
Submitted:
04 December 2024
Posted:
05 December 2024
You are already at the latest version
Abstract
The plant microbiome, encompassing the rhizosphere, phyllosphere, and endosphere, plays a critical role in nutrient acquisition, stress tolerance, and overall plant health. While extensive research exists on crop microbiomes, our meta-survey identified a significant gap in focused reviews addressing the rice plant microbiome. As the staple food for over half the global population, rice faces mounting challenges from climate change and environmental stresses. This review bridges this knowledge gap by synthesizing advancements in understanding the diversity and functional roles of rice-associated microbiomes. It highlights their contributions to plant growth, nutrient uptake, and resilience against biotic and abiotic stresses, emphasizing rhizosphere signalling mechanisms that mediate plant-microbe interactions. Advances in metagenomics have transformed our ability to map rice microbiomes, uncovering their composition and functional potential. The review also explores microbiome engineering strategies, including microbial inoculation, gene editing, and microbiome-shaping (M) genes, to develop stress-resistant rice varieties. These tools hold promise for improving rice productivity, reducing chemical inputs, and enhancing climate resilience. However, challenges remain in deciphering microbial interactions, environmental variability, and the scalable application of microbiome technologies. This review provides critical insights into leveraging microbiomes for sustainable rice production and outlines future directions for integrating advanced genomic tools and microbiome engineering into global agricultural practices.
Keywords: Microbiome Engineering, Rice Production, Biofertilization, Stress Tolerance, Microbiome-Shaping Genes, Metagenomics, Climate Resilience, Sustainable Agriculture
Keywords:
Microbiome Engineering
; Rice Production
; Biofertilization
; Stress Tolerance
; Microbiome-Shaping Genes
; Metagenomics
; Climate Resilience
; Sustainable Agriculture
1. Introduction
Microbiome engineering (ME) presents a transformative approach to boosting agricultural productivity and enhancing climate resilience [1,2]. By harnessing the power of microbial communities, ME optimizes ecosystem functions to promote plant growth and development [3,4]. This cutting-edge biotechnological strategy holds immense promise in addressing global food security challenges amid rapidly changing environmental conditions [5]. Symbiotic relationships with beneficial microbes boost plant resilience to biotic and abiotic stresses [6]. These interactions enhance plant health and productivity, essential for achieving sustainable agriculture [7,8]. Plant-associated microbiota includes epiphytes on the surface, endophytes in plant tissues, and the rhizosphere, endosphere, and phyllosphere environments. Many promising genera such as Rhizobium, Bacillus, Paraburkholderia, Delftia, Pseudomonas, Enterobacter, Lysobacter, Serratia, Alcaligenes, Burkholderia, Azotobacter, Azospirillum, Enterobacter, Brevibacterium, Clostridium etc. have demonstrated notable capabilities in augmenting plant growth, yield and curtailing defence mechanisms for sustainable agriculture [9,10]. These microbiomes can be customized to specific crop-soil-environment combinations, providing a targeted approach to plant support. This symbiotic relationship fosters improved crop yield, soil health, and ecological balance.
Agriculture faces significant environmental issues in the modern era, including water scarcity, climate change, and declining soil and water quality [11,12,13]. The United Nations (2019) projects that the global population will rise from 8.2 billion to 9.7 billion by 2050, creating a need for a 70-110% increase in food production to meet nutritional demands. Rice being a staple food for over 3.5 billion people worldwide will play a crucial role in addressing this need, particularly in developing countries [14,15]. The demand for rice is expected to rise continuously propelled by the steady growth of the global population [16,17]. The current use of agrochemicals and inorganic fertilizers has contributed to the degradation of soil health, water quality, dysbiosis of the microbiome, decline in beneficial insect populations, loss of biodiversity, disruption of ecosystem functions, increased vulnerability of rice plants to diseases, and an imbalance in microbial structures, causing irreversible harm to agriculture systems [18,19,20]. Only a limited percentage of sprayed chemical pesticides (about 1%) are reported to reach their intended targets due to volatilization, leaching, photolysis, and runoff [21]. With the combination of contemporary technology and environment-friendly management techniques, the application of plant microbiome for a sustainable, eco-friendly agricultural system has opened up new opportunities [5,22,23,24].
Recent plant microbiome research tools and techniques are essential for studying whole plant microbiota and its role in improving growth and productivity under stress [25]. Application of plant-beneficial bacteria improves the growth and yield of rice [26] by improving nutrient uptake and mitigating the impacts of both biotic and abiotic stresses [27,28,29]. Compant et al. [30] identified that Sphingomonas melonis naturally confers resistance to seed-borne pathogens like Burkholderia plantarii in rice. For “biological resistance” biopolymers like exopolysaccharides form a protective matrix around roots, while osmolytes, such as soluble sugars, amino acids, and organic solutes like proline and glycine betaine-help plants withstand stress, as observed in drought-resistant plants and halophytes colonized by Pseudomonas putida, Pantoea brenneri, and Acinetobacter calcoaceticus [31]. Rice plants inoculated with mycorrhiza exhibited improved photosynthetic efficiency under saline and drought stress conditions by direct uptake and translocation of water from the soil to the host plant via fungal hyphae, altering water retention properties of soil and higher adjustment of the osmotic potential of AM plants [32,33]. Additionally, microbes like Pseudomonas sp. and P. brenneri reduce oxidative stress through antioxidant enzymes and metabolites, and phytohormones or volatiles trigger induced systemic tolerance (IST) in plants, with ACC deaminase lowering ethylene levels to support stress resilience [30]. Microorganisms like Pseudomonas, Bacillus, Trichoderma, and arbuscular mycorrhizal fungi (AMF) can trigger induced systemic resistance (ISR) in plants, which helps protect them against various pathogens [34]. ISR activates defence-related genes, promoting the production of enzymes like phenylalanine ammonia-lyase, polyphenol oxidase, peroxidase, β-1,3-glucanase, and chitinase, along with reactive oxygen species accumulation [35]. Ashnaei (2019) [36] reported that Bacillus subtilis UTSP40 exhibited superior biocontrol efficacy against Magnaporthe oryzae FR1. Besides, Entomopathogenic fungi use virulence factors like chitinase enzymes and mycotoxins to effectively infect insects [37].
PGPR (plant growth-promoting rhizobacteria) enhances nutrient absorption, particularly nitrogen and phosphorus which support rice growth [38]. The rice-rhizobia symbiosis accounts for approximately 80% of total nitrogen fixation [39] where cyanobacteria can boost rice grain yield by 11% and contribute 25–50 % of nitrogen each season per hectare of fields [40]. Azotobacter and Azospirillum spp., has been reported to increase nitrogen availability by up to 0.4-0.9 tha-1 and 1.8 tha-1, respectively [41]. The bacterial genera Desulfobacteria, Entotheonella, and Algoriphagus, along with members of Anaerolineae and α-Proteobacteria are essential for sulfur oxidation and reduction, nitrogen fixation, and phosphorus metabolism, enhance nutrient availability and uptake in rice [42]. Inoculation of rice with various Rhizobium strains at different nitrogen levels resulted in a 4% to 19% increase in straw yield and an 8% to 22% increase in grain yield [43]. The Azolla-Anabaena system contributed 1.1 kg N ha-1day-1, providing 20–40 kg N ha-1 to rice in 20–25 days [44]. Zn-solubilizing bacteria (Burkholderia spp. and Acinetobacter spp.) exhibited enhanced growth and yield, with improved zinc acquisition [45]. These eco-friendly, cost-effective solutions not only improve plant yield, and defense systems but also protect against environmental stressors and offer advantages over harmful chemical fertilizers. Microbes in agroecosystems facilitate communication among plants, with plant genes playing distinct roles in shaping (Ms genes) or responding (Mr genes) to microbial communities. Understanding these gene–microbe interactions could reveal how certain genes offer new insights into gene–phenotype relationships. Ms genes exert a significant influence on plant-associated microbial communities by regulating the production of specific small peptides, secondary metabolites, and even through trans-kingdom miRNA transfer. This could allow plants to selectively recruit beneficial microbial communities to enhance their adaptability to various environments to support the host plant. Genome editing tools, such as CRISPR-Cas9, have rapidly advanced and are now indispensable genetic engineering tools for improving plant pathogen stress tolerance. Some research establishes a clear connection between a particular plant chemical and the health of the leaf microbiome, and suggested potential avenues for developing disease-resistant rice varieties through breeding.
Extensive studies and reviews have explored the rice microbiome, focusing on its diverse functions and potential applications in sustainable agriculture [46,47,48]. This review aims to critically analyze various aspects of the rice microbiome and outlines the following objectives: 1) to explore the diversity and dynamic structures of rice-associated microbiomes in the rhizosphere, phyllosphere, and endosphere, focusing on their roles in growth, nutrient acquisition, and stress resilience; 2) to examine signaling mechanisms in the rice rhizosphere and the contributions of microbes to growth, yield, and stress tolerance; 3) to highlight the role of metagenomics in mapping rice-associated microbial communities and their functional potential; 4) to discuss microbiome engineering strategies, including microbial inoculation, gene editing, and microbiome-shaping (M) genes, for developing stress-resistant rice cultivars and enhancing climate resilience; and 5) to identify current challenges in microbial interactions, scaling microbiome applications, and integrating these strategies into sustainable rice production, while offering insights into future research directions.
2. Understanding the Diversity and Functional Dynamics of Rice-Associated Microbiomes
Rice microbiomes exhibit diverse and dynamic structures, comprising intricate microbial communities that interact with host plants [49,50,51,52]. The biogeography of soil microbes significantly influences the composition of rice rhizosphere microbial communities across different planting locations [53]. Local environmental conditions, such as soil type and climate, directly determine the available microbial species pool and indirectly impact rhizosphere microbial communities by affecting plant and microbial physiology. Consequently, the relative contributions of plant genotype and planting location in shaping the rhizosphere microbiome remain a subject of ongoing debate [54]. The rhizosphere microbial community of Japonica rice and hybrid rice exhibits discernible structural differences, which are strongly correlated with rice yields [55]. The rice rhizosphere, a specific microbial environment, is marked by varying levels of oxidation due to the roots' oxygen release and intensive water control. This unevenness may contribute to the limited presence of certain microbial groups [56].
Bacteria in the rice field significantly enhance the geochemical cycling of essential nutrient elements and also enhance release of methane and other greenhouse gases that impact global warming [57,58]. Rice fields are rich ecosystems hosting diverse microorganisms [59,60], including nitrogen fixers, nitrifiers, and methanogens. These microbes play crucial roles in nutrient cycling, enhancing plant growth, and maintaining ecological balance [61].”The intricate interactions among these diverse components contribute to the overall health, nutrient cycling, and ecological balance within rice ecosystems [62] The composition and interactions within rice microbiomes can be influenced by factors such as soil type, genotype of rice, agricultural practices, and chemical, physical, and environmental conditions [56,63,64].
Using rice as a model, shows that there exist three different root niches hosting different microbial communities of eubacteria and methanogenic archaea. Tan et al. [65] explored the role of microbiomes in promoting healthy growth and yield of rice plants as part of climate-smart agricultural practices. This study emphasizes how microbial communities can be engineered for improved rice productivity under varying environmental conditions. Islam et al. [66] showed how root-associated microbes influence plant health, stress tolerance, and yield enhancement of rice. Adachi et al. [67] analysed the dynamics of root-associated microbiomes in rice and how these dynamics are influenced by soil nutrition. Their preprint on bioRxiv highlights the variability in microbial community composition based on nutrient availability in rice. Juliyanti et al. [68] analysed the microbial composition in the rhizosphere and endosphere of rice roots, finding no significant difference between weedy and cultivated rice lines. The endosphere had less diversity, with major groups from Proteobacteria, Myxococcota, Chloroflexota, and Actinobacteria.
These microbial communities are affected by geographical location, soil source, host genotype, and cultivation practice. Both biotic and abiotic variables have an impact on the microbial communities in rice field soils [69]. These include soil texture, the particular rice cultivar planted, pesticide applications, temperature, precipitation, humidity, pH levels, and anions and cations in balance [70].
Many species of oomycetes, archaea, and other microbes are crucial to the operation of many ecosystem services and ecological processes that support soil fertility and health of rice [71]. In these communities, bacteria and fungi hold prominent positions despite their diversity. Ascomycota, Basidiomycota, and Glomeromycota are the most common members of fungal communities, whereas Proteobacteria, Chloroflexi, Actinobacteria, Prevotella, and Acidobacteria usually dominate bacterial communities. Understanding this intricate microbial diversity is essential for effective soil management and sustainable agricultural practices in rice cultivation. Recently developed shot-gun metagenomics coupled with bioinformatics tools offer enormous opportunities for shedding light on population diversity and functional aspects of rice microbiome and their interactions with the host plants.
Rice plant-associated microbiota includes epiphytes on the surface, endophytes in plant tissues, and the rhizosphere, endosphere, and phyllosphere environments (Figure 1). Rice plant-associated microorganisms can be mutualists, commensals, or pathogens, with pathogenic ones being of concern due to their economic impact [72]. Multiple interactions between the host, microbes, and environment shape the plant-associated microbiome Proteobacteria and Firmicutes are enriched in plant endophytic communities, while Acidobacteria, Planctomycetes, Chloroflexi, and Verrucomicrobia are depleted. Ascomycota and Basidiomycota are the dominant fungal phyla in both above- and below-ground plant tissues. The phyllosphere community is mainly composed of Proteobacteria, Bacteroidetes, Firmicutes, and Actinomycetes [73].
3. The Role of Rhizospheric Microbes in Rice Health and Growth
The rhizosphere refers to the soil zone within 1–10 mm surrounding plant roots, where the plant exerts influence through the release of mucilage, root exudates, and deceased plant cells. The intricate interplay among the roots, rhizosphere microbiome, and abiotic factors culminates in a complex structure, which we commonly call as the rhizobiocomplex. Many studies indicate that under standard crop management, the central microbiome of rice rhizospheres includes prevalent bacterial genera such as Arenimonas, Arthrobacter, Anaeromyxobacter, Bacillus, Bellilinea, Prevotella, and mycorrhizae, PGPR, and biocontrol microbes all known for contributing to the growth and disease resistance of rice plants [74]. Approximately one million unique bacterial genomes could be present in only one gram of soil. Guo et al. [50] found that the rice rhizosphere microbiome exhibited elevated α-diversity, and reduced β-diversity due to the homeostatic influence of the roots. Rice bacterial leaf blight (BLB) is a destructive phyllosphere bacterial disease caused by Xanthomonas oryzae pv. oryzae (xoo) significantly alters the bacterial-fungal community in the rhizosphere, decreasing bacterial diversity but not fungal diversity. Bacterial Leaf Blight significantly impacts the rhizosphere microbiome's functional adaptation, increasing the abundance of functional genes involved in carbon, phosphorus, and methane metabolism [75]. Li et al. [76] found that inoculation with the R3 strain of Herbaspirillum altered the structure of native nitrogen-fixing microbial communities in the rhizosphere and endorhizosphere, increasing diversity and enhancing the abundance of key nitrogen-fixing genera (as Ralstonia, Azotobacter, Geobacter, Streptomyces, and Pseudomonas). Additionally, upregulation of nitrogen absorption and transport-related genes (OsNRT1 and OsPTR9) in rice roots may contribute to increased yield. Pang and his colleagues isolated bacteria and fungi from upland rice roots in Xishuangbanna, China in 2020 and they found root microbes differed from irrigated rice roots, with Firmicutes phylum members enriched by 28.54% [77]. Their study also found that fungi from upland rice roots can increase plant growth under irrigated and drought-stress conditions, making them effective microbial resources for sustainable agricultural production in arid regions. Zhang et al. [78] showed significant differences among the microbiomes between wild and cultivated rice, with indica rice having more similar bacterial and fungal communities. Indica rice had the lowest proportion of Actinobacteria and the highest relative abundance of Nitrospira. However, indica and japonica rice had higher relative abundances of Magnaporthales and Ustilaginales.
Different inter-cultural operations like herbicide application also significantly reduced the abundance and diversity of soil microbial communities [79]. They stated that the persistence of certain taxa, including Nitrosocosmicus, Nitrososphaera (archaea), Bacillus, Methyloceanibacter, Nitrospira, Thaurea (bacteria), and Acrophialophora, Aspergillus, Clonostachys, Emericellopsis, Exserohilum, Fusarium, Humicola, Nigrospora, Pyrenochaetopsis (fungi), improve the nutrient cycling and soil fertility maintenance in rice agroecosystems. A study conducted by Hester et al. [80] found that rice root microbiota is more sensitive to water management changes compared to bulk soil, with aerobic, PGPB being enriched under Alternate Wetting and Drying (AWD) treatment. In contrast, anaerobic microorganisms were depleted, highlighting the potential of AWD to influence soil and root microbial communities differently.
Despite extensive studies on the impact of numerous isolated fungi and bacteria on rice growth and fitness, the intricate interplay among the root and rhizosphere microbiomes, and their collective role in rice growth, yield, and tolerance to various stresses remains poorly understood. As certain microorganisms have been demonstrated to regulate host gene expression, it is imperative to explore the far-reaching implications of rhizosphere microbiome structures. This investigation is essential for utilizing them in a bio-rational manner to sustainably enhance rice production while maintaining soil health.
4. Phyllospheric Microbes and Their Contributions to Rice Growth and Disease Resistance
The above-ground parts of rice plants, particularly their leaves, stems and nodes, provide a habitat that harbours a diverse range of microorganisms, many of which are essential for the plant’s growth and health [52]. Unlike the nutrient-rich rhizosphere and endosphere, the phyllosphere, which encompasses the area around or on the leaves, tends to be relatively low in nutrients [70]. It is a dynamic environment where resident microbes encounter fluctuating environmental factors such as temperature, water availability, humidity, solar radiation, and altitude.
Wang et al. [81] investigated the phyllosphere microbial communities of a rice plant species at various elevations and growth stages. Their findings revealed a predominance of Proteobacteria, Actinobacteria, and Bacteroidetes among bacteria, and Ascomycota and Basidiomycota among fungi. These phyla exhibited significant variations across different elevations and growth stages. Elevation exerted a more pronounced influence on the α diversity of phyllosphere bacteria compared to fungi, while the growth stage significantly affected the α diversity of both bacterial and fungal communities. Additionally, the study demonstrated that the composition of both bacterial and fungal communities varied substantially along the elevation gradient within different growth stages.
The exploitation of rare earth minerals alters bacterial community structures in rice, reduce the diversity of phyllosphere dominant bacteria, (such as Burkholderia, Bacillus, Buttiauxella, Acinetobacter, Bradyrhizobium, Candida koribacter) but promoting dominant bacteria that help degrade pollutants and improve nutrient availability, supporting rice growth. However, increased REE content in rice from mining areas poses potential risks to human health, raising concerns about food safety [82]. Phyla Actinobacteria and Firmicutes are often found in arid environmental conditions. Methylotrophic bacteria, primarily from the genera Hyphomicrobium, Methylobacterium, Methylibium, Methylophilus, Methylocapsa, Methylocella, and Methylocystis, are the dominant organisms found in the phyllosphere. Qi et al. [83] identified Aspergillus as the most prevalent fungal genus in rice grains. Notably, the mycotoxin-producing species A. flavus and A. niger were significantly more abundant. Yeo et al. [84] recorded that the prokaryotic community in rice landraces showed high homogeneity across different landraces, with Proteobacteria being the most abundant phylum and an undefined genus under Cyanobacteria dominating. They found that rice plant organs, such as leaf blades and stems, primarily determine the composition of the prokaryotic community.
Roman-Reyna et al. [85] has identified 12 genera that characterize the microbial community on rice leaves, ranging from commensal or pathogenic to beneficial, including Clostridium, Bacillus, Helicobacter, Azotobacter, and Pseudomonas. Proteobacteria appear to be the predominant colonizers, with Bacteroidetes and Actinobacteria also being prevalent in phyllospheres. Both biological and environmental influences, along with human activities such as farming methods and fertilizer use, play essential roles in shaping both the taxonomical and functional aspects of phyllosphere microbiomes. These diverse factors contribute to the dynamic and heterogeneous nature of the rice phyllosphere environment.
5. Endospheric Microbes and Their Role in Enhancing Stress Tolerance in Rice
Certain microorganisms, known as endophytes, have the ability to penetrate and occupy the internal tissues of plants, giving rise to the endospheric microbiome. This pertains to the internal realm of plants, where endophytes, encompassing both bacteria, fungi and viruses, reside for most of their life cycle within plant tissues without inducing pathogenic symptoms [86]. The composition of microorganisms in the endosphere can exhibit temporal variations.
Tian et al. [87] identified 96 endophytic bacterial strains with plant growth-promoting (PGP) traits from Oryza officinalis, with 11 strains (Enterobacter mori, E. ludwigii, E. cloacae, Bacillus amyloliquefaciens, B. siamensis, Pseudomonas rhodesiae and Kosakonia oryzae) demonstrating enhanced root development, biomass accumulation, chlorophyll content, and nitrogen uptake in perennial rice seedlings. Wu et al. [88] demonstrated that under abiotic stress (drought stress), rice may specifically enrich certain bacterial taxa(Actinobacteriota, Gemmatimonadetes, and Patescibacteria,Bacteroidetes and Firmicutes) and enhance their positive interactions within its root system to improve adaptation. They are a good indicator of stress tolerance under stressful conditions. Wang et al. [52] found that other microorganisms like Xanthomonas sacchari JR3-14, enhance the diversity and complexity of rice-associated bacterial communities, particularly in the root and stem endosphere during early growth stages. The assembly of these bacterial communities can increase seed endophytes for crop production and breeding.
Zhang et al. [89] revealed that rice microbiome assemblages are primarily shaped by microhabitat rather than geographic location or rice variety, with diversity and complexity decreasing from soil to seed. Notably, core bacterial endophytes, especially Pantoea and Xanthomonas, were vertically transmitted across generations, displaying plant growth-promoting traits but limited antagonism to major rice pathogens.
