
Submitted:
31 January 2025
Posted:
03 February 2025
You are already at the latest version
Abstract
Recently, Fusarium root rot (FRR)-like symptoms were observed in Uganda’s agroecology zones, prompting the National Agricultural Organisation (NARO) to conduct a disease survey. The survey reports indicated FRR as the second most prevalent root rot disease of common bean in Uganda after Southern blight. Ninety nine Fusarium spp. strains were obtained from samples collected during the surveys. The strains were morphologically and pathogenically characterised and confirmed to cause Fusarium root rot as observed in the field. However, molecular characterization of the strains was not conducted. In this study therefore, 80 of the strains were characterized using partial sequences of translation elongation factor 1-alpha (TEF-1α) gene, beta tubulin (β tubulin) gene and internal transcribed spacers (ITS) region of ribosomal RNA to determine species diversity. High quality Sanger sequences from the target genes were compared to the sequences from Fusarium species available in the National Centre for Biotechnology Information coding sequences (NCBI-CDS) database to determine the most likely species the strains belonged. The sequences from our strains were deposited into the NCBI gene bank under ID#288420, 2883276, 2873058 for TEF-1α, β tubulin and ITS respectively. The Fusarium species identified included; F. Oxysporum, F. solani, F. equiseti F. delphinoides, F. commune, F. subflagellisporum, F. fabacearum, F. falciforme, F. brevicaudatum, F. serpentimum, F. fredkrugeri and F. brachygibbosum. The diversity of these Fusarium species needs to be taken into consideration when developing breeding programs for management of the disease since currently there is no variety of common bean resistant to FRR in Uganda.

Keywords:
Common bean
; Dry beans
; Fusarium root rot
; Genetic diversity
1. Introduction
The common bean (Phaseolus vulgaris L) is the most widely distributed Phaseolus species grown all over Africa [1]. According to FAO [2], Uganda produced 1,008,410 tons of common bean in 2016 making it the second largest producer after Tanzania at 1,200,000 tons. The production in Uganda is mostly done by small scale farmers with land holdings of between 0.1 to 4 hectares [3]. Several biotic and abiotic constraints face common bean production and among the biotic constraints, root rots are key [4,5].
Fusarium root rot disease hereafter referred to as FRR, was reported as the second most important bean root rot disease in Uganda after Southern blight caused by Sclerotium rolfsii Sacc. (teleomorph Arthelia rolfsii (Curzi) C. C. Tu & Kimbr.) [6]. The symptoms of FRR include longitudinal reddish-brown lesions on hypocotyls accompanied by longitudinal fissures or cracks with dying root tissues turning reddish brown. Infected plants are chlorotic beginning with the primary leaves, stunted and plants may wilt completely or undergo premature senescence. Bean yield losses due to FRR have been reported to reach 86% in severely infected soils [7]. The legumes program of National Agricultural Research Organisation conducted a survey of seven agroecological zones that included the South Western Highland (SWH), Western Mixed Farming system (WMFS), Lake Victoria Crescent and Mbale Farmlands (LVC), Eastern Highlands (EH), Northern Mixed Farming Syatem (NMFS), North Eastern Dry Lands (NEDL) and West Nile Mixed Farming System (WNMFS). During these surveys, wilting plants with Fusarium root rot like symptoms were collected and used for pathogen isolation. Fusarium species strains were obtained and characterised both morphologically in culture media and phenotypically through pathogenecity studies [8]. Whereas, understanding the molecular diversity among pathogen populations to facilitate development of host plant resistance is important, genetic diversity studies were not conducted in the earlier study.
Genetic diversity among Fusarium species has been studied using DNA-based markers such as Inter Simple Sequence Repeats (ISSR) and Single Sequence Repeats (SSR) [9], Amplified Fragment Length Polymorphism (AFLP) [10], Restriction Fragment Length Polymorphism (RFLP) and Randomly Amplified Polymorphic DNA (RAPD) [11]. Internal transcribed spacers region of the ribosomal RNA (ITS), beta tubulin (βtubulin) gene region [12], calmodulin gene region [13] and sequences from translation elongation factor 1 alpha (TEF1-α) gene have been widely used as taxonomic markers for fungal species identification [5,14,15].
Several studies have reported the effectiveness of TEF1-α gene in fungal species identification, disease diagnosis and postharvest fungal toxicity surveys in crops such as coffee (Coffea sp) [16], sugar beet (Beta vulgaris) [17], bread wheat (Triticum aestivum L.) [18], millet (Eleusine coracana Gaertn.), sorghum (Sorghum bicolor L. Moench.), maize (Zea mays L.), groundnuts/peanuts (Arachis hypogea L.) and sesame (Sesamum indicum L.) [19]. TEF-1α gene was used to identify and classify dermatophytes and it provided a high degree of differentiation between species that were closely related [15]. Similarly, partial sequences of β tubulin gene region have been used to study molecular diversity and identification of Fusarium species. Kalman et al. [12] used the β tubulin gene region to identify Fusarium species causing basal rot in Allium cepa. Several authors have used ITS for identification of fungal species [20]. Singha et al. [21] used ITS 1 and 4 to identify Fusarium species causing wilts in tomatoes and was able to detect several Fusarium species such as F. oxysporum, F. equiseti, F. proliferatum. Other authors have used other gene regions such as clamodulin (cam), RNA polymerase second largest subunit (rpb2) genes and Cytochrome oxydase 1 (COX 1) gene region for identification of Fusarium to species level. [13,22]. Though Calmodulin primers were ables to distiguish the Fusarium species [13], In the study by Gilmore [22], many of the species of Fusarium shared similar COX 1 partical gene sequences making COX 1 barcoding in Fusarium entirely infeasible.
The study by Paparu et al. [6] showed an increasing significance of FRR in Uganda’s agroecology zones. The disease was second most prevalent after Southern blight. Howevere, there is limited information on the diversity of Fusarium species causing the root rot disease in common bean. To fill the observed knowledge gap, we sought to identify Fusarium species causing bean root rot in Uganda. This information is useful in the development of host plant resistance which, is a key disease management strategy for smallholder farmers in sub saharan Africa.
2. Materials and Methods
2.1. Origin of Fusarium species Strains Used
A collection of 99 hyphal tipped Fusarium species strains previously stored on filter paper originated from 6 agroecological zones of Uganda. Strains were re-activated by growing them on Potato Dextrose Agar (PDA) media ( 39g PDA in 1L distilled water) for 14 days. Strains with growth rates of less than 0.6cm per day were selected since Fusarium species that cause common bean root rot were reported to have low growth rates [23].
2.2. DNA Extraction from Fusarium species Strains
DNA was extracted from two-week old mycelia of the 99 previously mentioned strains using a modified Cetyl trimethylammonium bromide (CTAB) protocol previously used by Joint Research Council (JRC), European commission [24]. Actively growing mycelia were harvested by scraping them off the surface of the PDA into sterile Petri dishes. The mycelia were oven dried over night at 30oC. About 0.02g of the mycelia was loaded into 2ml Eppendorf tubes containing beads. The mycelia were ground into a fine powder using an automated tissue homogenizer and cell lyser Geno Grinder (1600 MiniG,) for 3 minutes at 1450 strokes/minutes. Seven hundred microliters (700μl) of DNA extraction buffer (2% CTAB, 50mM EDTA pH 8.0, 100mM Tris-Base pH 8.0, 2% PVP-40, 1% NaSO3, 1.4M NaCl and 1% beta 2-mercaptoethanol) was added and the mycelia homogenized for another 2 minutes in the Geno grinder. Samples were incubated at 65oC for 30 minutes with occasional shaking. Tubes were then centrifuged at 12,000 strokes per minute for 10 minutes. Five hundred microliters (500μl) of the supernatant was picked and transferred into new 2ml Eppendorf tubes. Four hundred fifty microliters (450μl) of Chloroform and Iso amyl alcohol in the ratio of 24:1 was added to each sample and the tubes were shaken for 2 minutes. Samples were then centrifuged at 10,000 strokes per minute for 10 minutes. Four hundred microliters (400μl) of supernatant containing DNA was transferred into well-labeled 1.5ml Eppendorf tubes. Four hundred and fifty microliters (450μl) of Isopropanol (stored at -20oC) and 40µl of 3M Sodium Acetate solution were added to the DNA and incubated at -20oC for 2 hours to precipitate the DNA. The tubes were then centrifuged at 15,000 rpm for 15 minutes to separate the DNA from the Isopropanol. The supernatant was decanted and the pellet washed with 500µl of 70% ethanol by centrifuging at 7000 strokes per minute for 10 minutes. The supernatant was decanted and DNA pellets air dried for 1 hour at room temperature (25-30oC). DNA pellets were then resuspended in 100µl of elution buffer, DNA concentration assessed using a NanoDrop (ND-1000) and stored at -80oC. Generally, the concentration of all the samples was above 500ng/ μl while the A260/A280 ratios ranged between 1.9 to 2.1.
