
Submitted:
29 August 2023
Posted:
31 August 2023
You are already at the latest version
Abstract
Sarocladium and Fusarium species are commonly identified as causal agents of rice sheath rot disease worldwide. However, limited knowledge exists about their genetic, pathogenic, and toxigenic diversity in sub-Saharan African (SSA) countries, where an increasing incidence of this disease has been observed. This study aimed at identifying, characterizing, as well as assessing the genetic, pathogenic, and toxigenic diversity of the pathogens associated with rice sheath rot disease in Mali, Nigeria, and Rwanda. In this study seventy fungal isolates were obtained from rice plants displaying disease symptoms on research and farmer’s fields in Mali, Nigeria, and Rwanda. Thus, an extensive comparative analysis was conducted to assess their genetic, pathogenic, and toxigenic diversity. The Fusarium spp. were characterized using the translation elongation factor (EF-1α) region, while a concatenation of Internal Transcribed Spacer (ITS) and Actin-encoding regions were used to resolve Sarocladium species. Phylogenetic analysis revealed four Fusarium species complexes. The dominant complex in Nigeria was the Fusarium incarnatum-equiseti species complex (FIESC), comprising F. hainanense, F. sulawesiense, F. pernambucatum, and F. tanahbumbuense, while F. incarnatum was found in Rwanda. The Fusarium fujikuroi species complex (FFSC) was predominant in Rwanda and Mali, with species such as F. andiyazi, F. madaense, and F. casha in Rwanda, and F. annulatum and F. nygamai in Mali. F. marum was found in Nigeria. Furthermore, Fusarium oxysporum species complex (FOSC) members, F. callistephi and F. triseptatum, were found in Rwanda and Mali, respectively. Two isolates of F. acasiae-mearnsii, belonging to the Fusarium sambucinum species complex (FSAMSC) were obtained in Rwanda. Isolates of Sarocladium which were previously classified in three phylogenetic groups were resolved into three species which are: attenuatum, oryzae and sparsum. S. attenuatum was dominant in Rwanda, while S. oryzae and S. sparsum were found in Nigeria. Also, the susceptibility of FARO44, a rice cultivar released by Africa Rice Centre (AfricaRice) was tested against isolates from the four Fusarium species complexes and the three Sarocladium species. All isolates evaluated could induce typical sheath rot symptoms albeit with varying disease development levels. In addition, liquid chromatography-tandem mass spectrometry (LC-MS/MS) was used to determine variation in the in vitro mycotoxins of the Fusarium species. Regional differences were observed in the in vitro mycotoxins profiling. Out of the forty-six isolates tested, nineteen were able to produce one to four mycotoxins. Notably, very high zearalenone (ZEN) production was specific to the two F. hainanense isolates from Ibadan-Nigeria, while Fusarium nygamai isolates from Mali produced high amounts of fumonisins. To the best of our knowledge, it seems this study is the first to elucidate genetic, pathogenic, and toxigenic diversity of Fusarium species associated with the rice sheath rot disease complex in selected countries in SSA.
Keywords:
Fusarium species
; Sarocladium species
; rice
; pathogenic variability
; genetic diversity
; mycotoxins
; sub-Saharan Africa
1. Introduction
Rice (Oryza sativa) holds significant economic importance in Africa, with Nigeria being one of the leading contributors to the continent’s global rice production share (4.2%), accounting for 24% [1]. Despite this, Nigeria remains the second-largest importer of rice worldwide, trailing only behind China. In 2018 alone, Nigeria imported approximately 3 million metric tons of milled rice, struggling to meet its demand deficit for the past decade [2]. Mali, a landlocked country in West Africa, ranks fifth among African nations in terms of rice production. Its rice production is being managed through irrigated systems connected to the Niger River. In Rwanda, the demand for rice is estimated at 145,000 tons per year, while national supply accounts for about 40%, creating a 60% deficit that is met through imports [93]. While the rice cultivation area is expanding in Africa, the average yields (2.35 tons/ha in 2021) are low when compared to Asia (4.95 tons/ha in 2021) [1]. Rice production is constrained by various factors including biotic stresses such as pests and diseases. Africa has recorded a steady and substantial increase in the incidence of rice diseases such as Rice Yellow Mottle Virus (RYMV), Rice blast (Pyricularia oryzae), Bacterial Leaf Blight (Xanthomonas oryzae pv oryzae), Bacterial Leaf Streak (Xanthomonas oryzae pv oryzicola), and Rice Stripe Necrosis Virus (RSNV) [3,4,5,6,7].
Rice sheath rot is an emerging disease worldwide [8]. Sarocladium oryzae, [9], formerly Acrocylindrium oryzae, was the first organism to be associated with rice sheath rot symptoms in Taiwan in 1922 [10] Sarocladium attenuatum was originally described as a distinct species causing rice sheath rot, was then considered a synonym of Sarocladium oryzae [11], but has recently been re-established as a separate species causing rice sheath rot in Taiwan [12]. These authors also described a third species that causes sheath rot symptoms on rice called S. sparsum, which is closely related to S. oryzae and S. attenuatum. Sarocladium has been associated with rice sheath rot in thirty-eight countries [13].
Besides Sarocladium species, Fusarium spp. have been associated with the rice sheath rot complex. These mainly comprise isolates in the F. fujikuroi species complex (FFSC) including F. proliferatum, F. verticillioides, F. incarnatum and F. fujikuroi [14,15,16,17,18,19]. In addition, various bacterial species cause rice sheath rot symptoms. The most important one is Pseudomonas fuscovaginae which is known to cause sheath brown rot of rice at high altitudes. In Africa, this bacterium has been reported in Burundi [8] [20] and Madagascar.
In West Africa, rice sheath rot has been reported in Cote D’ivoire, Gambia, Niger, Nigeria, and Senegal [13]. However, no causative fungal strains were isolated and no detailed scientific information was provided except for Nigeria [21] where S. attenuatum was first reported in Nigeria as one of the causes of grain discoloration on rice [21]. In addition, an inhibitory effect of S. oryzae on seed germination was later observed [22]. Most information pertaining to the occurrence of S. oryzae in Africa relating to stored, marketed and field seeds especially with respect to mycotoxigenic potentials was enumerated by [23].
Rice sheath rot can cause high yield losses of 20-80% [24,25,26]. Furthermore, an extensive survey of rice fields across West Africa enabled the identification of sheath rot symptoms in Mali and Nigeria (AfricaRice disease database). Although yield losses due to the sheath rot disease has not been estimated in Mali and Nigeria, a field survey conducted in 2011 and 2013 revealed high incidence and severity of the disease in Rwanda [27].
Mycotoxin contamination of cereal products poses a serious concern for animal and human health. Several studies have reported Fusarium species as the major producers of mycotoxins contaminating cereals including rice [28,29,30,31,32,33,34]. In the African region, previous studies have reported several mycotoxins being synthesized by Fusarium species isolated from rice as a serious health threat to producers and consumer [35,36,37,38,39,40].
The comprehensive information regarding the incidence and distribution of sheath rot disease is the aspect notably lacking. Additionally, there has been a notable absence of research examining the genetic, pathogenic, and toxigenic variability of pathogens associated with this disease in East and West Africa. Acquiring this crucial information will offer valuable insights for disease control and enhance management strategies for breeding programs. Therefore, this study aimed at identifying, characterizing, as well as assessing the genetic, pathogenic, and toxigenic diversity of the pathogens associated with rice sheath rot disease in Mali, Nigeria, and Rwanda.
2. Materials and Methods
2.1. Collection of Samples
Samples were collected from rice research programs and farmer’s fields in Mali, Nigeria, and Rwanda (Figure 1). Naturally infected whole rice plants with sheath rot symptoms having sheath browning, necrosis, grain emptiness and rotting as indicated in Figure 2 were collected. Samples from farmer’s field were collected at the office of the rural development, Selingue village near Bamako, Mali and from two fields located at Ibadan, Oyo State and Katcha near Badeggi, Niger State in Nigeria during the 2017 rice growing season. Samples were randomly collected 25 m apart at each location. The samples collected were conserved in dry paper bags, while hands were disinfected with 70% alcohol after each sampling. Samples were later stored in dry bags in the refrigerator at 4°C in the laboratory. Isolates earlier collected from Rwanda in 2011 and 2013 were also included in this study. Agro-climatological details of the selected three countries with their various agroecologies are presented in Table 1. Isolation and identification of pathogens was carried out at the Phytopathology laboratory of Ghent University, Belgium.
2.1.1. Isolation and Purification of Sheath Rot-Associated Isolates
Infected sheath and seed samples showing symptoms of sheath rot were surface - sterilized in 2% sodium hypochlorite for two minutes and then rinsed thrice in sterile distilled water. They were drained using sterile paper towels, and thereafter cut into small pieces of about 0.5 cm2 and plated on 90 mm diameter Petri dishes containing Potato Dextrose Agar (PDA). The cultured Petri dishes were incubated at 28°C in darkness for 7-14 days. Cultures were further purified by plating on fresh PDA.
2.1.2. Identification of Pathogens
Identification of fungal pathogens was done based on their typical structure and basic characters as described by Barnett and Hunter (1972). The incidence and diversity of fungi was observed and recorded. After 5 days, all plates were examined under a compound microscope for the presence of several fungal pathogens. Conidia of these fungi were mounted on glass slides in water and examined under a compound microscope for identification at genus level. For Fusarium species, pure cultures were plated and stored on PDA slants at room temperature and maintained at -80°C with 40% glycerol. A similar method of storage was used for Sarocladium species except for the use of 20% glycerol.
2.2. Molecular Characterization of Isolates
2.2.1. DNA extraction, amplification, and sequencing
Fungal isolates were grown on potato dextrose broth (PDB) at 28 °C for seven days. Mycelia mats were harvested by filtration, dried by blotting using sterile paper towels, frozen in liquid nitrogen, and pulverized using a tissue lyser (MM400, Retsch GmbH, Haan, Germany).
Genomic DNA was extracted using the Wizard Genomic DNA Purification Kit (Promega). Quantification and purity were determined using Nanodrop 3000 (Thermo Scientific, Asheville, NC, USA) and diluted to a concentration of 20 ng µL-1.
Fusarium isolates were further identified by amplifying the Translation Elongation Factor (EF-1α), using primer pair TEF-1-F (5’-ATG GGT AAG GAA GAC AAG AC-3’) and TEF-2-R (5’-GGA AGT ACC AGT GAT CAT GTT-3’) [41]. PCR reactions were done in 25 µL of a solution consisting of 2 µL genomic DNA (100 ng µL-1), 5 µL PCR buffer (5x; Promega), 5 µL Q solution (Qiagen), 0.5 µL dNTPs (10 mM; Fermentas GmbH), 1.75 µL of each primer (10 µM), 0.15 µL Taq DNA polymerase (5 units µL−1; Fermentas GmbH) and 8.85 µL ultrapure sterile water. Amplification was performed with initial denaturation step at 94 °C for 1 min, followed by 35 cycles of denaturation at 94°C for 30 s, primer annealing at 53°C for 45 s and extension at 72 °C for 1 min. Cycling ended with a final extension step at 72 °C for 5 min [41]. The amplicons were separated by horizontal electrophoresis using 1.5 % agarose gels in TAE-buffer at 100 V for 25 min and visualized by ethidium bromide staining on a UV trans illuminator. Amplified products were purified with exosap and sequenced by LGC Genomics GmbH (Berlin, Germany) using Sanger sequencing.
For Sarocladium isolates, two genomic regions, the Internal Transcribed Spacer (ITS) and Actin, were amplified and sequenced. For the ITS region, primers ITS1 (5’-TCC GTA GGT GAA CCT GCG G-3’) and ITS4 (5’- TCC TCC GCT TAT TGA TAT GC -3’) were used (White et al., 1990). The actin region was amplified using ACT1 (5’- TGG GAC GAT ATG GAG AAG ATC TGG CA -3’) and ACT4 (5’- TCG TCG TAT TCT TGC TTG GAG ATC CAC AT-3’) [42].
For both primer pairs, PCR reactions were done in 25 µL of a solution consisting of 2 µL genomic DNA (100 ng µL-1), 5 µL PCR buffer (5x; Promega), 0.5 µL dNTPs (10 mM; Fermentas GmbH), 1.75 µL of each primer (10 µM), 0.15µL Taq DNA polymerase (5 units µL−1, Fermentas GmbH) and 13.85µL ultrapure sterile water. Amplification was performed using a Flexcycler PCR Thermal Cycler (Analytik Jena). For ITS amplification, the thermal profile consisted of an initial denaturation step at 94 °C for 10 min, followed by 35 cycles of denaturation at 94 °C for 1 min, primer annealing at 55 °C for 1 min and extension at 72 °C for 1 min. Cycling ended with a final extension step at 72 °C for 10 min [43]. ACT fragments were amplified using an initial denaturation step at 94°C for 1 min, followed by 39 cycles of denaturation at 94 °C for 45 sec, primer annealing at 59 °C for 30 sec and extension at 72 °C for 30 sec. Cycling ended with a final extension step at 72 °C for 8 min [42].
2.2.2. Phylogenetic Analysis
The nucleotide sequences generated by the forward and reverse primers were used to obtain consensus sequences after editing via BioEdit version 7.2.5. [44]. From each duplicate identical sequence, a representative sequence from each identical set of sequence was compared to other sequences available at GenBank. Sequences were first aligned via muscle alignment in Mega V.11. [45] after which a maximum-likelihood tree was constructed based on the matrix of pairwise distances obtained using the General Time Reversible (GTR) Model. Reference sequences of Fusarium (Table 5) and Sarocladium (Table 6) species representing the three countries were used for phylogenetic analysis. For Fusarium, Phylogenetic tree was constructed based on maximum-likelihood inferred from partial EF-1α sequences of four Fusarium species complexes using IQ-Tree with GTR + G + I model and annotated using iTol software [46]. Cylindrocarpon sp. AC2011 strain CPC 13531 was used as an outgroup. However, for Sarocladium characterization, concatenated alignments of ITS and ACT region were performed after which a single phylogenetic tree was generated. To root the tree, Sarocladium zeae strain CBS 800.69 was used as an outgroup.
