
Submitted:
20 March 2024
Posted:
22 March 2024
You are already at the latest version
Abstract
Soil-borne Trichoderma spp. have been extensively studied for their biocontrol activities against pathogens and growth promotion ability with plants. However, the beneficial effect of Trichoderma on inducing resistance against insect herbivores has been considered only recently. Among diverse Trichoderma species, consistent with previous reports, we showed that root colonization by T. virens triggered induced systemic resistance (ISR) to leaf-infecting fungal pathogens. Whether T. virens induces ISR to insect pests has not been tested before. In this study, we investigated whether T. virens affects jasmonic acid (JA) biosynthesis and defense against fall armyworm (FAW) and western corn rootworm (WCR). Unexpectedly, the results showed that T. virens colonization of maize seedlings grown in autoclaved soil suppressed wound-induced production of JA, resulting in reduced resistance to FAW. Similarly, the bacterial endophyte Pseudomonas chlororaphis 30-84 suppressed systemic resistance to FAW due to reduced JA. Further comparative analyses of the systemic effects of these endophytes when applied into sterile or non-sterile field soil showed that both T. virens and P. chlororaphis 30-84 triggered ISR against hemibiotrophic fungal pathogen Colletotrichum graminicola in both soil conditions, but only suppressed resistance to FAW in sterile soil. In contrast to the effect on FAW defense, T. virens colonization of maize roots suppressed WCR larvae survival and weight gain. This is the first report suggesting the potential role of T. virens as a biocontrol agent against WCR.
Keywords:
Trichoderma virens
; Pseudomonas chlororaphis
; induced systemic resistance
; jasmonic acid
; Spodoptera frugiperda
; Diabrotica virgifera virgifera
1. Introduction
Maize is the top-yielding cereal crop produced globally and a major source of livestock feed, biofuel, and staple food for many regions worldwide [1]. Pathogen infection and insect herbivory are constant threats to crop production causing over 20% yield losses of maize annually at a global level, resulting in substantial economic impact and food insecurity [2]. The hemibiotrophic fungal pathogen Colletotrichum graminicola is the causal agent of anthracnose leaf blight and stalk rot of maize and accounts for millions of dollars in yield losses annually in the United States [3-5]. Fall armyworm (FAW; Spodoptera frugiperda) originates in the Americas and has recently invaded Europe, Africa, Asia, and Australia causing serious damage [6, 7]. FAW larvae feed on all the aboveground tissue of maize, resulting in several billions of dollars in yield losses globally [6]. Western corn rootworm (WCR; Diabrotica virgifera virgifera) is the most economically important maize pest in the United States and causes losses in excess of US $1 billion annually in the United States and Europe [8-10]. WCR larvae cause the most significant damage through feeding on the corn root while WCR adult beetles damage the silks [8, 11]. Published evidence indicates that the plant phytohormone salicylic acid (SA) plays a critical role in maize defense against C. graminicola [12, 13], while resistance to chewing insects, including FAW, is mainly mediated by the lipid-derived phytohormone jasmonic acid (JA) [14-16]. However, resistance mechanisms against root-feeding WCR remain largely unknown [17].
Microbial biological control agents, including plant growth-promoting rhizobacteria and fungi (PGPR and PGPF, respectively), have been widely used to increase resistance in plants against pathogens and pests [18-20]. Isolates of soil-borne Trichoderma spp. and bacterium Pseudomonas chlororaphis are well-known biocontrol agents and have been extensively studied for their beneficial traits, including growth promotion and enhanced disease resistance of the host plants [21-24]. However, using biocontrol agents against pests has only recently been considered, and little is known about their impact on defense against chewing insects [25, 26]. Several studies have shown that T. virens induces systemic resistance (ISR) against leaf-infecting pathogens, the physiological process tightly regulated by fungal small secreted proteins, Sm1 and Sir1, which play pivotal but contrasting roles in triggering ISR responses [27-30]. Analyses of the T. virens sm1 deletion mutants revealed that they are unable to induce ISR against fungal pathogens in above-ground organs, suggesting that Sm1 peptide is a positive regulator of ISR [27, 28]. In sharp contrast, deletion of SIR1 resulted in much stronger ISR response [29]. Wang et al. (2020)[30] showed that T. virens colonization in maize roots upregulated genes involved in biosynthesis of JA precursors, including 12-OPDA (12-oxo-10(Z),15(Z)-phytodienoic acid) and OPC-4:0 {(Z)-4-[3-oxo-2-(pent-2-en-1-yl)cyclopentyl]butanoic acid}, yet downregulated genes downstream of 12-OPDA for JA biosynthesis. Whether T. virens colonization affects wound-induced JA production in leaves and confers resistance to leaf-feeding FAW and root-feeding WCR larvae remains unexplored.
The principal objective of this study was to investigate the effects of T. virens and its secreted peptide signals on the production of wound-induced JA and other defensive oxylipin metabolites and to test whether the beneficial effects of this endophyte extend to increasing resistance against FAW and WCR. Contrary to our expectation, the results demonstrated that maize seedlings grown in sterile soil amended with wild-type T. virens, the sm1 or sir1 single or the double mutants were found to suppress wound-induced JA biosynthesis, resulting in reduced resistance to FAW. We followed up by testing whether such a detrimental effect on defense against herbivory could be ascribed to another well-studied growth-promoting bacterial endophyte, P. chlororaphis. Colonization of roots by the P. chlororaphis 30-84 also reduced production of JA and defense against FAW. Further analysis showed that while both T. virens and P. chlororaphis induced ISR against C. graminicola when added in sterile or nonsterile soil, suppression of defense against FAW occurred only when maize seedlings were grown in sterile soil amended with these biocontrol agents. In addition, we demonstrated that T. virens reduced survival and growth (weight gain) of WCR larvae, making our study the first showing the potential of T. virens as a biocontrol agent against this devastating pest of maize.
2. Results
2.1. Colonization of Maize Roots by either Δsir1 or Δsm1Δsir1 Enhanced Greater Levels of ISR against C. Graminicola
Previously, we have shown that T. virens colonization triggered ISR against C. graminicola in a Sm1 peptide-dependent manner as the Δsm1 mutant lost its ISR-inducing capability [28, 30]. In contrast, plants treated with the deletion mutant of Sir1 peptide, Δsir1, exhibited enhanced systemic resistance [29, 30]. Moreover, Wang et al. (2020)[30] showed that the expression of SIR1 in Δsm1 remained similar to the level in wild-type T. virens strain (TvWT), while Δsir1 had elevated expression of SM1. These results suggested that one potential reason for increased ISR by Δsir1 is due to elevated SM1 expression in this strain. To test this possibility, we created Δsm1Δsir1 double mutant and tested its capability to induce resistance to C. graminicola. The result showed that colonization of both Δsir1 and Δsm1Δsir1 mutants triggered greater level of ISR against C. graminicola compared to TvWT (32%, 59%, and 56% reduction in lesion areas on TvWT-, Δsir1-, and Δsm1Δsir1-treated plants, respectively, compared to control) (Figure 1). These results suggest that a functional Sir1 suppresses disease resistance to C. graminicola by suppressing not only SM1, but other as yet unknown mechanisms.
2.2. T. virens Colonization Suppressed Insect Defense
Numerous studies have shown that beneficial Trichoderma spp. induce resistance against pathogens [31-33]. However, their effect on inducing insect resistance remains largely unexplored. Here, we tested whether different T. virens stains, including Δsm1, Δsir1, and Δsm1Δsir1, induce differential resistance to the foliar-feeding FAW. Surprisingly, the results showed that FAW larvae gained significantly more weight after feeding on T. virens-colonized plants for 7 days (60% to 95% increase) (Figure 2A) and 3rd-instar larvae consumed more leaf area on T. virens-colonized plants (64% to 83% increase) (Figure 2B). Treatment with T. virens reduced resistance to FAW, regardless of the mutation of these two signaling peptides.