2.3. Fusarium species Identification Using TEF1-α, β Tubulin and ITS Partial Sequences
A multi gene approach was used to identify the 99 Fusarium species strains. The primer sequences used to amplify partions of the target genes included TEF1-α gene forward primer (Ef 1: 5'-ATGGGTAAGGARGACAAGAC-3') and reverse primer
(Ff 2: 5’-GGARGTACCAGTSATCATGTT-3’) [25]. ITS forward primer ITS 1 (GGAAGTAAAAGTCGTAACAAGG) and reverse primer ITS 4 (TCCTCCGCTTATTGATATGC) was used for amplification of ITS region of ribosomal RNA [26]. Meanwhile β tubulin gene region was amplified using forward primer (T1-AACATGCGTGAGATTGTAAGT) and reverse primer (T2-TAGTGACCCTTGGCCCAGTTG) [12].
A PCR master mix (Bioneer Corporation, South Korea) was used in the amplification reactions according to the manufacturer’s instructions. A total reaction volume of 30µl was used and it consisted of 15µl premix, 1µl of each reverse and primer, 3µl of DNA and 10µl of DNase free water. The PCR conditions included an initial denaturation at 95oC for 5 min followed by 35 cycles of denaturation at 95oC for 3 min, annealing at the various annealing temperature for the respective primers for 40 sec, extension at 72oC for 1 min and a final extension at 72oC for 5 min. The annealing temperature for each primer was as follows; TEF1-α at 55oC, ITS at 53oC and β tubulin at 57 oC. For quality control, 5µl of PCR products from each sample were electrophoresed alongside the 100bp DNA ladder in a 1.5% agarose gel containing Gel-red fluorescent dye (Botium) in 1x TBE buffer at 100V for 40 min. Gels were documented using a bench top Transilluminator (BioDoc-ItTM Imaging System 8.0). However, out of the 99 strains, 80 of the strains were able to produce PCR bands with the various primers. The PCR products from the 80 strains that showed good amplification were purified using AccPrepTM Purification Kit (Bioneer Cooperation) following the manufacturer’s instructions. The products were sequenced using their respective reverse primers in an ABI13730XL Sanger sequencing machine (Applied Biosystems, USA) using BigDye Terminator v3.1 sequencing kit (Applied biosystems, USA) at Macrogen (Amsterdam, Netherlands).
2.4. Growth Rate, Disease Severity Index (DSI) and Morphological Characteristics of Fusarium species Strains
The average disease severity index (DSI) and growth rate of strains was obtained from Erima et al. [8]. In the study by Erima et al., the inoculum was prepared by cutting 1cm square agar plugs from two weeks old cultures on PDA and inoculating in 50g of sterile millet in a autoclave bag. Spore concentration could not be used to measure the inoculum because some of the isolates did not produce conidia. Bags were incubated at 25oC for two weeks until mycelia had fully covered the millet. Wooden trays of 100cm x 35cm x 10cm were used to set up the experiment in greenhouse. Ten grams of the inoculum was mixed with about 20kg of soil in the wooden trays. Then 16 seeds of each of the five test lines were planted in each tray with a replicate. A control tray which was un-inoculated was also planted with the test varieties. Virulence was then assessed at 28 days after planting using a scale of 1 to 9.
Meanwhile, growth rate was determined on PDA using 9cm daimeter Petri dishes. A cross was made on the bottom of Petri dish to mark its centre. Inoculum was picked from 2 week-old cultures by tapping the mycelia with a needle. The inoculum was then tranfered to the centre of the marked Petri dish. Each strain was replicated thrice. Growth data was collected from 2 days post inoculation by using a 30cm ruler to measure diameter of the colony until day 8 when mycelia for some isolates had reached the edge of the Petridish. Information on colony color was also recorded. Microscopy was then conducted using 2 weeks old cultures on PDA at X40 for selected strains of the different species. The shape and sizes of the macro and micro conidia were recorded and photos taken of the different strains.
2.5. Data Analysis
Sanger sequences were imported into chromas software for quality assessment. Low quality bases at the 5’- and 3’-end were trimmed off and high quality sequences exported as a fasta file. The high-quality reads representing 80 Fusarium species strains from different agro-ecology zones were obtained and used for downstream analysis. They included; 60 sequences from TEF1-α, 59 sequences from β tubulinand 58 sequences from ITS. The number of sequences of various strains varied because not all primer sets amplified the genes from same strains. To confirm the species, the strains’ sequences were compared to the coding sequences in the National Centre for Biotechnology Information coding sequences (NCBI-CDS) database using basic local alognment search tool for nucleotides (BLASTn). The sequences were analyzed for the presence of open reading frames, exons and introns. Concordance of species name between two independent databases as the top hit was used to assign the species identity to the strains. Sequences were imported into MEGA 11.0 and aligned. A phylogenetic tree was constructed using the Neighbor Joining method using the TEF1-α sequences since it resolved all the Fusarium species. Curated Fusarium species sequences were deposited to NCBI data base. ITS and β tubulin could not resolve some species from Fusarium solani and Fusarium oxysporum species complexes identifying all of them as F. solani and F. oxysporum respectively. Data on morphological characteristics such as growth rate, virulence and colony colour were obtained from Erima et al. [8]. Tukeys honestly significant difference (HSD) test was used to test the difference in virulence and growth rate between the different Fusarium species.
3. Results
Identification of Fusarium strains Using TEF1-α Gene, β Tubulin Gene and ITS Partial Sequences
. Partial sequences of about 700 bp, 580 bp and 560 pb were obtained after PCR amplification and sequencing of the PCR products of TEF1-α gene, β tubulin gene and ITS partial sequences, respectively (Figure 1). Sequences were successfully sequenced and processed for a combined total of 80 strains (TEF1-α= 60 strains, β tubulin= 59 strains and ITS= 58 strains). The sequences were deposited at National Centre for Biotechnology Information (NCBI) under accessions PQ363745 to PQ363805, PQ497178 to PQ497237, PQ497119 to PQ497177 for ITS, TEF1-α and β tubulin, respectively.
Comparison of the high-quality trimmed Sanger sequences to NCBI’s CDS database for TEF1-α, ITS and β tubulin identified 12 different Fusarium species with identities ranging from 99.9 to 100%. Thirty seven strains were most identical to F. oxysporum, 13 to F. solani, 7 to F. falciforme, 9 to F. equiseti and 4 to F. commune. Meanwhile, F. fabacearum and F. subflagellisporum were each represented by 2 strains. Single strains of F. delphinoides, F. brevicaudatum, F. serpentimum, F. fredkrugeri and F. brachygibbosum were identified. A strain belonging to Clonostachys rhizophaga was also identified (Table 1). TEF1-α was able to resolves the strains within the same species complex to species level meanwhile ITS and β tubulin were unable to resolve F. falciforme and F. serpentimum from Fusarium solani species complex identifying them as F. solani. The two primers were also unable to resolve F. fredkrugeri, F. commune, F. fabacearum, F. subflagellisporum and F. brachygibbosum from Fusarium oxysporum species complex identifying them as Fusarium oxysporum. A maximum likelihood phylogenetic tree was generated using neighbor joining method using the TEF1-α sequences since it had resolved all the species (Figure 2). The strains clustered according to their species. A consensus tree could not be generated by concatinating the sequences because not all the primers amplified all the sequences. A strain MitF-487-2 identified as Clonostachys rhizophaga could not be included in the tree because its sequences were too divergent to align with those of Fusarium species. The Fusarium species, their agroecology of origin and accession numbers are summarized in Table 1.
