
Submitted:
14 March 2025
Posted:
17 March 2025
You are already at the latest version
Abstract
Background/Objectives: Antibiotic therapy faces challenges due to rising acquired and biofilm-related antibiotic resistance rates. High resistance levels to commonly used antibiotics have been observed in methicillin-resistant Staphylococcus aureus (MRSA) strains among cystic fibrosis (CF) patients, indicating an urgent need for new antibacterial agents. For the first time, this study aimed to identify potential novel therapeutics with antibacterial and antibiofilm activities against an MRSA CF strain by screening a drug repurposing library. Methods/Results: Among the 3,386 compounds, high throughput screening identified 2,439 (72%), 654 (19.3%), and 426 (12.6%) drugs active against planktonic cells, biofilm formation, and preformed biofilm, respectively, although at different extents. The most active hits were 193 (5.7%) against planktonic cells, causing a 100% growth inhibition; 5 (0.14%) with excellent activity against biofilm formation (i.e., reduction ≥ 90%); and 4 showing high activity (i.e., 60% ≤ biofilm reduction < 90%) against preformed biofilms. The potential hits belonged to several primary research areas, with “cancer” as the most prevalent. After performing a literature review to identify other, already published, biological properties that could be relevant to the CF lung environment (i.e., activity against other CF pathogens, anti-inflammatory and anti-virulence potential), the most interesting hits were: 5-(N,N-Hexamethylene)-amiloride (diuretic), Toremifene (anticancer), and Zafirlukast (antiasthmatic), Fenretide (anticancer), and Montelukast (antiasthmatic) against planktonic S. aureus cells; Hemin against biofilm formation; and Heparin, Clemastine (antihistaminic), and Bromfenac (nonsteroidal anti-inflammatory) against established biofilms. Conclusions: These findings warrant further in vitro and in vivo studies to confirm the potential of repurposing these compounds for managing lung infections caused by S. aureus in CF patients.
Keywords:
High-throughput screening
; Staphylococcus aureus
; Cystic fibrosis
; Antibacterial
; Antibiofilm
1. Introduction
Staphylococcus aureus is the most prevalent bacterium isolated from the sputum of cystic fibrosis (CF) patients during their first decade of life. The incidence of methicillin-sensitive S. aureus (MSSA) is significantly lower than that of methicillin-resistant S. aureus (MRSA) in most CF patients (Craft et al., 2019). Chronic lung infections caused by methicillin-resistant S. aureus (MRSA) strains lead to worse clinical outcomes, including a more rapid decline in lung function (measured as forced expiratory volume in 1 second, FEV1) and increased mortality rates (Dasenbrook et al., 2008; Dasenbrook et al., 2010).
Antibiotic therapy presents challenges due to the rising rates of antibiotic resistance, particularly among biofilm-related infections (Craft et al., 2019). MRSA strains show high resistance to commonly used antibiotics in CF patients, such as ciprofloxacin, clindamycin, erythromycin, and gentamicin. This underscores the pressing need for new antibacterial agents (Xu et al., 2024).
Traditional drug discovery processes are often lengthy, labor-intensive, and expensive. In contrast, drug repurposing offers a more dynamic, cost-effective, and feasible approach to quickly addressing the declining drug discovery pipeline. This strategy involves exploring new applications for already approved pharmaceuticals, thus reducing the time required for drug development, lowering costs, and minimizing the inherent risks of drug innovation (Farha and Brown, 2019).
This study aimed to identify potential novel therapeutics against S. aureus by screening a drug repurposing library for antibacterial and antibiofilm activities for the first time.
2. Results
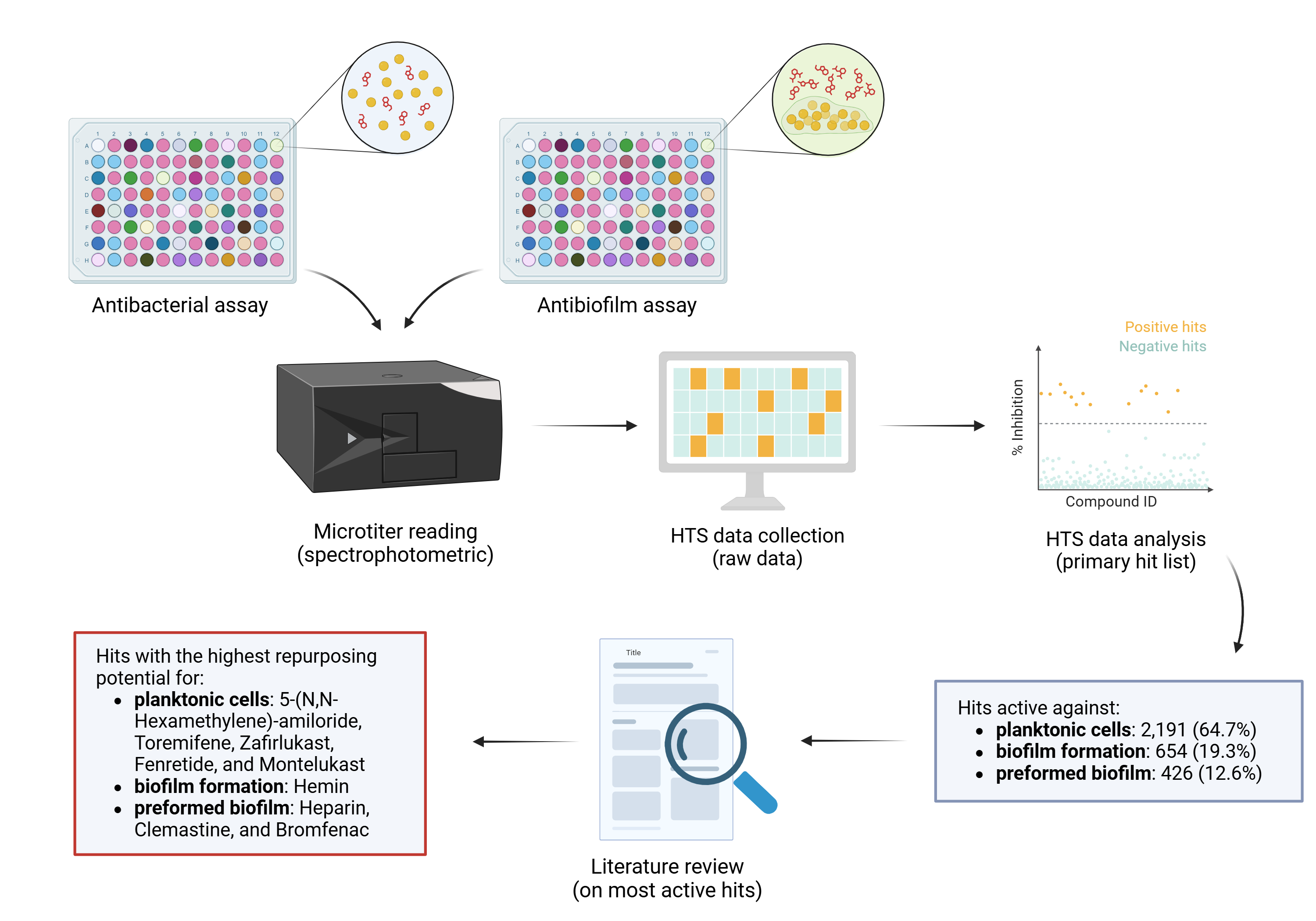
In the present study, we screened the Drug Repurposing Compound Library (MedChem Express) to identify hits with antibacterial and anti-biofilm activity toward S. aureus. The library consisted of 3,386 bioactive compounds (2,342 already launched; 1,044 have reached clinical trial stages in the USA: 1 drug in phase I, 606 drugs in phase II, 372 drugs in phase III, and 65 drugs in phase IV) (Figure 1A) used as drugs with several therapeutic indications, including cancer, neurodegenerative, infectious, and cardiovascular diseases (Figure 1B).
2.1. HTS Assay Validation
The screening quality was evaluated using the Z-factor, a standard measure of an HTS’s robustness and feasibility. The Z-factor estimates the magnitude of the difference between the positive and negative controls relative to the sum of the respective standard deviations.
The average Z-factor between the negative and positive control in the 96-well test plates was 0.675 (range: 0.500-0.843), well above 0.500 value, indicating the assay could reliably separate positive and negative controls (Figure 2). In addition to the Z-factor, also the coefficient of variation (< 10%) and the signal-to-background ratio (> 10-fold) further supported the feasibility of our anti-S. aureus drug screening assay.
2.2. Identification of Hits Inhibiting S. aureus Growth
We identified compounds active against the methicillin-resistant S. aureus Sa2 strain through a 96-well microtiter plate-based high-throughput screening (HTS) of 3,386 compounds belonging to the compound library. All compounds were initially tested at a single concentration of 0.1 mM to pinpoint active hits. The results were expressed as the percentage of inhibition of bacterial growth compared to that in untreated controls, estimated from spectrophotometric readings carried out on supernatant.
This primary screen initially identified 2,439 out of 3,386 (72%) compounds able to affect S. aureus Sa2 growth. Among those, 248 hits had “bacterial” as their primary target and were excluded since this work aimed to find new repositionable drugs. Most (n=146) of these excluded compounds showed high (i.e., 60% ≤ growth reduction < 90%) or excellent activity (i.e., growth reduction ≥ 90%), therefore confirming the robustness of HTS.
The remaining 2,191 (64.7%) hits showed a potential for repurposing against S. aureus, although at different extents: 617 (18.2%) with low activity (i.e., 10% < growth reduction < 25%); 809 (23.9%) with moderate activity (i.e., 25% ≤ growth reduction < 60%); 350 (10.3%) with high activity (i.e., 60% ≤ growth reduction < 90%); and, more interestingly, 415 (12.2%) compounds with excellent activity (i.e., growth reduction ≥ 90%).
The 415 hits showing excellent activity based on spectrophotometric readings were re-assessed by cell viable count, confirming the efficacy of 364 (10.7%) compounds, listed in Table S1 (in the Supplementary Material) and graphed in Figure 3. Among those, it is worth noting that 193 hits caused a 100% inhibition rate Based on their known pharmacological profiles, the hits were grouped into classes based on the primary research area (some compounds belonged to multiple areas). Most of the hits belonged to “cancer” (83 out of 193; 43.0%) as a primary research area, followed by “infection” (other than bacterial) (31; 16.1%), “neurological disease” (28; 14.5%), “inflammation” (17; 8.8%), “metabolic disease” and “cardiovascular disease” (12 each; 6.2%), “endocrinology” and "others” (5 each; 2.6%) (Figure 3). Dunnett's multiple comparison tests found no statistically significant differences in antibacterial activity among research areas.
2.3. Identification of Hits Active Against S. aureus Biofilm Formation
The compound library also underwent a 96-well microtiter plate-based HTS to identify hits that could affect biofilm formation by the MRSA S. aureus Sa2 strain. The results were expressed as the percentage of inhibition of biofilm biomass formation compared to that in untreated controls, estimated from spectrophotometric readings after a crystal violet assay. The compounds significantly affecting biofilm formation, with no known antibacterial activity and inactive against S. aureus Sa2 (i.e., growth inhibition ≤ 10%), were considered potential antibiofilm hits.
HTS revealed 654 out of 3,386 (19.3%) compounds can reduce biofilm formation by S. aureus Sa2, although at different extents: 128 out of 654 (19.6%) with low activity (i.e., 10% < biofilm reduction < 25%), 344 (52.6%) with moderate activity (i.e., 25% ≤ biofilm reduction < 60%); 168 (25.7%) with high activity (i.e., 60% ≤ biofilm reduction < 90%); and 14 (2.1%) compounds with excellent activity (i.e., biofilm reduction ≥ 90%). Overall, hits belonged to several primary research areas, with “cancer” as the most prevalent, as resumed in Figure 4. Dunnett's multiple comparison tests found no statistically significant differences in activity against biofilm formation among research areas.
