
Submitted:
22 January 2025
Posted:
23 January 2025
You are already at the latest version
Abstract
Methicillin-resistant Staphylococcus aureus (MRSA) poses a significant clinical challenge due to its resistance to multiple antibiotics. The urgent need for new therapeutic approaches has led to the exploration of natural compounds as potential treatments, particularly those targeting key bacterial proteins involved in antibiotic resistance. This study focuses on the Multidrug ABC Transporter and MecA proteins, which play crucial roles in MRSA's pathogenicity and resistance mechanisms.
Using computational techniques and molecular docking methods, we assessed the interactions of 80 natural compounds against S. aureus multidrug ABC transporter SAV1866 (SAV1866) and MecA proteins. Our analysis revealed 14 compounds with robust binding to SAV1866 and one compound with strong affinity for MecA. Notably, these compounds showed weaker affinities for MgrA, MepR, and arlR proteins, suggesting specificity in their interactions.
Among the 15 promising compounds identified, 1',2-Binaphthalen-4-one-2',3-dimethyl-1,8'-epoxy-1,4',5,5',8,8'-hexahydroxy-8-O-β-glucopyranosyl-5'-O-β-xylopyranosyl(1→6)-β-glucopyranoside, Cis-3,4-dihydrohamacanthin b, and Mamegakinone exhibited the highest binding affinity to S. aureus SAV1866. These compounds represent diverse chemical classes including alkaloids, indole derivatives, naphthalenes, and naphthoquinones, offering a range of structural scaffolds for further drug development.
Our findings provide valuable insights into potential new antibacterial agents targeting S. aureus SAV1866 and MecA proteins. These results lay the groundwork for future in vitro and in vivo studies to validate these compounds' efficacy in combating MRSA infections, potentially leading to the development of novel therapeutic strategies against antibiotic-resistant bacteria.
Keywords:
Methicillin-resistant Staphylococcus aureus (MRSA)
; Multidrug ABC Transporter
; MecA
; antibiotic resistance
; molecular docking
; and natural compounds
1. Introduction
Methicillin-resistant Staphylococcus aureus (MRSA) is a major cause of nosocomial and community-acquired infections worldwide (Brown, et al., 2021; Lee, et al., 2018; Morell and Balkin, 2010; Turner, et al., 2019). Overuse and misuse of antibiotics are among the factors that contribute to the development of antibiotic resistance (Dugassa and Shukuri, 2017; English and Gaur, 2010). MRSA poses a significant public health challenge, as it is resistant to many antibiotics that are commonly used to treat bacterial infections (Lee, et al., 2018; Morell and Balkin, 2010). This resistance is primarily due to the presence of a mecA gene, which encodes a protein called PBP2a that enables the bacteria to evade the effects of many antibiotics (Elal Mus, et al., 2019; Müller, et al., 2015; Otarigho and Falade, 2023; Wielders, et al., 2002). Another critical protein involved in antibiotic resistance is the multidrug ABC transporter SAV1866. This protein is responsible for the active efflux of a broad spectrum of antibiotics from the bacterial cell, utilizing energy derived from ATP hydrolysis. By actively pumping out antimicrobial agents, SAV1866 reduces the intracellular concentration of these drugs, thereby decreasing their efficacy and contributing significantly to the survival of bacteria in the presence of antibiotics (Akhtar and Turner, 2022; Dawson and Locher, 2006; Velamakanni, et al., 2008; Yao, 2011; Yoshikai, et al., 2016). Furthermore, the genome of S. aureus also contains other genes that encode antibiotic-resistance proteins, such as MgrA, MepR, and arlR, (Otarigho and Falade, 2018; Velamakanni, et al., 2008). Although some antibiotics such as vancomycin and linezolid are relatively effective against MRSA (Brown, et al., 2021), treatment of infections can be difficult due to the emergence of new resistant strains (Lowy, 2003; Morell and Balkin, 2010). Thus, there is an urgent need for alternative treatments and discovery of new medications or therapeutic approaches to combat MRSA (Kurlenda and Grinholc, 2012).
Natural agents, including plant extracts and essential oils, hold great promise as alternative therapies for treating bacterial infections (Langeveld, et al., 2014; Nascimento, et al., 2000). These agents have contributed an extensive history of traditional medicinal usage and have demonstrated substantial antibacterial potency (Langeveld, et al., 2014; Martin and Ernst, 2003; Nascimento, et al., 2000). A comprehensive body of research provides compelling evidence substantiating the efficacy of natural agents in treating bacterial pathogens (Nascimento, et al., 2000). Remarkably, these natural agents have displayed efficacy against both Gram-positive and Gram-negative bacterial strains, including antibiotic-resistant strains such as MRSA (Álvarez-Martínez, et al., 2021; Brown, et al., 2021; Dahiya and Purkayastha, 2012; Nascimento, et al., 2000).
The unique effectiveness of natural agents against bacterial infections often emanates from their distinct mechanisms compared to antibiotics (Álvarez-Martínez, et al., 2021; Dugassa and Shukuri, 2017). These mechanisms include disrupting bacterial cell membranes, suppressing enzyme activity, or interrupting bacterial DNA replication (Álvarez-Martínez, et al., 2021). By simultaneously targeting multiple cellular sites, these compounds aid in the disruption of vital cellular processes, thereby inhibiting bacterial proliferation, diminishing pathogenicity, and enhancing susceptibility to host immune responses an important arm of chemotherapeutic efficacy (Dugassa and Shukuri, 2017). Furthermore, natural agents have exhibited efficacy against a spectrum of bacterial infections, encompassing pneumonia, urinary tract infections, and cutaneous infections (Álvarez-Martínez, et al., 2021). Additionally, natural agents may yield fewer adverse effects when compared to antibiotics, largely stemming from their selective toxicity against bacterial cells while leaving human cells untouched (Álvarez-Martínez, et al., 2021; Dahiya and Purkayastha, 2012).
Consequently, the investigation of natural compounds as multi-target therapeutics against bacterial pathogens provides a promising area for identifying new drugs. These compounds exhibit a diverse array of chemical structures and mechanisms of action, making them attractive sources for novel chemotherapeutics (Álvarez-Martínez, et al., 2021; Dahiya and Purkayastha, 2012). Given these advantages, further research is required to identify natural compounds possessing antibacterial properties, particularly those demonstrating robust affinity to well-known MRSA antibiotic resistance-associated proteins such as MecA, SAV1866, MgrA, MepR, and arlR (Elmaidomy, et al., 2022; Nandhini, et al., 2023).
Molecular docking utilizes computational techniques to study the interaction between small molecules and macromolecules, such as proteins (Adelusi, et al., 2022; Agnihotry, et al., 2020; Ferreira, et al., 2015; Otarigho and Falade, 2023; Pinzi and Rastelli, 2019). In recent years, molecular docking has been used to evaluate the potential of natural agents as antibacterials (Nandhini, et al., 2023; Skariyachan, et al., 2011). This technique has been invaluable in providing helpful insights into the mechanisms of action of natural agents against bacterial proteins (Nandhini, et al., 2023). Molecular docking assists in predicting the binding affinity of a natural agent to a bacterial protein, a process important in the development of new antibacterial agents, as it can guide the selection of promising compounds for further study (Agnihotry, et al., 2020; Ferreira, et al., 2015; Meng, et al., 2011). In this study, we aimed to explore the efficacy of natural compounds derived from the South African Natural Compounds Database (SANCDB) (Diallo, et al., 2021) against major antibiotic-resistance proteins found in MRSA. Some of these proteins, namely MecA, SAV1866, MgrA, MepR, and arlR, are known to contribute significantly to the development of antibiotic resistance in MRSA strains (Otarigho and Falade, 2018). To assess the binding capabilities of these natural compounds, we utilized a molecular docking screening technique. Our analysis revealed that most of the natural compounds exhibited binding affinity towards the MRSA multidrug ABC transporter protein, which is a crucial component in the efflux of drugs from MRSA cells (Akhtar and Turner, 2022; Yoshikai, et al., 2016). These compounds primarily belonged to diverse chemical groups such as binaphthalenone, glycoside, organooxygen, alkaloid, indole, carboxylic acids, naphthoquinone, naphthalenes, and flavonoids (Chukwujekwu, et al., 2011; Diallo, et al., 2021; Kiplimo and Koorbanally, 2012; Lall, et al., 2017; Li, et al., 1998; Pendota, et al., 2015; Van der Kooy, et al., 2006; Zoraghi, et al., 2011). Additionally, our study identified a specific compound that displayed a remarkable affinity for the MecA protein, a key regulator of MRSA virulence (Elal Mus, et al., 2019; Kiplimo and Koorbanally, 2012; Müller, et al., 2015). The strong binding demonstrated between this natural compound and MecA indicates its potential as an inhibitor of MecA-mediated virulence, which could potentially reduce the pathogenicity of MRSA strains. In this study, the primary objective was to identify compounds exhibiting antibacterial, antimicrobial, and bacteriostatic activities. Our work provides valuable insights into using natural compounds as inhibitors of antibiotic-resistance proteins in MRSA. By targeting these proteins, natural compounds hold promise as potential therapeutic agents to combat MRSA infections and overcome antibiotic resistance.