Fusarium species have been reported to vary in their morphological characteristics such as growth rate, virulence and shape and sizes of microscopic structres and colony colour [23,30]. The average disease severity index (DSI), growth rate and colony colours of the strains was obtained from Erima et al. [8]. The average DSI and growth rate varied among the species (Table 1 and Table S1). All the different Fusarium speices varied significantly in disease severity index (DSI) caused to five common bean varieties (P<0.001). Fusarium solani was the least pathogenic with average DSI of 37.2% while F. subplagellisporum was the most pathogenic with average DSI of 66.6% (Table 1). The Fusarium species strains also varied significantly in average growth rate per day (P<0.001). Growth rates of F. brachygibosum and F. fredkrugeri were not significantly different while that of F. solani and F. oxysporum were also similar. There were also no significant differences between the growth rates of F. equiseti and F. fabacearum.
Many of the Fusarium species strains were multicoloured. Colony colorations such as white, white/purple, white/pink and white/cream were reported for F. oxysporum and F. solani though specific colorations such as white/yellow and white/brown were reported for F. solani. All the strains of F. equiseti were white on the top of the Petri dish. While F. falciforme and F. commune had strains which were white/pink, white/purple and white. The strains of F. brevicaudatum, F. serpentimum, F. brachygibbosum, F. subflagellisporum, C. rhizophaga, F. delphinoides, F. fabacearum and F. fredkrugeri were coloured white, white/purple white/purple, white/brown, white/purple, white/pink, purple and white respectively (Figure 3, Table S1. Photos of the symptoms caused by a few Fusarium species identified above were retrived from the archives at National Crops Resources Research Institute and were observed to varry among the species. The damage caused by F. oxysporum was along the vascular bundle and extended above the soil line while the lesions caused by F. equiseti, F. serpentimum and F. falciforme were restricted to the root area (Figure 4). Photos were not captured for every strain phenotyped, as we were not aware at that time if they belonged to different species or not.
Following microscopy, all the strains were observed to have septed hyphae. They either produced micro or macro conidia or both. The micro conidia were spherical while the macro conidia were rod shaped, oval or sickle shaped. The shape and sizes of micro and macro conidia are summerised in Figure 5 and Table 2.
4. Discussion
In this study, translation elongation factor 1-alpha (TEF1-α), β tubulin and ITS region of ribosomal RNA partial sequences were used to identify Fusarium species strains previously isolated from roots of wilting common bean. Fusarium species identified as the main pathogens causing Fusarium root rot (FRR) in Uganda included F. oxysporum, F. solani, F. equiseti, F. falciforme, F. flagellisporum, F. commune, F. brevicaudatum, F. brachyggibosum, F. serpentimum, F. frekrugeri, F. fabacearum and F. delphinoides. Identification of Fusarium species based on plant disease symptoms is quite challenging. In both the field and greenhouse, early symptoms of FRR and wilt are similar (wilting and yellowing of leaves) and sometimes root rots occur as disease complexes. Morphological identification and classification continue to be used but with enormous challenges as it requires experienced mycologists to identify fungi to species level [27]. Despite this, proper identification and classification of Fusarium spp. is important for monitoring changes in the species population and their impact on agriculture.
Several members of the Fusarium species complex have a wide host range and ecological niches but also differ in their characteristics. For example, F. equiseti was reported to cause seedling wilting, root tip discoloration and necrosis in sugar beet by Mohamad et al. [28]. Fusarium falciforme was also reported to cause root rot in Weigelia florida in China [29]. According to Rahma et al. [30], both F. oxysporum and F. brachygibbisum cause die back in olive trees and the two species clustered closely in the current study. The other species that clustered closely with Fusarium oxysporum included F. commune, F. frekrugeri, F. subflagellisporum, F. delphinoides, F. fabacearum, F. brachygibbosum and F. brevicaudatum. These species were resolved by TEF1-α while beta tubulin and ITS only reolved F. solani, F. oxysporum and F. equiseti. Namasaka [31] reported F. equiseti as a causal agent of cowpea root rot. Interestingly, the current study confirmed the specie as a causal agent of common bean root rot in Uganda. Marcelo et al. [32] recovered F. frekrugeri from soil under Musa acuminata from Kruger national park in South Africa in undisturbed forest soil. In this study, F. frekrugeri caused a DSI of 40.3%. Meanwhile, F. delphinoides strain GPK was reported to be pathogenic to chick peas and pigeon peas by Guruprasad et al. [33].
There are at least 20 species complexes in the genus Fusarium. Chehri et al. [34] and Coleman [35] reported F. falciforme as a specie under F. solani species complex. In the current study, the the strains of F. falciforme and F. solani clustered closely supporting the above argument. However, contradicting nomenclature of Fusarium species continues to be a challenge resulting in the lack of congruity between morphological and molecular phylogeny. For example, Sang et al. [36] reported F. cuneirostrum as a causal agent of FRR in common bean in Uganda, yet these strains were initially identified as F. solani f. sp. Phaseoli by Munkankusi [37], based on colony characteristics. Ji-wen et al. [38] also reported F. equiseti as members of F. incarnatum-equiseti species complex .
A strain MitF-487-2, identified as Clonostachys rhizophaga was obtained from LVC agroecology. It was detected by ITS and β tubulin while TEF1- α did not detect it. This is the first report of C. rhizophaga causing wilts in common bean in Uganda. The pathogen is reported to be pathogenic to several crops. It reportedly causes wilts and root rot in chick peas [34,39,41]. In water chestnut, C. rhizophaga causes longitudinal chlorotic streaks and black spots on the stem surface and vascular necrosis [42]. However, some other Clonostachys species such as C. rosea are reported to be mycoparasitic. They are aggressive parasites of fungi and research on their use for plant disease control is ongoing [43].
Secondary data on DSI was obtained from Erima et al. [8]. All the species differed significantly in DSI caused on common bean. Strains identified as F. oxysporum caused more disease than F. solani. However, in an earlier study, Chehri et al. [31] observed that F. solani caused more disease than F. oxysporum on potato tubers. Differences in DSI among different Fusarium species were equally observed by Siddique et al. [44] in common bean and Burlakoti et al. [45] in sugar beet, where F. graminearum strains were more pathogenic than F. oxysporum strains. The variation in virulence of a single Fusarium species in many different crops is an indicator that these species have their primary host on which they proliferate most. The colony colorations of the different species strains in the current study is also related to what other researchers obsereved. For example, Trebelsi et al. [30] reported purple coluration of Fusarium oxysporum causing olive trees die back. Tuiime [23] also reported white and brown coloration in Fusarium solani fsp phaseoli. All the F. equiseti isolates in this study had abundant white mycelia, similar findings were reported by Mohamed et al. [28]. Similarly, white colony coloration in F. falciforme was reported by Dong-Xia [29].
The Fusarium species strains in the current study were obtained from different agroecological zones of Uganda. This shows that Fusarium species can survive in a wide range of temperatures ranging from the cool humid South Western highlands to warm and less humid North Eastern Dryland. Tusiime [23], reported F. solani fsp phaseoli in the cool humid regions of South Western Highlands. However, In the current study, F. solani was reported in various agroecological zones including Northern Mixed farming system, Westerm Mixed farming system, Lake victoria crescent and Mbale farmlands, South Western Higlands and and North Eastern dry land which is generally warmer and less humid.
5. Conclusions
Fusarium species causing root rots and wilts of common bean in Uganda are morphologically, phenotypically and genetically diverse. This research has generated information on the molecular diversity of Fusarium species causing common bean root root. The diversity of Fusarium species observed in our study needs to be taken into consideration while developing new varieties in breeding program for management of the disease.
