Submitted:
01 January 2025
Posted:
03 January 2025
You are already at the latest version
Abstract
Panax ginseng C.A. Meyer is one of Korea's most popular traditional medicinal plants. Since ginseng seeds have morpho-physiological dormancy and a very short lifespan, the harvested seeds need outdoor warm and cold stratification for 100 days each. The seeds are covered by a fruit coat (endocarp), which should be opened during the warm stratification. Hence, farmers need to dehisce (open the endocarp) the seeds annually. The author examines conditions for embryo growth, dehiscence percentage, and endo-carp hardness as a factor of temperature, watering, stratification substances, solution scarification, microbial inoculation on the seed endocarp, etc. Watering, temperature (17.5°C), and aeration are crucial for embryo growth as a germination condition. Moreover, microbial-mediated decomposition of the endocarp is necessary for dehiscence and further embryo development. This study suggests that a combination of embryo growth and microbial-mediated decomposing of the endocarp during warm stratification is a prerequisite for the dehiscence of ginseng seeds, implying physical dormancy and morpho-physiological dormancy. The un-dehisced seeds of three P. ginseng varieties and one P. quinquefolius (American ginseng) were dehisced indoors with microbial inoculation instead of sand as a stratification substrate. This approach opens the door for year-round indoor dehiscence of ginseng seeds without sand.
Keywords:
embryo-to-endosperm length ratio
; endocarp hardness
; dehiscence
; Panax quinquefolius
1. Introduction
Panax ginseng C.A. Meyer is one of Korea’s most popular traditional medicinal plants. Despite their seed-propagating manner as self-pollinating species, Korean ginseng germplasm has been maintained in field collection due to its short lifespan and more extended cultivation period (3-4 years) for fruiting [1]. The dehiscence percentage of un-dehisced ginseng seeds decreased to 38.7% after one year of cold storage and did not germinate [2]. Therefore, farmers need seedlings from freshly harvested seeds a year before planting. Cryopreservation protocol and cryobanking of ginseng seeds at the after-ripening stage of un-dehisced (with or without fruits), dehisced, and cold-acclimated (ready to germinate) were established [3,4]. However, genebank staff encountered difficulties in germinating the cryopreserved seeds.
The ginseng seeds have immature embryos (0.35~0.4 mm long) at maturity in late Summer, germinating in the second spring after dispersal, i.e., 18-21 months in natural conditions [5,6]. After removing the fruits, the seeds were traditionally mixed with river sand (1:3 ratio) and subjected to underground watering 1~3 times a day for about 100 days (late July to early November) to shorten the after-ripening period. During the warm stratification at moderate temperature, embryos grow from the embryo-to-endosperm length ratio (EER) of 0.09 to 0.4, and endocarps are opened (dehiscence).
Following the warm stratification, the seeds need cold stratification for another 100 days at 4~5°C before germination due to their physiological dormancy [7,8]. In practice, seeds were sown in the field, allowing chilling in winter and germination in the following spring [8]. Pre-soaking in gibberellic acid (GA3) may improve ginseng seeds’ embryo development and dehiscence but increase the decaying of seeds [6,9].
In 2007, I aimed to develop an automatic dehiscence-stratification machinery like a laundry machine. However, I failed to dehiscence the ginseng seeds in chambers: embryos do not grow well due to oxygen limits, and endocarps do not open without microorganisms’ decomposing action. I modified the procedure by supplying the air into the chamber and inoculating the microorganisms in advance, which were placed in a chamber and then stratified.
Based on the seed dormancy classification, ginseng seeds have morpho-physiological (MDP) [10,11]. Ginseng seeds have been considered to have no physical dormancy [12], and microbial action-mediated endocarp opening is considered less likely [13]. However, the preliminary experiments raise the possibility that ginseng seeds have mechanical (physical) dormancy due to the fruit coat (endocarp). Kwon et al. [8] also argued that ginseng seeds have triple dormancy.
This study aimed to develop artificial dehiscence (warm stratification) of ginseng seeds approaching under automatic machinery instead of traditional stratification facilities. I examines conditions for endocarp hardness, embryo growth, and dehiscence percentage as a factor of temperature, watering, diverse microbial inoculation, etc. This study suggests that the combination of embryo growth and microbial-mediated decomposing of the endocarp during warm stratification is a prerequisite for the dehiscence of ginseng seeds.
2. Materials and Methods
2.1. Plant Material
Korean ginseng (Panax ginseng C.A. Meyer) landrace ‘Jakyungjong’ was used in this study. The red-purple-colored berries with seeds were harvested randomly from a population of 4-year-old plants in late July - early August (Figure 1-A). Harvested fruits were pulped, and un-dehiscent seeds were soaked in running water for a day. The seeds of no less than 4 mm in diameter were stored at 10°C before the experiments (Figure 1-B). For the year-round experiments, un-dehisced seeds were cryopreserved (vapor phase) at a moisture content (MC) of 6% [3]. After cryopreservation, seeds were moistened at MC 45% before the warm stratification experiments
2.2. Warm Stratification of Un-dehisced Seeds
2.2.1. Warm Stratification
The un-dehisced seeds surrounded by endocarp (hereafter “seeds”) were loaded into an onion net (10 x 20 cm) and placed in a pot (15 x 25 cm). The seeds were maintained in a ventilated room at 18-20°C and watered daily for 3-4 months until dehiscence (opening of the endocarp) was observed. Based on the experimental design, diverse options were tested, i.e., stratification temperatures, watering intervals, stratification substrates, solution scarifications, microbial inoculations, etc.
2.2.2. Warm Stratification Treatments
- Warm Stratification Temperature (Figure 2)
Un-dehisced seeds located within the onion net were stratified in incubators controlled at 10°C, 15°C, 17.5°C, 20°C, 22.5°C, 25°C or alternating temperature of 15-25°C (20°C, 8h→ 25°C, 4h→ 20°C, 8h→ 15°C, 4h) for 24 weeks. The seeds were watered once a day.
- Stratification Substrates and Fungi Inoculation (Figure 3)
The un-dehisced seeds were mixed with rice husk, vermiculite, or river sand (1:3 ratio). The intact seeds that did not mix with the substrates were prepared as a control. These four options of seeds were inoculated with a fungi mix (four strains of Talaromyces flavus), which were isolated from the dehisced seeds endocarp of KT&G farm (the government agency for ginseng and tabaco) (Table 1). The strains were cultured in rice bran obtained by milling rice for 1 week at 27°C. The mixture of seeds and fungal inoculum (1% w/v) was co-cultured for a day at 25°C and loaded in a pot maintained at 18-20°C room without water for 3-5 days. The seeds were watered once a day during the warm stratification. Watering thrice a day or once every three days was compared.
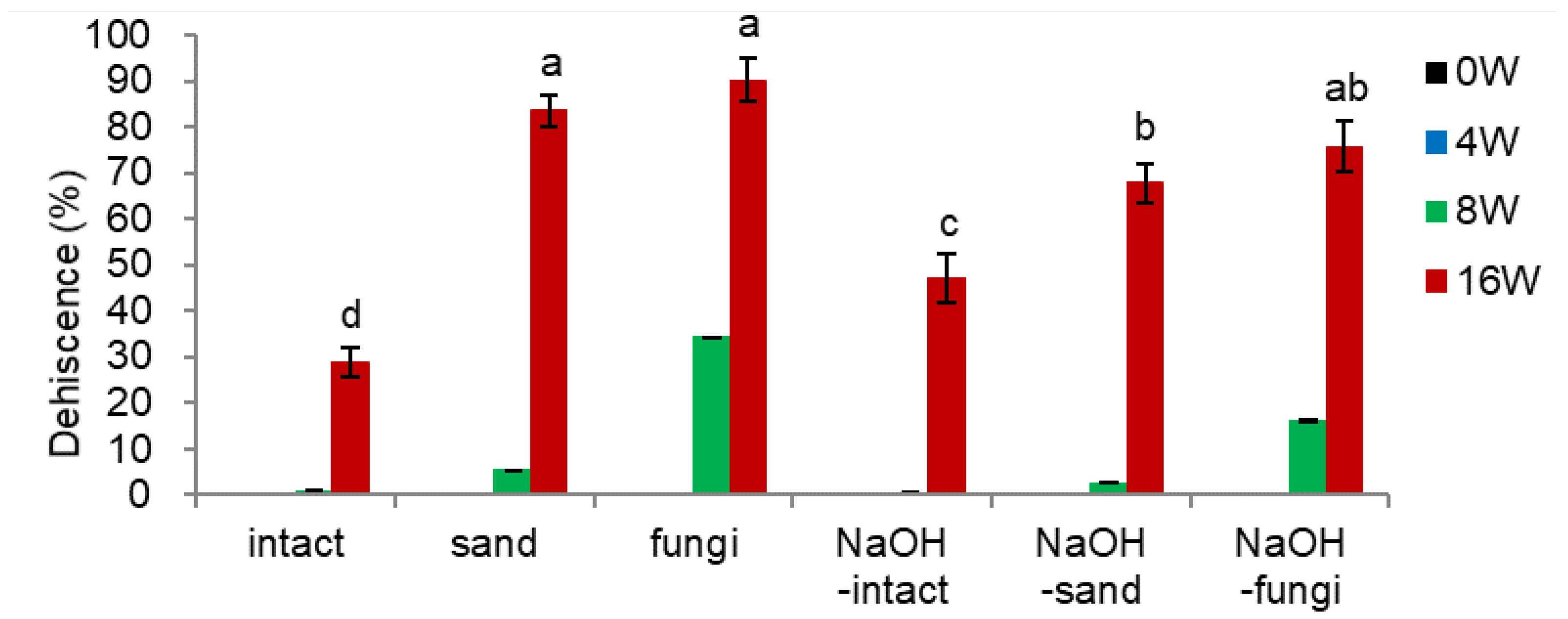
- NaOH soaking and Fugi Inoculation (Figure 4)
From the preliminary experiments, soaking the seeds in 5% sodium hydroxide (NaOH) for 25 min, followed by washing with running tap water overnight, was suitable to decrease the endocarp hardness. The combinational effect of NaOH-soaked seeds and three seed options of intact (intact), mixed with sand (sand), or inoculated with fungi-mix (fungi) were tested. The treated seeds were located within the onion net and watered once daily.
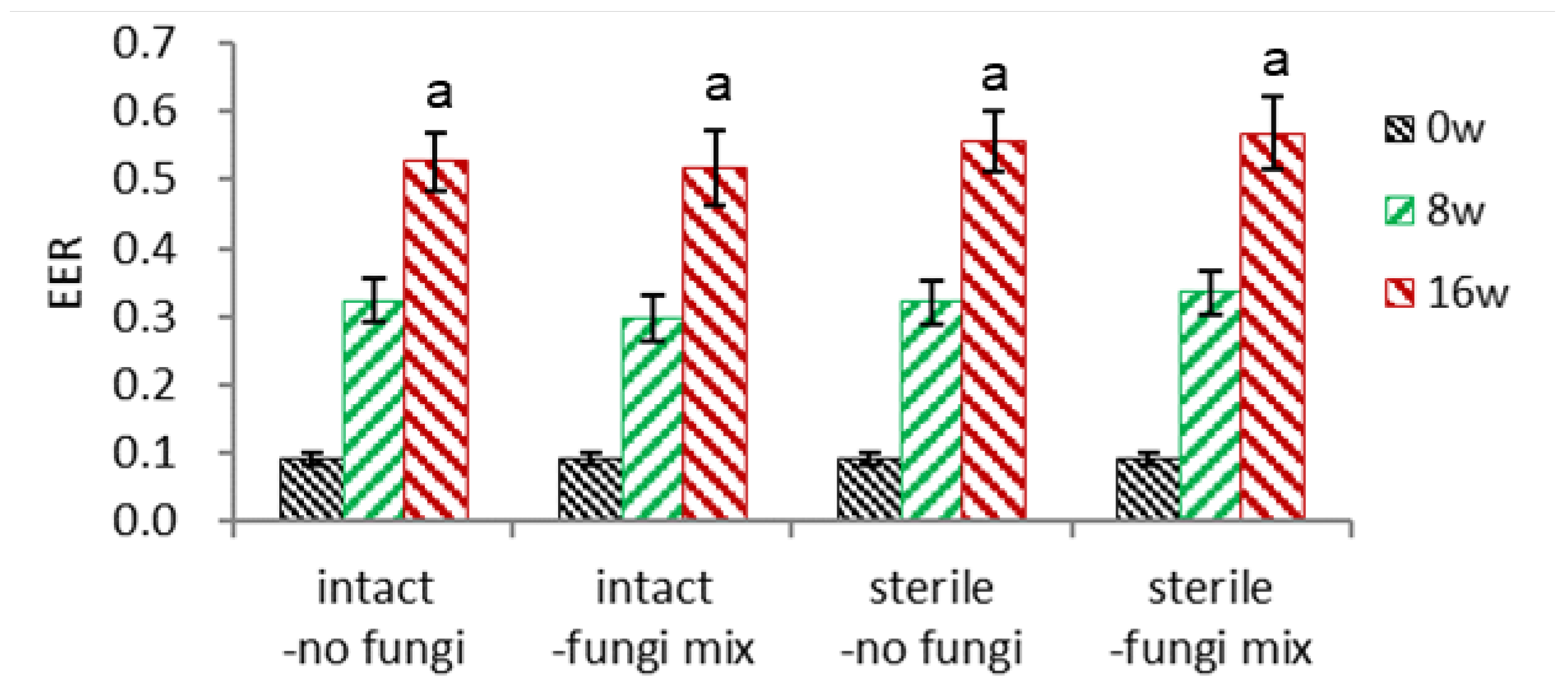
- Sterilization of Seeds and Fungi Inoculation (Figure 5)
The combinational effect of intact/sterile and no fungi/fungi-inoculated were tested. The seeds were sterilized with 1.5% NaOCl for 12 minutes and washed three times with tap water. The seeds were inoculated with a fungi mix of 1% w/v, co-cultured a day at 25°C, and loaded in a pot, maintained at 18-20°C room without watering for 3-5 days. The seeds were watered once a day during the warm stratification.