2.3. Pathogenicity Assay
Location of the isolates, and genetic groupings were used to select a subset of twenty-nine isolates for pathogenicity studies on rice plants. For pathogenicity tests with Fusarium species, representative isolates from the four Fusarium species complexes recorded in all the locations (FIESC, FFSC, FOSC, FSAMSC) were used for rice inoculation. To evaluate the pathogenicity of Sarocladium species, thirteen isolates comprising S. attenuatum (3), S. oryzae (6), and S. sparsum (4) were used. Indica rice cultivar (FARO 44) released by Africa Rice Center was used for the assay.
Inoculum was prepared according to the standard grain inoculum technique [47]. Briefly, rice grains were soaked in water for 60 min, excess water was removed, and the grains were autoclaved twice on two different days. For 4 g of rice grains, 1 plug (diameter = 5 mm) from the edge of a 14-day-old fungal colony was added together with 1 ml of sterile distilled water. Every two days, the grain inoculum was shaken to prevent the formation of clumps. After 10 days of incubation at 28°C, the inoculum was fully colonized.
The rice seeds were dehulled and surface sterilized in 2% sodium hypochlorite solution for 25 min, rinsed five times in sterile distilled water, and placed in Petri dishes containing sterile moistened filter papers (Whatman, grade 3). Following seedling emergence, six seedlings were transplanted into perforated plastic trays (22 x 15 x 6 cm) containing potting soil (Structural; Snebbout, Kaprijke, Belgium). Plants were watered daily, fertilized weekly with 0.2% iron sulphate and 0.1% ammonium sulphate, and maintained in a growth chamber (28°C, 60% relative humidity). Six-week-old plants were used for the inoculation.
One fully colonized fungal grain was introduced in the junction point between the sheath of the second youngest plant leaf and the stem. Inoculation points were covered with moist cotton wool and wrapped with parafilm to maintain humidity. High humidity was maintained for 24 h post inoculation by incubating in a controlled room at 28°C day and night, 16/12 light regime, and 85 % relative humidity. Subsequently, incubation was maintained at 65 % relative humidity, temperature, and light regime as above for 2-10 days. The disease development was evaluated eight days after inoculation by measuring the lesion length on the flag leaf sheath. Each treatment was replicated thrice while the experiment was conducted once.
2.4. Statistical Analysis
Lesion length was used as a measure of isolate virulence on rice plants. Since the conditions of normality were not met, a non-parametric analysis was carried out. Lesion length of the infected sheath were quantified using Kruskal-Wally’s Rank Sum test followed by a post hoc Mann-Whitney test. Statistical significance was defined as P= 0.05.
2.5. Mycotoxin analysis
2.5.1. Culture Preparation
Pure cultures of seventy-seven identified isolates (Fusarium - 46, Sarocladium – 31) were sub-cultured on Petri dishes with PDA. The medium was poured into 90 mm Petri dishes. Two mm of clean and pure cultured isolates were sub-cultured on fresh PDA plates and incubated at 25 °C for 3 weeks. Each isolate was grown in triplicates.
2.5.2. Reagents and Standards
Ethyl acetate and dichloromethane (DCM) were purchased from (Thermofisher scientific, Merelbeke, Belgium). Analytical grade formic acid (100%) and ammonium acetate were from (Merck, Darmstadt, Germany). Purified water was from the Arium pro VF system (Millipore, Belgium). LC-MS grade acetic acid and methanol were from (Biosolve, Valkenswaard, the Netherlands).
Certified mycotoxin standard solutions, more specifically aflatoxin mix (AFB1, AFB2, AFG1 and AFG2), deoxynivalenol (DON), fumonisin mix (FB1 and FB2), nivalenol (NIV), neosolaniol (NEO), OTA, T2, HT2, 3-acetyldeoxynivalenol (3-ADON), diacetoxyscirpenol (DAS), 15- acetyldeoxynivalenol (15-ADON), fusarenon-X (F-X), sterigmatocystin (STC), zearalenone (ZEN) and deepoxy-deoxynivalenol (DOM) were purchased from Biopure (RomerLabs, Getzersdorf, Austria). Fumonisin B3 (FB3) and enniatin B (ENN B) were obtained from Fermentek (Jerusalem, Israel). Alternariol (AOH), alternariol monomethylether (AME), were purchased from Sigma-Aldrich (Bornem, Belgium) and roquefortine (ROQ-C) from Alexis Biochemicals (Enzo Life Sciences BVBA, Zandhoven, Belgium).
Working solutions were prepared by diluting the stock solutions in methanol and stored at -20°C. A standard mixture consisting of the above mycotoxins (without DOM) in a concentration range between 0.5 ng/µl and 40 ng/µl was prepared as well and stored at -20°C.
Mobile phase A (94% water, 5% methanol, 1% acetic acid and 5mM ammonium acetate) and mobile phase B (97% methanol, 2% water, 1% acetic acid and 5mM ammonium acetate) were prepared.
2.5.3. Sample Preparation and Extraction
The extraction process started with the preparation of the quality control samples. Briefly, three plugs each of blank agar (uninoculated) were removed and placed into each of the three 50 ml Falcon tubes (spike 1, spike 2, and blank) and were macerated into pieces using a sterile scalpel blade. Then 50 µL DOM internal standard (50 ng/µl) was added into each tube, after which 25 µL and 100 µL of the standard mixture were added to spike 1 and spike 2, respectively. The mixtures (spikes and blank) were left in the dark for 15 minutes.
Following the control sample preparation, Fusarium mycotoxins were extracted from pure cultures of different isolates by using a sterile 9 mm cork borer and scalpel to take three plugs (2 sides + centre). The plugs were transferred into 50 ml Falcon tubes and macerated into pieces using a sterile scalpel blade. Then 50 µL DOM internal standard (50 ng/µl) was added into each tube and left in the dark for 15 minutes. The samples together with the quality control samples were extracted by adding ethyl acetate + 1% formic acid. The content was agitated gently on a vertical shaker for 20 min and centrifuged at 3000 g for 15 min. Then a folded filter paper (VWR International, Zaventem, Belgium) moistened with ethyl acetate + 1% formic was placed on a new extraction tube to collect the upper layer of the filtrate. Thereafter, 5 ml of dichloromethane (DCM) was added to each of the samples. The mixtures were agitated on a vertical shaker for 20 min and centrifuged at 3000 g for 15 min. Following centrifugation, the bottom layer (DCM phase) was collected in the same Falcon tube with the same filter paper. The filtrates were evaporated to dryness at 40 °C under a gentle nitrogen stream. The dissolved residue was reconstituted in 200 µl injection solvent (60% mobile phase A and 40% mobile phase B), well vortexed, and ultra centrifuged for 5 min at 10000 rpm. Finally, 100 µl of the filtrates were transferred into HPLC vials for LC-MS/MS analysis.
2.5.4. Multi-metabolite analysis (LC-MS/MS)
The samples were analysed using a Quattro Premier XE triple quadrupole mass spectrometer coupled with a Waters Acquity UPLC system (Waters, Milford, MA, USA).
Liquid chromatography conditions and MS parameters were followed as described by [48].The analytical column used was a symmetry C18, 5 µm, 2.1 x 150 mm with a guard column of the same material (3.5 µm, 10 mm x 2.1 mm) (Waters, Zellik, Belgium) kept at room temperature. The injection volume was 10 µl. Capillary voltage was set at 3.2 kV with a source block temperature and desolvation temperature of 120 and 400 °C respectively. Data processing was done using the Masslynx and Quanlynx software.
3. Results
3.1. Sampling and Isolation
Information on Fusarium isolates obtained from diseased rice plants in Nigeria, Mali and Rwanda is presented in Table 2. Of the 46 Fusarium isolates evaluated in this study, nine isolates were obtained from seeds, while 37 isolates originated from the rice sheath. The highest number was obtained from Rwanda (24 isolates), followed by Nigeria (15 isolates), while Mali (seven isolates) had the least.
Information on Sarocladium isolates obtained from diseased rice plants in Nigeria, Mali and Rwanda is given in Table 3. Out of the 24 Sarocladium isolates obtained, four were from seeds and 20 from the rice sheath. The highest number of isolates were obtained from Nigeria (nine from Katcha, seven from Ibadan), six isolates were from Rwanda, and only two from Mali (Table 3).
Altogether, seventy isolates comprising Sarocladium species (24) and Fusarium species (46) were obtained. Four ITS characterized CBS Sarocladium species were added, and we sequenced their actin region in this study (Table 2).
3.2. Phylogenetic Analysis of Fusarium and Sarocladium-like spp.
3.2.1. Fusarium Species
Partial sequences of TEF-1a gene revealed the identity of all the 46 Fusarium isolates used. Similarities to DNA sequences in the Fusaroid-ID and GenBank database ranged from 99–100%. Members of four species complexes were identified: F. fujikuroi species complex (FFSC – 48%), F. incarnatum-equiseti species complex (FIESC – 35%), F. oxysporum species complex (FOSC – 13%) and F. sambucinum species complex (FSAMSC – 4%) (Table 2). The phylogenetic analysis of the 46 Fusarium isolated is presented in Figure 3. The origin and Genbank accession numbers of the reference isolates used in given in Table A1.
FFSC isolates from Rwanda clustered with F. andiyazi (7 isolates), F. madaense (6 isolates) and F. casha (1 isolate). FFSC isolates from Mali were identified as F. nygamai (5 isolates) and F. annulatum (1 isolate), while in Nigeria, 2 F. marum isolates were found.
FIESC isolates were found in Nigeria and Rwanda and belong to five different species (sulawesiense, pernambucatum, tanahbumbuense, hainanense, and incarnatum) previously classified from rice, cereals, insects, and human samples [31], [35], [36], [50,51,52,53] (Figure 3). Eight of our FIESC isolates were nested within the F. sulawesiense clade including 7 isolates from Ibadan and an isolate from Katcha. One isolate from Ibadan clustered with members of F. pernambucatum, two isolates from Katcha nested within the F. tanahbumbuense clade, and two isolates from Ibadan nested within the F. hainanense group. Three isolates from Rwanda were found in F. incarnatum (Figure 3). None of our isolates clustered with the F. equiseti species clade (Figure 3).
The FOSC was found in Rwanda and Mali with members belonging to F. callistephi (5 isolates from Rwanda), and F. triseptum (1 isolate from Mali). Two isolates belonging to F. acasia-mearnsii in the FSAMSC were obtained from Rwanda.
3.2.2. Sarocladium Species
Thirty-one S. oryzae-like isolates were used for phylogenetic analysis (Nigeria=16, Mali=2, Rwanda=6, reference isolates =7). Partial sequences and concatenation of both ITS and ACT regions showed the identities of all the isolates. They were further subjected to BLASTn comparison with isolates in GenBank. Results revealed that all the 31 isolates had 98 -100% identity with Sarocladium species. A concatenated tree, in which reference sequences from GenBank were included (see Table A2), clearly delineated the Sarocladium isolates into three distinct phylogenetic groups with high bootstrap values (Figure 4). Most isolates from Nigeria (11 of 16), one isolate from Mali (SEMA0013A) and one isolate from Rwanda (RFRG2) clustered together with reference isolates CBS 180.74 from India, CBS 361.75 from Kenya, and CBS120.817 from Panama and were identified as S. oryzae. The second Mali isolate (SEMA0029) and five of the six Rwandan isolates clustered with reference isolates CBS 101.61 from Mexico and CBS 399.73 from India and belong to the S. attenuatum lineage. Five isolates from Nigeria clustered with reference isolate CBS 414.81 from Nigeria and the S. sparsum isolate 18042 from Taiwan (Figure 4). They mostly occurred in Ibadan-Nigeria (DS) except for an isolate (BDNG0025) found on infected seed in Katcha (SGS). Finding a substantial number of this group in Ibadan was not strange because it clustered with a reference isolate CBS 414.81 of Ibadan origin, collected, and reported [21], and later deposited into the GenBank [42], and recently reclassified as S. sparsum.
3.3. Pathogenicity Testing
Representative isolates from the dominant Fusarium groups recorded in Nigeria, Rwanda and Mali were used for pathogenicity testing on the FARO 44 rice variety. All the four Fusarium species complexes could induce rice sheath rot symptoms on the rice cultivar, but the degree of virulence of all the Fusarium species tested showed significant variations on the rice cultivar (Figure 5). Specifically, one of the two isolates of F. marum IBNGF0016 from Ibadan in Nigeria had the highest disease severity on FARO 44 followed by the second F. marum isolate IBNGF0012 and an FIESC isolate F. sulawesiense BDNGF0002 from Katcha in Nigeria. On the contrary, F. nygamai originated from Mali and one of the FIESC isolate F. tanahbumbuense BDNGF0001 from Ibadan-Nigeria were the least virulent isolates.
For the pathogenicity tests with Sarocladium species, disease evaluation at 8 days post inoculation (DPI) showed that all the thirteen isolates tested could induce typical sheath rot symptoms on FARO 44 albeit with varying disease development levels (Figure 6). Isolates affiliated with S. sparsum, all of Nigeria origin, were the most aggressive (p < 0.05). In contrast, isolates affiliated with S. oryzae and S. attenuatum were less virulent (Figure 6). S. oryzae isolate IBNG0011 from Nigeria is the most aggressive among the group while SEMA0013A from Mali appears to be the least virulent.
3.4. Mycotoxin Profiling (In vitro)
Forty-six Fusarium and thirty-one Sarocladium species obtained from rice with sheath rot symptoms were investigated for multi-mycotoxin production using LC-MS/MS . None of the Sarocladium isolates produced mycotoxins. Our results revealed that the Fusarium species were able to produce eight mycotoxins. The most detected mycotoxins include type A (DAS and NEO) and B (NIV and FUS-X) trichothecenes which were produced by FIESC isolates obtained across the two regions of Nigeria albeit at different concentrations (Table 4). Furthermore, zearalenone (ZEN) at a very high concentration of 26,173 and 32,529 µg/kg was detected in the two Ibadan-Nigerian F. hainanense isolates IBNGF0005 and IBNGF0003, respectively. Two Rwandan isolates F. madaense RFRM18 and F. acasiae-mearnsii RFRM19 produced ZEN at the concentration of 1349 and 329 µg/kg, respectively. The predominant mycotoxins detected from Malian F. nygamai isolates were fumonisins (FB1, FB2, and FB3), which occurred at a very high concentration. FB1 concentrations ranged from 53,118 – 141,102 µg/kg followed by FB2 ranging from 3,391 - 5,122 µg/kg while FB3 occurred at a lower concentration which ranged between 355 – 692 µg/kg. Besides F. nygamai, F. annulatum SEMAF17-225A from Mali produced DAS, FUS-X, NIV, and NEO at different concentrations (Table 4).