Supplementary Materials
Partial sequences of Fusarium species strains can be obtained at NCBI under ID#2884260, 2883276, 2873058 for TEF-1α, β tubulin and ITS respectively
Author Contributions
Conceptualization, P.P. and S.E.; methodology, P.P. and S.E.; A.N.; A.C.; N.H.; formal analysis, S.E. and M.N.; resources, P.P.; writing- First draft, S.E.; writing- review and editing, M.N., P.P. and R.E.; supervision, P.P. and R.E. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Bill & Melinda Gates Foundation. Opportunity/Contract ID: OPP1084135
Institutional Review Board Statement
Not applicable
Informed Consent Statement
Not applicable
Data Availability Statement
The partial sequences of TEF 1α, βtubulin and ITS can be obtained from NCBI data base under ID; 288260, 288276 and 2873058 for TEF 1α, β tubulin and ITS respectively. The rest of the data are within the article
Acknowledgments
We acknowledge funding from the Bill & Melinda Gates Foundation through Grant Ref OPP1084135. We are grateful to staff of the Legumes Program at the National Crops Resources Research Institute, who provided us with the strains. Our appreciation also goes to the management and staff of Makerere University Centre for Crop Improvement who hosted and supported the molecular work.
Conflict of Interest
The authors have no conflict of intertest. The funders did not participate in conceptualization of the research.
References
- Melotto, M.; Monteiro-Vitorello, C.B.; Bruschi, A.G.; Camargo, L.E. Comparative bioinformatic analysis of genes expressed in common bean (Phaseolus vulgaris L.) seedlings. Genome 2005, 48, 562–570. [Google Scholar] [CrossRef] [PubMed]
- Food and Agricultural Organisation of United Nations (FAO). World food and agricultural statistical pocket book. Rome, Italy, 2018, pp 28.
- CASA. Bean Sector Strategy-Uganda. CASA Uganda country team, Kampala, Uganda. 2020, pp 4. Unpublished.
- Miklas, P.N.; Kelly, J.D.; Beebe, S.E.; Blair, M.W. Common bean breeding for resistance against biotic and abiotic stresses: From classical to MAS breeding. Euphytica 2006, 147, 105–131. [Google Scholar] [CrossRef]
- Li, Y.P.; You, M.P.; Barbetti, M.J. Species of Pythium Associated with Seedling Root and Hypocotyl Disease on Common Bean (Phaseolus vulgaris) in Western Australia. Plant Dis. 2014, 98, 1241–1247. [Google Scholar] [CrossRef] [PubMed]
- Paparu, P.; Acur, A.; Kato, F.; Acam, C.; Nakibuule, J.; Musoke, S.; Nkalubo, S.; Mukankusi, C. PREVALENCE AND INCIDENCE OF FOUR COMMON BEAN ROOT ROTS IN UGANDA. Exp. Agric. 2017, 54, 888–900. [Google Scholar] [CrossRef]
- Abawi, G. S.; Pastor-Corrale, M. A. Root rots of beans in Latin America and Africa:Diagnosis, research methodologies, and management strategies: CIAT, Cali, Colombia, 1990, p.114. https://hdl.handle.net/10568/54258.
- Erima, S.; Nyine, M.; Edema, R.; Nkuboye, A.; Nakibuule, J.; Paparu, P. Morphological and pathogenic characterization of Fusarium species causing common bean root rot in Uganda. J. Sci. Agric. 2024, 7–14. [Google Scholar] [CrossRef]
- Zietkiewicz, E.; Rafalski, A.; Labuda, D. Genome Fingerprinting by Simple Sequence Repeat (SSR)-Anchored Polymerase Chain Reaction Amplification. Genomics 1994, 20, 176–183. [Google Scholar] [CrossRef]
- Puri, K.D.; Saucedo, E.S.; Zhong, S. Molecular Characterization of Fusarium Head Blight Pathogens Sampled from a Naturally Infected Disease Nursery Used for Wheat Breeding Programs in China. Plant Dis. 2012, 96, 1280–1285. [Google Scholar] [CrossRef]
- Fourie, G.; Steenkamp, E.T.; Ploetz, R.C.; Gordon, T.R.; Viljoen, A. Current status of the taxonomic position of Fusarium oxysporum formae specialis cubense within the Fusarium oxysporum complex. Infect. Genet. Evol. 2011, 11, 533–542. [Google Scholar] [CrossRef]
- Kalman, B.; Abraham, D.; Graph, S.; Perl-Treves, R.; Harel, Y.M.; Degani, O. Isolation and Identification of Fusarium spp., the Causal Agents of Onion (Allium cepa) Basal Rot in Northeastern Israel. Biology 2020, 9, 69. [Google Scholar] [CrossRef]
- Nuangmek, W.; Kumla, J.; Khuna, S.; Lumyong, S.; Suwannarach, N. Identification and Characterization of Fusarium Species Causing Watermelon Fruit Rot in Northern Thailand. Plants 2023, 12, 956. [Google Scholar] [CrossRef]
- Kristensen, R.; Torp, M.; Kosiak, B.; Holst-Jensen, A. Phylogeny and toxigenic potential is correlated in Fusarium species as revealed by partial translation elongation factor 1 alpha gene sequences. Mycol. Res. 2005, 109, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Mirhendi, H.; Makimura, K.; de Hoog, G.S.; Rezaei-Matehkolaei, A.; Najafzadeh, M.J.; Umeda, Y.; Ahmadi, B. Translation elongation factor 1-α gene as a potential taxonomic and identification marker in dermatophytes. Med Mycol. 2014, 53, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Olal, S.; Olango, N.; Kiggundu, A.; Ochwo, S.; Adriko, J.; Nanteza, A.; Matovu, E.; Lubega, G. W.; Kagezi, G.; Hakiza, G. J.; Wagoire, W. Using translation elongation factor gene to specifically detect and diagnose Fusarium xylaroides, a causative agent of coffee wilt disease in Ethiopia, East and Central Africa. J. Plant Pathol. Microbiol. 2018, 9, 440. [Google Scholar] [CrossRef]
- Nitschke, E.; Nihlgard, M.; Varrelmann, M.; Hafez, M.; Abdelmagid, A.; Adam, L.R.; Daayf, F.; Covey, P.A.; Kuwitzky, B.; Hanson, M.; et al. Differentiation of Eleven Fusarium spp. Isolated from Sugar Beet, Using Restriction Fragment Analysis of a Polymerase Chain Reaction–Amplified Translation Elongation Factor 1α Gene Fragment. Phytopathology® 2009, 99, 921–929. [Google Scholar] [CrossRef]
- Kheseli, O.P.; Susan, I.S.; Sheila, O.; Otipa, M.; Wafula, W.V. Prevalence and Phylogenetic Diversity of Pathogenic Fusarium Species in Genotypes of Wheat Seeds in Three Rift Valley Regions, Kenya. Adv. Agric. 2021, 2021, 1–13. [Google Scholar] [CrossRef]
- Wokorach, G.; Landschoot, S.; Audenaert, K.; Echodu, R.; Haesaert, G. Genetic Characterization of Fungal Biodiversity in Storage Grains: Towards Enhancing Food Safety in Northern Uganda. Microorganisms 2021, 9, 383. [Google Scholar] [CrossRef]
- Fajarningsih, N.D. Internal Transcribed Spacer (ITS) as Dna Barcoding to Identify Fungal Species: a Review. Squalen Bull. Mar. Fish. Postharvest Biotechnol. 2016, 11, 37–44. [Google Scholar] [CrossRef]
- Singha, I.M.; Kakoty, Y.; Unni, B.G.; Das, J.; Kalita, M.C. Identification and characterization of Fusarium sp. using ITS and RAPD causing fusarium wilt of tomato isolated from Assam, North East India. J. Genet. Eng. Biotechnol. 2016, 14, 99–105. [Google Scholar] [CrossRef]
- Gilmore, S.R.; Gräfenhan, T.; Louis-Seize, G.; Seifert, K.A. Multiple copies of cytochrome oxidase 1 in species of the fungal genus Fusarium. Mol. Ecol. Resour. 2009, 9, 90–98. [Google Scholar] [CrossRef]
- Tusiime, G. Variation and detection of Fusarium solani f. sp. phaseoli and quantification of soil inoculum in common bean fields. PhD Thesis, Makerere University, Kampala, Uganda. 2004.