The fourteen hits showing excellent activity were retested in a secondary screen - carried out in two independent experiments, each in triplicate - confirming an antibiofilm potential for five compounds (Table S2 in the Supplementary Material). These repositionable hit candidates comprised three anticancer agents (Tipifarnib, Olaparib, and Acefylline), Hemin (“cardiovascular disease” research area), and TMC647055 (Choline salt) (“infection” research area).
2.4. Identification of Hits Active Against Preformed Biofilm by S. aureus
Each of the 3,386 compounds was tested at a single concentration of 0.1 mM to pinpoint hits effective towards preformed, 24 h-old biofilm formed by the MRSA S. aureus Sa2 strain. The results were expressed as the percentage of biofilm biomass dispersion compared to that in untreated controls, estimated from spectrophotometric readings after a crystal violet assay. The compounds with significant activity against mature biofilms, with no known antibacterial activity and not active against S. aureus Sa2, were considered potential antibiofilm hits.
The primary screen identified 426 out of 3,386 (12.6%) compounds with the potential for repurposing against mature biofilms by S. aureus Sa2, although at different extents. Most of the hits showed low (i.e., 10% < biofilm reduction < 25%) (216 out of 426; 56.1%) or moderate (i.e., 25% ≤ biofilm reduction < 60%) (202, 40.7%) activity, whereas only 8 (3.1%) showed high activity (i.e., 60% ≤ biofilm reduction < 90%). No compound was able to disperse at least 90% of biofilm biomass.
Overall, hits belonged to several primary research areas, although those with low and moderate activity mainly belonged to “neurological disease” and “cancer” areas (Figure 5). Dunnett's multiple comparison tests found no statistically significant differences in activity against mature biofilm among research areas.
The eight hits showing high activity in the primary screening were re-assessed in a secondary screen - carried out in two independent experiments, each in triplicate - confirming 4 repositionable hit candidates, comprising Flumatinib (mesylate) with anticancer activity, the anti-inflammatory Bromfenac (sodium hydrate), Clemastine (fumarate) (“neurological disease”), and Heparin (Lithium salt) (Table S3 in the Supplementary Material).
3. Discussion
In the present study, we screened the Drug Repurposing Compound Library (MedChem Express) to identify hits with relevant potential for antibacterial and antibiofilm activities against an S. aureus strain causing a long-term pulmonary infection in a CF patient.
The library we tested in the present study consists of 3,386 bioactive compounds, most already launched and used as drugs with several therapeutic indications, including cancer, neurodegenerative, infectious, and cardiovascular diseases. The library structure could explain the surprisingly high hit rates we observed during the first HTS: 72% (2,440 out of 3,386) for antibacterial activity, 19.3% (654 out of 3,386) for activity against biofilm formation, and 12.6% (426 out of 3,386) for activity against preformed biofilm. Indeed, less than 0.1% hit rates were reported in other HTS studies evaluating large, random chemical libraries of small synthetic molecules that commonly contain numerous non-drug-like molecules (Nilakantan et al., 2002; Langer et al., 2009). The calculation of the Z-factor, the coefficient of variation, and the signal-to-background ratio validated the robustness and feasibility of our anti-S. aureus drug HTS assay.
An HTS was initially performed to identify compounds active against planktonic cells of S. aureus Sa2 strain. Among the 3,386 compounds screened, 2,191 compounds - with a primary target different from “bacterial” - caused a decrease in Sa2 strain growth, although with different effectiveness. We focused on selected 193 compounds with the highest potential for repurposing since they exhibited maximum activity (i.e., 100% growth reduction), as confirmed by viable cell counts.
A revision of the scientific literature indicated that 135 out of 193 compounds were never reported active against S. aureus in previous studies and could be further investigated for repurposing potential. Trying to individuate drugs with a high repurposing potential relevant to the CF lung infection, the selected 135 potential hits underwent a literature review to reveal other biological effects critical to the interplay between the host and pathogens that shape the course of the disease, i.e., activity against other CF pathogens (e.g., Pseudomonas aeruginosa, Burkholderia cepacia, Haemophilus influenzae), anti-inflammatory and anti-virulence potential (Bhagirath et al., 2016). In this way, 37 compounds were identified and are listed in Table 1.
Here, we reported an antibacterial activity for 31 out of 37 selected compounds for the first time. Previous studies referred to the activity of the remaining six compounds against species other than S. aureus: the anticancer Toremifene and the antiasthmatic Zafirlukast vs. Porphyromonas gingivalis (Geritis et al., 2017a; Geritis et al., 2017b), Zafirlukast, the anticancer Napabucasin and the antipsychotic Perphenazine vs. oral streptococci (Mortensen et al., 1992; Geritis et al., 2017b; Kuang et al., 2020), Zafirlukast and the anticancer Linsitinib vs. Mycobacterium tuberculosis (Pinault et al., 2013; Wang et al., 2022), and Perphenazine vs. Neisseria meningitidis, Enterobacteriaceae, and Listeria monocytogenes (Mortensen et al., 1992).
Opportunistic polymicrobial bacterial airway infection is a hallmark of CF lung disease and early mortality (VanDevanter et al., 2024). From birth, the airways of individuals with CF are susceptible to infections from microbial opportunists. Over time, more concerning bacterial species often appear in respiratory secretions throughout their shortened lifetimes. The failure of bacterial clearance generates a dominant chronic inflammation, resulting in a toxic pro-inflammatory local microenvironment that damages the lung and the innate immunity, further facilitating infections and thus resulting in the predominant cause of morbidity and mortality in CF patients. In this frame, it is worth noting the previously reported activity of the diuretic 5-(N,N-Hexamethylene)-amiloride and Perphenazine, respectively, against Pseudomonas cepacia (Cohn and Rudzienski, 1994) and Haemophilus influenzae (Mortensen et al., 1992), since these species have the potential to cause pulmonary exacerbations and lung function decline in people suffering from CF (LiPuma, 2010; Gutiérrez Santana et al., 2024).
Others recent evidence suggest that viridans group streptococci (VGS) may play a crucial role in maintaining a stable microbial ecology within the CF lung, enhancing virulence associated with polymicrobial interactions and directly contributing to the pathology of the lung. Streptococcus pneumoniae has recently been associated with an increased severe decline in FEV1 (Paganin et al., 2015). In addition, VGS may have the ability to aggregate with other important CF bacterial pathogens, such as S. aureus and P. aeruginosa, promoting the initial attachment and eventual colonization with these pathogens and the development of multispecies biofilms in the CF airways (Rickard et al., 2003). In this regard, the previously reported activity of Perphenazine against S. pneumoniae (Mortensen et al., 1992) and Toremifene against Streptococcus mutans (Geritis et al., 2017a) adds value to their potential for repurposing, warranting further studies.
The prolonged use of antibiotics has been essential in improving the survival rates of CF patients. However, this approach leads to complications, such as the rise of adaptative antibiotic resistance, adverse effects, and allergic reactions. An attractive alternative recently explored for treating bacterial infections is targeting or reducing the production of virulence factors, such as inhibiting quorum sensing, biofilm formation, iron uptake, and efflux pumps (Carullo et al., 2023; Mudgil et al., 2024). Conversely to antimicrobial therapies, this strategy does not affect bacterial growth and is less likely to lead to resistance, exerting low selective pressure. In this frame, five drugs among the hits we identified in the present study have previously been shown to exhibit anti-virulence potential against S. aureus and P. aeruginosa, increasing their repurposing potential in CF patients (Table 1). S. aureus hemolysis activity has been observed to be significantly affected in over 200 clinical isolates by the antihistamine compound Loratadine (Zheng et al., 2022) and by the antiparasitic Miltefosine in the invertebrate Galleria mellonella and a murine model of pneumonia (Koch et al., 2017). The virulence potential of S. aureus is also decreased by the synthetic estrogen Diethylstilbestrol - causing reduction of alpha-toxin, coagulase, deoxyribonuclease, and penicillinase production (Yotis et al., 1977) – and Loratadine, affecting biofilm formation, pigmentation (Zheng et al., 2022) and exotoxin production (Balogh et al., 2024). In a rat subcutaneous catheter model, biofilm formation was also inhibited by the anticancer Toremifene (De Cremer et al., 2014). Miltefosine and the anti-hypercholesterolemic agent Lovastatin showed anti-virulence potential in P. aeruginosa, respectively, inhibiting phospholipase C/sphingomyelinase with a protective effect against murine pneumonia (Wargo et al., 2011) and decreasing swarming motility (Hennessy et al., 2013). Other studies revealed synergistic activity of Toremifene and the anticancer Mitotane combined with polymyxin B against multidrug-resistant P. aeruginosa (Hussein et al., 2017; Tran et al., 2018) and the percutaneous enhancer Laurocapram combined with cephalosporins against MRSA (Wang et al., 2024).
The main hallmark of CF pathophysiology is excessive inflammation and the inability to resolve lung infections, contributing to morbidity and, eventually, mortality. Therefore, anti-inflammatory properties could be desirable to improve the repurposing potential of the hits found in the present study. Most of the 37 selected hit compounds were reported for anti-inflammatory potential in the literature, although in clinical settings or models not pertinent for infective diseases (e.g., inflammatory bowel disease (Verhaar et al., 2013), allergic dermatitis (Heo et al., 2020), pulmonary arterial hypertension (Geerdink et al., 2017), osteoarthritis (Aizman et al., 2014; Hu et al., 2016), asthma (Towsend et al., 2023), Parkinson's disease (Tian et al., 2022), cigarette smoke (Yang et al., 2021), vascular diseases (fan et al., 2024), thyroid eye disease (Gulbins et al., 2023), and amyotrophic lateral sclerosis (Ketabforoush et al., 2023) (Table 1). Conversely, a few hits have previously been reported for anti-inflammatory potential in models that could be relevant to CF lung infections, thus increasing their potential for repurposing: 5-(N,N-Hexamethylene)-amiloride in lipopolysaccharide (LPS)-exposed alveolar epithelial cells (Haddad and Land, 2002), Zafirlukast in LPS-exposed mice (Xue et al., 2024), GW 501516 (Cardarine) – aromatic ether under investigation for lipid metabolism’s disorder (phase 2) – and the anticancer Ricolinostat protect against LPS-activated macrophages inflammation (; Zhang et al., 2019; Lim and Kwak, 2024), Bardoxololone - semisynthetic triterpenoids under investigation for lymphoma - Omaveloxolone - a potent anti-inflammatory in treating Friedreich's ataxia – and Verteporfin – used as a photosensitizer in photodynamic therapy – alleviate LPS-triggered acute lung injury in mice (Pei et al., 2019; Wu et al., 2024; Cen et al., 2024), and Vandertanib – approved for unresectable and disseminated diseases - in SARS-CoV2-infected mice (Puhl et al., 2022) (Table 1). Interestingly, the literature already reported anti-inflammatory properties in CF patients or models for three hits. Specifically, Fenretide increases IL-1β expression in Cftr-knockout mice, improving their ability to combat P. aeruginosa lung infection (Guilbalt et al., 2008); in addition, it normalizes the fatty acid imbalance by reducing arachidonic acid and increasing docosahexaenoic acid in CF patients (Guilbault et al., 2009). Zafirlukast, a leukotriene receptor antagonist licensed for asthma prevention, significantly improves NIH clinical score observed in a pilot study involving CF patients (Conway et al., 2003), probably due to ameliorated lung tissue pathology and reduced inflammatory cell infiltration reported in LPS-induced lung inflammation, both in vitro and in mice (Xue et al., 2024). Finally, the quinoline Montelukast, approved for asthma and allergic rhinitis, decreases eosinophil cationic protein and IL-8 serum and sputum levels and sputum levels of myeloperoxidase in CF patients (Schmitt-Grohé and Zielen, 2005).