2. Materials and Methods
2.1. Data mining and Retrieval of Antibiotic Resistance Proteins
This study focused on antibiotic-resistance proteins, which play significant roles as global regulators and major virulence determinants in S. aureus (Elal Mus, et al., 2019; Müller, et al., 2015; Velamakanni, et al., 2008; Wielders, et al., 2002; Yao, 2011; Yoshikai, et al., 2016). Specifically, the multidrug ABC transporter SAV1866, MecA, MgrA, MepR, and arlR were of interest, and their structural information was obtained from the Protein Data Bank (PDB), accessible at https://www.rcsb.org/ (Berman, et al., 2000; Burley, et al., 2017). The downloaded experimental data snapshots of the protein structures from the PDB provide specific details about the methods employed for structure determination, along with relevant metrics. The experimental data snapshot of SAV1866 (PDB ID: 2HYD) and MecA (PDB ID: 1MWS) indicates an X-ray diffraction method with a resolution of 3.00 Å and 2.00 Å respectively. The R-Value Free, R-Value Work, and R-Value reported for SAV1866 were reported as 0.272, 0.255, and 0.255, respectively (Dawson and Locher, 2006), while the R-Value Free, R-Value Work, and R-Value reported for MecA were reported as 0.274, 0.234, and 0.234, respectively (Lim and Strynadka, 2002) . Similarly, for MgrA (PDB ID: 2BV6), the experimental data snapshot reveals an X-ray diffraction method with a resolution of 2.80 Å, and the R-Value Free, R-Value Work, and R-Value Observed were reported as 0.292, 0.250, and 0.250, respectively (Chen, et al., 2006). For MepR (PDB ID: 4L9N), the experimental data snapshot indicates an X-ray diffraction method with a resolution of 1.60 Å, and the R-Value Free, R-Value Work, and R-Value Observed were reported as 0.217, 0.190, and 0.191, respectively (Birukou, et al., 2013). For arIR (PDB ID: 6IS3), the experimental data snapshot indicates an X-ray diffraction method with a resolution of 1.55 Å, and the R-Value Free, R-Value Work, and R-Value Observed were reported as 0.206, 0.173, and 0.176, respectively (Ouyang, et al., 2019). These experimental data snapshots offer valuable insights into the quality and reliability of the obtained protein structures. The resolution values indicate the level of detail at which the structures were determined, while the R-values provide an assessment of the agreement between the experimental data and the refined protein models (Feig, 2017; Gore, et al., 2017).
2.2. Data Mining and Retrieval of Natural Compounds with Known Antibiotic Activities
The exploration for antibacterial compounds was carried out using the South African Natural Compounds Database (SANCDB) (Diallo, et al., 2021; Hatherley, et al., 2015), which can be accessed at https://sancdb.rubi.ru.ac.za/. The compounds chosen for the search were already recognized for their antibiotic properties against a diverse range of bacteria (Diallo, et al., 2021). The SANCDB serves as a comprehensive and freely accessible repository of natural chemical compounds originating from South African biodiversity (Diallo, et al., 2021; Hatherley, et al., 2015). Eighty compounds were selected and mined in the PDB format. The 2D chemical structure was analyzed and obtained using Marvin JS (https://marvinjs-demo.chemaxon.com/latest/index.html), an online chemical editor developed by ChemAxon that enables users to draw and visualize chemical structures on the web.
2.3. Docking of the Natural Compounds Against the MRSA Antibiotic-Resistance Protein
Each of the 80 compounds obtained from the previous step was subjected to molecular docking against the SAV1866, MecA, MgrA, MepR, and arlR using the CB-Dock2 server, accessible at https://cadd.labshare.cn/cb-dock2/php/blinddock.php (Liu, et al., 2022). Before the molecular docking, we anticipated potential binding sites on distinct MRSA antibiotic resistance proteins for the diverse compounds using the CB-Dock2 server. Only compounds that exhibited a strong affinity, with Vina scores of -10 kcal/mol and below, were selected for further analysis. The selection of compounds with high binding affinity is crucial as it indicates a stronger potential for specific interactions with the target proteins. These compounds are more likely to exhibit favorable binding and potentially possess greater therapeutic efficacy against the target proteins.
2.4. Analyses of the Binding Affinity of the Antibiotic Resistance Proteins to the Natural Compounds
In this study, we utilized computational tools to analyze and visualize the binding affinities of 80 natural compounds against the SAV1866, MecA, MgrA, MepR, and arlR. The vina scores were visualized using the Datawrapper tool (https://www.datawrapper.de/). By inputting the vina scores into Datawrapper, we generated a chart to represent the binding affinities of the 80 compounds against the target proteins. This visualization provided an overview of the compound-protein interactions and allowed for easy comparison and interpretation of the data. The significant binding affinities with a Vina Score of less than -10 kcal/mol were further analyzed with Datawrapper. This visualization provided a more detailed and focused representation of the interaction between the identified compounds and proteins. Additionally, we employed the TBtool (Chen, et al., 2018), which was downloaded from https://github.com/CJ-Chen/TBtools, to create a heatmap to visualize the 15 natural compounds that were significantly bound to SAV1866 and MecA. The affinities of these 15 compounds to the MgrA, and MepR proteins were also included in this analysis. The heatmap representation allowed us to observe patterns and trends in the binding affinities of these compounds across multiple proteins simultaneously. This visualization provided a comprehensive view of the compound-protein interactions and highlighted the compounds that exhibited high binding affinity to multiple target proteins.
3. Results
After mining the SANCDB (South African Natural Compound Database), we identified 80 compounds sourced from tropical plants (Table S1-5). These compounds were sourced from different plant species, these include, Scutia myrtina, Erythrina abyssinica, Eucalyptus globulus, and Aloe vera which have the highest number of compounds with antibacterial activities (Figure 1A). Among these naturally derived compounds, 68 have confirmed antibacterial properties, while 11 and 1 respectively exhibit antimicrobial and bacteriostatic properties (Figure 1B and Table S1-5). This suggests that the activities described for these plants make them a rich repository of bioactive compounds with antibacterial potential.
We used molecular docking to evaluate the interactions of the 80 compounds with the five protein structures, SAV1866, MecA, MgrA, MepR, arlR, which were obtained from the PDB database (Figure 2). We calculated binding affinities with vina scores less than -10 kcal/mol used for selecting active compounds (Table S1-5). The 2D structure of the 15 compounds that demonstrated significant binding affinity is shown in Figure 3. Among these, out of the compounds tested, 14 compounds displayed robust interaction with SAV1866, with only one compound exhibiting substantial interaction with MecA (Figure 3, Figure 4, Figure 5, Figure 6 and Table S1-7). The specific residues involved in the interactions of the compounds with SAV1866 and MecA are depicted in Figure 6.
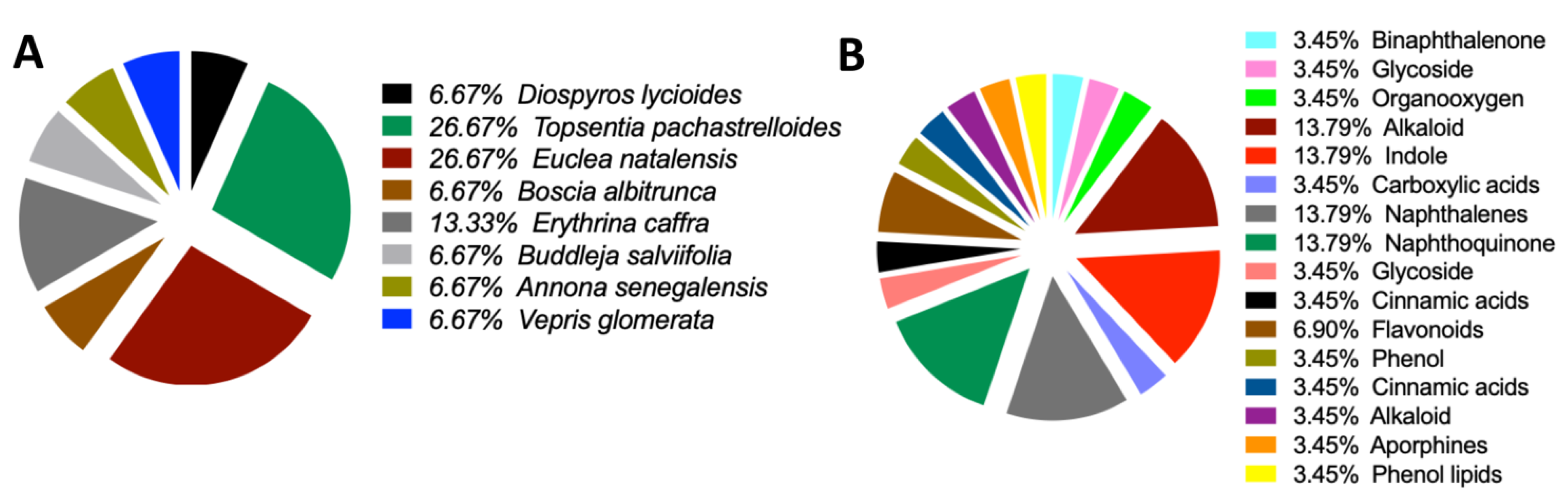
Consequently, none of the 80 compounds docked show strong affinity for MgrA, MepR, and ArlR, and thus were not further investigated because of their high vina score, which was greater than -10 kcal/mol. To provide further insights into the activity of the docked compounds that have the strongest affinity, we determined to explore their plant sources. Our work revealed that the following plant species: Diospyros lycioides, Topsentia pachastrelloides, Euclea natalensis, Boscia albitrunca, Erythrina caffra, Buddleja salviifolia, Annona senegalensis, and Vepris glomerata, were the sources of the identified compounds (Figure 7A and Table S7). Each of these plant species is known to produce compounds that exhibit antibacterial properties (Chukwujekwu, et al., 2011; Kiplimo and Koorbanally, 2012; Lall, et al., 2017; Li, et al., 1998; Pendota, et al., 2015; Van der Kooy, et al., 2006; Zoraghi, et al., 2011). Among the identified plant species, Topsentia pachastrelloides and Euclea natalensis stood out as they had the highest number of compounds with antibacterial activities, with each species contributing four compounds (Figure 7A). The compounds with the highest binding affinities were SANC00524 (1',2-Binaphthalen-4-one-2',3-dimethyl-1,8'-epoxy-1,4',5,5',8,8'-hexahydroxy-8-O-β-glucopyranosyl-5'-O-β-xylopyranosyl(1→6)-β-glucopyranoside), SANC00416 (Cis-3,4-dihydrohamacanthin b), and SANC00436 (Mamegakinone). Their Vina scores were -12.2 kcal/mol, -11.3 kcal/mol, and -11.1 kcal/mol, respectively (Figure 4 and Table S6-7). These strong binding affinities suggest their therapeutic potential. Further analyses showed that most of the compounds identified belonged to various chemical classes, such as alkaloids, indole derivatives, naphthalenes, and naphthoquinones (Figure 7B and Table S7). Previous work has demonstrated that these diverse chemical classes exhibit antibacterial properties against various bacteria and some fungi (Barbieri, et al., 2017; Elmaidomy, et al., 2022; Pollo, et al., 2021; Porras, et al., 2020).