The AM fungi and other endophytic fungi are the primary inhabitants of the endosphere increase nutritional acquisition and improve the stress resilience of the host plants [90,91]. During the vegetative phase of rice plants, bacteria capable of producing siderophores, notably from the genera Sphingomonas, Pseudomonas, Burkholderia, and Enterobacter, are predominantly found in plant tissues. These bacteria play a vital role to facilitate the plant's acquisition of iron and other essential elements from the soil. Besides colonizing plant organs, endophytic bacteria are also associated with rice seeds. Genera belonging to the Alpha- and Gammaproteobacteria, Flavobacteria, Bacilli, and Actinobacteria classes have been observed inhabiting rice seeds. Some of these endophytic microorganisms engage in beneficial interactions with their host plants. Endophytic microorganisms contribute to enhance plant health, performance, and adaptation to both biotic and abiotic stresses. Moreover, the composition and diversity of the endophytic microbiome within above-ground and below-ground tissues may vary within the same plant.
6. Decoding Signaling Pathways in the Rice Rhizosphere
The signalling mechanisms in the rhizosphere can be classified into three main types. (i) Plants secrete low-molecular-weight compounds to communicate with microbes, fostering complex interactions essential for nutrient acquisition and stress resilience. (ii) Interspecies and intraspecies microbial signalling primarily occur through quorum-sensing (QS) and other signalling processes. It allows microorganisms to coordinate and adjust their behaviour based on population density and their functional behaviour. (iii) Microorganisms to plant signalling involve compounds produced by microorganisms that influence various aspects of plant physiology and immunity that ultimately impact root system architecture, plant defence mechanisms, responses to both biotic and abiotic stresses, and gene expression [92].
For instance, several studies have investigated the role of acyl-homoserine lactones (AHLs) and other signalling molecules in shaping plant-microbe interactions. Updated research highlights how these molecules not only regulate quorum sensing (QS) among microbial populations but also influence plant physiological responses, such as stress tolerance and nutrient uptake. Chen et al. [93] found that AHL-mediated signaling in the rice rhizosphere promotes the recruitment of beneficial microbes, enhance nitrogen fixation and root growth. Similarly, Zhang et al. [58] demonstrated that certain AHLs trigger systemic, improving the plant's defense against pathogens.
Additionally, beyond AHLs, other signalling molecules like volatile organic compounds (VOCs) and lipo-chitooligosaccharides (LCOs) have been shown to play critical roles in plant-microbe signalling. Wang et al. [81] uncovered that VOCs released by Pseudomonas spp. in the rice rhizosphere can alter root architecture, improving water and nutrient acquisition. Kelbessa et al. [94] (2022) expanded on the role of LCOs in promoting symbiotic relationships, particularly in phosphate-solubilizing bacteria, which significantly enhances nutrient cycling in paddy fields.
These recent publications underscore the complex and multi-faceted nature of microbial signalling in rice ecosystems. Integrating these studies would provide a richer, more nuanced discussion of how microbial communication contributes to plant health, nutrient efficiency, and resilience. Addressing these aspects would not only align better with the objectives of this review but also offer a more up-to-date and robust understanding of the microbial plant signalling networks that are crucial in rice production systems.
According to Santosh Kumar et al. [95], rice seedling roots interact with Sinorhizobium meliloti 1021, which secretes bioactive signalling chemicals recognized by rice receptor proteins (PRRs, FLS2, and LRR-RLKs), triggers gene expression changes in the plant. These bioactive molecules interact with rice root cells close to where bacteria colonize and initially infect. Once these bacterial signals are identified and transmitted, the expression of DEGs associated with producing phytohormones that control plant growth and development, like auxins, gibberellins, and cytokinins, changes in the rice seedling shoots. Subsequently, the bacteria migrate within the plant to other rice tissues where they coexist symbiotically, continuing to influence plant gene expression.
Xiao et al. [96] reported that the combined application of Rhodopseudomonas palustris and Bacillus subtilis resulted in a substantial increase in rice yields, reaching up to 17.73%. This synergistic effect was attributed to the significant modification of the soil bacterial community structure. Furthermore, the study demonstrated an enhancement of membrane transporters and signal transduction pathways, coupled with an increase in certain essential metabolic pathways.
Acyl-homoserine lactones (AHLs) are the primary signalling molecules involved in quorum sensing (QS) and are typically species-specific [58]. AHLs function by interacting with bacterial receptors, leading to alterations in gene expression. This enables different microbial populations to synchronize their activities, thereby allowing them to function as a unified and coordinated community [97]. Viswanath et al. [97] demonstrated that QS systems using AHLs are integral to communication among bacterial species within the rhizosphere. They highlighted how AHL-mediated QS can influence the interactions between microbes, affecting traits like biofilm formation, motility, and the production of secondary metabolites. These interactions are vital for establishing a balanced microbial community that can adapt to environmental changes and compete with other microbes in the soil environment.
QS plays a crucial role in structuring the rhizosphere microbiome, enabling the formation of microbial consortia that can have either beneficial effects, such as promoting plant growth and resilience, or pathogenic impacts [5]. Recent findings have reported the production of AHLs for the first time in the rice rhizosphere by species such as Acinetobacter lactucae, Aeromonas popofi, Serratia oryzae, and Rhizobium wuzhouense. The presence of diverse AHLs among these rhizobacterial groups suggests that these signalling molecules might be key regulators in coordinating rhizobacterial behaviour and promoting symbiotic interactions between microbes and plants. This discovery sheds light on the potential role of AHLs in regulating the dynamic interactions within the rice rhizosphere, contributing to enhanced plant health and growth. The establishment of a beneficial microbial community in the rice rhizosphere relies on signalling events between the host plant and bacterial symbionts. Molecular signals, including the well-known Nod factor, facilitate the selective recruitment of specific microbes by the plant. This process involves the exchange of signals, such as flavonoids inducing bacterial Nod factor production. Tricarboxylic acid secretion also contributes to recruiting other beneficial microbes, enhancing overall plant health. Cooperative interactions among different microbes shape the plant's well-being. It is a common occurrence that volatile organic compounds (VOCs) produced by microorganisms can also stimulate both plant growth and tolerance to biotic stresses.
Plants have the ability to detect and respond to various VOC signals originating from PGPR or plant growth-promoting fungi (PGPF), including compounds like undecanone and heptanol. The VOCs such as 2-heptanol and 2-undecanone, released by Bacillus subtilis and B. amyloliquefaciens, have been identified to enhance the growth performances of Arabidopsis thaliana when grown in the presence of these PGPR strains. This underscores the significant role of microbially produced VOCs in influencing plant physiology, fostering growth, and fortifying resistance against biotic stress [98]. VOCs also induce systemic resistance of rice plants to biotic and abiotic stresses.
Plants activate signalling pathways to defend against various threats, such as insects and pathogens. Different pathways along with jasmonic acid (JA)/Ethylene (ET) dependent Induced Systemic Resistance (ISR) and salicylic acid (SA) dependent SAR, are associated in plant defence responses [99,100]. Herbivore feeding habits induce JA-SA signalling, activating defence responses in distal organs. Aboveground herbivory can recruit rhizobacteria and induce systemic signalling throughout the plant. Plant immune signalling influences microbial communities in the root microbiome, with changes in JA signalling impacting the rhizosphere community. Signalling compounds, such as hormones (JA, ET, SA, ABA, CK, GA, auxin), are essential. Plants release root exudates in response to changes in JA signalling, influencing bacterial and archaeal abundances in the rhizosphere. The production of signalling compounds is triggered by stimuli like herbivory, pathogen attack, and environmental stress. Signalling pathways involve specific receptors, initiating cascades leading to the synthesis and release of compounds.
Rice root exudates indeed consist of a diverse array of primary and secondary metabolites, encompassing sugars, amino acids, organic acids, phenolics, and flavonoids [101,102]. The composition and stoichiometric ratios of these exudates have been shown to impact microbial activity, carbon mineralization, and the microbial biomass in the rhizosphere, thereby modulating the biotic interactions and nutrient cycling in the soil ecosystem. Some beneficial endophytes like Azoarcus activate JA and SA signalling, promoting root colonization and shaping the root microbiome of rice [103]. Endophytic fungus Acrocalymma vagum enhanced rice yield by 5.73% and induced 83.24%resistance against rice. Likewise, Some Lactobacillus and Nigrospora strains possessed significantly inhibitive activity on rice blast pathogen Magnaporthe oryzae. Additionally, some fungal strains of Sarocladium and Nigrospora genera promoted rice growth [104].
This allows plants to communicate with microbes and regulate various physiological processes, including defence responses and growth. The production of plant hormones by bacteria is considered for plant growth promotion, though evidence in the phyllosphere is limited. Overall, understanding these intricate signalling networks provides insights into plant-microbe interactions for biofertilization and stress resilience in rice.
7. How Microbes Enhance Rice Growth and Yield: Mechanisms and Benefits
Microbes play diverse roles in influencing the morphology, physiology, growth, resilience, and gene expression patterns of plants [71]. The application of microorganisms in rice production is regarded as a sustainable and environment friendly approach that has gained attention in agricultural practices without the use of agrochemicals or mechanical interventions. Absolutely, and this phenomenon highlights the importance of beneficial bacteria in promoting plant growth through various mechanisms beyond nitrogen fixation. While nitrogen-fixing bacteria (for instance Rhizobium) are traditionally connected with leguminous plants, other groups of bacteria, like PGPR, can positively influence the growth and yield of non-leguminous crops, including rice. Biological nitrogen-fixation with Rhizobium sp. can significantly increase shoot and root growth, grain yield, increased their biomass, nutrient uptake and nitrogen-use efficiency in rice plants by activating the conversion of atmospheric nitrogen (N₂) into ammonia (NH₃) via the nitrogenase enzyme, which plants can readily assimilate, enhance their nitrogen availability [95]. Rice yields improved by 17.73% with the combined use of Rhodopseudomonas palustris and Bacillus subtilis due to changes in the soil bacterial community [96]. The use of microbial inoculants did not affect GHG emissions but enhanced grain yields by 6.0%–9.1%, reducing overall GHG intensity [105]. Kalkhajeh et al. [106] demonstrated that combining basal nitrogen fertilizer and straw-decomposing microbial inoculant (SDMI) significantly enhances wheat straw decomposition and rice yield in rice soil as application boosted microbial activity and respiration, particularly in the tillering stage, leading to higher straw decomposition and biomass.
Co-inoculation with nitrogen-fixing and PGPB, such as Rhodopseudomonas palustris and Bacillus subtilis, resulted in yield increases of up to 13.7% due to improved seed setting rates and altered microbial community structures [107]. The use of biofertilizers allows for a reduction in nitrogen fertilizer application while maintaining or even increasing rice yields by up to 26%, thus mitigating nitrogen losses and environmental impacts [108]. Methane-derived microbial biostimulants have been found to increase rice grain yield while simultaneously reducing greenhouse gas emissions, demonstrating a dual benefit for food security and environmental sustainability [109]. Different crop establishment methods combined with microbial formulations showed improved soil microbial properties and increased rice yield, particularly with specific combinations of nutrients and microbial inoculants [110]. The use of consortia of PGPB (P. agglomerans strain O4, P. putida strain P13 + P. agglomerans strain P5, P. koreensis strain S14 + P. vancouverensis strain S19) significantly enhanced rice grain yield, protein content, and nutrient uptake compared to untreated plants [111]. Coinoculation with Azospirillum brasilense and Pseudomonas fluorescens significantly increases rice yields while reducing mineral nitrogen use (30 kg of N ha–1), enhance nitrogen efficiency and economic production (37%). Inoculating rice with effective microbial agents increase crop performance by mobilizing nutrients, producing phytohormones, and providing protection against biotic and abiotic stresses, ultimately improve yield. The combination of Trichoderma microbial inoculant and rice straw compost significantly increased rice yield while reducing soil copper and lead concentrations in contaminated rice [112].
8. The Role of Microbes in Alleviating Biotic Stresses in Rice
The metaorganism concept, which encompasses plants and their associated microorganisms, has proven valuable in enhancing agricultural practices and disease management. In rice cultivation, this concept has been instrumental to promote biotic stress tolerance, and reduce the need for chemical pesticides, which, although effective, pose risks to the environment and human health [113]. Beneficial microbes in the rice rhizosphere establish mutualistic relationships that support plant growth and resilience against pathogens through mechanisms like competition, antibiosis, and the production of extracellular enzymes [114]. As biocontrol agents, these microbes mitigate plant diseases responsible for up to 25% global crop losses, particularly in developing regions [115]. Their interactions with rice plants facilitate adaptation to biotic stressors and enhance tolerance, providing a sustainable alternative to chemical interventions in agriculture [5].
Beneficial microorganisms, such as bacteria, fungi, and other microbes, play a pivotal role in safeguarding plants from pests, diseases, and pathogens. These microbes exert antagonistic and biocontrol effects, thereby enhancing plant defence mechanisms. Their interactions with plants, both direct and indirect, contribute significantly to the plant's resilience against infections and infestations. For instance, many researchers like [116,117,118,119,120,121,122,123] proved the efficacy of several strains of Bacillus subtilis to control the most devastating blast disease of rice caused by rice Magnaporthe oryzae. This bacterium produces antibiotics like surfactins and iturins that suppress the growth of blast fungal pathogen. Besides, these strains exhibit the ability to synthesize cell wall-degrading enzymes such as chitinase, protease, and β-1,3-glucanases, as well as antifungal metabolites such as siderophores. Known for inducing systemic resistance in rice, the bacterium Pseudomonas fluorescens can help manage bacterial blight caused by Xoo. P. fluorescens induces systemic resistance in rice by activating defence pathways through jasmonic acid and ethylene signalling, while producing antibiotics like DAPG and lytic enzymes that directly inhibit Xoo. Additionally, it promotes plant growth through phytohormone production and competes with pathogens for resources in the rhizosphere by producing siderophores and forming biofilms [124,125]. This beneficial fungus T. harzianum is used in rice fields for the management of Rhizoctonia solani, the causal agent of sheath blight in rice. T. harzianum acts through direct parasitism of the pathogen by secreting cell wall-degrading hydrolytic enzymes, and it also competes with the pathogen for nutrients. Several strains of Trichoderma have been proven effective against rice blast [126,127,128] and root rot of rice caused by Fusarium spp. [129,130], brown leaf spot disease caused by Bipolaris oryzae [131], rice sheath blight disease caused by Rhizoctonia solani [132]; sheath rot caused by S. oryzae [133]. Furthermore, the T. asperellum isolate demonstrated a pronounced ability to inhibit the growth of R. solani by means of hyphal coiling [134]. The nematophagous fungus Paecilomyces spp has shown efficacy in controlling rice root-knot nematodes (Meloidogyne graminicola), by parasitizing the nematode eggs and reducing their population. Pochonia chlamydosporia also works similar way [135]. PGPRs like Azospirillum brasilense is not only used to enhance rice growth but also to induce systemic resistance against diseases like rice blast and bacterial blight by priming the plant’s immune system [136]. PGPRs, such as Bacillus velezensis, B. megaterium, and B. toyonensis, significantly improved germination, seedling vigor, and dry weight. They also demonstrated the lowest disease incidence, relative lesion length, delayed sclerotia formation, and recorded maximum grain yield by suppressing pathogens through mechanisms like nutrient competition, production of antimicrobial compounds, and induction of systemic resistance [137]. A Bradyrhizobium japonicum strain capable of abscisic acid biosynthesis demonstrates potential as a biological control agent for bacterial wilt disease induced by Ralstonia solanacearum [138].
These biotic stresses, caused by the aforementioned devastating pathogens, lead to significant yield losses in rice production systems. Table 1 highlights the role of antagonistic microbes in suppressing phytopathogens, reducing diseases, promoting plant health, and enhancing resilience, offering sustainable and eco-friendly strategies for managing biotic stress challenges in rice cultivation.
9. Harnessing Microbes to Combat Abiotic Stresses in Rice
Rice is severely susceptible to abiotic stresses like salinity, drought, heat, cold, acidity, and sodicity, as well as biotic stresses caused by insects and pathogens (Figure 2). Plants experiencing stress can leverage beneficial microbial endophytes for survival. By emitting chemical signals, plants essentially "cry for help" activating these microbes to mitigate stress-related damage [139]. Microbes like Bacillus, Pseudomonas, and Trichoderma can alleviate these abiotic stresses by producing antioxidants, enzymes, antibiotics, and phytohormones (Table 2). Microbial involvement in rice plants is multifaceted, contributing to stress tolerance through various mechanisms. Whether through direct interactions with pathogens, modulation of plant physiology, or improvements in soil conditions, the plant-microbe partnership plays an essential role in enhancing the resilience of rice crops to both biotic and abiotic stressors [95].
Microbes from the genera Bacillus, Pseudomonas, Enterobacter, Ochrobactrum, Alcaligens, Paecilomyces, Burkholderia, Achromobacter, Azospirillum, and Glomus have been shown to mitigate abiotic stress in rice. These beneficial microorganisms achieve this by enhancing the production of antioxidants, hormones (ascorbate, proline, ethylene, and auxin), and stomatal conductance, as well as by synthesizing compounds like 1-aminocyclopropane-1-carboxylate deaminase, β-aminobutyric acid, salicylic acid, and siderophores. This collective action contributes significantly to the plant's resilience against environmental challenges [28].
Different microbial agents (endophytic and rhizospheric bacteria) exhibit varying mechanisms of salinity stress alleviation in rice, potentially enhancing photosynthetic efficiency, root architecture, and antioxidant enzyme activity viz. CAT, SOD, PO, PPO, APX, and PAL activity along with the effect on proline levels and also inhibits the activities of superoxide dismutase and lipid peroxidation. Bacillus haynesii 2P2 demonstrated the most significant improvement in biomass accumulation and tiller number, indicates a possible cultivar-specific microbial consortium for climate-resilient rice cultivation by stimulating tress responsive genes i.e. CATa, cAPX, MnSOD1 [140]. Furthermore, the rice plant's tolerance to salt was enhanced, and its capacity to generate glycine betaine was elevated. Bacillus subtilis and B. pumilus enhance plant growth in saline environments through their ability to solubilize phosphate and secrete the phytochemicals hydrogen cyanide (HCN), indole-3-acetic acid (IAA), and ammonia. Bacillus amyloliquefaciens is known to induce salt tolerance in rice plants by stimulating the production of phytohormones, including auxins and abscisic acid (ABA), as documented by Shahzad et al. [141] found that Nitrogenase activity and IAA was significantly increased by the application of PGPR (Azospirillum amazoense). In addition to this, Stenotrophomonas maltophilia application increase ACC deaminase, and phosphate solubilization in rice [142]. Both of the hose improves stress tolerance in rice plant. Curtobacterium sp, Enterobacter ludwigii, Bacillus cereus, and Micrococcus yunnanensis useful mitigate salinity stress in rice. Several investigations have demonstrated the beneficial impact of ACC deaminase-producing microorganisms, like Bacillus pumilus strain TUAT-1, on plant growth in saline conditions, as exemplified by their positive influence on rice [143]. Pseudomonas sp. inoculation enhance rice tolerance to salt stress [144], while Bacillus pumilus inoculation mitigate the adverse effects of both salt and high boron stress on rice plants [145].
Trichoderma and Pseudomonas species mitigated the adverse effects of water scarcity on rice plants. Microbial inoculants enhance the accumulation of polyphenolic compounds, known for their antioxidant properties. The activation of superoxide dismutase, coupled with the accumulation of hydrogen peroxide, was associated with hypersensitive cell death in leaves. Microbial inoculation also upregulates the activities of peroxidase, ascorbate peroxidase, glutathione peroxidase, and glutathione reductase enzymes, contribute to the reduction of reactive oxygen species (ROS) burden. Genes involved in key metabolic pathways, including phenylpropanoid biosynthesis, superoxide dismutation, hydrogen peroxide peroxidation, and oxidative defence, were overexpressed. These findings demonstrate the efficacy of microbial inoculants in bolstering the intrinsic biochemical and molecular capacities of rice plants to cope with drought stress [146]. Foliar spraying of Bacillus megaterium PB50, produce indole-3-acetic acid (IAA) which induces the synthesis of 1-aminocyclopropane-1-carboxylic acid (ACC) deaminase in the rice plants that enhance drought stress tolerance in the rice plants [147]. ACC deaminase-producing rhizobacteria have demonstrated their efficacy in mitigating drought stress in rice. Overexpressing the trehalose-6-phosphate synthase gene OsTPS1 in rice plants, using Escherichia coli or Saccharomyces cerevisiae, significantly enhanced their drought tolerance. The presence of Patescibacteria and the genera Massilia, Nocardioides, Frateuria, and Angustibacter, along with fungi belonging to Talaromyces, may enhance rice drought tolerance [148].
Chieb and Gachomo, [149] stated that the increased proline content in rice plants, due to inoculation with PGPR consortia, enhances the plants' tolerance to water stress. Application of Pseudomonas sp. K32 improves the Pb and Cd stress tolerance in rice. Pseudomonas species demonstrate the ability to remediate heavy metal contamination by detoxifying cadmium citrate and the Fe (III)-zinc complex through biotransformation processes. Saha et al. [150] reported that bacteria-produced siderophores can chelate not only ferric ions but also other metals, thereby assisting in phytoremediation. Consequently, PGPR can mitigate the adverse effects of heavy metal stress on plants. Inoculation with the Azotobacter brasilense strain A39 facilitated the accumulation of polyamines in rice seedlings subjected to osmotic stress, as demonstrated by [151]. Psychrophilic bacteria, including Arthrobacter nicotianae, Brevundimonas terrae, and Pseudomonas cedrina, demonstrated their efficacy to enhance plant growth and development even in adverse, and frigid environments. Extensive research has been undertaken to identify bacteria capable of producing organic acids that can solubilize insoluble phosphate [152]. In environments characterized by elevated temperatures, the bacterium Paecilomyces formosus exhibited traits that positively influenced plant growth. Similar to bacterial counterparts, endophytic and symbiotic fungi, including Curvularia protuberata, demonstrated the capacity to bolster plant heat tolerance and positively impact plant development.