Bacterial persistence in CF lungs is influenced by local host defense impairments and the pathogens' ability to adapt to a challenging environment due to selective pressures, such as hyperinflammation, oxidative stress, limited nutrients, anaerobiosis, increased acidity, and antibiotic exposure. Biofilm formation is a key adaptive strategy for bacteria, allowing them to persist even when antibiotics are administered. Nowadays, CF is considered a biofilm-associated disease, and controlling these biofilms is crucial for effective antimicrobial strategies. Several studies reported direct S. aureus biofilm visualization in CF patients by scanning electron microscopy and fluorescence electron in situ hybridization using a specific peptide nucleic acid and further viability evaluation by confocal laser scanning microscopy (Jean-Pierre et al., 2022). Biofilm formation was a common trait of both MRSA and MSSA strains in CF patients, and high biofilm-formation ability has been associated with fewer pulmonary exacerbations and, conversely, exacerbations negatively impacted biofilm formation (Wieneke et al., 2021).
In this frame, in the second step of the present study, the compound library also underwent a 96-well microtiter plate-based HTS to identify hits that could affect biofilm formation and preformed–mature - biofilm by the MRSA Sa2 strain. Our findings indicated that 654 compounds affect biofilm formation, although to different extents. Notably, five compounds – i.e., the anticancer Tipifarnib and Olaparib, the bronchodilator Acefylline, Hemin, and the NS5b polymerase inhibitor TMC647055, under investigation for treating hepatitis C – exhibited, for the first time, the highest potential for repurposing due to excellent activity against biofilm formation, which resulted in a reduction of ≥ 90% compared to untreated control (Table 2). A careful revision of the literature aimed at identifying additional biological properties of interest in managing CF patients revealed that Olaparib, Acefylline, and Hemin were previously reported for anti-inflammatory activity in the LPS-induced acute lung injury model (Kapoor et al., 2015; Elzagar et al., 2023; Cheng et al., 2023). Hemin can also affect S. aureus virulence by downregulating hemolysins expression (Schmitt et al., 2012). In disagreement with our findings, Tipifarnib and Hemin have previously been shown to inhibit S. aureus growth (Ladan et al., 1993; Weber et al., 2021).
Regarding hits active against preformed biofilms, four compounds exhibited the highest activity, i.e., 60% ≤ biofilm reduction < 90% (Table 3). The anti-biofilm activity exhibited by Clemastine (fumarate), a histamine receptor H1 antagonist, is probably due to a decrease in the transcriptional level of the biofilm formation relevant fnbB, icaA, and icaB genes in S. aureus (Shang et al., 2022). Conflicting findings come from the literature concerning the antibiofilm potential of Heparin, a highly sulfated glycosaminoglycan with uneven chain length, routinely used in central venous catheters to prevent thrombosis. Our findings disagree with a previous study reporting Heparin augments biofilm formation in S. aureus (Shanks et al., 2005; Najarzadeh et al., 2021), probably due to extracellular DNA-binding proteins on the S. aureus surface that mediates the incorporation of Heparin into the biofilm matrix (Mishra and Horswill, 2017). A similar pro-biofilm effect was observed for Escherichia coli (Wu et al., 2021), Staphylococcus epidermidis (Shanks et al., 2005), and in patients with colorectal cancer where Heparin increased the biofilm formed by intestinal flora, thus providing a protective layer in the intestinal tract of patients (Chen et al., 2022). In agreement with our findings, heparinoids, glycosaminoglycans chemically and pharmacologically related to heparin, were found to suppress biofilm formation in Cutibacterium acnes by inhibiting the AI-2-mediated QS (Hamada et al., 2024). No evidence of activity against preformed biofilms has been previously published for the anticancer Flumatinib (mesylate) and nonsteroidal anti-inflammatory drug Bromfenac (sodium hydrate). All the five hits here showing high potential for dispersing established biofilm have also been reported to trigger anti-inflammatory activity (Kida et al., 2014; Yi et al., 2015; Chimenti et al., 2017; Motawi et al., 2023), although only Heparin under experimental settings relevant to CF patients, such as human bronchial cells (Yi et al., 2015) and LPS-induced lung injury (Chimenti et al., 2017).
4. Materials and Methods
4.1. Compound Library
The “Drug Repurposing Compound Library” was purchased from MedChem Express (cat. no. HY-L035; Monmouth Junction, NJ, USA). The library was provided in a 96-well plate format with aliquots of 10 mM stocks of drugs in DMSO or water, stored at -80°C.
4.2. Bacterial Strain and Growth Conditions
HTS was performed using S. aureus Sa2, an MRSA strain known for causing long-term persistence in a CF patient. Some colonies grew on Tryptone Soya Agar (TSA; Oxoid, Milan, Italy) following overnight incubation at 37°C were suspended in sterile saline 0.9% (Fresenius Kabi Italia, Verona, Italy) to reach an optical density at 550 nm (OD550) of 0.3. This suspension was diluted 1:10 in sterile saline to achieve a final 1-2 x 107 CFU/mL concentration. This standardized inoculum was used for all the assays unless differently indicated.
4.3. Antibacterial HTS Assay
The library was screened at a single concentration point to identify antibacterial hit compounds against the S. aureus Sa2 strain. Briefly, 5 µL of the standardized inoculum (corresponding to 0.5-1 x 105 CFU/well) were added to each well of a 96-well polystyrene microtiter plate containing 100 µL of Tryptone Soya Broth (TSB; Oxoid) and 1 µL of a 10 mM compound stock solution from the MedChem library, achieving a final drug concentration of 0.1 mM. Uninoculated samples with 1% (v/v) DMSO (final background in each well) were considered blank. Negative control was also prepared with 50% (v/v) DMSO to yield 100% killing. The content of each well was mixed by pipetting, and plates were incubated at 37°C under an aerobic atmosphere. After 24 h-incubation, the survival rate of planktonic cells was assessed spectrophotometrically by measuring the broth culture supernatants OD550 using a microplate reader (Tecan Infinite® M PLEX; Tecan Group Ltd Mannedorf, Switzerland). This value was corrected by subtracting the average OD550 value of the uninoculated wells (blank). The percentage of growth rate was calculated compared to the inoculated, but not treated, control sample (100% growth). The antibacterial activity of library drugs was classified based on the growth reduction compared to the untreated control sample: i) low efficacy: 10% < x < 25%; ii) moderate efficacy: 25% ≤ x < 60%; iii) high efficacy: 60% ≤ x < 90%; and iv) excellent efficacy: 90% ≤ x ≤ 100%. Only drugs causing a ≥ 90% reduction of bacterial burden, subsequently confirmed by cell viable count, were considered potential anti-S. aureus hit compounds and underwent further characterization. DMSO at the final concentration of 0.1 mM did not show any activity against S. aureus Sa2, as assessed by cell viable count, thus indicating that the antibacterial effects observed were due to the compound only.
4.4. HTS Assay Validation
The results from each HTS microplate were validated by calculating the Z-factor. To validate the degree of separation, the Z-factor and the percent inhibition of the positive and negative controls were determined using the formula:
where σp and σn are the standard deviations of the positive and negative controls, respectively, and μp and μn are the corresponding mean values. A Z-factor between 0.5 and 1.0 indicates an excellent assay and statistically reliable separation between the positive and negative controls.
4.5. Biofilm Inhibition and Dispersion HTS Assays
To evaluate the effect of the drugs in preventing biofilm formation, 5 μL of the standardized inoculum (1-2 x 107 CFU/mL) was added in each well of a TC-treated microplate (Falcon) containing 100 μL of TSB with 1 μL of a 10 mM compound stock solution from the MediChem library, achieving a final drug concentration of 0.1 mM. Control samples were exposed to TSB only. After incubating at 37°C for 24 h, planktonic cells were gently removed by washing twice with 100 μL PBS. After fixing samples (60°C, 1 h), the biofilm biomass was quantified spectrophotometrically. Briefly, 100 μL of 1% (w/v) Hucker’s crystal violet was added to each well, and after incubation at room temperature for 5 min, each well was washed using tap water. After drying at 37°C, 100 μL 33% (v/v) glacial acetic acid was added to dissolve the stained dye for 15 min. The biofilm biomass was determined by measuring the absorbance at 492 nm using a Tecan Infinite® M PLEX microplate reader (Tecan).
To evaluate the efficacy of drugs against preformed biofilms, 24 h-old biofilms were treated with 100 μL of TSB with the selected drug at a final concentration of 0.1 mM, or TSB only (controls). Biofilm samples were incubated at 37°C for another 24 h and then washed with PBS. As previously described, crystal violet staining finally measured the biofilm biomass.
A potential “antibiofilm hit” had to affect biofilm formation or disperse preformed biofilms, whereas it had no antibacterial activity (growth reduction ≤ 10%). The magnitude of the hit’s activity against biofilm formation or preformed biofilms was categorized as follows: low (i.e., 10% < biofilm reduction < 25%), moderate (i.e., 25% ≤ biofilm reduction < 60%), high (i.e., 60% ≤ biofilm reduction < 90%), and excellent (i.e., biofilm reduction ≥ 90%).
4.6. Statistical Analysis
All assays were conducted in triplicate and repeated twice (n=6). Statistical analysis was performed using GraphPad Prism 7.0 software (GraphPad Software, San Diego, CA, USA). Data were tested for normal distribution using the Shapiro–Wilk test. Ordinary one-way ANOVA with Dunnett’s multiple comparisons test was applied when assessing differences among three or more groups of unpaired data. The statistical analysis assumed a confidence level of ≥ 95%, thus considering p values < 0.05 statistically significant.
5. Conclusions
The present study aimed to identify compounds with high repurposing potential against planktonic and sessile cells of an S. aureus strain causing chronic lung infection in a CF patient. In this regard, HTS of a library compound performed in 96-well microtiter plates with a spectrophotometric assessment of the results revealed a helpful platform. We identified 193 hits causing 100% killing of planktonic cells, five causing biofilm reduction of at least 90%, and four effectively dispersing from 60 to 90% of established biofilm. Most hits were anticancer agents, followed by anti-infective (other than antibacterial) and anti-inflammatory drugs. Also considering other, already published, biological properties that could be relevant to the CF lung environment, among the hits active against planktonic S. aureus cells, those with the most interesting repurposing potential are 5-(N,N-Hexamethylene)-amiloride (active against other CF pathogens and with anti-inflammatory potential), Toremifene (active against other CF pathogens and with anti-virulence potential), and Zafirlukast, Fenretide, and Montelukast (with anti-inflammatory properties already observed in CF patients). Among hits affecting biofilm formation, Hemin also showed anti-virulence and anti-inflammatory effects. Heparin, Clemastine, and Bromfenac are the most interesting hits that significantly affect established biofilms.
Further in vitro and in vivo studies are needed to confirm the potential of repurposing these compounds for managing S. aureus lung infections in CF patients.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Hits (n = 364) showing excellent activity (i.e., growth reduction ≥ 90% vs. unexposed control) against S. aureus Sa2 strain, based on spectrophotometric readings and cell viable count; Table S2: Hits (n = 5) showing excellent activity (i.e., biofilm reduction ≥ 90% vs. unexposed control) against biofilm formation by S. aureus Sa2 strain, based on spectrophotometric readings; Table S3: Hits (n = 4) showing high activity (i.e., 60% ≤ biofilm reduction < 90% vs. unexposed control) against preformed biofilm by S. aureus Sa2 strain, based on spectrophotometric readings.
Author Contributions
A.P.: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Validation, Writing – original draft, Writing – review and editing; V.L.: Investigation, Writing – review and editing; V.P.: Investigation, Writing – review and editing; G.D.B.: Conceptualization, Data curation, Formal analysis, Funding acquisition, Methodology, Supervision, Writing – original draft, Writing – review and editing.