- Inoculation of Fungi, Actinomycetes, Bacteria (Figure 6)
The seeds were inoculated with 1% (w/v) of fungi-mix, actinomycetes-mix, and bacteria-mix (Table 1) and co-cultured for a day at 25°C, loaded in a pot, maintained at 18-20°C room without water for 3-5 days. The seeds were watered once a day during the warm stratification.
- Inoculation of Fungi, Actinomycetes, and Bacteria of Cellulose Dissociation Activity (Figure 7)
The seeds were inoculated with seven different options of microbial strains of high and low cellulose dissociation activity (Table 1). The seeds were inoculated individually or in the mixture (total concentration of 1%, w/v) of fungus, actinomyces, or bacteria and co-cultured for a day at 25°C, loaded in a pot, and maintained at 18-20°C room without watering for 3-5 days. Later on, the seeds were watered once a day during the warm stratification.
2.2.2. Application of Fungi Inoculation to Ginseng Varieties (Figure 8)
Three varieties of Korean ginseng (Panax ginseng) landrace ‘Jakyeong (JK)’, the newly released variety ‘Yeonpoong (YP)’, and Cheonpoong (CP)’ seeds and one American ginseng variety (Panax quinquefolius) were stratified as non-treated (intact) or fungi #4 inoculated.
2.2.3. Measurement of Endocarp Hardness, Embryo Growth and Dehiscence
- Endocarp Hardness
The hardness (kg cm−1) of the seed endocarp was measured by a Rockwell Hardness Tester (Korea). Seeds (with endocarp) were mounted vertically on the tester plate. It measured the hardness when the endosperm was split. The hardness of the endocarp opened (dehisced) seeds was recorded as zero.
- Embryo-to-Endosperm Length Ration (EER)
The length of the embryo and endosperm was measured periodically at 0, 4, 8, 12, 16, 20, and 24 weeks. Fifteen seeds were longitudinally cut in half using a surgical knife, and embryo length was measured using the analytical software loaded in the Leica microscope (Wetzlar, Germany). The embryo-to-endosperm length ratio (EER) was expressed as the ratio of the length of the embryo to the endosperm.
- Dehiscence percentage
Seeds were considered to be dehisced when the endocarp was opened, no less than 3-4 mm. Dehiscence was expressed as the percentage of seeds with an open endocarp out of the total number of seeds.
2.3. Statistical Analysis
For the measurement of endocarp hardness and EER of all treatments, 15 seeds were randomly chosen for each period of 0, 4, 8, 12, and 16 weeks during the warm stratification. One hundred seeds with five replications (n = 500) were monitored for dehiscence percentage. At each period, dehisced seeds were removed from the onion net.
The data from all experiments of the last period (16 weeks) were analyzed by analysis of variance (ANOVA) and Duncan’s multiple range test (p < 0.05) using SAS on Demand for Academics software (SAS Institute Inc., USA).
3. Results
3.1. Effect of Warm Stratification Treatments
3.1.1. Effect of Warm Stratification Temperature
- Embryo-to-Endosperm Length Ratio (EER)
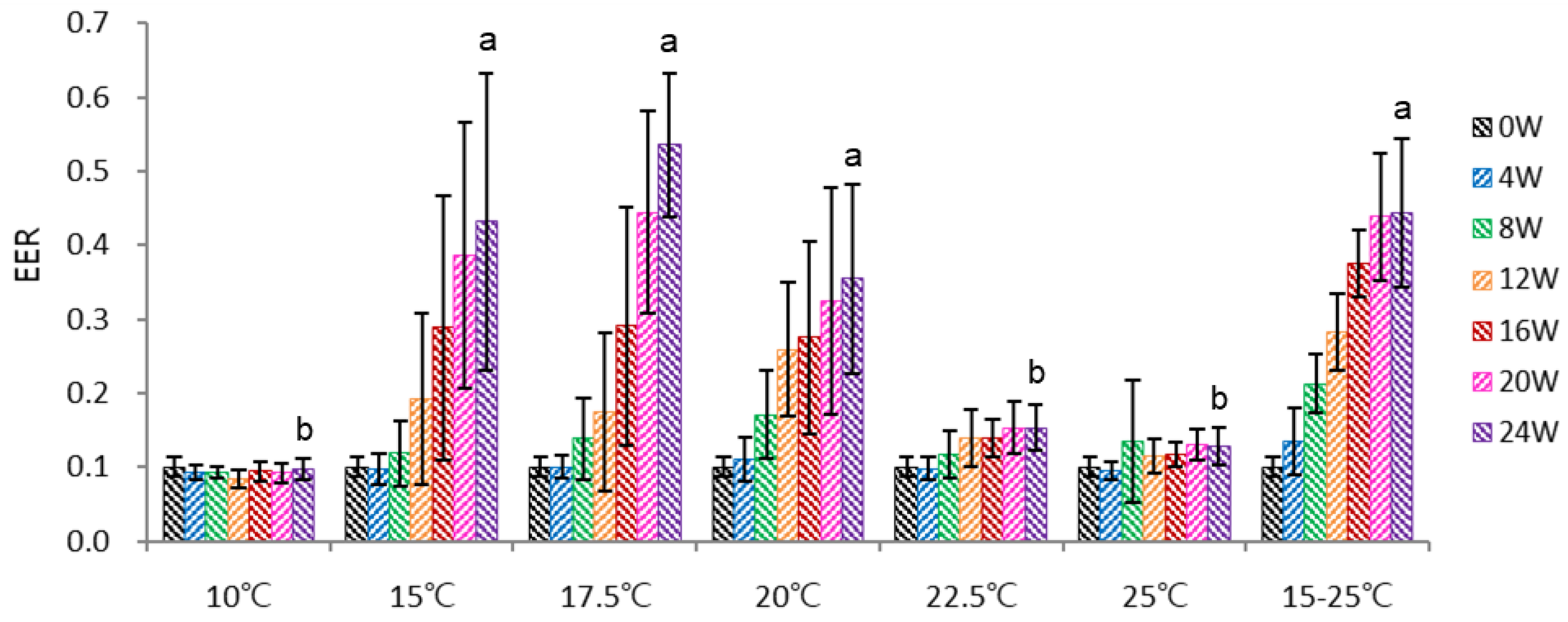
After pulping, un-dehisced ginseng seeds were incubated in the growth chamber at 10-25°C for 24 weeks and watered once a day. The embryo size is less than 0.4 mm, and the embryo-to-endosperm length ratio (EER) was 0.11 at the harvest stage.
The warm stratification temperature critically affected embryo development. The optimum temperature for embryo growth was 15-20°C, with the highest growth at 17.5°C; below (10°C) or above (22.5°C, 25°C) this temperature, embryo growth was highly retarded (Figure 2). At 15°C and 17.5°C, EER after 16-24 weeks increased under the increase in dehiscence (data not shown).
The temperature alternating cycle at 15-25 °C (20°C, 8h→25°C, 4h→20°C, 8h→15 °C, 4h) was beneficial for embryo growth at the early stage (4-16 weeks); thus, EER reached 0.38 after 16 weeks. However, afterward, the EER of alternating temperature of 15-25 °C was retarded to 0.44 at 24 weeks. This retardation was partly on account of a lower dehiscence percentage compared to the 15-20°C. It is likely that fluctuating temperature was not beneficial for microbial growth and eventually lowered dehiscence, resulting in slower embryo growth. In contrast, rapid embryo growth was observed at 17.5 °C, and the EER reached 0.54 at 24 weeks due to a higher dehiscence percentage benefit.
Overall, embryo growth in the non-ventilated chambers was slower than in the case of an outdoor traditional warm stratification facility. From the additional side experiments, I identified aeration (oxygen supply) in the chamber was crucial for embryo development. For instance, dipping the seeds in shallow water retarded embryo growth, but forced aeration in the controlled room facilitated embryo growth (data not provided).
Figure 2.
Effect of temperature on ginseng seeds’ embryo-to-endosperm length ratio (EER) during the warm stratification. *15-25°C, temperature fluctuation cycle of 20°C for 8 h→25°C for 4 h→20°C for 8 h→15°C for 4 h.
Figure 2.
Effect of temperature on ginseng seeds’ embryo-to-endosperm length ratio (EER) during the warm stratification. *15-25°C, temperature fluctuation cycle of 20°C for 8 h→25°C for 4 h→20°C for 8 h→15°C for 4 h.
3.1.2. Effect of Dehiscence Material, Fungi Inoculation, and Watering Period
- Embryo-to-Endosperm Length Ratio (EER)
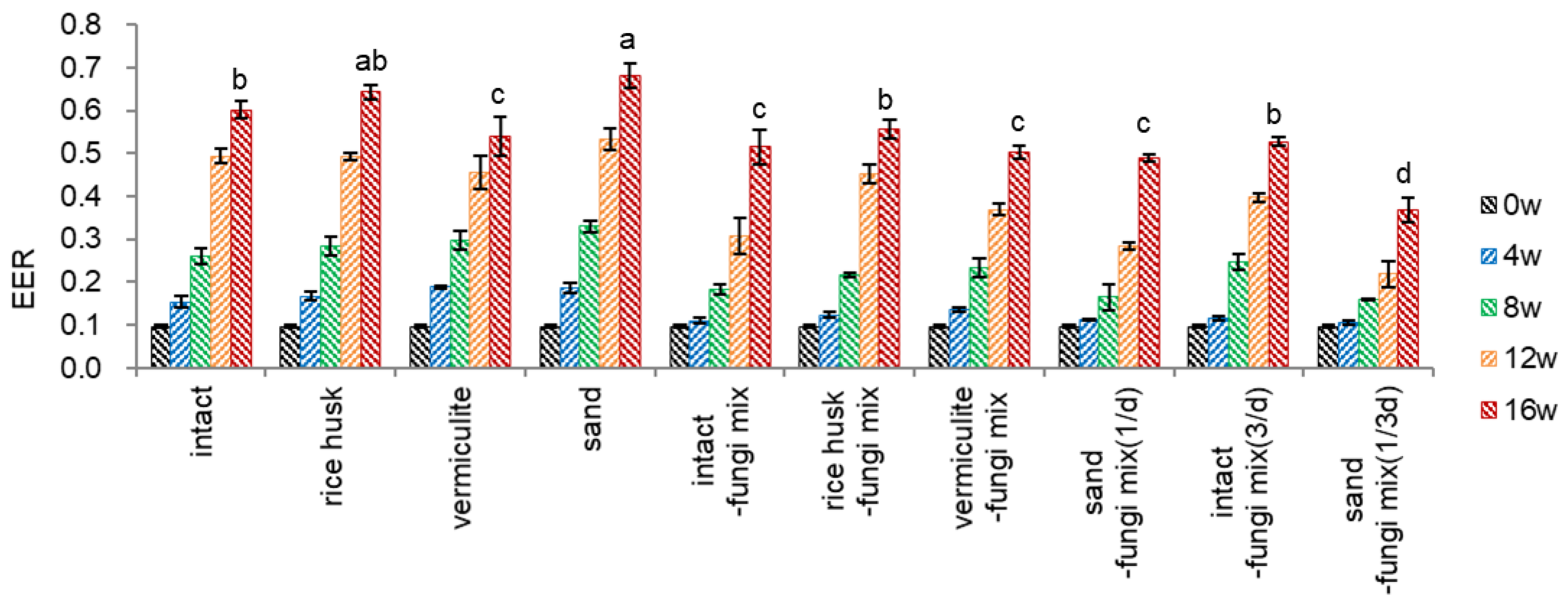
I selected effective dehiscence materials (stratification substrates) to increase the dehiscence (endocarp opening) since dehiscence is a prerequisite for germinating ginseng seeds. Un-dehisced ginseng seeds mixed with dehiscence materials were inoculated with fungi mix and incubated in a growth room at 18-20°C for 16 weeks with watering once a day.
During the warm stratification using dehiscence materials, embryos free of dehiscence material (intact) grew well comparably to rice husk and vermiculite, and the EER reached 0.60 at 16 weeks (Figure 3-1). Sand effectively stimulated embryo growth among the dehiscence materials tested; in contrast, vermiculite was inadequate for embryo growth, implying aeration is critical for embryo growth. The faster embryo growth with sand-treated seeds may relate to the fact that sand provides sufficient ventilation to the seeds, and the surface of sand holds more microbial organisms, which plays a pivotal role in endocarp opening. Moreover, faster embryo growth with sand-treated seeds may be linked to a higher dehiscence percentage than other conditions. In Figure 3-2, the dehiscence of sand-treated seeds started at 8 weeks (23.3%) and reached 69.5% at 12 weeks since dehisced seeds may facilitate water supply and aeration and permit more expansive space for embryo growth.
The surface of the endocarp treated with fungi (Figure 5-3) was darker than that of non-inoculated seeds. Inoculation of fungi on the surface of seed endocarps slightly inhibited embryo growth (EER) in dehiscence material treatments compared to the non-inoculated conditions at 16 weeks (0.54~0.64 → 0.49~0.56). Especially in sand treatment, inoculation of fungi inhibited embryo growth at 16 weeks (0.68 → 0.49). Despite a variable EER range of 0.29-0.45 at 12 weeks, the embryo growth reached a stable EER range of 0.49-0.56 at 16 weeks
Watering three times a day for seeds without dehiscence material and fungi inoculated {intact-fungi mix(3/d)} significantly stimulated embryo growth at 8-12 weeks compared to once a day (intact-fungi mix), but no difference at 16 weeks. Significant inhibition of embryo growth was observed in watering once in three days for sand-treated seeds and fungi-inoculated {sand-fungi mix(1/3d)}, compared to once a day (sand-mix) (0.49→0.37).
Figure 3.
Effect of dehiscence material, fungi inoculation, and watering period on embryo to endosperm length ratio (EER) of un-dehisced ginseng seeds during the warm-stratification.
