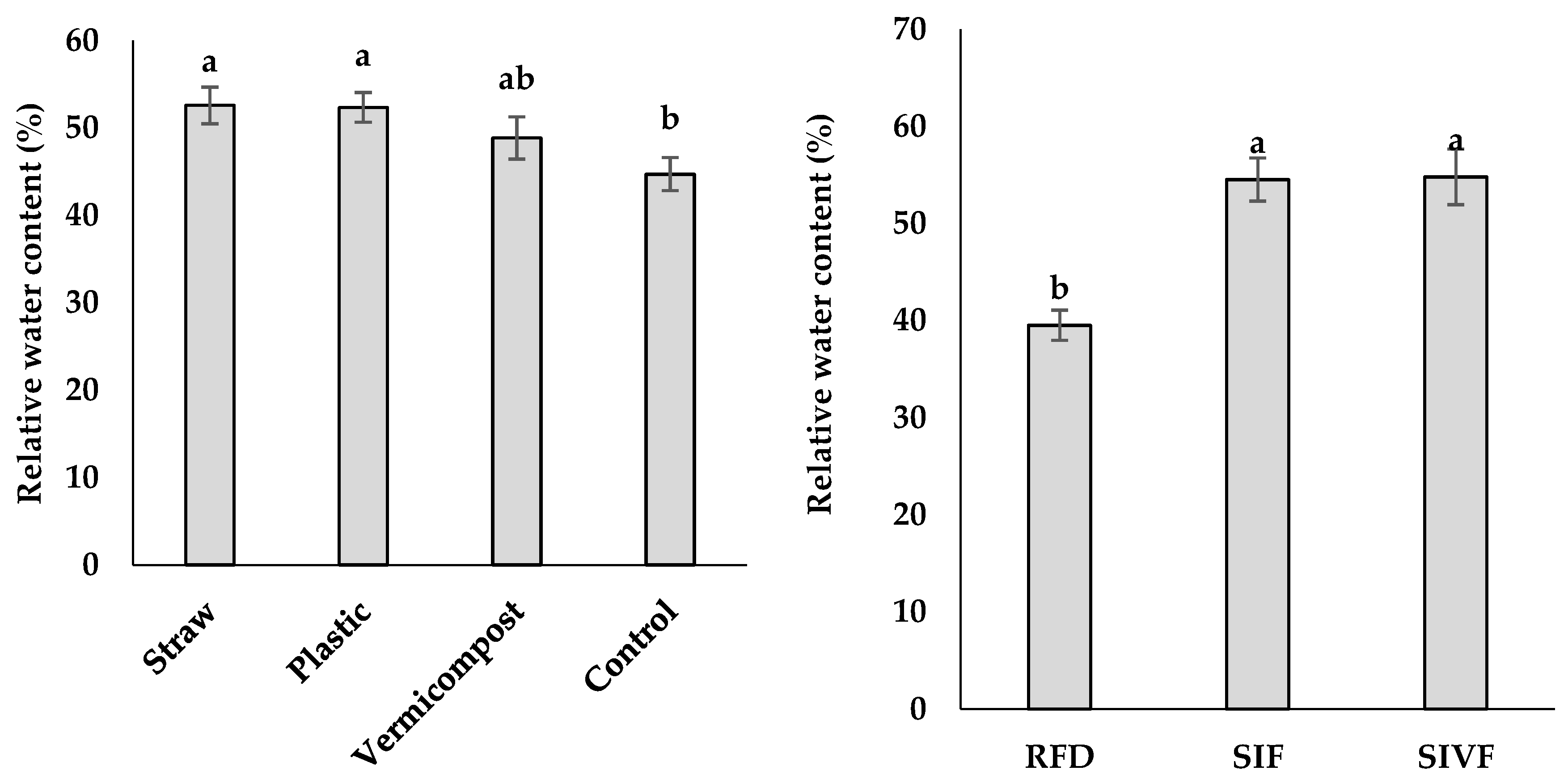
Figure 6.
Interaction between mulch type and irrigation regime on linseed chlorophyll a, b, and carotenoid contents. RFD: rainfed condition; SIF: one supplemental irrigation in the flowering stage; SIVF: two supplemental irrigations in the vegetative and flowering stages. Lowercase letters above the bars indicate significant differences between treatments at p ≤ 0.05.