Funding
This work was partly funded by the “G. D’ANNUNZIO” UNIVERSITY OF CHIETI-PESCARA, grant number FAR “ex-60%”, 2022.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data is contained within the article or Supplementary Material.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| CF | Cystic Fibrosis |
| MSSA | Methicillin-Sensitive Staphylococcus aureus |
| MRSA | Methicillin-Resistant Staphylococcus aureus |
| FEV1 | Forced Expiratory Volume in 1 second |
| HTS | High-Throughput Screening |
| VGS | Viridans Group Streptococci |
References
- Aizman, E., Blacher, E., Ben-Moshe, O., Kogan, T., Kloog, Y., Mor, A., 2014. Therapeutic effect of farnesylthiosalicylic acid on adjuvant-induced arthritis through suppressed release of inflammatory cytokines. Clin. Exp. Immunol., 175(3), 458-467. [CrossRef]
- Alnfakh, Z.A., Al-Mudhafar, D.H., Al-Nafakh, R.T., Jasim, A.E., Hadi, N.R., 2022. The anti-inflammatory and antioxidant effects of Montelukast on lung sepsis in adult mice. J. Med. Life, 15(6), 819-827. [CrossRef]
- Alyousef, A.A., Divakar, D.D., Muzaheed., 2017. Chemically modified tetracyclines an emerging host modulator in chronic periodontitis patients: A randomized, double-blind, placebo-controlled, clinical trial. Microb. Pathog., 110, 279-284. [CrossRef]
- Balogh, H., Anthony, A.K., Stempel, R., Vossen, L., Federico, V.A., Valenzano, G.Z., Blackledge, M.S., Miller, H.B., 2024. Novel anti-virulence compounds disrupt exotoxin expression in MRSA. Microbiol. Spectr., 12(12), e0146424. [CrossRef]
- Bhagirath, A.Y., Li, Y., Somayajula, D., Dadashi, M., Badr, S., Duan, K., 2016. Cystic fibrosis lung environment and Pseudomonas aeruginosa infection. BMC Pulm. Med., 16(1), 174. [CrossRef]
- Capecchi, P.L., Laghi Pasini, F., Ceccatelli, L., Di Perri, T., 1993. Isradipine inhibits PMN leukocyte function. A possible interference with the adenosine system. Immunopharmacol. Immunotoxicol., 15(2-3), 133-149. [CrossRef]
- Carullo, G., Di Bonaventura, G., Rossi, S., Lupetti, V., Tudino, V., Brogi, S., Butini, S., Campiani, G., Gemma, S., Pompilio, A., 2023. Development of Quinazolinone Derivatives as Modulators of Virulence Factors of Pseudomonas aeruginosa Cystic Fibrosis Strains. Molecules, 28(18), 6535. [CrossRef]
- Cen, H., Sun, M., Zheng, B., Peng, W., Wen, Q., Lin, Z., Zhang, X., Zhou, N., Zhu, G., Yu, X., Zhang, L., Liang, L., 2024. Hyaluronic acid modified nanocarriers for aerosolized delivery of verteporfin in the treatment of acute lung injury. Int. J. Biol. Macromol., 267(Pt 1), 131386. [CrossRef]
- Chen, Q., Xu, L., Wu, T., Li, J., Hua, L., 2022. Analysis of abnormal intestinal flora on risk of intestinal cancer and effect of heparin on formation of bacterial biofilm. Bioengineered, 13(1), 894-904. [CrossRef]
- Cheng, K.C., Hsu, J.Y., Fu, L.S., Chang, W.C., Chu, J.J., Chi, C.S., 2006. Influence of cetirizine and loratadine on granulocyte-macrophage colony-stimulating factor and interleukin-8 release in A549 human airway epithelial cells stimulated with interleukin-1beta. J. Microbiol. Immunol. Infect., 39(3), 206-211.
- Cheng, X., Yin, M., Sun, X., Zhang, Z., Yao, X., Liu, H., Xia, H., 2023. Hemin attenuated LPS-induced acute lung injury in mice via protecting pulmonary epithelial barrier and regulating HO-1/NLRP3-mediated pyroptosis. Shock, 59(5):744-753. [CrossRef]
- Chimenti, L., Camprubí-Rimblas, M., Guillamat-Prats, R., Gomez, M.N., Tijero, J., Blanch, L., Artigas, A., 2017. Nebulized Heparin Attenuates Pulmonary Coagulopathy and Inflammation through Alveolar Macrophages in a Rat Model of Acute Lung Injury. Thromb. Haemost., 117(11), 2125-2134. [CrossRef]
- Cohn, R.C., Rudzienski, L., 1994. In vitro suppression of Pseudomonas cepacia after limited exposure to subinhibitory concentrations of amiloride and 5-(N,N-hexamethylene) amiloride. Pediatr. Pulmonol., 17(6), 366-369. [CrossRef]
- Conway, S.P., Etherington, C., Peckham, D.G., Whitehead, A., 2003. A pilot study of zafirlukast as an anti-inflammatory agent in the treatment of adults with cystic fibrosis. J. Cyst. Fibros., 2(1), 25-28. [CrossRef]
- Craft, K.M., Nguyen, J.M., Berg, L.J., Townsend, S.D., 2019. Methicillin-resistant Staphylococcus aureus (MRSA): antibiotic-resistance and the biofilm phenotype. Medchemcomm., 10(8), 1231-1241. [CrossRef]
- Dasenbrook, E.C., Merlo, C.A., Diener-West, M., Lechtzin, N., Boyle, M.P., 2008. Persistent methicillin-resistant Staphylococcus aureus and rate of FEV1 decline in cystic fibrosis. Am. J. Respir. Crit. Care Med., 178(8), 814-821. [CrossRef]
- Dasenbrook, E.C., Checkley, W., Merlo, C.A., Konstan, M.W., Lechtzin, N., Boyle, M.P., 2010. Association between respiratory tract methicillin-resistant Staphylococcus aureus and survival in cystic fibrosis. JAMA, 303(23), 2386-2392. [CrossRef]
- De Cremer, K., Delattin, N., De Brucker, K., Peeters, A., Kucharíková, S., Gerits, E., Verstraeten, N., Michiels, J., Van Dijck, P., Cammue, B.P., Thevissen, K., 2014. Oral administration of the broad-spectrum antibiofilm compound toremifene inhibits Candida albicans and Staphylococcus aureus biofilm formation in vivo. Antimicrob. Agents Chemother., 58(12), 7606-7610. [CrossRef]
- Elgazar, A.A., El-Domany, R.A., Eldehna, W.M., Badria, F.A., 2023. Theophylline-based hybrids as acetylcholinesterase inhibitors endowed with anti-inflammatory activity: synthesis, bioevaluation, in silico and preliminary kinetic studies. RSC Adv., 13(36), 25616-25634. [CrossRef]
- Fan, T., Wang, W., Wang, Y., Zeng, M., Liu, Y., Zhu, S., Yang, L., 2024. PDE4 inhibitors: potential protective effects in inflammation and vascular diseases. Front. Pharmacol., 15, 1407871. [CrossRef]
- Farghaly, T.A., Alqurashi, R.M., Masaret, G.S., Abdulwahab, H.G., 2024. Recent Methods for the Synthesis of Quinoxaline Derivatives and their Biological Activities. Mini Rev. Med. Chem., 24(9), 920-982. [CrossRef]
- Farha, M.A., Brown, E.D., 2019. Drug repurposing for antimicrobial discovery. Nat. Microbiol., 4(4), 565-577. [CrossRef]
- Geerdink, L.M., Bertram, H., Hansmann, G., 2017. First-in-child use of the oral selective prostacyclin IP receptor agonist selexipag in pulmonary arterial hypertension. Pulm. Circ., 7(2), 551-554. [CrossRef]
- Gerits, E., Defraine, V., Vandamme, K., De Cremer, K., De Brucker, K., Thevissen, K., Cammue, B.P., Beullens, S., Fauvart, M., Verstraeten, N., Michiels, J., 2017a. Repurposing Toremifene for Treatment of Oral Bacterial Infections. Antimicrob. Agents Chemother., 61(3), e01846-16. [CrossRef]
- Gerits, E., Van der Massen, I., Vandamme, K., De Cremer, K., De Brucker, K., Thevissen, K., Cammue, B.P.A., Beullens, S., Fauvart, M., Verstraeten, N., Michiels, J., 2017b. In vitro activity of the antiasthmatic drug zafirlukast against the oral pathogens Porphyromonas gingivalis and Streptococcus mutans. FEMS Microbiol. Lett., 2017b, 364(2). [CrossRef]
- Guilbault, C., De Sanctis, J.B., Wojewodka, G., Saeed, Z., Lachance, C., Skinner, T.A., Vilela, R.M., Kubow, S., Lands, L.C., Hajduch, M., Matouk, E., Radzioch, D., 2008. Fenretinide corrects newly found ceramide deficiency in cystic fibrosis. Am. J. Respir. Cell. Mol. Biol., 38(1), 47-56. [CrossRef]
- Guilbault, C., Wojewodka, G., Saeed, Z., Hajduch, M., Matouk, E., De Sanctis, J.B., Radzioch, D., 2009. Cystic fibrosis fatty acid imbalance is linked to ceramide deficiency and corrected by fenretinide. Am. J. Respir. Cell. Mol. Biol., 41(1), 100-6. [CrossRef]
- Gulbins, A., Horstmann, M., Daser, A., Flögel, U., Oeverhaus, M., Bechrakis, N.E., Banga, J.P., Keitsch, S., Wilker, B., Krause, G., Hammer, G.D., Spencer, A.G., Zeidan, R., Eckstein, A., Philipp, S., Görtz, G.E., 2023. Linsitinib, an IGF-1R inhibitor, attenuates disease development and progression in a model of thyroid eye disease. Front. Endocrinol. (Lausanne), 14, 1211473. [CrossRef]
- Gutiérrez Santana, J.C., Coria Jiménez, V.R., 2024. Burkholderia cepacia complex in cystic fibrosis: critical gaps in diagnosis and therapy. Ann. Med., 56(1), 2307503. [CrossRef]
- Haddad, J.J., Land, S.C., 2002. Amiloride blockades lipopolysaccharide-induced proinflammatory cytokine biosynthesis in an IkappaB-alpha/NF-kappaB-dependent mechanism. Evidence for the amplification of an antiinflammatory pathway in the alveolar epithelium. Am. J. Respir. Cell. Mol. Biol., 26(1), 114-126. [CrossRef]
- Hamada, S., Minami, S., Gomi, M., 2024. Heparinoid enhances the efficacy of a bactericidal agent by preventing Cutibacterium acnes biofilm formation via quorum sensing inhibition. J. Microorg. Control., 29(1), 27-31. [CrossRef]
- He, J.X., Zhu, C.Q., Liang, G.F., Mao, H.B., Shen, W.Y., Hu, J.B., 2024. Targeted-lung delivery of bardoxolone methyl using PECAM-1 antibody-conjugated nanostructure lipid carriers for the treatment of lung inflammation. Biomed. Pharmacother., 178, 116992. [CrossRef]
- He, Y., Gong, G., Quijas, G., Lee, S.M., Chaudhuri, R.K., Bojanowski, K., 2025. Comparative activity of dimethyl fumarate derivative IDMF in three models relevant to multiple sclerosis and psoriasis. FEBS Open Bio. [CrossRef]
- Hennessy, E., Mooij, M.J., Legendre, C., Reen, F.J., O'Callaghan, J., Adams, C., O'Gara, F., 2013. Statins inhibit in vitro virulence phenotypes of Pseudomonas aeruginosa. J. Antibiot. (Tokyo), 66(2), 99-101. [CrossRef]
- Heo, M.J., Choi, S.Y., Lee, C., Choi, Y.M., An, I.S., Bae, S., An, S., Jung, J.H., 2020. Perphenazine Attenuates the Pro-Inflammatory Responses in Mouse Models of Th2-Type Allergic Dermatitis. Int. J. Mol. Sci., 21(9), 3241. [CrossRef]
- Hu, H., Yang, B., Li, Y., Zhang, S., Li, Z., 2016. Blocking of the P2X7 receptor inhibits the activation of the MMP-13 and NF-κB pathways in the cartilage tissue of rats with osteoarthritis. Int. J. Mol. Med., 38(6), 1922-1932. [CrossRef]
- Hussein, M.H., Schneider, E.K., Elliott, A.G., Han, M., Reyes-Ortega, F., Morris, F., Blastovich, M.A.T., Jasim, R., Currie, B., Mayo, M., Baker, M., Cooper, M.A., Li, J., Velkov, T., 2017. From Breast Cancer to Antimicrobial: Combating Extremely Resistant Gram-Negative "Superbugs" Using Novel Combinations of Polymyxin B with Selective Estrogen Receptor Modulators. Microb. Drug Resist., 23(5), 640-650. [CrossRef]
- Ishikawa, S., Sawamoto, A., Okuyama, S., Nakajima, M., 2021. T-Cell Activation-Inhibitory Assay to Screen Caloric Restriction Mimetics Drugs for Drug Repositioning. Biol. Pharm. Bull., 44(4), 550-556. [CrossRef]
- Jean-Pierre, V., Boudet, A., Sorlin, P., Menetrey, Q., Chiron, R., Lavigne, J.P., Marchandin, H., 2022. Biofilm Formation by Staphylococcus aureus in the Specific Context of Cystic Fibrosis. Int. J. Mol. Sci., 24(1), 597. [CrossRef]
- Kapoor, K., Singla, E., Sahu, B., Naura, A.S., 2015. PARP inhibitor, olaparib ameliorates acute lung and kidney injury upon intratracheal administration of LPS in mice. Mol. Cell. Biochem., 400(1-2), 153-162. [CrossRef]
- Ketabforoush, A.H.M.E., Chegini, R., Barati, S., Tahmasebi, F., Moghisseh, B., Joghataei, M.T., Faghihi, F., Azedi, F., 2023. Masitinib: The promising actor in the next season of the Amyotrophic Lateral Sclerosis treatment series. Biomed. Pharmacother., 160, 114378. [CrossRef]
- Kida, T., Kozai, S., Takahashi, H., Isaka, M., Tokushige, H., Sakamoto, T., 2014. Pharmacokinetics and efficacy of topically applied nonsteroidal anti-inflammatory drugs in retinochoroidal tissues in rabbits. PLoS One, 9(5), e96481. [CrossRef]
- Koch, G., Wermser, C., Acosta, I.C., Kricks, L., Stengel, S.T., Yepes, A., Lopez, D., 2017. Attenuating Staphylococcus aureus Virulence by Targeting Flotillin Protein Scaffold Activity. Cell. Chem. Biol., 24(7), 845-857.e6. [CrossRef]
- Kuang, X., Yang, T., Zhang, C., Peng, X., Ju, Y., Li, C., Zhou, X., Luo, Y., Xu, X., 2020. Repurposing Napabucasin as an Antimicrobial Agent against Oral Streptococcal Biofilms. Biomed. Res. Int., 2020, 8379526. [CrossRef]
- Ladan, H., Nitzan, Y., Malik, Z., 1993. The antibacterial activity of haemin compared with cobalt, zinc and magnesium protoporphyrin and its effect on potassium loss and ultrastructure of Staphylococcus aureus. FEMS Microbiol. Lett., 112(2), 173-7. [CrossRef]
- Langer, T., Hoffmann, R., Bryant, S., Lesur, B., 2009. Hit finding: towards ‘smarter’ approaches. Curr. Opin. Pharmacol., 9, 589–593. [CrossRef]
- Lim, H.J., Kwak, H.J., 2024. Selective PPARδ Agonist GW501516 Protects Against LPS-Induced Macrophage Inflammation and Acute Liver Failure in Mice via Suppressing Inflammatory Mediators. Molecules, 29(21), 5189. [CrossRef]
- LiPuma, J.J., 2010. The changing Microbial Epidemiology in cystic fibrosis. Clin. Microbiol. Rev., 23(2), 299–323.