- https://joint-research-centre.ec.europa.eu/tools-and-laboratories/standardisation_en.
- O'Donnell, K. Molecular Phylogeny of the Nectria haematococca-Fusarium solani Species Complex. Mycologia 2000, 92, 919. [Google Scholar] [CrossRef]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; et al.; Fungal Barcoding Consortium; Fungal Barcoding Consortium Author List; Bolchacova, E Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed]
- Raja, H.A.; Miller, A.N.; Pearce, C.J.; Oberlies, N.H. Fungal Identification Using Molecular Tools: A Primer for the Natural Products Research Community. J. Nat. Prod. 2017, 80, 756–770. [Google Scholar] [CrossRef]
- Khan, M.F.; Liu, Y.; Bhuiyan, M.Z.R.; Lakshman, D.; Liu, Z.; Zhong, S.; Bhuyian, Z.R.; Lashman, D. First Report of Fusarium equiseti Causing Seedling Death on Sugar Beet in Minnesota, U.S.A. Plant Dis. 2021, 105, 2017. [Google Scholar] [CrossRef] [PubMed]
- Shen, D.-X.; Song, Z.-W.; Lu, Y.-M.; Fan, B. First Report of Fusarium falciforme (FSSC 3+4) Causing Root Rot in Weigela florida in China. Plant Dis. 2020, 104, 981–981. [Google Scholar] [CrossRef]
- Trabelsi, R.; Sellami, H.; Gharbi, Y.; Krid, S.; Cheffi, M.; Kammoun, S.; Dammak, M.; Mseddi, A.; Gdoura, R.; Triki, M.A. Morphological and molecular characterization of Fusarium spp. associated with olive trees dieback in Tunisia. 3 Biotech 2017, 7, 28. [Google Scholar] [CrossRef]
- Namasaka, R. W. 2017 Inheritence of resistance to Fusarium root rot (Fusarium Redolense) disease of cow peases in Uganda. MSc. Thesis, Makerere University, Kampala. [CrossRef]
- Sandoval-Denis, M.; Swart, W.J.; Crous, P.W. New Fusarium species from the Kruger National Park, South Africa. MycoKeys 2018, 34, 63–92. [Google Scholar] [CrossRef]
- Kulkarni, G.B.; Sajjan, S.S.; Karegoudar, T.B. Pathogenicity of indole-3-acetic acid producing fungus Fusarium delphinoides strain GPK towards chickpea and pigeon pea. Eur. J. Plant Pathol. 2011, 131, 355–369. [Google Scholar] [CrossRef]
- Chehri, K.; Mohamed, N.; Salleh, B.; Latiffah, Z. Occurrence and pathogenicity of Fusarium spp. on potato tubers in Malaysia. African J. Agric. Res. 2011, 6(16), 3706–3712. [Google Scholar] [CrossRef]
- Coleman, J.J. The Fusarium solani species complex: ubiquitous pathogens of agricultural importance. Mol. Plant Pathol. 2015, 17, 146–158. [Google Scholar] [CrossRef]
- Sang, H.; Jacobs, J.L.; Wang, J.; Mukankusi, C.M.; Chilvers, M.I. First Report of Fusarium cuneirostrum Causing Root Rot of Common Bean (Phaseolus vulgaris L.) in Uganda. Plant Dis. 2018, 102, 2639–2639. [Google Scholar] [CrossRef]
- Mukankusi, C. M. Improving resistance to Fusarium root rot [Fusarium solani (Mart.) Sacc. f. sp. phaseoli (Burkholder) W.C. Snyder & H.N. Hans.] in Common bean (Phaseolus vulgaris L.). PhD Thesis. University of KwaZulu- Natal, 2008 ,pp. 200.
- Xia, J.; Sandoval-Denis, M.; Crous, P.; Zhang, X.; Lombard, L. Numbers to names - restyling the Fusarium incarnatum-equiseti species complex. Persoonia - Mol. Phylogeny Evol. Fungi 2019, 43, 186–221. [Google Scholar] [CrossRef] [PubMed]
- Dugan, F.; Lupien, S.; Chen, W. Clonostachys rhizophaga and other fungi from chickpea debris in the Palouse region of the Pacific Northwest, USA. North Am. Fungi 2012, 7. [Google Scholar] [CrossRef]
- Cota-Barreras, C.I.; García-Estrada, R.S.; León-Félix, J.; Valenzuela-Herrera, V.; Mora-Romero, G.A.; Leyva-Madrigal, K.Y.; Tovar-Pedraza, J.M. First report of Clonostachys chloroleuca causing chickpea wilt in Mexico. New Dis. Rep. 2022, 46. [Google Scholar] [CrossRef]
- Gibert, S.; Edel-Hermann, V.; Gautheron, E.; Gautheron, N.; Bernaud, E.; Sol, J.; Capelle, G.; Galland, R.; Bardon-Debats, A.; Lambert, C.; et al. Identification, pathogenicity and community dynamics of fungi and oomycetes associated with pea root rot in northern France. Plant Pathol. 2022, 71, 1550–1569. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, A.-F.; Gu, C.-Y.; Zang, H.-Y.; Chen, Y. First Report of Clonostachys rhizophaga as a Pathogen of Water Chestnut (Eleocharis dulcis) in Anhui Province of China. Plant Dis. 2019, 103, 151–151. [Google Scholar] [CrossRef]
- Funck, J.D.; Dubey, M.; Jensen, B.; Karlsson, M. Clonostachys rosea to control plant diseases. 2022. [CrossRef]
- Siddique, S.; Bhuiyan, M.; Momotaz, R.; Bari, G.; Rahman, M. Cultural Characteristics, Virulence and In-vitro Chemical Control of Fusarium oxysporum f. sp. phaseoli of Bush bean (Phaseolus vulgaris L.). Agric. 2014, 12, 103–110. [Google Scholar] [CrossRef]
- Burlakoti, P.; Rivera, V.; Secor, G.A.; Qi, A.; Del Rio-Mendoza, L.E.; Khan, M.F.R. Comparative Pathogenicity and Virulence of Fusarium Species on Sugar Beet. Plant Dis. 2012, 96, 1291–1296. [Google Scholar] [CrossRef]
Figure 1.
PCR product bands of Fusarium species strains following amplification using ITS (A), β tubulin (B) and TEF1-α primers (C). Some strains without bands were not detected by the primers.
Figure 1.
PCR product bands of Fusarium species strains following amplification using ITS (A), β tubulin (B) and TEF1-α primers (C). Some strains without bands were not detected by the primers.