Figure 3.
Effect of dehiscence material, fungi inoculation, and watering period on embryo to endosperm length ratio (EER) of un-dehisced ginseng seeds during the warm-stratification.
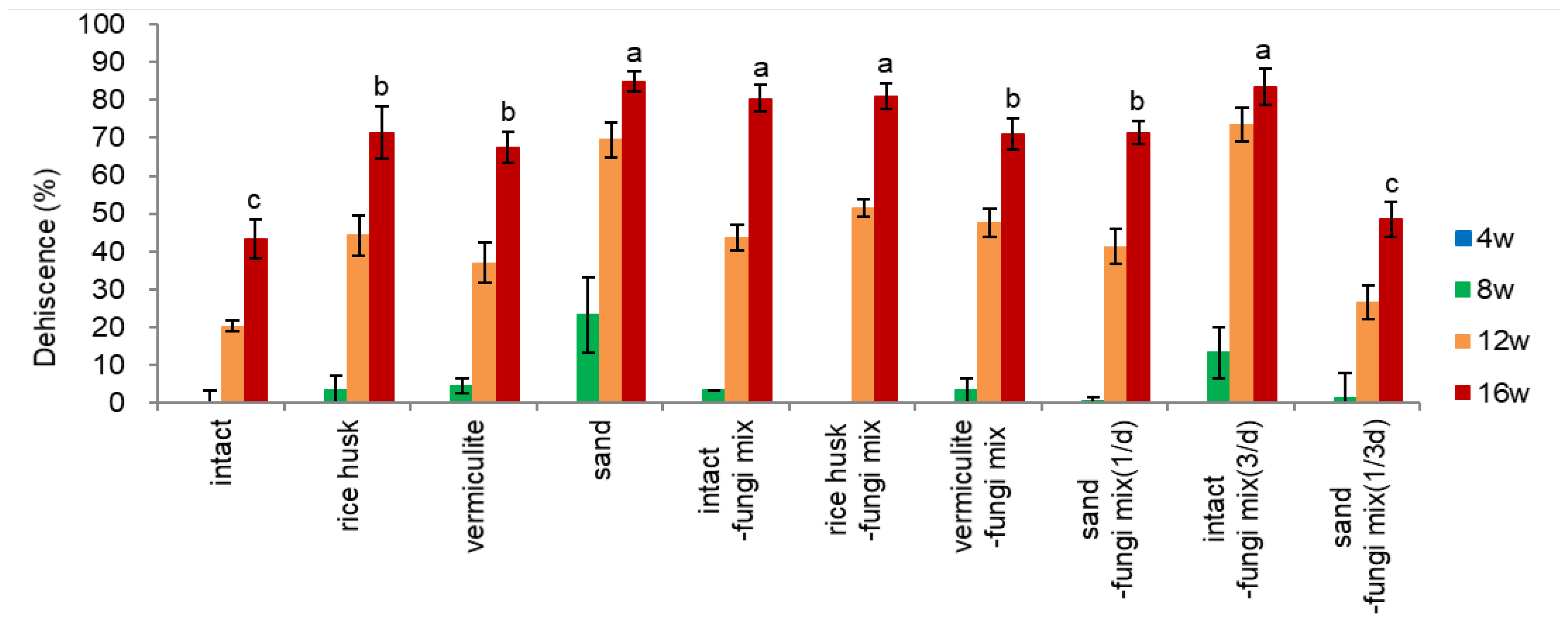
- Dehiscence Percentage
The dehiscence materials (rice husk, vermiculite, sand) were effective in increasing the dehiscence percentage of ginseng seeds over non-treated (intact) seeds (at 16 weeks, 67.5-85.0% vs. 43.3% in Figure 3-2). Among the dehiscence materials, sand was the most effective at 8-12 weeks and, finally, the highest dehiscence at 16 weeks (85%), followed by rice husk (71.5%) and vermiculite (67.5%). Sand may facilitate drainage and provide aeration.
Inoculation of fungi with non-material treated (intact-fungi mix) seeds drastically increased the dehiscence percentage from 43.3% to 80.5% after 16 weeks (Figure 3-2), which is comparable to the dehiscence percentage of dehiscence material treated and fungi inoculated seeds (67.5-85.0%).
The inoculation of fungi with seeds of dehiscence materials was not so effective on dehiscence; only a slight increase of dehiscence was observed in rice husk treated (rice husk-fungi mix, 71.5→81.0%) and vermiculite treated (vermiculite-fungi mix, 67.5→71.0%) seeds, but even significantly decrease in sand treated (sand-fungi mix) seeds (85.0%→71.5%). The effect of dehiscence materials during warm stratification is primarily related to providing a beneficial environment for fungi growth (watering and aeration) and opening of endocarp rather than embryo growth per se.
The fungi inoculated may break down the fibrin near the endocarp hole (hilum) and promote endocarp dehiscence. There is also an interaction between embryo growth and dehiscence percentage since the embryo growth and expansion of endosperm stimulated endocarp opening and vice versa by providing aeration and moisture in the opened endocarp.
Watering three times a day {intact-fungi mix(3/d)} improved embryo growth and dehiscence at 8-12 weeks, compared to once in a day {intact-fungal mix(1/d)}: at 12 weeks EER 0.31→0.40, dehiscence 43.5%→73.5%. But, at 16 weeks, there were no significant effects on EER (Figure 3-1) or dehiscence percentage (Figure 3-2). Watering once in three days {sand-fungi mix(1/3d)} decreased embryo growth (Figure 3-1) and dehiscence percentage (Figure 3-2) compared to watering daily {sand-fungi mix(1/d)}. In this treatment, the surface of the endocarp turned darker than in other treatments, indicating that the higher fungi population induced by sparse watering inhibited embryo growth and finally decreased dehiscence.
This result implies that the dehiscence of ginseng seed is modulated by the interaction of embryo growth and microbial (fungal) decomposing action.
Figure 3.
Effect of dehiscence material and fungi inoculation on dehiscence percentage of un-dehisced ginseng seeds during the warm-stratification.
Figure 3.
Effect of dehiscence material and fungi inoculation on dehiscence percentage of un-dehisced ginseng seeds during the warm-stratification.
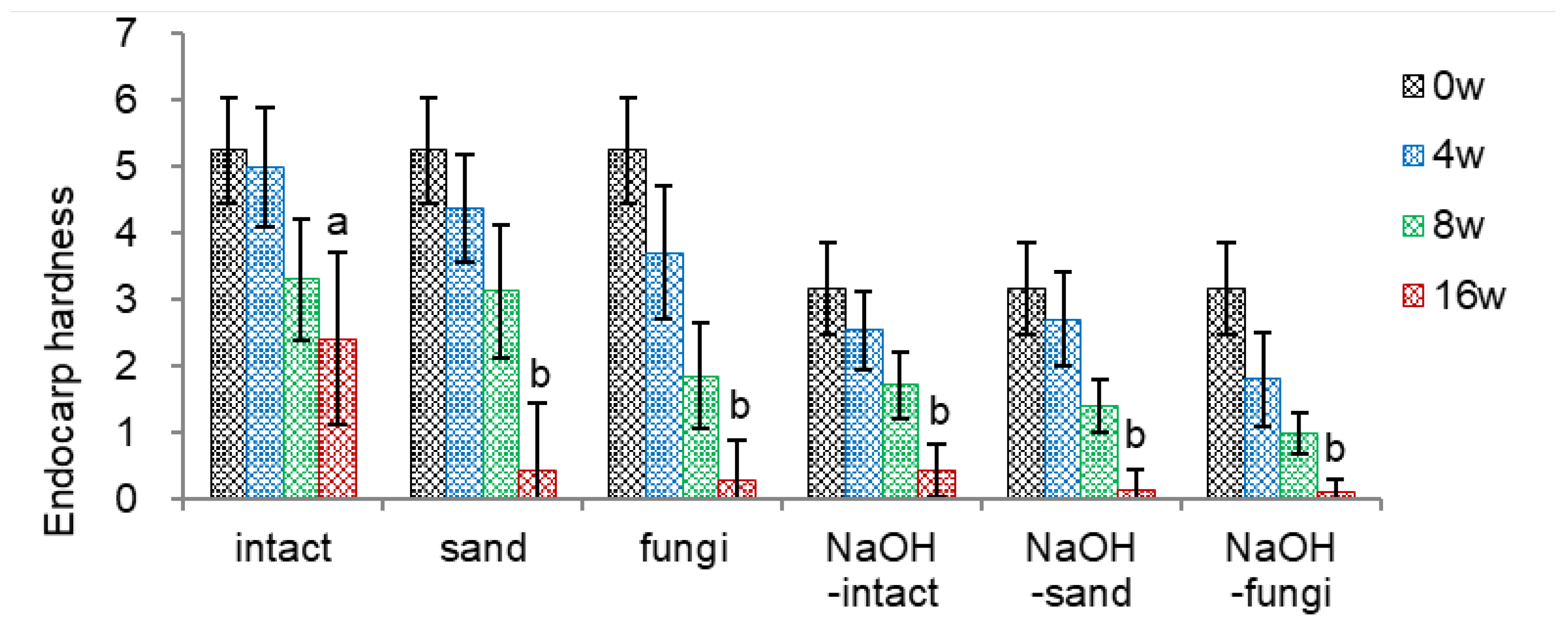
3.1.3. Effect of Sand, Fungi Inoculation, and NaOH
- Endocarp Hardness
The endocarp hardness of intact seeds decreased from 5.25 to 2.41 at 16 weeks of warm stratification. Inoculating fungi mix before the warm stratification was the most effective in reducing the endocarp hardness (0.27 at 16 weeks), followed by applying sand as dehiscence material (0.43 at 16 weeks) (Figure 4-1).
To facilitate the endocarp opening by decreasing the endocarp hardness, I soaked un-dehisced seeds in sodium hydroxide (NaOH) since sulfonic acid (H2SO4) was ineffective in reducing the endocarp hardness. Based on the preliminary screening, un-dehisced seeds were soaked in 5% NaOH for 25 min, which decreased the endocarp hardness from 5.25 (intact) to 3.16 (NaOH-intact). NaOH-soaking reduces the endocarp hardness, reaching 0.09-0.43 at 16 weeks.
Combining NaOH-soaking with inoculating fungi mix (NaOH-fungi) was the most effective in decreasing endocarp hardness (0.99 and 0.09 at 8 and 16 weeks, respectively) among the treatments tested.
Figure 4.
Effect of NaOH and fungi inoculation on endocarp hardness of un-dehisced ginseng seeds during the warm-stratification.
Figure 4.
Effect of NaOH and fungi inoculation on endocarp hardness of un-dehisced ginseng seeds during the warm-stratification.
- Embryo-to-Endosperm Length Ratio (EER)
NaOH soaking facilitated embryo growth at 8 weeks during the warm stratification. However, EER at 16 weeks was not significantly different among the conditions (Figure 4-2).
Figure 4.
Effect of NaOH and fungi inoculation on embryo growth (EER) of un-dehisced ginseng seeds during the warm-stratification.
Figure 4.
Effect of NaOH and fungi inoculation on embryo growth (EER) of un-dehisced ginseng seeds during the warm-stratification.
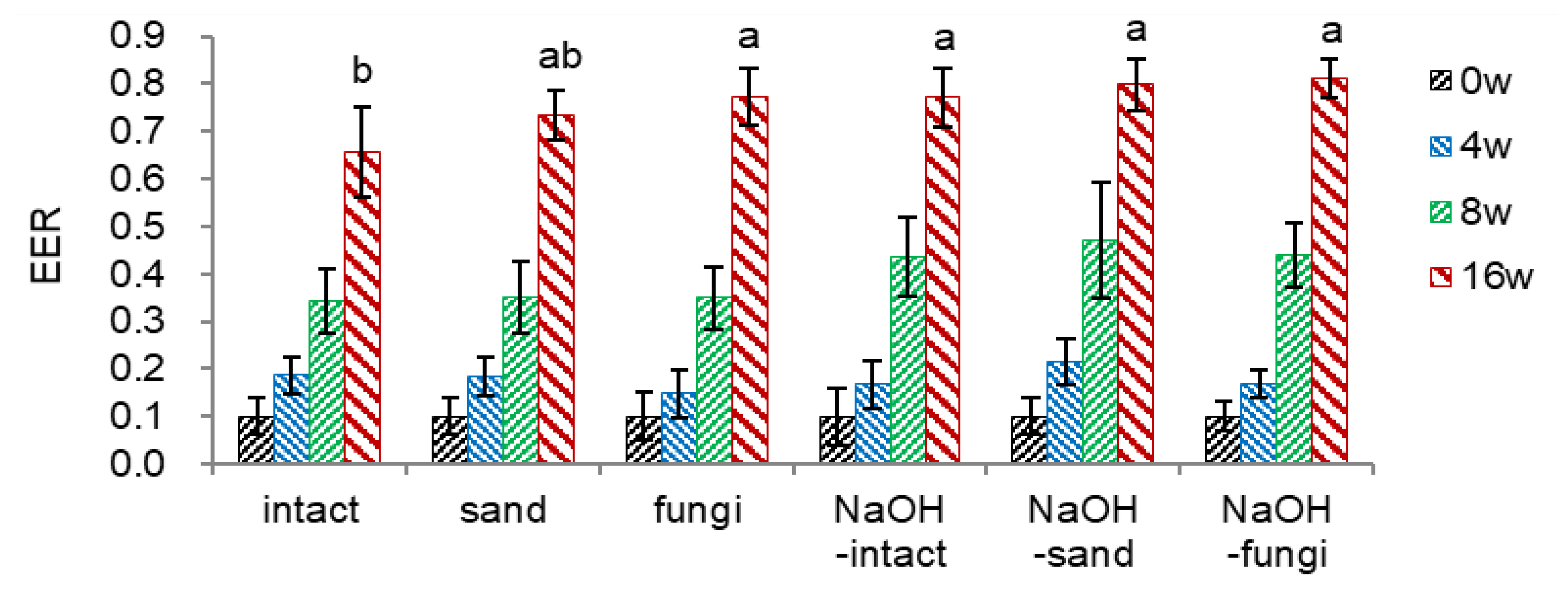
- Dehiscence Percentage
Inoculating fungi before warm stratification showed the fastest and the highest dehiscence percentage (90.3% at 16 weeks) compared to the intact seeds (28.8%) (Figure 4-3). Mixing with sand as a dehiscence material also facilitated the dehiscence.