4. Discussion
The current study employed molecular docking techniques to investigate the interaction between natural agents and different bacterial proteins namely SAV1866, MecA, MgrA, MepR.and arlR. Here, we screened a total of 80 natural compounds against major MRSA antibiotic resistance proteins. The results revealed that most of these compounds exhibited high affinity towards the MRSA multidrug ABC transporter and one compound demonstrated strong affinity against MecA. Our current work emphasizes the potential of natural agents as antibacterial therapies and provides valuable insights into their mechanisms of action.
SAV1866 is a multidrug ABC transporter protein that plays a crucial role in expelling antibiotics and toxic substances from bacterial cells. This efflux activity of SAV1866 contributes to multidrug resistance and reduces the effectiveness of antimicrobial treatments (Akhtar and Turner, 2022; Dawson and Locher, 2006; Velamakanni, et al., 2008; Yoshikai, et al., 2016). Compounds that bind strongly to SAV1866 have the potential to inhibit its function and counteract antibiotic efflux (Dashtbani-Roozbehani and Brown, 2021). By targeting SAV1866 with potent inhibitors, it is possible to increase the intracellular concentration of antibiotics, thus restoring their efficacy against resistant bacteria, a process akin to using drug-resistant reversal agents (Akhtar and Turner, 2022; Dawson and Locher, 2006; Marquez, et al., 2005; Mun, et al., 2014; Velamakanni, et al., 2008; Yoshikai, et al., 2016). On the other hand, MecA is a regulatory protein that plays a crucial role in regulating virulence in MRSA (Elal Mus, et al., 2019; Müller, et al., 2015; Wielders, et al., 2002). Targeting MecA provides an opportunity to develop anti-virulence therapies against MRSA, reducing its pathogenicity and enhancing the effectiveness of antibiotic treatments (Kane, et al., 2018; Lade and Kim, 2021; Vestergaard, et al., 2019). This could provide another chemotherapeutic approach to the treatment of MRSA infections (Elal Mus, et al., 2019). Furthermore, compounds that strongly bind to MecA can inhibit its regulatory activities, disrupt the expression of virulence factors, and alleviate the severity of MRSA infections (Elal Mus, et al., 2019; Müller, et al., 2015; Wielders, et al., 2002). The identification of compounds with strong binding to both SAV1866 and MecA highlights the potential of natural compounds in developing novel therapeutic strategies (Lade and Kim, 2021; Vestergaard, et al., 2019).
One of the highly effective natural compounds identified in this study is 1',2-Binaphthalen-4-one-2',3-dimethyl-1,8'-epoxy-1,4',5,5',8,8'-hexahydroxy-8-O-β-glucopyranosyl-5'-O-β-xylopyranosyl(1→6)-β-glucopyranoside (SANC00524). This compound was obtained through the methanol extraction of Diospyros lycioides twigs, and was shown to possess significant antimicrobial activity against S. sanguis and Streptococcus mutans (Li, et al., 1998). Furthermore, we found that several other compounds, namely Cis-3,4-dihydrohamacanthin b (SANC00416), Bromodeoxytopsentin (SANC00415), (Bromotopsentin) SANC00413, and Spongotine A (SANC00414) (Zoraghi, et al., 2011), displayed strong affinity towards the MRSA SAV1866 protein. These Bis-indole Alkaloids were extracted from Topsentia pachastrelloides and have been reported to possess antibacterial properties against MRSA. Additionally, these compounds were observed to disrupt MRSA's cell membranes and inhibit MRSA pyruvate kinase's enzymatic activity (Zoraghi, et al., 2011). In our study, we demonstrated the potent inhibitory effect of these compounds on the MRSA multidrug ABC transporter protein. Similarly, other compounds, including Mamegakinone (SANC00436), Diospyrin (SANC00434), Isodiospyrin (SANC00435), and Neodiospyrin (SANC00438), exhibited strong affinity against the MRSA SAV1866 protein (Van der Kooy, et al., 2006). The Work by Van der Kooy and colleagues demonstrated the inhibitory role of these compounds, derived from the roots of Euclea natalensis, against Mycobacterium tuberculosis (Van der Kooy, et al., 2006). Our current work provides evidence of their robust inhibition of the MRSA multidrug ABC transporter protein. The compounds (Martynoside) SANC01067 and Acteoside (SANC00370), extracted from the leaves of Boscia albitrunca (Pendota, et al., 2015), displayed remarkable affinity towards the MRSA SAV1866 protein. Additionally, Burttinone (SANC00940) and Abyssinone-V 4'-methyl ether (SANC00941), derived from the stem bark of Erythrina caffra Thunb (Chukwujekwu, et al., 2011), exhibited a strong affinity for MRSA SAV1866 protein. These compounds like the others mentioned above, possess potent activity against various Gram-negative and Gram-positive bacteria as well as fungi. We also identified an alkaloid compound, Nornantenine (SANC01101) (Lall, et al., 2017), which strongly binds to MRSA SAV1866. This compound was extracted from the aerial parts of Annona senegalensis and showed efficacy against Streptococcus mutans (Lall, et al., 2017).
Lastly, Limonin (SANC01041), which we demonstrated to have a strong affinity against MRSA MecA but not SAV1866 was isolated from the root and stem bark of V. glomerata (Kiplimo and Koorbanally, 2012). This compound had been shown to possess inhibitory activity against the growth of Staphylococcus aureus and Shigella dysentrieae. These findings highlight the diverse range of natural compounds with potent antimicrobial properties, specifically against MRSA and other bacterial pathogens.
5. Conclusions
We have identified natural compounds that potentially inhibit SAV1866 and MecA proteins of MRSA. Further work is required to understand their mechanisms of action and investigate their chemotherapeutic capacity as antibacterial agents. A need for a better understanding of the specific interactions between these compounds and their target proteins is important in providing valuable insights into their effectiveness against MRSA infections. In addition, conducting preclinical and clinical studies to assess their safety, efficacy, and feasibility as potential antibacterial agents is of the utmost importance. Although identifying these compounds is significant, more research is necessary to uncover their mechanisms of action and evaluate their suitability for future development.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1. Table S1: Binding Affinity of MRSA MecA Protein to 80 Natural Compounds. Table S2: Binding Affinity of MRSA SAV1866 Protein to 80 Natural Compounds. Table S3: Binding Affinity of MRSA mgrA Protein to 80 Natural Compounds. Table S4: Binding Affinity of MRSA MepR Protein to 80 Natural Compounds. Table S5: Binding Affinity of MRSA arlR Protein to 80 Natural Compounds. Table S6: Summary of Binding Affinity of MRSA Antibiotic Resistance Proteins to 80 Natural Compounds. Table S7: Origin, Classification, and Significant Binding Affinity of MRSA SAV1866 and MecA Proteins to 15 Natural Compounds.
Author Contributions
B.O. and M.O.F. contributed significantly to the conceptualization, methodology, formal analysis, and investigation. All authors, B.O. P.M.D and M.O.F., carried out data curation, wrote the original draft, reviewed and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Bingham Start-up Program of Transylvania University to MOF.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data underlying the results are included as part of the published article.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Adelusi, T.I., et al. Molecular modeling in drug discovery. Informatics in Medicine Unlocked 2022:100880.
- Agnihotry, S., et al. Molecular docking and structure-based drug design. Computer-aided drug design 2020:115-131.
- Akhtar, A.A. and Turner, D.P. The role of bacterial ATP-binding cassette (ABC) transporters in pathogenesis and virulence: Therapeutic and vaccine potential. Microbial Pathogenesis 2022;171:105734.