4. Discussion
This study provides new insights into the genetic, pathogenic, and toxigenic diversity of Fusarium and Sarocladium species associated with rice sheath rot disease in SSA. Molecular characterization using the EF-1α gene enabled the delineation of Fusarium isolates into four distinct Fusarium species complexes, whereas concatenation of ITS and ACT sequences delineated Sarocladium into three species. It was discovered that Fusarium species are the dominant species associated with rice sheath rot disease in Mali and Rwanda, while in Nigeria, Fusarium and Sarocladium species were equally represented. Phylogenetic analysis showed that isolates grouped differently according to their geographical location (Figure 3 & 4).
Four Fusarium species complexes (FIESC, FFSC, FOSC, and FSAMSC) were found to be associated with rice sheath rot in SSA. Previous research has resolved FIESC species as a mere complex indicated by numbers, but current studies further updated them according to the new nomenclature well elucidated from numbers to names [49,50,51]. Additionally, they were characterized using the recently updated Fusarium ID database (Fusarioid-ID). Dominant in our findings were members of the FIESC previously classified from rice, cereals, insects, and human samples [29,33,52,53,54,55] which include F. sulawesiense (FIESC 16), F. pernambucatum (FIESC 17), F. tanahbumbuense (FIESC 24), F. hainanense (FIESC 26), and F. incarnatum (FIESC 38). This is similar to those found associated with rice sheath rot in Indonesia [19], India [17] and the USA [18]. Larger part of the isolates clustered with F. sulawesiense which supports the findings in Brazil and China [29,56], while the abundance of F. hainanense and few others such as F. pernambucatum, F. tanahbumbuense, were among the FIESC reported on Brazilian rice. There is a wide variation among the species obtained within the FIESC complex and the observed variation could be correlated with variation in agro-ecological zones. Notable is the fact that some of the F. sulawesiense isolates from Ibadan are closely related to strains NTB 1 (rice sheath rot, Indonesia), LC6936 (rice, China), F1 (Sweet potato, US), BT48, and PRT6 (oil palm, Indonesia), and ITEM7547 (Musa, Bahamas) all of which originated from climates characterised by high temperature and humidity. Isolate F1 originated from Louisiana, USA that is known for its humid subtropical climate with long, hot, and humid summer, similar to the climate of ITEM7547 from Bahamas [57,58]. Based on our findings we can hypothesize that environmental factors could be the driving forces to be considered in the distribution of the FIESC isolates. Notable differences in climate and farming practice could not be underestimated as the three F. incarnatum isolates from a higher altitude in Rwanda formed part of recently classified FIESC 38 isolates from a similar altitude and climate in Brazil [29]. Within the FIESC clades, none of the studied isolates was found among F. equiseti clade which disagrees with the previous studies of [54,55,59] whereby variable percentage of both F. equiseti and F. incarnatum were reported from rice samples. Members of F. equiseti clade are frequent in cereals grown in Western Europe, Turkey, and North America [34].
FFSC species have been implicated as another causal agent of various rice diseases including sheath rot [51,60]. F. andiyazi and F. madaense represent the principal species recovered in Rwanda; F. nygamai was the dominant species isolated from Mali, while F. marum was recovered from Nigeria. An isolate each of F. casha (Rwanda) and F. annulatum (Mali) were also found among our FFSC. The peculiarity in the clustering of F. andiyazi, F. madaense and F. marum, clades observed in our studies is comparable to the typical phylogenetic pattern observed in [61]. F. andiyazi and F. madaense are typically associated with tropical grasses including sorghum, maize, millet, and rice in various parts of the world [61] The two virulent F. marum isolates from Nigeria clearly resolved into a separate clade and clustered with two F. marum isolates obtained from sorghum in Cameroon [61]. Isolates clustering with F. nygamai, the most dominant species in Mali, are closely related to isolates obtained from cereals from regions with similar warm and dry climate (Fig 4), such as Australia [62], Italy [28], Mexico [63], and Tunisia [36]. These FIESC and FFSC findings are consistent with the recent report of rice sheath rot disease in Indonesia, [19], and rice disease in China [56,64].
Furthermore, isolates clustering with F. callistephi were found in Rwanda, while F. triseptatum was obtained in Mali. Both are members of the FOSC. F. callistephi is mainly known as a wilt pathogen on ornamentals from the Asteraceae family [65]. Lastly, members of FSAMSC which includes two isolates clustering with F. acasiae-mearnsii of Rwanda origin were also identified (Fig. 4). F. acasiae-mearnsii isolates have previously be found in Australia and South-Africa and can cause head blight on wheat [66].
Reports on mycotoxins produced by Fusarium species isolated from rice sheath rot disease in sub-Saharan Africa are very limited. In consequence, this is the first study to investigate the toxigenic potentials of Fusarium isolates from rice sheath rot disease in Mali, Nigeria, and Rwanda. The predominant mycotoxins found in FIESC isolates from Nigeria were trichothecenes, while the three F. incarnatum isolates from Rwanda did not produce mycotoxins. Among the 12 FIESC isolates collected from Ibadan (derived savannah region) in Nigeria, type A (DAS, NEO) and B (NIV, FUS-X) trichothecenes were detected in 7 samples. This confirms the previous mycotoxins findings on cereals [29,59,67]. In addition, a huge ZEN production of 32,529 and 26,173 µg/kg was detected from the two F. hainanense isolates collected from Ibadan. Similar results for F. hainanense were obtained in Brazilian rice [29]. This also corroborates the study of [68] which demonstrated that ZEN production by Fusarium species is greater in moldy samples which is favoured by wet climates with high rainfall and high humidity. Within the FFSC, F. andiyazi isolates did not produce mycotoxins, while the F. annulatum isolate and 3 out of the 5 F. nygamai isolates from the dry and hot Sudan Guinea Savannah of Mali produced trichothecenes and fumonisins (FB1, FB2, and FB3), respectively. This is consistent with the findings of [69] reporting high levels of toxins for F. nygamai while F. andiyazi isolates produced little or no mycotoxins. Moreover, fumonisin-producers were not detected among the isolates collected from Nigeria and Rwanda. Thus, fumonisin contamination may be expected to be higher in samples collected in the Sahel with a warm and dry climate. The development of fumonisins in cereal crops prior to harvest might increase due to heat and water stress that characterized the environmental drought [70]. Only two of the 23 Fusarium isolates from Rwanda produced mycotoxins: trichothecenes (NIV and Fus-X) and ZEN were detected in a F. madaense isolate and a F. acaciae-mearnsii isolate, both obtained from the Rwamagana district. It has been shown before that F. acacia-mearnsii isolates can produce NIV [71] and ZEN [72].
According to the research of [73], which elucidated the presence and absence of biosynthetic gene clusters responsible for the synthesis of mycotoxins and secondary metabolites in FIESC, further studies are necessary to investigate if the mycotoxin production potentials of our isolates agree with their genetic profile or assess if there are differences in expression level. In conclusion, mycotoxin production is common in rice-derived Fusarium isolates from Nigeria (12 out of 15) and Mali (5 out of 7), but rare in Rwanda (2 out of 25).
In contrast to the heterogeneity observed among Fusarium species, three clearly delineated Sarocladium species were recovered from the three countries of study, however with lower frequency of occurrence in Mali and Rwanda. Following the characterization of Sarocladium species causing rice sheath rot in Taiwan by [12] and using a concatenation of two genes, we were able to resolve our isolates into three species namely attenuatum, oryzae and sparsum (Fig. 4). Isolates belonging to S. sparsum were only found in Nigeria and mostly originated from Ibadan (DS) except for an isolate (BDNG0025) from an infected seed in Katcha (SGS). Finding larger part of this group in Ibadan was not strange because it clusters with an Ibadan origin reference isolate CBS 414.81, collected and reported as S. attenuatum [21], and later deposited into the Genbank [42]. Our results clearly show, however, that isolate CBS 414.81 belongs to sparsum species. Surprisingly, this group was not found in Mali and Rwanda. There occurs a notable correlation between this group and the collection region, which proved that geographical area and climate are the most crucial factors that influence occurrence of these pathotypes and their virulence. This agrees with the hypothesis that isolates from different locations may also vary in their level of aggressiveness [74] [75]. S. oryzae isolates (Fig. 4) showed a strong intra-species similarity that is not phylogeographic based. Isolates in this species were the most predominant and widely distributed. It consists of 18 similar isolates from nearly all the rice-growing regions in the world. Most isolates from Nigeria (11 of 16) belongs to this group. An isolate from Mali (SEMA0013A) generated from this study, isolates from previous studies on rice such as 13017 from Taiwan [12] CBS180.74 from India, African isolates CBS361.75 from Kenya [76] RFRG2 from Rwanda (this study), Central American CBS120.817 from Panama and CBS485.80 Australia [77] are part of this group. They were found in two agro-ecological zones of Nigeria, although more frequently from Katcha than Ibadan.
The presence of S. oryzae in all the rice growing regions of the world is a signal of its flexibility to adjust to various agro-ecological zones. It also suggests a link between its dispersal, rice movement and international trade. A potential quarantine threat is of a great concern with the rapid distribution of this group. This might also imply that the origin of the isolate may be connected to Asia. It should be noted that African countries including Mali, Nigeria and Rwanda are major importers of rice from Asia despite Nigeria being the highest producer in the continent.
A third distinct group, S. attenuatum, was dominated by Rwandan isolates, the second Mali isolate, SEMA0029 also formed part of the group.
It is important to note that the two Sarocladium isolates obtained from Mali formed two of the three species found in this study despite the small sample size. Several reasons might be responsible, from accession variability, as local rice is mostly cultivated in Mali, [78], to toxigenic variability among the competing Fusarium species, and multiple cropping system variation, among others. Larger scale surveys across the country are necessary to have a wider knowledge of genetic diversity and distribution.
It is a well-known phenomenon that several factors such as variation in climate, topography, farming practices among others are drivers of variation in pathogen populations. To verify this claim, this study has revealed a strong ecology driven diversification among the Sarocladium species used. It also exposed how environmental variation was able to influence the genetic and virulence relatedness. The S. sparsum isolates that are mainly found in Ibadan (DS) are clearly more aggressive on the rice FARO44 cultivar used in this study than the S. oryzae and S. attenuatum strains, which are more common in the savannah region in Katcha-Badeggi and in Rwanda. This confirms previous work showing that group 3 strains (= S. sparsum) are more aggressive on the rice japonica cultivar Kitaake than group 1 (= S. oryzae) or group 2 (= S. attenuatum) strains. S. sparsum isolates also produce high amounts of the toxin helvolic acid in planta which is clearly correlated with disease severity [79].
The results showed that isolates of Sarocladium species were able to induce sheath rot symptoms and they are the most aggressive pathogens on FARO44 cultivar used in this study with higher pathogenicity index. We also observed a noticeable correlation between isolate’s country of origin and virulence, which tends to agree with earlier findings where isolates were found to be widely diverse in virulence within countries and within the same population. The aggressiveness of Sarocladium species collected from the two locations in Nigeria, and Selingue in Mali seems to be influenced by their location. The same trend of aggressiveness was reported in the study of [21] who used four isolates (presumably S. sparsum) collected from the southern region (DS and Humid Forest) of Nigeria. The similarity between isolates from Katcha (SGS) in Nigeria and the two Mali (NGS) isolates that were classified as moderate and low virulence may not be unconnected with the similarity in their ecological origin.
5. Conclusion
In conclusion, our study showed that diversity occurs within the East (Rwanda) and West African (Mali and Nigeria) isolates of Fusarium and Sarocladium. Notable correlation was observed between the virulent group and collection area. The toxigenic profile of both pathogens was elucidated, and we found that most Fusarium isolates from Nigeria and Mali were able to produce one or more mycotoxins. In contrast, only two out of 24 isolates from Rwanda were able to produce mycotoxins. Further investigations with a broader geographic scope and a larger collection of samples are necessary to examine pathogenic variability and the population genetic structure. Meanwhile, based on the distinct groups of isolates from different regions, breeders in various agroecological regions should take note of the variations in virulence. This information can serve as a basis for selecting strains useful for identification and selecting effective sources of resistance for local rice breeding programs.
Moreover, additional studies are required to determine whether Sarocladium and Fusarium individually contribute to the observed symptoms in the field or if there is an interplay between both pathogens in the rice sheath rot complex. To the best of our knowledge, this research provides the first comprehensive dataset on the distribution, genetics, pathogenicity, and toxigenic profile of Fusarium species associated with rice sheath rot disease in sub-Saharan Africa.
Author Contributions
Conceptualization,V.B.; O.A.; O.I.N; and M.H.; methodology, O.A.; V.B.; M.D.V, F.O. and G.K.H.H.; validation, O.A.; V.B.; formal analysis, O.A. investigation, O.A.; V.B.; L.B, and G.K.H.H.; Resources, O.A.; V.B.; J.O.; O.E.O.; A.O.; curation, O.A; writing—original draft preparation, O.A. writing, review, and editing, M.H.; visualization, O.A.; V.B.; supervision, M.H, and S.D.S project administration, O.A.; V.B.; M.H.; funding acquisition, O.A.; V.B.; S.D.S.; and M.H.; All authors have read and agreed to the published version of the manuscript.
Funding
V.P. Bigirimana received a doctoral grant from the Belgian Technical Cooperation (BTC) (project reference: 10RWA/0018). This work was funded by a grant from the Special Research Fund of Ghent University (GOA 01GB3013) and by the Fund for Scientific Research Flanders (FWO G031317N).