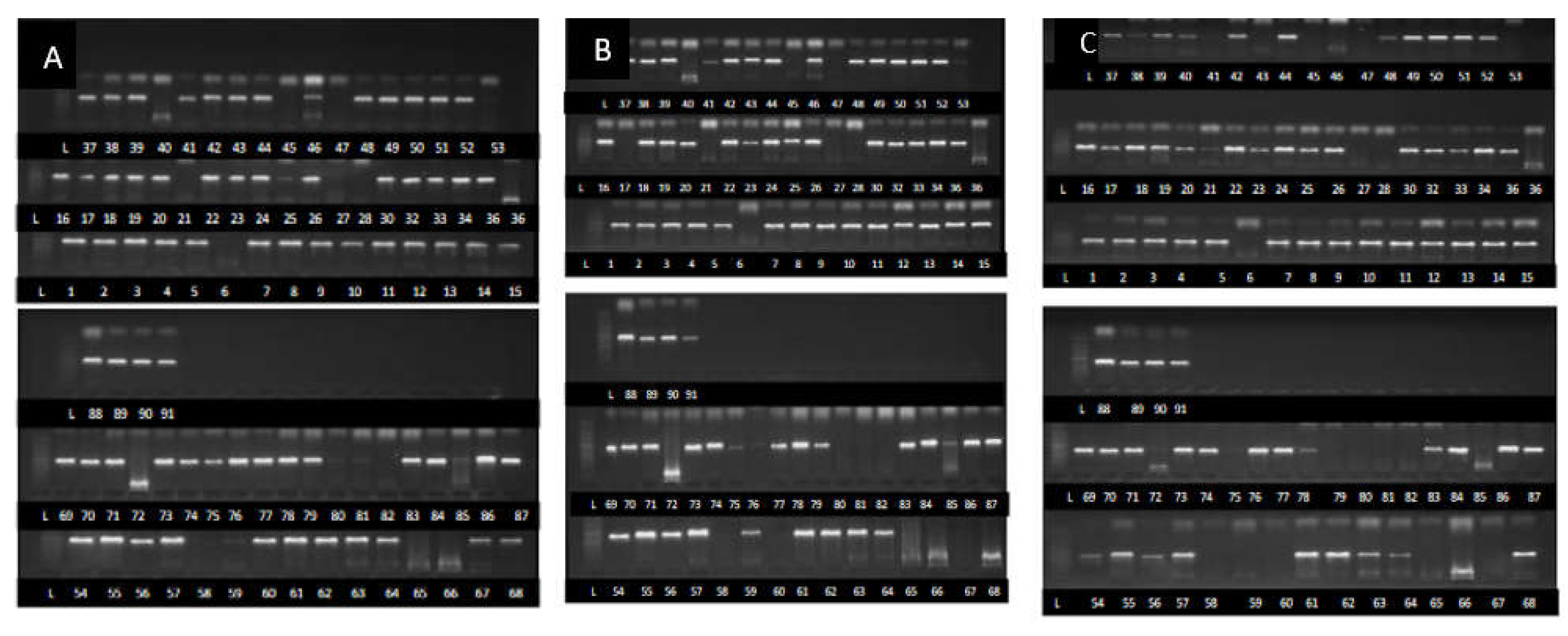
Figure 2.
The maximum likelihood phylogenetic tree constructed using the Neighbor Joining method for Fusarium species strains collected from six Ugandan agroecology zones.
Figure 2.
The maximum likelihood phylogenetic tree constructed using the Neighbor Joining method for Fusarium species strains collected from six Ugandan agroecology zones.

Figure 3.
Colony coloration of different Fusarium species strain on the top and bottom of Petridish; a- MorF-119 F. fabacearum, b- Apaf-560 F. subflagellisporum, c- MitF-487-2 C. rhizophaga, d- Kolf-563 F. brachygibbosum, e- Kirf-418 F. falciforme, f- Nakf-521 F. equiseti, g-Gulf-451-1 F. oxysporum, h- SirF-358 F. falciforme, i- Nakf-106-1 F. solani, j- Masf-403 F. fredkrugeri.
Figure 3.
Colony coloration of different Fusarium species strain on the top and bottom of Petridish; a- MorF-119 F. fabacearum, b- Apaf-560 F. subflagellisporum, c- MitF-487-2 C. rhizophaga, d- Kolf-563 F. brachygibbosum, e- Kirf-418 F. falciforme, f- Nakf-521 F. equiseti, g-Gulf-451-1 F. oxysporum, h- SirF-358 F. falciforme, i- Nakf-106-1 F. solani, j- Masf-403 F. fredkrugeri.
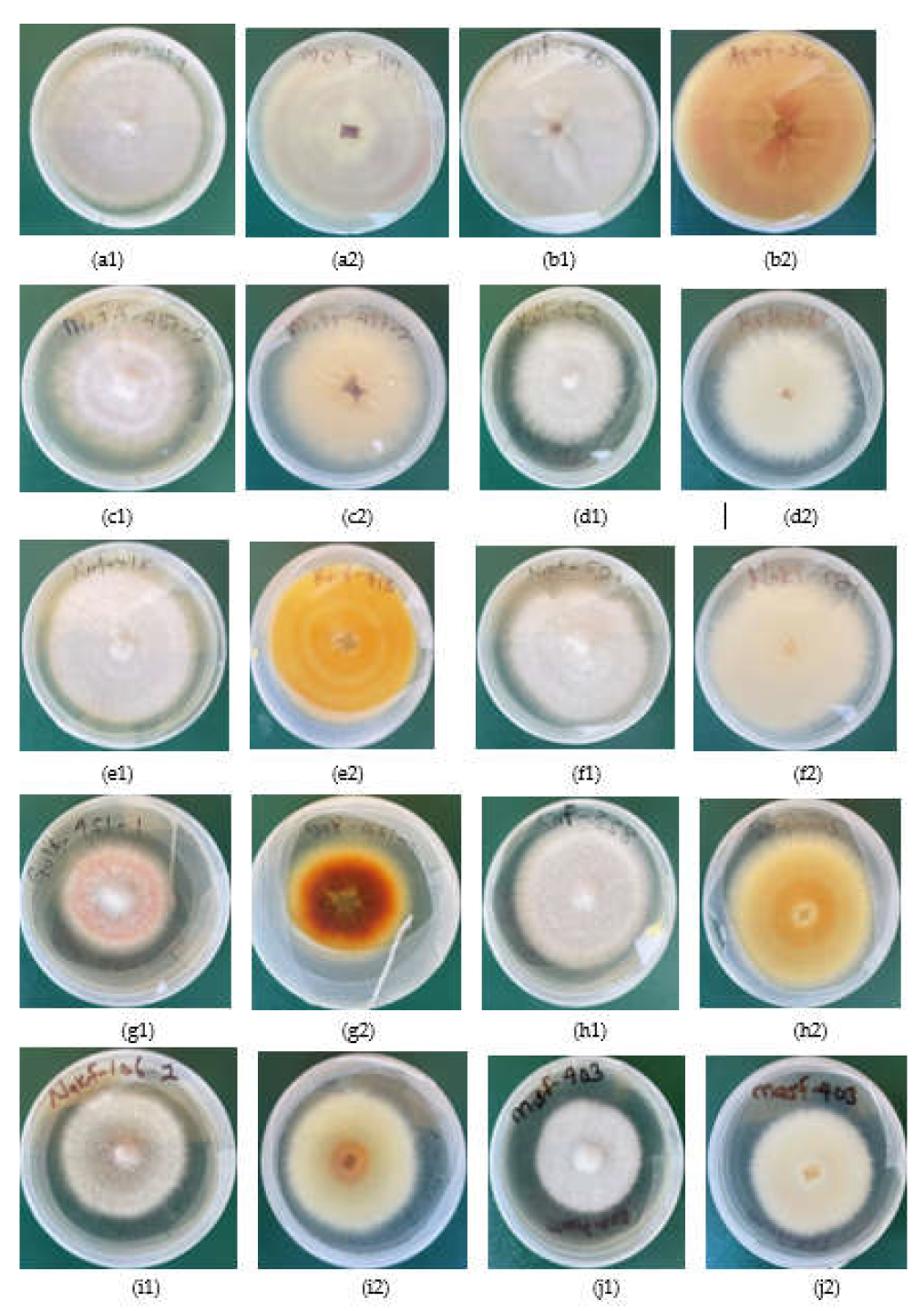
Figure 4.
Symptoms caused by some of the strains during pathogenicity by Erima et al. [8]. a- Strain NapF-375 identified as F. oxysporum, b- Strain ApaF-551 identified as F. equiseti, c- Strain NapF-520-2 identified as F. serpentimum.
Figure 4.
Symptoms caused by some of the strains during pathogenicity by Erima et al. [8]. a- Strain NapF-375 identified as F. oxysporum, b- Strain ApaF-551 identified as F. equiseti, c- Strain NapF-520-2 identified as F. serpentimum.

Figure 5.