- Álvarez-Martínez, F., et al. Antibacterial plant compounds, extracts and essential oils: An updated review on their effects and putative mechanisms of action. Phytomedicine 2021;90:153626.
- Barbieri, R., et al. Phytochemicals for human disease: An update on plant-derived compounds antibacterial activity. Microbiological research 2017;196:44-68.
- Berman, H.M., et al. The protein data bank. Nucleic acids research 2000;28(1):235-242.
- Birukou, I., et al. The molecular mechanisms of allosteric mutations impairing MepR repressor function in multidrug-resistant strains of Staphylococcus aureus. MBio 2013;4(5):e00528-00513.
- Brown, N.M., et al. Treatment of methicillin-resistant Staphylococcus aureus (MRSA): updated guidelines from the UK. JAC-Antimicrobial Resistance 2021;3(1):dlaa114.
- Burley, S.K., et al. Protein Data Bank (PDB): the single global macromolecular structure archive. Protein crystallography: methods and protocols 2017:627-641.
- Chen, C., et al. TBtools, a toolkit for biologists integrating various biological data handling tools with a user-friendly interface. BioRxiv 2018;289660(10.1101):289660.
- Chen, P.R., et al. An oxidation-sensing mechanism is used by the global regulator MgrA in Staphylococcus aureus. Nature chemical biology 2006;2(11):591-595.
- Chukwujekwu, J., Van Heerden, F. and Van Staden, J. Antibacterial activity of flavonoids from the stem bark of Erythrina caffra thunb. Phytotherapy Research 2011;25(1):46-48.
- Dahiya, P. and Purkayastha, S. Phytochemical screening and antimicrobial activity of some medicinal plants against multi-drug resistant bacteria from clinical isolates. Indian journal of pharmaceutical sciences 2012;74(5):443.
- Dashtbani-Roozbehani, A. and Brown, M.H. Efflux pump mediated antimicrobial resistance by staphylococci in health-related environments: challenges and the quest for inhibition. Antibiotics 2021;10(12):1502.
- Dawson, R.J. and Locher, K.P. Structure of a bacterial multidrug ABC transporter. Nature 2006;443(7108):180-185.
- Diallo, B., et al. SANCDB: an update on South African natural compounds and their readily available analogs. Journal of Cheminformatics 2021;13(1):1-14.
- Dugassa, J. and Shukuri, N. Review on antibiotic resistance and its mechanism of development. Journal of Health, Medicine and Nursing 2017;1(3):1-17.
- Elal Mus, T., et al. Investigation of mecA gene, virulence traits and antibiotic resistance profiles in methicillin-resistant Staphylococcus aureus isolates from dairy products. Journal of Food Safety 2019;39(3):e12620.
- Elmaidomy, A.H., et al. Antimicrobial potentials of natural products against multidrug resistance pathogens: A comprehensive review. RSC advances 2022;12(45):29078-29102.
- English, B.K. and Gaur, A.H. The use and abuse of antibiotics and the development of antibiotic resistance. Hot topics in infection and immunity in children VI 2010:73-82.
- Feig, M. Computational protein structure refinement: almost there, yet still so far to go. Wiley Interdisciplinary Reviews: Computational Molecular Science 2017;7(3):e1307.
- Ferreira, L.G., et al. Molecular docking and structure-based drug design strategies. Molecules 2015;20(7):13384-13421.
- Gore, S., et al. Validation of structures in the Protein Data Bank. Structure 2017;25(12):1916-1927.
- Hatherley, R., et al. SANCDB: a South African natural compound database. Journal of cheminformatics 2015;7:1-9.
- Kane, T.L., Carothers, K.E. and Lee, S.W. Virulence factor targeting of the bacterial pathogen Staphylococcus aureus for vaccine and therapeutics. Current drug targets 2018;19(2):111-127.
- Kiplimo, J.J. and Koorbanally, N.A. Antibacterial activity of an epoxidised prenylated cinnamaldehdye derivative from Vepris glomerata. Phytochemistry Letters 2012;5(3):438-442.
- Kurlenda, J. and Grinholc, M. Alternative therapies in Staphylococcus aureus diseases. Acta Biochimica Polonica 2012;59(2).
- Lade, H. and Kim, J.-S. Bacterial targets of antibiotics in methicillin-resistant Staphylococcus aureus. Antibiotics 2021;10(4):398.
- Lall, N., et al. Alkaloids from aerial parts of Annona senegalensis against Streptococcus mutans. Natural Product Research 2017;31(16):1944-1947.
- Langeveld, W.T., Veldhuizen, E.J. and Burt, S.A. Synergy between essential oil components and antibiotics: a review. Critical reviews in microbiology 2014;40(1):76-94.
- Lee, A.S., et al. Methicillin-resistant Staphylococcus aureus. Nature reviews Disease primers 2018;4(1):1-23.
- Li, X.-C., van der Bijl, P. and Wu, C.D. Binaphthalenone Glycosides from African Chewing Sticks, Diospyros l ycioides. Journal of natural products 1998;61(6):817-820.
- Lim, D. and Strynadka, N.C. Structural basis for the β lactam resistance of PBP2a from methicillin-resistant Staphylococcus aureus. Nature structural biology 2002;9(11):870-876.
- Liu, Y., et al. CB-Dock2: Improved protein–ligand blind docking by integrating cavity detection, docking and homologous template fitting. Nucleic Acids Research 2022;50(W1):W159-W164.
- Lowy, F.D. Antimicrobial resistance: the example of Staphylococcus aureus. The Journal of clinical investigation 2003;111(9):1265-1273.
- Marquez, B., et al. Multidrug resistance reversal agent from Jatropha elliptica. Phytochemistry 2005;66(15):1804-1811.
- Martin, K.W. and Ernst, E. Herbal medicines for treatment of bacterial infections: a review of controlled clinical trials. Journal of Antimicrobial Chemotherapy 2003;51(2):241-246.
- Meng, X.-Y., et al. Molecular docking: a powerful approach for structure-based drug discovery. Current computer-aided drug design 2011;7(2):146-157.
- Morell, E.A. and Balkin, D.M. Methicillin-resistant Staphylococcus aureus: a pervasive pathogen highlights the need for new antimicrobial development. The Yale journal of biology and medicine 2010;83(4):223.
- Müller, S., et al. Poorly cross-linked peptidoglycan in MRSA due to mecA induction activates the inflammasome and exacerbates immunopathology. Cell host & microbe 2015;18(5):604-612.
- Mun, S.-H., et al. Curcumin reverse methicillin resistance in Staphylococcus aureus. Molecules 2014;19(11):18283-18295.
- Nandhini, P., et al. In-Silico molecular screening of natural compounds as a potential therapeutic inhibitor for Methicillin-resistant Staphylococcus aureus inhibition. Chemico-Biological Interactions 2023;374:110383.
- Nascimento, G.G., et al. Antibacterial activity of plant extracts and phytochemicals on antibiotic-resistant bacteria. Brazilian journal of microbiology 2000;31:247-256.
- Otarigho, B. and Falade, M.O. Analysis of antibiotics resistant genes in different strains of Staphylococcus aureus. Bioinformation 2018;14(3):113.
- Otarigho, B. and Falade, M.O. Computational Screening of Approved Drugs for Inhibition of the Antibiotic Resistance Gene mecA in Methicillin-Resistant Staphylococcus aureus (MRSA) Strains. BioTech 2023;12(2):25.
- Ouyang, Z., et al. Deciphering the activation and recognition mechanisms of Staphylococcus aureus response regulator ArlR. Nucleic Acids Research 2019;47(21):11418-11429.
- Pendota, S., Aderogba, M. and Van Staden, J. In vitro antimicrobial activity of extracts and an isolated compound from Boscia albitrunca leaves. South African Journal of Botany 2015;96:91-93.
- Pinzi, L. and Rastelli, G. Molecular docking: shifting paradigms in drug discovery. International journal of molecular sciences 2019;20(18):4331.
- Pollo, L.A., et al. Search for antimicrobial activity among fifty-two natural and synthetic compounds identifies anthraquinone and polyacetylene classes that inhibit Mycobacterium tuberculosis. Frontiers in Microbiology 2021;11:622629.
- Porras, G., et al. Ethnobotany and the role of plant natural products in antibiotic drug discovery. Chemical reviews 2020;121(6):3495-3560.
- Skariyachan, S., et al. Computer aided screening and evaluation of herbal therapeutics against MRSA infections. Bioinformation 2011;7(5):222.
- Turner, N.A., et al. Methicillin-resistant Staphylococcus aureus: an overview of basic and clinical research. Nature Reviews Microbiology 2019;17(4):203-218.
- Van der Kooy, F., Meyer, J. and Lall, N. Antimycobacterial activity and possible mode of action of newly isolated neodiospyrin and other naphthoquinones from Euclea natalensis. South African Journal of Botany 2006;72(3):349-352.
- Velamakanni, S., et al. Multidrug transport by the ABC transporter Sav1866 from Staphylococcus aureus. Biochemistry 2008;47(35):9300-9308.
- Vestergaard, M., Frees, D. and Ingmer, H. Antibiotic resistance and the MRSA problem. Microbiology spectrum 2019;7(2):7.2. 18.
- Wielders, C., et al. mecA gene is widely disseminated in Staphylococcus aureus population. Journal of clinical microbiology 2002;40(11):3970-3975.