- Lv, S.L., Zeng, Z.F., Gan, W.Q., Wang, W.Q., Li, T.G., Hou, Y.F., Yan, Z., Zhang, R.X., Yang, M., 2021. Lp-PLA2 inhibition prevents Ang II-induced cardiac inflammation and fibrosis by blocking macrophage NLRP3 inflammasome activation. Acta Pharmacol. Sin., 42(12), 2016-2032. [CrossRef]
- Marcuzzi, A., De Leo, L., Decorti, G., Crovella, S., Tommasini, A., Pontillo, A., 2011. The farnesyltransferase inhibitors tipifarnib and lonafarnib inhibit cytokines secretion in a cellular model of mevalonate kinase deficiency. Pediatr. Res., 70(1), 78-82. [CrossRef]
- Mishra, S., Horswill, A.R., 2017. Heparin Mimics Extracellular DNA in Binding to Cell Surface-Localized Proteins and Promoting Staphylococcus aureus Biofilm Formation. mSphere, 2(3), e00135-17. [CrossRef]
- Mortensen, I., Kristiansen, J.E., Christensen, A.V., Hvidberg, E.F., 1992. The antibacterial effect of some neuroleptics on strains isolated from patients with meningitis. Pharmacol. Toxicol., 71(6), 449-51. [CrossRef]
- Motawi, T.K., El-Maraghy, S.A., Kamel, A.S., Said, S.E., Kortam, M.A., 2023. Modulation of p38 MAPK and Nrf2/HO-1/NLRP3 inflammasome signaling and pyroptosis outline the anti-neuroinflammatory and remyelinating characters of Clemastine in EAE rat model. Biochem. Pharmacol., 209, 115435. [CrossRef]
- Mudgil, U., Khullar, L., Chadha, J., Prerna, Harjai, K., 2024. Beyond antibiotics: Emerging antivirulence strategies to combat Pseudomonas aeruginosa in cystic fibrosis. Microb. Pathog., 193, 106730. [CrossRef]
- Najarzadeh, Z., Zaman, M., Sereikaite, V., Strømgaard, K., Andreasen, M., Otzen, D.E., 2021. Heparin promotes fibrillation of most phenol-soluble modulin virulence peptides from Staphylococcus aureus. J. Biol. Chem., 297(2), 100953. [CrossRef]
- Nilakantan, R., Immermann, F., Haraki, K., 2002. A novel approach to combinatorial library design. Comb. Chem. High Throughput Screen., 5, 105–110. [CrossRef]
- Paganin, P., Fiscarelli, E.V., Tuccio, V., Chiancianesi, M., Bacci, G., Morelli, P., Dolce, D., Dalmastri, C., De Alessandri, A., Lucidi, V., Taccetti, G., Mengoni, A., Bevivino, A., 2015. Changes in cystic fibrosis airway microbial community associated with a severe decline in lung function. PLoS One, 10(4), e0124348. [CrossRef]
- Patti, J.M., Allen, B.L., McGavin, M.J., Höök, M., 1994. MSCRAMM-mediated adherence of microorganisms to host tissues. Annu. Rev. Microbiol., 48, 585-617. [CrossRef]
- Pei, X., Zhang, X.J., Chen, H.M., 2019. Bardoxolone treatment alleviates lipopolysaccharide (LPS)-induced acute lung injury through suppressing inflammation and oxidative stress regulated by Nrf2 signaling. Biochem. Biophys. Res. Commun., 516(1), 270-277. [CrossRef]
- Pinault, L., Han, J.S., Kang, C.M., Franco, J., Ronning, D.R., 2013. Zafirlukast inhibits complexation of Lsr2 with DNA and growth of Mycobacterium tuberculosis. Antimicrob. Agents Chemother., 57(5), 2134-2140. [CrossRef]
- Planagumà, A., Pfeffer, M.A., Rubin, G., Croze, R., Uddin, M., Serhan, C.N., Levy, B.D., 2010. Lovastatin decreases acute mucosal inflammation via 15-epi-lipoxin A4. Mucosal Immunol., 3(3), 270-9. [CrossRef]
- Prieto, J.M., Rapún-Araiz, B., Gil, C., Penadés, J.R., Lasa, I., Latasa, C., 2020. Inhibiting the two-component system GraXRS with verteporfin to combat Staphylococcus aureus infections. Sci. Rep., 10(1), 17939. [CrossRef]
- Puhl, A.C., Gomes, G.F., Damasceno, S., Fritch, E.J., Levi, J.A., Johnson, N.J., Scholle, F., Premkumar, L., Hurst, B.L., Lee-Montiel, F., Veras, F.P., Batah, S.S., Fabro, A.T., Moorman, N.J., Yount, B.L., Dickmander, R.J., Baric, R.S., Pearce, K.H., Cunha, F.Q., Alves-Filho, J.C., Cunha, T.M., Ekins, S., 2022. Vandetanib Blocks the Cytokine Storm in SARS-CoV-2-Infected Mice. ACS Omega, 7(36), 31935-31944. [CrossRef]
- Rajendran, R., Rajendran, V., Böttiger, G., Stadelmann, C., Shirvanchi, K., von Au, L., Bhushan, S., Wallendszus, N., Schunin, D., Westbrock, V., Liebisch, G., Ergün, S., Karnati, S., Berghoff, M., 2023. The small molecule fibroblast growth factor receptor inhibitor infigratinib exerts anti-inflammatory effects and remyelination in a model of multiple sclerosis. Br. J. Pharmacol., 180(23), 2989-3007. [CrossRef]
- Ravi, L.I., Li, L., Wong, P.S., Sutejo, R., Tan, B.H., Sugrue, R.J., 2013. Lovastatin treatment mitigates the pro-inflammatory cytokine response in respiratory syncytial virus infected macrophage cells. Antiviral. Res., 98(2), 332-43. [CrossRef]
- Raza, M., Al-Shabanah, O.A., El-Hadiyah, T.M.H., Qureshi, S., 2000. Thermoregulatory and In-vivo Anti-inflammatory Effects of Vigabatrin In Rat and Mice. Scientia Pharmac., 68(4), 379-388. [CrossRef]
- Rickard, A.H., Gilbert, P., High, N.J., Kolenbrander, P.E., Handley, P.S., 2003. Bacterial coaggregation: an integral process in the development of multi-species biofilms. Trends Microbiol., 11(2), 94-100. [CrossRef]
- Rosett, W., Hodges, G.R., 1980. Antimicrobial activity of heparin. J. Clin. Microbiol., 11(1), 30-4. [CrossRef]
- Schmitt, J., Joost, I., Skaar, E.P., Herrmann, M., Bischoff, M., 2012. Haemin represses the haemolytic activity of Staphylococcus aureus in an Sae-dependent manner. Microbiology (Reading), 158(Pt 10), 2619-2631. [CrossRef]
- Schmitt-Grohé, S., Zielen, S., 2005. Leukotriene receptor antagonists in children with cystic fibrosis lung disease: anti-inflammatory and clinical effects. Paediatr. Drugs, 7(6), 353-363. [CrossRef]
- Shang, Y., Guo, J., Zhao, Y., Chen, J., Meng, Q., Qu, D., Zheng, J., Yu, Z., Wu, Y., Deng, Q., 2002. Clemastine Inhibits the Biofilm and Hemolytic of Staphylococcus aureus through the GdpP Protein. Microbiol. Spectr., 10(2), e0054121. [CrossRef]
- Shanks, R.M., Donegan, N.P., Graber, M.L., Buckingham, S.E., Zegans, M.E., Cheung, A.L., O'Toole, G.A., 2005. Heparin stimulates Staphylococcus aureus biofilm formation. Infect. Immun., 73(8), 4596-4606. [CrossRef]
- Talmon, M., Rossi, S., Pastore, A., Cattaneo, C.I., Brunelleschi, S., Fresu, L.G., 2018. Vortioxetine exerts anti-inflammatory and immunomodulatory effects on human monocytes/macrophages. Br. J. Pharmacol., 175(1):113-124. [CrossRef]
- Tian, Q., Yang, X., Du, J., Huang, H., Liu, W., Zhao, P., 2022. Translocator Protein Ligand Etifoxine Attenuates MPTP-Induced Neurotoxicity. Front. Mol. Neurosci., 15, 850904. [CrossRef]
- Tou, J.S., Urbizo, C., 2008. Diethylstilbestrol inhibits phospholipase D activity and degranulation by stimulated human neutrophils. Steroids, 73(2), 216-221. [CrossRef]
- Townsend, E.A., Guadarrama, A., Shi, L., Roti Roti, E., Denlinger, L.C., 2023. P2X7 signaling influences the production of pro-resolving and pro-inflammatory lipid mediators in alveolar macrophages derived from individuals with asthma. Am. J. Physiol. Lung Cell. Mol. Physiol., 325(4), L399-L410. [CrossRef]
- Tran, T.B., Wang, J., Doi, Y., Velkov, T., Bergen, P.J., Li, J., 2018. Novel Polymyxin Combination With Antineoplastic Mitotane Improved the Bacterial Killing Against Polymyxin-Resistant Multidrug-Resistant Gram-Negative Pathogens. Front. Microbiol., 9, 721. [CrossRef]