Figure 6.
Interaction between mulch type and irrigation regime on linseed chlorophyll a, b, and carotenoid contents. RFD: rainfed condition; SIF: one supplemental irrigation in the flowering stage; SIVF: two supplemental irrigations in the vegetative and flowering stages. Lowercase letters above the bars indicate significant differences between treatments at p ≤ 0.05.
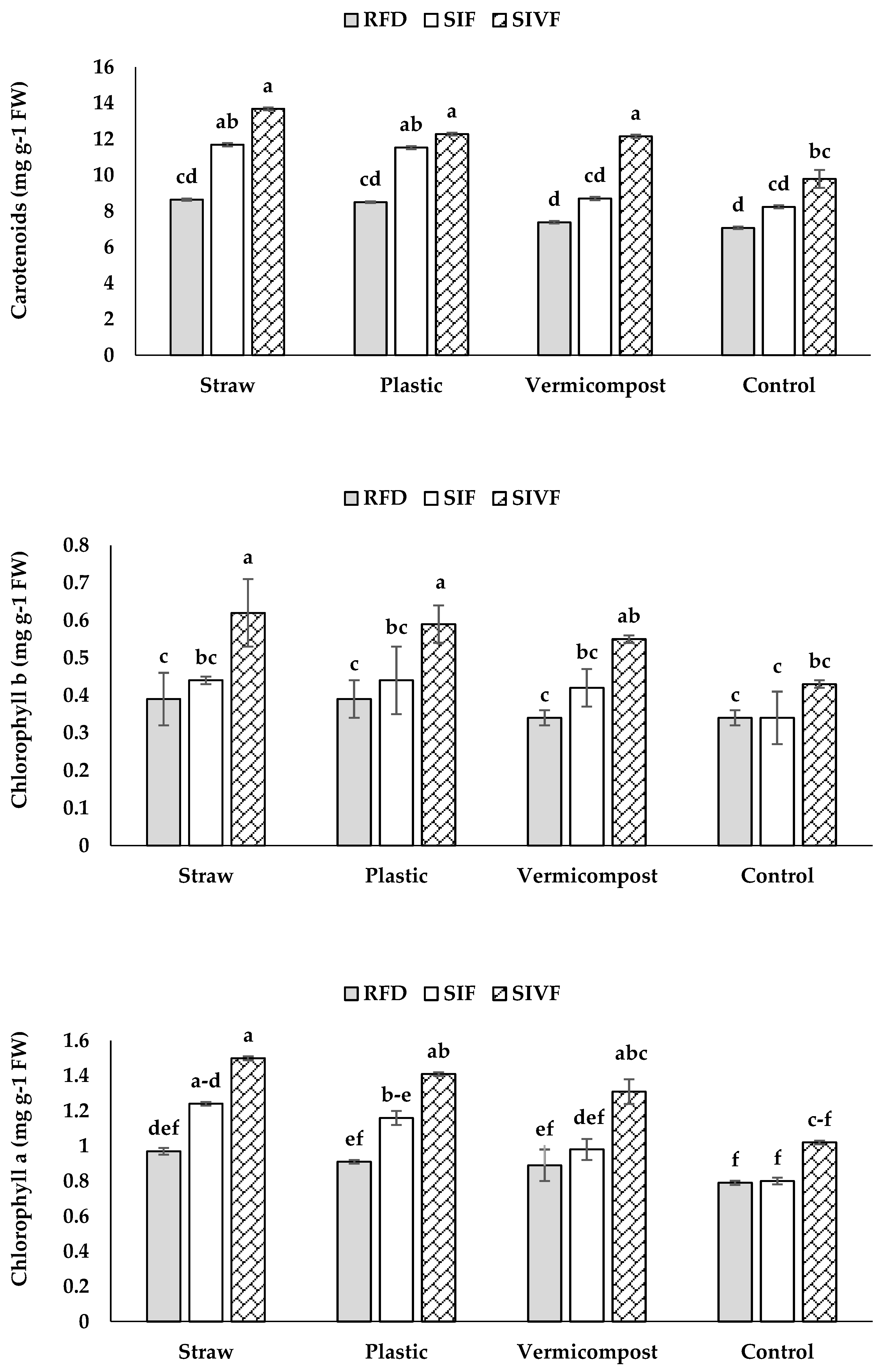
Figure 7.