2.3. T. virens Treatment Reduced Wound-Induced JA Accumulation by Suppressing Biosynthesis and Enhancing Catabolism
A previous study showed that treatment with T. virens downregulated JA biosynthesis genes in colonized maize roots [30] and that JA dominantly modulates defense responses against chewing insects [14]. In the present study, we found that T. virens treatment decreased insect defense, which prompted us to test whether T. virens colonization regulates wound-induced JA production in leaves. Phytohormone and oxylipin profiling showed that significantly lower amounts of jasmonates, including JA precursors, 12-OPDA, OPC-4:0, JA, and the bioactive JA derivatives, JA-Isoleucine (JA-Ile), JA-leucine (JA-Leu), and JA-Valine (JA-Val) conjugates, were accumulated in the T. virens-treated plants at 1 and 2 hours post wounding (hpw) (Figure 3A). Specifically, JA accumulation was reduced by 23% to 37% and 33% to 38% and JA-Ile content was reduced by 8% to 37% and 44% to 50% compared to the amounts accumulated in the control plants at 1 and 2 hpw, respectively (Figure 3B,C). Notably, we also found that one of the JA catabolites, 12OH-JA, accumulated to a significantly higher level in the T. virens-treated plants in response to mechanical wounding (83% to 171% and 23% to 82% increase compared to the contents in the control plants at 1 and 2 hpw, respectively) (Figure 3A and 3D). This latter result suggests that one potential reason behind decreased JA content may be due to increased catabolism of JA. To confirm the reduced insect defense by T. virens colonization was modulated by JA signaling, we tested the effect of T. virens on the JA-deficient opr7opr8 mutant [14] to FAW. The result showed that T. virens colonization did not increase susceptibility to FAW on opr7opr8 double mutants, suggesting that attenuated JA signaling plays an essential role in T. virens-reduced insect defense (Figure 4). Interestingly, the levels of the α-ketol, 9,10-KODA (9-hydroxy-10-oxo-12(Z),15(Z)-octadecadienoic acid), have been significantly reduced at 1 hpw (24% to 37% reduction compared to the contents in the control plants) by all strains of T. virens (Figure 3E). This molecule has been recently shown to play a major JA-dependent signaling role in defense against FAW [34]. Together, these results indicate that T. virens colonization suppressed defense against FAW via downregulating JA biosynthesis and/or enhancing JA catabolism.
2.4. T. virens and P. chlororaphis Colonization Enhanced Growth but Suppressed Insect Defense against FAW in Sterile Soil While Increasing Resistance to C. graminicola in both Sterile and Nonsterile Soil
Bacterial biocontrol agent P. chlororaphis 30-84 was previously reported to promote growth and suppress growth of fungal pathogens [35, 36], yet its role in insect defense against foliar-feeding insects remains mostly unexplored. Therefore, we also tested the impact of P. chlororaphis 30-84 on plant growth promotion, defense against FAW, and pathogen resistance and the effect of combining the two beneficial microorganisms, T. virens and P. chlororaphis 30-84, on these physiological processes. Because the beneficial microorganisms were commonly tested in an agricultural setting, we further tested their effect in both sterile and nonsterile soil mixtures of field soil and potting mix. Similar to T. virens treatment, colonization by P. chlororaphis 30-84 promoted plant growth (12% to 15% increase in plant height) but suppressed defense against FAW (41% to 51% increase in weight gain) in the sterile soil mixture (Figure 5A,B). Plant growth and insect defense were not significantly affected when seedlings were grown in a nonsterile soil mixture upon treatment with T. virens and P. chlororaphis 30-84. However, both T. virens and P. chlororaphis 30-84 were able to increase resistance to C. graminicola in both soil types (53% to 66% reduction and 44% to 55% reduction in lesion areas in nonsterile and sterile soils, respectively) (Figure 5C).
2.5. T. virens and P. chlororaphis 30-84 Colonization Reduced Wound-Induced JA Accumulation by Suppressing Biosynthesis and Enhancing Catabolism in Plants Grown in Sterile Soil
Decreased FAW resistance found with the amendment of P. chlororaphis 30-84 in this study prompted us to investigate whether this biocontrol agent impacts JA homeostasis in response to wounding in sterile soil. Phytohormone and oxylipin profiling revealed that P. chlororaphis 30-84 treatment reduced wound-induced accumulation of jasmonates, including 12-OPDA, JA, JA-Ile, and JA-Leu in sterile soil (Figure 6A). More specifically, treatments with T. virens, P. chlororaphis 30-84, and the mixture of T. virens and P. chlororaphis 30-84 resulted in 25%, 21%, and 19% reduction in JA accumulation and 27%, 32%, and 27% reduction in JA-Ile level at 1 hpw, respectively (Figure 6B,C). Moreover, JA catabolism was increased by 50%, 13%, and 21% in the plants treated with T. virens, P. chlororaphis 30-84, and the mixture of T. virens and P. chlororaphis 30-84, respectively (Figure 6D). Interestingly, suppression of wound-induced JA biosynthesis (Figure 6) and FAW insect defense (Figure 5) by either T. virens or P. chlororaphis 30-84 was only observed in the sterile soil condition. A similar accumulation pattern was observed for 9,10-KODA (17% to 34% reduction) in sterile soil (Figure 6E). Together, these results suggest that T. virens and P. chlororaphis 30-84 suppress foliar-feeding insect defense via regulating JA homeostasis in response to wounding only in plants grown in sterile soil amended with either of these biocontrol agents.
2.6. T. virens Colonization Suppressed WCR Larval Survival and Weight Gain
We also tested whether T. virens plays a role in conferring resistance to root-feeding WCR larvae. The results showed that significantly fewer larvae were recovered (50% fewer) (Figure 7A), and WCR larvae gained significantly less weight after feeding on T. virens-colonized plants (57% less) (Figure 7B). However, T. virens treatment did not reduce tissue consumption (Figure 7C,D). These results suggest that T. virens may be a potent biocontrol agent against WCR by suppressing WCR larval survival and weight gain.
3. Discussion
Soil-borne Trichoderma spp. are ubiquitous in soil worldwide, and have been used as biocontrol agents against pathogens for decades [23, 31, 37-39]. However, the potential utility of Trichoderma spp. as biocontrol agents against insect pests has just begun to be addressed [26], and information on their effects against foliar-feeding and root-feeding insects is scarce. Accumulating evidence suggests that root colonization by Trichoderma spp. triggered ISR against insects, such as aphids [40-43], whiteflies [44], thrips [45], and Lepidoptera [41, 46-50], and also enhanced defense against spider mites [51] and nematodes [52-54]. Previous reports [27, 28, 30] and this study showed that T. virens confers resistance to C. graminicola. In addition, these studies have identified a fungal-secreted peptide, Sm1, as a signal necessary for inducing systemic resistance by showing that the Δsm1 deletion mutant of T. virens is unable to induce ISR [27, 28, 30]. In contrast to Δsm1, another signaling peptide mutant of T. virens, Δsir1 [29, 30], triggered enhanced ISR against C. graminicola. As this mutant overexpressed SM1 transcripts [30], we hypothesized that the increased ISR following colonization by this mutant is due to elevated SM1 expression. We tested this hypothesis by deleting SM1 in the Δsir1 mutant background and found that the Δsm1Δsir1 double mutant still displayed stronger resistance as compared to the wild-type strain. We are suggesting that Sir1 peptide acts to suppress ISR not only by downregulating SM1 expression, but also by employing other as-yet-unknown mechanisms of ISR suppression.