In response to salt stress, T. harzianum increases rice plants' resistance by improving physiological parameters under saline conditions [95]. B. pumilus enhances tolerance to salt-stress tolerance in rice by up-regulating antioxidant enzymatic activities and accelerating certain soil enzyme activities [153]. Piriformospora indica regulates salt tolerance in rice by enhancing specific genes associated with salt-stress responses [154]. Bacterial endophyte P. alhagi enhances antioxidant enzyme activities and overall plant performance in salt-stress conditions [155].
10. Metagenomics: Unraveling the Complexities of Rice Microbial Communities
Metagenomics has emerged as a pivotal tool in understanding the intricate associations between microbes and plants, providing unprecedented insights into the complex dynamics of plant-associated microbial communities. DNA extraction from plant tissues is challenging due to the dominance of plant DNA over microbial DNA. Plant-associated microbes, especially endophytes, are under-investigated sources of bioactive molecules [156]. Metagenomics studies can help uncover beneficial functions of endophytes and identify potential beneficial species. DNA extraction methods for plant-associated microbial communities include surface sterilization and the use of commercial kits. Metagenomic techniques allow for the analysis of endophyte communities and their functional roles in plants. Metagenomic techniques involve profiling the microbial community using genomic samples from the natural habitat [72]. Comparative studies utilizing metagenomics have allowed the classification of microbes into different taxa based on gene libraries, highlighting significant differences among them [157]. In addition to metagenomic studies, other omics technologies, such as metabolomics and proteomics, have been employed to assess microbial diversity [158]. The success achieved through Next Generation Sequencing (NGS) in accessing metagenome sequences has been remarkable in various fields, including medicine, agriculture, pharmaceuticals, food, and environmental studies, facilitated by the use of different metagenomes. Metagenomic studies provide a comprehensive understanding of the taxonomic classification and functional/metabolic pathways of active endophytic microorganisms [159]. This approach is particularly valuable for determining the total microbiomes, including yet-to-be-cultured environmental microbes. Despite the computational challenges involved in the analysis and interpretation of metagenome sequenced data, the interest of scientists in metagenomics studies remains high.
The generation of longer sequence reads in metagenomics studies significantly influences the analysis of metagenomic sequences. Notably, real-time nanopore sequencing analysis has emerged in clinical metagenomic studies and has been applied in the detection of viral pathogens [160]. This technology allows for the direct sequencing of DNA without the need for amplification and holds promise for enhancing our understanding of microbial communities in diverse environments.
Metagenomic analyses provide a sophisticated approach for elucidating the intricate microbial communities that engage with plants. By leveraging the advantageous characteristics of beneficial microorganisms, this technology offers prospects for augmenting crop yield and robustness. High-throughput sequencing has become a prevalent tool for investigating the microbial communities associated with various crops and their interactions with environmental challenges [161,162]. Metagenomic binning offers a precise method for characterizing the taxonomic diversity and functional capabilities of microbial communities at a fine-grained level [163]. Numerous researchers [164,165,166,167,168] utilized Metagenomic to identify and analyse the genetic material of all the microorganisms present in a given environment. This helps in understanding the composition, functions, and interactions within the microbial communities. Cheng et al. [163] integrated metagenomics and machine learning to investigate the root-associated microbiome of two rice cultivars to identify clues for enhancing crop resilience to Cd stresses. Similarly, Li et al. [169] combined metagenomics with metabolomic analyses and found that Bt rice (T1C-1) planting increased soil microbiome diversity, and network stability, and influenced carbon and nitrogen cycling without adversely affecting probiotic or phytopathogenic microorganisms, though significant differences were observed in the rhizosphere compared to non-Bt cultivars. Additionally, Bt rice selectively modulated rhizosphere microbiota through altered root exudates, impacting soil metabolite profiles and providing mechanistic insights into the plant-microbe-environment interactions of genetically modified crops. Proteomics analysis of rice where Pseudomonas alcaliphila Ej2 was used as a biocontrol of blast under salt stress revealed rice proteomic profile, including metabolism, plant-pathogen interactions, and biosynthesis of unsaturated fatty acids significantly influenced by the applied bacterial strain [170]. Proteomic analysis of rice treated with Pseudomonas alcaliphila Ej2, as a biological control agent for blast disease under salt stress conditions, identified significant alterations in rice protein expression profiles related to metabolic processes, plant-pathogen interactions, and the biosynthesis of unsaturated fatty acids. These findings underscore the substantial influence of the applied bacterial strain on rice physiology [170]. Transcriptomics analysis is also used to map the functions of rice plant associated microbiome. Patel et al. [171] conducted a transcriptomic analysis of the endophytic bacterium Microbacterium testaceum and found that it can inhibit the growth of the rice blast fungus Magnaporthe oryzae through the release of volatile organic compounds and the upregulation of the OsNPR1 and OsCERK genes. A comparative analysis of microbiome composition in rice leaves using metabarcoding and culture-based methods identifies bacterial communities (Pantoea, Enterobacter, Pseudomonas, and Erwinia) associated with blast disease resistance. Quantitative RT-PCR analysis revealed elevated expression of defense-related genes, including OsCEBiP, OsCERK1, and phytohormone-associated genes like OsPAD4, OsEDS1, OsPR1.1, OsNPR1, OsPDF2.2, and OsFMO, in rice seedlings [172]. Nanfack et al. [63] utilized Illumina-based 16S rRNA gene sequencing to analyse the bacterial communities in two rainfed rice varieties, NERICA 3 and NERICA 8. Their findings revealed that healthy seedlings exhibited a higher abundance of beneficial genera, such as Brevundimonas, Sphingomonas, and Exiguobacterium. Conversely, abnormal seedlings displayed an increase in potentially harmful genera originating from the seed-associated microbiome. Krishnappa et al. [173] utilized 22 meta-barcoded NGS datasets to map the rice foliar microbiome across various plant surfaces, revealing a diverse microbiome of 157 genera, with Proteobacteria and Actinobacteria as dominant phyla. Besides their culturomics confirmed the prevalence of beneficial genera like Pantoea and Pseudomonas, highlighting potential microbial resources for microbiome-assisted rice cultivation. Sondo et al. [174] used similar techniques to analyse the diversity and plant growth promoting ability of rice root-associated bacteria in Burkina-Faso.
11. Microbiome Engineering: A Pathway to Sustainable Rice Cultivation
Plant microbiome engineering refers to the manipulation and modification of the microbial communities associated with plants [175]. Ongoing research in this field aims to optimize microbial interactions to benefit rice cultivation and contribute to global food security. There are different approaches to rice microbiome engineering, including biotechnological and conventional methods (Figure 3).
Liu et al. [176] outlined a protocol for CRISPR/Cas9 techniques, such as designing target sequences, constructing expression vectors, and transforming Pyricularia (Magnaporthe oryzae) for genome editing. These methods enable efficient gene disruption, base editing, and reporter gene knock-in without altering host components. Additionally, this protocol can facilitate the application of CRISPR/Cas technologies in various functional genomics studies of P. oryzae. The study conducted by Li et al. [177] successfully reported the increased plant resistance against blast disease caused by Magnaporthe oryzae by using targeted CRISPR/Cas9 mutation in ethylene responsive factor (ERF), OsERF922 in rice. For example, a recent study by Zafar et al. [178] revealed that a solitary amino acid substitution in the ALS gene, specifically the replacement of tryptophan with leucine at position 548, significantly improved the resistance of basmati rice to bacterial blight. OsSWEET14 gene editing by Sam et al. [179] improved BB resistance in rice. [180] enhanced blast disease resistance in rice through CRISPR/Cas9-mediated editing of the OsHDT701 gene. Similarly, [181] successfully improved disease resistance by targeting the Mildew Locus O (CaMLO2) gene using CRISPR/Cas9 technology [182] successfully developed rice plants resistant to the bacterial blight pathogen xoo by silencing the OsSWEET11, OsSWEET13, and OsSWEET14 genes, which regulate sugar transport within the plant. Ji et al. [183] demonstrated that the function of the bacterial blight resistance gene, xa23, could be restored by correcting gene mutations using the CRISPR-Cas9 system. This restored gene function triggered effective defense responses against Xoo infection. Engineering the endogenous xa23 gene by EBE stacking in its promoter broadens Xa23's defensive role against diverse bacterial pathogens, addressing the short-lived resistance typically conferred by a single R gene, which pathogens often overcome through mutations in effector genes [184]. CRISPR/Cas9-mediated editing of OsSWEET13, a susceptibility gene for bacterial blight in rice caused by Xanthomonas oryzae pv. Oryzae, generated two knockout mutants targeting its promoter, leading to enhanced tolerance against bacterial blight in rice [185]. IR64 rice lines were engineered to exhibit resistance to rice tungro spherical virus (RTSV) through targeted mutagenesis of the eukaryotic initiation factor 4G (eIF4G) gene. CRISPR/Cas9-mediated mutagenesis generated novel eIF4G alleles, leading to the development of RTSV-resistant rice varieties [186]. Plant annexins play a noteworthy role in plant improvement and defense against various environmental stresses, as highlighted by Shen et al. [187] who demonstrated the significance of the rice annexin gene OsAnn3 under cold stress through studies on OsAnn3 CRISPR knockouts. Shen et al. [188] employed CRISPR-based QTL editing to elucidate the roles of grain number (Gn1a) and grain size (GS3) QTLs in rice varieties. Shen et al. [189] previously engineered 571 OsALS genes and identified three mutant variants—ALSS627N and 1884G-A, ALSS627N, and ALSS627N/G628E—that demonstrated resistance to imidazole ethylnicotinic acid. The OsDST gene, engineered in the rice cultivar MTU1010 via CRISPR/Cas technology, exhibited augmented tolerance to both drought and salt stress, primarily due to increased leaf retention under drought conditions [190]. Furthermore, genetic engineering of genes such as OsBADH2, OsMPK2, SAPK2, and OsPDS has demonstrated potential for enhancing abiotic stress tolerance in rice [191]. Gene editing of OsHAK1, OsNramp5, and OsARM1 in rice has resulted in reduced accumulation of arsenic, cadmium, and calcium, respectively [192]. Furthermore, [193] showed that CRISPR-mediated editing of the OsRAV2 gene in rice improved salt stress tolerance. Some other research has utilized CRISPR-Cas9 to develop rice mutants resistant to salt stress by targeting OsDST [194], OsNAC45[195], AGO2 [196], and OsBBS1[197]. Table 3 further highlights innovative strategies for rice microbiome engineering aimed at enhancing rice productivity, stress tolerance, and climate resilience. These strategies include advanced tools such as gene editing, microbial inoculation, traditional rice plant breeding techniques, and high-throughput metagenomic sequencing. By integrating these approaches, the potential for sustainable improvements in rice cultivation to address global agricultural challenges is significantly amplified.
12. Microbiome-Shaping (M) Genes: Unlocking New Avenues for Stress-Resilient Traits
The rhizosphere (interface between roots and soil), phyllosphere (surface of leaves), and endosphere (internal tissues) are the three main compartments into which the plant microbiota can be divided [198]. The phyllosphere, or biological niche, which is made up of a variety of microorganisms, is the greatest biological habitat found in plant leaves. Controlling pathogenic bacteria and preserving the host's overall health depends on the existence of beneficial microbial members in the phyllosphere [139,199]. These beneficial microbes support innate immunity by serving as an additional line of defense [200]. Increasing microbiome homeostasis and manipulating the microbiota specifically have emerged as viable strategies for long-term preventative suppression of plant diseases [201,202].
A large body of literature indicates that a variety of plants, including Arabidopsis, barley, cucumbers, maize, and rice, have the ability to modify their microbiome through the expression of particular genes [203,204,205] According to reports, these genes that shape the microbiome help plants perform better and become more resilient to a variety of stresses. These stresses include defence against soilborne pathogens [206], coping with nitrogen deficiency, enhancing nitrogen acquisition [207] and striking a balance between growth and defence [208]. These genes all have the trait of being changed by the bacteria found in the rhizosphere or roots.
Su et al. [46] observed distinct phyllosphere microbial communities between indica and japonica rice varieties at various taxonomic levels, suggesting a potential influence of host genetics. They found four bacterial orders (Pseudomonadales, Burkholderiales, Xanthomonadales, and Enterobacterales) were particularly responsive to specific host genetic backgrounds, notably those linked to the phenylpropanoid biosynthesis pathway. Further investigation revealed haplotype differences in the OsPAL02 gene between indica and japonica, leading to differential 4-hydroxycinnamic acid (4-HCA) production, a key precursor for lignin biosynthesis. Plant-emitted VOCs can shape the phyllosphere microbial community by acting as antimicrobial agents or carbon sources, while the shaped microbial community can in turn influence plant physiology [208]. A recent study has revealed that rice employs a secondary metabolite biosynthetic gene to regulate the balance of its leaf microbiome. This finding suggests a promising avenue for developing disease-resistant crop varieties by manipulating microbiome-influencing genes [209]. This suggests that Ms genes, which are involved in the synthesis or release of secondary metabolites, may be particularly sensitive to environmental changes. This sensitivity could allow plants to modulate their interactions with beneficial microbes, thereby enhancing their ability to adapt to changing environmental conditions [161].
All of these data to the likelihood that the genes that shape the microbiome are spread and preserved in plant hosts, which were defined as "M genes” by [78]. In general, the use of M genes in crop cultivars that are naturally enriched in useful microbial taxa may provide new opportunities for the molecular breeding of desirable traits [210]. The M genes allow for direct disease-suppressive effects as well as the enrichment of disease-suppressive microbiota in the context of disease-resistant breeding. Rapidly evolving diseases are posing an increasing challenge to current breeding tactics, which rely on the accumulation of resistance genes [210]. However, a different kind of breeding relies on modifying the host's susceptibility genes. On the other hand, using M genes becomes a tactic that goes after the microbiota. This strategy might make it easier to produce crop varieties with longer-lasting disease resistance and a wider range of applications. Understanding how the M genes work in connection with the host plant's innate immunity to maintain microbiome homeostasis is still developing, even though the notion of the M gene has filled a knowledge gap regarding the interactions between pathogens, host plants, and the resident microbiota [78]. Using gene editing tools (such CRISPR-Cas9) to alter or incorporation of the M genes to the crop genome could be a rapid strategy to develop disease resistant cultivars. The discovery of M genes in various plants, including rice genotypes, presents an intriguing resource for engineering microbiomes through molecular breeding. This approach aims to maximize the benefits derived from the associated microorganisms. Further research is essential to elucidate the role of M genes in rice genotypes.
13. Overcoming Challenges and Exploring Future Prospects in Rice Microbiome Engineering
Microbial communities associated with rice are dynamic and influenced by various environmental factors such as soil type, climate, and plant genotype [211,212]. Notably, the rice microbiome is primarily shaped by microhabitat rather than geographic location, emphasizing the need for targeted interventions that consider the specific growing environments of rice [78]. Core endophytes, including Pantoea and Xanthomonas, play vital roles in nutrient uptake and stress tolerance; however, their capacity to antagonize pathogens is generally limited [78]. Moreover, different rice genotypes display distinct root microbiome architectures, which significantly influence plant growth and stress responses [213]. The rhizosphere of rice supports a complex ecosystem of microorganisms with intricate interactions, making the understanding and manipulation of these relationships a prominent challenge in microbiome engineering [214,215].
One of the key challenges is predicting and managing the dynamics of these microbial communities. While major rhizobacterial phyla can be cultivated for genome sequencing and phenotypic characterization, it remains difficult to assign specific functional roles to individual microbes or groups due to within-species variability. The study of plant-microbial interactions is further complicated by the predominance of unculturable microorganisms, which may harbor untapped potential for beneficial applications in sustainable agriculture, highlighting a relevant area for continued exploration.
Numerous methodologies have been developed to study soil microorganisms, including the traditional approach of cultivating microbes on solid or liquid media. This method allows for the isolation of specific microbial strains for biochemical and physiological evaluation. Recent advancements in culture-based techniques, such as dilution-to-extinction culturing, have contributed to a growing inventory of known soil microbial isolates, facilitating the identification of hundreds of heterotrophic bacterial taxa. Additionally, cutting-edge high-throughput cultivation methods, termed 'culturomics', harness machine learning and robotics to accelerate the isolation of microorganisms from microtiter plates [216].
Metagenomic techniques hold significant promise but also face limitations in accurately characterizing microbial communities within specific soil types and in detecting ecologically important, low-abundance species [217]. While DNA sequencing, such as 16S rRNA amplicon sequencing for bacteria and archaea, and 18S rRNA for eukaryotes, provides a comprehensive view of the soil microbiome—capturing both active and dormant cells—this method tends to include dead cells and primarily identifies taxonomic signatures without detailing functional potential [218]. Although high-throughput sequencing technologies ameliorate some depth-of-coverage challenges, they also impose substantial computational requirements for managing extensive datasets. Quantitative analyses become problematic without absolute abundance data, as many sequencing studies predominantly rely on relative abundance, making it difficult to discern true abundance shifts within communities. Additionally, the instability of RNA, combined with the complexities of extracting it from soil, presents further obstacles, as does the pre-eminence of ribosomal RNA that can obscure the sequencing of messenger RNA transcripts, which are critical for understanding functional dynamics. Inefficient protein extraction hinders the recovery and subsequent analysis of proteins expressed by soil microbial communities, thus limiting the depth of metaproteomic studies. However, emerging bioinformatics tools—such as DRAM, MEMPIS, and XCMS—provide promising avenues for the analysis and integration of omics datasets, enhancing our ability to identify specific species, genes, enzymes, and functions within the soil microbiome that could be leveraged for engineering applications.
Our understanding of the complex interactions between host plants, pathogens, and their associated microbiota is still evolving, particularly concerning the role of M genes and their interplay with the host plant’s innate immunity. Future research should investigate how these M genes might not only regulate metabolite levels but also influence overall microbiome composition, ultimately aiming to enhance crop yield and disease resistance. Climate change poses additional threats by disrupting microbial populations and influencing plant development, and urbanization introduces pollutants that predominantly impact phyllosphere microbiota. Nonetheless, microbiome engineering has the potential to foster sustainable agricultural practices through the enhancement of beneficial plant-microbial relationships. Investigating the rice microbiome may yield valuable insights into biofertilization and stress resilience. Continued research should focus on elucidating the complexities of microbial interactions, discovering novel strains, and integrating microbiome engineering with traditional breeding techniques to promote enhanced stress resilience and nutritional efficiency in rice varieties.
14. Conclusion
Rice-associated microbial communities play a pivotal role in advancing sustainable agricultural practices by enhancing crop productivity and resilience to environmental stresses. This review has underscored the critical functions of these microbiomes in promoting growth, managing biotic and abiotic stressors, and reducing dependence on chemical inputs. Through a detailed exploration of microbial diversity, signalling mechanisms, and stress management strategies, this study highlights the potential of microbiome engineering in transforming rice cultivation. Advances in metagenomics and microbial inoculation approaches provide valuable insights into the interactions between rice plants and their microbiomes, offering innovative solutions for enhancing biofertilization, stress tolerance, and crop health. While significant progress has been made, challenges remain in understanding the complexities of microbial interactions, environmental variability, and plant genotype influences. Future research should focus on characterizing microbial functions, exploring novel microbial associations, and integrating microbiome-based approaches with traditional breeding methods. By harnessing the potential of microbiome engineering and integrating it with advanced genomics and breeding, we can develop rice varieties capable of thriving in a changing climate. This collaborative approach paves the way for sustainable food production systems that address global challenges and secure food security for future generations.
Author Contributions
Conceptualization: TI, MNI and MNS; Original Draft Preparation: IJM and MOK; Review, Editing, and Improvement: MTI, MOK, MNI, DRN; Supervision: TI; All authors have read and agreed to the published version of the manuscript.