Interaction between mulch type and irrigation regime on total soluble sugars and proline content in linseed. RFD: rainfed condition; SIF: one supplemental irrigation in the flowering stage; SIVF: two supplemental irrigations in the vegetative and flowering stages. Lowercase letters above the bars indicate significant differences between treatments at p ≤ 0.05.
Figure 7.
Interaction between mulch type and irrigation regime on total soluble sugars and proline content in linseed. RFD: rainfed condition; SIF: one supplemental irrigation in the flowering stage; SIVF: two supplemental irrigations in the vegetative and flowering stages. Lowercase letters above the bars indicate significant differences between treatments at p ≤ 0.05.
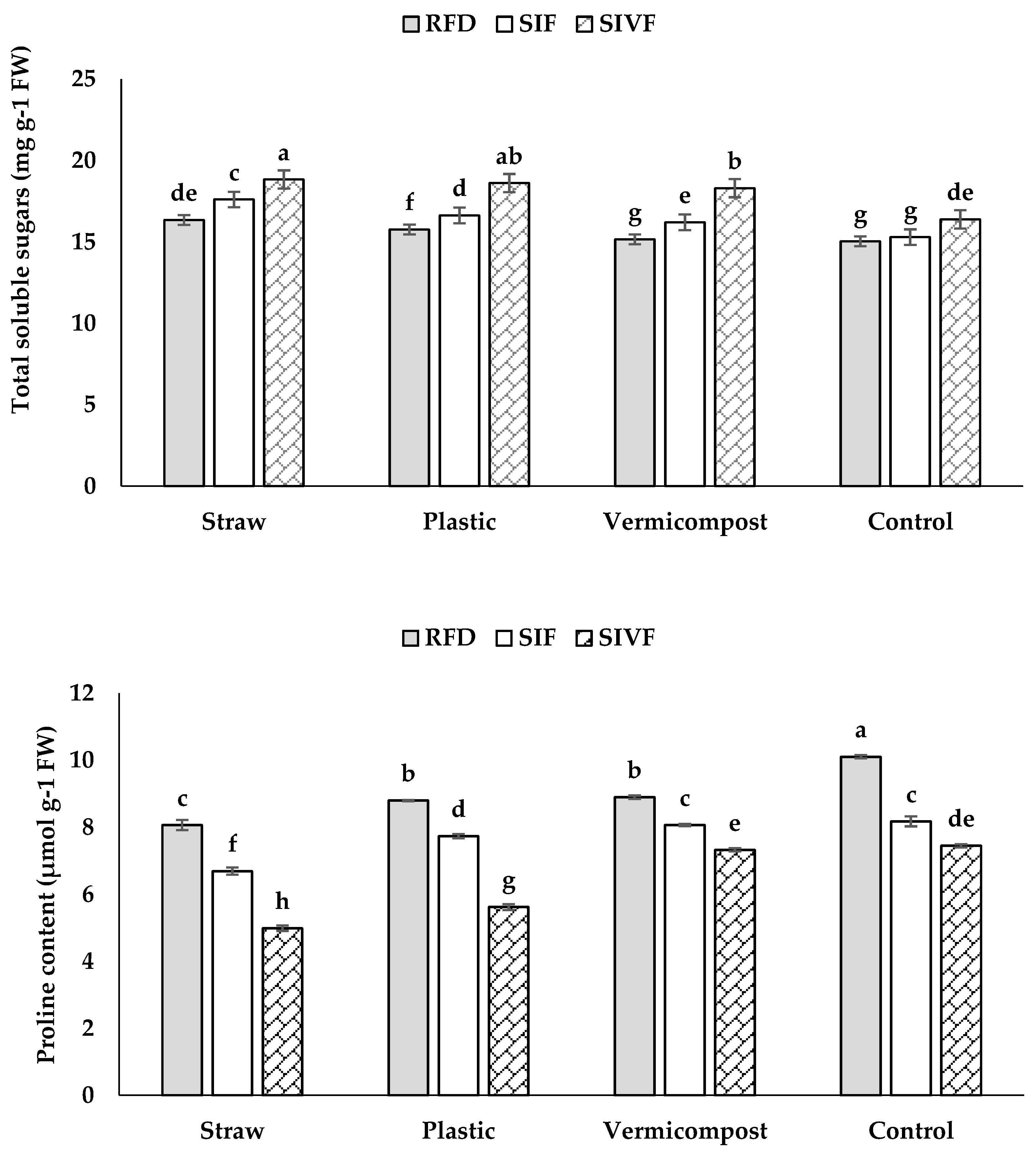
Figure 8.