Soaking intact seeds with NaOH (NaOH-intact) increased the dehiscence percentage by 18% compared to intact seeds. However, soaking the un-dehisced seeds with NaOH and then applying sand as a dehiscence material (NaOH-sand) or inoculating fungi (NaOH-fungi) negatively affected the dehiscence percentage, i.e., about a 15% decrease at 16 weeks. Although soaking the seeds with NaOH effectively reduced endocarp hardness, it sterilized the seeds’ surface and thus negatively affected the dehiscence percentage, implying a crucial role of fungi in endocarp opening. Therefore, inoculating fungi with un-dehisced seeds or mixing with sand before warm stratification is a prerequisite for dehiscencing ginseng seeds.
Figure 4.
Effect of NaOH and fungi inoculation on dehiscence percentage of un-dehisced ginseng seeds during the warm-stratification.
Figure 4.
Effect of NaOH and fungi inoculation on dehiscence percentage of un-dehisced ginseng seeds during the warm-stratification.
3.1.4. Combinational Effect of Fngi Inoculation and Sterilization of Unhehisced Seeds
- Embryo-to-Endosperm Length Ratio (EER)
I inoculated a mixture of fungi with non-sterilized (intact) or sterilized ginseng seeds to confirm the effect of fungi for dehiscence. With these four conditions, embryo growth was not significantly different (Figure 5-1).
Figure 5.
Effect of surface sterilization of seeds and inoculation of fungi on embryo-to-endosperm length ratio (EER) of un-dehisced ginseng seeds during warm stratification.
Figure 5.
Effect of surface sterilization of seeds and inoculation of fungi on embryo-to-endosperm length ratio (EER) of un-dehisced ginseng seeds during warm stratification.
- Dehiscence
Inoculation of fungi was very beneficial for the dehiscence of intact ginseng seeds without dehiscence materials (20.6% vs. 83.0%, Figure 5-2). The dehiscence percentage of surface sterilized seeds without fungi inoculation was 31.5%, and that of fungi inoculation was 66.7% after 16 weeks (Figure 5-2). Sterilization of the seed surface before warm stratification negatively affected fungi inoculation (growth) on the seed surface and negatively impacted endocarp opening.
Figure 5.
Effect of surface sterilization of seeds and inoculation of fungi on dehiscence of un-dehisced ginseng seeds during warm stratification.
Figure 5.
Effect of surface sterilization of seeds and inoculation of fungi on dehiscence of un-dehisced ginseng seeds during warm stratification.
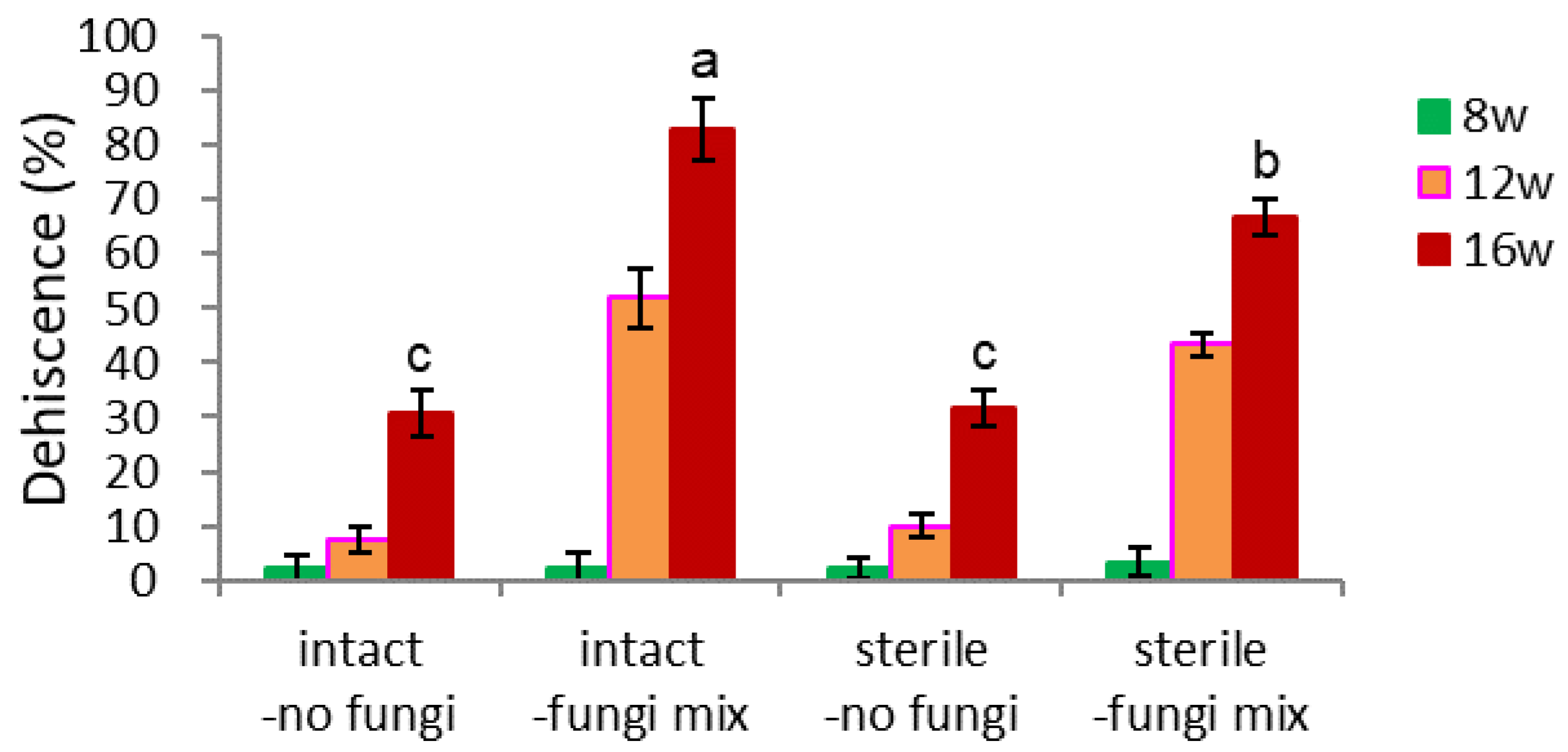
The surface area of non-sterilized and fungi-inoculated (intact-fungi mix) seeds turned dark brown during the warm stratification. Sterilizing the seed’s surface and later fungi inoculation (sterile-fungi mix) also turned the endocarp dark brown (Figure 5-3-A). In a broad sense, the endocarp color of the dehisced (opened) seeds was often light black, while that of the un-dehisced seeds was often light brown.
Figure 5.
Prior fungi inoculation turned the endocarp surface dark brown during the warm stratification. A; The combinational treatments of no-sterile(intact)/sterile and no-fungi/fungi-inoculated make the endocarp surface different after 16 weeks. upper, non-sterilized; below, sterilized with NaOCl; left, non-fungi inoculated; right, fungi inoculated. B-C; No-fungi inoculated ginseng seeds mixed with sand as a substrate kept light yellowish brown at 4 weeks (B). In contrast, fungi-inoculated ginseng seeds without sand turned brown at 4 weeks (C).
Figure 5.
Prior fungi inoculation turned the endocarp surface dark brown during the warm stratification. A; The combinational treatments of no-sterile(intact)/sterile and no-fungi/fungi-inoculated make the endocarp surface different after 16 weeks. upper, non-sterilized; below, sterilized with NaOCl; left, non-fungi inoculated; right, fungi inoculated. B-C; No-fungi inoculated ginseng seeds mixed with sand as a substrate kept light yellowish brown at 4 weeks (B). In contrast, fungi-inoculated ginseng seeds without sand turned brown at 4 weeks (C).

3.1.5. Inoculation of Fungi, Actinomycetes, and Bacteria (Bacillus) Mix
- Endocarp Hardness, EER, and Dehiscence
To compare the efficiency of microorganisms on the dehiscence of ginseng seed, a mixture of fungi, actinomycetes, and bacteria was inoculated with the seeds. The inoculation of these three microbial mixtures fastened the decrease in endocarp hardness (Figure 6-1) and had no significant effect on EER (Figure 6-2). All the inoculation treatments significantly increased the dehiscence percentage (Figure 6-3) compared to the control seeds (75.2~79.9% vs. 47.2%). However, there was no significant difference among the three microbial mixtures.
Figure 6.
Effect of fungi, antinomycetes, and bacteria inoculation on endocarp hardness of un-dehisced ginseng seeds during the warm-stratification.
Figure 6.
Effect of fungi, antinomycetes, and bacteria inoculation on endocarp hardness of un-dehisced ginseng seeds during the warm-stratification.
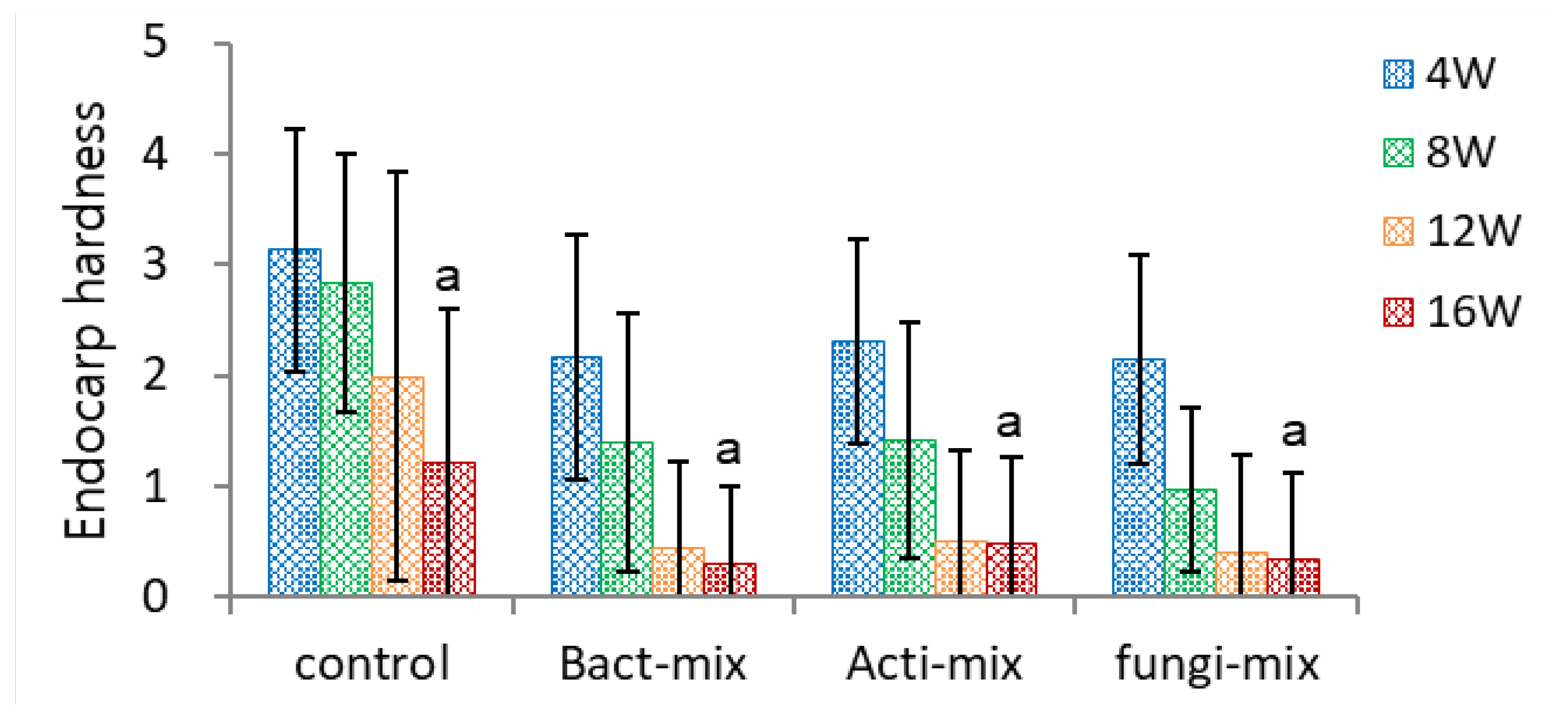
Figure 6.
Effect of fungi, antinomycetes, and bacteria inoculation on embryo-to-endosperm ratio (EER) of un-dehisced ginseng seeds during the warm-stratification.
Figure 6.
Effect of fungi, antinomycetes, and bacteria inoculation on embryo-to-endosperm ratio (EER) of un-dehisced ginseng seeds during the warm-stratification.