Funding and Acknowledgments
This manuscript is published as a featured article in the special issue (Microbiome-Driven Agriculture: Transforming Sustainability and Resilience through Functional Microbial Insights and Metagenomics) of MDPI Microbiology Journal, following an invitation from the editorial team of the journal and the special issue editors. The publication was honored with a full waiver of the manuscript processing fee.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding authors.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hornstein, E.D.; Sederoff, H. Back to the Future: Re-Engineering the Evolutionarily Lost Arbuscular Mycorrhiza Host Trait to Improve Climate Resilience for Agriculture. Crit. Rev. Plant Sci. 2024, 43(1), 1–33. [CrossRef]
- Thakur, N.; Nigam, M.; Mann, N.A.; Gupta, S.; Hussain, C.M.; Shukla, S.K.; Shah, A.A.; Casini, R.; Elansary, H.O.; Khan, S.A. Host-Mediated Gene Engineering and Microbiome-Based Technology Optimization for Sustainable Agriculture and Environment. Funct. Integr. Genomics 2023, 23(1), 57. [CrossRef]
- Silverstein, M.R.; Segrè, D.; Bhatnagar, J.M. Environmental Microbiome Engineering for the Mitigation of Climate Change. Glob. Change Biol. 2023, 29(8), 2050–2066. [CrossRef]
- Albright, M.B.N.; Louca, S.; Winkler, D.E.; et al. Solutions in Microbiome Engineering: Prioritizing Barriers to Organism Establishment. ISME J. 2022, 16(2), 331–338. [CrossRef]
- Afridi, M.S.; Ali, S.; Salam, A.; César Terra, W.; Hafeez, A.; Sumaira; Ali, B.; S. AlTami, M.; Ameen, F.; Ercisli, S.; et al. Plant Microbiome Engineering: Hopes or Hypes. Biology 2022, 11(12), 1782. [CrossRef]
- Zhao, J.; Yu, X.; Zhang, C.; Hou, L.; Wu, N.; Zhang, W.; Wang, Y.; Yao, B.; Delaplace, P.; Tian, J. Harnessing Microbial Interactions with Rice: Strategies for Abiotic Stress Alleviation in the Face of Environmental Challenges and Climate Change. Sci. Total Environ. 2023, 168847. [CrossRef]
- Beattie, G.A.; Bayliss, K.L.; Jacobson, D.A.; Broglie, R.; Burkett-Cadena, M.; Sessitsch, A.; Kankanala, P.; Stein, J.; Eversole, K.; Lichens-Park, A. From Microbes to Microbiomes: Applications for Plant Health and Sustainable Agriculture. Phytopathology® 2024, 114(8), 1742–1752. [CrossRef]
- Islam, M.T.; Rahman, M.M.; Pandey, P.; Boehme, M.H.; Haesaert, G. (Eds.) Bacilli and Agrobiotechnology: Phytostimulation and Biocontrol; Springer: Cham, 2019. [CrossRef]
- Mia, M.B.; Momotaj, A.; Islam, T. Consortia of Probiotic Bacteria and Their Potentials for Sustainable Rice Production. Sustainable Agrobiology: Design and Development of Microbial Consortia 2023, 151–176. [CrossRef]
- Fatema, K.; Mahmud, N.U.; Gupta, D.R.; Siddiqui, M.N.; Sakif, T.I.; Sarker, A., ... & Islam, T. Enhancing rice growth and yield with weed endophytic bacteria Alcaligenes faecalis and Metabacillus indicus under reduced chemical fertilization. Plos one 2024, 19(5), e0296547. [CrossRef]
- Saleem, A.; Anwar, S.; Nawaz, T.; Fahad, S.; Saud, S.; Ur Rahman, T.; Khan, M.N.; Nawaz, T. Securing a Sustainable Future: The Climate Change Threat to Agriculture, Food Security, and Sustainable Development Goals. J. Umm Al-Qura Univ. Appl. Sci. 2024, 1–7. [CrossRef]
- Yang, Y.; Tilman, D.; Jin, Z.; Smith, P.; Barrett, C.B.; Zhu, Y.G.; Burney, J.; D’Odorico, P.; Fantke, P.; Fargione, J.; Finlay, J.C. Climate Change Exacerbates the Environmental Impacts of Agriculture. Science 2024, 385(6713), eadn3747. [CrossRef]
- Uniyal, B.; Kosatica, E.; Koellner, T. Spatial and Temporal Variability of Climate Change Impacts on Ecosystem Services in Small Agricultural Catchments Using the Soil and Water Assessment Tool (SWAT). Sci. Total Environ. 2023, 875, 162520. [CrossRef]
- Alam, M.; Lou, G.; Abbas, W.; Osti, R.; Ahmad, A.; Bista, S.; Ahiakpa, J.K.; He, Y. Improving rice grain quality through ecotype breeding for enhancing food and nutritional security in Asia–Pacific region. Rice 2024, 17(1), 47. [CrossRef]
- Zhao, J.; Yu, X.; Zhang, C.; Hou, L.; Wu, N.; Zhang, W.; Wang, Y.; Yao, B.; Delaplace, P.; Tian, J. Harnessing microbial interactions with rice: Strategies for abiotic stress alleviation in the face of environmental challenges and climate change. Science of The Total Environment. 2023 Nov 28:168847. [CrossRef]
- He, X.; Batáry, P.; Zou, Y.; Zhou, W.; Wang, G.; Liu, Z.; Bai, Y.; Gong, S.; Zhu, Z.; Settele, J.; Zhang, Z. Agricultural Diversification Promotes Sustainable and Resilient Global Rice Production. Nat. Food 2023, 4(9), 788–796. [CrossRef]
- Jamal, M.R.; Kristiansen, P.; Kabir, M.J.; Lobry de Bruyn, L. Challenges and Adaptations for Resilient Rice Production under Changing Environments in Bangladesh. Land 2023, 12(6), 1217. [CrossRef]
- Sarkar, S.; Jaswal, A.; Singh, A. Sources of Inorganic Nonmetallic Contaminants (Synthetic Fertilizers, Pesticides) in Agricultural Soil and Their Impacts on the Adjacent Ecosystems. In Bioremediation of Emerging Contaminants from Soils; Elsevier: 2024; pp 135–161. [CrossRef]
- Khan, B.A.; Nadeem, M.A.; Nawaz, H.; Amin, M.M.; Abbasi, G.H.; Nadeem, M.; Ali, M.; Ameen, M.; Javaid, M.M.; Maqbool, R.; Ikram, M. Pesticides: Impacts on Agriculture Productivity, Environment, and Management Strategies. In Emerging Contaminants and Plants: Interactions, Adaptations, and Remediation Technologies; Springer: Cham, 2023; pp 109–134. [CrossRef]
- Hossain, M.E.; Shahrukh, S.; Hossain, S.A. Chemical Fertilizers and Pesticides: Impacts on Soil Degradation, Groundwater, and Human Health in Bangladesh. In Environmental Degradation: Challenges and Strategies for Mitigation; Springer International Publishing, 2022; pp 63–92. [CrossRef]
- Mohd Nizam, S.N.; Haji Baharudin, N.S.; Ahmad, H. Application of Pesticide in Paddy Fields: A Southeast Asia Case Study Review. Environ. Geochem. Health 2023, 45(8), 5557–5577. [CrossRef]
- Krishna, S.; Bhutia, D.D.; Chaubey, R.K.; Sudhir, I. Exploring Plant Microbiome: A Holistic Approach to Sustainable Agriculture. In Microbiome Drivers of Ecosystem Function; Academic Press: 2024; pp 61–77. [CrossRef]
- Sena, L.; Mica, E.; Valè, G.; Vaccino, P.; Pecchioni, N. Exploring the Potential of Endophyte-Plant Interactions for Improving Crop Sustainable Yields in a Changing Climate. Front. Plant Sci. 2024, 15, 1349401. [CrossRef]
- Vimal, S.R.; Singh, J.S.; Prasad, S.M. Crop Microbiome Dynamics in Stress Management and Green Agriculture. In Microbiome Drivers of Ecosystem Function; Academic Press: 2024; pp 341–366. [CrossRef]
- Mukherjee, A.; Singh, B.N.; Kaur, S.; Sharma, M.; de Araújo, A.S.F.; de Araujo Pereira, A. P., ... & Verma, J.F. Unearthing the power of microbes as plant microbiome for sustainable agriculture. Microbiological Research, 2024, 286, 127780. [CrossRef]
- Chang, W.; Chen, W.; Hu, Y.; Wang, Z. Bacillus altitudinis LZP02 improves rice growth by reshaping the rhizosphere microbiome. Plant and Soil. 2024 May;498(1), 279-94. [CrossRef]
- Ramakrishnan, B.; Maddela, N.R.; Venkateswarlu, K.; Megharaj, M. Potential of Microalgae and Cyanobacteria to Improve Soil Health and Agricultural Productivity: A Critical View. Environ. Sci. Adv. 2023, 2(4), 586–611. [CrossRef]
- Kumar, U.; Kaviraj, M.; Kundu, S.; Rout, S.; Priya, H.; Nayak, A.K. Microbial Alleviation of Abiotic and Biotic Stresses in Rice. In Sustainable Agriculture Reviews 60: Microbial Processes in Agriculture; Springer Nature Switzerland: 2023; pp 243–268. [CrossRef]
- Haney, C.H.; Samuel, B.S.; Bush, J.; Ausubel, F.M. Associations with Rhizosphere Bacteria Can Confer an Adaptive Advantage to Plants. Nat. Plants 2015, 1, 15051. [CrossRef]
- Compant, S.; Cassan, F.; Kostić, T.; Johnson, L.; Brader, G.; Trognitz, F.; Sessitsch, A. Harnessing the Plant Microbiome for Sustainable Crop Production. Nat. Rev. Microbiol. 2024, 1–5.
- Leontidou, K.; et al. Plant Growth Promoting Rhizobacteria Isolated from Halophytes and Drought-Tolerant Plants: Genomic Characterization and Exploration of Phyto-Beneficial Traits. Sci. Rep. 2020, 10, 14857. [CrossRef]
- Etesami, H.; Li, Z.; Maathuis, F.J.; Cooke, J. The Combined Use of Silicon and Arbuscular Mycorrhizas to Mitigate Salinity and Drought Stress in Rice. Environ. Exp. Bot. 2022, 201, 104955. [CrossRef]
- Das, D.; Ullah, H.; Himanshu, S.K.; Tisarum, R.; Cha-Um, S.; Datta, A. Arbuscular Mycorrhizal Fungi Inoculation and Phosphorus Application Improve Growth, Physiological Traits, and Grain Yield of Rice under Alternate Wetting and Drying Irrigation. J. Plant Physiol. 2022, 278, 153829. [CrossRef]
- Samaras, A.; Roumeliotis, E.; Ntasiou, P.; Karaoglanidis, G. Bacillus Subtilis MBI600 Promotes Growth of Tomato Plants and Induces Systemic Resistance Contributing to the Control of Soilborne Pathogens. Plants 2021, 10 (1113). [CrossRef]
- Yu, Y.; et al. Induced Systemic Resistance for Improving Plant Immunity by Beneficial Microbes. Plants 2022, 11 (386). [CrossRef]
- Ashnaei, S.P. Plant growth promoting rhizobacteria and Rhizophagus irregularis: Biocontrol of rice blast in wild type and mycorrhiza-defective mutant. J. Plant Prot. Res. 2019, 3, 362–375. [CrossRef]
- Islam, W.; et al. Insect-Fungal-Interactions: A Detailed Review on Entomopathogenic Fungi Pathogenicity to Combat Insect Pests. Microb. Pathog. 2021, 159, 105122. [CrossRef]
- Pathak, A.R. Exploring Microbial Dynamics in India’s Rice Fields: Investigation of Plant Growth-Promoting Rhizobacteria for Sustainable Agriculture. Int. J. Multidiscip. Res. 2024, 6(3). [CrossRef]
- Li, P.; Tian, Y.; Yang, K.; Tian, M.; Zhu, Y.; Chen, X.; Hu, R.; Qin, T.; Liu, Y.; Peng, S.; Yi, Z. Mechanism of Microbial Action of the Inoculated Nitrogen-Fixing Bacterium for Growth Promotion and Yield Enhancement in Rice (Oryza Sativa L.). Adv. Biotechnol. 2024, 2(4), 1–6.
- Purwani, J.; Pratiwi, E.; Sipahutar, I.A. The Effect of Different Species of Cyanobacteria on the Rice Yield and Nitrogen Use Efficiency Under Different Levels of Nitrogen Fertilizer on Alluvial West Java. IOP Conf. Ser. Earth Environ. Sci. 2021, 648(1), 012196. [CrossRef]
- Aasfar, A.; Bargaz, A.; Yaakoubi, K.; Hilali, A.; Bennis, I.; Zeroual, Y.; Meftah Kadmiri, I. Nitrogen fixing Azotobacter species as potential soil biological enhancers for crop nutrition and yield stability. Front. Microbiol. 2021, 12, 628379. [CrossRef]
- Singh, A.; Chauhan, P.S. N-Acyl Homoserine Lactone Mediated Quorum Sensing Exhibiting Plant Growth-Promoting and Abiotic Stress Tolerant Bacteria Demonstrates Drought Stress Amelioration. J. Pure Appl. Microbiol. 2022, 16(1). [CrossRef]
- Sammauria, R.; Kumawat, S.; Kumawat, P.; Singh, J.; Jatwa, T.K. Microbial Inoculants: Potential Tool for Sustainability of Agricultural Production Systems. Arch. Microbiol. 2020, 202, 677–693. [CrossRef]
- Setiawati, M.R.; Damayani, M.; Herdiyantoro, D.; Suryatmana, P.; Anggraini, D.; Khumairah, F.H. The Application Dosage of Azolla Pinnata in Fresh and Powder Form as Organic Fertilizer on Soil Chemical Properties, Growth and Yield of Rice Plant. AIP Conf. Proc. 2018, 1927, 030017.
- Vaid, S.K.; Kumar, B.; Sharma, A.; Shukla, A.; Srivastava, P. Effect of Zn Solubilizing Bacteria on Growth Promotion and Zn Nutrition of Rice. J. Soil Sci. Plant Nutr. 2014, 14, 889–910. [CrossRef]
- Su, P.; Kang, H.; Peng, Q.; Wicaksono, W.A.; Berg, G.; Liu, Z.; Ma, J.; Zhang, D.; Cernava, T.; Liu, Y. Microbiome Homeostasis on Rice Leaves Is Regulated by a Precursor Molecule of Lignin Biosynthesis. Nat. Commun. 2024, 15(1), 23. [CrossRef]
- Nanfack, A.D.; Nguefack, J.; Musonerimana, S.; La China, S.; Giovanardi, D.; Stefani, E. Exploiting the Microbiome Associated with Normal and Abnormal Sprouting Rice (Oryza sativa L.) Seed Phenotypes Through a Metabarcoding Approach. Microbiol. Res. 2024, 279, 127546. [CrossRef]
- Rana, R.; Nayak, P.K.; Madhavan, V.N.; Sonti, R.V.; Patel, H.K.; Patil, P.B. Comparative Genomics-Based Insights into Xanthomonas indica, a Non-Pathogenic Species of Healthy Rice Microbiome with Bioprotection Function. Appl. Environ. Microbiol. 2024, 90(9), e00848-24. [CrossRef]
- Eyre, A.W.; Wang, M.; Oh, Y.; Dean, R.A. Identification and Characterization of the Core Rice Seed Microbiome. Phytobiomes, J. 2019, 3(2), 148–157. [CrossRef]
- Guo, Y.; Kuzyakov, Y.; Li, N.; Song, B.; Liu, Z.; Adams, J.M.; Yang, L. Rice Rhizosphere Microbiome Is More Diverse but Less Variable Along Environmental Gradients Compared to Bulk Soil. Plant Soil 2024, 1–9. [CrossRef]
- Iniesta-Pallarés, M.; Brenes-Álvarez, M.; Lasa, A.V.; Fernández-López, M.; Álvarez, C.; Molina-Heredia, F.P.; Mariscal, V. Changes in Rice Rhizosphere and Bulk Soil Bacterial Communities in the Doñana Wetlands at Different Growth Stages. Appl. Soil Ecol. 2023, 190, 105013. [CrossRef]
- Wang, X.; He, S.W.; He, Q.; Ju, Z.C.; Ma, Y.N.; Wang, Z.; Han, J.C.; Zhang, X.X. Early Inoculation of an Endophyte Alters the Assembly of Bacterial Communities across Rice Plant Growth Stages. Microbiol. Spectrum 2023, 11(5), e04978-22. [CrossRef]
- Edwards, J.A.; Santos-Medellín, C.M.; Liechty, Z.S.; Nguyen, B.; Lurie, E.; Eason, S.; Phillips, G.; Sundaresan, V. Compositional Shifts in Root-Associated Bacterial and Archaeal Microbiota Track the Plant Life Cycle in Field-Grown Rice. PLoS Biol. 2018, 16(2), e2003862. [CrossRef]
- Xu, S.; Qiu, Q.; Yan, C.; Xiong, J. Structure, Acquisition, Assembly, and Function of the Root-Associated Microbiomes in Japonica Rice and Hybrid Rice. Agric. Ecosyst. Environ. 2024, 373, 109122. [CrossRef]
- Xiong, J.; Lu, J.; Li, X.; Qiu, Q.; Chen, J.; Yan, C. Effect of Rice (Oryza sativa L.) Genotype on Yield: Evidence from Recruiting Spatially Consistent Rhizosphere Microbiome. Soil Biol. Biochem. 2021, 161, 108395. [CrossRef]
- Wei, X.; Zhu, Z.; Wei, L.; Wu, J.; Ge, T. Biogeochemical Cycles of Key Elements in the Paddy-Rice Rhizosphere: Microbial Mechanisms and Coupling Processes. Rhizosphere 2019, 10, 100145. [CrossRef]
- Ding, Y.; Gao, X.; Shu, D.; Siddique, K.H.; Song, X.; Wu, P.; Li, C.; Zhao, X. Enhancing Soil Health and Nutrient Cycling through Soil Amendments: Improving the Synergy of Bacteria and Fungi. Sci. Total Environ. 2024, 923, 171332. [CrossRef]
- Zhang, F.; Deng, Y.; Peng, R.; Jiang, H.; Bai, L. Bioremediation of Paddy Soil with Amphitropic Mixture Markedly Attenuates Rice Cadmium: Effect of Soil Cadmium Removal and Fe/S-Cycling Bacteria in Rhizosphere. Sci. Total Environ. 2024, 915, 169876. [CrossRef]
- Murase, J.; Asiloglu, R. Protists: The Hidden Ecosystem Players in a Wetland Rice Field Soil. Biol. Fertil. Soils 2024, 60(6), 773–787. [CrossRef]
- Zhang, C.; Mi, W.; Xu, Y.; Zhou, W.; Bi, Y. Long-Term Integrated Rice-Crayfish Culture Disrupts the Microbial Communities in Paddy Soil. Aquac. Rep. 2023, 29, 101515. [CrossRef]
- Islam, M.T.; Deora, A.; Hashidoko, Y.; Rahman, A.; Ito, T.; Tahara, S. Isolation and Identification of Potential Phosphate Solubilizing Bacteria from the Rhizoplane of Oryza sativa, L. cv. BR29 of Bangladesh. Z. Naturforsch. C 2007, 62(1–2), 103–110. [CrossRef]
- Cao, X.; Liu, L.; Ma, Q.; Lu, R.; Kong, H.; Kong, Y.; Zhu, L.; Zhu, C.; Tian, W.; Jin, Q.; Wu, L. Optimum Organic Fertilization Enhances Rice Productivity and Ecological Multifunctionality via Regulating Soil Microbial Diversity in a Double Rice Cropping System. Field Crops Res. 2024, 318, 109569. [CrossRef]
- Khatibi, S.M.; Dimaano, N.G.; Veliz, E.; Sundaresan, V.; Ali, J. Exploring and Exploiting the Rice Phytobiome to Tackle Climate Change Challenges. Plant Commun. 2024, 3, 101078. [CrossRef]
- Nunna, S.A.; Balachandar, D. Rhizobacterial Community Structure Differs Between Landrace and Cultivar of Rice Under Drought Conditions. Curr. Microbiol. 2024, 81(10), 1–3. [CrossRef]
- Tan, L.T.; Dailin, D.J.; Hanapi, S.Z.; Rahman, R.A.; Mehnaz, S.; Shahid, I.; Ho, T.; El Ensahsy, H.A. Role of Microbiome on Healthy Growth and Yield of Rice Plant. In Plant Holobiome Engineering for Climate-Smart Agriculture; Springer Nature Singapore: 2024; pp 141–161.
- Islam, M.M.; Jana, S.K.; Sengupta, S.; Mandal, S. Impact of Rhizospheric Microbiome on Rice Cultivation. Curr. Microbiol. 2024, 81(7), 188.