Interaction between mulch type and irrigation regime on catalase (CAT) and superoxide dismutase (SOD) activities in linseed. RFD: rainfed condition; SIF: one supplemental irrigation in the flowering stage; SIVF: two supplemental irrigations in the vegetative and flowering stages. Lowercase letters above the bars indicate significant differences between treatments at p ≤ 0.05.
Figure 8.
Interaction between mulch type and irrigation regime on catalase (CAT) and superoxide dismutase (SOD) activities in linseed. RFD: rainfed condition; SIF: one supplemental irrigation in the flowering stage; SIVF: two supplemental irrigations in the vegetative and flowering stages. Lowercase letters above the bars indicate significant differences between treatments at p ≤ 0.05.
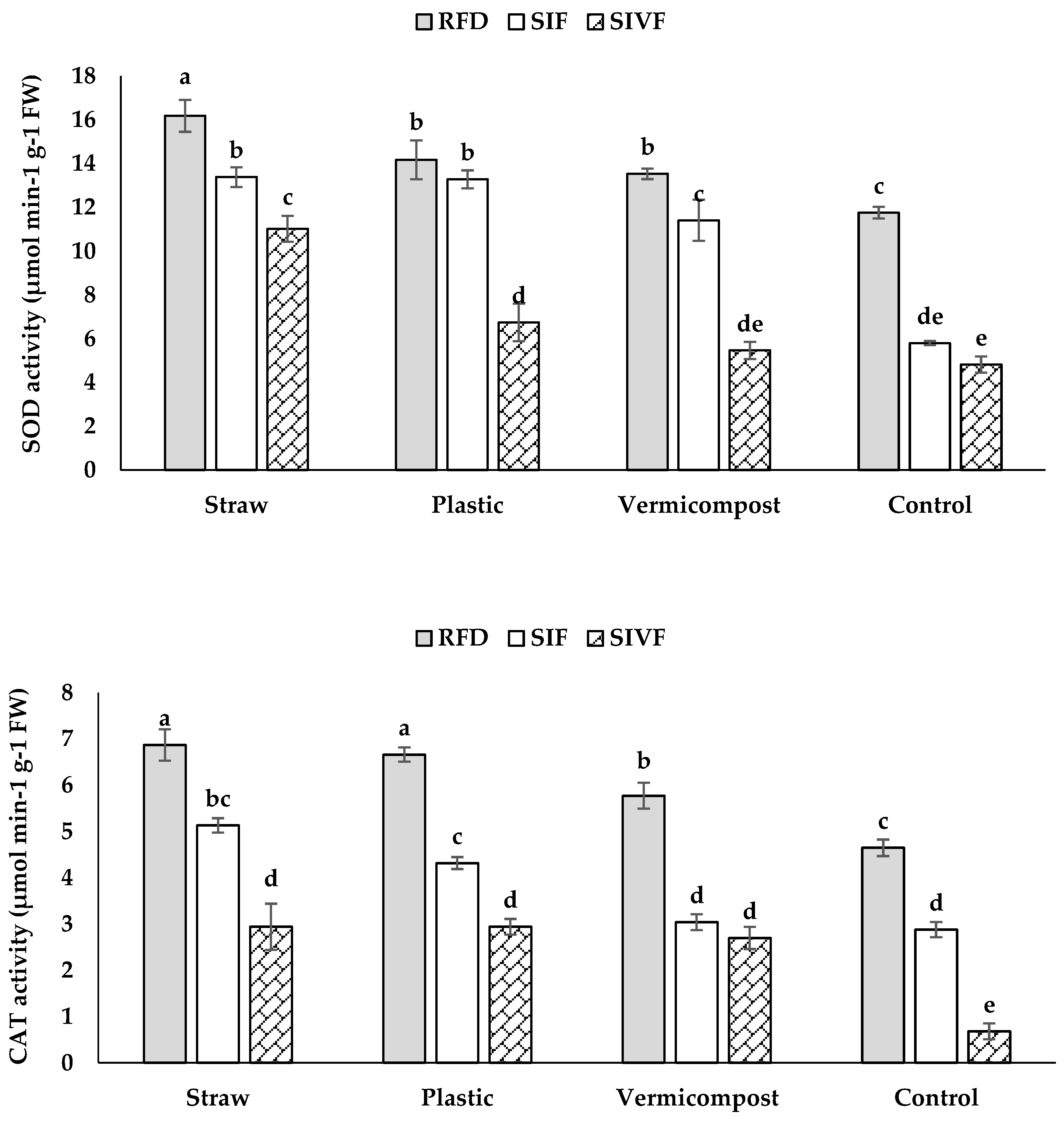
Figure 9.