Several reports showed that several Trichoderma spp. directly impact Lepidoptera spp. by producing antifeedant compounds and toxic secondary metabolites [26, 55-59] or indirectly impact them via activation of JA-mediated systemic defense, production of parasitoid-attracting volatile compounds, or disruption of insect gut microbiota and metabolome [41, 42, 46-48, 50, 60]. Here, we demonstrated that the treatment with T. virens suppressed defense against FAW by reducing wound-induced JA production and increased JA catabolism in plants grown in sterile soil. Consistent with our findings, Lelio et al. (2021)[47] showed that when grown at cool temperatures (20°C as compared to 25°C), tomato plants colonized with T. afroharzianum T22 had less negative impact on Spodoptera littoralis larvae and this was accompanied with suppressed JA biosynthesis and signaling pathways. Also consistent with our results, Kinyungu et al. (2023)[49] showed that the treatment with T. atroviride F2S21 strain resulted in a higher leaf defoliation rate by FAW larvae. The fact that some studies showed increased resistance to certain chewing insects due to Trichoderma spp. treatment [41, 46, 50] while others, including the current study, show opposite effect of Trichoderma treatment, suggest Trichoderma species- and strain-specific effects as well an environment-dependent effect on insect defense. Interestingly, our results indicate that the reduced insect defense is independent of the fungal secreted peptides, Sm1 and Sir1, suggesting that fungal signals regulating ISR against pathogens and increased induced susceptibility against chewing insects involve different molecular signals to be identified in the future.
Previous reports showed that root colonization by T. virens induced biosynthesis of the JA precursor, 12-OPDA, and ketols [30, 61]. Both 12-OPDA and ketols were shown to act as xylem-mobile long-distance signals that prime the above-ground organs for greater resistance against pathogens in maize [30, 61]. In contrast to these oxylipins, JA-Ile levels in xylem sap were not significantly increased in response to T. virens treatment and transfusion of maize seedlings with xylem sap supplemented with JA-Ile increased susceptibility to C. graminicola [30]. The role of JA as a susceptibility factor against anthracnose leaf blight caused by C. graminicola was further supported by the studies of Gorman et al. (2020)[12] and Huang et al. (2023)[13]. Moreover, transcriptome analysis showed that maize roots responded to colonization by T. virens by increased expression of 12-OPDA biosynthesis genes but suppressed expression of the genes required for 12-OPDA conversion to JA-Ile [30]. Because JA is widely reported to suppress growth in numerous plant species [62-64], it is tempting to speculate that attenuated synthesis of JAs in roots colonized by T. virens is a potential mechanism accounting for the strong growth promotion effect by this symbiont observed in this study and previous reports. Similar to the T. virens-mediated induction of ISR against pathogens and growth promotion, we showed that root colonization by the bacterial symbiont, P. chlororaphis 30-84, resulted in increased seedling growth and systemic resistance against C. graminicola, suggesting a similar mechanism underlying these two beneficial responses. However, as in the case of T. virens, P. chlororaphis 30-84 treatment reduced JA production and decreased resistance to herbivory by FAW when treated plants were grown in sterile soil. Therefore, it is likely that growth promotion by both beneficial microorganisms, T. virens and P. chlororaphis 30-84, and ISR against the hemibiotrophic pathogen C. graminicola is linked to their ability to suppress JA synthesis and JA-mediated signaling. Interestingly, wound-induced JA accumulation and insect defense were not affected when maize seedlings were treated with T. virens or P. chlororaphis 30-84 in nonsterile soil, suggesting that natural microbial communities dampened JA suppression activities by both endophytes.
WCR root feeding causes substantial economic damage to corn production in the United States and Europe [65, 66] and the insect has adapted to overcome most of the available management techniques, including chemical insecticides and Bacillus thuringiensis (Bt) toxins [66, 67]. So far, there is a lack of efficient management of this notorious pest and, thus, there is an urgent need for a sustainable and economical management approach, such as utilizing efficient biocontrol agents. In this study, we showed that T. virens colonization significantly enhanced mortality and suppressed weight gain of WCR larvae. Our previous report showed that increased WCR mortality was correlated with greater levels of herbivory-induced production of insecticidal death acids and ketols [16]. Therefore, one of the potential mechanisms of T. virens-mediated WCR defense may be to activate the production of insecticidal defensive compounds. The other potential mechanism behind the detrimental effect on larvae survival and growth conferred by T. virens may be the production of some fungus-derived insecticidal metabolites as reported in several studies [26, 55-59]. Further experiments are needed to explore whether T. virens has a direct insecticidal impact on the WCR root-feeding larvae or has an indirect effect due to the induction of the host -mediated synthesis of toxic compounds.
4. Materials and Methods
4.1. Plant, Growth Medium, and Plant-Beneficial Inoculants
The maize (Zea mays) inbred line B73 and opr7opr8 double mutant (Yan et al., 2012)[14] were used in this study. Plants were grown in conical pots (20.5 × 4 cm) filled with autoclaved (121°C for 60 minutes, 2 cycles) commercial potting mix (Jolly Gardener Pro Line C/20) on light shelves at room temperature (22–24°C) with a 16-h light period. To test the efficacy of the beneficial microorganisms in an agricultural setting, maize B73 seeds were grown in a mixture of field soil (collected from Texas A&M University Research Farm in the fall of 2022 after maize was harvested) and potting mix (Jolly Gardener Pro Line C/20) at the ratio of 1 to 3 that was either autoclaved (121°C for 60 minutes, 2 cycles; henceforth referred to as sterile) or untreated (hence referred to as nonsterile) after mixing. Chlamydospores of T. virens strains TvWT (Gv29-8; Baek and Kenerly, 1998)[68], Δsm1 (Djonovic´ et al., 2006; Djonovic et al., 2007)[27, 28], Δsir1 (formerly D77560; Lamdan et al., 2015)[29], and Δsm1Δsir1 (this study) were obtained from liquid cultures grown in molasses-yeast extract medium as described in Wang et al. (2020)[30]. P. chlororaphis 30-84 was grown in Luria-Bertani (LB) medium containing 5 g of NaCl per liter at pH 7 at 28°C with agitation at 200 rpm, as described in Yuan et al. (2020)[36]. Seven days after sowing, seedlings were treated with 0.05 g of the T. virens chlamydospores, or 300 μl of P. chlororaphis 30-84 suspension at OD620 = 0.8, or a mixture of both T. virens and P. chlororaphis with the same amount described above (added to the soil at a depth of approximately 2 cm around the seeds), or left untreated.
4.2. Anthracnose Leaf Blight Assay
C. graminicola stain 1.001 was grown on half strength potato dextrose agar (PDA) plates for 2-3 weeks to allow sporulation. Two weeks after being colonized by beneficial microorganisms, the plants were inoculated with 10 μl of conidial suspension (106 conidia/ml) with six droplets of spore suspensions per leaf collected from hemibiotrophic fungal pathogen C. graminicola plates as described in Huang et al. (2023a)[13]. The leaves were scanned and the lesion areas measured using ImageJ software [69] seven days post-inoculation.
4.3. Fall Armyworm Assay
The laboratory strain of fall armyworm (FAW; Spodoptera frugiperda) was purchased from Benzon Research (Carlisle, PA, USA). Two weeks after incubation with the beneficial microorganisms, six FAW neonates were placed to the whorls of maize seedlings contained in individual plastic jars and allowed to move and feed on the plants freely for 7 days, as described in Huang et al. (2023b)[16]. FAW larvae were removed from the plants, and the total weight was determined 7 days post-infestation. To evaluate insect resistance and leaf damaged area, one maize leaf was individually caged in a handmade clip-cage and infested with one 3rd-instar FAW larva per spot for approximately one hour, then moved toward the base of the plant. Leaves were then scanned, and eaten areas were measured with ImageJ software[69].
4.4. Oxylipin Profiling of Wounded Leaf Tissue
For the wounding treatment, the third fully expanded leaf of seedlings grown in sterile or nonsterile soil conditions that were previously colonized with beneficial microorganisms for two weeks or left untreated was wounded seven times using a hemostat, with three wound sites on one side and four on the other side of the midvein with the wound sites approximately 1 cm apart. The wounded regions were then harvested and placed in 2 mL screw-cap Fast-Prep tubes (Qbiogene, Carlsbad, CA, U. S. A.) in liquid nitrogen and stored in a -80 °C freezer. Phytohormone and oxylipin extraction and profiling of wounded leaf tissue were performed as described previously [13].