- Adachi, A.; Utami, Y.D.; Dominguez, J.J.; Fuji, M.; Kirita, S.; Imai, S.; Murakami, T.; Hongoh, Y.; Shinjo, R.; Kamiya, T.; Fujiwara, T. Soil nutrition-dependent dynamics of the root-associated microbiome in paddy rice. bioRxiv 2024, 2024-09. [CrossRef]
- Juliyanti, V.; Itakura, R.; Kotani, K.; et al. Comparative Analysis of Root-Associated Microbes in Tropical Cultivated and Weedy Rice (Oryza spp.) and Temperate Cultivated Rice. Sci. Rep. 2024, 14, 9656. [CrossRef]
- Kim, H.; Lee, Y.H. The Rice Microbiome: A Model Platform for Crop Holobiome. Phytobiomes, J. 2020, 4(1), 5–18. [CrossRef]
- Dastogeer, K.M.; Tumpa, F.H.; Sultana, A.; Akter, M.A.; Chakraborty, A. Plant Microbiome–An Account of the Factors that Shape Community Composition and Diversity. Curr. Plant Biol. 2020, 23, 100161. [CrossRef]
- Doni, F.; Mispan, M.S.; Suhaimi, N.S.; Ishak, N.; Uphoff, N. Roles of Microbes in Supporting Sustainable Rice Production Using the System of Rice Intensification. Appl. Microbiol. Biotechnol. 2019, 103, 5131–5142. [CrossRef]
- Fadiji, A.E.; Babalola, O.O. Metagenomics Methods for the Study of Plant-Associated Microbial Communities: A Review. J. Microbiol. Methods 2020, 170, 105860. [CrossRef]
- Trivedi, Pankaj, Jan E. Leach, Susannah G. Tringe, Tongmin Sa, and Brajesh K. Singh. Plant–microbiome interactions: from community assembly to plant health. Nature reviews microbiology 18, no. 11 (2020):607-621. [CrossRef]
- Zhou, X.; Wang, J.T.; Zhang, Z.F.; Li, W.; Chen, W.; Cai, L.; 2020. Microbiota in the rhizosphere and seed of rice from China, with reference to their transmission and biogeography. Front. Microbiol. 11, 529571. [CrossRef]
- Jiang, H.; Luo, J.; Liu, Q.; Ogunyemi, S.O.; Ahmed, T.; Li, B.; Yu, S.; Wang, X.; Yan, C.; Chen, J.; Li, B. Rice Bacterial Leaf Blight Drives Rhizosphere Microbial Assembly and Function Adaptation. Microbiol. Spectr. 2023, 11, e01059-23. [CrossRef]
- Li, P.; Tian, Y.; Yang, K.; et al. Mechanism of Microbial Action of the Inoculated Nitrogen-Fixing Bacterium for Growth Promotion and Yield Enhancement in Rice (Oryza sativa L.). Adv. Biotechnol. 2024, 2, 32. [CrossRef]
- Pang, Z.; Zhao, Y.; Xu, P.; Yu, D. Microbial Diversity of Upland Rice Roots and Their Influence on Rice Growth and Drought Tolerance. Microorganisms 2020, 8, 1329. [CrossRef]
- Zhang, J.; Yao, Z.; Chen, Y.; Zhang, J.; Luo, S.; Tian, C.; Tian, L. Study of Rhizosphere Microbial Community Structures of Asian Wild and Cultivated Rice Showed That Cultivated Rice Had Decreased and Enriched Some Functional Microorganisms in the Process of Domestication. Diversity 2022, 14, 67. [CrossRef]
- Bhardwaj, L.; Reddy, B.; Nath, A.J.; Dubey, S.K. Influence of Herbicide on Rhizospheric Microbial Communities and Soil Properties in Irrigated Tropical Rice Field. Ecol. Indic. 2024, 158, 111534. [CrossRef]
- Hester, E.R.; Vaksmaa, A.; Valè, G.; Monaco, S.; Jetten, M.S.; Lüke, C. Effect of Water Management on Microbial Diversity and Composition in an Italian Rice Field System. FEMS Microbiol. Ecol. 2022, 98(3), fiac018. fiac018. [CrossRef]
- Wang, P.; Dai, J.; Luo, L.; Liu, Y.; Jin, D.; Zhang, Z.; Li, X.; Fu, W.; Tang, T.; Xiao, Y.; Hu, Y. Scale-Dependent Effects of Growth Stage and Elevational Gradient on Rice Phyllosphere Bacterial and Fungal Microbial Patterns in the Terrace Field. Front. Plant Sci. 2022, 12, 766128. [CrossRef]
- Zhang, R.S.; Wang, F.G.; Qi, Z.Q.; Qiao, J.Q.; Du, Y.; Yu, J.J.; Yu, M.N.; Liang, D.; Song, T.Q.; Yan, P.X.; Cao, H.J. Iturins Produced by Bacillus velezensis Jt84 Play a Key Role in the Biocontrol of Rice Blast Disease. Biol. Control 2022, 174, 105001. [CrossRef]
- Qi, Z.; Zhou, X.; Tian, L.; Zhang, H.; Cai, L.; Tang, F. Temporal and Spatial Variation of Microbial Communities in Stored Rice Grains from Two Major Depots in China. Food Res. Int. 2022, 152, 110876. [CrossRef]
- Yeo, F.K.; Cheok, Y.H.; Wan Ismail, W.N.; Kueh-Tai, F.F.; Lam, T.T.; Chong, Y.L. Genotype and Organ Effect on the Occupancy of Phyllosphere Prokaryotes in Different Rice Landraces. Arch. Microbiol. 2022, 204(10), 600. [CrossRef]
- Roman-Reyna, V.; Pinili, D.; Borja, F.N.; Quibod, I.L.; Groen, S.C.; Mulyaningsih, E.S.; Rachmat, A.; Slamet-Loedin, I.H.; Alexandrov, N.; Mauleon, R.; Oliva, R. The Rice Leaf Microbiome Has a Conserved Community Structure Controlled by Complex Host-Microbe Interactions. BioRxiv 2019, 615278. [CrossRef]
- Adeleke, B.S.; Ayilara, M.S.; Akinola, S.A.; Babalola, O.O. Biocontrol mechanisms of endophytic fungi. Egypt. J. Biol. Pest Control 2022, 32(1), 46. [CrossRef]
- Tian, Q.; Gong, Y.; Liu, S.; Ji, M.; Tang, R.; Kong, D.; Xue, Z.; Wang, L.; Hu, F.; Huang, L.; Qin, S. Endophytic bacterial communities in wild rice (Oryza officinalis) and their plant growth-promoting effects on perennial rice. Frontiers in Plant Science. 2023 Aug 14;14:1184489. [CrossRef]
- Wu, C.; Zhang, X.; Liu, Y.; Tang, X.; Li, Y.; Sun, T.; Yan, G.; Yin, C. Drought Stress Increases the Complexity of the Bacterial Network in the Rhizosphere and Endosphere of Rice (Oryza sativa L.). Agronomy 2024, 14, 1662. [CrossRef]
- Zhang, X.; Hu, Z.; Pan, H.; et al. Effects of Rare Earth Elements on Bacteria in Rhizosphere, Root, Phyllosphere and Leaf of Soil–Rice Ecosystem. Sci. Rep. 2022, 12, 2089. [CrossRef]
- Bao, X.; Zou, J.; Zhang, B.; Wu, L.; Yang, T.; Huang, Q. Arbuscular Mycorrhizal Fungi and Microbes Interaction in Rice Mycorrhizosphere. Agronomy 2022, 12(6), 1277. [CrossRef]
- Chareesri, A.; De Deyn, G.B.; Sergeeva, L.; Polthanee, A.; Kuyper, T.W. Increased Arbuscular Mycorrhizal Fungal Colonization Reduces Yield Loss of Rice (Oryza sativa L.) Under Drought. Mycorrhiza 2020, 30, 315–328. [CrossRef]
- Altaf, M.M.; Khan, M.S.; Abulreesh, H.H.; Ahmad, I. Quorum sensing in plant growth-promoting rhizobacteria and its impact on plant-microbe interaction. In Plant-Microbe Interactions in Agro-Ecological Perspectives: Volume 1: Fundamental Mechanisms, Methods and Functions; 2017; pp 311–331. [CrossRef]
- Chen, Y.; Liu, P.; Wu, J.; Yan, W.; Xie, S.; Sun, X.; Ye, B.C.; Chu, X. N-Acylhomoserine Lactonase-Based Hybrid Nanoflowers: A Novel and Practical Strategy to Control Plant Bacterial Diseases. J. Nanobiotechnol. 2022, 20(1), 347. [CrossRef]
- Kelbessa, B.G.; Dubey, M.; Catara, V.; Ghadamgahi, F.; Ortiz, R.; Vetukuri, R.R. Potential of Plant Growth-Promoting Rhizobacteria to Improve Crop Productivity and Adaptation to a Changing Climate. CABI Rev. 2023, 13(2023).
- Doni, F.; Suhaimi, N.S.M.; Mispan, M.S.; Fathurrahman, F.; Marzuki, B.M.; Kusmoro, J.; Uphoff, N. Microbial Contributions for Rice Production: From Conventional Crop Management to the Use of ‘Omics’ Technologies. Int. J. Mol. Sci. 2022, 23(2), 737. [CrossRef]
- Xiao, X.; Zhu, Y.; Gao, C.; Zhang, Y.; Gao, Y.; Zhao, Y. Microbial Inoculations Improved Rice Yields by Altering the Presence of Soil Rare Bacteria. Microbiol. Res. 2022, 254, 126910. [CrossRef]
- Viswanath, G.; Sekar, J.; Ramalingam, P.V. Detection of Diverse N-Acyl Homoserine Lactone Signaling Molecules Among Bacteria Associated with Rice Rhizosphere. Curr. Microbiol. 2020, 77(11), 3480–3491.
- Jamil, F.; Mukhtar, H.; Fouillaud, M.; Dufossé, L. Rhizosphere Signaling: Insights into Plant–Rhizomicrobiome Interactions for Sustainable Agronomy. Microorganisms 2022, 10(5), 899. [CrossRef]
- Frankenberger, W.T.; Arshad, M. Phytohormones in Soils: Microbial Production and Function; CRC Press: Boca Raton, FL, USA, 2020. [CrossRef]
- Khan, N.; Bano, A.; Ali, S.; Babar, M.A. Crosstalk Amongst Phytohormones from Planta and PGPR under Biotic and Abiotic Stresses. Plant Growth Regul. 2020, 90, 189–203. [CrossRef]
- Jiang, O.; Li, L.; Duan, G.; Gustave, W.; Zhai, W.; Zou, L.; An, X.; Tang, X.; Xu, J. Root Exudates Increased Arsenic Mobility and Altered Microbial Community in Paddy Soils. J. Environ. Sci. 2023, 127, 410–420. [CrossRef]
- Cai, G.; Shahbaz, M.; Ge, T.; Hu, Y.; Li, B.; Yuan, H.; Wang, Y.; Liu, Y.; Liu, Q.; Shibistova, O.; Sauheitl, L. Root Exudates with Low C/N Ratios Accelerate CO2 Emissions from Paddy Soil. Land Degrad. Dev. 2022, 33(8), 1193–1203. [CrossRef]
- Wang, L.; Qin, T.; Liu, T.; Guo, L.; Li, C.; Zhai, Z. Inclusion of Microbial Inoculants with Straw Mulch Enhances Grain Yields from Rice Fields in Central China. Food Energy Secur. 2020, 9(4), e230. [CrossRef]
- Zeng, Q.; Man, X.; Huang, Z.; Zhuang, L.; Yang, H.; Sha, Y. Effects of Rice Blast Biocontrol Strain Pseudomonas alcaliphila Ej2 on the Endophytic Microbiome and Proteome of Rice under Salt Stress. Front. Microbiol. 2023, 14, 1129614. [CrossRef]
- Wang, R.; Wang, H.L.; Tang, R.P.; Sun, M.Y.; Chen, T.M.; Duan, X.C.; Lu, X.F.; Liu, D.; Shi, X.C.; Laborda, P.; Wang, S.Y. Pseudomonas putida Represses JA- and SA-Mediated Defense Pathways in Rice and Promotes an Alternative Defense Mechanism Possibly through ABA Signaling. Plants 2020, 9(12), 1641. [CrossRef]
- Kalkhajeh, Y.K.; He, Z.; Yang, X.; Lu, Y.; Zhou, J.; Gao, H.; Ma, C. Co-Application of Nitrogen and Straw-Decomposing Microbial Inoculant Enhanced Wheat Straw Decomposition and Rice Yield in a Paddy Soil. J. Agric. Food Chem. 2021, 4, 100134. [CrossRef]
- Huang, K.; Yin, H.; Zheng, Q.; Lv, W.; Shen, X.; Ai, M.; Zhao, Y. Microbial Inoculation Alters Rhizoplane Bacterial Community and Correlates with Increased Rice Yield. Pedobiologia 2024, 104, 150945. [CrossRef]
- Hu, M.; Xue, H.; Wade, A.J.; Gao, N.; Qiu, Z.; Long, Y.; Shen, W. Biofertilizer Supplements Allow Nitrogen Fertilizer Reduction, Maintain Yields, and Reduce Nitrogen Losses to Air and Water in China Paddy Fields. Agric. Ecosyst. Environ. 2024, 362, 108850. [CrossRef]
- Kumar, S.R.; David, E.M.; Pavithra, G.J.; Sajith, G.K.; Lesharadevi, K.; Akshaya, S.; Bassavaraddi, C.; Navyashree, G.; Arpitha, P.S.; Sreedevi, P.; Zainuddin, K. Enabling Greenhouse Gas Emission Reduction while Improving Rice Yield with a Methane-Derived Microbial Biostimulant. bioRxiv 2024, . [CrossRef]
- Shahne, A.A.; Shivay, Y.S. Interactions of Microbial Inoculations with Fertilization Options and Crop Establishment Methods on Modulation of Soil Microbial Properties and Productivity of Rice. Int. J. Bio-resour. Stress Manag. 2024, 15(4), 1–17. [CrossRef]
- Ashouri, R.; Fallah, H.; Niknezhad, Y.; Barari Tari, D. Grain Quality and Yield Response of Rice to Application of Plant Growth-Promoting Bacteria and Amino Acids. J. Plant Nutr. 2023, 46(20), 4698–4709. [CrossRef]
- Cuevas, V.C.; Banaay, C.G. In Situ Bioremediation and Crop Growth Promotion Using Trichoderma Microbial Inoculant (TMI) Ameliorate the Effects of Cu Contamination in Lowland Rice Paddies. Philipp. J. Sci. 2022, 151(3), 1255–1265.
- Rani, L.; Thapa, K.; Kanojia, N.; Sharma, N.; Singh, S.; Grewal, A.S.; Srivastav, A.L.; Kaushal, J. An Extensive Review on the Consequences of Chemical Pesticides on Human Health and Environment. J. Clean. Prod. 2021, 283, 124657. [CrossRef]
- Raymaekers, K.; Ponet, L.; Holtappels, D.; Berckmans, B.; Cammue, B.P. Screening for Novel Biocontrol Agents Applicable in Plant Disease Management–A Review. Biol. Control 2020, 144, 104240. [CrossRef]
- Dean, R.A.; Talbot, N.J.; Ebbole, D.J.; Farman, M.L.; Mitchell, T.K.; Orbach, M.J.; Thon, M.; Kulkarni, R.; Xu, J.R.; Pan, H.; et al. The Genome Sequence of the Rice Blast Fungus Magnaporthe grisea. Nature 2005, 434, 980–986. [CrossRef]
- He, Y.; Zhu, M.; Huang, J.; Hsiang, T.; Zheng, L. Biocontrol Potential of a Bacillus subtilis Strain BJ-1 Against the Rice Blast Fungus Magnaporthe oryzae. Can. J. Plant Pathol. 2019, 41(1), 47–59. [CrossRef]
- Zhang, X.; Ma, Y.N.; Wang, X.; Liao, K.; He, S.; Zhao, X.; Guo, H.; Zhao, D.; Wei, H.L. Dynamics of Rice Microbiomes Reveal Core Vertically Transmitted Seed Endophytes. Microbiome 2022, 10(1), 216. [CrossRef]
- Koné, Y.; Alves, E.; da Silveira, P.R.; Cruz-Magalhães, V.; Botelho, F.B.; Ferreira, A.N.; Guimarães, S.D.; de Medeiros, F.H. Microscopic and Molecular Studies in the Biological Control of Rice Blast Caused by Pyricularia oryzae with Bacillus sp. BMH under Greenhouse Conditions. Biol. Control 2022, 172, 104983. [CrossRef]
- Lei, L.Y.; Xiong, Z.X.; Li, J.L.; Yang, D.Z.; Li, L.; Chen, L.; Zhong, Q.F.; Yin, F.Y.; Li, R.X.; Cheng, Z.Q.; Xiao, S.Q. Biological Control of Magnaporthe oryzae Using Natively Isolated Bacillus subtilis G5 from Oryza officinalis Roots. Front. Microbiol. 2023, 14, 1264000. [CrossRef]
- Zhu, Y.; Wang, Y.; He, X.; Li, B.; Du, S.; 2023. Plant growth-promoting rhizobacteria: A good companion for heavy metal phytoremediation. Chemosphere, 139475.
- Mirara, F.; Dzidzienyo, D.K.; Mwangi, M. Bacillus amyloliquefaciens D203 Ameliorates Rice Growth and Resistance to Rice Blast Disease. Cogent Food Agric. 2024, 10(1), 2371943. [CrossRef]
- Yang, Y.; Zhang, Y.; Zhang, L.; Zhou, Z.; Zhang, J.; Yang, J.; Gao, X.; Chen, R.; Huang, Z.; Xu, Z.; Li, L. Isolation of Bacillus siamensis B-612, a Strain That Is Resistant to Rice Blast Disease and an Investigation of the Mechanisms Responsible for Suppressing Rice Blast Fungus. Int. J. Mol. Sci. 2023, 24(10), 8513. [CrossRef]
- Chaowanaprasert, A.; Thanwisai, L.; Siripornadulsil, W.; Siripornadulsil, S. Biocontrol of Blast Disease in KDML105 Rice by Root-Associated Bacteria. Eur. J. Plant Pathol. 2024, 1–8. [CrossRef]
- Behera, S.; Behera, S.; Nayak, B.S. Management of Bacterial Leaf Blight of Rice Caused by Xanthomonas oryzae pv. oryzae Through an Integrated Approach in Western Undulating Zones of Odisha. J. Cereal Res. 2024, 16(1).
- Masnilah, R.; Nurmala, F.; Pradana, A.P. In Vitro Study of Phyllosphere Bacteria as Promising Biocontrol Agents Against Bacterial Leaf Blight Disease (Xanthomonas oryzae pv. oryzae) in Rice Plants. J-PEN Borneo: J. Ilmu Pertanian 2023, 6(2). [CrossRef]
- Rabnawaz, M.; Irshad, G.; Majeed, A.; Yousaf, M.; Javaid, R.A.; Siddique, F.; Rehman, A.; Hanif, A. Trichoderma harzianum as Growth Stimulator and Biological Control Agent Against Bacterial Leaf Blight (BLB) and Blast of Rice. Pakistan, J. Phytopathol. 2023, 35(2), 317–326. [CrossRef]
- Mahamadou, D.; Adounigna, K.; Amadou, H.B.; Oumarou, H.; Fousseyni, C.; Abdoulaye, H. Isolation and In-Vitro Assessment of Antagonistic Activity of Trichoderma spp. Against Magnaporthe oryzae Longorola Strain Causing Rice Blast Disease in Mali. Afr. J. Microbiol. Res. 2022, 16(2), 67–75. [CrossRef]
- Arellano, A.D.; da Silva, G.M.; Guatimosim, E.; da Rosa Dorneles, K.; Moreira, L.G.; Dallagnol, L.J. Seeds coated with Trichoderma atroviride and soil amended with silicon improve the resistance of Lolium multiflorum against Pyricularia oryzae. Biol. Control 2021, 154, 104499. [CrossRef]
- Prismantoro, D.; Akbari, S.I.; Permadi, N.; Dey, U.; Anhar, A.; Miranti, M.; Mispan, M.S.; Doni, F. The Multifaceted Roles of Trichoderma in Managing Rice Diseases for Enhanced Productivity and Sustainability. J. Agric. Food Res. 2024, 18, 101324. [CrossRef]
- Asad, S.A. Mechanisms of action and biocontrol potential of Trichoderma against fungal plant diseases—A review. Ecol. Complex. 2022, 49, 100978. [CrossRef]
- Priyadarshani, T.D.; Karunarathna, D.D.; Menike, G.D.; Weerasinghe, P.A.; Madushani, M.A.; Madhushan, K.W. Efficiency of Locally Isolated Trichoderma virens for Controlling Rice Brown Leaf Spot Disease Caused by Bipolaris oryzae. Int. Congr. Turk. Sci. Technol. Publ. 2023, 9–13.
- Safari Motlagh, M.R.; Jahangiri, B.; Kulus, D.; Tymoszuk, A.; Kaviani, B. Endophytic fungi as potential biocontrol agents against Rhizoctonia solani JG Kühn, the causal agent of rice sheath blight disease. Biology. 2022 Aug 29;11(9):1282. [CrossRef]
- Bora, B.; Ali, M.S. Evaluation of Microbial Antagonists Against Sarocladium oryzae Causing Sheath Rot Disease of Rice (Oryza sativa L.). Int. J. Curr. Microbiol. Appl. Sci. 2019, 8(7), 1755–1760. [CrossRef]
- Nurhayati, Y.; Suryanti, S.; Wibowo, A. In Vitro Evaluation of Trichoderma asperellum Isolate UGM-LHAF Against Rhizoctonia solani Causing Sheath Blight Disease of Rice. J. Perlind. Tanam. Indones. 2021, 25(1), 64–73. [CrossRef]
- Haque, Z.; Khan, M.R.; Zamir, S.; Pandey, K.; Rajana, R.N.; Gupta, N. Fungal Antagonists and Their Effectiveness to Manage the Rice Root-Knot Nematode, Meloidogyne graminicola. In Sustainable Management of Nematodes in Agriculture, Vol. 2: Role of Microbes-Assisted Strategies; Springer International Publishing, 2024; pp 237–247. [CrossRef]
- Ullah, Q.; Munir, T.; Mumtaz, T.; Chawla, M.; Amir, M.; Ismail, M.; Qasim, M.; Ghaffar, A.; Haidri, I. Harnessing Plant Growth-Promoting Rhizobacteria (PGPRs) for Sustainable Management of Rice Blast Disease Caused by Magnaporthe oryzae: Strategies and Remediation Techniques in Indonesia. Indonesian, J. Agric. Environ. Anal. 2024, 3(2), 65–76. [CrossRef]
- Patil, N.; Raghu, S.; Mohanty, L.; et al. Rhizosphere Bacteria Isolated from Medicinal Plants Improve Rice Growth and Induce Systemic Resistance in Host Against Pathogenic Fungus. J. Plant Growth Regul. 2024, 43, 770–786. [CrossRef]
- Chattopadhyay, P.; Banerjee, G.; Handique, P.J. Use of an Abscisic Acid-Producing Bradyrhizobium japonicum Isolate as Biocontrol Agent Against Bacterial Wilt Disease Caused by Ralstonia solanacearum. J. Plant Dis. Prot. 2022, 129, 869–879. [CrossRef]
- Liu, H.; Brettell, L.E.; Singh, B. Linking the Phyllosphere Microbiome to Plant Health. Trends Plant Sci. 2020, 25(9), 841–844. [CrossRef]
- Gupta, A.; Singh, A.N.; Tiwari, R.K.; Sahu, P.K.; Yadav, J.; Srivastava, A.K.; Kumar, S. Salinity Alleviation and Reduction in Oxidative Stress by Endophytic and Rhizospheric Microbes in Two Rice Cultivars. Plants 2023, 12, 976. [CrossRef]
- Shahzad, R.; Khan, A.L.; Bilal, S.; Waqas, M.; Kang, S.M.; Lee, I.J. Inoculation of Abscisic Acid-Producing Endophytic Bacteria Enhances Salinity Stress Tolerance in Oryza sativa. Environ. Exp. Bot. 2017, 136, 68–77. [CrossRef]
- Li, P.; Ye, S.; Chen, J.; Wang, L.; Li, Y.; Ge, L.; Wu, G.; Song, L.; Wang, C.; Sun, Y.; Wang, J. Combined Metagenomic and Metabolomic Analyses Reveal That Bt Rice Planting Alters Soil CN Metabolism. ISME Commun. 2023, 3(1), 4. [CrossRef]
- Win, K.T.; Oo, A.Z.; Yokoyama, T. Plant Growth and Yield Response to Salinity Stress of Rice Grown Under the Application of Different Nitrogen Levels and Bacillus pumilus Strain TUAT-1. Crops 2022, 2(4), 435–444. [CrossRef]
- Sen, S.; Chandrasekhar, C.N. Effect of PGPR on Growth Promotion of Rice (Oryza sativa L.) under Salt Stress. Asian, J. Plant Sci. Res. 2014, 4(5), 62–67.