Acknowledgments
The authors wish to appreciate Christ’l Detavernier, Frédéric Dumoulin (UGent, Faculty of Pharmaceutical Sciences) for their help with the LC-MS/MS analysis. We also acknowledged the contributions of Aderonke Oludare and Opeyemi Ogedengbe (AfricaRice) for their coordination of rice sampling and transportation to Belgium.
Conflicts of Interest
The authors declare no conflict of interest.

Table A1.
Reference strains of Fusarium obtained from GenBank used for building phylogenetic tree.
| Species complex | Species | Isolatename | Host | Origin | Accession number | References |
| FIESC | F. equiseti | NL19-045005 | Soil | Netherlands | MZ921835 | [50] |
| CPC 35262 | Human toenail | Czech republic | QED42271 | [49] | ||
| F. hainanense (26) | LC11638 | Oryza sp | China | MK289581 | [80] | |
| 15Ar057 | Rice | Brazil | MK298120 | [29] | ||
| LB2 | Oryza sativa | Philippines | JF715935 | [31] | ||
| PCO2 | Oil palm | Indonesia | HM770725 | [58] | ||
| NRRL 28714 | Clinical samples | USA | GQ505604 | [54] | ||
| JBR 10 | Oryza sativa sheath | Indonesia | MT138474 | [19] | ||
| F. nanum (25) | F685 | Wheat | Spain | KF962950 | [81] | |
| NRRL 22244 | Clinical samples | USA | GQ505596 | [54] | ||
| F. tanahbumbuense (24) | LC13726 | Digitaria sp | China | MW594396 | [80] | |
| NTT 6 | Oryza sativa sheath | Indonesia | MT138460 | [19] | ||
| F. incarnatum (38) | ITEM 7155 | Trichosanthe dioica | Malawi | LN901581 | [34] | |
| F. pernambucanum (17) | NRRL 32864 | Clinical samples | USA | GQ505613 | [54] | |
| CBS 791.70 | Musa sampientum | Netherlands* | MN170491 | [49] | ||
| F. sulawesiense (16) | CBS 622.87 | Bixa orellana | Brazil* | MN170503 | [49] | |
| ITEM7547 | Musa sampientum | Bahamas | LN901580 | [34] | ||
| LC12173 | Luffa aegyptica | China | MK289605 | [80] | ||
| MS3369 | Wild rice | Brazil | MT682685 | [82] | ||
| LC6936 | Oryza sativa | China | MK289621 | [80] | ||
| F1 | Sweet potato | USA | KC820972 | [83] | ||
| BT48 | Oil palm | Indonesia | HM770722 | [58] | ||
| PRT6 | Oil palm | Indonesia | HM770723 | [58] | ||
| NTB 1 | Indonesia | Oryza sativa sheath | MT138458 | [19] | ||
| FFSC | F. andiyazi | LLC 1152 | Striga hermonthica seed | Ethiopia | OP486864 | [84] |
| MO5-1946S-3_PCNB | Sorghum grain | USA | KM462919 | [85] | ||
| F.marum | KSU 15077 | Sorghum | Cameroun | MT374735 | [61] | |
| KSU15074 | Sorghum | Cameroun | MT374736 | [61] | ||
| E432 | Rice seeds | Italy | GU827420 | [86] | ||
| E439 | Rice seeds | Italy | GU827419 | [86] | ||
| 30ALH | Oryza sativa seed | China | FN252387 | [15] | ||
| F. madaense | CBS 146669 | Arachis hypogaea | Nigeria | MW402098 | [40] | |
| 44ALH | Oryza sativa seed | Tanzania | FN252390 | [15] | ||
| IALH | Oryza sativa seed | Burkina Faso | FN252388 | [15] | ||
| CML3853 | Sorghumbicolor | Nigeria | MK895723 | [61] | ||
| CML3895 | Sorghumbicolor | Tanzania | MK895727 | [61] | ||
| PB-2 | Sugarcane | China | KP314282 | [87] | ||
| 38ALH | Oryza sativa seed | India | FN252389 | [15] | ||
| F. casha | PPRI20462 | Amaranthus cruentus | South Africa | MF787262 | [88] | |
| F. nygamai | B5Hp1g3B1 | Barley | Tunisia | MG452941 | [36] | |
| KC 13 | Tomato | Kenya | KT357537 | [89] | ||
| ENTO90 | Wild rice | Australia | MG873156 | [62] | ||
| M7491 | Rice | Italy | HM243236 | [28] | ||
| F. annulatum | LC11670 | Oryza sativa | China | MW580517 | [64] | |
| 34ALH | Oryza sativa seed | China | FN252396 | [15] | ||
| LC13675 | Syzygium samarangense | China | MW580542 | [64] | ||
| F. proliferatum | CBS 480.96 | Soil | Papua New Guinea | MN534059 | [90] | |
| FOSC | F. triseptatum | LC13771 | Deep sea sediment | China | MW594358 | [64] |
| F. oxysporum | Foc230 | Banana | Nigeria | AY217161 | Upublished | |
| F. callistephi | CBS 187.53 | Callistephus chinensis | Netherlands | MH484966 | [84] | |
| SRRC1630 | Cooked rice | Nigeria | KT950251 | [91] | ||
| CBS 115423 | Agathosma betulina | South Africa | MH484996 | [84] | ||
| FSAMSC | F. acaciae-mearnsii | LC13786 | Musa nana | China | MW620091 | [64] |
Table A2.
Sequences of reference strains of Sarocladium oryzae on rice obtained from GenBank used for building phylogenetic trees.
Table A2.
Sequences of reference strains of Sarocladium oryzae on rice obtained from GenBank used for building phylogenetic trees.
| Genus | Species | Isolate | Origin | Accession number ITS | Reference ITS | Accession number ACT | Reference ACT |
| Sarocladium | Attenuatum | CBS 399.73 | India | HG965027 | [42] | HG964979 | [42] |
| 2-5 | Taiwan | LC461444 | [12] | LC464336 | [12] | ||
| oryzae | CBS 180.74 | India | HG965026 | [42] | HG964978 | [42] | |
| 13017 | Taiwan | LC461506 | [12] | LC464380 | [12] | ||
| sparsum | CBS 414.81 | Nigeria | HG965028 | [42] | HG964980 | [42] | |
| 18042 | Taiwan | LC461520 | [12] | LC464308 | [12] | ||
| Sarocladium | zeae | CBS 800.69 | USA | FN691451 | [92] | HG965000 | [42] |
References
- FAOSTAT. Available online: https://www.fao.org/faostat/en/#home (accessed on 10 August 2022).
- KPMG Nigeria Rice Industry Review. 2019.
- Afolabi, O.; Milan, B.; Amoussa, R.; Koebnik, R.; Poulin, L.; Szurek, B.; Habarugira, G.; Bigirimana, J.; Silue, D.; Ongom, J.; et al. First Report of Xanthomonas oryzae pv oryzicola Causing Bacterial Leaf Streak of Rice in Uganda. Plant Dis 2014, 98, 1579–1579. [Google Scholar] [CrossRef] [PubMed]
- Afolabi, O.; Amoussa, R.; Bilé, M.; Oludare, A.; Gbogbo, V.; Poulin, L.; Koebnik, R.; Szurek, B.; Silué, D. First Report of Bacterial Leaf Blight of Rice Caused by Xanthomonas oryzae pv. oryzae in Benin. Plant Dis 2016, 100, 515–515. [Google Scholar] [CrossRef]
- Oludare, A.; Sow, M.; Afolabi, O.; Pinel-Galzi, A.; Hébrard, E.; Silué, D. First Report of Rice stripe necrosis virus Infecting Rice in Benin. Plant Dis 2015, 99, 735–735. [Google Scholar] [CrossRef]
- Séré, Y.; Onasanya, A.; Afolabi, A.; Abo, E. Evaluation and Potential of Double Immunodifusion Gel Essay for Serological Characterization of Rice yellow mottle virus Isolates in West Africa. Afr J Biotechnol 2005, 4, 197–205. [Google Scholar]
- Traoré, O.; Pinel-Galzi, A.; Sorho, F.; Sarra, S.; Rakotomalala, M.; Sangu, E.; Kanyeka, Z.; Séré, Y.; Konaté, G.; Fargette, D. A Reassessment of the Epidemiology of Rice yellow mottle virus Following Recent Advances in Field and Molecular Studies. Virus Res 2009, 141, 258–267. [Google Scholar] [CrossRef]
- Bigirimana, V. de P.; Hua, G.K.H.; Nyamangyoku, O.I.; Höfte, M. Rice Sheath Rot: An Emerging Ubiquitous Destructive Disease Complex. Front Plant Sci 2015, 6. [Google Scholar] [CrossRef]
- Gams, W.; Hawksworth, D. L. The Identity of Acrocylindrium oryzae Sawada and a Similar Fungus Causing Sheath-Rot of Rice. Kavaka 1976, 57–61. [Google Scholar]
- Mew, T.W.; Gonzales, P. A Handbook of Rice Seedborne Fungi; IRRI, 2002; ISBN 1578082552.
- Bridge, P.D.; Hawksworth, D.L.; Kavishe, D.F.; Farnell, P.A. A Revision of the Species Concept in Sarocladium, the Causal Agent of Sheath-rot in Rice and Bamboo Blight, Based on Biochemical and Morphometric Analyses. Plant Pathol 1989, 38, 239–245. [Google Scholar] [CrossRef]
- Ou, J.H.; Lin, G.C.; Chen, C.Y. Sarocladium Species Associated with Rice in Taiwan. Mycol Prog 2020, 19, 67–80. [Google Scholar] [CrossRef]
- CABI Sarocladium oryzae (Rice Sheath Rot). Available online: https://www.cabi.org/isc/datasheet/48393 (accessed on 10 August 2022).
- Shier, W.T.; Bird, C.B.; Rice, L.G.; Abouzied, M.M.; Meredith, F.I.; Cartwright, R.D.; Sciumbato, G.L.; Abbas, H.K.; Ross, P.F. Natural Occurrence of Fumonisins in Rice with Fusarium Sheath Rot Disease. Plant Dis 2007, 82, 22–25. [Google Scholar] [CrossRef]
- Wulff, E.G.; Sørensen, J.L.; Lübeck, M.; Nielsen, K.F.; Thrane, U.; Torp, J. Fusarium spp. Associated with Rice Bakanae: Ecology, Genetic Diversity, Pathogenicity and Toxigenicity. Environ Microbiol 2010, 12, 649–657. [Google Scholar] [CrossRef]
- Abbas, H.K.; Cartwright, R.D.; Shier, W.T.; Abouzied, M.M.; Bird, C.B.; Rice, L.G.; Ross, P.F.; Sciumbato, G.L.; Meredith, F.I. Natural Occurrence of Fumonisins in Rice with Fusarium Sheath Rot Disease. Plant Dis 1998, 82, 22–25. [Google Scholar] [CrossRef] [PubMed]
- Prabhukarthikeyan, S.R.; Keerthana, U.; Nagendran, K.; Yadav, M.K.; Parameswaran, C.; Panneerselvam, P.; Rath, P.C. First Report of Fusarium proliferatum Causing Sheath Rot Disease of Rice in Eastern India. Plant Dis 2021, 105, 704. [Google Scholar] [CrossRef]
- Imran, M.; Khanal, S.; Zhou, X. Disease Note. 2022, 106, 3206. [Google Scholar]
- Pramunadipta, S.; Widiastuti, A.; Wibowo, A.; Suga, H.; Priyatmojo, A. Identification and Pathogenicity of Fusarium Spp. Associated with the Sheath Rot Disease of Rice (Oryza sativa) in Indonesia. J Plant Pathol 2022, 104, 251–267. [Google Scholar] [CrossRef]
- Musonerimana, S.; Bez, C.; Licastro, D.; Habarugira, G.; Bigirimana, J.; Venturi, V. Pathobiomes Revealed That Pseudomonas Fuscovaginae and Sarocladium oryzae Are Independently Associated with Rice Sheath Rot. Microb Ecol 2020, 80, 627–642. [Google Scholar] [CrossRef] [PubMed]
- Ngala, G. Sarocladium attenuatum as One of the Causes of Rice Grain Spotting in Nigeria. Plant Pathol 1983, 32. [Google Scholar] [CrossRef]
- Ekan, E. J, Ibrnenge, Vange, T. Effect of Seed Treatment with Tolchlofos-Methyl on Percentage Incidence of Seed-Borne Fungi and Gernlination of Rice (Oryza sativa) in Makurdi, Nigeria. Indian J Agric Sci 2006, 76, 450–453. [Google Scholar]
- Makun, H.A.; Dutton, M.F.; Njobeh, P.B.; Phoku, J.Z.; Yah, C.S. Incidence, Phylogeny and Mycotoxigenic Potentials of Fungi Isolated from Rice in Niger State, Nigeria. J Food Saf 2011, 31, 334–349. [Google Scholar] [CrossRef]
- Ghosh, M.K.; Amudha, R.; Jayachandran, S.; Sakthivel, N. Detection and Quantification of Phytotoxic Metabolites of Sarocladium oryzae in Sheath Rot-Infected Grains of Rice. Lett Appl Microbiol 2002, 34, 398–401. [Google Scholar] [CrossRef]
- Naeimi, S.; Okhovvat, S.M.; Hedjaroude, G.A.; Khosravi, V. Sheath Rot of Rice in Iran. Abstract. Commun Agric Appl Biol Sci 2003, 68, 681–684. [Google Scholar] [PubMed]
- Mvuyekure, S.M.; Sibiya, J.; Derera, J.; Nzungize, J.; Nkima, G. Genetic Analysis of Mechanisms Associated with Inheritance of Resistance to Sheath Rot of Rice. Plant Breeding 2017, 136, 509–515. [Google Scholar] [CrossRef]
- Bigirimana, V.P. 2016. Characterization of sheath rot pathogens from major rice-growing areas in Rwanda. PhD Thesis, Ghent University.