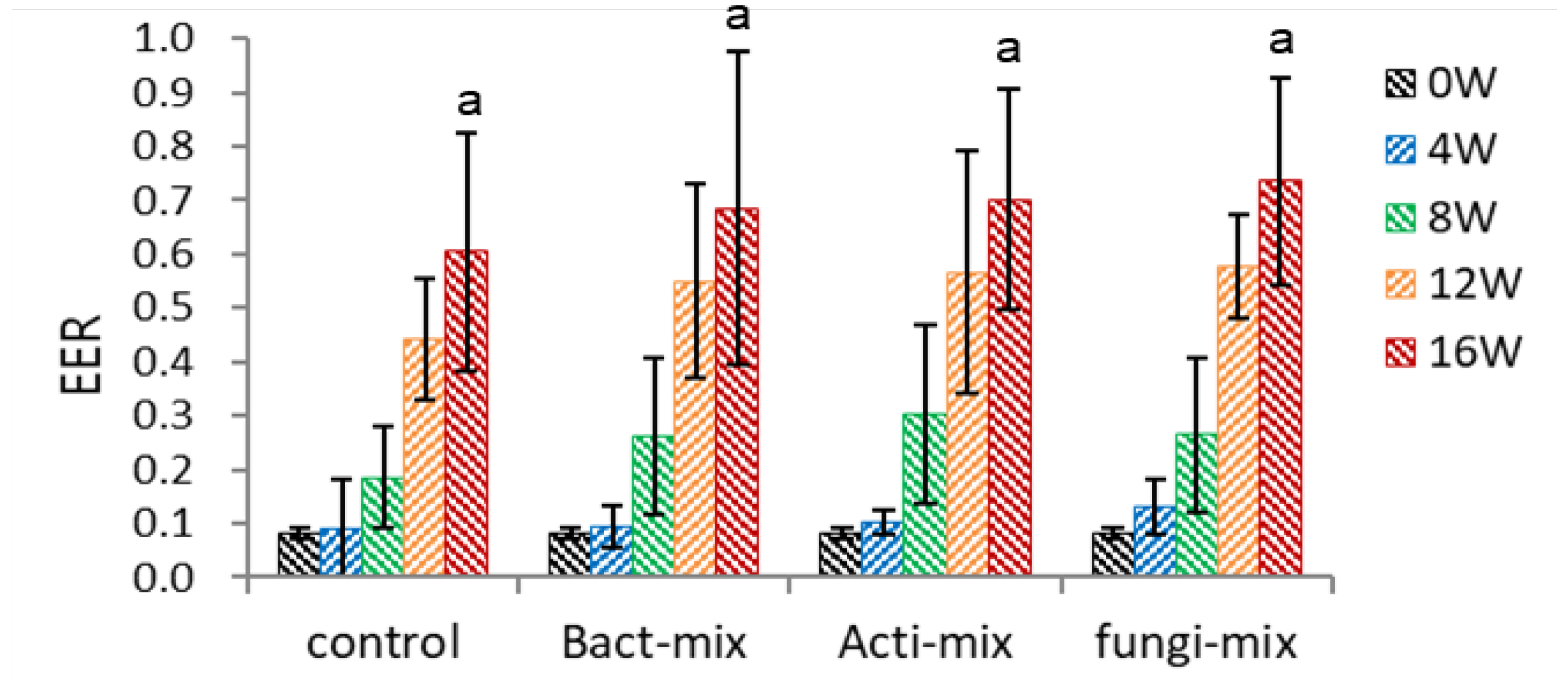
Figure 6.
Effect of fungi, antinomycetes, and bacteria inoculation on dehiscence of un-dehisced ginseng seeds during the warm-stratification.
Figure 6.
Effect of fungi, antinomycetes, and bacteria inoculation on dehiscence of un-dehisced ginseng seeds during the warm-stratification.
3.1.6. Inoculation of Fungi, Actinomycetes, and Bacteria with High or Low Cellulose Dissociation Activity
- Endocarp Hardness
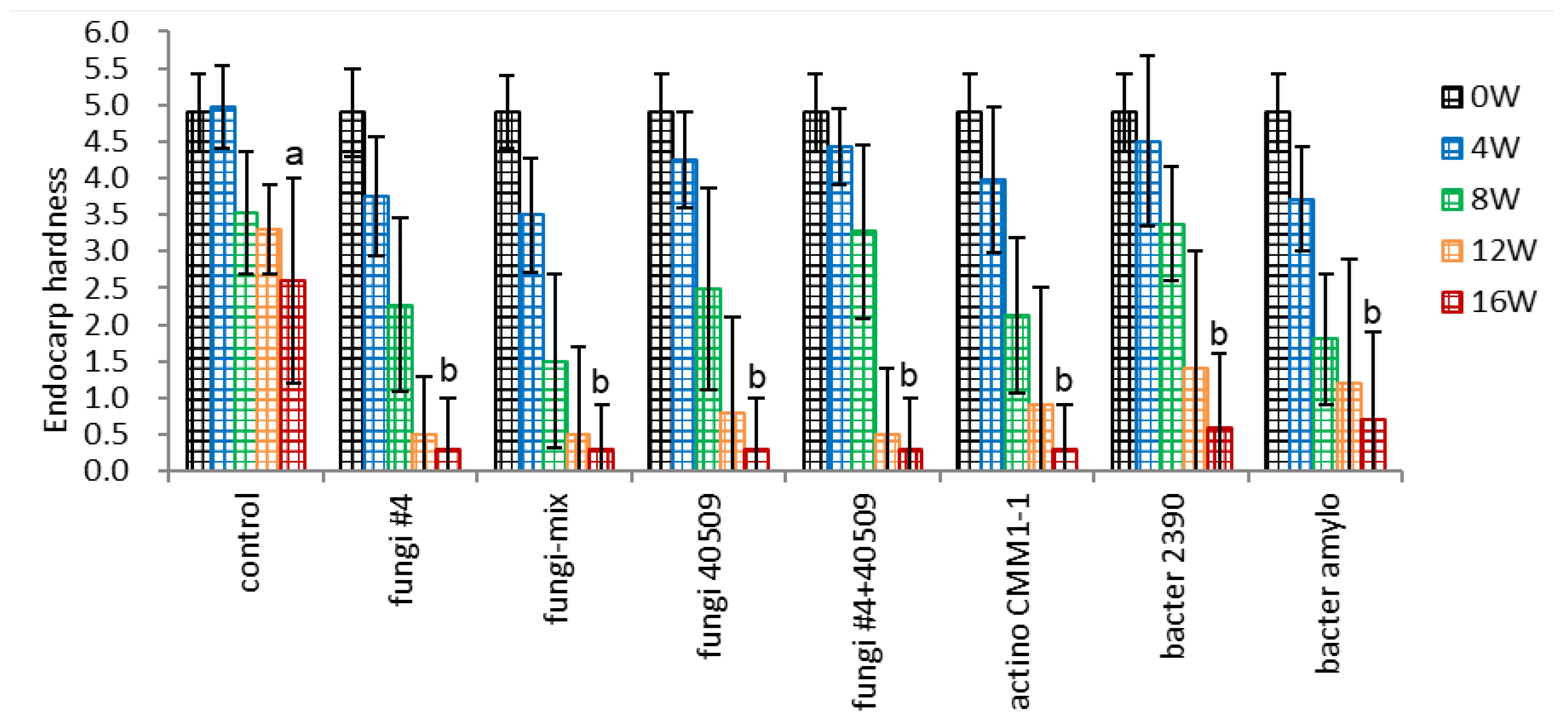
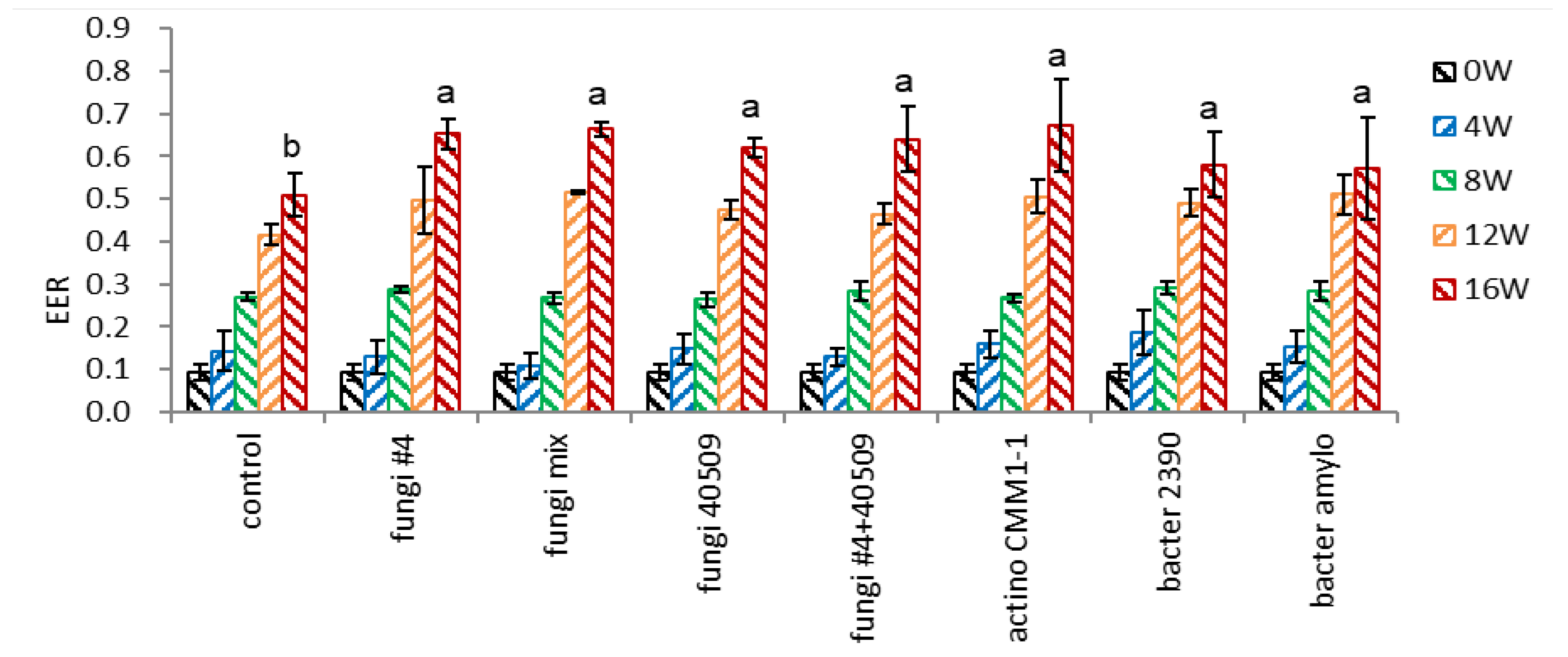
The seeds were inoculated with a broader range of microorganisms with high or low cellulose dissociation activity to validate the effect of microbial inoculation. All the bacterial inoculation treatments tested, including fungi, actinomycetes, and bacteria, significantly decreased the endocarp hardness, and there were no significant differences among the three groups tested (Figure 7-1). Interestingly, no significant difference was observed in endocarp hardness between the strains of high cellulose dissociation activity in the field (fungi #4) or in vitro assay (actino CMM1-1, bacter 2390) and two strains of low cellulose dissociation activity in in vitro assay (fungi 40509, bacter amylo). The mixture of four fungi strains (fungi-mix) more effectively decreased the endocarp hardness after 8 weeks (1.50 vs. control 3.54).
Figure 7.
Effect of microbial inoculation on endocarp hardness of un-dehisced ginseng seeds during the warm-stratification.
Figure 7.
Effect of microbial inoculation on endocarp hardness of un-dehisced ginseng seeds during the warm-stratification.
- Embryo-to-Endosperm Length Ratio (EER)
The EER of intact (control) seeds linearly increased, and there was no difference in the EER of control and microbial-inoculated seeds during 0-8 weeks (Figure 7-2). However, the EER in microbial inoculated seeds rose rapidly, compared to the non-inoculated seeds, during 12-16 weeks. This rapid embryo growth at 12-16 weeks coincides with the rapid increase of dehiscence percentage in microbial inoculated seeds during this period (Figure 10-3). There was no difference in the EER among the microbial-inoculated seeds, implying the cellulose dissociation activity of microorganisms may not be specific to decomposing the seed endocarp.
Figure 7.
Effect of microbial inoculation on EER of un-dehisced ginseng seeds during the warm-stratification.
Figure 7.
Effect of microbial inoculation on EER of un-dehisced ginseng seeds during the warm-stratification.
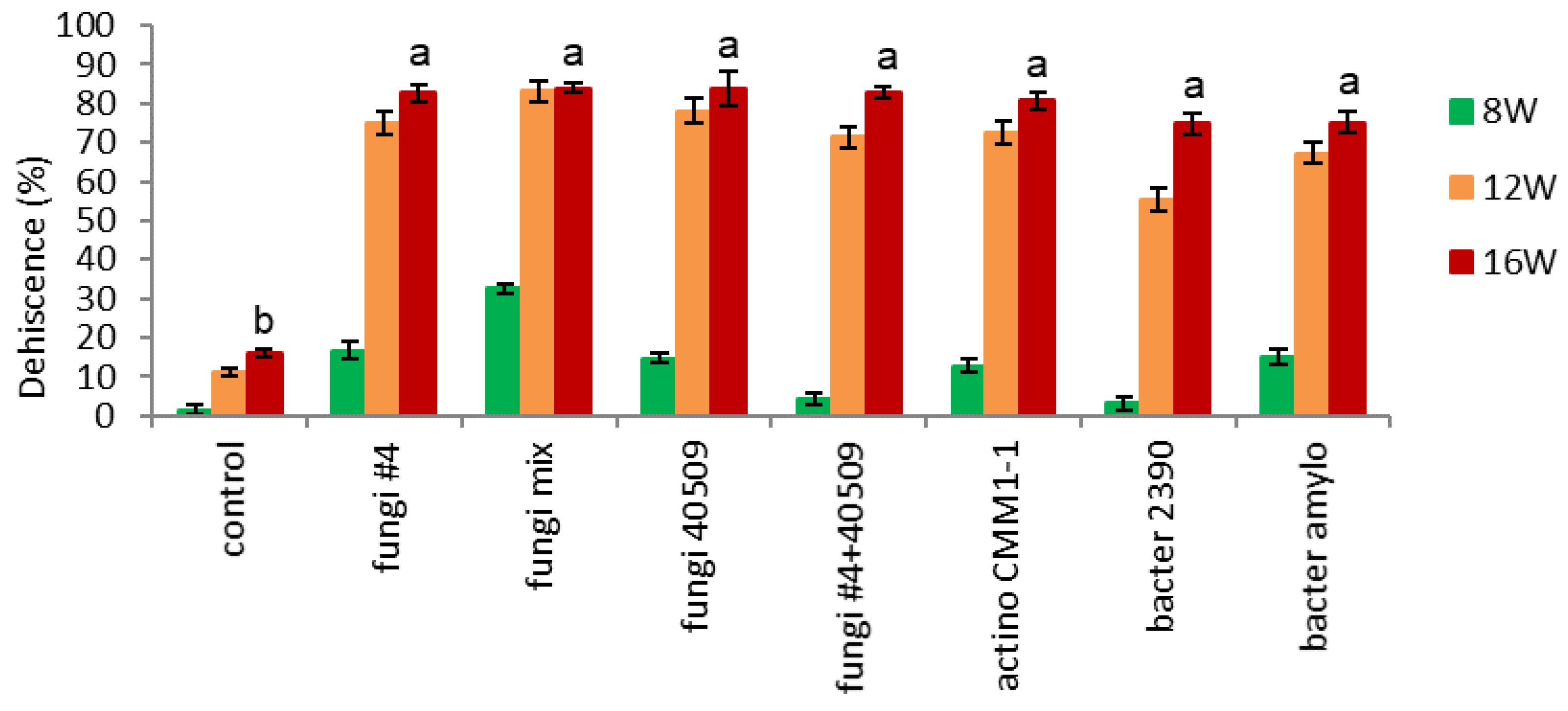
- Dehiscence.