- Khan, A.; Zhao, S.; Javed, M.; Khan, K.; Bano, A.; Shen, R.; Masood, S. Bacillus pumilus Enhances Tolerance in Rice (Oryza sativa L.) to Combined Stresses of NaCl and High Boron Due to Limited Uptake of Na. Environ. Exp. Bot. 2016, 124, 120–129. [CrossRef]
- Singh, D.P.; Singh, V.; Gupta, V.K.; et al. Microbial Inoculation in Rice Regulates Antioxidative Reactions and Defense-Related Genes to Mitigate Drought Stress. Sci. Rep. 2020, 10, 4818. [CrossRef]
- Devarajan, A.K.; Muthukrishanan, G.; Truu, J.; Truu, M.; Ostonen, I.; Kizhaeral, S.S.; et al. The Foliar Application of Rice Phyllosphere Bacteria Induces Drought-Stress Tolerance in Oryza sativa (L). Plants 2021, 10(2), 387. [CrossRef]
- Zuliang Lei, Yexin Ding, Weifeng Xu, Yingjiao Zhang, Microbial community structure in rice rhizosheaths under drought stress, Journal of Plant Ecology, Volume 16, Issue 5, October 2023, rtad012, . [CrossRef]
- Chieb, M.; Gachomo, E.W. The Role of Plant Growth Promoting Rhizobacteria in Plant Drought Stress Responses. BMC Plant Biol. 2023, 23, 407. [CrossRef]
- Saha, C.; Mukherjee, G.; Agarwal-Banka, P.; Seal, A. A Consortium of Non-Rhizobial Endophytic Microbes from Typha angustifolia Functions as Probiotic in Rice and Improves Nitrogen Metabolism. Plant Biol. 2016, 18(6), 938–946.
- Cassán, F.; Maiale, S.; Masciarelli, O.; Vidal, A.; Luna, V.; Ruiz, O. Cadaverine Production by Azospirillum brasilense and Its Possible Role in Plant Growth Promotion and Osmotic Stress Mitigation. Eur. J. Soil Biol. 2009, 45, 12–19. [CrossRef]
- de Almeida Leite, R.; Martins da Costa, E.; Cabral Michel, D.; do Amaral Leite, A.; de Oliveira-Longatti, S.M.; de Lima, W.; Konstantinidis, K.T.; de Souza Moreira, F.M. Genomic Insights into Organic Acid Production and Plant Growth Promotion by Different Species of Phosphate-Solubilizing Bacteria. World, J. Microbiol. Biotechnol. 2024, 40(10), 1–4.
- Kumar, A.; Singh, S.; Mukherjee, A.; Rastogi, R.P.; Verma, J.P. Salt-Tolerant Plant Growth-Promoting Bacillus pumilus Strain JPVS11 to Enhance Plant Growth Attributes of Rice and Improve Soil Health under Salinity Stress. Microbiol. Res. 2021, 242, 126616. [CrossRef]
- Jogawat, A.; Vadassery, J.; Verma, N.; Oelmüller, R.; Dua, M.; Nevo, E.; Johri, A.K. PiHOG1, a Stress Regulator MAP Kinase from the Root Endophyte Fungus Piriformospora indica, Confers Salinity Stress Tolerance in Rice Plants. Sci. Rep. 2016, 6(1), 36765. [CrossRef]
- Sun, L.; Lei, P.; Wang, Q.; Ma, J.; Zhan, Y.; Jiang, K.; Xu, Z.; Xu, H. The Endophyte Pantoea alhagi NX-11 Alleviates Salt Stress Damage to Rice Seedlings by Secreting Exopolysaccharides. Front. Microbiol. 2020, 10, 3112. [CrossRef]
- Adeleke, R.A.; Obieze, C.C.; Mukoro, C.; Chikere, C.B.; Tsipinana, S.; Nciizah, A. Phosphorus fertilizer application and tillage practices influence bacterial community composition: implication for soil health. Arch. Agron. Soil Sci. 2023, 69(5), 803–820. [CrossRef]
- Mandal, S.; Van Treuren, W.; White, R.A.; Eggesbø, M.; Knight, R.; Peddada, S.D. Analysis of Composition of Microbiomes: A Novel Method for Studying Microbial Composition. Microb. Ecol. Health Dis. 2015, 26(1), 27663. [CrossRef]
- Li, Z.; Shen, J.; Xu, Y.; Zhu, W. Metagenomic Analysis Reveals Significant Differences in Microbiome and Metabolic Profiles in the Rumen of Sheep Fed Low N Diet with Increased Urea Supplementation. FEMS Microbiol. Ecol. 2020, 96(10), fiaa117. [CrossRef]
- Forbes, J.D.; Knox, N.C.; Ronholm, J.; Pagotto, F.; Reimer, A. Metagenomics: The Next Culture-Independent Game Changer. Front. Microbiol. 2017, 8, 1069. [CrossRef]
- Hamza, I.A.; Bibby, K. Critical Issues in Application of Molecular Methods to Environmental Virology. J. Virol. Methods 2019, 266, 11–24. [CrossRef]
- Lv, L.; Huang, H.; Lv, J.; Xu, X.; Cao, D.; Rao, Z.; Geng, F.; Kang, Y. Unique Dissolved Organic Matter Molecules and Microbial Communities in Rhizosphere of Three Typical Crop Soils and Their Significant Associations Based on FT-ICR-MS and High-Throughput Sequencing Analysis. Sci. Total Environ. 2024, 919, 170904. [CrossRef]
- Feng, X.; Wang, Z.; Li, X.; Wang, W.; Gu, A.; Liu, Y. Analysis of Endophytic Bacterial Diversity in Rice Seeds with Regional Characteristics in Yunnan Province, China, Based on High-Throughput Sequencing Technology. Curr. Microbiol. 2023, 80(9), 287. [CrossRef]
- Cheng, Z.; Zheng, Q.; Shi, J.; He, Y.; Yang, X.; Huang, X.; Wu, L.; Xu, J. Metagenomic and Machine Learning-Aided Identification of Biomarkers Driving Distinctive Cd Accumulation Features in the Root-Associated Microbiome of Two Rice Cultivars. ISME Commun. 2023, 3(1), 14. [CrossRef]
- Masuda, S.; Gan, P.; Kiguchi, Y.; Anda, M.; Sasaki, K.; Shibata, A.; Iwasaki, W.; Suda, W.; Shirasu, K. Uncovering Microbiomes of the Rice Phyllosphere Using Long-Read Metagenomic Sequencing. Commun. Biol. 2024, 7(1), 357. [CrossRef]
- Ahmed, T.; Guo, J.; Noman, M.; Lv, L.; Manzoor, N.; Qi, X.; Li, B. Metagenomic and biochemical analyses reveal the potential of silicon to alleviate arsenic toxicity in rice (Oryza sativa L.). Environ. Pollut. 2024, 345, 123537. [CrossRef]
- Zhu, S.; Zhao, W.; Sun, S.; Yang, X.; Mao, H.; Sheng, L.; Chen, Z.; 2024. Community metagenomics reveals the processes of cadmium resistance regulated by microbial functions in soils with Oryza sativa root exudate input. Sci. Total Environ. 949, 175015. [CrossRef]
- Wang, C.; Chang, J.; Tian, L.; Sun, Y.; Wang, E.; Yao, Z.; Ye, L.; Zhang, H.; Pang, Y.; Tian, C. A Synthetic Microbiome Based on Dominant Microbes in Wild Rice Rhizosphere to Promote Sulfur Utilization. Rice. 2024 Dec;17(1):18. DOI. [CrossRef]
- Ge, L.; Mao, C.; Wu, Y.; Wang, L.; Chao, S.; Lv, B.; Ye, S.; Wang, X.; Zhao, K.; Chen, J.; Li, P. Soil Nutrient Cycling and Microbiome Responses to Bt Rice Cultivation. Plant Soil 2024, 1–6. [CrossRef]
- Li, J.; Wu, H.; Pu, Q.; Zhang, C.; Chen, Y.; Lin, Z.; Hu, X.; Li, O. Complete Genome of Sphingomonas paucimobilis ZJSH1, an Endophytic Bacterium from Dendrobium officinale with Stress Resistance and Growth Promotion Potential. Arch. Microbiol. 2023, 205(4), 132. [CrossRef]
- Zeng, Y.-L.; Lu, X.-J.; Wang, M.-R.; Zhu, J.-N.; Li, Q.-X.; Zhu, Q.-P.; Wang, L.; et al. Endophytic Acrocalymma vagum Drives the Microbiome Variations for Rice Health. 2023. [CrossRef]
- Patel, A.; Sahu, K.P.; Mehta, S.; Javed, M.; Balamurugan, A.; Ashajyothi, M.; Sheoran, N.; Ganesan, P.; Kundu, A.; Gopalakrishnan, S.; et al. New Insights on Endophytic Microbacterium-Assisted Blast Disease Suppression and Growth Promotion in Rice: Revelation by Polyphasic Functional Characterization and Transcriptomics. Microorganisms 2023, 11, 362. [CrossRef]
- Sahu, K.P.; Patel, A.; Kumar, M.; Sheoran, N.; Mehta, S.; Reddy, B.; Eke, P.; Prabhakaran, N.; Kumar, A. Integrated Metabarcoding and Culturomic-Based Microbiome Profiling of Rice Phyllosphere Reveal Diverse and Functional Bacterial Communities for Blast Disease Suppression. Front. Microbiol. 2021, 12, 780458.
- Krishnappa, C.; Mushineni, A.; Reddy, B.; et al. Fine-Scale Mapping of the Microbiome on Phylloplane and Spermoplane of Aromatic and Non-Aromatic Rice Genotypes. Folia Microbiol. 2023, 68, 889–910. [CrossRef]
- Sondo, M.; Wonni, I.; Koïta, K.; Rimbault, I.; Barro, M.; Tollenaere, C.; Moulin, L.; Klonowska, A. Diversity and Plant Growth Promoting Ability of Rice Root-Associated Bacteria in Burkina Faso and Cross-Comparison with Metabarcoding Data. PLOS ONE 2023, 18(11), e0287084. [CrossRef]
- Santoyo, G. How Plants Recruit Their Microbiome? New Insights into Beneficial Interactions. J. Adv. Res. 2022, 40, 45–58. [CrossRef]
- Liu, J.; Zhang, W.; Liu, Y.; Zhu, W.; Yuan, Z.; Su, X.; Ding, C. Differences in Phyllosphere Microbiomes Among Different Populus spp. in the Same Habitat. Front. Plant Sci. 2023, 14, 1143878. [CrossRef]
- Li, T.; Liu, B.; Spalding, M.H.; Weeks, D.P.; Yang, B. High-Efficiency TALEN-Based Gene Editing Produces Disease-Resistant Rice. Nat. Biotechnol. 2012. [CrossRef]
- Zafar, K.; Khan, M.Z.; Amin, I.; Mukhtar, Z.; Zafar, M.; Mansoor, S. Employing Template-Directed CRISPR-Based Editing of the OsALS Gene to Create Herbicide Tolerance in Basmati Rice. AoB Plants 2023, 15(2), plac059.
- Sam, V.H.; Van, P.T.; Ha, N.T.; Ha, N.T.T.; Huong, P.T.T.; Hoi, P. X., ... & Le Quyen, C. DESIGN AND TRANSFER OF OsSWEET14-EDITING T-DNA CONSTRUCT TO BAC THOM 7 RICE CULTIVAR. Journal of Biology/TẠp chí Sinh HỌc 2021, 43(1).
- Mathsyaraja, S.; Lavudi, S.; Vutukuri, P.R. Enhancing Resistance to Blast Disease Through CRISPR/Cas9 Gene Editing Technology in OsHDT701 Gene in RPBio-226 Rice Cv. (Oryza sativa L.). J. Appl. Biol. Biotechnol. 2024. [CrossRef]
- Park, J.H.; Kim, H. Harnessing CRISPR/Cas9 for Enhanced Disease Resistance in Hot Peppers: A Comparative Study on CAMLO2-Gene-Editing Efficiency Across Six Cultivars. Int. J. Mol. Sci. 2023, 24(23), 16775.
- Zafar, K.; Khan, M.Z.; Amin, I.; Mukhtar, Z.; Yasmin, S.; Arif, M.; Ejaz, K.; Mansoor, S. Precise CRISPR-Cas9 Mediated Genome Editing in Super Basmati Rice for Resistance Against Bacterial Blight by Targeting the Major Susceptibility Gene. Front. Plant Sci. 2020, 11, 575.
- Ji, Z.; Sun, H.; Wei, Y.; Li, M.; Wang, H.; Xu, J.; Lei, C.; Wang, C.; Zhao, K. Ectopic Expression of Executor Gene Xa23 Enhances Resistance to Both Bacterial and Fungal Diseases in Rice. Int. J. Mol. Sci. 2022, 23(12), 6545.
- Kumar, A.; Dubey, A. Rhizosphere Microbiome: Engineering Bacterial Competitiveness for Enhancing Crop Production. J. Adv. Res. 2020, 24, 337–352. [CrossRef]
- Xu, Z.; Xu, X.; Wang, Y.; Liu, L.; Li, Y.; Yang, Y.; Liu, L.; Zou, L.; Chen, G. A Varied AvrXa23-like TALE Enables the Bacterial Blight Pathogen to Avoid Being Trapped by Xa23 Resistance Gene in Rice. J. Adv. Res. 2022, 42, 263–272.
- Macovei, A.; Sevilla, N.R.; Cantos, C.; Jonson, G.B.; Slamet-Loedin, I.; Čermák, T.; Voytas, D.F.; Choi, I.R.; Chadha-Mohanty, P. Novel Alleles of Rice eIF4G Generated by CRISPR/Cas9-Targeted Mutagenesis Confer Resistance to Rice Tungro Spherical Virus. Plant Biotechnol. J. 2018, 16(11), 1918–1927.
- Shen, C.; Que, Z.; Xia, Y.; Tang, N.; Li, D.; He, R.; Cao, M. Knock Out of the Annexin Gene OsAnn3 via CRISPR/Cas9-Mediated Genome Editing Decreased Cold Tolerance in Rice. J. Plant Biol. 2017, 60, 539–547.
- Shen, L.; Wang, C.; Fu, Y.; Wang, J.; Liu, Q.; Zhang, X.; et al. QTL Editing Confers Opposing Yield Performance in Different Rice Varieties. J. Integr. Plant Biol. 2018, 60, 89–93. [CrossRef]
- Liying, Y.; Yuanye, Z.; Rongtian, L.; et al. Improvement of Herbicide Resistance in Rice by Using CRISPR/Cas9 System. Chin. J. Rice Sci. 2022, 36(5), 459.
- Ganie, S.A.; Wani, S.H.; Henry, R.; Hensel, G. Improving Rice Salt Tolerance by Precision Breeding in a New Era. Curr. Opin. Plant Biol. 2021, 60, 101996. 101996. [CrossRef]
- Ashokkumar, S.; Jaganathan, D.; Ramanathan, V.; Rahman, H.; Palaniswamy, R.; Kambale, R.; Muthurajan, R. Creation of Novel Alleles of Fragrance Gene OsBADH2 in Rice through CRISPR/Cas9 Mediated Gene Editing. PLoS One 2020, 15(8), e0237018. [CrossRef]
- Tang, L.; Mao, B.; Li, Y.; Lv, Q.; Zhang, L.; Chen, C.; et al. Knockout of OsNramp5 Using the CRISPR/Cas9 System Produces Low Cd-Accumulating Indica Rice without Compromising Yield. Sci. Rep. 2017, 7, 14438. [CrossRef]
- Duan, Y.B.; Li, J.; Qin, R.Y.; Xu, R.F.; Li, H.; Yang, Y.C.; Ma, H.; Li, L.; Wei, P.C.; Yang, J.B. Identification of a Regulatory Element Responsible for Salt Induction of Rice OsRAV2 through Ex Situ and In Situ Promoter Analysis. Plant Mol. Biol. 2016, 90, 49–62. DOI:. [CrossRef]
- Santosh Kumar, V.; Verma, R.K.; Yadav, S.K.; Yadav, P.; Watts, A.; Rao, M.; & Chinnusamy, V.CRISPR-Cas9 mediated genome editing of drought and salt tolerance (OsDST) gene in indica mega rice cultivar MTU1010. Physiology and Molecular Biology of Plants, 2020,26, 1099-1110. DOI:. [CrossRef]
- Zhang, X.; Long, Y.; Huang, J.; & Xia, J. OsNAC45 is involved in ABA response and salt tolerance in rice. Rice, 2020, 13, 1-13. DOI:. [CrossRef]
- Yin, W.; Xiao, Y.; Niu, M.; Meng, W.; Li, L.; Zhang, X., ... & Tong, H. ARGONAUTE2 enhances grain length and salt tolerance by activating BIG GRAIN3 to modulate cytokinin distribution in rice. The Plant Cell 2020, 32(7), 2292-2306. [CrossRef]
- Zeng, Y.; Cui, Y.; Zhang, Y.; Zhang, Y.; Liang, M.; Chen, H.; Lan, J.; Song, G.; Lou, J. The Initiation, Propagation and Dynamics of CRISPR-SpyCas9 R-Loop Complex. Nucleic Acids Res. 2018, 46(1), 350–361.
- Fitzpatrick, C.R.; Salas-González, I.; Conway, J.M.; Finkel, O.M.; Gilbert, S.; Russ, D.; Teixeira, P.J.; Dangl, J.L. The Plant Microbiome: From Ecology to Reductionism and Beyond. Annu. Rev. Microbiol. 2020, 74, 81–100. [CrossRef]
- Liu, Q.; Cheng, L.; Nian, H.; Jin, J.; Lian, T. Linking Plant Functional Genes to Rhizosphere Microbes: A Review. Plant Biotechnol. J. 2023, 21, 902–917. [CrossRef]
- Ma, L.; Rocha, F.I.; Lee, J.; Choi, J.; Tejera, M.; Sooksa-Nguan, T.; Boersma, N.; VanLoocke, A.; Heaton, E.; Howe, A. The impact of stand age and fertilization on the soil microbiome of Miscanthus× giganteus. Phytobiomes Journal. 2021 Apr 30;5(1):51-9. [CrossRef]
- Matsumoto, H.; Fan, X.; Wang, Y.; Kusstatscher, P.; Duan, J.; Wu, S.; Chen, S.; Qiao, K.; Wang, Y.; Ma, B.; Zhu, G. Bacterial Seed Endophyte Shapes Disease Resistance in Rice. Nat. Plants 2021, 7(1), 60–72. [CrossRef]
- Goossens, P.; Spooren, J.; Baremans, K.C.; Andel, A.; Lapin, D.; Echobardo, N.; Pieterse, C.M.; Van Den Ackerveken, G.; Berendsen, R.L. Congruent Downy Mildew-Associated Microbiomes Reduce Plant Disease and Function as Transferable Resistobiomes. bioRxiv 2023, . [CrossRef]
- Zhang, X.; Li, L.; Butcher, J.; Stintzi, A.; Figeys, D. Advancing Functional and Translational Microbiome Research Using Meta-Omics Approaches. Microbiome 2019, 7, 1–2. [CrossRef]
- Song, Y.; Wilson, A.J.; Zhang, X.C.; Thoms, D.; Sohrabi, R.; Song, S.; Geissmann, Q.; et al. FERONIA Restricts Pseudomonas in the Rhizosphere Microbiome via Regulation of Reactive Oxygen Species. Nat. Plants 2021, 7, 644–654. [CrossRef]
- Song, S.; Morales Moreira, Z.; Briggs, A.L.; Zhang, X.C.; Diener, A.C.; Haney, C.H. PSKR1 Balances the Plant Growth–Defence Trade-off in the Rhizosphere Microbiome. Nat. Plants 2023, 9(12), 2071–2084. [CrossRef]
- Song, S.; Liu, Y.; Wang, N.R.; Haney, C.H. Mechanisms in Plant–Microbiome Interactions: Lessons from Model Systems. Curr. Opin. Plant Biol. 2021, 62, 102003. [CrossRef]
- Farré-Armengol, G.; Filella, I.; Llusia, J.; Peñuelas, J. Bidirectional Interaction Between Phyllospheric Microbiotas and Plant Volatile Emissions. Trends Plant Sci. 2016, 21(10), 854–860.