- Amatulli, M.T.; Spadaro, D.; Gullino, M.L.; Garibaldi, A. Conventional and Real-Time PCR for the Identification of Fusarium fujikuroi and Fusarium proliferatum from Diseased Rice Tissues and Seeds. Eur J Plant Pathol 2012, 134, 401–408. [Google Scholar] [CrossRef]
- Avila, C.F.; Moreira, G.M.; Nicolli, C.P.; Gomes, L.B.; Abreu, L.M.; Pfenning, L.H.; Haidukowski, M.; Moretti, A.; Logrieco, A.; Del Ponte, E.M. Fusarium incarnatum-equiseti species complex Associated with Brazilian Rice: Phylogeny, Morphology and Toxigenic Potential. Int J Food Microbiol 2019, 306, 108267. [Google Scholar] [CrossRef]
- Choi, J.H.; Lee, S.; Nah, J.Y.; Kim, H.K.; Paek, J.S.; Lee, S.; Ham, H.; Hong, S.K.; Yun, S.H.; Lee, T. Species Composition of and Fumonisin Production by the Fusarium fujikuroi species complex Isolated from Korean Cereals. Int J Food Microbiol 2018, 267, 62–69. [Google Scholar] [CrossRef]
- Cruz, A.; Marín, P.; González-Jaén, M.T.; Aguilar, K.G.I.; Cumagun, C.J.R. Phylogenetic Analysis, Fumonisin Production and Pathogenicity of Fusarium fujikuroi Strains Isolated from Rice in the Philippines. J Sci Food Agric 2013, 93, 3032–3039. [Google Scholar] [CrossRef]
- Desjardins, A.E.; Manandhar, H.K.; Plattner, R.D.; Manandhar, G.G.; Poling, S.M.; Maragos, C.M. Fusarium Species from Nepalese Rice and Production of Mycotoxins and Gibberellic Acid by Selected Species. Appl Environ Microbiol 2000, 66, 1020–1025. [Google Scholar] [CrossRef]
- Leyva-Madrigal, K.Y.; Larralde-Corona, C.P.; Apodaca-Sánchez, M.A.; Quiroz-Figueroa, F.R.; Mexia-Bolaños, P.A.; Portillo-Valenzuela, S.; Ordaz-Ochoa, J.; Maldonado-Mendoza, I.E. Fusarium species from the Fusarium fujikuroi Species Complex Involved in Mixed Infections of Maize in Northern Sinaloa, Mexico. J Phytopathol 2015, 163, 486–497. [Google Scholar] [CrossRef]
- Villani, A.; Moretti, A.; Saeger, S.; Han, Z.; Mavungu, J.D.; Soares, C.M.G.; Proctor, R.H.; Venâncio, A.; Lima, N.; Stea, G.; et al. A Polyphasic Approach for Characterization of a Collection of Cereal Isolates of the Fusarium incarnatum-equiseti species complex. Int J Food Microbiol 2016, 234. [Google Scholar] [CrossRef]
- Amadi, J.; Adeniyi, D. Mycotoxin Production by Fungi Isolated from Stored Grains. Afr J Biotechnol 2009, 8, 1219–1221. [Google Scholar]
- Jedidi, I.; Soldevilla, C.; Lahouar, A.; Marín, P.; González-Jaén, M.T.; Said, S. Mycoflora Isolation and Molecular Characterization of Aspergillus and Fusarium species in Tunisian cereals. Saudi J Biol Sci 2018, 25, 868–874. [Google Scholar] [CrossRef] [PubMed]
- Madbouly, A.K.; Ibrahim, M.I.M.; Sehab, A.F.; Abdel-Wahhab, M.A. Co-Occurrence of Mycoflora, Aflatoxins and Fumonisins in Maize and Rice Seeds from Markets of Different Districts in Cairo, Egypt. Food Addit Contam: Part B 2012, 5, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Makun, H.; Gbodi, T.; Akanya, O. Fungi and Some Mycotoxins Contaminating Rice (Oryza sativa) in Niger State, Nigeria. Afr J Biotechnol 2007, 6, 99–108. [Google Scholar]
- Rofiat, A.-S.; Fanelli, F.; Atanda, O.; Sulyok, M.; Cozzi, G.; Bavaro, S.; Krska, R.; Logrieco, A.F.; Ezekiel, C.N. Fungal and Bacterial Metabolites Associated with Natural Contamination of Locally Processed Rice (Oryza sativa L.) in Nigeria. Food Addit Contam Part A Chem Anal Control Expo Risk Assess 2015, 32, 950–959. [Google Scholar] [CrossRef]
- Ezekiel, C.N.; Kraak, B.; Sandoval-Denis, M.; Sulyok, M.; Oyedele, O.A.; Ayeni, K.I.; Makinde, O.M.; Akinyemi, O.M.; Krska, R.; Crous, P.W.; et al. Diversity and Toxigenicity of Fungi and Description of Fusarium madaense Sp. Nov. From Cereals, Legumes and Soils in North-Central Nigeria. MycoKeys 2020, 67, 95–124. [Google Scholar] [CrossRef]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple Evolutionary Origins of the Fungus Causing Panama Disease of Banana: Concordant Evidence from Nuclear and Mitochondrial Gene Genealogies. Proc Nat Acad Sci USA 1998, 95. [Google Scholar] [CrossRef] [PubMed]
- Giraldo, A.; Gené, J.; Sutton, D.A.; Madrid, H.; de Hoog, G.S.; Cano, J.; Decock, C.; Crous, P.W.; Guarro, J. Phylogeny of Sarocladium (Hypocreales). Persoonia 2015, 34, 10–24. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A guide to methods and applications; Innes, M.A., Gefland, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press Inc.: San Diego, US-CA, 1990; pp. 315–322. [Google Scholar]
- Hall, T. Bioedit Version 7 2 5 | Bioedit Company | Bioz Available online:. Available online: https://www.bioz.com/result/bioedit%20version%207%202%205/product/Bioedit%20Company (accessed on 10 August 2022).
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol Biol Evol 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (ITOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Sakthivel, N.; Gnanamanickam, S.S. Evaluation of Pseudomonas fluorescens for Suppression of Sheath Rot Disease and for Enhancement of Grain Yields in Rice (Oryza sativa L.). Appl Envir Microbiol 1987, 53, 2056–2059. [Google Scholar] [CrossRef]
- Sofie, M.; Van Poucke, C.; Detavernier, C.T.L.; Dumoultn, F.; Van Velde, M.D.E.; Schoeters, E.; Van Dyck, S.; Averkieva, O.; Van Peteghem, C.; De Saeger, S. Occurrence of Mycotoxins in Feed as Analyzed by a Multi-Mycotoxin LC-MS/MS Method. J Agric Food Chem 2010, 58, 66–71. [Google Scholar] [CrossRef]
- Xia, J.W.; Sandoval-Denis, M.; Crous, P.W.; Zhang, X.G.; Lombard, L. Numbers to Names – Restyling the Fusarium incarnatum-equiseti species complex. Persoonia 2019, 43, 186–221. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Lombard, L.; Sandoval-Denis, M.; Seifert, K.A.; Schroers, H.J.; Chaverri, P.; Gené, J.; Guarro, J.; Hirooka, Y.; Bensch, K.; et al. Fusarium: More than a Node or a Foot-Shaped Basal Cell. Stud Mycol 2021, 98. [Google Scholar] [CrossRef]
- Han, S.L.; Wang, M.M.; Ma, Z.Y.; Raza, M.; Zhao, P.; Liang, J.M.; Gao, M.; Li, Y.J.; Wang, J.W.; Hu, D.M.; et al. Fusarium Diversity Associated with Diseased Cereals in China, with an Updated Phylogenomic Assessment of the Genus. Stud Mycol 2023. [Google Scholar] [CrossRef]
- Wang, J.-H.; Zhang, J.-B.; Li, H.-P.; Gong, A.-D.; Xue, S.; Agboola, R.S.; Liao, Y.-C. Molecular Identification, Mycotoxin Production and Comparative Pathogenicity of Fusarium temperatum Isolated from Maize in China. Journal of Phytopathology 2014, 162, 147–157. [Google Scholar] [CrossRef]
- Castellá, G.; Cabañes, F.J. Phylogenetic Diversity of Fusarium incarnatum-equiseti species complex Isolated from Spanish Wheat. Anton Leeuw Int J Gen Mol Microbiol 2014, 106, 309–317. [Google Scholar] [CrossRef]
- O’Donnell, K.; Gueidan, C.; Sink, S.; Johnston, P.R.; Crous, P.W.; Glenn, A.; Riley, R.; Zitomer, N.C.; Colyer, P.; Waalwijk, C.; et al. A Two-Locus DNA Sequence Database for Typing Plant and Human Pathogens within the Fusarium oxysporum Species Complex. Fungal Genet Biol 2009, 46, 936–948. [Google Scholar] [CrossRef]
- O’Donnell, K.; Humber, R.A.; Geiser, D.M.; Kang, S.; Park, B.; Robert, V.A.R.G.; Crous, P.W.; Johnston, P.R.; Aoki, T.; Rooney, A.P.; et al. Phylogenetic Diversity of Insecticolous Fusaria Inferred from Multilocus DNA Sequence Data and Their Molecular Identification via FUSARIUM-ID and Fusarium MLST. Mycologia 2012, 104, 427–445. [Google Scholar] [CrossRef]
- Lu, Y.; Qiu, J.; Wang, S.; Xu, J.; Ma, G.; Shi, J.; Bao, Z. Species Diversity and Toxigenic Potential of Fusarium incarnatum-equiseti species complex isolates from Rice and Soybean in China. Plant Dis 2021, 105. [Google Scholar] [CrossRef]
- Silva, W.L.; Clark, C.A. Infection of Sweetpotato by Fusarium solani and Macrophomina phaseolina Prior to Harvest. Plant Dis 2013, 97, 1636–1644. [Google Scholar] [CrossRef]
- Suwandi; Akino, S. ; Kondo, N. Common Spear Rot of Oil Palm in Indonesia. Plant Dis 2012, 96, 537–543. [Google Scholar] [CrossRef]
- Kosiak, E.B.; Holst-Jensen, A.; Rundberget, T.; Gonzalez Jaen, M.T.; Torp, M. Morphological, Chemical and Molecular Differentiation of Fusarium equiseti Isolated from Norwegian Cereals. Int J Food Microbiol 2005, 99, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Nicolli, C.P.; Haidukowski, M.; Susca, A.; Gomes, L.B.; Logrieco, A.; Stea, G.; Del Ponte, E.M.; Moretti, A.; Pfenning, L.H. Fusarium fujikuroi Species Complex in Brazilian Rice: Unveiling Increased Phylogenetic Diversity and Toxigenic Potential. Int J Food Microbiol 2020, 330. [Google Scholar] [CrossRef]
- Costa, M.M.; Saleh, A.A.; Melo, M.P.; Guimarães, E.A.; Esele, J.P.; Zeller, K.A.; Summerell, B.A.; Pfenning, L.H.; Leslie, J.F. Fusarium mirum Sp. Nov, Intertwining Fusarium madaense and Fusarium andiyazi, Pathogens of Tropical Grasses. Fungal Biol 2022, 126, 250–266. [Google Scholar] [CrossRef] [PubMed]
- Petrovic, T.; Burgess, L.W.; Cowie, I.; Warren, R.A.; Harvey, P.R. Diversity and Fertility of Fusarium Sacchari from Wild Rice (Oryza Australiensis) in Northern Australia, and Pathogenicity Tests with Wild Rice, Rice, Sorghum and Maize. Eur J Plant Pathol 2013, 136, 773–788. [Google Scholar] [CrossRef]
- Velarde-Félix, S.; Garzón-Tiznado, J.A.; Hernández-Verdugo, S.; López-Orona, C.A.; Retes-Manjarrez, J.E. Occurrence of Fusarium oxysporum Causing Wilt on Pepper in Mexico. Can J Plant Pathol 2018, 40, 238–247. [Google Scholar] [CrossRef]
- Wang, M.M.; Crous, P.W.; Sandoval-Denis, M.; Han, S.L.; Liu, F.; Liang, J.M.; Duan, W.J.; Cai, L. Fusarium and Allied Genera from China: Species Diversity and Distribution. Persoonia 2022, 48, 1–53. [Google Scholar] [CrossRef]
- Edel-Hermann, V.; Lecomte, C. Current Status of Fusarium oxysporum Formae Speciales and Races. Phytopathology 2019, 109, 512–530. [Google Scholar] [CrossRef]
- Starkey, D.E.; Ward, T.J.; Aoki, T.; Gale, L.R.; Kistler, H.C.; Geiser, D.M.; Suga, H.; Tóth, B.; Varga, J.; O’Donnell, K. Global Molecular Surveillance Reveals Novel Fusarium Head Blight Species and Trichothecene Toxin Diversity. Fungal Genet Biol 2007, 44, 1191–1204. [Google Scholar] [CrossRef]
- Hartman, G.L.; McCormick, S.P.; O’donnell, K. Trichothecene-Producing Fusarium species Isolated from Soybean Roots in Ethiopia and Ghana and Their Pathogenicity on Soybean. Plant Dis 2019, 103, 2070–2075. [Google Scholar] [CrossRef]
- Makun, H.A.; Hussaini, A.M.; Timothy, A.G.; Olufunmilayo, H.A.; Ezekiel, A.S.; Godwin, H.O. Fungi and Some Mycotoxins Found in Mouldy Sorghum in Niger State, Nigeria. World J Agric Sci 5, 5–17.