Microscopic features of Fusarium species strains at x40 magnification; a- F. oxysporum, b- F. solani, c- F. falciforme, d- equiseti, e- F. brachygibosum, f- C. rhizophaga, g- F. fredkrugeri, h- F. subflagellisporum, i- F. fabacearum, j- F. delphinoides, k- F. commune and l- F. equiseti macro and micro conidia. The scale in the pictures represents 100µm.
Figure 5.
Microscopic features of Fusarium species strains at x40 magnification; a- F. oxysporum, b- F. solani, c- F. falciforme, d- equiseti, e- F. brachygibosum, f- C. rhizophaga, g- F. fredkrugeri, h- F. subflagellisporum, i- F. fabacearum, j- F. delphinoides, k- F. commune and l- F. equiseti macro and micro conidia. The scale in the pictures represents 100µm.
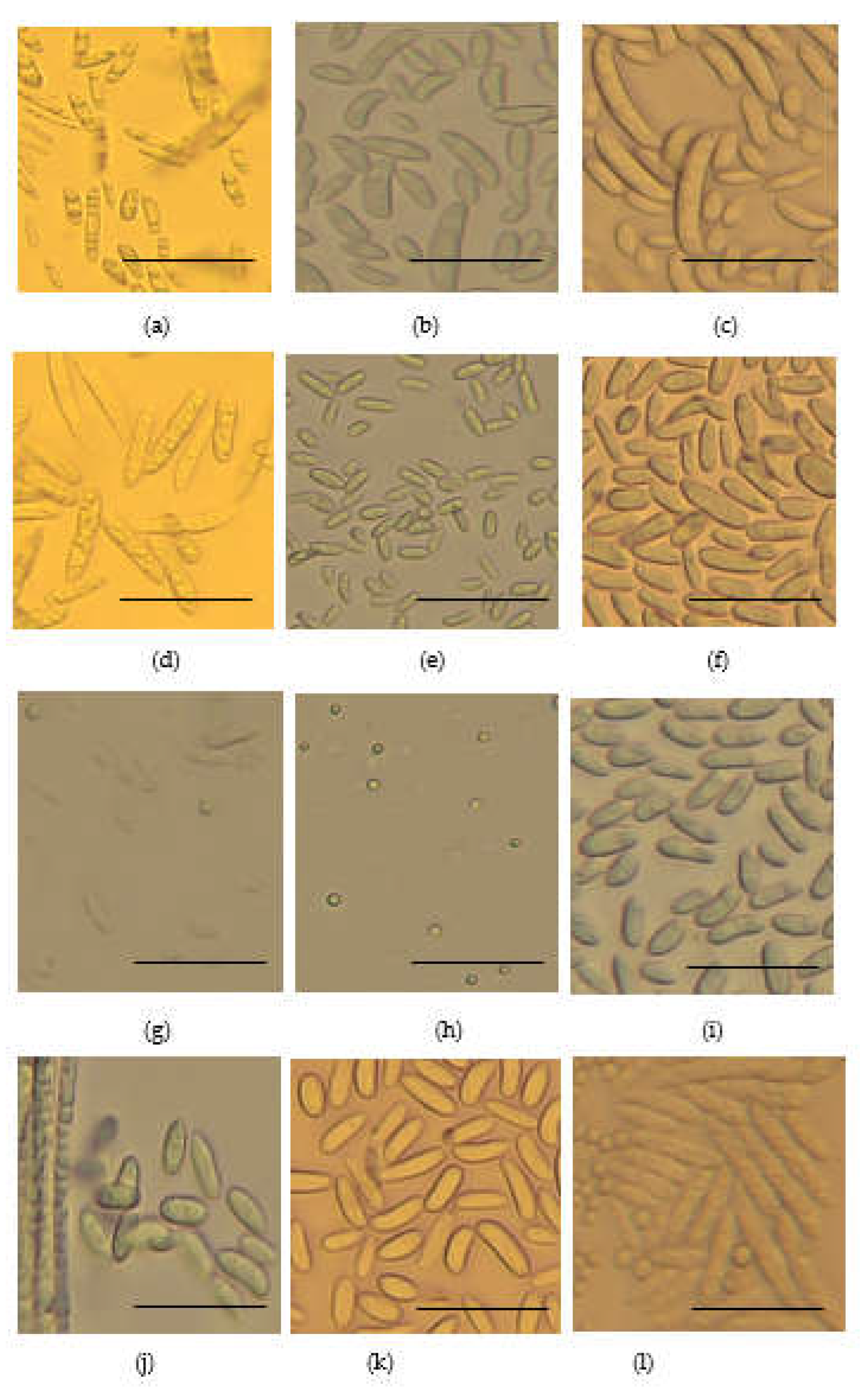

Table 1.
Fusarium species strains and accession numbers at the NCBI database. Gene regions that were amplified have accession numbers while those that failed to amplified do not have accession numbers.
Table 1.
Fusarium species strains and accession numbers at the NCBI database. Gene regions that were amplified have accession numbers while those that failed to amplified do not have accession numbers.
| S/no | Strains | Agroecology | Species | Accession numbers | ||
|---|---|---|---|---|---|---|
| TEF1-α | Β tubulin | ITS | ||||
| 1 | MbrF-119 | WMFS | F. fabacearum | PQ497180 | PQ497177 | PQ363745 |
| 2 | NakF-106-2 | NEDL | F. oxysporum | PQ497191 | PQ497142 | PQ363764 |
| 3 | KabF-103 | SWH | F. oxysporum | PQ497198 | PQ497143 | PQ363757 |
| 4 | SheF-250-1 | WMFS | F. oxysporum | PQ497207 | PQ497144 | PQ363766 |
| 5 | GomF-492 | LVC | F. oxysporum | PQ497213 | PQ497145 | PQ363773 |
| 6 | KabF-108-1 | SWH | F. oxysporum | PQ497226 | PQ497146 | PQ363790 |
| 7 | KamF-290-2 | WMFS | F. oxysporum | PQ497234 | PQ497147 | PQ363797 |
| 8 | LweF-507 | LVC | F. oxysporum | - | PQ497148 | PQ363804 |
| 9 | MubF-442-2 | LVC | F. oxysporum | PQ497182 | PQ497148 | - |
| 10 | MitF-489 | LVC | F. oxysporum | PQ497181 | PQ497150 | PQ363746 |
| 11 | AmuF-513-3 | NEDL | F. falciforme | PQ497183 | PQ497151 | - |
| 12 | MubF-463 | LVC | F. oxysporum | PQ497184 | PQ497152 | PQ363747 |
| 13 | KabF-114 | SWH | F. solani | PQ497185 | PQ497153 | - |
| 14 | GulF-451-1 | NMFS | F. solani | PQ497186 | PQ497154 | - |
| 15 | SheF-249 | WMFS | F. oxysporum | PQ497187 | PQ497155 | PQ363748 |
| 16 | KolF-563 | NMFS | F. brachygibbosum | PQ497188 | PQ497138 | PQ363749 |
| 17 | LweF-504 | LVC | F. oxysporum | PQ497189 | PQ497176 | PQ363750 |
| 18 | MubF-463-1 | LVC | F. oxysporum | PQ497190 | PQ497156 | PQ363751 |
| 19 | KabF-109-1 | SWH | F. delphinoides | PQ497192 | - | PQ363752 |
| 20 | OyaF-541-3 | NMFS | F. falciforme | PQ497193 | - | - |
| 21 | MubF-234 | LVC | F. oxysporum | PQ497194 | PQ497175 | PQ363753 |
| 22 | KolF-557-4 | NMFS | F. equiseti | PQ497195 | PQ497174 | PQ363756 |
| 23 | LweF-497 | LVC | F. oxysporum | PQ497196 | PQ497173 | - |
| 24 | ApaF-548 | NMFS | F. solani | - | PQ497172 | PQ363755 |
| 25 | MitF-491-1 | LVC | F. falciforme | PQ497197 | PQ497171 | PQ363791 |
| 26 | NakF-521 | NEDL | F. equiseti | PQ497205 | PQ497164 | PQ363765 |
| 27 | MubF-462-2 | LVC | F. oxysporum | PQ497199 | PQ497170 | PQ363758 |
| 28 | ApaF-560 | NMFS | F. subflagellisporum | PQ497200 | PQ497169 | PQ363759 |
| 29 | NakF-520 | NEDL | F. equiseti | PQ497201 | PQ497168 | PQ363760 |
| 30 | MubF-465 | LVC | F. oxysporum | PQ497202 | PQ497167 | PQ363761 |
| 31 | ApaF-551 | NEDL | F. equiseti | PQ497203 | PQ497166 | PQ363762 |
| 32 | LirF-602-2 | NEDL | F. equiseti | - | - | PQ363763 |
| 33 | NakF-106 | NEDL | F. solani | PQ497204 | PQ497165 | PQ363764 |
| 34 | KyeF-323 | WMFS | F. solani | PQ497206 | PQ497163 | - |
| 35 | LweF-223 | LVC | F. equiseti | PQ497208 | - | - |
| 36 | KapF-372 | EH | F. oxysporum | - | PQ497162 | PQ363767 |
| 37 | SirF-349-1 | LVC | F. oxysporum | PQ497209 | PQ497161 | - |
| 38 | KamF-289 | WMFS | F. solani | - | - | PQ363768 |
| 39 | IbaF-270 | WMFS | F. oxysporum | PQ497210 | PQ497160 | PQ363769 |
| 40 | ApaF-546 | NMFS | F. equiseti | - | PQ497159 | PQ363770 |
| 41 | LweF-393 | LVC | F. oxysporum | PQ497211 | PQ497158 | PQ363771 |
| 42 | SirF-358 | LVC | F. falciforme | PQ497212 | PQ497157 | PQ363772 |
| 43 | KabF-113-2 | SWH | F. fabacearum | PQ497214 | PQ497141 | PQ363774 |
| 44 | BusF-258 | WMFS | F. oxysporum | - | PQ497140 | PQ363775 |