- Zhan, C.; Wang, M. Disease Resistance Through M Genes. Nat. Plants 2024, 10, 352–353. [CrossRef]
- Zhan, C.; Matsumoto, H.; Liu, Y.; Wang, M. Pathways to Engineering the Phyllosphere Microbiome for Sustainable Crop Production. Nat. Food 2022, 3(12), 997–1004. [CrossRef]
- Huang, S.; Zha, X.; Fu, G. Affecting Factors of Plant Phyllosphere Microbial Community and Their Responses to Climatic Warming—A Review. Plants 2023, 12(16), 2891. [CrossRef]
- Shankar, N.; Shetty, P.; Melo, T.C.; Kesseli, R. Multi-Generation Ecosystem Selection of Rhizosphere Microbial Communities Associated with Plant Genotype and Biomass in Arabidopsis thaliana. Microorganisms 2023, 11(12), 2932. [CrossRef]
- Singh, A.; Kumar, M.; Chakdar, H.; Pandiyan, K.; Kumar, S.C.; Zeyad, M.T.; Singh, B.N.; Ravikiran, K.T.; Mahto, A.; Srivastava, A.K.; Saxena, A. Influence of Host Genotype in Establishing Root Associated Microbiome of Indica Rice Cultivars for Plant Growth Promotion. Front. Microbiol. 2022, 13, 1033158. [CrossRef]
- Haldar, S.; Mondal, S.; Kumari, A.; Ghosh, A.; Chattopadhyay, D.; Ghosh, A. Rhizosphere Microbiome Engineering. In Current Developments in Biotechnology and Bioengineering; Elsevier, 2022; pp 377–396. [CrossRef]
- Kumar, S.; Verma, R.K.; Yadav, S.K.; Yadav, P.; Watts, A.; Rao, M.; Chinnusamy, V. CRISPR-Cas9 Mediated Genome Editing of Drought and Salt Tolerance (OsDST) Gene in Indica Mega Rice Cultivar MTU1010. Physiol. Mol. Biol. Plants 2020, 26, 1099–1110.
- Huang, Y.; et al. High-Throughput Microbial Culturomics Using Automation and Machine Learning. Nat. Biotechnol. 2023. [CrossRef]
- McArdle, A.J.; Kaforou, M. Sensitivity of Shotgun Metagenomics to Host DNA: Abundance Estimates Depend on Bioinformatic Tools and Contamination Is the Main Issue. Access Microbiol. 2020, 2, acmi000104. [CrossRef]
- Jansson, J.K.; McClure, R.; Egbert, R.G. Soil Microbiome Engineering for Sustainability in a Changing Environment. Nat. Biotechnol. 2023, 41(12), 1716–1728. [CrossRef]
- Jain, A.; Chatterjee, A.; Das, S. Synergistic Consortium of Beneficial Microorganisms in Rice Rhizosphere Promotes Host Defense to Blight-Causing Xanthomonas oryzae pv. oryzae. Planta 2020, 252(6), 106. [CrossRef]
- Sukanya, S.; Thammasittirong, A.; Kittakoop, P.; Prachya, S.; Na-Ranong, S.; Thammasittirong, T. Antagonistic Activity Against Dirty Panicle Rice Fungal Pathogens and Plant Growth-Promoting Activity of Bacillus amyloliquefaciens BAS23. J. Microbiol. Biotechnol. 2018, 28(9), 1527–1535. [CrossRef]
- Masum, M.M.I.; Liu, L.; Yang, M.; Hossain, M.M.; Siddiqa, M.M.; Supty, M.E.; Ogunyemi, S.O.; Hossain, A.; An, Q.; Li, B. Halotolerant Bacteria Belonging to Operational Group Bacillus amyloliquefaciens in Biocontrol of the Rice Brown Stripe Pathogen Aidovorax oryzae. J. Appl. Microbiol. 2018, 125(6), 1852–1867. [CrossRef]
- Saikia, K.; Bora, L.C. Exploring Actinomycetes and Endophytes of Rice Ecosystem for Induction of Disease Resistance Against Bacterial Blight of Rice. Eur. J. Plant Pathol. 2021, 159, 67–79. [CrossRef]
- Elshakh, A.S.; Anjum, S.I.; Qiu, W.; Almoneafy, A.A.; Li, W.; Yang, Z.; Cui, Z.Q.; Li, B.; Sun, G.C.; Xie, G.L. Controlling and Defence-Related Mechanisms of Bacillus Strains Against Bacterial Leaf Blight of Rice. J. Phytopathol. 2016, 164(7–8), 534–546. [CrossRef]
- Yasmin, S.; Hafeez, F.Y.; Mirza, M.S.; Rasul, M.; Arshad, H.M.; Zubair, M.; Iqbal, M. Biocontrol of Bacterial Leaf Blight of Rice and Profiling of Secondary Metabolites Produced by Rhizospheric Pseudomonas aeruginosa BRp3. Front. Microbiol. 2017, 8, 287562. [CrossRef]
- Suárez-Moreno, Z.R.; Vinchira-Villarraga, D.M.; Vergara-Morales, D.I.; Castellanos, L.; Ramos, F.A.; Guarnaccia, C.; Degrassi, G.; Venturi, V.; Moreno-Sarmiento, N. Plant-Growth Promotion and Biocontrol Properties of Three Streptomyces spp. Isolates to Control Bacterial Rice Pathogens. Front. Microbiol. 2019, 10, 290. [CrossRef]
- de Sousa Oliveira, M.I.; Chaibub, A.A.; Sousa, T.P.; Cortes, M.V.; de Souza, A.C.; da Conceição, E.C.; de Filippi, M.C. Formulations of Pseudomonas fluorescens and Burkholderia pyrrocinia Control Rice Blast of Upland Rice Cultivated Under No-Tillage System. Biol. Control 2020, 144, 104153. [CrossRef]
- McNeely, D.; Chanyi, R.M.; Dooley, J.S.; Moore, J.E.; Koval, S.F. Biocontrol of Burkholderia cepacia Complex Bacteria and Bacterial Phytopathogens by Bdellovibrio bacteriovorus. Can. J. Microbiol. 2017, 63(4), 350–358. [CrossRef]
- Campos-Soriano, L.I.; García-Martínez, J.; Segundo, B.S. The Arbuscular Mycorrhizal Symbiosis Promotes the Systemic Induction of Regulatory Defence-Related Genes in Rice Leaves and Confers Resistance to Pathogen Infection. Mol. Plant Pathol. 2012, 13(6), 579–592. [CrossRef]
- Abbas, A.; Fu, Y.; Qu, Z.; Zhao, H.; Sun, Y.; Lin, Y.; Xie, J.; Cheng, J.; Jiang, D. Isolation and evaluation of the biocontrol potential of Talaromyces spp. against rice sheath blight guided by soil microbiome. Environ. Microbiol. 2021, 23(10), 5946–5961. [CrossRef]
- Someya, N.; Nakajima, M.; Watanabe, K.; Hibi, T.; Akutsu, K. Potential of Serratia marcescens Strain B2 for Biological Control of Rice Sheath Blight. Biocontrol Sci. Technol. 2005, 15(1), 105–109. [CrossRef]
- Vidhyasekaran, P.; Rabindran, R.; Muthamilan, M.; Nayar, K.; Rajappan, K.; Subramanian, N.; Vasumathi, K. Development of Powder Formulation of Pseudomonas fluorescens for Control of Rice Blast. Plant Pathol. 1997, 46, 291–297. [CrossRef]
- Yang, D.; Wang, B.; Wang, J.; Chen, Y.; Zhou, M. Activity and Efficacy of Bacillus subtilis Strain NJ-18 Against Rice Sheath Blight and Sclerotinia Stem Rot of Rape. Biol. Control 2009, 51(1), 61–65. [CrossRef]
- Kumar, K.V.K.; Yellareddygari, S.K.; Reddy, M.S.; Kloepper, J.W.; Lawrence, K.S.; Zhou, X. G.; ... & Miller, M. E. Efficacy of Bacillus subtilis MBI 600 Against Sheath Blight Caused by Rhizoctonia solani and on Growth and Yield of Rice. Rice Sci. 2012, 19(1), 55–63.
- Zhu, F.; Wang, J.; Jia, Y.; Tian, C.; Zhao, D.; Wu, X.; Liu, Y.; Wang, D.; Qi, S.; Liu, X.; Li, L.; 2021. Bacillus subtilis GB519 promotes rice growth and reduces the damages caused by rice blast fungus Magnaporthe oryzae. PhytoFrontiers™ 1(4), 330-338. [CrossRef]
- Hossain, M.T.; Khan, A.; Chung, E.J.; Rashid, M.H.; Chung, Y.R. Biological Control of Rice Bakanae by an Endophytic Bacillus oryzicola YC7007. Plant Pathol. J. 2016, 32(3), 228. [CrossRef]
- Chung, E.J.; Hossain, M.T.; Khan, A.; Kim, K.H.; Jeon, C.O.; Chung, Y.R. Bacillus oryzicola sp. nov.; an Endophytic Bacterium Isolated from the Roots of Rice with Antimicrobial, Plant Growth Promoting, and Systemic Resistance Inducing Activities in Rice. Plant Pathol. J. 2015, 31(2), 152. [CrossRef]
- Shao, Z.; Li, Z.; Fu, Y.; Wen, Y.; Wei, S. Induction of Defense Responses Against Magnaporthe oryzae in Rice Seedling by a New Potential Biocontrol Agent Streptomyces JD211. J. Basic Microbiol. 2018, 58(8), 686–697. [CrossRef]
- Chaibub, A.A.; de Carvalho, J.C.B.; de Sousa Silva, C.; Collevatti, R.G.; Gonçalves, F.J.; de Carvalho Barros Cortes, M.V.; de Filippi, M.C.C.; de Faria, F.P.; Lopes, D.C.B.; de Araújo, L.G. Defence Responses in Rice Plants in Prior and Simultaneous Applications of Cladosporium sp. During Leaf Blast Suppression. Environ. Sci. Pollut. Res. 2016, 23, 21554–21564. [CrossRef]
- Chaibub, A.A.; de Sousa, T.P.; de Araújo, L.G.; de Filippi, M.C.C. Molecular and Morphological Characterization of Rice Phylloplane Fungi and Determination of the Antagonistic Activity Against Rice Pathogens. Microbiol. Res. 2020, 231, 126353. [CrossRef]
- Subedi, S.; Bohara, A.K.; Thapa, S.; Timilsena, K. Safeguarding Rice Crops in Nepal: Unveiling Strategies Against the Yellow Stem Borer (Scirpophaga incertulas). Discover Agriculture 2024, 2(1), 64.
- Liu, X.; Matsumoto, H.; Lv, T.; Zhan, C.; Fang, H.; Pan, Q.; Xu, H.; Fan, X.; Chu, T.; Chen, S.; Qiao, K. Phyllosphere Microbiome Induces Host Metabolic Defence Against Rice False-Smut Disease. Nat. Microbiol. 2023, 8(8), 1419–1433. [CrossRef]
- Isawa, T.; Yasuda, M.; Awazaki, H.; Minamisawa, K.; Shinozaki, S.; Nakashita, H. Azospirillum sp. Strain B510 Enhances Rice Growth and Yield. Microbes Environ. 2010, 25(1), 58–61. [CrossRef]
- Yasuda, M.; Dastogeer, K.M.; Sarkodee-Addo, E.; Tokiwa, C.; Isawa, T.; Shinozaki, S.; Okazaki, S. Impact of Azospirillum sp. B510 on the Rhizosphere Microbiome of Rice under Field Conditions. Agronomy 2022, 12(6), 1367. [CrossRef]
- Harsonowati, W.; Astuti, R.I.; Wahyudi, A.T. Leaf Blast Disease Reduction by Rice-Phyllosphere Actinomycetes Producing Bioactive Compounds. J. Gen. Plant Pathol. 2017, 83, 98–108. [CrossRef]
- Krishanti, N.I.P.R.A.; Wahyudi, A.T.; Nawangsih, A.A. (2015) Non-Pathogenic Phyllosphere Bacteria Producing Bioactive Compounds as Biological Control of Xanthomonas oryzae pv. oryzae.
- Glick, B.R.; Gamalero, E. Recent Developments in the Study of Plant Microbiomes. Microorganisms 2021, 9(7), 1533. [CrossRef]
- Ke, J.; Wang, B.; Yoshikuni, Y. Microbiome Engineering: Synthetic Biology of Plant-Associated Microbiomes in Sustainable Agriculture. Trends Biotechnol. 2021, 39(3), 244–261. [CrossRef]
- Uroz, S.; Courty, P.E.; Oger, P. Plant Symbionts Are Engineers of the Plant-Associated Microbiome. Trends Plant Sci. 2019, 24(10), 905–916. [CrossRef]
- Sheth, R.U.; Cabral, V.; Chen, S.P.; Wang, H.H. Manipulating Bacterial Communities by In Situ Microbiome Engineering. Trends Genet. 2016, 32(4), 189–200. [CrossRef]
- Pozo, M.J.; Zabalgogeazcoa, I.; de Aldana, B.R.; Martinez-Medina, A. Untapping the potential of plant mycobiomes for applications in agriculture. Current opinion in plant biology. 2021 Apr 1;60: 102034. [CrossRef]
- Hartman, K.; van der Heijden, M.G.; Wittwer, R.A.; Banerjee, S.; Walser, J.C.; Schlaeppi, K. Cropping Practices Manipulate Abundance Patterns of Root and Soil Microbiome Members Paving the Way to Smart Farming. Microbiome 2018, 6, 1–4. [CrossRef]
- Shukla, N.; Awasthi, R.P.; Rawat, L.; Kumar, J. Biochemical and Physiological Responses of Rice (Oryza sativa L.) as Influenced by Trichoderma harzianum under Drought Stress. Plant Physiol. Biochem. 2012, 54, 78–88. [CrossRef]
- Pandey, V.; Ansari, M.W.; Tula, S.; Yadav, S.; Sahoo, R.K.; Shukla, N.; Bains, G.; Badal, S.; Chandra, S.; Gaur, A.K.; Kumar, A. Dose-Dependent Response of Trichoderma harzianum in Improving Drought Tolerance in Rice Genotypes. Planta 2016, 243, 1251–1264. [CrossRef]
- Bashyal, B.; Parmar, P.; Zaidi, N.; Aggarwal, R. Molecular Programming of Drought-Challenged Trichoderma harzianum-Bioprimed Rice (Oryza sativa L.). Front. Microbiol. 2021, 12. [CrossRef]
- Zaidi, N.; Singh, M.; Kumar, S.; Sangle, U.N.; Singh, R.S.; Prasad, R.; Singh, S.; Singh, S.; Yadav, A.; Singh, A.; Waza, S.; Singh, U. Trichoderma harzianum Improves the Performance of Stress-Tolerant Rice Varieties in Rainfed Ecologies of Bihar, India. Field Crops Res. 2017, 220, 97–104. [CrossRef]
- Morán-Diez, E.; Rubio, B.; Dominguez, S.; Hermosa, R.; Monte, E.; Nicolás, C. Transcriptomic Response of Arabidopsis thaliana After 24 h Incubation with the Biocontrol Fungus Trichoderma harzianum. J. Plant Physiol. 2012, 169(6), 614–620. [CrossRef]
- Rawat, L.; Singh, Y.; Shukla, N.; Kumar, J. Seed Biopriming with Salinity Tolerant Isolates of Trichoderma harzianum Alleviates Salt Stress in Rice: Growth, Physiological, and Biochemical Characteristics. J. Plant Pathol. 2012, 94, 353–365. [CrossRef]
- Jha, Y.; Subramanian, R.B. Characterization of Root-Associated Bacteria from Paddy and Its Growth-Promotion Efficacy. 3 Biotech 2014, 4, 325–330.
- Srivastava, S.; Bist, V.; Srivastava, S.; Singh, P.; Trivedi, P.; Asif, M.; Chauhan, P.; Nautiyal, C. Unraveling Aspects of Bacillus amyloliquefaciens Mediated Enhanced Production of Rice under Biotic Stress of Rhizoctonia solani. Front. Plant Sci. 2016, 7, 587. [CrossRef]
- Rajkumar, M.; Bruno, L.; Jeyakumar, R. Alleviation of Environmental Stress in Plants: The Role of Beneficial Pseudomonas spp. Crit. Rev. Environ. Sci. Technol. 2017, 47, 372–407. [CrossRef]
- Jha, Y.; Subramanian, R.; Patel, S. Combination of Endophytic and Rhizospheric Plant Growth Promoting Rhizobacteria in Oryza sativa Shows Higher Accumulation of Osmoprotectant Against Saline Stress. Acta Physiol. Plantarum 2011, 33, 797–802. [CrossRef]
- Ashwitha, K.; Rangeshwaran, R.; Vajid, N.; Sivakumar, G.; Jalali, S.; Rajalaksmi, K.; Manjunath, H. Characterization of Abiotic Stress Tolerant Pseudomonas spp. Occurring in Indian Soils. J. Biol. Control 2013, 27, 319–328.
- Tiwari, S.; Prasad, V.; Chauhan, P.; Lata, C. Bacillus amyloliquefaciens Confers Tolerance to Various Abiotic Stresses and Modulates Plant Response to Phytohormones through Osmoprotection and Gene Expression Regulation in Rice. Front. Plant Sci. 2017, 8, 1510. [CrossRef]
- Lee, Y.; Kim, B.; Lee, B.; Jo, K.; Lee, N.; Chung, C.; Lee, Y.; Lee, J. Purification and Characterization of Cellulase Produced by Bacillus amyoliquefaciens DL-3 Utilizing Rice Hull. Bioresour. Technol. 2008, 99(2), 378–386. [CrossRef]
- Kakar, K.; Ren, X.; Nawaz, Z.; Cui, Z.; Li, B.; Xie, G.; Hassan, M.; Ali, E.; Sun, G. A Consortium of Rhizobacterial Strains and Biochemical Growth Elicitors Improve Cold and Drought Stress Tolerance in Rice (Oryza sativa L.). Plant Biol. 2016, 18(3), 471–483. [CrossRef]
- Bisht, N.; Mishra, S.; Chauhan, P. Bacillus amyloliquefaciens Inoculation Alters Physiology of Rice (Oryza sativa, L. var. IR-36) through Modulating Carbohydrate Metabolism to Mitigate Stress Induced by Nutrient Starvation. Int. J. Biol. Macromol. 2019. [CrossRef]
- Naser, I.B.; Mahmud, N.U.; Sarker, A.; Hoque, M.N.; Islam, T. A Highly Salt-Tolerant Bacterium Brevibacterium sediminis Promotes the Growth of Rice (Oryza sativa L.) Seedlings. Stresses 2022, 2(3), 275–289. [CrossRef]
- Choi, J.; Choudhury, A.; Walitang, D.; Lee, Y.; Sa, T. ACC Deaminase-Producing Brevibacterium linens RS16 Enhances Heat-Stress Tolerance of Rice (Oryza sativa L.). Physiol. Plant. 2021. [CrossRef]
- Singh, N.; Marwa, N.; Mishra, J.; Verma, P.C.; Rathaur, S.; Singh, N. Brevundimonas diminuta Mediated Alleviation of Arsenic Toxicity and Plant Growth Promotion in Oryza sativa, L. Ecotoxicol. Environ. Saf. 2016, 125, 25–34. [CrossRef]
- Udayashankar, A.C.; Nayaka, S.C.; Reddy, M.S.; Srinivas, C. Plant Growth-Promoting Rhizobacteria Mediate Induced Systemic Resistance in Rice against Bacterial Leaf Blight Caused by Xanthomonas oryzae pv. oryzae. Biol. Control 2011, 59(2), 114–122. [CrossRef]
- Narayanasamy, S.; Thangappan, S.; Uthandi, S. Plant Growth-Promoting Bacillus sp. Cahoots Moisture Stress Alleviation in Rice Genotypes by Triggering Antioxidant Defense System. Microbiol. Res. 2020, 239, 126518. [CrossRef]
- Xu, X.; Xu, Z.; Ma, W.; et al. Tale-Triggered and ITale-Suppressed Xa1 Resistance to Bacterial Blight Is Independent of OsTfiiaγ1 or OsTfiiaγ5 in Rice. J. Exp. Bot. 2021. [CrossRef]
- Zhang, J.; Zhang, N.; Liu, Y.X.; Zhang, X.; Hu, B.; Qin, Y.; Xu, H.; Wang, H.; Guo, X.; Qian, J.; Wang, W. Root Microbiota Shift in Rice Correlates with Resident Time in the Field and Developmental Stage. Sci. China Life Sci. 2018, 61, 613–621. [CrossRef]
Figure 1.
Major types of rice plant microbiomes (endospheric, phyllospheric and rhizospheric).
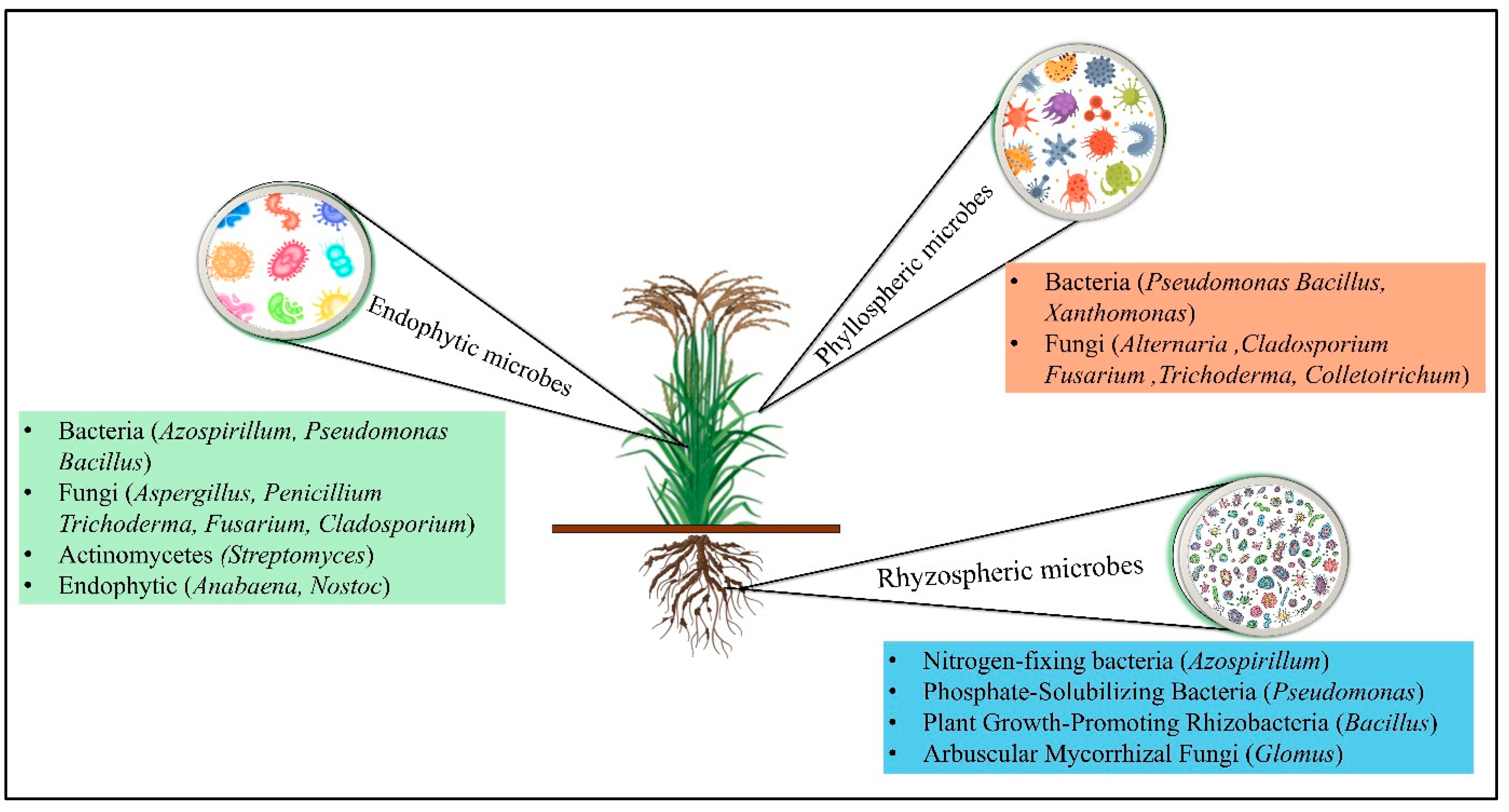
Figure 2.