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Blackwell Publishing Ltd.: Oxford. 2006. [Google Scholar]
- Atehnkeng, J.; Ojiambo, P.S.; Donner, M.; Ikotun, T.; Sikora, R.A.; Cotty, P.J.; Bandyopadhyay, R. Distribution and Toxigenicity of Aspergillus species Isolated from Maize Kernels from Three Agro-Ecological Zones in Nigeria. Int J Food Microbiol 2008, 122, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Goswami, R.S.; Kistler, H.C. Pathogenicity and in Planta Mycotoxin Accumulation among Members of the Fusarium graminearum Species Complex on Wheat and Rice. Phytopathology 2005, 95, 1397–1404. [Google Scholar] [CrossRef]
- Beukes, I.; Rose, L.J.; van Coller, G.J.; Viljoen, A. Disease Development and Mycotoxin Production by the Fusarium graminearum Species Complex Associated with South African Maize and Wheat. Eur J Plant Pathol 2018, 150, 893–910. [Google Scholar] [CrossRef]
- Villani, A.; Proctor, R.H.; Kim, H.S.; Brown, D.W.; Logrieco, A.F.; Amatulli, M.T.; Moretti, A.; Susca, A. Variation in Secondary Metabolite Production Potential in the Fusarium incarnatum-equiseti species Complex Revealed by Comparative Analysis of 13 Genomes. BMC Genomics 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Ayyadurai, N.; Kirubakaran, S.I.; Srisha, S.; Sakthivel, N. Biological and Molecular Variability of Sarocladium oryzae, the Sheath Rot Pathogen of Rice (Oryza sativa L.). Curr Microbiol 2005, 50, 319–323. [Google Scholar] [CrossRef]
- Tajul Islam Chowdhury, M.; Salim Mian, M.; Taher Mia, M.A.; Rafii, M.Y.; Latif, M.A. Agro-Ecological Variations of Sheath Rot Disease of Rice Caused by Sarocladium oryzae and DNA Fingerprinting of the Pathogen’s Population Structure. Genet Mol Res 2015, 14, 18140–18152. [Google Scholar] [CrossRef]
- Bills, G.F.; Platas, G.; Gams, W. Conspecificity of the Cerulenin and Helvolic Acid Producing “Cephalosporium caerulens”, and the Hypocrealean Fungus Sarocladium oryzae. Mycol Res 2004, 108, 1291–1300. [Google Scholar] [CrossRef]
- Lanoiselet, V.; You, M.P.; Li, Y.P.; Wang, C.P.; Shivas, R.G.; Barbetti, M.J. First Report of Sarocladium oryzae Causing Sheath Rot on Rice (Oryza sativa) in Western Australia. Plant Dis 2012, 96, 1382. [Google Scholar] [CrossRef]
- Demont, M.; Ndour, M. Upgrading Rice Value Chains: Experimental Evidence from 11 African Markets. Glob Food Sec 2015, 5, 70–76. [Google Scholar] [CrossRef]
- Peeters, K.J.; Haeck, A.; Harinck, L.; Afolabi, O.O.; Demeestere, K.; Audenaert, K.; Höfte, M. Morphological, Pathogenic and Toxigenic Variability in the Rice Sheath Rot Pathogen Sarocladium oryzae. Toxins 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.M.; Chen, Q.; Diao, Y.Z.; Duan, W.J.; Cai, L. Fusarium incarnatum-equiseti Complex from China. Persoonia 2019, 43, 70–89. [Google Scholar] [CrossRef]
- Castella, G.; Cabañes, F.J. Phylogenetic Diversity of Fusarium incarnatum-equiseti species Complex Isolated from Spanish Wheat. Antonie Van Leeuwenhoek 2014, 106. [Google Scholar] [CrossRef] [PubMed]
- Tralamazza, S.M.; Piacentini, K.C.; Savi, G.D.; Carnielli-Queiroz, L.; de Carvalho Fontes, L.; Martins, C.S.; Corrêa, B.; Rocha, L.O. Wild Rice (O. Latifolia) from Natural Ecosystems in the Pantanal Region of Brazil: Host to Fusarium incarnatum-equiseti species Complex and Highly Contaminated by Zearalenone. Int J Food Microbiol 2021, 345. [Google Scholar] [CrossRef] [PubMed]
- da Silva, W.L.; Clark, C.A. Infection of Sweetpotato by Fusarium solani and Macrophomina phaseolina Prior to Harvest. Plant Dis 2013, 97, 1536–1664. [Google Scholar] [CrossRef]
- Lombard, L.; Sandoval-Denis, M.; Lamprecht, S.C.; Crous, P.W. Epitypification of Fusarium oxysporum – Clearing the Taxonomic Chaos. Persoonia 2019, 43, 1–47. [Google Scholar] [CrossRef]
- Funnell-Harris, D.L.; Sattler, S.E.; O’Neill, P.M.; Eskridge, K.M.; Pedersen, J.F. Effect of Waxy (Low Amylose) on Fungal Infection of Sorghum Grain. Phytopathology 2015, 105, 786–796. [Google Scholar] [CrossRef]
- Prà, M.D.; Tonti, S.; Pancaldi, D.; Nipoti, P.; Alberti, I. First Report of Fusarium andiyazi Associated with Rice Bakanae in Italy. Plant Dis 2010, 94, 1070–1070. [Google Scholar] [CrossRef]
- Yao, Z.; Zou, C.; Peng, N.; Zhu, Y.; Bao, Y.; Zhou, Q.; Wu, Q.; Chen, B.; Zhang, M. Virome Identification and Characterization of Fusarium sacchari and F. andiyazi: Causative Agents of Pokkah Boeng Disease in Sugarcane. Front Microbiol 2020, 11. [Google Scholar] [CrossRef]
- Vermeulen, M.; Rothmann, L.A.; Swart, W.J.; Gryzenhout, M. Fusarium casha Sp. Nov. and F. curculicola Sp. Nov. in the Fusarium fujikuroi Species Complex Isolated from Amaranthus cruentus and Three Weevil Species in South Africa. Diversity 2021, 13. [Google Scholar] [CrossRef]
- Bogner, C.W.; Kariuki, G.M.; Elashry, A.; Sichtermann, G.; Buch, A.K.; Mishra, B.; Thines, M.; Grundler, F.M.W.; Schouten, A. Fungal Root Endophytes of Tomato from Kenya and Their Nematode Biocontrol Potential. Mycol Prog 2016, 15. [Google Scholar] [CrossRef]
- Africa, S. Environmental India BBA 69553; DAOM 225121; FRC O-1116; 2021; Vol. 46.
- Moore GG and Fapohunda, SO. Molecular Investigations of Food-Borne Cladosporium and Fusarium. J Bacteriol Mycol 2016, 3, 1024. [Google Scholar]
- Perdomo, H.; Sutton, D.A.; García, D.; Fothergill, A.W.; Cano, J.; Gené, J.; Summerbell, R.C.; Rinaldi, M.G.; Guarro, J. Spectrum of Clinically Relevant Acremonium species in the United States. J Clin Microbiol 2011, 49, 243–256. [Google Scholar] [CrossRef] [PubMed]
- MINICOM 2021 Ministere de commerce trade report (2021).
Figure 1.
Locations in Mali, Nigeria, and Rwanda where rice samples were taken. A. Locations of Mali, Nigeria, and Rwanda in Africa; B, location of the area where samples were collected in Mali; C, locations in Nigeria; D, locations in Rwanda.
Figure 1.
Locations in Mali, Nigeria, and Rwanda where rice samples were taken. A. Locations of Mali, Nigeria, and Rwanda in Africa; B, location of the area where samples were collected in Mali; C, locations in Nigeria; D, locations in Rwanda.
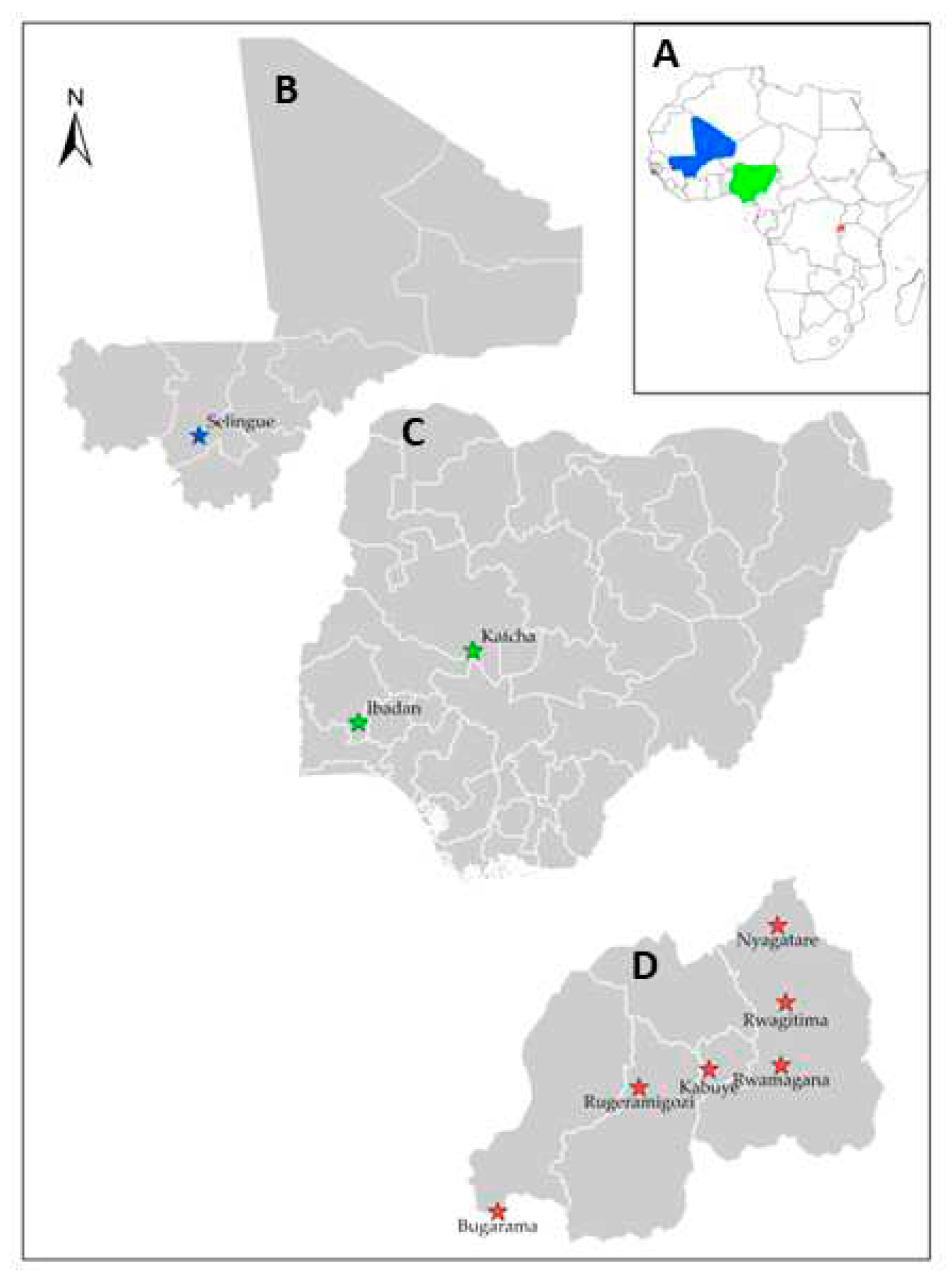
Figure 2.
Diseased rice plants showing typical sheath rot disease symptoms. A: Typical sheath browning characteristic of sheath rot disease on rice field at AfricaRice experimental field, Ibadan-Nigeria. B Greyish-brown lesions on the leaf flags enclosing the panicle observed during the screenhouse experiment. C: Emerged brownish panicles, chaffy and sterile grains showing typical sheath rot disease symptoms.
Figure 2.
Diseased rice plants showing typical sheath rot disease symptoms. A: Typical sheath browning characteristic of sheath rot disease on rice field at AfricaRice experimental field, Ibadan-Nigeria. B Greyish-brown lesions on the leaf flags enclosing the panicle observed during the screenhouse experiment. C: Emerged brownish panicles, chaffy and sterile grains showing typical sheath rot disease symptoms.

Figure 3.
Phylogenetic tree based on maximum-likelihood inferred from partial EF-1α sequences of four Fusarium species complexes using IQ-Tree with GTR + G + I model and annotated using the iTOL software. Cylindrocarpon sp. AC2011 strain CPC 13531 was used as an outgroup. Isolates in colour and bold were obtained in this study (Blue – Mali, Green – Nigeria and Red – Rwanda). FIESC: Fusarium incarnatum-equiseti species complex; FSAMSC: Fusarium sambucinum species complex; FOSC: Fusarium oxysporum species complex; FFSC: Fusarium fujikuroi species complex.
Figure 3.
Phylogenetic tree based on maximum-likelihood inferred from partial EF-1α sequences of four Fusarium species complexes using IQ-Tree with GTR + G + I model and annotated using the iTOL software. Cylindrocarpon sp. AC2011 strain CPC 13531 was used as an outgroup. Isolates in colour and bold were obtained in this study (Blue – Mali, Green – Nigeria and Red – Rwanda). FIESC: Fusarium incarnatum-equiseti species complex; FSAMSC: Fusarium sambucinum species complex; FOSC: Fusarium oxysporum species complex; FFSC: Fusarium fujikuroi species complex.