- Yao, Y.: University of Cambridge; 2011. Multidrug transport by the ABC transporter Sav1866 from Staphylococcus aureus.
- Yoshikai, H., et al. Multidrug-resistance transporter AbcA secretes Staphylococcus aureus cytolytic toxins. The Journal of Infectious Diseases 2016;213(2):295-304.
- Zoraghi, R., et al. Methicillin-resistant Staphylococcus aureus (MRSA) pyruvate kinase as a target for bis-indole alkaloids with antibacterial activities. Journal of Biological Chemistry 2011;286(52):44716-44725.
Figure 1.
Scientific depiction of the biological and botanical characteristics of natural compounds found within the South African Natural Compounds Database. A: The plant sources that yield the compounds highlighting the diverse botanical sources present in the South African Natural Compounds Database. B: The different types of activity exhibited by the compounds.
Figure 1.
Scientific depiction of the biological and botanical characteristics of natural compounds found within the South African Natural Compounds Database. A: The plant sources that yield the compounds highlighting the diverse botanical sources present in the South African Natural Compounds Database. B: The different types of activity exhibited by the compounds.
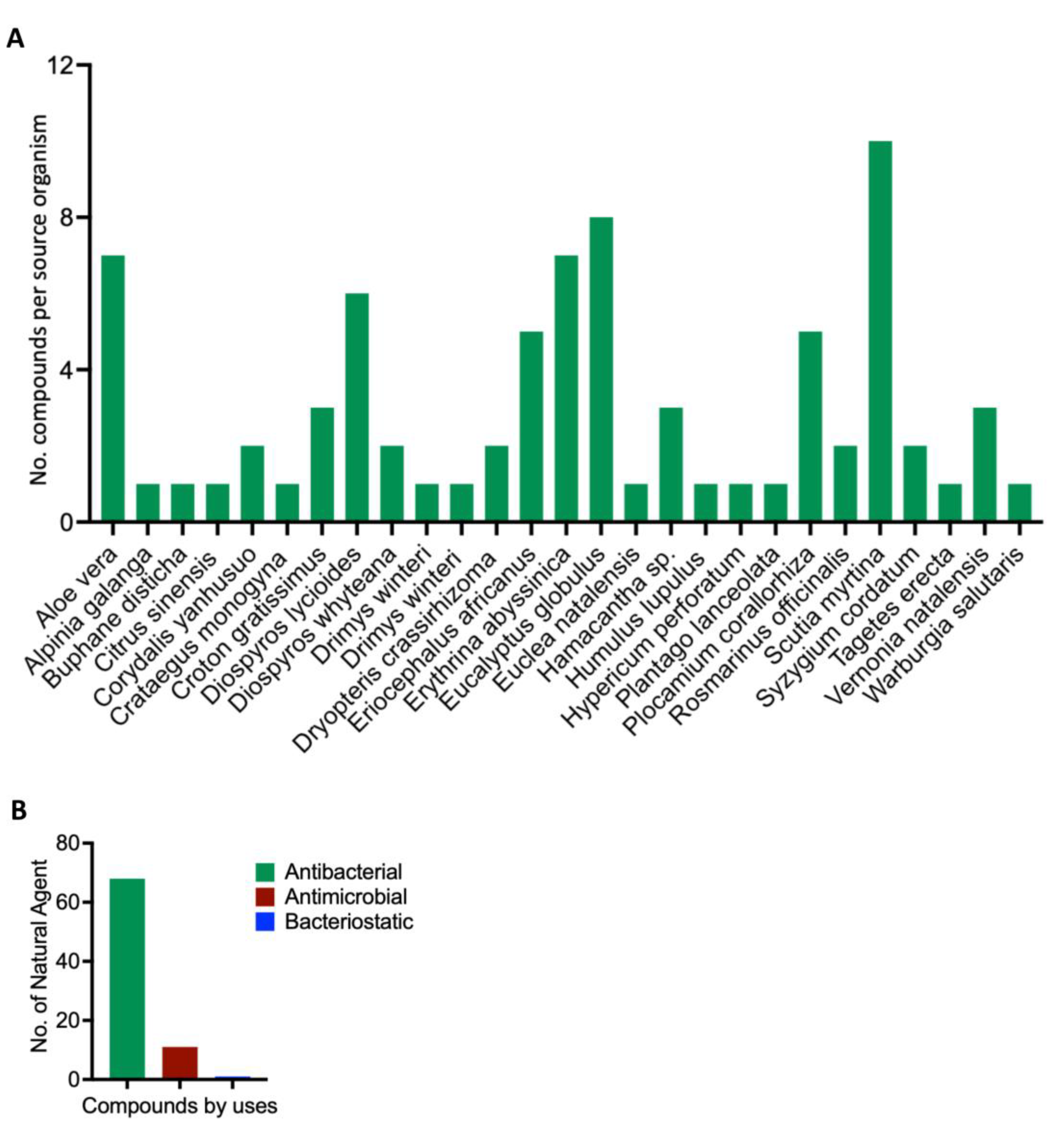
Figure 2.
Three-dimensional configuration of proteins responsible for conferring antibiotic resistance to Methicillin-Resistant Staphylococcus aureus (MRSA). A: Side view of MRSA SAV1866: This view showcases the MRSA SAV1866 protein from a lateral perspective, highlighting its structural chains of transmembrane proteins. B: Dorsal view of an MRSA SAV1866: This view presents MRSA SAV1866 protein from a top-down or dorsal perspective, showing the aperture of the transmembrane proteins. C: Structure of MRSA MgrA displaying the detailed structure of MRSA MgrA protein. D: Structure of MRSA MecA. E: Structure of MRSA MepR. F: Structure of MRSA arIR.
Figure 2.
Three-dimensional configuration of proteins responsible for conferring antibiotic resistance to Methicillin-Resistant Staphylococcus aureus (MRSA). A: Side view of MRSA SAV1866: This view showcases the MRSA SAV1866 protein from a lateral perspective, highlighting its structural chains of transmembrane proteins. B: Dorsal view of an MRSA SAV1866: This view presents MRSA SAV1866 protein from a top-down or dorsal perspective, showing the aperture of the transmembrane proteins. C: Structure of MRSA MgrA displaying the detailed structure of MRSA MgrA protein. D: Structure of MRSA MecA. E: Structure of MRSA MepR. F: Structure of MRSA arIR.
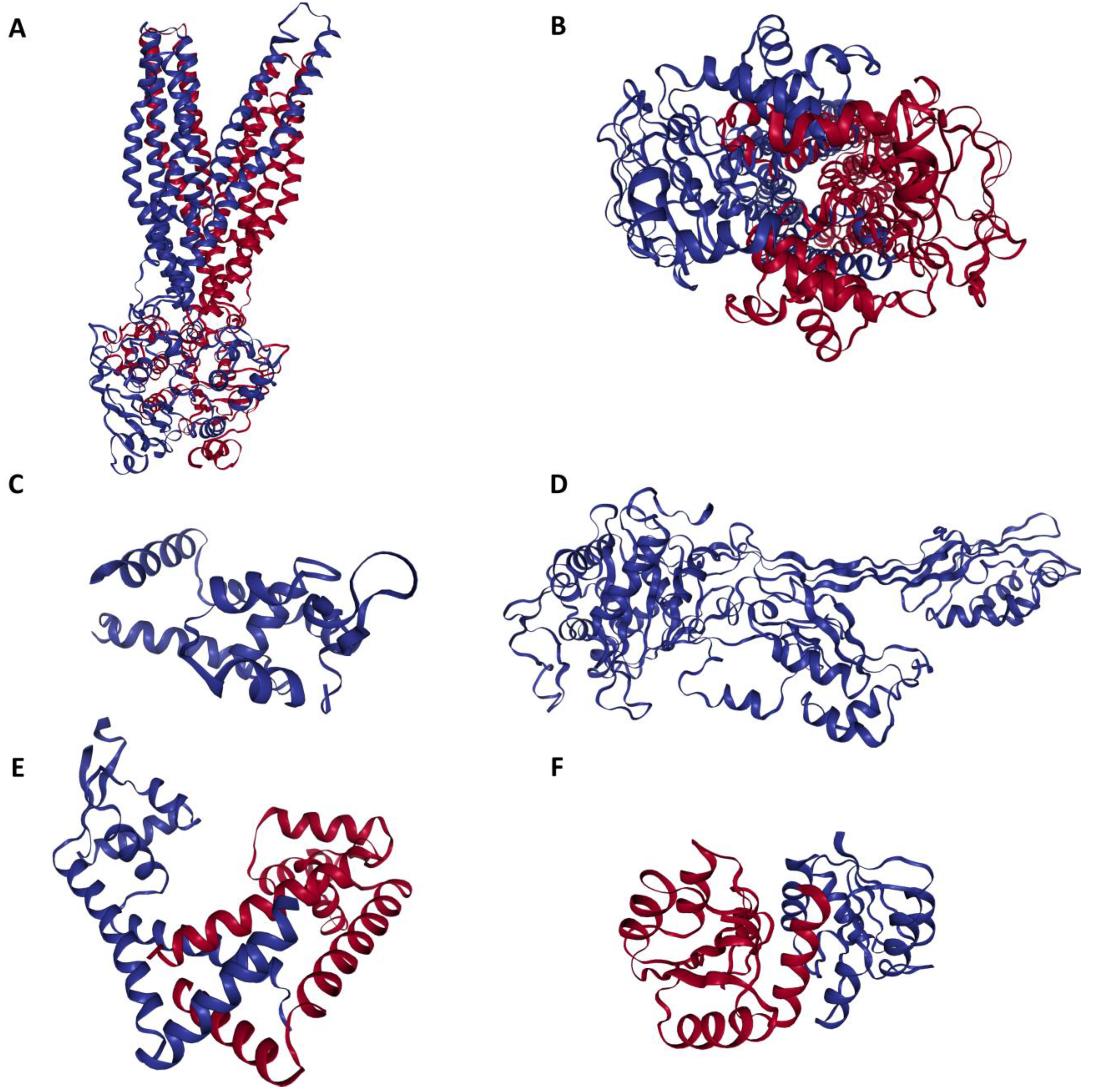
Figure 3.