Interaction between mulch type and irrigation regime on total phenol content and DPPH radical scavenging capacity of linseed. RFD: rainfed condition; SIF: one irrigation at the flowering stage; SIVF: two irrigations at the vegetative and flowering stages. Lowercase letters above the bars indicate significant differences between treatments at p ≤ 0.05.
Figure 9.
Interaction between mulch type and irrigation regime on total phenol content and DPPH radical scavenging capacity of linseed. RFD: rainfed condition; SIF: one irrigation at the flowering stage; SIVF: two irrigations at the vegetative and flowering stages. Lowercase letters above the bars indicate significant differences between treatments at p ≤ 0.05.
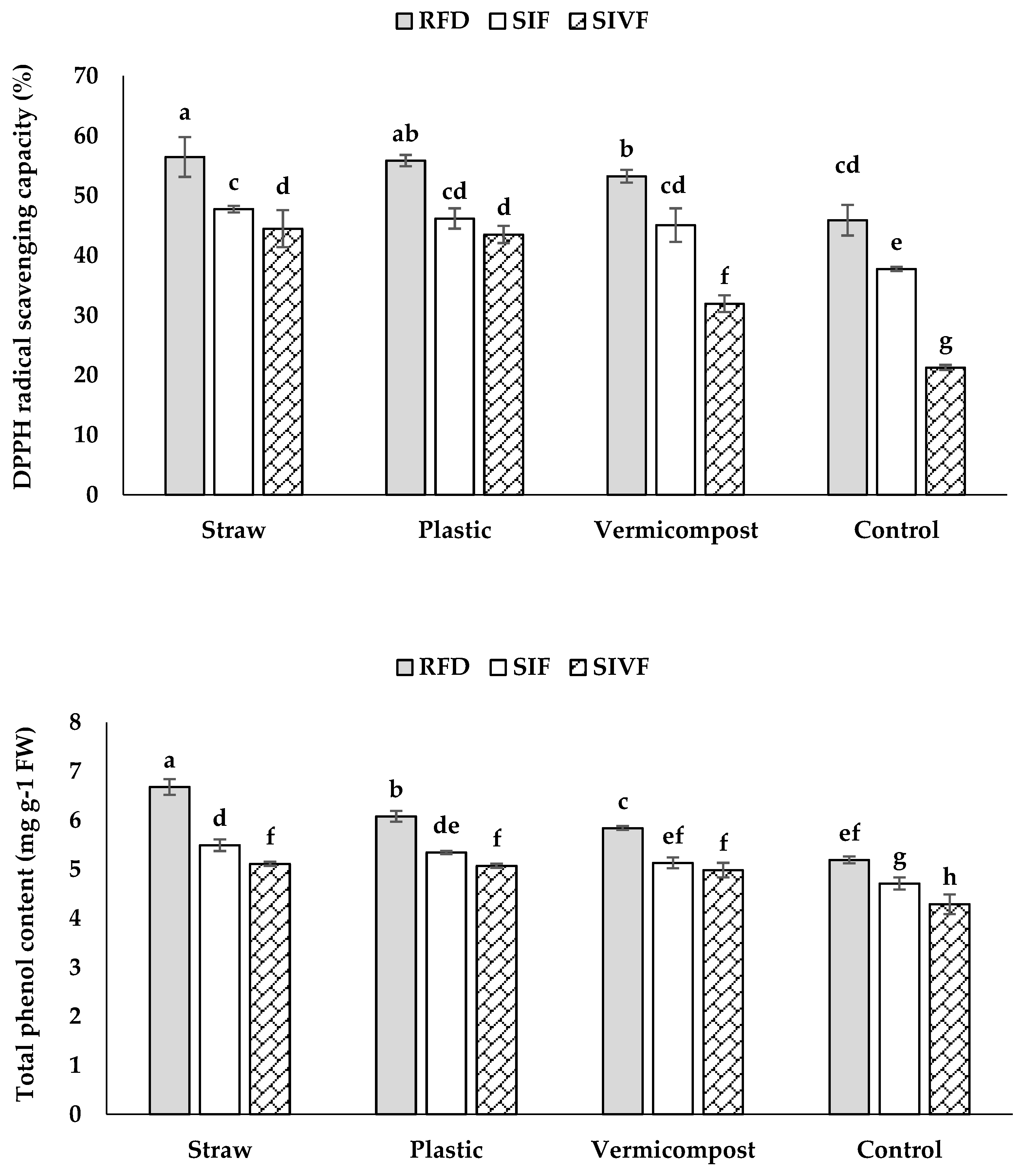
Table 2.
Composition of linseed oil fatty acids in mulch and stress modifier applications.
| No. | Component | Treatment | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RFD | RFD | RFD | RFD | SIF | SIF | SIF | SIF | SIVF | SIVF | SIVF | SIVF | ||
| C | V | PM | SM | C | V | PM | SM | C | V | PM | SM | ||
| 1 | Palmitic acid | 6.09 | 6.18 | 6.19 | 6.41 | 6.3 | 6.49 | 7.38 | 6.96 | 5.08 | 5.1 | 4.49 | 4.68 |
| 2 | Stearic acid | 5.08 | 5.29 | 5.78 | 5.53 | 5.61 | 5.67 | 5.5 | 5.66 | 5.15 | 5.09 | 4.97 | 5.45 |
| 3 | Oleic acid | 21.54 | 21.93 | 23.02 | 23.28 | 23.65 | 24.22 | 24.12 | 24.78 | 23.11 | 25.02 | 24.96 | 25.99 |