| 45 | MitF-490 | LVC | F. commune | PQ497215 | PQ497139 | PQ363776 |
| 46 | AmuF-518-2 | NEDL | F. equiseti | PQ497216 | PQ497137 | PQ363777 |
| 47 | KamF-493-3 | WMFS | F. oxysporum | PQ497217 | PQ497136 | PQ363778 |
| 48 | KirF-416 | WMFS | F. solani | PQ497218 | PQ497135 | PQ363779 |
| 49 | SirF-358-1 | LVC | F. falciforme | PQ497219 | PQ497134 | PQ363780 |
| 50 | NakF-102-2 | NEDL | F. solani | - | - | PQ363781 |
| 51 | KabF-91-1 | SWH | F. oxysporum | - | - | PQ363782 |
| 52 | BusF-255-1 | WMFS | F. oxysporum | PQ497220 | PQ497120 | PQ363783 |
| 53 | MubF-464-2 | LVC | F. oxysporum | PQ497221 | PQ497133 | PQ363784 |
| 54 | LweF-296 | LVC | F. commune | PQ497222 | PQ497119 | PQ363785 |
| 55 | NakF-520-1 | NEDL | F. serpentimum | PQ497223 | - | PQ363786 |
| 56 | MbarF-229 | WMFS | F. commune | PQ497224 | - | PQ363787 |
| 57 | ApaF-560-1 | NMFS | F. subflagellisporum | PQ497200 | PQ497121 | - |
| 58 | NakF-105-1 | NEDL | F. oxysporum | PQ497227 | - | - |
| 59 | MitF-487-2 | LVC | C. rhizophaga | PQ363792 | - | PQ363792 |
| 60 | MitF-481 | LVC | F. commune | PQ497225 | PQ497132 | PQ363789 |
| 61 | KyeF-320-2 | WMFS | F. solani | PQ497227 | PQ497131 | - |
| 62 | ApaF-548-2 | NMFS | F. solani | PQ497228 | - | - |
| 63 | SheF-250 | WMFS | F. oxysporum | PQ497207 | - | - |
| 64 | MasF-403 | WMFS | F. fredkrugeri | PQ497229 | - | - |
| 65 | MitF-491-2 | LVC | F. falciforme | PQ497230 | PQ497171 | PQ363788 |
| 66 | KirF-418 | WMFS | F. falciforme | PQ497231 | PQ497128 | PQ363793 |
| 67 | HoiF-385 | WMFS | F. oxysporum | PQ497232 | PQ497127 | PQ363794 |
| 68 | SirF-349-3 | LVC | F. oxysporum | PQ497233 | PQ497126 | PQ363795 |
| 69 | MitF-487 | LVC | F. oxysporum | - | PQ497129 | PQ363788 |
| 70 | MubF-466 | LVC | F. oxysporum | - | - | PQ363798 |
| 71 | ApaF-546 | NMFS | F. equiseti | - | PQ497159 | - |
| 72 | KolF-562 | NMFS | F. solani | PQ497236 | PQ497124 | PQ363799 |
| 73 | KamF-290 | WMFS | F. oxysporum | - | - | PQ363800 |
| 74 | LweF-496 | LVC | F. oxysporum | - | - | PQ363801 |
| 75 | KolF-562-1 | NMFS | F. solani | - | PQ497125 | - |
| 76 | NakF-375 | NEDL | F. oxysporum | PQ497237 | PQ497123 | PQ363802 |
| 77 | Apaf-551-1 | NMFS | F. brevicaudatum | PQ497233 | - | - |
| 78 | LweF-215 | LVC | F. oxysporum | PQ497179 | PQ497122 | PQ363805 |
| 79 | ApaF-560 | NMFS | F. oxysporum | PQ497178 | PQ497121 | - |
| 80 | HoiF-385-1 | WMFS | F. solani | PQ497219 | - | PQ363803 |
Table 2.
Average DSI, growth rate and microscopic structures of different Fusarium species from Ugandan agroecology zones.
Table 2.
Average DSI, growth rate and microscopic structures of different Fusarium species from Ugandan agroecology zones.
| S/no | Organism Name |
No. of strains | DSI (%) | Growth rate (cm/day) | Microscopic structues at x40 magnification |
|---|---|---|---|---|---|
| 1 | F. delphinoides | 1 | 46.8 | 0.96 | Rod shaped non septate macro conidia about 5 to 50µm lond and spherical micro conidia |
| 2 | F. solani | 13 | 36.3 | 0.79 | Sickle shaped non septate macro conidia about 5 to 50 µm long. |
| 3 | F. oxysporum | 37 | 44.4 | 0.79 | Rod shaped sepate micro conidia about 20 to 50µm long |
| 4 | F. equiseti | 9 | 47.3 | 0.7 | Rod shaped non sepate macro conidia about 50 to 100µm long. Spherical micro conidia 2 to 10µm long |
| 5 | C. rhizophaga | 1 | 31.3 | 0.37 | Oval and rod shaped macro conidia about 10 to 50µm long |
| 6 | F. subflagellisporum | 2 | 66.6 | 1.2 | Sperical micro conidia about 2 to 5µm long. No macro conidia |
| 7 | F. fabacearum | 2 | 40.24 | 0.70 | Rod shaped non septate macro conidia about 5 to 50µm long. Spheical micro conidia |
| 8 | F. falciforme | 8 | 32.3 | 0.74 | Rod shaped macro conidia about 50 to 150µm long. Spherical micro conidia |
| 9 | F. brachygibbosum | 1 | 65.8 | 0.87 | Oval non sepate macro conidia about 10 to 30µm long |
| 10 | F. brevicaudatum | 1 | 59.3 | 0.6 | Isolates in storage failed to regenerate for microscopy |
| 11 | F. commune | 4 | 62.5 | 0.88 | Rod shaped nonsepate macro conidia about 5 to 50µm, |
| 12 | F. serpentimum | 1 | 45.1 | 0.17 | Isolates in storage failed to regenerate for microscopy |
| 13 | F. frekrugeri | 1 | 40.3 | 0.87 | Oval non septate macro conidia up to about 40µm long. Spherical micro conidia 5 to 10µm |
*Disease severity indecs (DSI) and growth rate data were obtained from Erima et al. [8].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