Structural representation of the natural compounds obtained from the South African Natural Compounds Database that exhibit strong binding affinity to two specific proteins, namely MRSA SAV1866 and MecA, which are associated with Methicillin-Resistant Staphylococcus aureus (MRSA) resistance.
Figure 3.
Structural representation of the natural compounds obtained from the South African Natural Compounds Database that exhibit strong binding affinity to two specific proteins, namely MRSA SAV1866 and MecA, which are associated with Methicillin-Resistant Staphylococcus aureus (MRSA) resistance.
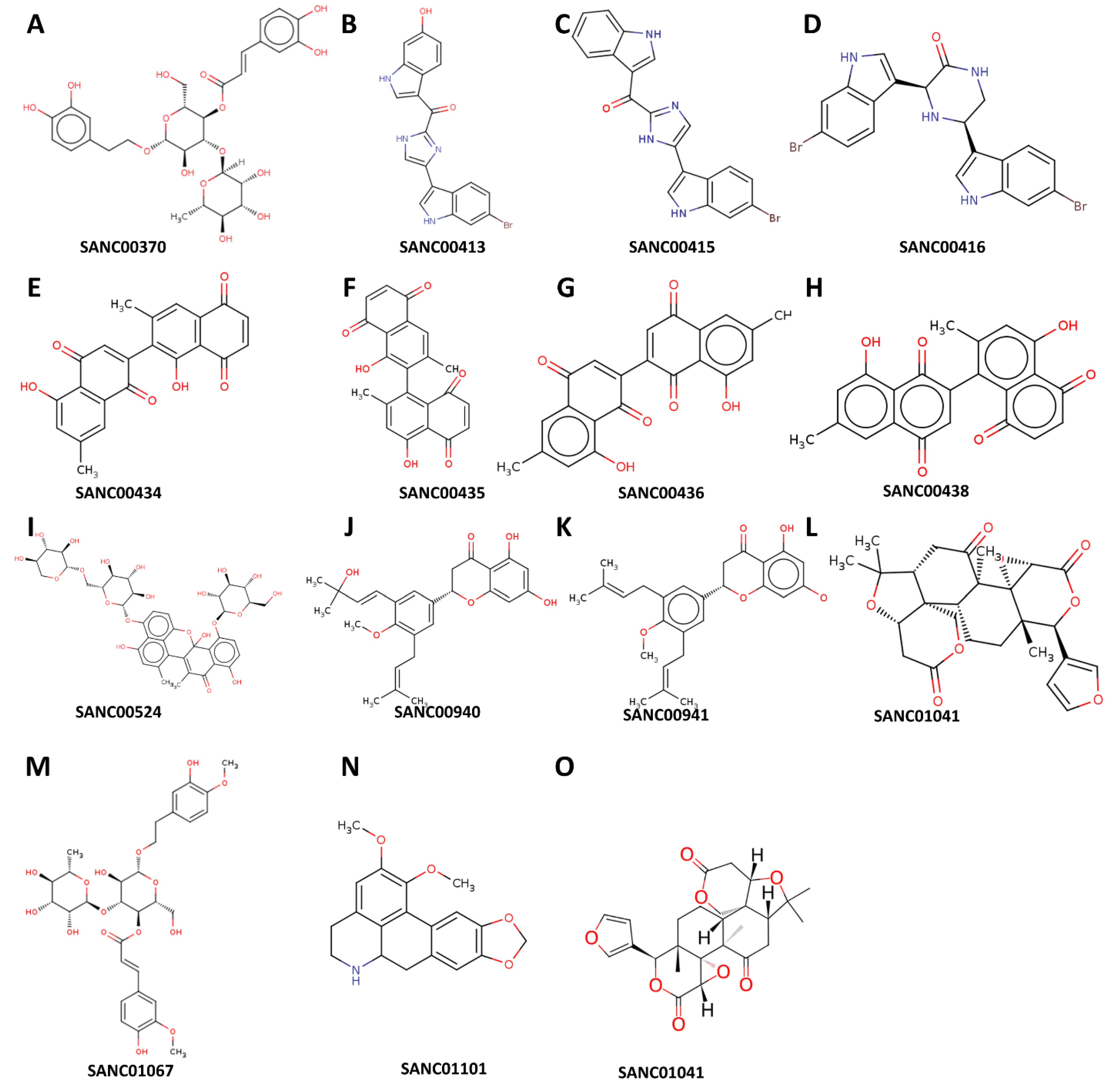
Figure 4.
A comprehensive view of the natural compounds that exhibit robust binding affinity to MRSA SAV1866, MecA, arlR, MgrA, and MepR proteins. A: Inverted bar chart illustrating the bonding of 80 compounds to MRSA SAV1866, MecA, arlR, MgrA, and MepR. B: Inverted bar chart depicting the bonding of 14 compounds to MRSA SAV1866. Since only SAV1866 proteins demonstrate strong bonding with more than one compound. C: Heatmap visualizing the bonding of 80 compounds to MRSA SAV1866, MecA, arlR, MgrA, and MepR. The heatmap employs a color gradient to indicate the strength of the bonding interaction, enabling easy identification of compounds with high or low affinity for each protein. D: Heatmap illustrating the bonding of 15 compounds to MRSA SAV1866, MecA, arlR, MgrA, and MepR: This component specifically focuses on the heatmap representation of the binding affinity of 15 compounds with MRSA SAV1866, MecA, arlR, MgrA, and MepR proteins. The color-coded heatmap visually represents the compounds' bonding strengths with MRSA SAV1866, MecA, arlR, MgrA, and MepR.
Figure 4.
A comprehensive view of the natural compounds that exhibit robust binding affinity to MRSA SAV1866, MecA, arlR, MgrA, and MepR proteins. A: Inverted bar chart illustrating the bonding of 80 compounds to MRSA SAV1866, MecA, arlR, MgrA, and MepR. B: Inverted bar chart depicting the bonding of 14 compounds to MRSA SAV1866. Since only SAV1866 proteins demonstrate strong bonding with more than one compound. C: Heatmap visualizing the bonding of 80 compounds to MRSA SAV1866, MecA, arlR, MgrA, and MepR. The heatmap employs a color gradient to indicate the strength of the bonding interaction, enabling easy identification of compounds with high or low affinity for each protein. D: Heatmap illustrating the bonding of 15 compounds to MRSA SAV1866, MecA, arlR, MgrA, and MepR: This component specifically focuses on the heatmap representation of the binding affinity of 15 compounds with MRSA SAV1866, MecA, arlR, MgrA, and MepR proteins. The color-coded heatmap visually represents the compounds' bonding strengths with MRSA SAV1866, MecA, arlR, MgrA, and MepR.