| 4 | Linoleic acid | 12.06 | 12.57 | 12.18 | 12.28 | 12.05 | 12.15 | 12.32 | 12.66 | 10.67 | 12.85 | 12.58 | 12.91 |
| 5 | Linolenic acid | 50.26 | 52.56 | 51.99 | 51.71 | 50.73 | 50.86 | 49.05 | 51.46 | 41.09 | 41.3 | 38.31 | 40.46 |
| 6 | Arachidic acid | 0.65 | 0.21 | 0.21 | 0.21 | 0.21 | 0.21 | 0.21 | 0.21 | 0.21 | 0.21 | 0.21 | 0.21 |
C, V, PM, and SM correspond to control, vermicompost, plastic, and straw mulch, respectively. RFD, SIF, and SIVF correspond to rainfed, with one supplementary irrigation at the flowering stage and two supplementary irrigations in the vegetative and flowering stages, respectively. The main components are bold.
Table 3.
Analysis of variance of linseed physiological traits as influenced by mulch type and irrigation regime.
Table 3.
Analysis of variance of linseed physiological traits as influenced by mulch type and irrigation regime.
| Source of variation | Relative water content | Chlorophyll a | Chlorophyll b | Carotenoids | Total soluble sugars | Proline content | CAT | SOD | Total phenol content | DPPH radical scavenging capacity |
|---|---|---|---|---|---|---|---|---|---|---|
| Mulch (M) | ** | ** | ** | * | ** | ** | ** | ** | ** | ** |
| Irrigation regime (IR) | * | * | n.s. | ** | ** | ** | ** | * | ** | ** |
| M x IR | n.s. | ** | * | ** | ** | ** | ** | ** | ** | ** |
** and *: significant at 1% and 5% probability levels, respectively; n.s., not significant.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