4.5. Western Corn Rootworm Bioassays
Western corn rootworm (WCR; Diabrotica virgifera virgifera) eggs were provided by USDA-ARS North Central Agriculture Research Laboratory (Brookings, SD, USA). Two weeks after being colonized by beneficial microorganisms, ten WCR neonates were placed on the soil surface in individual cone-tainers with a maize seedling and allowed to burrow and feed for 10 days. WCR larvae were removed from the soil, and the total weight was determined 10 days post-infestation. The fresh weights of shoot and root tissues was measured after the soil was removed and the ratio was analyzed as larvae damaged over control root mass, as described in Huang et al., (2023)[16].
4.6. Statistical Methods
Statistical analyses were performed using the software programs JMP Pro 17 (SAS Institute Inc., Cary, NC, USA). Lesion areas, FAW larval weight, leaf eaten areas, wound-induced accumulation of metabolites, and plant heights were analyzed using One-way Analysis of Variance (ANOVA) with post-hoc Tukey HSD (Honestly Significant Difference). FAW larval weight after feeding on opr7opr8 double mutants and wild-type seedlings treated with T. virens or left untreated and root and shoot ratios after WCR infestation were analyzed using Two-way ANOVA with Tukey HSD. Effect of sterile and nonsterile mixture was analyzed using Student’s t-test.
5. Conclusions
The elevated level of ISR against C. graminicola triggered by both T. virens Δsir1 and Δsm1Δsir1 double mutants revealed that a functional Sir1 signal peptide reduces disease resistance by downregulating not only SM1 (as demonstrated previously by Wang et al. (2020)) but also involves other unknown mechanisms as well. Colonization of plants grown in sterile soil by different T. virens mutants suppressed wound-induced JA accumulation, resulting in reduced defense against FAW, which is independent of both signal peptides, Sm1 and Sir1. When testing both beneficial microorganisms, T. virens and P. chlororaphis 30-84, using both sterile and nonsterile mixtures of field soil and potting mix, both biocontrol agents triggered ISR against C. graminicola in both soil types. However, these agents reduced defense against FAW only when the treated plants were grown in sterile soil due to reduced JA. In contrast to the effect on leaf-feeding FAW, T. virens colonization suppressed root-feeding WCR larvae survival and weight gain, the first report revealing the potential of T. virens as a biocontrol agent against WCR.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: title; Table S1: title; Video S1: title.
Author Contributions
Designed the research and analyzed the data, P.-C.H. and M.V.K.; performed most of the experiments and drafted the article, P.-C.H.; treatment with P. chlororaphis 30-84, P.Y.; conducted the WCR bioassays, J.M.G and P.-C.H..; created Δsm1Δsir1 double mutant, J.T.; helped design the experiments and wrote the article with input from and revisions by J.M.G., J.T., E.-O.T., E.A.P., J.S.B., and C.M.K. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported in part by United States Department of Agriculture (USDA)-National Institute of Food and Agriculture (NIFA) 2017-67013- 26524 to M.V.K and 2021-67013-33568 grants awarded to M.V.K and J.S.B.
Data Availability Statement
The datasets used and analyzed during the current study are available from the corresponding author upon reasonable request.
Acknowledgments
We thank Andie Miller and Dr. Greg Sword, Department of Entomology, Texas A&M University, for kindly sharing FAW eggs/neonates.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Erenstein, O.; Jaleta, M.; Sonder, K.; Mottaleb, K.; Prasanna, B. M. , Global maize production, consumption and trade: trends and R&D implications. Food Security 2022, 14(5), 1295–1319. [Google Scholar]
- Savary, S.; Willocquet, L.; Pethybridge, S. J.; Esker, P.; McRoberts, N.; Nelson, A., The global burden of pathogens and pests on major food crops. Nature ecology & evolution 2019, 3 (3), 430-439.
- Belisário, R.; Robertson, A. E.; Vaillancourt, L. J. , Maize Anthracnose Stalk Rot in the Genomic Era. Plant Disease 2022, 106(9), 2281–2298. [Google Scholar] [CrossRef] [PubMed]
- Bergstrom, G. C.; Nicholson, R. L. , The Biology of Corn Anthracnose: Knowledge to Exploit for Improved Management. Plant Disease 1999, 83(7), 596–608. [Google Scholar] [CrossRef] [PubMed]
- Dean, R.; Van Kan, J. A. L.; Pretorius, Z. A.; Hammond-Kosack, K. E.; Di Pietro, A.; Spanu, P. D.; Rudd, J. J.; Dickman, M.; Kahmann, R.; Ellis, J.; Foster, G. D. , The Top 10 fungal pathogens in molecular plant pathology. Molecular plant pathology 2012, 13(4), 414–430. [Google Scholar] [CrossRef]
- Kenis, M. B. , Giovanni; Biondi, Antonio; Calatayud, Paul-André; Day, Roger; Desneux, Nicolas; Harrison, Rhett D; Kriticos, Darren; Rwomushana, Ivan; van den Berg, Johnnie; Verheggen, François; Zhang, Yong-Jun; Agboyi, Lakpo Koku; Ahissou, Régis Besmer; Ba, Malick N; Wu, Kongming, Invasiveness, biology, ecology, and management of the fall armyworm, Spodoptera frugiperda. Entomologia Generalis 2023, 43(2), 55. [Google Scholar]
- Overton, K.; Maino, J. L.; Day, R.; Umina, P. A.; Bett, B.; Carnovale, D.; Ekesi, S.; Meagher, R.; Reynolds, O. L. , Global crop impacts, yield losses and action thresholds for fall armyworm (Spodoptera frugiperda): A review. Crop Protection 2021, 145, 105641. [Google Scholar] [CrossRef]
- Gray, M. E.; Sappington, T. W.; Miller, N. J.; Moeser, J.; Bohn, M. O. , Adaptation and invasiveness of western corn rootworm: intensifying research on a worsening pest. Annu Rev Entomol 2009, 54, 303–21. [Google Scholar] [CrossRef]
- Darlington, M.; Reinders, J. D.; Sethi, A.; Lu, A. L.; Ramaseshadri, P.; Fischer, J. R.; Boeckman, C. J.; Petrick, J. S.; Roper, J. M.; Narva, K. E.; Vélez, A. M. , RNAi for Western Corn Rootworm Management: Lessons Learned, Challenges, and Future Directions. Insects 2022, 13 (1). [Google Scholar] [CrossRef]
- Wechsler, S.; Smith, D. , Has resistance taken root in US corn fields? Demand for insect control. American Journal of Agricultural Economics 2018, 100(4), 1136–1150. [Google Scholar] [CrossRef]
- Gyeraj, A.; Szalai, M.; Pálinkás, Z.; Edwards, C. R.; Kiss, J. , Effects of adult western corn rootworm (Diabrotica virgifera virgifera LeConte, Coleoptera: Chrysomelidae) silk feeding on yield parameters of sweet maize. Crop Protection 2021, 140, 105447. [Google Scholar] [CrossRef]
- Gorman, Z.; Christensen, S. A.; Yan, Y.; He, Y.; Borrego, E.; Kolomiets, M. V. , Green leaf volatiles and jasmonic acid enhance susceptibility to anthracnose diseases caused by Colletotrichum graminicola in maize. Molecular plant pathology 2020, 21(5), 702–715. [Google Scholar] [CrossRef]
- Huang, P. C.; Tate, M.; Berg-Falloure, K. M.; Christensen, S. A.; Zhang, J.; Schirawski, J.; Meeley, R.; Kolomiets, M. V. , A non-JA producing oxophytodienoate reductase functions in salicylic acid-mediated antagonism with jasmonic acid during pathogen attack. Molecular plant pathology 2023, 24(7), 725–741. [Google Scholar] [CrossRef]
- Yan, Y.; Christensen, S.; Isakeit, T.; Engelberth, J.; Meeley, R.; Hayward, A.; Emery, R. J.; Kolomiets, M. V. , Disruption of OPR7 and OPR8 reveals the versatile functions of jasmonic acid in maize development and defense. Plant Cell 2012, 24(4), 1420–36. [Google Scholar] [CrossRef]
- Christensen, S. A.; Nemchenko, A.; Borrego, E.; Murray, I.; Sobhy, I. S.; Bosak, L.; DeBlasio, S.; Erb, M.; Robert, C. A.; Vaughn, K. A.; Herrfurth, C.; Tumlinson, J.; Feussner, I.; Jackson, D.; Turlings, T. C.; Engelberth, J.; Nansen, C.; Meeley, R.; Kolomiets, M. V. , The maize lipoxygenase, ZmLOX10, mediates green leaf volatile, jasmonate and herbivore-induced plant volatile production for defense against insect attack. The Plant journal : for cell and molecular biology 2013, 74 (1), 59–73. [Google Scholar] [CrossRef]
- Huang, P. C.; Grunseich, J. M.; Berg-Falloure, K. M.; Tolley, J. P.; Koiwa, H.; Bernal, J. S.; Kolomiets, M. V. , Maize OPR2 and LOX10 Mediate Defense against Fall Armyworm and Western Corn Rootworm by Tissue-Specific Regulation of Jasmonic Acid and Ketol Metabolism. Genes 2023, 14(9). [Google Scholar] [CrossRef]
- Castano-Duque, L.; Loades, K. W.; Tooker, J. F.; Brown, K. M.; Paul Williams, W.; Luthe, D. S., A Maize Inbred Exhibits Resistance Against Western Corn Rootwoorm, Diabrotica virgifera virgifera. Journal of chemical ecology 2017, 43 (11-12), 1109-1123.