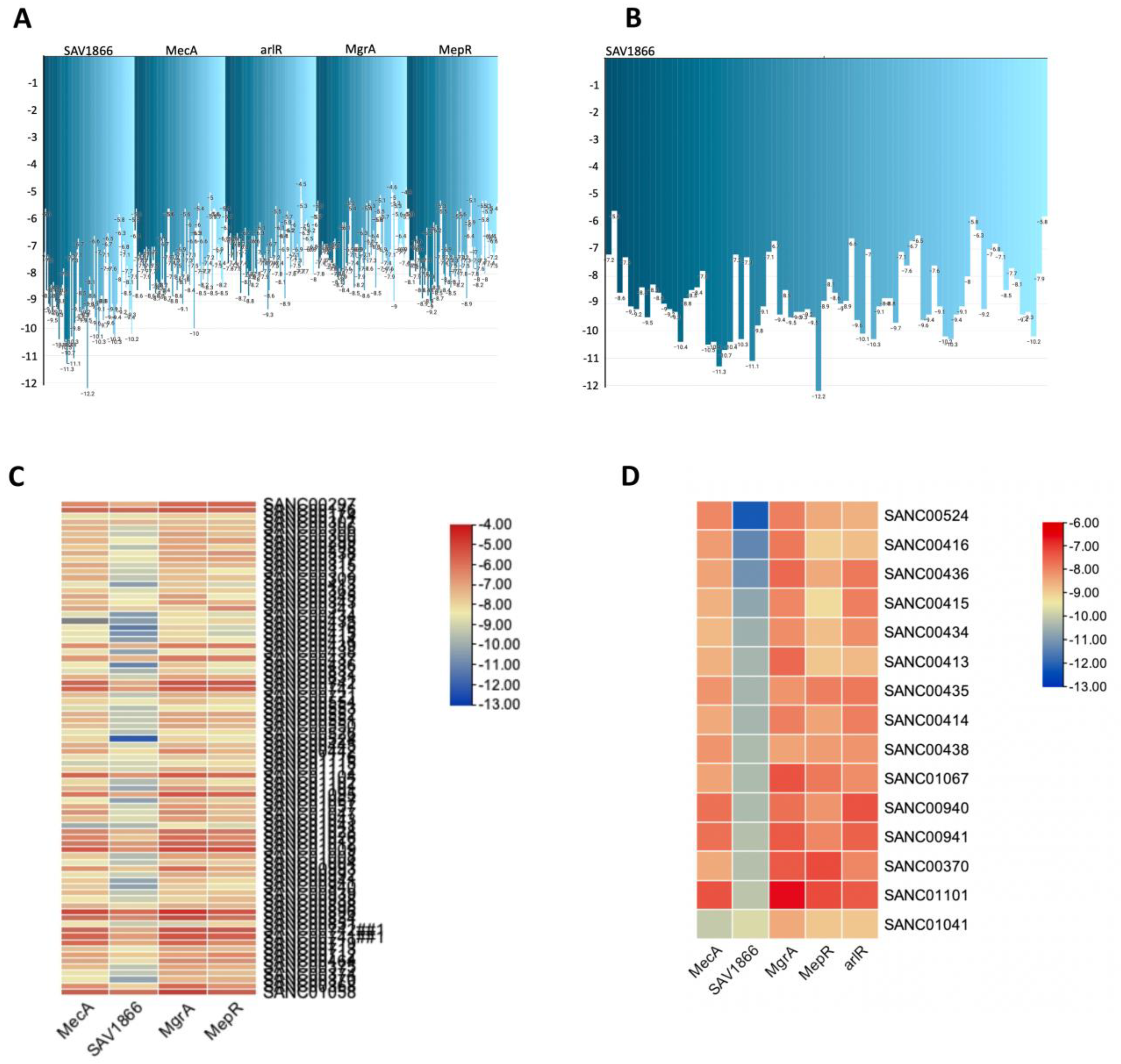
Figure 5.
Three-dimensional binding structure of 14 natural compounds to SAV1866 and the compound SANC01041 (Limonin) to MecA proteins of Methicillin-Resistant Staphylococcus aureus (MRSA).
Figure 5.
Three-dimensional binding structure of 14 natural compounds to SAV1866 and the compound SANC01041 (Limonin) to MecA proteins of Methicillin-Resistant Staphylococcus aureus (MRSA).
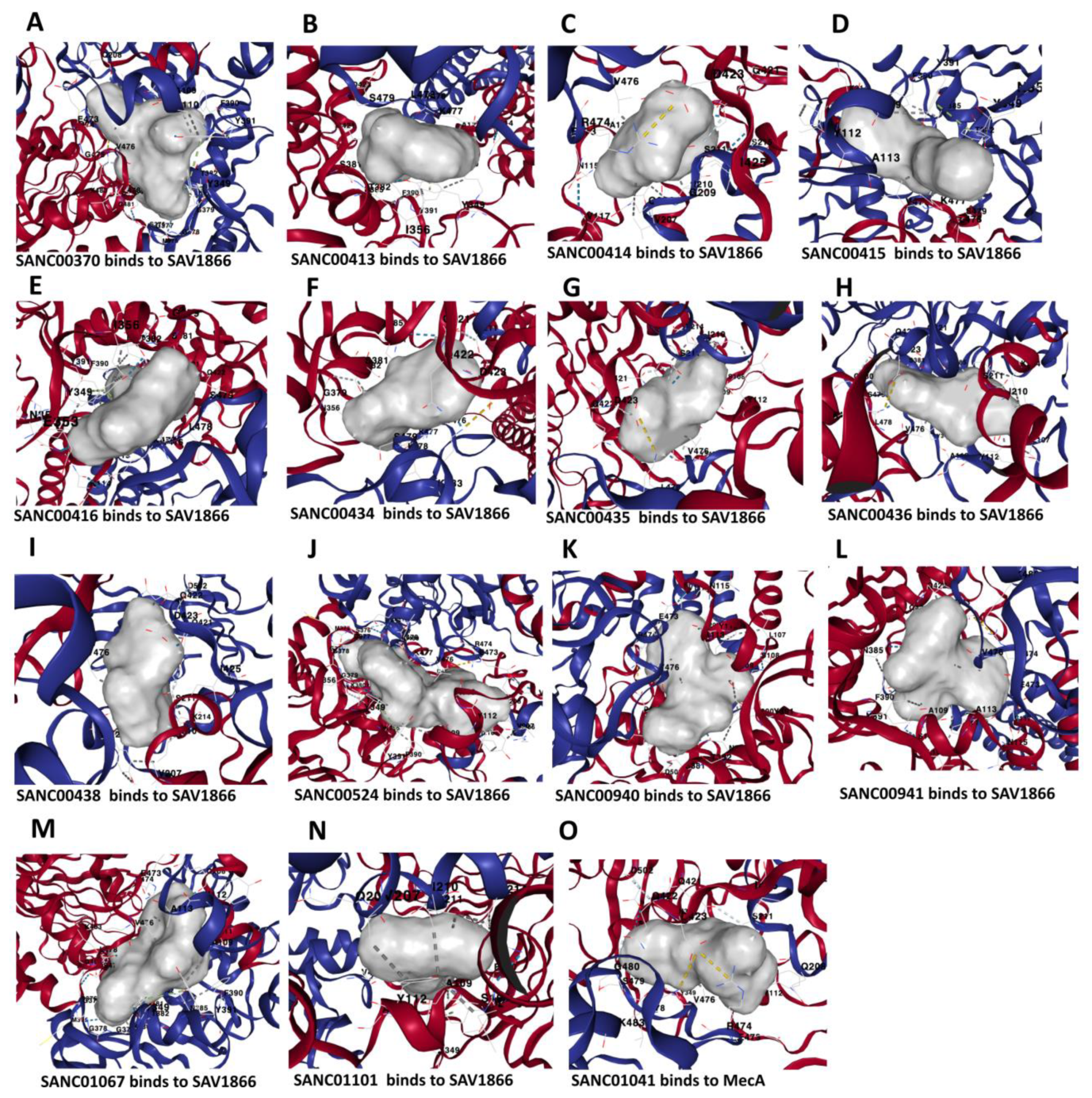
Figure 6.
Interacting residues between the multidrug ATP-binding cassette (ABC) transporter and each of the 14 compounds, as well as the interaction between MecA proteins and a specific natural compound. A: Interacting residues of SAV1866 that bind SANC00370. Interacting residues of Chain A: ALA109, ARG110, TYR112, ALA113, ASN114, PHE347, TYR349, ASN350, SER381, THR382, ASN385, PHE390, TYR391, GLN421, GLN422, and ASP423. Interacting residues of Chain B: VAL207, GLN208, GLY209, ILE210, SER211, GLU473, ARG474, GLY475, VAL476, LYS477, LEU478, SER479, GLY480, and LYS483. B: Interacting residues of SAV1866 that bind SANC00413. Interacting residues of Chain A: ALA109, ALA113, ASN114, TYR349, ASN350, ILE356, SER381, THR382, ASN385, PHE390, TYR391, GLN421, ASP423. Interacting residues of Chain B: SER211, VAL476, LYS477, LEU478, and SER479. C: Interacting residues of SAV1866 that bind SANC00414. Interacting residues of Chain A: SER108, ALA109, TYR112, ALA113, ASN115, VAL117, PHE390, TYR391, GLN421, ASP423, ILE425. Interacting residues of Chain B: VAL207, GLN208, GLY209, ILE210, SER211, LYS214, SER215, GLU473, ARG474, VAL476. D: Interacting residues of SAV1866 that bind SANC00415. Interacting residues of Chain A: SER108, ALA109, TYR112, ALA113, TYR349, THR382, ASN385, PHE390, TYR391, GLN421. Interacting residues of Chain B: ILE210, SER211, LYS214, VAL476, LYS477, LEU478, SER479. E: Interacting residues of SAV1866 that bind SANC00416. Interacting residues of Chain A: ALA109, ARG110, ALA113, ASN114, TYR349, ASN350, GLU353, ILE356, GLY379, SER381, THR382, ASN385, PHE390, TYR391, GLN421. Interacting residues of Chain B: VAL476, LYS477, LEU478, SER479. F: Interacting residues of SAV1866 that bind SANC00434. Interacting residues of Chain A: SER211, GLY475, VAL476, LYS477, LEU478, SER479, LYS483. Interacting residues of Chain B: TYR349, ILE356, GLY377, SER381, THR382, ASN385, TYR391, GLN421, GLN422, ASP423. G: Interacting residues of SAV1866 that bind SANC00436. Interacting residues of Chain A: ILE210, SER211, VAL212, LYS214, SER215, VAL476, LEU478, SER479, GLY480, LYS483. Interacting residues of Chain B: LEU107, SER108, ALA109, TYR112, ALA113, TYR349, SER381, ASN385, PHE390, TYR391, GLN421, GLN422, ASP423. H: Interacting residues of SAV1866 that bind SANC00438. Interacting residues of Chain A: LEU107, SER108, ALA109, TYR112, ALA113, SER381, ASN385, PHE390, TYR391, GLN421, GLN422, ASP423. Interacting residues of Chain B: ILE210, SER211, LYS214, VAL476, LEU478, SER479. I: Interacting residues of SAV1866 that bind SANC00524. Interacting residues of Chain A: ARG206, VAL207, GLN208, GLY209, ILE210, SER211, GLU473, ARG474, VAL476, LYS477, LEU478, SER479, GLY480, GLY481, GLN482, LYS483, and ALA507. Interacting residues of Chain B: ALA109, TYR112, ALA113, VAL117, TYR349, ILE356, GLY374, MET375, SER376, GLY377, GLY378, GLY379, LYS380, SER381, THR382, ASN385, PHE390, TYR391, GLN421, and GLN422. J: Interacting residues of SAV1866 that bind SANC00940. Interacting residues of Chain A: LEU107, SER108, ALA109, TYR112, ALA113, ASN115, VAL117, SER381, THR382, ASN385, PHE390, TYR391, GLN421, GLN422, ASP423 and ASP502. Interacting residues of Chain B: VAL207, GLN208, ILE210, SER211, LYS214, GLU473, ARG474, and VAL476. K: Interacting residues of SAV1866 that bind SANC00941. Interacting residues of Chain A: LEU107, SER108, ALA109, TYR112, ALA113, ASN115, VAL117, ASN385, PHE390, TYR391, GLN421, GLN422, and ASP423. Interacting residues of Chain B: VAL207, GLN208, ILE210, SER211, LYS214, GLU473, ARG474, VAL476, LEU478, and LYS483. L: Interacting residues of SAV1866 that bind SANC00435. Interacting residues of Chain A: SER108, ALA109, TYR112, ALA113, TYR349, ASN385, PHE390, TYR391, GLN421, GLN422, and ASP423. Interacting residues of Chain B: ILE210, SER211, LYS214, VAL476, LYS477, LEU478, SER479, and LYS483. M: Interacting residues of SAV1866 that bind SANC01067. Interacting residues of Chain A: GLN208, SER211, ARG474, GLY475, VAL476, LEU478, SER479, GLY480, GLY481, LYS483, and ALA507. Interacting residues of Chain B: ALA109, TYR112, TYR349, MET375, SER376, GLY377, GLY378, GLY379, LYS380, SER381, THR382, ASN385, PHE390, TYR391, LEU419, GLN421, GLN422, ASP423, ASP502, GLU503, VAL532, and HIS534. N: Interacting residues of SAV1866 that bind SANC01101. Interacting residues of Chain A: LEU107, SER108, ALA109, TYR112, ALA113, TYR349, ASN385, PHE390, TYR391, and GLN421. Interacting residues of Chain B: VAL207, GLN208, ILE210, SER211, LYS214, and VAL476. O: Interacting residues of MecA that bind MecA. Interacting residues of LYS148, SER149, GLU150, ARG151, ASN164, THR165, LYS239, SER240, ARG241, VAL256, GLY257, PRO258, VAL277, HIS293, MET372, and TYR373.