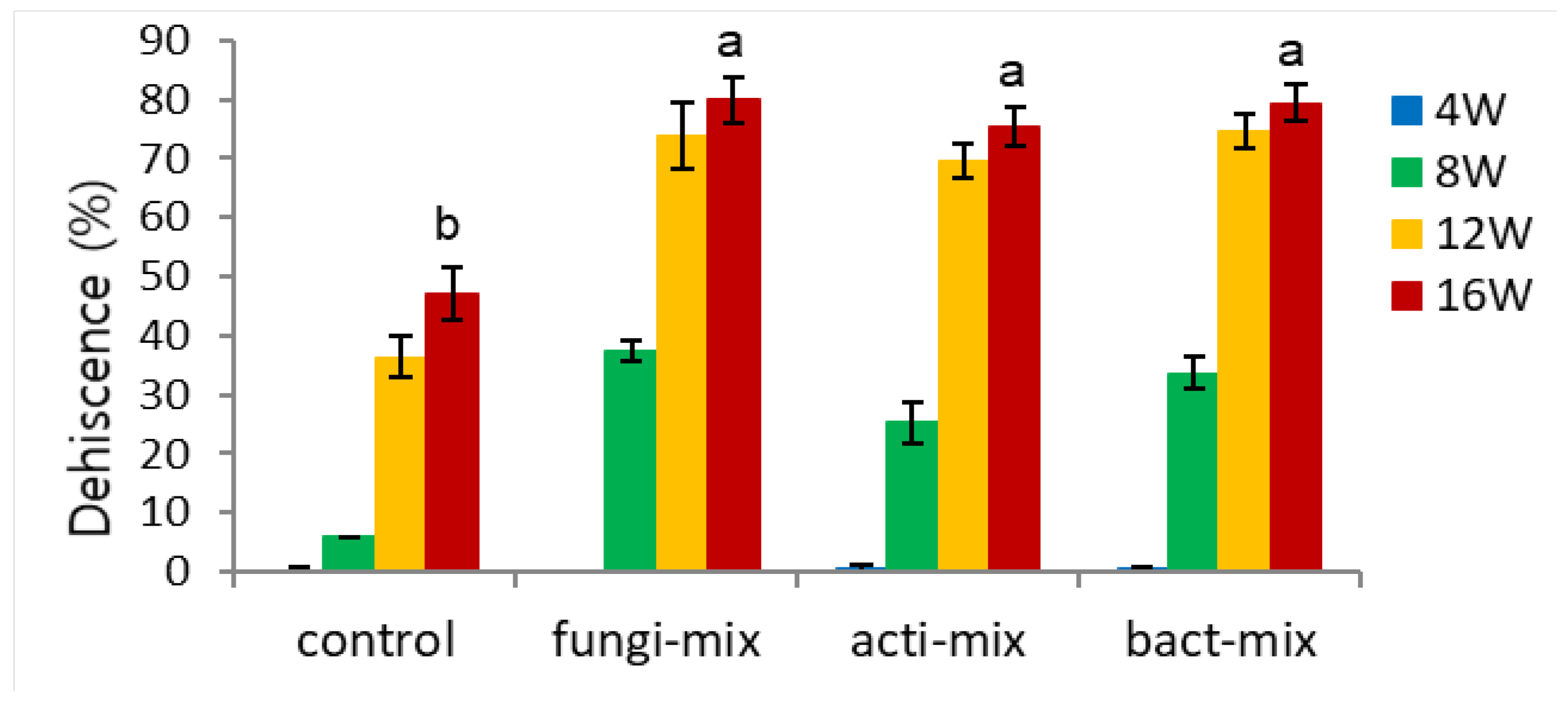
Endocarp opening (dehiscence) started after 8 weeks, particularly in inoculation with a mixture of four fungi strains (fungi-mix 32% vs. other treatments 2-17%) (Figure 7-3). This result coincides with Figure 7-1, which showed a sharp decrease in endocarp hardness after 8 weeks. Dehiscence rapidly increased in all the microbial-inoculated seeds during 12 weeks (67-83%), except bacteria-inoculated seeds (bacter 2390), which showed relatively lower dehiscence (56%) (Figure 7-3). After 16 weeks, all the microbial inoculated seeds resulted in 60% higher dehiscence than the control (75-83 vs. 17%).
Figure 7.
Effect of microbial infection on dehiscence of un-dehisced ginseng seeds during the warm-stratification.
Figure 7.
Effect of microbial infection on dehiscence of un-dehisced ginseng seeds during the warm-stratification.
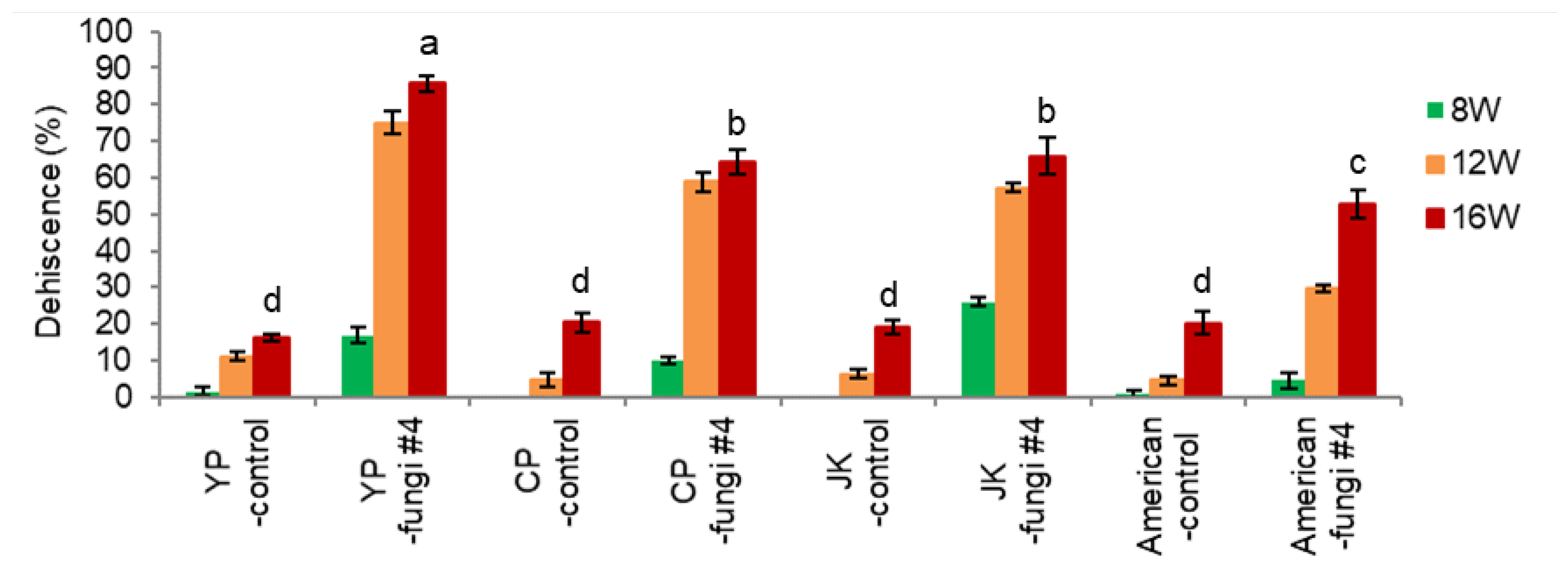
3.2. Application of Fungi Inoculation to three Varieties of Ginseng and One American Ginseng
- Endocarp Hardness
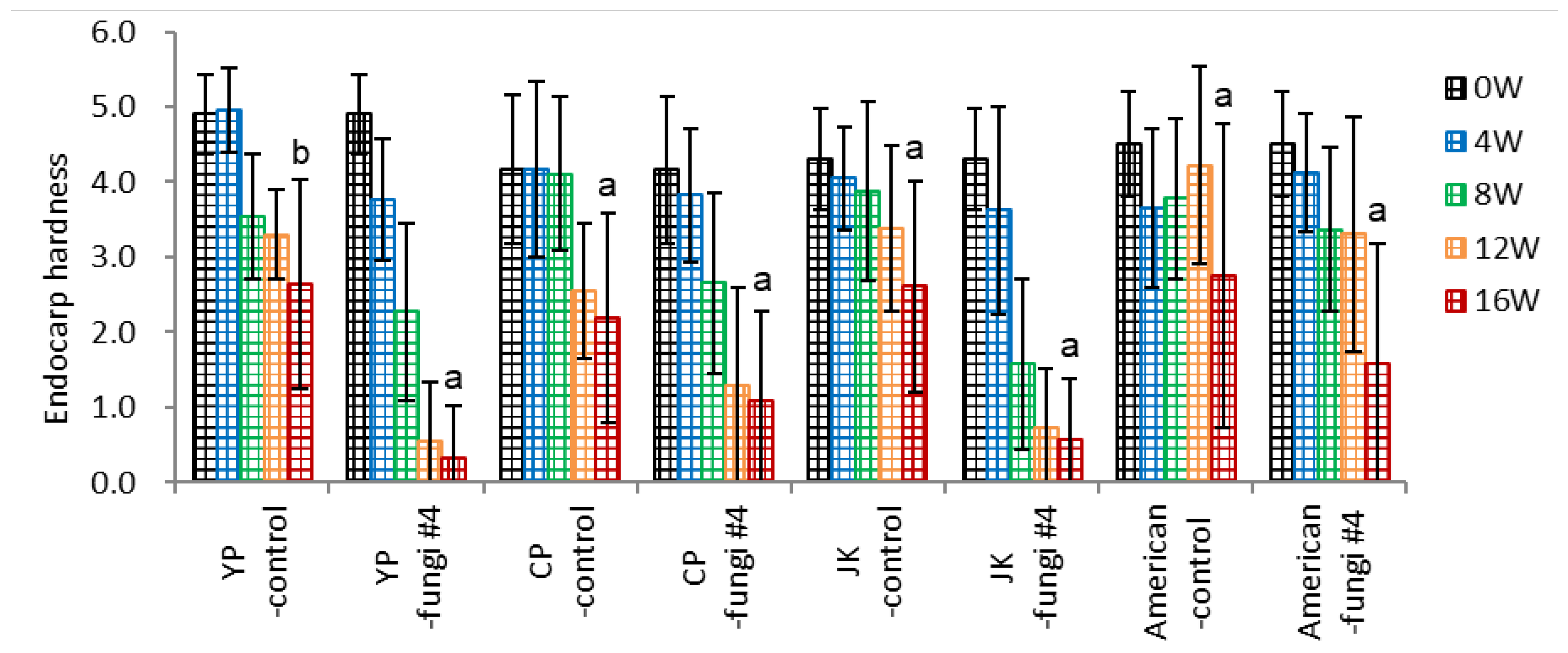
Three varieties of P. ginseng seeds (one landrace ‘JK’, two varieties of ‘YP’ and ‘CP’) and P. quinquefolius seeds with (-fungi #4) or without (-control) fungi inoculation were subject to warm stratification in pots.
Inoculation of fungi facilitated a decline in endocarp hardness in ‘YP’, ‘JP’ varieties of P. ginseng seeds (2.3 and 2.0, respectively) (Figure 8-1). The drop of endocarp hardness in ‘CP’ and P. quinquefolius (American) seeds (1.1 and 1.2, respectively) was slower than the former, which is related to the lower endocarp opening (dehiscence percentage).
Figure 8.
Effect of ginseng variety on endocarp hardness of un-dehisced ginseng seeds with or without fungi inoculation during the warm-stratification.
Figure 8.
Effect of ginseng variety on endocarp hardness of un-dehisced ginseng seeds with or without fungi inoculation during the warm-stratification.