- Köhl, J.; Kolnaar, R.; Ravensberg, W. J. , Mode of Action of Microbial Biological Control Agents Against Plant Diseases: Relevance Beyond Efficacy. Frontiers in plant science 2019, 10. [Google Scholar] [CrossRef]
- Tiénébo, E. O.; Harrison, K.; Abo, K.; Brou, Y. C.; Pierson, L. S., 3rd; Tamborindeguy, C.; Pierson, E. A.; Levy, J. G., Mycorrhization Mitigates Disease Caused by "Candidatus Liberibacter solanacearum" in Tomato. Plants (Basel, Switzerland) 2019, 8 (11).
- El-Saadony, M. T.; Saad, A. M.; Soliman, S. M.; Salem, H. M.; Ahmed, A. I.; Mahmood, M.; El-Tahan, A. M.; Ebrahim, A. A. M.; Abd El-Mageed, T. A.; Negm, S. H.; Selim, S.; Babalghith, A. O.; Elrys, A. S.; El-Tarabily, K. A.; AbuQamar, S. F. , Plant growth-promoting microorganisms as biocontrol agents of plant diseases: Mechanisms, challenges and future perspectives. Frontiers in plant science 2022, 13, 923880. [Google Scholar] [CrossRef] [PubMed]
- Zin, N. A.; Badaluddin, N. A. , Biological functions of Trichoderma spp. for agriculture applications. Annals of Agricultural Sciences 2020, 65(2), 168–178. [Google Scholar] [CrossRef]
- Tyśkiewicz, R.; Nowak, A.; Ozimek, E.; Jaroszuk-Ściseł, J., Trichoderma: The Current Status of Its Application in Agriculture for the Biocontrol of Fungal Phytopathogens and Stimulation of Plant Growth. International journal of molecular sciences 2022, 23 (4).
- Yao, X.; Guo, H.; Zhang, K.; Zhao, M.; Ruan, J.; Chen, J. , Trichoderma and its role in biological control of plant fungal and nematode disease. Frontiers in microbiology 2023, 14, 1160551. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A. J.; Kim, Y. C. , Insights into plant-beneficial traits of probiotic Pseudomonas chlororaphis isolates. Journal of medical microbiology 2020, 69(3), 361–371. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Cornejo, H. A.; Macías-Rodríguez, L.; del-Val, E.; Larsen, J. , The root endophytic fungus Trichoderma atroviride induces foliar herbivory resistance in maize plants. Applied Soil Ecology 2018, 124, 45–53. [Google Scholar] [CrossRef]
- Poveda, J. , Trichoderma as biocontrol agent against pests: New uses for a mycoparasite. Biological Control 2021, 159, 104634. [Google Scholar] [CrossRef]
- Djonović, S.; Pozo, M. J.; Dangott, L. J.; Howell, C. R.; Kenerley, C. M., Sm1, a proteinaceous elicitor secreted by the biocontrol fungus Trichoderma virens induces plant defense responses and systemic resistance. Molecular plant-microbe interactions : MPMI 2006, 19 (8), 838-53.
- Djonovic, S.; Vargas, W. A.; Kolomiets, M. V.; Horndeski, M.; Wiest, A.; Kenerley, C. M. , A proteinaceous elicitor Sm1 from the beneficial fungus Trichoderma virens is required for induced systemic resistance in maize. Plant physiology 2007, 145(3), 875–89. [Google Scholar] [CrossRef]
- Lamdan, N. L.; Shalaby, S.; Ziv, T.; Kenerley, C. M.; Horwitz, B. A., Secretome of Trichoderma interacting with maize roots: role in induced systemic resistance. Molecular & cellular proteomics : MCP 2015, 14 (4), 1054-63.
- Wang, K. D.; Borrego, E. J.; Kenerley, C. M.; Kolomiets, M. V. , Oxylipins Other Than Jasmonic Acid Are Xylem-Resident Signals Regulating Systemic Resistance Induced by Trichoderma virens in Maize. Plant Cell 2020, 32(1), 166–185. [Google Scholar] [CrossRef]
- Khan, R. A. A.; Najeeb, S.; Chen, J.; Wang, R.; Zhang, J.; Hou, J.; Liu, T. , Insights into the molecular mechanism of Trichoderma stimulating plant growth and immunity against phytopathogens. Physiologia plantarum 2023, 175(6), e14133. [Google Scholar] [CrossRef] [PubMed]
- Dehbi, I.; Achemrk, O.; Ezzouggari, R.; El Jarroudi, M.; Mokrini, F.; Legrifi, I.; Belabess, Z.; Laasli, S. E.; Mazouz, H.; Lahlali, R., Beneficial Microorganisms as Bioprotectants against Foliar Diseases of Cereals: A Review. Plants (Basel, Switzerland) 2023, 12 (24).
- Saldaña-Mendoza, S. A.; Pacios-Michelena, S.; Palacios-Ponce, A. S.; Chávez-González, M. L.; Aguilar, C. N., Trichoderma as a biological control agent: mechanisms of action, benefits for crops and development of formulations. World journal of microbiology & biotechnology 2023, 39 (10), 269.