- VanDevanter, D.R., LiPuma, J.J., Konstan, M.W., 2024. Longitudinal bacterial prevalence in cystic fibrosis airways: Fact and artifact. J. Cyst. Fibros., 23(1), 58-64. [CrossRef]
- Verhaar, A.P., Wildenberg, M.E., Velde, A.A., Meijer, S.L., Vos, A.C., Duijvestein, M., Peppelenbosch, M.P., Hommes, D.W., van den Brink, G.R., 2013. Miltefosine suppresses inflammation in a mouse model of inflammatory bowel disease. Inflamm. Bowel Dis., 19(9), 1974-1982. [CrossRef]
- Voynikov, Y., Valcheva, V., Momekov, G., Peikov, P., Stavrakov, G., 2014. Theophylline-7-acetic acid derivatives with amino acids as anti-tuberculosis agents. Bioorg. Med. Chem. Lett., 24(14), 3043-3045. [CrossRef]
- Wang, H., Bi, J., Zhang, Y., Pan, M., Guo, Q., Xiao, G., Cui, Y., Hu, S., Chan, C.K., Yuan, Y., Kaneko, T., Zhang, G., Chen, S., 2022. Human Kinase IGF1R/IR Inhibitor Linsitinib Controls the In Vitro and Intracellular Growth of Mycobacterium tuberculosis. ACS Infect. Dis., 8(10), 2019-2027. [CrossRef]
- Wang, J., Feng, H., Li, Z., Zhang, X., 2019. Napabucasin prevents brain injury in neuronal neonatal rat cells through suppression of apoptosis and inflammation. Microb. Pathog., 128, 337-341. [CrossRef]
- Wang, N., Li, W., Yu, H., Huang, W., Qiao, Y., Wang, Q., Wei, Y., Deng, X., Wang, J., Cui, M., Zhang, P., Zhou, Y., 2024. Laurocapram, a transdermal enhancer, boosts cephalosporin's antibacterial activity against Methicillin-resistant Staphylococcus aureus. Biochem. Pharmacol., 227, 116404. [CrossRef]
- Wargo, M.J., Gross, M.J., Rajamani, S., Allard, J.L., Lundblad, L.K., Allen, G.B., Vasil, M.L., Leclair, L.W., Hogan, D.A., 2011. Hemolytic phospholipase C inhibition protects lung function during Pseudomonas aeruginosa infection. Am. J. Respir. Crit. Care Med., 184(3), 345-354. [CrossRef]
- Weber, L., Hagemann, A., Kaltenhäuser, J., Besser, M., Rockenfeller, P., Ehrhardt, A., Stuermer, E., Bachmann, H.S., 2021. Bacteria Are New Targets for Inhibitors of Human Farnesyltransferase. Front. Microbiol., 12, 628283. [CrossRef]
- Wieneke, M.K., Dach, F., Neumann, C., Görlich, D., Kaese, L., Thißen, T., Dübbers, A., Kessler, C., Große-Onnebrink, J., Küster, P., Schültingkemper, H., Schwartbeck, B., Roth, J., Nofer, J.R., Treffon, J., Posdorfer, J., Boecken, J.M., Strake, M., Abdo, M., Westhues, S., Kahl, B.C., 2021. Association of Diverse Staphylococcus aureus Populations with Pseudomonas aeruginosa Coinfection and Inflammation in Cystic Fibrosis Airway Infection. mSphere, 6(3), e0035821. [CrossRef]
- Williams, J.P., Hernady, E., Johnston, C.J., Reed, C.M., Fenton, B., Okunieff, P., Finkelstein, J.N., 2004. Effect of administration of lovastatin on the development of late pulmonary effects after whole-lung irradiation in a murine model. Radiat. Res., 161(5), 560-567. [CrossRef]
- Wu, D., Li, X., Yu, Y., Gong, B., Zhou, X., 2021. Heparin stimulates biofilm formation of Escherichia coli strain Nissle 1917. Biotechnol. Lett., 43(1), 235-246. [CrossRef]
- Wu, J.J., Yuan, X.M., Huang, C., An, G.Y., Liao, Z.L., Liu, G.A., Chen, R.X., 2019. Farnesyl thiosalicylic acid prevents iNOS induction triggered by lipopolysaccharide via suppression of iNOS mRNA transcription in murine macrophages. Int. Immunopharmacol., 68, 218-225. [CrossRef]
- Wu, Y., Zhang, Y., Ge, L., He, S., Zhang, Y., Chen, D., Nie, Y., Zhu, M., Pang, Q., 2024. RTA408 alleviates lipopolysaccharide-induced acute lung injury via inhibiting Bach1-mediated ferroptosis. Int. Immunopharmacol., 142(Pt B), 113250. [CrossRef]
- Xu, X., Zhang, X., Zhang, G., Abbasi Tadi, D., 2024. Prevalence of antibiotic resistance of Staphylococcus aureus in cystic fibrosis infection: a systematic review and meta-analysis. J. Glob. Antimicrob. Resist., 36, 419-425. [CrossRef]
- Xue, T., Zhang, Q., Zhang, T., Meng, L., Liu, J., Chai, D., Liu, Y., Yang, Z., Jiao, R., Cui, Y., Gao, J., Li, X., Xu, A., Zhou, H., 2024. Zafirlukast ameliorates lipopolysaccharide and bleomycin-induced lung inflammation in mice. BMC Pulm. Med., 24(1), 456. [CrossRef]
- Yang, D., Xu, D., Wang, T., Yuan, Z., Liu, L., Shen, Y., Wen, F., 2021. Mitoquinone ameliorates cigarette smoke-induced airway inflammation and mucus hypersecretion in mice. Int. Immunopharmacol., 90, 107149. [CrossRef]
- Ye, B., Xiong, X., Deng, X., Gu, L., Wang, Q., Zeng, Z., Gao, X., Gao, Q., Wang, Y., 2017. Meisoindigo, but not its core chemical structure indirubin, inhibits zebrafish interstitial leukocyte chemotactic migration. Pharm. Biol., 55(1), 673-679. [CrossRef]
- Ye, Y., Jin, T., Zhang, X., Zeng, Z., Ye, B., Wang, J., Zhong, Y., Xiong, X., Gu, L., 2019. Meisoindigo Protects Against Focal Cerebral Ischemia-Reperfusion Injury by Inhibiting NLRP3 Inflammasome Activation and Regulating Microglia/Macrophage Polarization via TLR4/NF-κB Signaling Pathway. Front. Cell. Neurosci., 13, 553. [CrossRef]
- Yi, N.Y., Newman, D.R., Zhang, H., Morales Johansson, H., Sannes, P.L., 2015. Heparin and LPS-induced COX-2 expression in airway cells: a link between its anti-inflammatory effects and GAG sulfation. Exp. Lung Res., 41(9), 499-513. [CrossRef]
- Yin, Z., Wang, Y., Whittell, L.R., Jergic, S., Liu, M., Harry, E., Dixon, N.E., Kelso, M.J., Beck, J.L., Oakley, A.J., 2014. DNA replication is the target for the antibacterial effects of nonsteroidal anti-inflammatory drugs. Chem. Biol., 21(4), 481-487. [CrossRef]
- Yotis, W., 1977. Effects of diethylstilbestrol on the production of various extracellular products of Staphylococcus aureus. Experientia, 33(3), 325-326. [CrossRef]
- Yu, H., Valerio, M., Bielawski, J., 2013. Fenretinide inhibited de novo ceramide synthesis and proinflammatory cytokines induced by Aggregatibacter actinomycetemcomitans. J. Lipid Res., 54(1), 189-201. [CrossRef]
- Zappala, C., Chandan, S., George, N., Faoagali, J., Boots, R.J., 2007. The antimicrobial effect of heparin on common respiratory pathogens. Crit. Care Resusc., 9(2), 157-160.
- Zhang, W.B., Yang, F., Wang, Y., Jiao, F.Z., Zhang, H.Y., Wang, L.W., Gong, Z.J., 2019. Inhibition of HDAC6 attenuates LPS-induced inflammation in macrophages by regulating oxidative stress and suppressing the TLR4-MAPK/NF-κB pathways. Biomed. Pharmacother., 117, 109166. [CrossRef]
- Zheng, J., Shang, Y., Wu, Y., Zhao, Y., Chen, Z., Lin, Z., Li, P., Sun, X., Xu, G., Wen, Z., Chen, J., Wang, Y., Wang, Z., Xiong, Y., Deng, Q., Qu, D., Yu, Z., 2022. Loratadine inhibits Staphylococcus aureus virulence and biofilm formation. iScience, 25(2), 103731. [CrossRef]
Figure 1.
Drug Repurposing Compound Library (MedChem Express) structure: clinical information (a) and research area (b). Reported are the number of drugs and relative percentage. Some drugs belong to more research areas.
Figure 1.
Drug Repurposing Compound Library (MedChem Express) structure: clinical information (a) and research area (b). Reported are the number of drugs and relative percentage. Some drugs belong to more research areas.
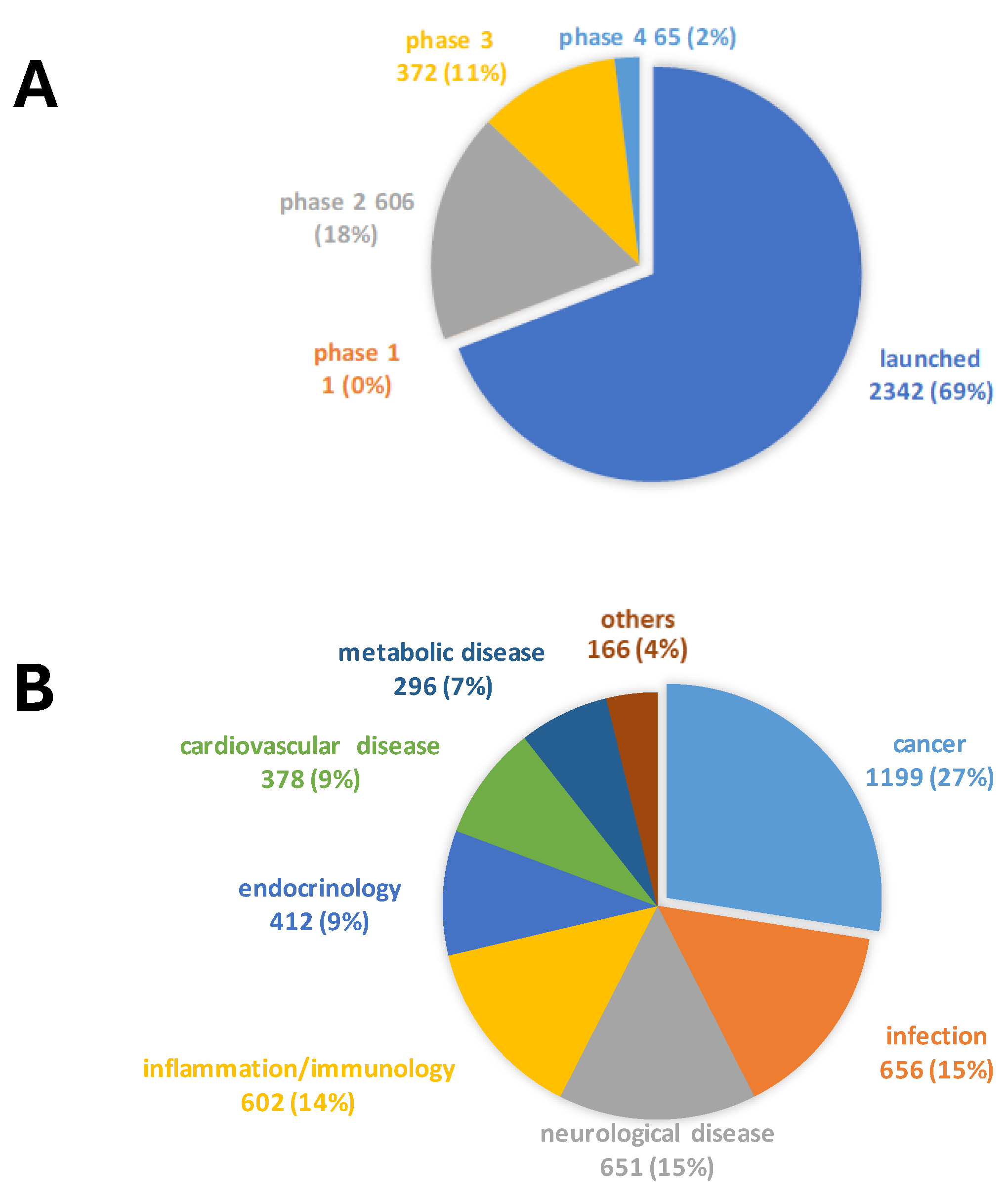
Figure 2.