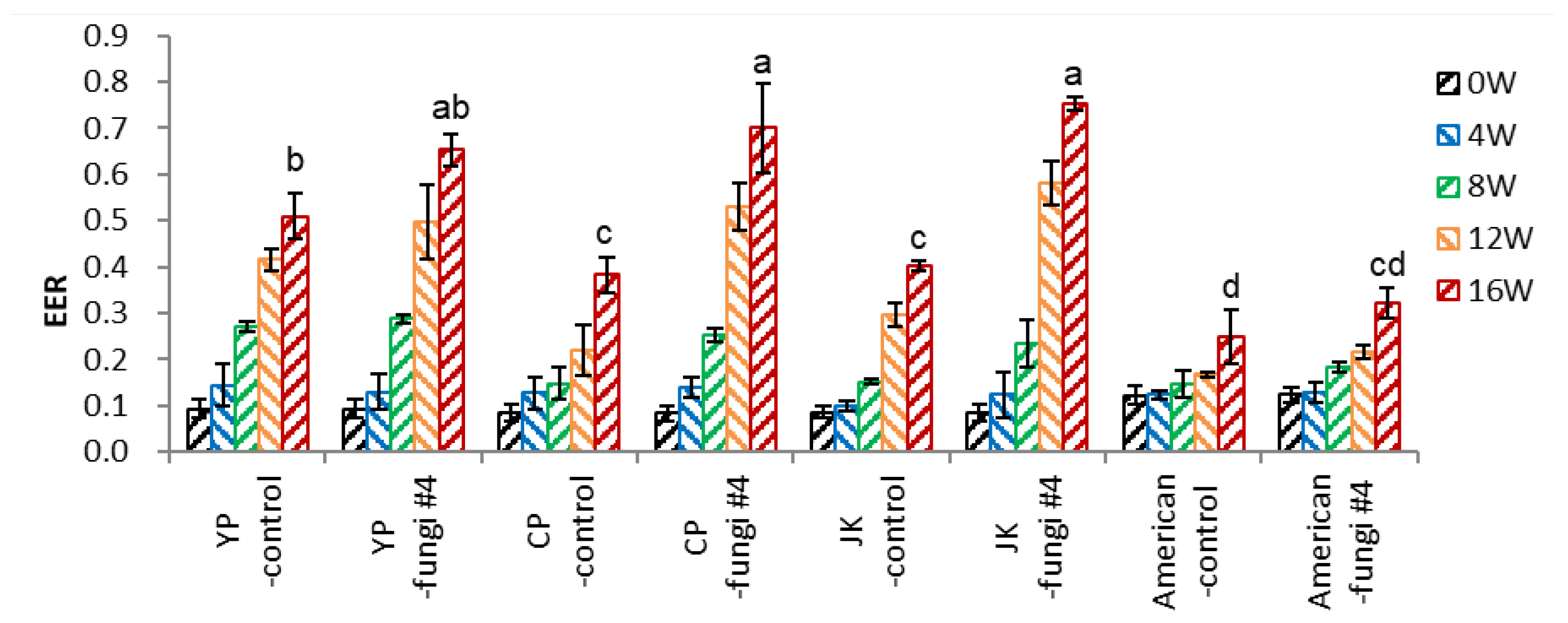
- Embryo-to-Endosperm Length Ratio (EER)
Inoculation of fungi on seeds of P. ginseng significantly increased the EER at 16 weeks; ‘YP’ 0.51 → 0.65, ‘CP’ 0.38→ 0.70, ‘JK’ 0.40→ 0.75 (Figure 8-2). The significant embryo growth at 8-16 weeks was associated with the opening of the seeds (dehiscence), facilitating water and aeration supply. However, fungi inoculation in P. quinquefolius seeds (American-fungi #4) slightly increased the EER (0.25→0.32), reflecting the lower dehiscence (Figure 8-3).
Figure 8.
Effect of ginseng varieties on EER of un-dehisced ginseng seeds with or without fungi inoculation during the warm-stratification.
Figure 8.
Effect of ginseng varieties on EER of un-dehisced ginseng seeds with or without fungi inoculation during the warm-stratification.
- Dehiscence
Intact seeds of P. ginseng (two varieties of ‘YP’ and ‘CP’, one landrace ‘JK’) and P. quinquefolius without fungi inoculation showed lower dehiscence at 16 weeks (15.2-20.5%) (Figure 8-3). Inoculation of fungi (#4) significantly facilitated the dehiscence of P. ginseng seeds (‘YP’ 82.7%, ‘CP’ 53.3%, ‘JK’ 58.8%) and P. quinguefolius seeds (41.5%). Facilitation of endocarp opening in fungi-inoculated seeds was associated with the decline of endocarp hardness by microbial inoculation during the 0-12 weeks, and opening of endocarp accelerates embryo growth.
Figure 8.
Effect of ginseng varieties on dehiscence of un-dehisced ginseng seeds with or without fungi inoculation during the warm-stratification.
Figure 8.
Effect of ginseng varieties on dehiscence of un-dehisced ginseng seeds with or without fungi inoculation during the warm-stratification.
4. Discussion
Ginseng seeds have a short life span, and it takes 3-4 years to seed harvest. After three years of storage at −2°C, the ginseng seed’s dehiscence percentage dropped to 45% from the 84% fresh unstored seeds [14]. The hydration window for cryopreservation of ginseng seeds (exclude endocarp) is variable during the after-ripening stage; un-dehisced seeds (EER 0.1) 4-8%, dehisced (EER 0.45) 7-10%, and fully developed embryos (EER 0.9) 9-11% moisture content [4]. The damaged seeds from desiccation and cryo-injury failed to dehisce. However, dehisced seeds germinated after cold stratification, implying dehiscence is a significant hurdle for germinating ginseng seeds [4].
The moisture content of fresh seeds (embryo and endosperm) increased from 45.9% for un-dehisced seeds after harvest to 58.5% after dehisced and 66.3% for fully developed embryos. It implies that the endocarp hinders moisture imbibition heavily at the un-dehisced stage and slightly at the dehisced stage; the physiological state of seeds varied during the after-ripening stages.
The dormancy of ginseng seeds has been classified as morpho-physiological (MDP) [10,11]. Therefore, the seeds need warm and cold stratification for further embryo development and germination [7,8]. As physical dormancy is defined as water-impermeable layers of palisade cells in the seed or fruit coat [12], ginseng seeds have been considered to have no physical dormancy. However, this study suggests that ginseng seeds have mechanical (physical) dormancy due to the fruit coat (endocarp). The hard endocarp of ginseng seeds has a hole (hilum) and partially allows for the imbibition of water and gas exchange. Therefore, the endocarp can be opened from the hilum through microbial decomposing in the traditional outdoor dehiscence facility conditions in summer and autumn. However, the endocarp may not open indoors during winter, failing to further embryo growth and germination.
Though embryo growth may facilitate the opening of endocarp, endocarp opening is not directly determined by embryo growth. In the summer, the sand used as a dehiscence material (stratification substrate) contains sufficient microorganisms in the traditional farming practice of endocarp opening. During the warm stratification period, the microorganisms are inoculated into the endocarp of the ginseng seed and eat away the fibers of the endocarp, thereby allowing the opening to occur in association with the expansion of the seed embryo and endosperm.
The beneficial effects of sand as a stratification substrate may include 1) providing aeration (oxygen supply) via facilitated drainage, 2) providing fungi inoculation to the endocarp surface, and 3) balancing the inhabitation of microorganisms and after-ripening of the seeds. Oxygen is a pivotal factor for embryo development and the germination of seeds. The high-pressure oxygen favored dormancy breakage and germination of Crataegus mollis seeds [15].
These findings suggest that endocarp dehiscence needs embryo development and microbial-assisted decomposing of the endocarp hole (hilum). Therefore, I proposes a dehiscence model of ginseng seeds at the traditional dehiscence facility during the warm stratification (Figure 10). The three conditions (water, oxygen, and temperature) are essential for stratification and germination of seeds. This study emphasizes the importance of microbial inoculation on the surface area of the endocarp for microbial-assisted decomposing of the endocarp (from hilum to raphae) and, finally, dehiscence (opening of endocarp). Dehiscence is a combinational action of microbial-assisted decomposing of the endocarp and extension of seed embryo and endosperm.
Figure 10.
A dehiscence model of ginseng seeds at the traditional dehiscence facility was used during the warm stratification.
Figure 10.
A dehiscence model of ginseng seeds at the traditional dehiscence facility was used during the warm stratification.
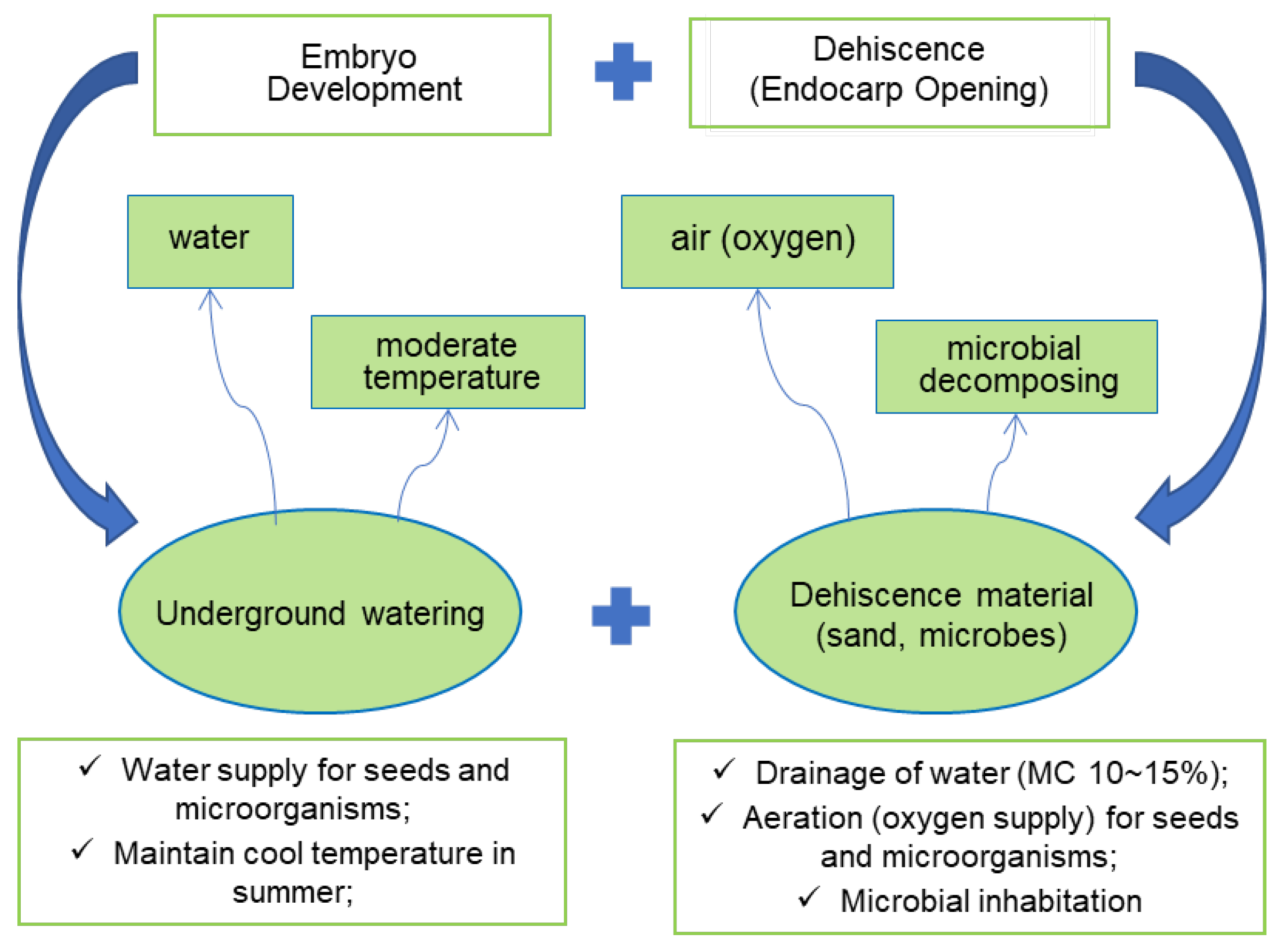
In traditional outdoor warm stratification, sand, as a stratification substrate, may provide a source of fungi inoculum and appropriate conditions for moisturizing and aeration (supplying oxygen). Seed embryos can grow under such conditions, and fungi on the endocarp surface can inhabit them. It is hypothesized that moisture and aeration with appropriate temperature are prerequisites for embryo growth, just like germination conditions. With this condition, fungi can also multiply, and they may decompose the cellulose of the endocarp and finally allow it to open.
During the warm stratification, seed embryos grow with water and oxygen and prefer moderate temperatures. The temperature effect is critical for developing immature embryos, and 17.5°C was the optimum temperature. Contrary to previous studies [8], the constant optimum temperature was superior to alternating temperature for embryo development and germination, reflecting earlier endocarp opening was beneficial for further embryo development. Moreover, the dehiscence of the endocarp facilitates embryo growth at a later stage of warm stratification, implying that the endocarp is a limiting factor for embryo development.
The optimum after-ripening temperature for ginseng seeds is variable during development stages: warming stratification (dehiscence treatment, 100 days) 17°C, middle stage (30 days after dehiscence) 10°C, physiological dormancy at terminal stage (60-90 days) 5°C, germination 15°C [16,17].
Ginseng seeds and endocarp’s abscisic acid (ABA) content gradually decreased, while gibberellic acid (GA3) contents increased during the warm stratification [18]. However, the effect of exogenous treatment of GA3 before the stratification was controversial. Soaking of Eleutherococus senticosus (Araliaceae family) seeds in around 300 mg L−1 GA3 for a day increased the dehiscence percentage (from 58.8% to 80.7%), but the seed decay rate also increased [19,20].
The ginseng seed endocarp is water-permeable through the hilum (Figure 1). It may partly allow respiratory gas exchange, too. However, the endocarp impedes embryo development during the initial warm stratification and prevents further cellular extension of embryo development, failing to germinate. When the endocarp was removed manually by the nail cutter, most of the seeds decayed during the warm stratification (data not shown). It raises the concept that the ginseng seed endocarp works as a seed coat, inhibits embryo development, and protects the seeds.
The ginseng endocarp is composed of cellulose and hemicellulose. The hard endocarp (fruit coat) of Lannea microcarpa seeds contains lignin and hemicellulose [21]. The split (dehiscence) and germination of American ginseng seeds depended more on plumpness (seed kernel area/seed coat area) than on the seed size [22].