- Yuan, P.; Borrego, E.; Park, Y. S.; Gorman, Z.; Huang, P. C.; Tolley, J.; Christensen, S. A.; Blanford, J.; Kilaru, A.; Meeley, R.; Koiwa, H.; Vidal, S.; Huffaker, A.; Schmelz, E.; Kolomiets, M. V. , 9,10-KODA, an α-ketol produced by the tonoplast-localized 9-lipoxygenase ZmLOX5, plays a signaling role in maize defense against insect herbivory. Molecular plant 2023. [Google Scholar] [CrossRef]
- Yu, J. M.; Wang, D.; Pierson, L. S., 3rd; Pierson, E. A. , Effect of Producing Different Phenazines on Bacterial Fitness and Biological Control in Pseudomonas chlororaphis 30-84. The plant pathology journal 2018, 34(1), 44–58. [Google Scholar] [CrossRef]
- Yuan, P.; Pan, H.; Boak, E. N.; Pierson, L. S., 3rd; Pierson, E. A. , Phenazine-Producing Rhizobacteria Promote Plant Growth and Reduce Redox and Osmotic Stress in Wheat Seedlings Under Saline Conditions. Frontiers in plant science 2020, 11, 575314. [Google Scholar] [CrossRef]
- Harman, G. E.; Howell, C. R.; Viterbo, A.; Chet, I.; Lorito, M. , Trichoderma species — opportunistic, avirulent plant symbionts. Nature Reviews Microbiology 2004, 2(1), 43–56. [Google Scholar] [CrossRef] [PubMed]
- Lorito, M.; Woo, S. L.; Harman, G. E.; Monte, E. , Translational research on Trichoderma: from 'omics to the field. Annual review of phytopathology 2010, 48, 395–417. [Google Scholar] [CrossRef] [PubMed]
- Woo, S. L.; Hermosa, R.; Lorito, M.; Monte, E. , Trichoderma: a multipurpose, plant-beneficial microorganism for eco-sustainable agriculture. Nature Reviews Microbiology 2023, 21(5), 312–326. [Google Scholar] [CrossRef]
- Coppola, M.; Cascone, P.; Chiusano, M. L.; Colantuono, C.; Lorito, M.; Pennacchio, F.; Rao, R.; Woo, S. L.; Guerrieri, E.; Digilio, M. C. , Trichoderma harzianum enhances tomato indirect defense against aphids. Insect science 2017, 24(6), 1025–1033. [Google Scholar] [CrossRef] [PubMed]
- Coppola, M.; Cascone, P.; Lelio, I. D.; Woo, S. L.; Lorito, M.; Rao, R.; Pennacchio, F.; Guerrieri, E.; Digilio, M. C. , Trichoderma atroviride P1 Colonization of Tomato Plants Enhances Both Direct and Indirect Defense Barriers Against Insects. Frontiers in physiology 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Coppola, M.; Diretto, G.; Digilio, M. C.; Woo, S. L.; Giuliano, G.; Molisso, D.; Pennacchio, F.; Lorito, M.; Rao, R. , Transcriptome and Metabolome Reprogramming in Tomato Plants by Trichoderma harzianum strain T22 Primes and Enhances Defense Responses Against Aphids. Frontiers in physiology 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Islam, M. S.; Subbiah, V. K.; Siddiquee, S. , Efficacy of Entomopathogenic Trichoderma Isolates against Sugarcane Woolly Aphid, Ceratovacuna lanigera Zehntner (Hemiptera: Aphididae). Horticulturae 2022, 8(1), 2. [Google Scholar] [CrossRef]
- Jafarbeigi, F.; Samih, M. A.; Alaei, H.; Shirani, H. , Induced Tomato Resistance Against Bemisia tabaci Triggered by Salicylic Acid, β-Aminobutyric Acid, and Trichoderma. Neotropical entomology 2020, 49(3), 456–467. [Google Scholar] [CrossRef]
- Muvea, A. M.; Meyhöfer, R.; Subramanian, S.; Poehling, H. M.; Ekesi, S.; Maniania, N. K. , Colonization of onions by endophytic fungi and their impacts on the biology of Thrips tabaci. PLoS One 2014, 9(9), e108242. [Google Scholar] [CrossRef]
- Contreras-Cornejo, H. A.; del-Val, E.; Macías-Rodríguez, L.; Alarcón, A.; González-Esquivel, C. E.; Larsen, J. , Trichoderma atroviride, a maize root associated fungus, increases the parasitism rate of the fall armyworm Spodoptera frugiperda by its natural enemy Campoletis sonorensis. Soil Biology and Biochemistry 2018, 122, 196–202. [Google Scholar] [CrossRef]
- Di Lelio, I.; Coppola, M.; Comite, E.; Molisso, D.; Lorito, M.; Woo, S. L.; Pennacchio, F.; Rao, R.; Digilio, M. C. , Temperature Differentially Influences the Capacity of Trichoderma Species to Induce Plant Defense Responses in Tomato Against Insect Pests. Frontiers in plant science 2021, 12, 678830. [Google Scholar] [CrossRef] [PubMed]
- Papantoniou, D.; Vergara, F.; Weinhold, A.; Quijano, T.; Khakimov, B.; Pattison, D. I.; Bak, S.; van Dam, N. M.; Martínez-Medina, A., Cascading Effects of Root Microbial Symbiosis on the Development and Metabolome of the Insect Herbivore Manduca sexta L. Metabolites 2021, 11 (11).
- Kinyungu, S. W.; Agbessenou, A.; Subramanian, S.; Khamis, F. M.; Akutse, K. S. , One stone for two birds: Endophytic fungi promote maize seedlings growth and negatively impact the life history parameters of the fall armyworm, Spodoptera frugiperda. Frontiers in physiology 2023, 14, 1253305. [Google Scholar] [CrossRef]
- Di Lelio, I.; Forni, G.; Magoga, G.; Brunetti, M.; Bruno, D.; Becchimanzi, A.; De Luca, M. G.; Sinno, M.; Barra, E.; Bonelli, M.; Frusciante, S.; Diretto, G.; Digilio, M. C.; Woo, S. L.; Tettamanti, G.; Rao, R.; Lorito, M.; Casartelli, M.; Montagna, M.; Pennacchio, F. , A soil fungus confers plant resistance against a phytophagous insect by disrupting the symbiotic role of its gut microbiota. Proceedings of the National Academy of Sciences of the United States of America 2023, 120(10), e2216922120. [Google Scholar] [CrossRef]
- Pappas, M. L.; Samaras, K.; Koufakis, I.; Broufas, G. D. , Beneficial Soil Microbes Negatively Affect Spider Mites and Aphids in Pepper. Agronomy 2021, 11(9), 1831. [Google Scholar] [CrossRef]
- Medeiros, H. A. d.; Araújo Filho, J. V. d.; Freitas, L. G. d.; Castillo, P.; Rubio, M. B.; Hermosa, R.; Monte, E. , Tomato progeny inherit resistance to the nematode Meloidogyne javanica linked to plant growth induced by the biocontrol fungus Trichoderma atroviride. Scientific Reports 2017, 7(1), 40216. [Google Scholar] [CrossRef]
- Fan, H.; Yao, M.; Wang, H.; Zhao, D.; Zhu, X.; Wang, Y.; Liu, X.; Duan, Y.; Chen, L. , Isolation and effect of Trichoderma citrinoviride Snef1910 for the biological control of root-knot nematode, Meloidogyne incognita. BMC Microbiology 2020, 20(1), 299. [Google Scholar] [CrossRef]
- Saharan, R.; Patil, J. A.; Yadav, S.; Kumar, A.; Goyal, V. , The nematicidal potential of novel fungus, Trichoderma asperellum FbMi6 against Meloidogyne incognita. Scientific Reports 2023, 13(1), 6603. [Google Scholar] [CrossRef] [PubMed]
- Berini, F.; Caccia, S.; Franzetti, E.; Congiu, T.; Marinelli, F.; Casartelli, M.; Tettamanti, G. , Effects of Trichoderma viride chitinases on the peritrophic matrix of Lepidoptera. Pest Management Science 2016, 72(5), 980–989. [Google Scholar] [CrossRef] [PubMed]
- Vijayakumar, N.; Alagar, S.; Madanagopal, N. , Effects of chitinase from Trichoderma viride on feeding, growth and biochemical parameters of the rice moth, Corcyra cephalonica Stainton. Journal of Entomology and Zoology Studies 2016, 4(4), 520–523. [Google Scholar]
- Massry, S.; Shokry, H. M.; Hegab, M. In EFFICIENCY OF Trichoderma harzianum AND SOME ORGANIC ACIDS ON THE COTTON BOLLWORMS, Earias insulana AND Pectinophora gossypiella, 2016.