Microbial involvement in abiotic/biotic stress tolerance in rice. Abbreviations, ABA (Abscisic acid), ROS (Reactive oxygen species), SAR (Systemic acquired resistance), ISR (Induced systemic resistance), ACC (1-Aminocyclopropane 1-carboxylic acid).
Figure 2.
Microbial involvement in abiotic/biotic stress tolerance in rice. Abbreviations, ABA (Abscisic acid), ROS (Reactive oxygen species), SAR (Systemic acquired resistance), ISR (Induced systemic resistance), ACC (1-Aminocyclopropane 1-carboxylic acid).
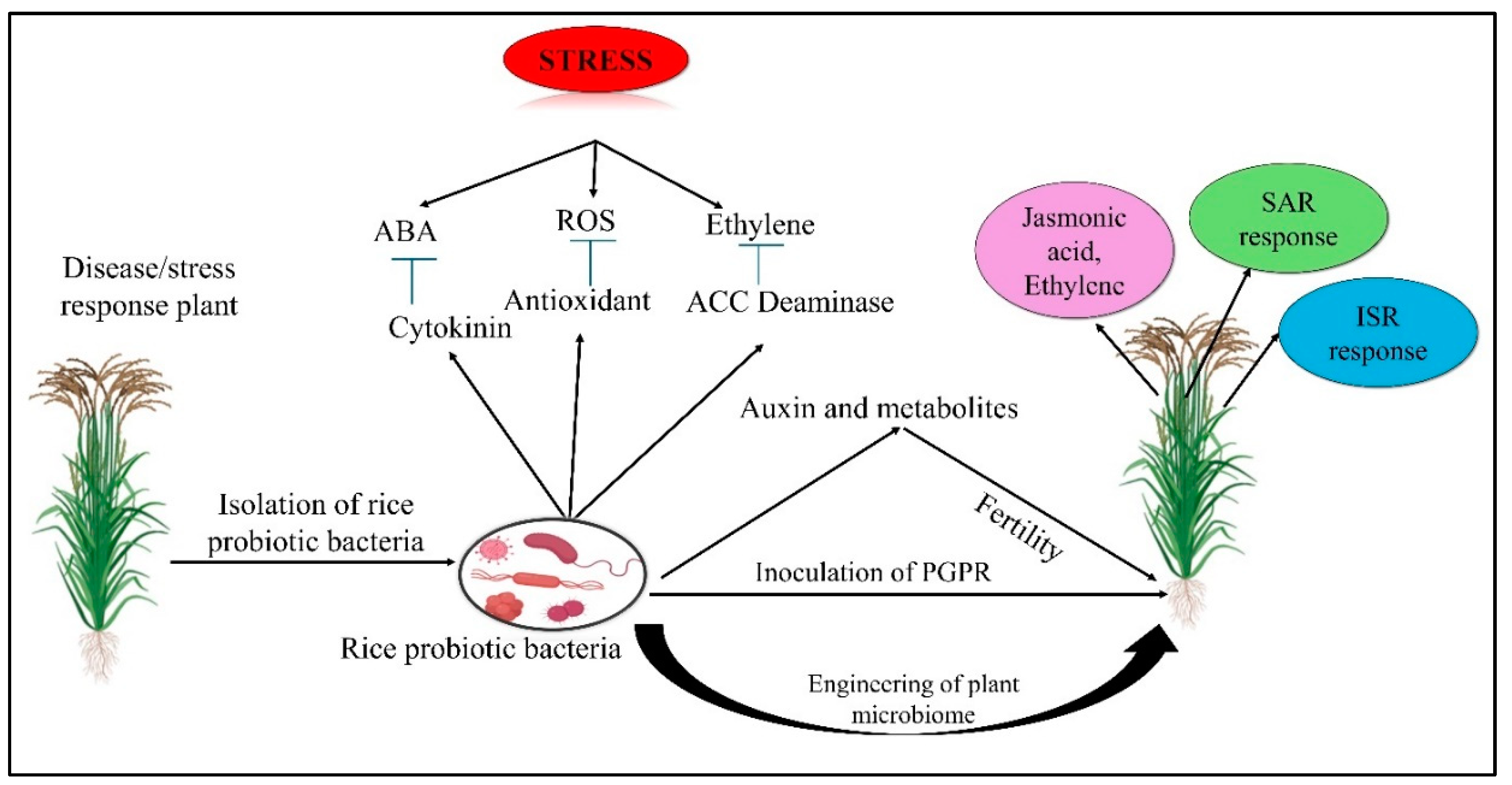
Figure 3.
Approaches to microbiome engineering in rice (including conventional and biotechnological methods).
Figure 3.
Approaches to microbiome engineering in rice (including conventional and biotechnological methods).
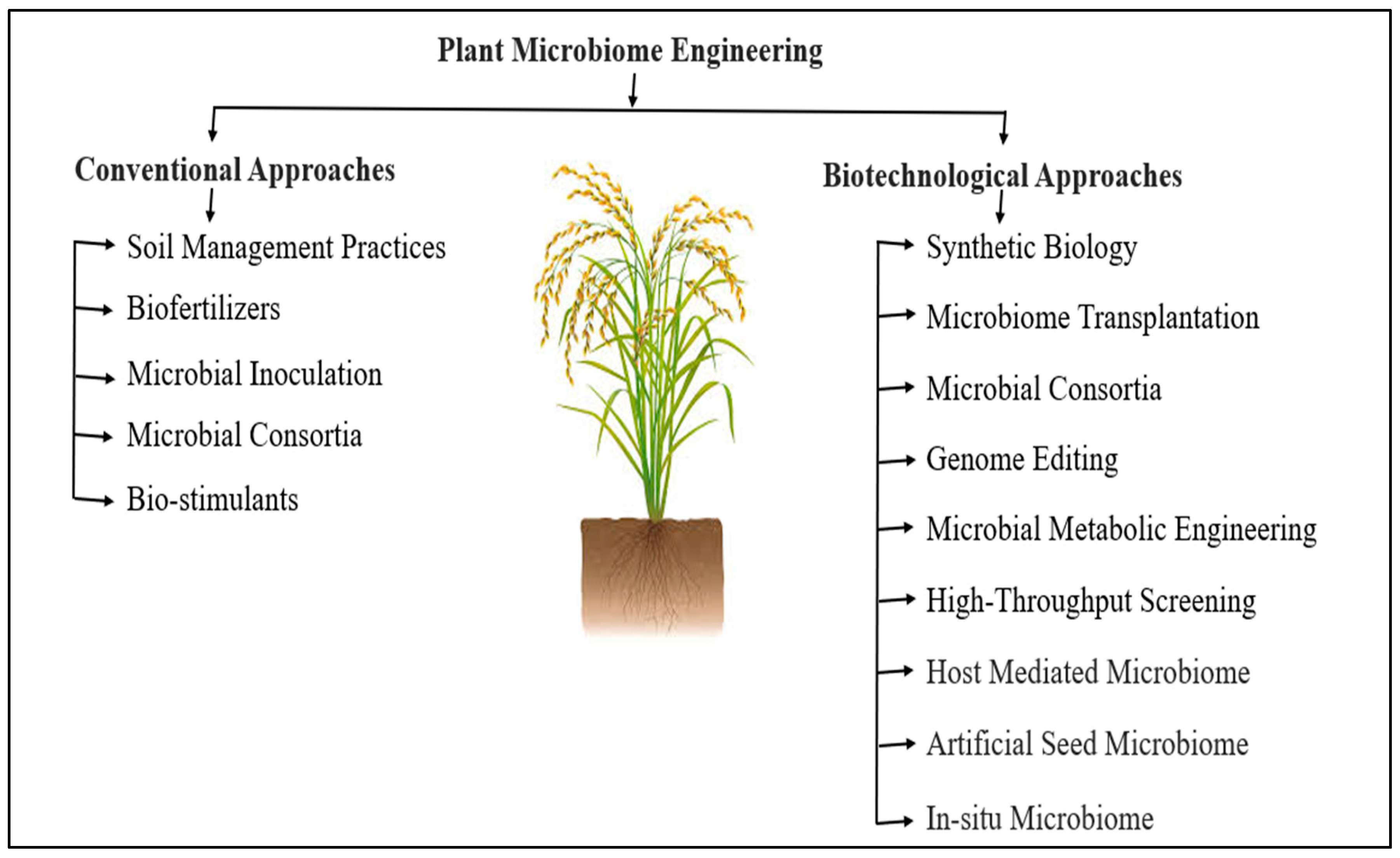

Table 1.
Beneficial microbes (bacteria) and their roles in biocontrol of rice diseases against numerous phytopathogens.
Table 1.
Beneficial microbes (bacteria) and their roles in biocontrol of rice diseases against numerous phytopathogens.
| Beneficial antagonistic microbes | Phytopathogens | Observed effects | References |
|---|---|---|---|
|
Bacillus amyloliquefaciens and Aspergillus pseudoporous |
Xanthomonas oryzae pv. oryzae |
Increase expression of defense related enzymes, proteins, and elevated levels of total phenols. | [219] |
| Curvularia lunata, Fusarium semitectum, and Helminthosporium oryzae | Suppress the germ tube elongation and mycelial development in fungal infections. | [220] | |
| Acidovorax oryzae | Cell membrane damage results in decreased cell count, biofilm development, and impaired swimming capability. | [221] | |
| Consortium of S. fimicarius, S. laurentii, P. putida, and Metarhizium anisopliae |
X. oryzae pv. oryzae |
Decrease the occurrence of leaf blight. | [222] |
|
B. subtilis, B.amyloliquefaciens, and B. methyltrophicus |
X. oryzae pv. oryzae | Active the defense-related enzymes. | [223] |
| P. aeruginosa |
X. oryzae pv. oryzae |
Active the defense enzymes | [224] |
| Streptomyces spp. | B. glumae | Inhibit the growth of B. glumae and promote plant growth. | [225] |
| P. fluorescens | M. oryzae | Reduce the physical damage caused by M. oryzae | [226] |
| Streptomyces spp. | M. oryzae | Increase in the activity of defensive enzymes. | [227] |
| Glomus intraradices | M. oryzae | Enhance the expression of defense-related genes. | [228] |
| Talaromyces spp. | R. solani | Increase the expression of defense-related genes and defense enzymes synthesis. | [229] |
| Serratia marcescens | R. solani | Decrease the occurrence of sheath blight | [230] |
| P. fluorescens | P. oryzae | Induce integrated stress response (ISR) in rice against P. oryzae | [231] |
| Serratia marcescens | R. solani | Reduce the frequency of sheath blight | [232] |
| R. solani | Decrease the occurrence of sheath blight and enhanced plant growth | [233,234] | |
| Bacillus subtilis |
M. oryzae (Rice blast) |
Reduce (Over 50%) blast disease, enhances systemic resistance, improves plant resilience | [78,114,116,143,235] |
| B. oryzicola | Gibberella fujikuroi | Decrease Bakanae severity by 46–78%. | [227] |
| B. glumae | Stimulate the resistance and enhancement of plant development. | [236] | |
| Glomus intraradices | M. oryzae | Increase expression of defense-response genes like OsNPR1, OsAP2, OsEREBP, and OsJAmyb. | [229] |
| Streptomyces spp. | M. oryzae | Enhance the defensive enzyme activity | [237] |
| Cladosporium cladosporioides | M. oryzae | Increase the enzyme activity and expression of defense-related genes like JIOsPR10, LOX-RLL, and PR1b. | [238,239] |
| Talaromyces spp. | R. solani | Increase expression of defense-related genes and activity of defense enzymes | [230] |
| B. subtilis; B. amyloliquefaciens; and B. methyltrophicus | X. oryzae pv. oryzae | Activate ISR leads to increase the activity of defense-related enzymes | [224] |
| P. aeruginosa | X. oryzae pv. oryzae | Increase the functions of defense-associated enzymes | [225] |
| Streptomyces spp. | B. glumae | Suppress B. glumae development | [226] |
| Consortium of S. fimicarius, S. laurentii, P. putida, and Metarhizium anisopliae | X. oryzae pv. oryzae | Decrease the occurrence of leaf blight | [223] |
| Bacillus thuringiensis |
Scirpophaga excerptalis (Rice stem borer) |
Reduces pest damage, enhances plant growth and yield | [240] |
| Pseudomonas fluorescens | Xanthomonas oryzae pv. oryzae (Bacterial blight) | Reduces bacterial blight, promotes plant growth, increases disease resistance | [124,125] |
| Azospirillum brasilense |
Magnaporthe oryzae (Rice blast), Xanthomonas oryzae (Bacterial blight) |
Enhances growth, induces resistance against multiple pathogens, improves disease tolerance | [136] |
| Bacillus velezensis |
Pyricularia oryzae (Rice blast), Bipolaris oryzae (Brown leaf spot) |
Reduces fungal disease incidence, enhances plant growth, improves grain yield | [137] |
| Bacillus megaterium | Fusarium spp. (Root rot) | Suppresses root rot, promotes plant growth, increases disease resistance | [137] |
| Bacillus toyonensis | Bipolaris oryzae (Brown leaf spot) | Reduces disease incidence, promotes growth, and improves yield | [137] |
| Bradyrhizobium japonicum | Ralstonia solanacearum (Bacterial wilt) | Controls bacterial wilt, enhances plant health and disease resistance | [138] |
| Trichoderma spp. |
Rhizoctonia solani (Sheath blight), Pyricularia oryzae (Rice blast), Fusarium spp. (Root rot), Bipolaris oryzae (Brown leaf spot) |
Suppresses fungal pathogens, promotes growth, reduces disease incidence | [126,127,128] |
| Trichoderma asperellum |
Rhizoctonia solani (Sheath blight) |
Suppresses fungal growth, reduces disease severity | [134] |
|
Lactobacillus spp. and Aspergillus spp. |
Ustilaginoidea virens (false-smut disease) | Reduced pathogen infection and disease severity in rice panicle | [241] |
| Azospirillum sp. | Various soilborne pathogens | Increased rice growth and yield, enhanced stress resistance | [242,243] |
| Saccharothrix spp. | [63,244,245] |
Table 2.
Microbes contributing to the improve tolerance to abiotic stresses in rice.
| Beneficial Microbes | Mechanism | References |
|---|---|---|
| Trichoderma harzianum | Improve root development in water scarcity like salinity stress | [246] |
| Increase the expression of aquaporin, dehydrin, and malondialdehyde genes, as well as other physiological factors. | [247] | |
| Improves seed germination and seedling growth at different stress conditions decreasing oxidative damage and lipid peroxidation. | [248] | |
| Enhance the phenol levels, peroxidase activity, lignin content, and cell membrane integrity | [249] | |
| Increase the levels of antioxidant enzymes and secondary metabolites in plants | [250] | |
| Improve gene expression associated with stress response | [251] | |
| Enhance efficiency of photosynthetic, antioxidant enzymes, and physiological adaptation in saline environments. | [252] | |
| Pseudomonas pseudoalcaligenes and Bacillus pumilus | Decrease the toxicity of reactive oxygen species (ROS) | [253] |
| Increase the amount of osmoprotectants in rice, like glycine betaine-like quaternary compounds, to help shoots grow more when they are under saline stress. | [145] | |
| Inhibit the absorption of Na+ ions.Synthesize growth related metabolites and enzymes | [104,254] | |
| Reduce sodium uptake in roots under saline stress conditions | [255] | |
| Protect cells from saline stress. | [256] | |
| Enhance the synergistic interaction among several PGPR strains. | [256] | |
| Reduce abiotic stress by increasing plant hormone, osmolytes, antioxidants, and growth-regulated genes. | [257] | |
| Bacillus amyloliquefaciens | Improve photosynthesis, hormone signalling, stress response, and carbohydrate metabolism | [258] |
| Enhance the synthesis of secondary metabolites, hormones, and enzymes. | [259,260] | |
| Increase the production of indole-3-acetic acid, siderophores, and cellulase under stress condition | [260] | |
| Increase biomass, relative water and proline content at stress conditions | [258,261] | |
| Brevibacterium sp. | Increase tolerance to salinity stress | [262] |
| Increase expression of stress linked genes | [263,264] | |
| Reduce ethylene release, and reactive oxygen species levels in rice | [263] | |
| Decrease arsenic absorption in rice plants and lower stress-related enzyme activity | [264] | |
| Bacillus sp. | Enhanced levels of phenylalanine ammonia lyase, peroxidase, and polyphenol oxidase to combat bacterial leaf blight | [264] |
| Enhance tolerance to water stress | [263] | |
| Inhibit sodium ion absorption and enhance antioxidant enzyme | [265] | |
| Enhance resistance to cold and drought stress | [260] |
Table 3.
Engineering rice microbiome using gene editing, inoculation, traditional breeding and metagenomic approaches for crop improvement.
Table 3.
Engineering rice microbiome using gene editing, inoculation, traditional breeding and metagenomic approaches for crop improvement.
| Approach | Method | Outcome | References |
|---|---|---|---|
| CRISPR/Cas9 Gene Editing | Gene editing to enhance disease resistance | CRISPR/Cas9 protocol for genome editing of Pyricularia oryzae (rice blast fungus). Enables gene disruption, base editing, and functional genomics | [176] |
| Bacterial blight resistance | Developed bacterial blight-resistant rice by silencing OsSWEET11, OsSWEET13, and OsSWEET14 genes, which regulate sugar transport in the plant. | [266] | |
| Resistance enhancement via CRISPR | Improved rice resistance to Magnaporthe oryzae by editing the OsHDT701 gene | [181] | |
| Blast disease resistance | Amino acid substitution in ALS gene of basmati rice significantly improves resistance to bacterial blight | [179] | |
| CRISPR editing for enhanced stress tolerance | Improved drought and salt stress tolerance in rice by editing the OsDST gene. | [191] | |
| Salt and abiotic stress tolerance | Improved salt stress tolerance in rice via CRISPR editing of OsRAV2 and OsDST genes. | [194,195] | |
| Reduced heavy metal accumulation | Reduced arsenic, cadmium, and calcium accumulation in rice by editing OsHAK1, OsNramp5, and OsARM1 genes. | [192] | |
| Microbial Inoculation | Beneficial microbial strains | Introduction of beneficial microbes (e.g., Pseudomonas, Bacillus) to enhance nitrogen fixation and suppress pathogens in rice. | [5] |
| Endophytic microbial engineering | Modification of endophytic bacteria to promote plant health and stress tolerance by enhancing the plant microbiome. | [175] | |
| Rhizosphere microbial community manipulation | Modulation of the plant rhizosphere microbiome to improve disease resistance and nutrient uptake in rice. | [181] | |
| Synthetic Biology | Engineering synthetic microbial consortia | Design and application of synthetic microbial communities to optimize plant-microbe interactions and improve plant resilience. | [176] |
| Microbe-engineered growth-promoting substances | Engineering microbes to produce beneficial compounds (e.g., antimicrobial peptides, growth regulators) to enhance plant health. | [5] | |
| Traditional Breeding | Selection for microbiome-supportive traits | Traditional breeding to select rice varieties that support beneficial microbial communities through exudate production or root architecture. | [189] |
| Breeding for enhanced plant-microbe interactions | Breeding rice varieties with traits that favor beneficial plant-microbe interactions, such as improved exudate profiles that attract beneficial microbes | [186] | |
| Metagenomics / Microbiome Profiling | High-throughput sequencing of microbiomes | Profiling rice microbiomes to identify beneficial microbes and determine how plant varieties impact microbial communities. | [189] |
| Microbial community analysis and optimization | Metagenomic analysis to identify microbial communities that enhance plant resilience to stresses like drought and disease. | [5] | |
| Environmental Modification | Soil amendments to enhance microbial diversity | Use of biochar, organic fertilizers, and other soil amendments to promote beneficial microbiomes in the rhizosphere of rice. | [196] |
| Fertilizer application to modulate microbiome | Modulation of the plant microbiome through strategic fertilizer application, enhancing nutrient availability and plant health. | [267] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