Figure 6.
Interacting residues between the multidrug ATP-binding cassette (ABC) transporter and each of the 14 compounds, as well as the interaction between MecA proteins and a specific natural compound. A: Interacting residues of SAV1866 that bind SANC00370. Interacting residues of Chain A: ALA109, ARG110, TYR112, ALA113, ASN114, PHE347, TYR349, ASN350, SER381, THR382, ASN385, PHE390, TYR391, GLN421, GLN422, and ASP423. Interacting residues of Chain B: VAL207, GLN208, GLY209, ILE210, SER211, GLU473, ARG474, GLY475, VAL476, LYS477, LEU478, SER479, GLY480, and LYS483. B: Interacting residues of SAV1866 that bind SANC00413. Interacting residues of Chain A: ALA109, ALA113, ASN114, TYR349, ASN350, ILE356, SER381, THR382, ASN385, PHE390, TYR391, GLN421, ASP423. Interacting residues of Chain B: SER211, VAL476, LYS477, LEU478, and SER479. C: Interacting residues of SAV1866 that bind SANC00414. Interacting residues of Chain A: SER108, ALA109, TYR112, ALA113, ASN115, VAL117, PHE390, TYR391, GLN421, ASP423, ILE425. Interacting residues of Chain B: VAL207, GLN208, GLY209, ILE210, SER211, LYS214, SER215, GLU473, ARG474, VAL476. D: Interacting residues of SAV1866 that bind SANC00415. Interacting residues of Chain A: SER108, ALA109, TYR112, ALA113, TYR349, THR382, ASN385, PHE390, TYR391, GLN421. Interacting residues of Chain B: ILE210, SER211, LYS214, VAL476, LYS477, LEU478, SER479. E: Interacting residues of SAV1866 that bind SANC00416. Interacting residues of Chain A: ALA109, ARG110, ALA113, ASN114, TYR349, ASN350, GLU353, ILE356, GLY379, SER381, THR382, ASN385, PHE390, TYR391, GLN421. Interacting residues of Chain B: VAL476, LYS477, LEU478, SER479. F: Interacting residues of SAV1866 that bind SANC00434. Interacting residues of Chain A: SER211, GLY475, VAL476, LYS477, LEU478, SER479, LYS483. Interacting residues of Chain B: TYR349, ILE356, GLY377, SER381, THR382, ASN385, TYR391, GLN421, GLN422, ASP423. G: Interacting residues of SAV1866 that bind SANC00436. Interacting residues of Chain A: ILE210, SER211, VAL212, LYS214, SER215, VAL476, LEU478, SER479, GLY480, LYS483. Interacting residues of Chain B: LEU107, SER108, ALA109, TYR112, ALA113, TYR349, SER381, ASN385, PHE390, TYR391, GLN421, GLN422, ASP423. H: Interacting residues of SAV1866 that bind SANC00438. Interacting residues of Chain A: LEU107, SER108, ALA109, TYR112, ALA113, SER381, ASN385, PHE390, TYR391, GLN421, GLN422, ASP423. Interacting residues of Chain B: ILE210, SER211, LYS214, VAL476, LEU478, SER479. I: Interacting residues of SAV1866 that bind SANC00524. Interacting residues of Chain A: ARG206, VAL207, GLN208, GLY209, ILE210, SER211, GLU473, ARG474, VAL476, LYS477, LEU478, SER479, GLY480, GLY481, GLN482, LYS483, and ALA507. Interacting residues of Chain B: ALA109, TYR112, ALA113, VAL117, TYR349, ILE356, GLY374, MET375, SER376, GLY377, GLY378, GLY379, LYS380, SER381, THR382, ASN385, PHE390, TYR391, GLN421, and GLN422. J: Interacting residues of SAV1866 that bind SANC00940. Interacting residues of Chain A: LEU107, SER108, ALA109, TYR112, ALA113, ASN115, VAL117, SER381, THR382, ASN385, PHE390, TYR391, GLN421, GLN422, ASP423 and ASP502. Interacting residues of Chain B: VAL207, GLN208, ILE210, SER211, LYS214, GLU473, ARG474, and VAL476. K: Interacting residues of SAV1866 that bind SANC00941. Interacting residues of Chain A: LEU107, SER108, ALA109, TYR112, ALA113, ASN115, VAL117, ASN385, PHE390, TYR391, GLN421, GLN422, and ASP423. Interacting residues of Chain B: VAL207, GLN208, ILE210, SER211, LYS214, GLU473, ARG474, VAL476, LEU478, and LYS483. L: Interacting residues of SAV1866 that bind SANC00435. Interacting residues of Chain A: SER108, ALA109, TYR112, ALA113, TYR349, ASN385, PHE390, TYR391, GLN421, GLN422, and ASP423. Interacting residues of Chain B: ILE210, SER211, LYS214, VAL476, LYS477, LEU478, SER479, and LYS483. M: Interacting residues of SAV1866 that bind SANC01067. Interacting residues of Chain A: GLN208, SER211, ARG474, GLY475, VAL476, LEU478, SER479, GLY480, GLY481, LYS483, and ALA507. Interacting residues of Chain B: ALA109, TYR112, TYR349, MET375, SER376, GLY377, GLY378, GLY379, LYS380, SER381, THR382, ASN385, PHE390, TYR391, LEU419, GLN421, GLN422, ASP423, ASP502, GLU503, VAL532, and HIS534. N: Interacting residues of SAV1866 that bind SANC01101. Interacting residues of Chain A: LEU107, SER108, ALA109, TYR112, ALA113, TYR349, ASN385, PHE390, TYR391, and GLN421. Interacting residues of Chain B: VAL207, GLN208, ILE210, SER211, LYS214, and VAL476. O: Interacting residues of MecA that bind MecA. Interacting residues of LYS148, SER149, GLU150, ARG151, ASN164, THR165, LYS239, SER240, ARG241, VAL256, GLY257, PRO258, VAL277, HIS293, MET372, and TYR373.

Figure 7.
Origin Sources of the compounds that demonstrate strong binding to the MRSA antibiotics resistance protein. A: Sources of the compounds that strongly bind to MRSA multidrug ATP-binding cassette (ABC) and MecA. B: Classification of the compounds that strongly bind to MRSA multidrug ATP-binding cassette (ABC) and MecA.
Figure 7.
Origin Sources of the compounds that demonstrate strong binding to the MRSA antibiotics resistance protein. A: Sources of the compounds that strongly bind to MRSA multidrug ATP-binding cassette (ABC) and MecA. B: Classification of the compounds that strongly bind to MRSA multidrug ATP-binding cassette (ABC) and MecA.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