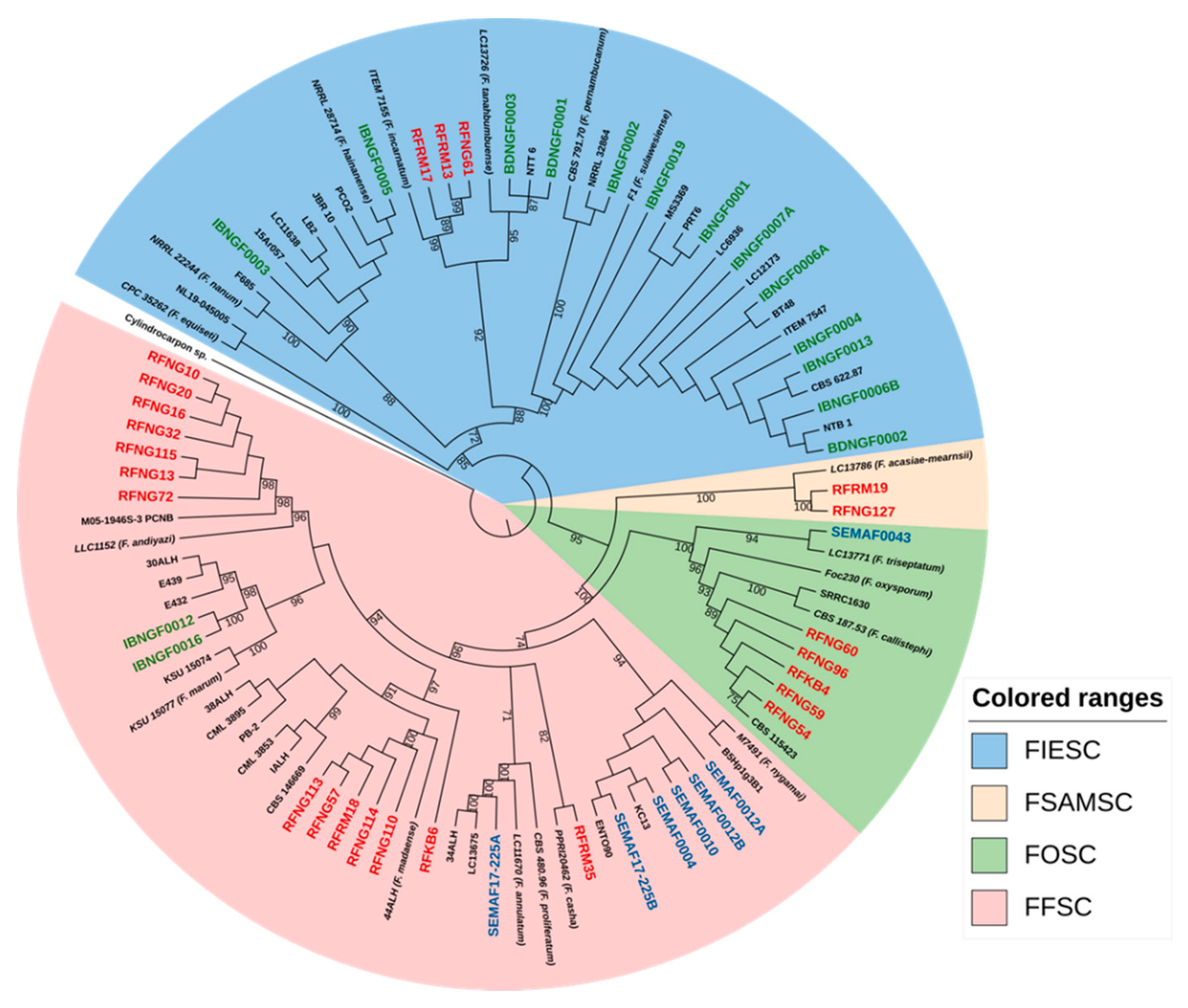
Figure 4.
Phylogenetic tree based on the concatenation of both ITS and Actin region of Sarocladium species obtained in this study. The tree was generated using the Jukes Cantor model and the maximum likelihood method in MEGA. Sarocladium zeae strain CBS 800.69 was used as an outgroup. Sequences in colour and bold were obtained in this study (Blue – Mali, Green – Nigeria and Red – Rwanda).
Figure 4.
Phylogenetic tree based on the concatenation of both ITS and Actin region of Sarocladium species obtained in this study. The tree was generated using the Jukes Cantor model and the maximum likelihood method in MEGA. Sarocladium zeae strain CBS 800.69 was used as an outgroup. Sequences in colour and bold were obtained in this study (Blue – Mali, Green – Nigeria and Red – Rwanda).
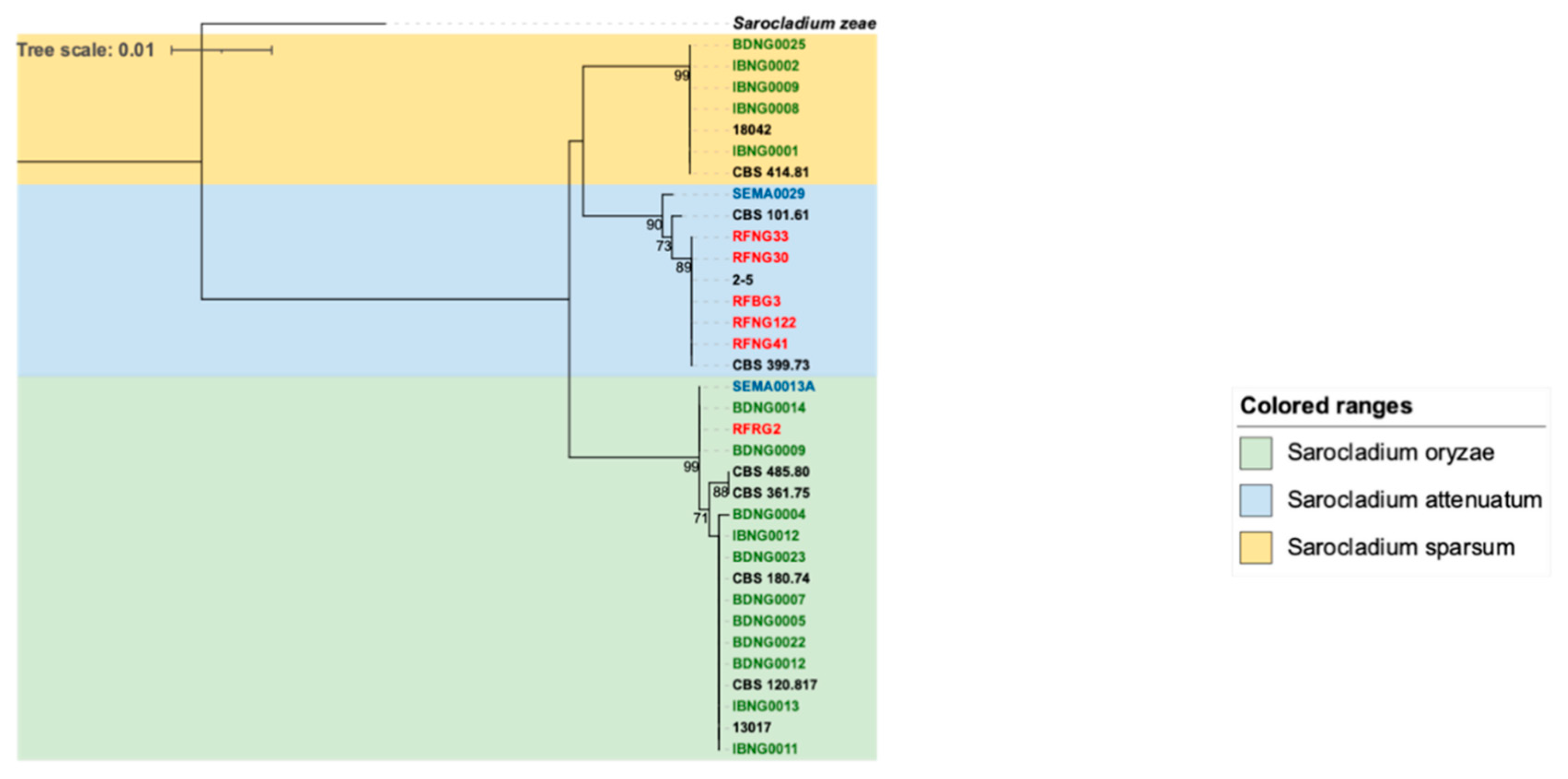
Figure 5.
Mean lesion length (millimeters) at 8 dpi on FARO44 rice cultivar inoculated with isolates from four Fusarium species 6 weeks after planting. Different letters indicate statistically significant differences based on Kruskal-Wallis Rank Sum test followed by a post hoc Mann-Whitney test. Statistical significance was defined as P= 0.05.
Figure 5.
Mean lesion length (millimeters) at 8 dpi on FARO44 rice cultivar inoculated with isolates from four Fusarium species 6 weeks after planting. Different letters indicate statistically significant differences based on Kruskal-Wallis Rank Sum test followed by a post hoc Mann-Whitney test. Statistical significance was defined as P= 0.05.
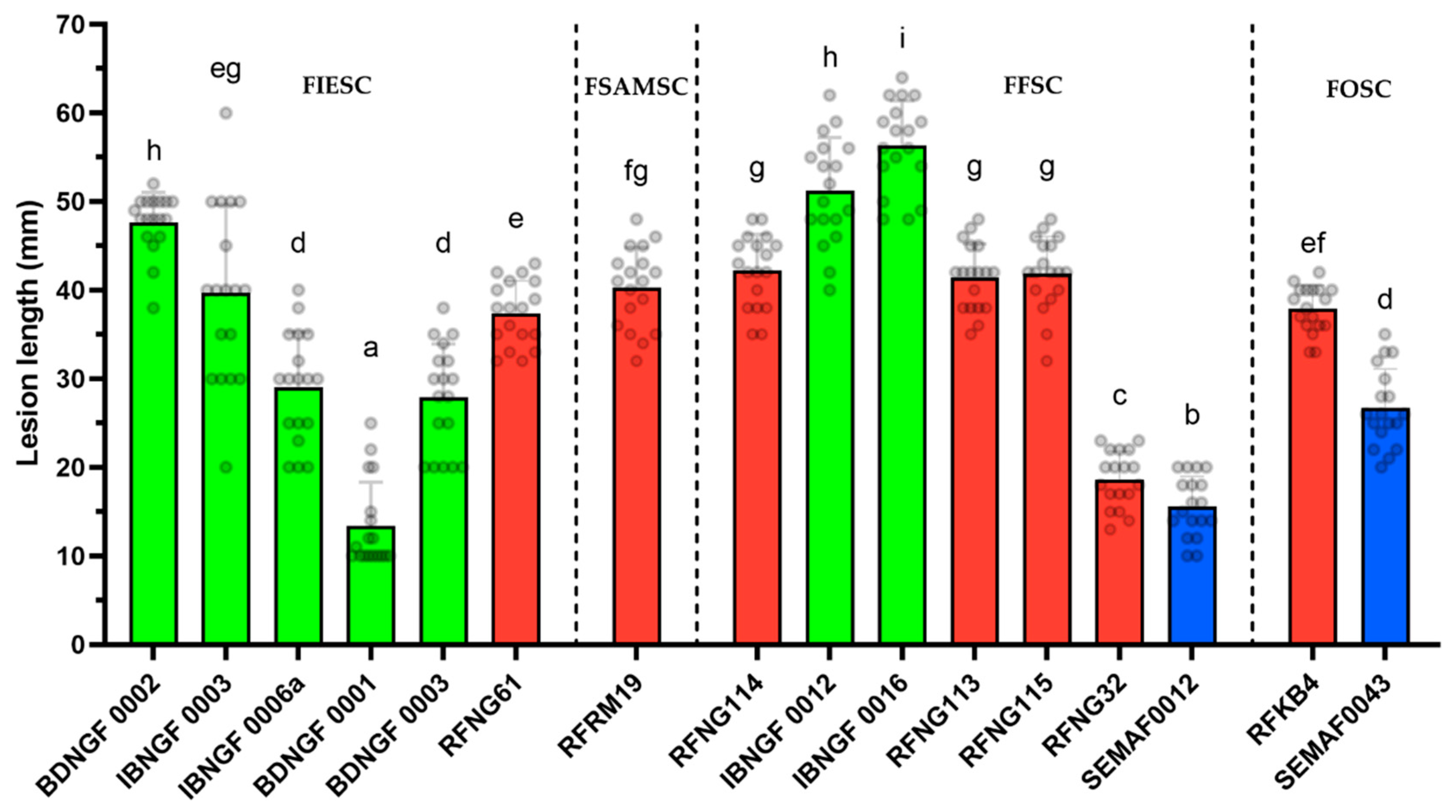
Figure 6.
Mean lesion length (millimeters) at 8 dpi on FARO44 rice cultivar inoculated with Sarocladium species after 6 weeks of planting. Different letters indicate statistically significant differences based on Kruskal-Wally’s Rank Sum test followed by a post hoc Mann-Whitney test. Statistical significance was defined as P= 0.05.
Figure 6.
Mean lesion length (millimeters) at 8 dpi on FARO44 rice cultivar inoculated with Sarocladium species after 6 weeks of planting. Different letters indicate statistically significant differences based on Kruskal-Wally’s Rank Sum test followed by a post hoc Mann-Whitney test. Statistical significance was defined as P= 0.05.
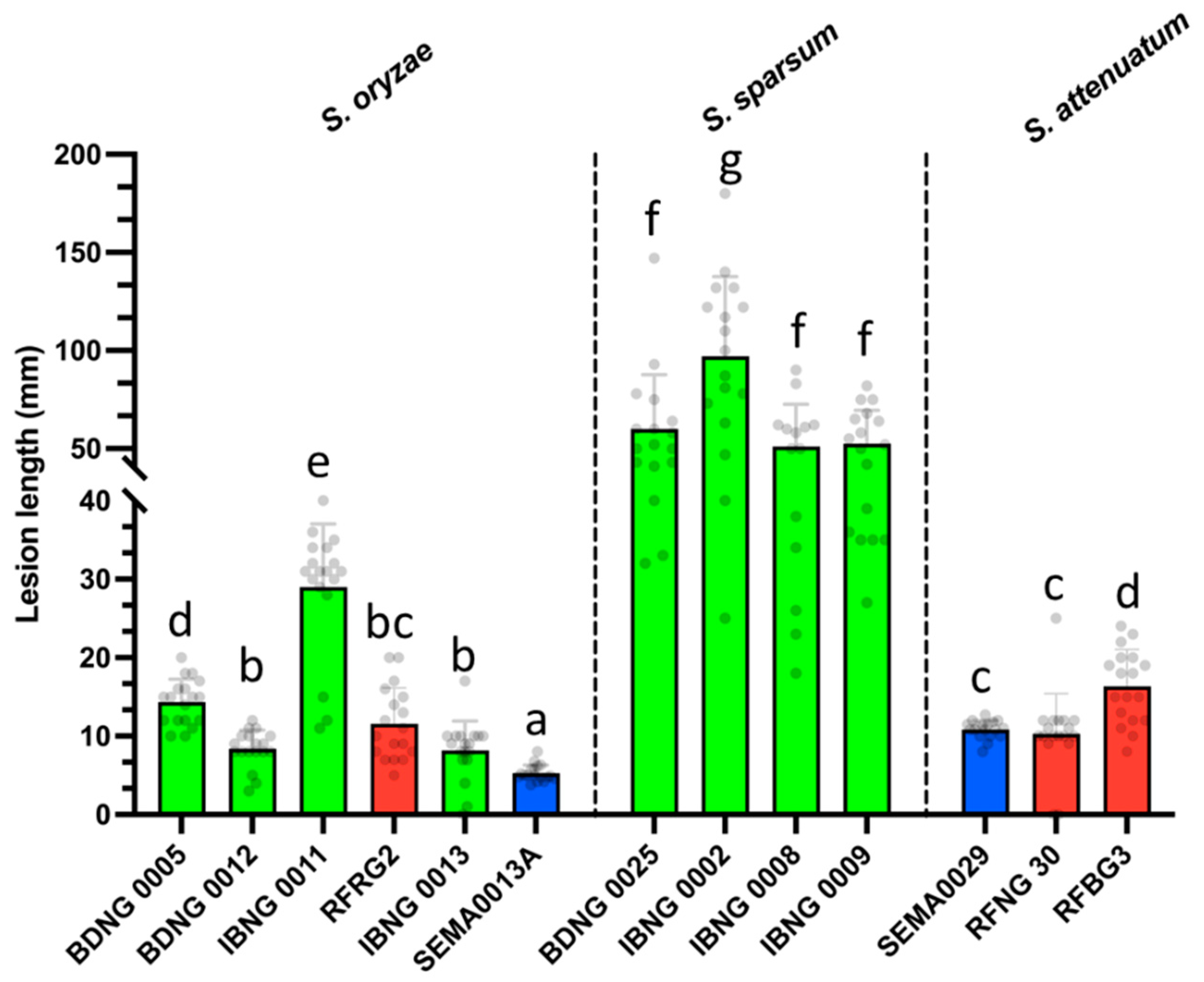
Table 1.