- Chinnaperumal, K.; Govindasamy, B.; Paramasivam, D.; Dilipkumar, A.; Dhayalan, A.; Vadivel, A.; Sengodan, K.; Pachiappan, P. , Bio-pesticidal effects of Trichoderma viride formulated titanium dioxide nanoparticle and their physiological and biochemical changes on Helicoverpa armigera (Hub.). Pesticide Biochemistry and Physiology 2018, 149, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S. K.; Pal, S. , Entomopathogenic potential of Trichoderma longibrachiatum and its comparative evaluation with malathion against the insect pest Leucinodes orbonalis. Environmental Monitoring and Assessment 2015, 188(1), 37. [Google Scholar] [CrossRef]
- Zhou, D.; Huang, X.-F.; Guo, J.; dos-Santos, M. L.; Vivanco, J. M. , Trichoderma gamsii affected herbivore feeding behaviour on Arabidopsis thaliana by modifying the leaf metabolome and phytohormones. Microbial Biotechnology 2018, 11(6), 1195–1206. [Google Scholar] [CrossRef]
- Wang, K. D.; Gorman, Z.; Huang, P. C.; Kenerley, C. M.; Kolomiets, M. V. , Trichoderma virens colonization of maize roots triggers rapid accumulation of 12-oxophytodienoate and two ᵧ-ketols in leaves as priming agents of induced systemic resistance. Plant signaling & behavior 2020, 15 (9), 1792187. [Google Scholar]
- Yan, Y.; Huang, P.-C.; Borrego, E.; Kolomiets, M. , New perspectives into jasmonate roles in maize. Plant signaling & behavior 2014, 9 (10), e970442–e970442. [Google Scholar]
- Huang, H.; Liu, B.; Liu, L.; Song, S. , Jasmonate action in plant growth and development. Journal of Experimental Botany 2017, 68(6), 1349–1359. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Song, L.; Gong, X.; Xu, J.; Li, M. , Functions of Jasmonic Acid in Plant Regulation and Response to Abiotic Stress. International journal of molecular sciences 2020, 21 (4). [Google Scholar] [CrossRef]
- Bažok, R.; Lemić, D.; Chiarini, F.; Furlan, L. , Western Corn Rootworm (Diabrotica virgifera virgifera LeConte) in Europe: Current Status and Sustainable Pest Management. Insects 2021, 12 (3). [Google Scholar] [CrossRef]
- Paddock, K. J.; Robert, C. A. M.; Erb, M.; Hibbard, B. E. , Western Corn Rootworm, Plant and Microbe Interactions: A Review and Prospects for New Management Tools. Insects 2021, 12 (2). [Google Scholar] [CrossRef]
- Carrière, Y.; Brown, Z.; Aglasan, S.; Dutilleul, P.; Carroll, M.; Head, G.; Tabashnik, B. E.; Jørgensen, P. S.; Carroll, S. P. , Crop rotation mitigates impacts of corn rootworm resistance to transgenic Bt corn. Proceedings of the National Academy of Sciences 2020, 117(31), 18385–18392. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.; Kenerley, C. , Detection and enumeration of a genetically modified fungus in soil environments by quantitative competitive polymerase chain reaction. FEMS Microbiology Ecology 2006, 25, 419–428. [Google Scholar] [CrossRef]
- Schneider, C. A.; Rasband, W. S.; Eliceiri, K. W. , NIH Image to ImageJ: 25 years of image analysis. Nature Methods 2012, 9(7), 671–675. [Google Scholar] [CrossRef]
Figure 1.
Both T. virens Δsir1 and Δsm1Δsir1 treatments enhanced greater level of ISR against C. graminicola than TvWT. Lesion areas of maize seedlings colonized by different T. virens strains after inoculation with C. graminicola for 7 days. Bars are means ± SE. Different letters indicate statistically significant differences (Tukey’s HSD test, P < 0.05).
Figure 1.
Both T. virens Δsir1 and Δsm1Δsir1 treatments enhanced greater level of ISR against C. graminicola than TvWT. Lesion areas of maize seedlings colonized by different T. virens strains after inoculation with C. graminicola for 7 days. Bars are means ± SE. Different letters indicate statistically significant differences (Tukey’s HSD test, P < 0.05).
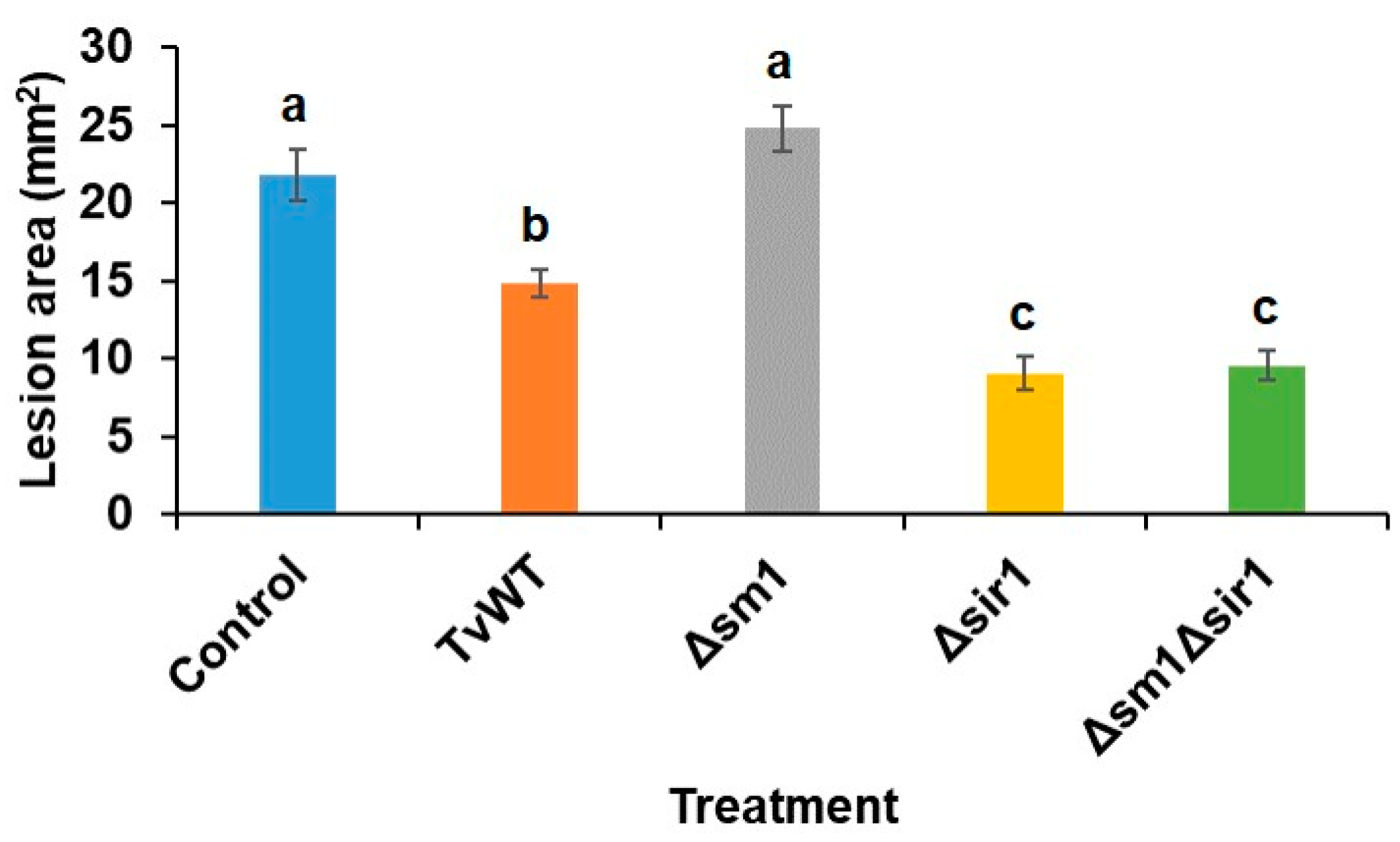
Figure 2.
T. virens colonization suppressed insect defense. One-week old maize seedlings were colonized by T. virens strains for 14 days before FAW infestation. (A) FAW larval weight was determined after neonates feeding on maize plants for 7 days. (B) Maize leaf consumed area by 3rd-instar FAW larvae for 6 hours. Bars are means ± SE. Different letters indicate statistically significant differences (Tukey’s HSD test, P < 0.05).
Figure 2.