Z-factor plot from high-throughput screening performed in 96-well plates. Solid dots represent the Z′ factor from thirty-four 96-well plates. The dashed line indicates the threshold of 0.5.
Figure 2.
Z-factor plot from high-throughput screening performed in 96-well plates. Solid dots represent the Z′ factor from thirty-four 96-well plates. The dashed line indicates the threshold of 0.5.
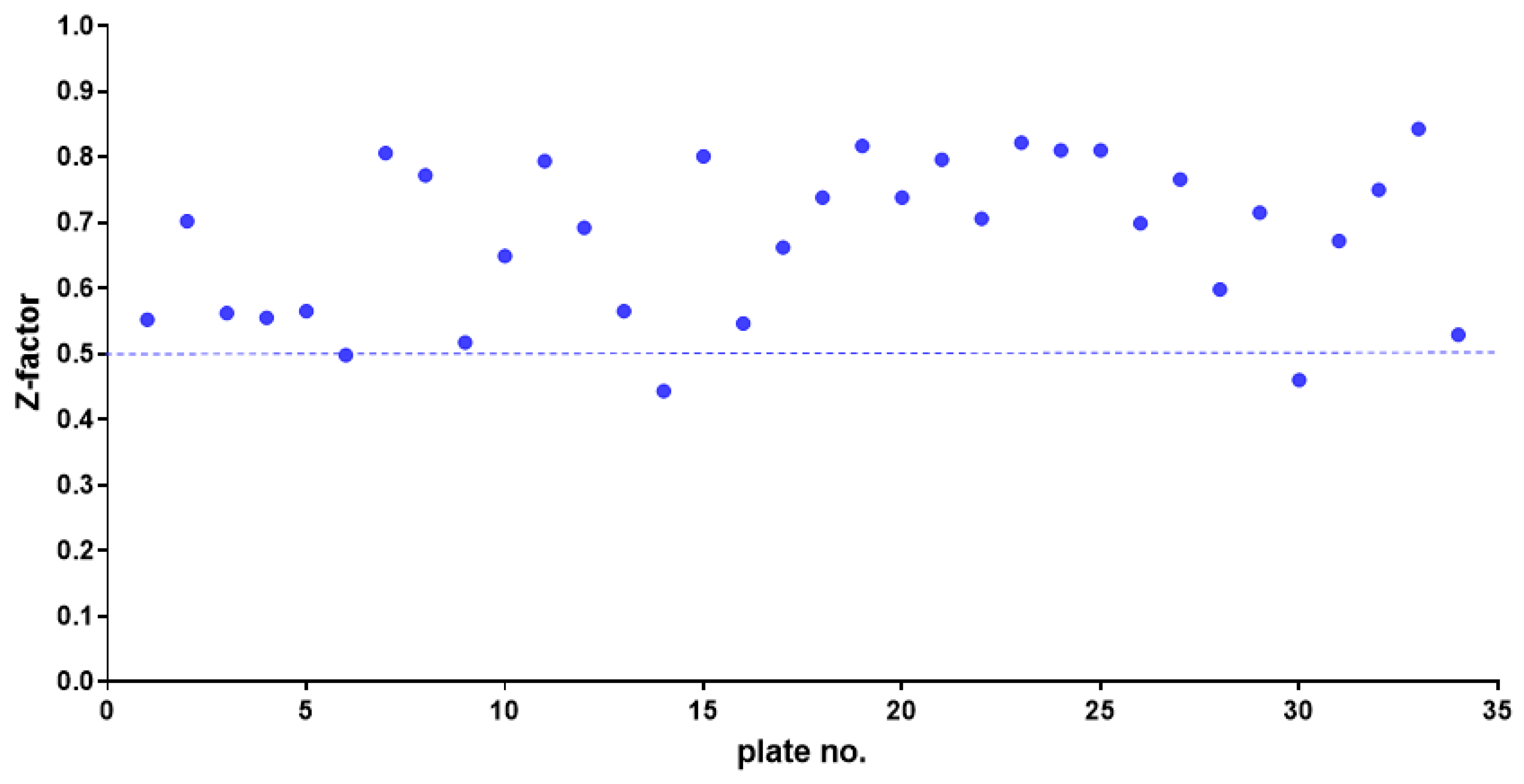
Figure 3.
HTS of the compound library to identify active hits against planktonic S. aureus Sa2 cells. The antibacterial activity of compounds in several research areas is shown as a percentage referred to as the untreated control.
Figure 3.
HTS of the compound library to identify active hits against planktonic S. aureus Sa2 cells. The antibacterial activity of compounds in several research areas is shown as a percentage referred to as the untreated control.
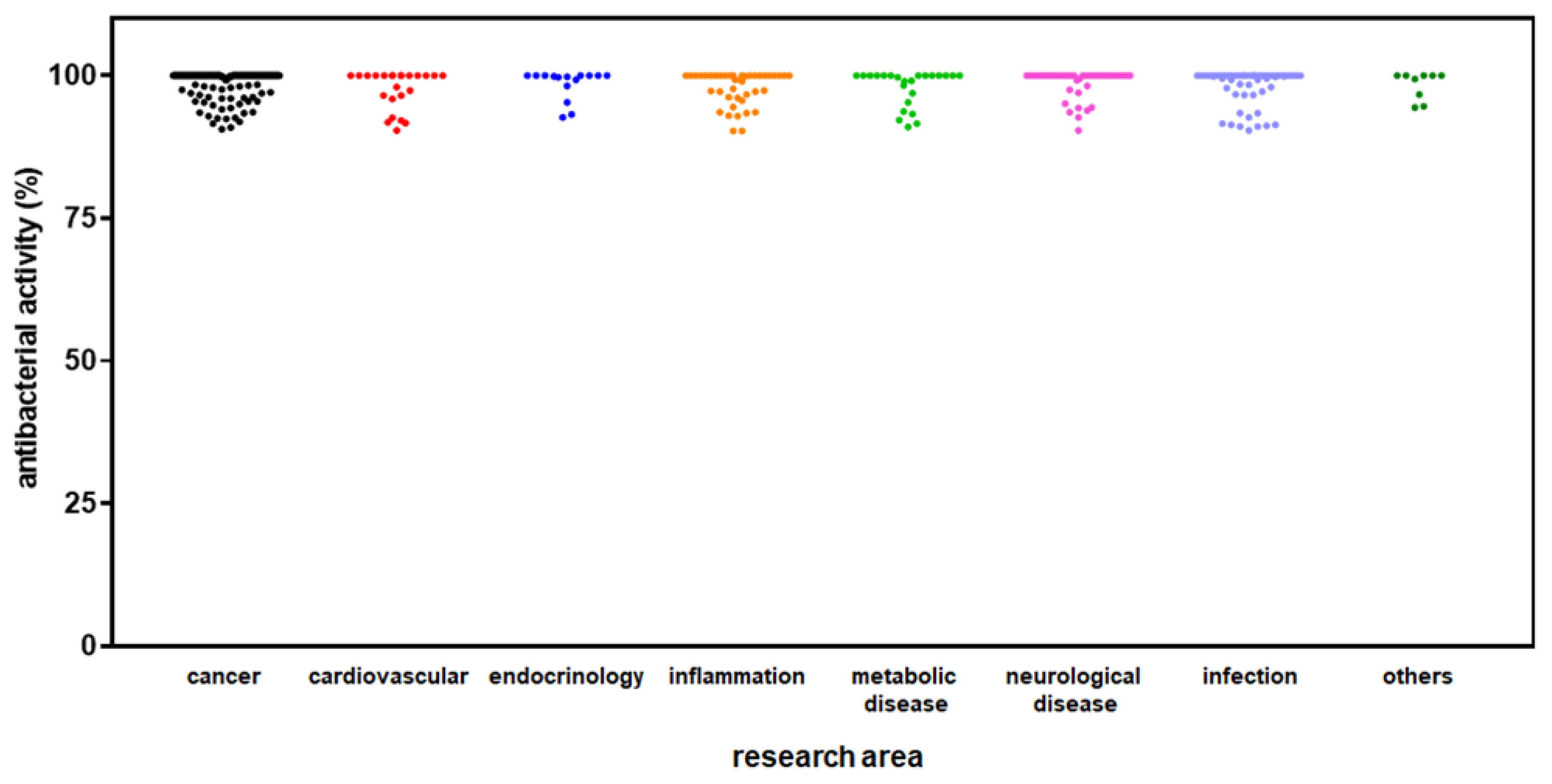
Figure 4.
HTS of the compound library to identify active hits against biofilm formation by S. aureus Sa2 strain. The anti-biofilm activity of compounds in several research areas is shown as a percentage referred to as the untreated control. The magnitude of the anti-biofilm activity is categorized as follows: low (i.e., 10% < biofilm reduction < 25%), moderate (i.e., 25% ≤ biofilm reduction < 60%), high (i.e., 60% ≤ biofilm reduction < 90%), and excellent (i.e., biofilm reduction ≥ 90%).
Figure 4.
HTS of the compound library to identify active hits against biofilm formation by S. aureus Sa2 strain. The anti-biofilm activity of compounds in several research areas is shown as a percentage referred to as the untreated control. The magnitude of the anti-biofilm activity is categorized as follows: low (i.e., 10% < biofilm reduction < 25%), moderate (i.e., 25% ≤ biofilm reduction < 60%), high (i.e., 60% ≤ biofilm reduction < 90%), and excellent (i.e., biofilm reduction ≥ 90%).