Agro-ecological details of the sampling regions in Mali, Nigeria, and Rwanda
| Location | Ecology | Annual precipitation (mm) | Temperature (°C) | Ecosystem | Elevation (m) |
| Nigeria | |||||
| Ibadan | Derived savannah | 1300-1500 | 25-35 | Irrigated lowland | 225 |
| Katcha | Southern Guinea Savannah | 900-1000 | 28-40 | Rainfed lowland | 123 |
| Mali | |||||
| Selingue | Sudan Guinea Savannah | 600 | 35-50 | Irrigated lowland | 351 |
| Rwanda | |||||
| Bugarama | Mosaic Vegetation and Forest (West) | 1098 | 24 | Irrigated marshland | 900 |
| Kabuye | Mosaic Vegetation and Forest (Central) | 951 | 22 | Irrigated marshland | 1270 |
| Nyagatare | Savannah (East) | 783 | 20 | Irrigated marshland | 1470 |
| Rwamagana | Savannah (East) | 979 | 19 | Irrigated marshland | 1680 |
| Rugeramigozi | Mosaic Vegetation and Forest (South) | 1154 | 19 | Irrigated marshland | 1706 |
Table 2.
Fusarium isolates obtained from rice plants showing sheath rot disease symptoms in Nigeria, Mali and Rwanda.
Table 2.
Fusarium isolates obtained from rice plants showing sheath rot disease symptoms in Nigeria, Mali and Rwanda.
| Origin | Strain code | Species | Species complex | Host part | Year of isolation | Genbank EF-1α |
| Nigeria | ||||||
| Ibadan | IBNGF0001 | F. sulawesiense | FIESC 16 | Seed | 2017 | MN539083 |
| Ibadan | IBNGF0002 | F. pernambucanum | FIESC 17 | Sheath | 2017 | MN539084 |
| Ibadan | IBNGF0003 | F. hainanense | FIESC 26 | Seed | 2017 | MN539085 |
| Ibadan | IBNGF0004 | F. sulawesiense | FIESC 16 | Sheath | 2017 | MN539086 |
| Ibadan | IBNGF0005 | F. hainanense | FIESC 26 | Sheath | 2017 | MN539087 |
| Ibadan | IBNGF0006A | F. sulawesiense | FIESC 16 | Sheath | 2017 | MN539088 |
| Ibadan | IBNGF0006B | F. sulawesiense | FIESC 16 | Sheath | 2017 | MN539089 |
| Ibadan | IBNGF0007A | F. sulawesiense | FIESC 16 | Sheath | 2017 | MN539090 |
| Ibadan | IBNGF0012 | F. marum | FFSC | Sheath | 2017 | MN539096 |
| Ibadan | IBNGF0013 | F. sulawesiense | FIESC 16 | Sheath | 2017 | MN539091 |
| Ibadan | IBNGF0016 | F. marum | FFSC | Sheath | 2017 | MN539097 |
| Ibadan | IBNGF0019 | F. sulawesiense | FIESC 16 | Sheath | 2017 | MN539092 |
| Katcha | BDNGF0001 | F. tanahbumbuense | FIESC 24 | Sheath | 2017 | MN539091 |
| Katcha | BDNGF0002 | F. sulawesiense | FIESC 16 | Seed | 2017 | MN539094 |
| Katcha | BDNGF0003 | F. tanahbumbuense | FIESC 24 | Seed | 2017 | MN539095 |
| Mali | ||||||
| Selingue | SEMAF0004 | F. nygamai | FFSC | Seed | 2017 | MN539098 |
| Selingue | SEMAF0010 | F. nygamai | FFSC | Seed | 2017 | MN539099 |
| Selingue | SEMAF0012A | F. nygamai | FFSC | Seed | 2017 | MN539100 |
| Selingue | SEMAF0012B | F. nygamai | FFSC | Sheath | 2017 | MN539101 |
| Selingue | SEMAF17-225A | F. annulatum | FFSC | Sheath | 2017 | MN539103 |
| Selingue | SEMAF17-225B | F. nygamai | FFSC | Seed | 2017 | MN539102 |
| Selingue | SEMAF0043 | F. triseptatum | FOSC | Sheath | 2017 | MN539104 |
| Rwanda | ||||||
| Kabuye | RFKB4 | F. callistephi | FOSC | Seed | 2013 | KX424544 |
| Kabuye | RFKB6 | F. madaense | FFSC | Sheath | 2013 | KX424545 |
| Nyagatare | RFNG10 | F. andiyazi | FFSC | Sheath | 2011 | KX424546 |
| Nyagatare | RFNG13 | F. andiyazi | FFSC | Sheath | 2011 | KX424552 |
| Nyagatare | RFNG16 | F. andiyazi | FFSC | Sheath | 2011 | KX424553 |
| Nyagatare | RFNG20 | F. andiyazi | FFSC | Sheath | 2011 | KX424554 |
| Nyagatare | RFNG32 | F. andiyazi | FFSC | Sheath | 2011 | KX424555 |
| Nyagatare | RFNG54 | F. callistephi | FOSC | Sheath | 2011 | OQ909428 |
| Nyagatare | RFNG57 | F. madaense | FFSC | Sheath | 2011 | KX424556 |
| Nyagatare | RFNG59 | F. callistephi | FOSC | Sheath | 2011 | KX424557 |
| Nyagatare | RFNG60 | F. callistephi | FOSC | Sheath | 2011 | OQ909429 |
| Nyagatare | RFNG61 | F. incarnatum | FIESC 38 | Sheath | 2011 | OQ909431 |
| Nyagatare | RFNG72 | F. andiyazi | FFSC | Sheath | 2011 | OQ909425 |
| Nyagatare | RFNG96 | F. callistephi | FOSC | Sheath | 2011 | OQ909430 |
| Nyagatare | RFNG110 | F. madaense | FFSC | Sheath | 2011 | OQ909426 |
| Nyagatare | RFNG113 | F. madaense | FFSC | Sheath | 2011 | KX424548 |
| Nyagatare | RFNG114 | F. madaense | FFSC | Sheath | 2011 | KX424549 |
| Nyagatare | RFNG115 | F. andiyazi | FFSC | Sheath | 2011 | KX424550 |
| Nyagatare | RFNG127 | F. acasiae mearnsii | FSAMSC | Sheath | 2013 | KX424551 |
| Rwamagana | RFRM13 | F. incarnatum | FIESC 38 | Sheath | 2013 | OQ867255 |
| Rwamagana | RFRM17 | F. incarnatum | FIESC 38 | Sheath | 2013 | OQ909427 |
| Rwamagana | RFRM18 | F. madaense | FFSC | Sheath | 2013 | KX424559 |
| Rwamagana | RFRM19 | F. acasiae-mearnsii | FSAMSC | Sheath | 2013 | KX424560 |
| Rwamagana | RFRM35 | F. casha | FFSC | Sheath | 2013 | KX424561 |
Table 3.
Sarocladium isolates obtained from rice plants showing sheath rot disease symptoms in Nigeria, Mali and Rwanda.
Table 3.
Sarocladium isolates obtained from rice plants showing sheath rot disease symptoms in Nigeria, Mali and Rwanda.
| Strain code | Species | Host/Part | Year of Isolation | Genbank ITS | Genbank ACTIN | |
| Nigeria | ||||||
| Ibadan | IBNG0001 | S. sparsum | Sheath | 2017 | MN389594 | MN783308 |
| Ibadan | IBNG0002 | S. sparsum | Sheath | 2017 | MN389595 | MN783309 |
| Ibadan | IBNG0008 | S. sparsum | Sheath | 2017 | MN389596 | MN783310 |
| Ibadan | IBNG0009 | S. sparsum | Seed | 2017 | MN389597 | MN783311 |
| Ibadan | IBNG0011 | S. oryzae | Sheath | 2017 | MN389589 | MN783312 |
| Ibadan | IBNG0012 | S. oryzae | Sheath | 2017 | MN389590 | MN783313 |
| Ibadan | IBNG0013 | S. oryzae | Sheath | 2017 | MN389591 | MN783314 |
| Katcha | BDNG0004 | S. oryzae | Seed | 2017 | MN389581 | MN783299 |
| Katcha | BDNG0005 | S. oryzae | Sheath | 2017 | MN389582 | MN783300 |
| Katcha | BDNG0007 | S. oryzae | Seed | 2017 | MN389583 | MN783301 |
| Katcha | BDNG0009 | S. oryzae | Sheath | 2017 | MN389584 | MN783302 |
| Katcha | BDNG0012 | S. oryzae | Sheath | 2017 | MN389585 | MN783303 |
| Katcha | BDNG0014 | S. oryzae | Sheath | 2017 | MN389586 | MN783304 |
| Katcha | BDNG0022 | S. oryzae | Sheath | 2017 | MN389587 | MN783305 |
| Katcha | BDNG0023 | S. oryzae | Sheath | 2017 | MN389588 | MN783306 |
| Katcha | BDNG0025 | S. sparsum | Seed | 2017 | MN389593 | MN783307 |
| Mali | ||||||
| Selingue | SEMA0013A | S. oryzae | Sheath | 2017 | MN641009 | MN783315 |
| Selingue | SEMA0029 | S. attenuatum | Sheath | 2017 | MN641010 | MN783316 |
| Rwanda | ||||||
| Bugarama | RFBG3 | S. attenuatum | Sheath | 2011 | KX424828 | OP374130 |
| Nyagatare | RFNG30 | S. attenuatum | Sheath | 2011 | KX424536 | OP374131 |
| Nyagatare | RFNG33 | S. attenuatum | Sheath | 2011 | KX424537 | OP374132 |
| Nyagatare | RFNG41 | S. attenuatum | Sheath | 2011 | KX424538 | OP374133 |
| Nyagatare | RFNG122 | S. attenuatum | Sheath | 2011 | KX424531 | OP374134 |
| Rugeramigozi | RFRG2 | S. oryzae | Sheath | 2013 | KX424542 | OP374135 |
| CBS isolates | ||||||
| Mexico | CBS 101.61 | S. attenuatum | NA | 1959 | MN389592 | MN783317 |
| Kenya | CBS 361.75 | S. oryzae | NA | NA | MN389580 | MN783318 |
| Panama | CBS 120.817 | S. oryzae | NA | NA | MN389579 | MN783319 |
| Australia | CBS 485.80 | S. oryzae | Sheath | 1980 | MN389598 | MN783320 |
Table 4.
Mycotoxins produced by the Fusarium spp in vitro. The values are concentration in (µg/kg) using LC-MS/MS. All strains were tested, only mycotoxins producers are reported.
Table 4.
Mycotoxins produced by the Fusarium spp in vitro. The values are concentration in (µg/kg) using LC-MS/MS. All strains were tested, only mycotoxins producers are reported.
| Fusarium sp. | Strain code | Mycotoxin (µg/kg) | |||||||
| NIV | NEO | FX | DAS | FB1 | FB2 | FB3 | ZEN | ||
| FIESC | |||||||||
| F. hainanense | IBNGF0003 | 32432 | |||||||
| F. hainanense | IBNGF0005 | 29681 | |||||||
| F. sulawesiense | IBNGF0004 | 73 | 106 | ||||||
| F. sulawesiense | IBNGF0006A | 115 | 20 | 196 | |||||
| F. sulawesiense | IBNGF0006B | 95 | |||||||
| F. sulawesiense | IBNGF0007A | 56 | |||||||
| F. sulawesiense | IBNGF0013 | 47 | 16 | 211 | |||||
| F. sulawesiense | IBNGF00019 | 2575 | 118 | 1370 | 53 | ||||
| F. sulawesiense | BDNGF0002 | 81 | |||||||
| F. pernambucanum | IBNGF0002 | 122 | 25 | 355 | |||||
| F. tanahbumbuense | BDNGF0001 | 12 | |||||||
| F. tanahbumbuense | BDNGF0003 | 53 | |||||||
| FFSC | |||||||||
| F. annulatum | SEMAF17-225A | 567 | 34 | 159 | 77 | ||||
| F. madaense | RFRM18 | 195 | 1120 | 1349 | |||||
| F. nygamai | SEMAF0010 | 69679 | 4234 | 573 | |||||
| F. nygamai | SEMAF0012A | 118024 | 9325 | 702 | |||||
| F. nygamai | SEMAF0012B | 53118 | 3389 | 355 | |||||
| FOSC | |||||||||
| F. triseptatum | SEMAF0043 | 40 | |||||||
| FSAMSC | |||||||||
| F. acasiae-mearnsii | RFRM19 | 82 | 178 | 330 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