Kim et al. [23] observed that the water supply for embryo growth is accomplished by the vascular bundles entered into the endocarp through the opened hilum. The thickness of the raphae region of the endocarp is thinner (200 µm) than the other regions (300~600 µm), and sclereids in that region are regularly arranged. Hence, the seed coat of the ginseng seed opens, and it always occurs along the raphae, centering from the hilum [23].
Eight endophytic bacteria and 10 fungi were isolated from the endocarp of dehisced ginseng seeds [24,25]. Kim et al. [26] also identified five fungal strains of Talaromyces flavus from the dehisced seeds in traditional ginseng seed stratification facilities. They found that T. flavus-treated seeds showed more than 40% higher dehiscence than untreated seeds. The T. flavus (Penicillium dangeardii or P. vermiculatum) is a notable biocontrol agent for controlling soil-borne plant pathogens [27]. This species is also known for its ability to decompose various organic materials, including lignocellulosic substrates of plant materials [27]. Specific studies directly investigating its activity on endocarp (the hard inner layer of fruit) decomposition were not extensively documented. Fungi like Talaromyces are generally crucial in nutrient cycling within ecosystems, and they can participate in decomposing rigid plant materials. These ginseng seed-associated fungi were rarely colonized on ginseng roots in soil [28].
Watering of inoculated seeds should start a few days after the microbial inoculation to allow inoculation to the surface areas of the endocarp. Too frequent watering at the initial stage failed inoculation. However, watering that is too scarce increases the microbial density, resulting in the decay of seeds.
Some studies reported dormancy breaking of seeds with water-impermeable coats by soil-microbial action. For example, microbial inoculation promotes the germination of Rosa seeds [29]. However, Baskin and Baskin [13] found “only weak support for breakage physical dormancy by soil microbial action”. In Yang et al.’s study [19] using the traditional warm stratification facility in the summer-autumn season, microbial inoculation of bacteria or fungi on the ginseng seed surface lowered the dehiscence percentage compared to the intact seeds. In that case, additional inoculation of microorganisms may hurt the embryo and hinder dehiscence since the authors already used sand as a substrate.
When the seeds were treated with fungicide Captan, the hardness of the seed endocarp merely decreased from 10.9 kg cm−1 to 8.0 kg cm−1, compared to the non-treated seeds, which dropped to 4.9 kg cm−1 during the 80 days of warm stratification [25]. The EER and dehiscence for fungicide-treated seeds were 0.34 and 5%, in contrast to non-treated seeds EER of 0.43 and dehiscence 79% [25]. Lee et al. [25] argued that embryo growth aligned with the decreased seed hardness. Moreover, the frequency of fungi isolation from the endocarp was negatively correlated (r= −0.984) with the hardness of the seed endocarp [25]. The color of the endocarp surface for fungicide-treated seeds remained yellowish brown compared to the non-treated seeds, which turned dark brown [25].
5. Conclusions
Panax ginseng seeds have triple dormancy: morpho-physiological dormancy and physical dormancy. Therefore, the seeds need warm and cold stratification. During warm stratification, three conditions are necessary for embryo development: moisture, oxygen, and moderate temperature. Moreover, microbial inoculation is crucial for endocarp opening (dehiscence) and embryo development. River sand as a dehiscence substrate was effective on embryo growth and dehiscence. However, fungi inoculation was more reliable in indoor warm stratification. In contrast, soaking the seeds with 5% NaOH decreases the endocarp hardness but hinders microbial inhabitation, resulting in lower dehiscence.
These findings suggest that the cellulose decomposing activity of microorganisms is crucial to opening the endocarp of ginseng seeds associated with embryo development. Any microbes (fungi, bacteria, actinomycetes) tested with high or low cellulose decomposing ability increased dehiscence percentage by 66% compared to the untreated control. Three varieties of P. ginseng and one variety of P. quinquefolius seeds were successfully dehisced using the fungal inoculation of seeds.
Author Contributions
Conceptualization, investigation, data analysis, writing, and funding acquisition —H.K. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Research Promotion Program of Sunchon National University (SCNU).
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author/s.
Acknowledgments
Thanks to lab members and Mrs. EJ Han for excellent technical support.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Lee, J.H.; Lee, S.S.; Ahn, I.O.; Kang, J.Y.; Lee, M.G. Relation between storage periods and germination ability of dehisced seeds of Panax ginseng C.A Meyer. Korean J Ginseng Sci. 2004, 28, 215-218. (in Korean).
- Lee, M.K.; Jung, C.M.; Lee, J.H.; Jung, Y.Y.; Kang, J.Y.; Kim, M.S.; Choi, K.T. (1995) in Annual Report of Korean Ginseng and Tobacco Research Institute, KT&G, pp. 341-417. (in Korean).
- Kim, H.H.; Lee, J.H.; Shin, D.J.; Ko, H.C.; Hwang, H.S.; Kim, T.; Cho, E.G. Engelmann, F. Desiccation sensitivity and cryopreservation of Korean ginseng seeds. CryoLetters 2008, 29, 419-426.
- Han, E.; Popova, E.; Cho, G.; Park, S.; Lee, S.; Pritchard, H.W.; Kim, H.H. Post-harvest embryo development in ginseng seeds increases desiccation sensitivity and narrows the hydration window for cryopreservation. CryoLetters 2016, 37, 284-294.
- Kondo, T.; Sato, C.; Baskin, J.M.; Baskin, C.C. Post-dispersal embryo development, germination phenology, and seed dormancy in Cardiocrinum cordatum var. glehnii (Liliaceae s. str.), a perennial herb of the broadleaved deciduous forest in Japan. Am. J Bot. 2006, 93, 849-859. [CrossRef]
- Lee, J.W.; Jo, I.H.; Kim, J.U.; Hong, C.E.; Kim, Y.C.; Kim, D.H.; Park, Y.D. Improvement of seed dehiscence and germination in ginseng by stratification, gibberellin, and/or kinetin treatments. Hortic. Environ. Biote. 2018, 59, 293-301. [CrossRef]
- Li, T.S.C.; Bedford, K.E.; Sholberg, P.L. Improved germination of American ginseng seeds under controlled environments. HortTechnology 2000, 10, 131–135. [CrossRef]
- Kwon, W.S.; Lee, J.H.; Lee, M.G. Optimum chilling terms for germination of the dehisced ginseng (Panax ginseng C. A. Meyer) seed. J Ginseng Res. 2001, 25, 167–170.
- Kim, S.J.; Kim, J.U.; Lee, J.W. Improvement of seed dehiscence using plant growth regulators and its effect on subsequent germination and growth of Panax ginseng C. C. Meyer. Korean J. Medicinal Crop Sci. 2024, 32, 295-304. [CrossRef]
- Nikolaeva, M.G. 1977. Factors controlling the seed dormancy pattern. In A. A. Khan [ed.] The physiology and biochemistry of seed dormancy and germination p. 51–74 North-Holland, Amsterdam, Netherlands.
- Baskin, C.C.; Baskin, J.M. 1998. Seeds: ecology, biogeography, and evolution of dormancy and germination Academic Press, San Diego, California, USA.
- Baskin, J.M.; Baskin, C.C. A classification system for seed dormancy. Seed Sci Res. 2004, 14, 1–16. [CrossRef]
- Baskin, J.M.; Baskin, C.C. Evolutionary considerations of claims for physical dormancy-break by microbial action and abrasion by soil particles. Seed Sci. Res. 2000, 10, 409-413. [CrossRef]
- Kim, Y.C.; Kim, Y.B.; Park, H.W.; Bang, K.H.; Kim, J.U.; Jo, I.H.; Kim, K.H.; Song, B.H.; Kim, D.H. Optimal harvesting time of ginseng seeds and effect of gibberellic acid(GA3) treatment for improving stratification rate of ginseng (Panax ginseng C. A. Meyer) seeds. Korean J. Medicinal Crop Sci. 2014, 22, 423-428. [CrossRef]
- Davis, W.E.; Rose, R.C. The Effect of External Conditions upon the After-Ripening of the Seeds of Crataegus mollis. Botanical Gazette 1912, 54, 49–62. http://www.jstor.org/stable/2468391.
- Lee, J.C.; Byun, J.S.; Proctor, J.T.A. Dormancy of ginseng seed as influenced by temperature and gibberellic acid. J. Korean Soc. Crop Sci. 1986, 31, 220-225. (in Korean).
- Won, J.Y.; Jo, J.S.; Kim, H.H. Studies on the germination of Korean ginseng (Panax ginseng C.A. Meyer) seed. J. Korean Soc. Crop Sci. 1988, 33, 59-63. (in Korean).
- Choi, S.Y.; Lee, K.S. Studies on the physiological chemistry of dormancy and germination in Panax ginseng seeds. Kor. J. Crop Sci. 1987, 32, 277-286. (in Korean).
- Yang, D.C.; Cheon, S.K.; Yang, D.C.; Kim, H.J. The effect of various dehiscence materials, growth regulators and fungicides on the ginseng seeds (Panax ginseng C.A. Meyer). Korean J. Ginseng Sci. 1982, 6, 56-66. (in Korean).
- Lim, S.H.; Jeong, H.N.; Kang, A.S.; Joen, M.S. Influence of GA3 soak and seed dressing with oros (tolclofos methyl) wp. on the dehiscence of Eleutherococcus senticosus Maxim seeds. Korean J. Med. Crop Sci. 2008, 16, 106-111.
- Neya, O.; Hoekstra, F.A.; Golovina, E.A. Mechanism of endocarp-imposed constraints of germination of Lannea microcarpa seeds. Seed Scie. Res. 2008, 8, 13-24. [CrossRef]
- Xu, S.Q.; Zhang, H.; Hou, Z.F.; Wang, Y.P. Characteristics of seed size and its relationship to germination in American ginseng (Panax quinquefolius L.). J. Medicinal Plant Sci. 2017, 5, 4-8.
- Kim, W.K.; Kim, E.S.; Jeong, B.K. A study on structure and differentiation of seed coat of Panax ginseng C. A. Meyer. Korean J. Bot. 1986, 29, 295-315.
- Um, Y.; Kim, B.R.; Jeong, J.J.; Chung, C.M.; Lee, Y. Identification of endophytic bacteria in Panax ginseng seeds and their potential for plant growth promotion. Korean J Medicinal Crop Sci. 2014, 22, 306–312. (in Korean). [CrossRef]
- Lee, J.C.; Chung, Y.R.; Park, H.; Ohh, S.H. Influence of seed dressing with Captan wp. on the dehiscence of Panax ginseng seeds. Korean J. Crop Sci. 1983, 28, 262-266. (in Korean).
- Kim, M.J.; Shim, C.K.; Kim, Y.K.; Hong, S.J.; Park, J.H.; Han, E.J. S.C. Enhancement of seed dehiscence by seed treatment with Talaromyces flavus GG01 and GG04 in ginseng (Panax ginseng). Plant Pathology J. 2017, 33, 1-8. [CrossRef]
- Fahima, T.; Henis, Y. Quantitative assessment of the interaction between the antagonistic fungus Talaromyces flavus and the wilt pathogen Verticillium dahliae on eggplant roots. Plant Soil 1995, 176, 129–137. [CrossRef]
- Peterson, E.A. Seed-borne fungi in relation to colonization of roots. Can. J. Microbiol. 1959, 5, 579-582. [CrossRef]
- Kazaz, S.; Erbas, S.; Baydar, H. Breaking seed dormancy in oil rose (Rosa damascena Mill.) by microbial inoculation. African J. Biotech. 2010, 9, 6503-6508.
Figure 1.
The ginseng seeds before and after dehiscence. A; de-pulped but un-dehisced ginseng seeds in late July. The Arrow of the hilum indicates an endocarp hole (less than 1 mm in diameter) that allows water imbibition and aeration. B; dehisced seeds in November. The Arrow of the raphae area is relatively thinner than the other areas of the endocarp. C-D; longitudinally bisected un-dehisced seed after harvest with the immature embryo (0.4 mm) on the right side (C) and fully developed embryo after warm and cold stratification (D).
Figure 1.
The ginseng seeds before and after dehiscence. A; de-pulped but un-dehisced ginseng seeds in late July. The Arrow of the hilum indicates an endocarp hole (less than 1 mm in diameter) that allows water imbibition and aeration. B; dehisced seeds in November. The Arrow of the raphae area is relatively thinner than the other areas of the endocarp. C-D; longitudinally bisected un-dehisced seed after harvest with the immature embryo (0.4 mm) on the right side (C) and fully developed embryo after warm and cold stratification (D).


Table 1.
List of microbial strains inoculated with un-dehisced ginseng seeds.
| Group | No. | Microbial strains | Source | Code |
|---|---|---|---|---|
| Fungi | 1 2 3 4 5 6 |
Talaromyces flavus (F1) Talaromyces flavus (F2) Talaromyces flavus (F3) Talaromyces flavus (F4) Mix of the four strains of T. flavus Trichoderma harzianum (40509) |
Isolated Isolated Isolated Isolated Isolated Non-decomposition |
fungi #1 fungi #2 fungi #3 fungi #4 fungi-mix fungi 40509 |
| Actinomycetes | 1 2 3 4 |
Kitasatospora gansuensis (CMM1-1) Micromonospora matsumotoense (CMM7-5) Strptomyces sp. (CMM1-28) Mix of three strains of Actinomycetes |
Decomposing Decomposing Decomposing Decomposing |
actino CMM1-1 actino CMM7-5 actino CMM1-28 actino-mix |
| Bacteria | 1 2 3 4 5 |
Bacillus sp. (B5001) Bacillus sp. (B5002) Bacillus sp. (B2390) Mix of three strains of bacteria Bacillus amyloliquefaciens (amylo 12067) |
Decomposing Decomposing Decomposing Decomposing non-decomposition |
bacter B5001 bacter B5002 bacter B2390 bacter-mix bacter amylo |
* Microbial strains were cultured in rice bran in vitro. The 1% (w/v) microbial inoculum was inoculated with ginseng seeds by co-culture at 25°C for a day and then kept at 18-20°C room for 3-5 days without watering. .
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