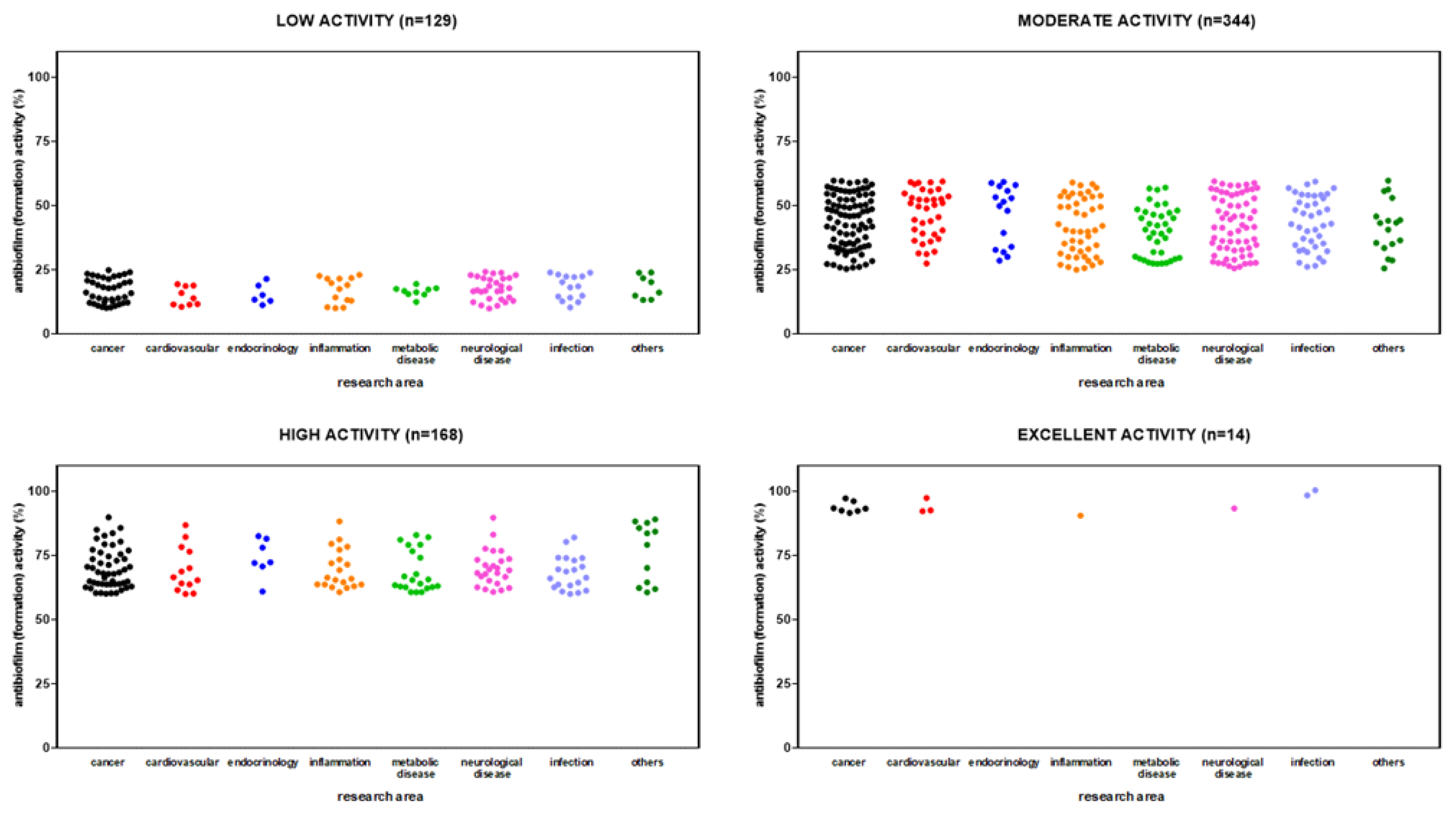
Figure 5.
HTS of the compound library to identify active hits against preformed biofilm by S. aureus Sa2 strain. The anti-biofilm activity of compounds in several research areas is shown as a percentage referred to as the untreated control. The magnitude of the anti-biofilm activity is categorized as follows: low (i.e., 10% < biofilm reduction < 25%), moderate (i.e., 25% ≤ biofilm reduction < 60%), high (i.e., 60% ≤ biofilm reduction < 90%), and excellent (i.e., biofilm reduction ≥ 90%).
Figure 5.
HTS of the compound library to identify active hits against preformed biofilm by S. aureus Sa2 strain. The anti-biofilm activity of compounds in several research areas is shown as a percentage referred to as the untreated control. The magnitude of the anti-biofilm activity is categorized as follows: low (i.e., 10% < biofilm reduction < 25%), moderate (i.e., 25% ≤ biofilm reduction < 60%), high (i.e., 60% ≤ biofilm reduction < 90%), and excellent (i.e., biofilm reduction ≥ 90%).
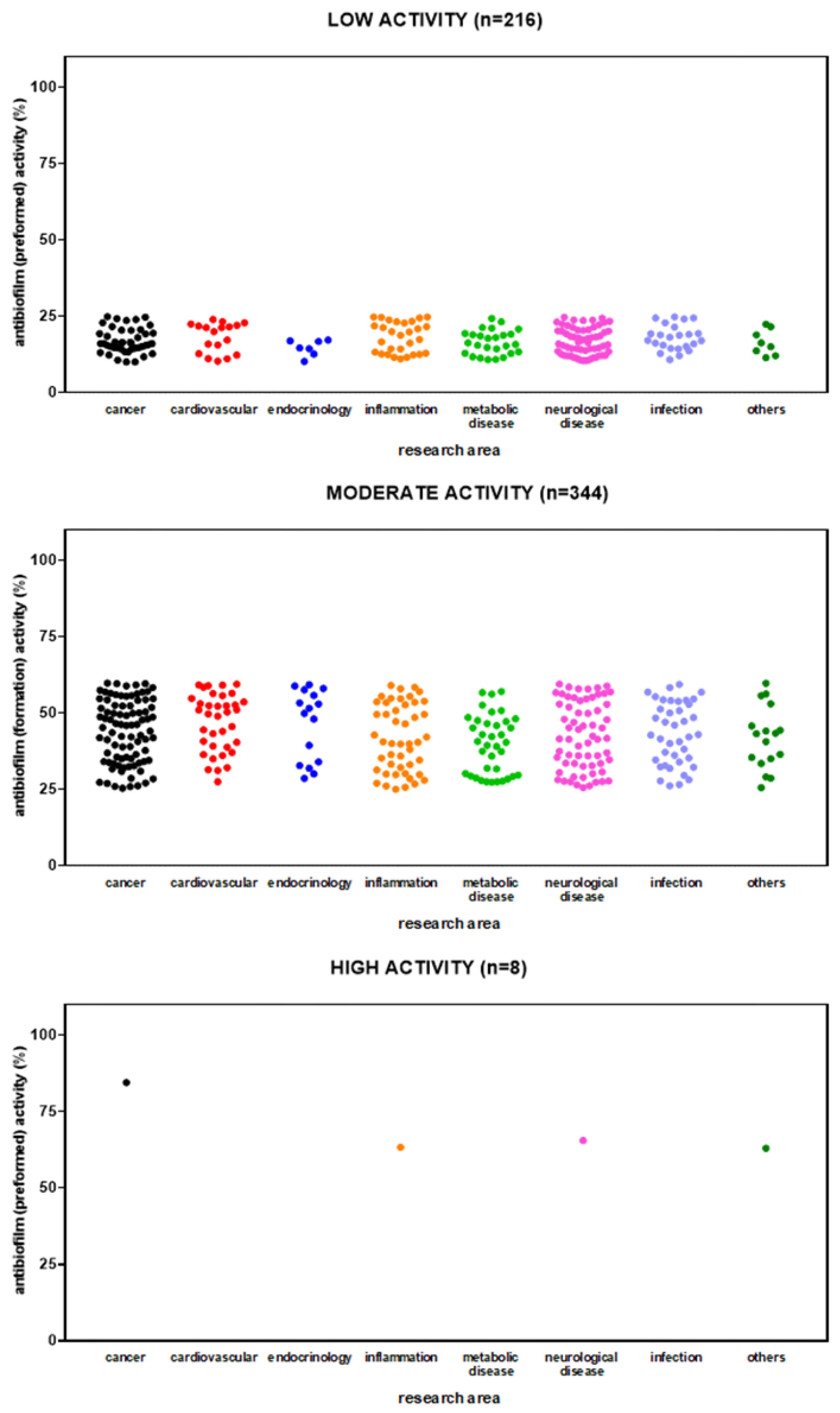

Table 1.
Compounds (n = 37) with maximum activity against planktonic cells (i.e., 100% growth reduction) of S. aureus Sa2 at HTS. Classification, therapeutic category, and mechanism of action are listed, along with other biological properties relevant to cystic fibrosis (CF).
Table 1.
Compounds (n = 37) with maximum activity against planktonic cells (i.e., 100% growth reduction) of S. aureus Sa2 at HTS. Classification, therapeutic category, and mechanism of action are listed, along with other biological properties relevant to cystic fibrosis (CF).
| Compound | Classification Therapeutic category and indication(s) Mechanism(s) of action |
Anti-bacterial activity | Anti-virulence activity | Anti-inflammatory activity |
|---|---|---|---|---|
| Perphenazine |
|
|
|
|
| Miltefosine |
|
|
|
|
| Diethylstilbestrol |
|
|
|
|
| Selexipag |
|
|
||
| AZD-9056 |
|
|
||
| Vortioxetine |
|
|
||
| Lovastatin |
|
|
|
|
| Fenretinide |
|
|
||
| Napabucasin |
|
|
|
|
| Loratadine |
|
|
|
|
| Toremifene |
|
|
|
|
| Etifoxine |
|
|
||
| 5-(N,N-Hexamethylene)-amiloride |
|
|
|
|
| Zafirlukast |
|
|
|
|
| Mitoquinone (mesylate) |
|
|
||
| Crisaborole |
|
|
||
| Linsitinib |
|
|
||
| GW 501516 (Cardarine) |
|
|
||
| Darapladib |
|
|
||
| Bardoxolone |
|
|
||
| Meisoindigo |
|
|
||
| Epinastine |
|
|
||
| Salirasib |
|
|
||
| Omaveloxolone |
|
|
||
| Incyclinide |
|
|
||
| Glecaprevir |
|
|
||
| Isradipine |
|
|
||
| Laurocapram |
|
|
||
| Masitinib |
|
|
||
| Mitotane |
|
|
||
| Montelukast |
|
|
||
| Ricolinostat |
|
|
||
| Vandetanib |
|
|
||
| Verteporfin |
|
|
||
| Vigabatrin |
|
|
||
| Diroximel (fumarate) |
|
|
||
| Infigratinib |
|
|
Data source: PubChem (https://pubchem.ncbi.nlm.nih.gov/). The literature review was performed using PubMed (https://pubmed.ncbi.nlm.nih.gov/) using the following keywords: [compound name] and “antibacterial”, “aureus”, “aeruginosa”, “inflammatory”, “inflammatory lung”, “virulence” and “cystic fibrosis”.
Table 2.
Compounds (n = 5) with excellent activity against biofilm formation (i.e., biofilm reduction ≥ 90%) by S. aureus Sa2 at HTS. Classification, therapeutic category, and mechanism of action are listed, along with other biological properties relevant to cystic fibrosis (CF).
Table 2.
Compounds (n = 5) with excellent activity against biofilm formation (i.e., biofilm reduction ≥ 90%) by S. aureus Sa2 at HTS. Classification, therapeutic category, and mechanism of action are listed, along with other biological properties relevant to cystic fibrosis (CF).
| Compound | Classification Therapeutic category and indication(s) Mechanism(s) of action |
Anti-bacterial activity | Anti-virulence activity | Anti-inflammatory activity |
|---|---|---|---|---|
| Tipifarnib |
|
|
|
|
| Olaparib |
|
|
||
| Acefylline |
|
|
|
|
| Hemin |
|
|
|
|
| TMC647055 (Choline salt) |
|
Data source: PubChem (https://pubchem.ncbi.nlm.nih.gov/). The literature review was performed using PubMed (https://pubmed.ncbi.nlm.nih.gov/) using the following keywords: [compound name] and “antibacterial”, “aureus”, “aeruginosa”, “inflammatory”, “inflammatory lung”, “virulence” and “cystic fibrosis”.
Table 3.
Compounds (n=4) with good activity against established biofilm (i.e., 60% ≤ biofilm dispersion < 90%) by S. aureus Sa2 at HTS. Classification, therapeutic category, and mechanism of action are listed, along with other biological properties relevant to cystic fibrosis (CF).
Table 3.
Compounds (n=4) with good activity against established biofilm (i.e., 60% ≤ biofilm dispersion < 90%) by S. aureus Sa2 at HTS. Classification, therapeutic category, and mechanism of action are listed, along with other biological properties relevant to cystic fibrosis (CF).
| Compound | Classification Therapeutic category and indication(s) Mechanism(s) of action |
Anti-bacterial activity | Anti-virulence activity | Anti-inflammatory activity |
|---|---|---|---|---|
| Clemastine (fumarate) |
|
|
|
|
| Heparin |
|
|
|
|
| Flumatinib (mesylate) |
|
|||
| Bromfenac (sodium hydrate) |
|
|
|
Data source: PubChem (https://pubchem.ncbi.nlm.nih.gov/). The literature review was performed using PubMed (https://pubmed.ncbi.nlm.nih.gov/) using the following keywords: [compound name] and “antibacterial”, “aureus”, “aeruginosa”, “inflammatory”, “inflammatory lung”, “virulence” and “cystic fibrosis”.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