T. virens colonization suppressed insect defense. One-week old maize seedlings were colonized by T. virens strains for 14 days before FAW infestation. (A) FAW larval weight was determined after neonates feeding on maize plants for 7 days. (B) Maize leaf consumed area by 3rd-instar FAW larvae for 6 hours. Bars are means ± SE. Different letters indicate statistically significant differences (Tukey’s HSD test, P < 0.05).
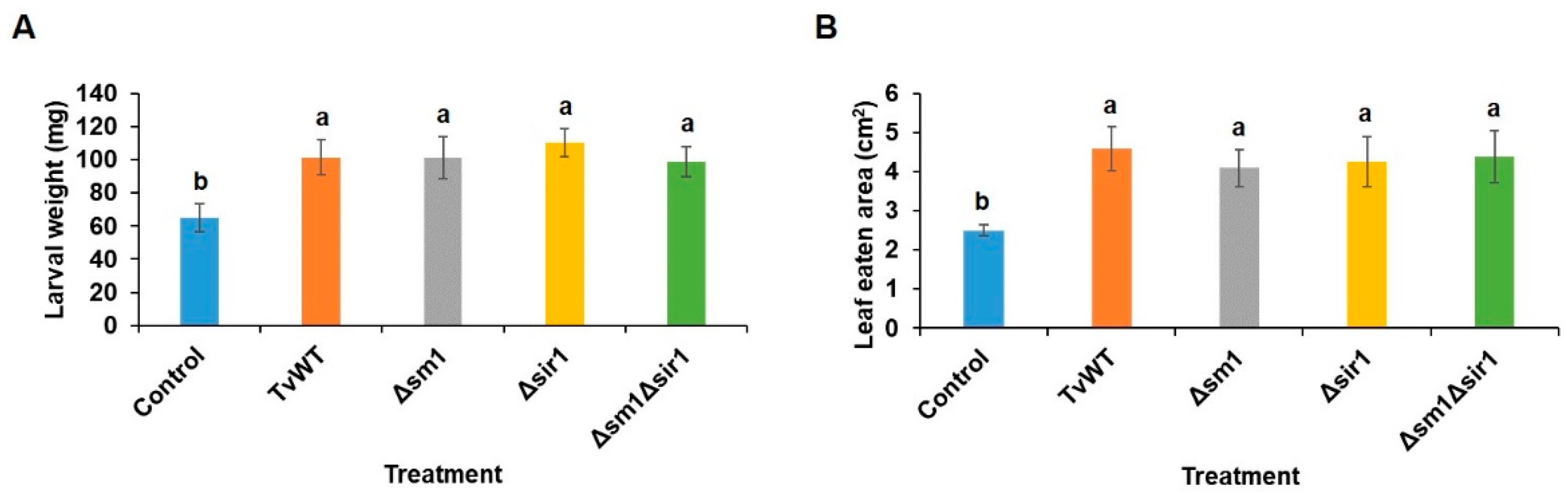
Figure 3.
T. virens colonization suppressed wound-induced JA biosynthesis and enhanced JA catabolism. (A) Heatmap showing relative accumulation of oxylipins and phytohormones in leaves of B73 seedlings colonized by different T. virens stains or untreated control at 0, 1, and 2 hours post wounding. Contents of (B) JA, (C) JA-Ile, (D) 12OH-JA, and (E) 9,10-KODA in different T. virens strains colonized or control plants at 0, 1, and 2 hours post wounding. Abbreviation for compounds were listed in Supplemental Table 1. Bars are means ± SE. Different letters indicate statistically significant differences within the same time points (Tukey’s HSD test, P < 0.05).
Figure 3.
T. virens colonization suppressed wound-induced JA biosynthesis and enhanced JA catabolism. (A) Heatmap showing relative accumulation of oxylipins and phytohormones in leaves of B73 seedlings colonized by different T. virens stains or untreated control at 0, 1, and 2 hours post wounding. Contents of (B) JA, (C) JA-Ile, (D) 12OH-JA, and (E) 9,10-KODA in different T. virens strains colonized or control plants at 0, 1, and 2 hours post wounding. Abbreviation for compounds were listed in Supplemental Table 1. Bars are means ± SE. Different letters indicate statistically significant differences within the same time points (Tukey’s HSD test, P < 0.05).
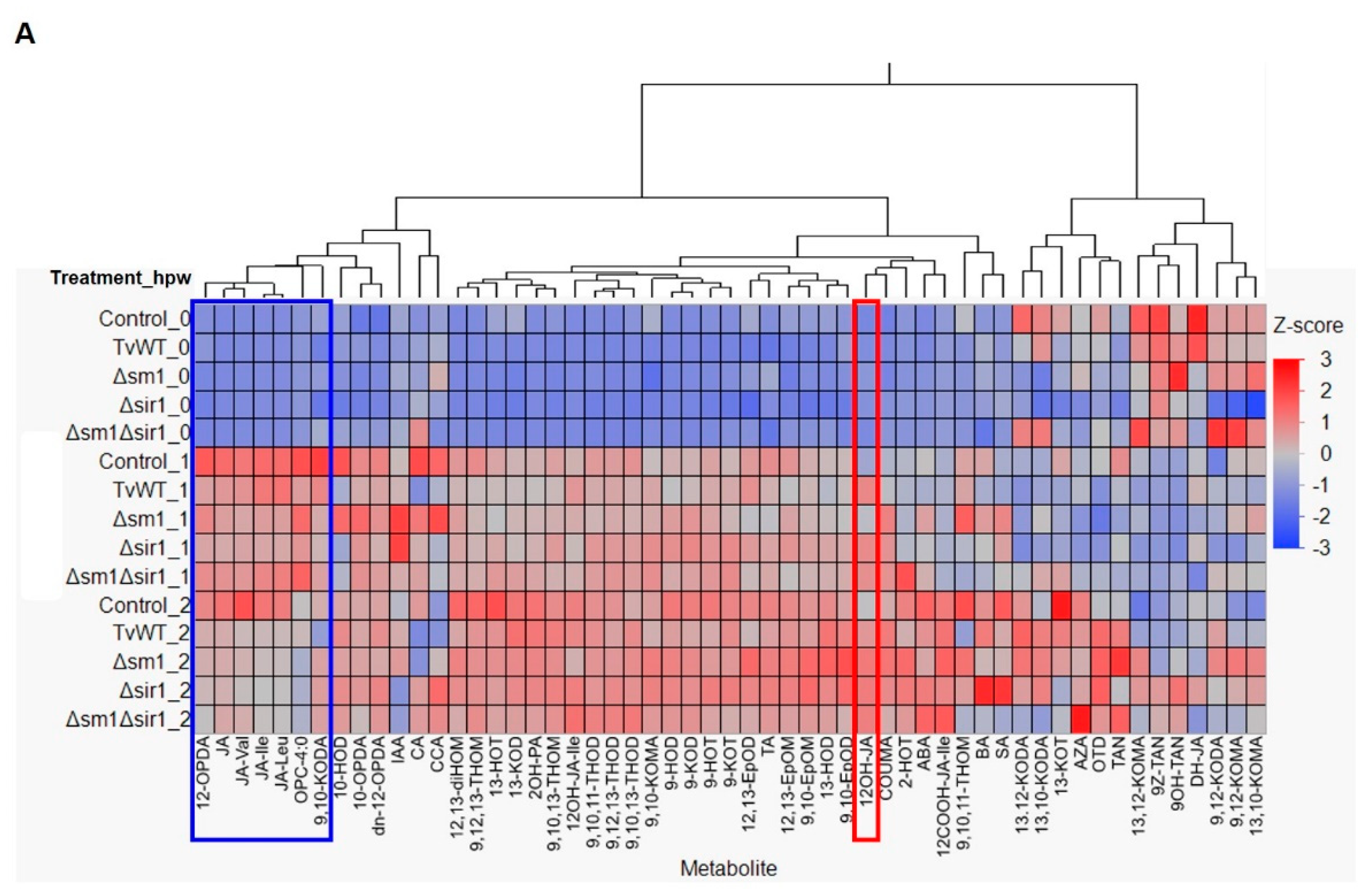
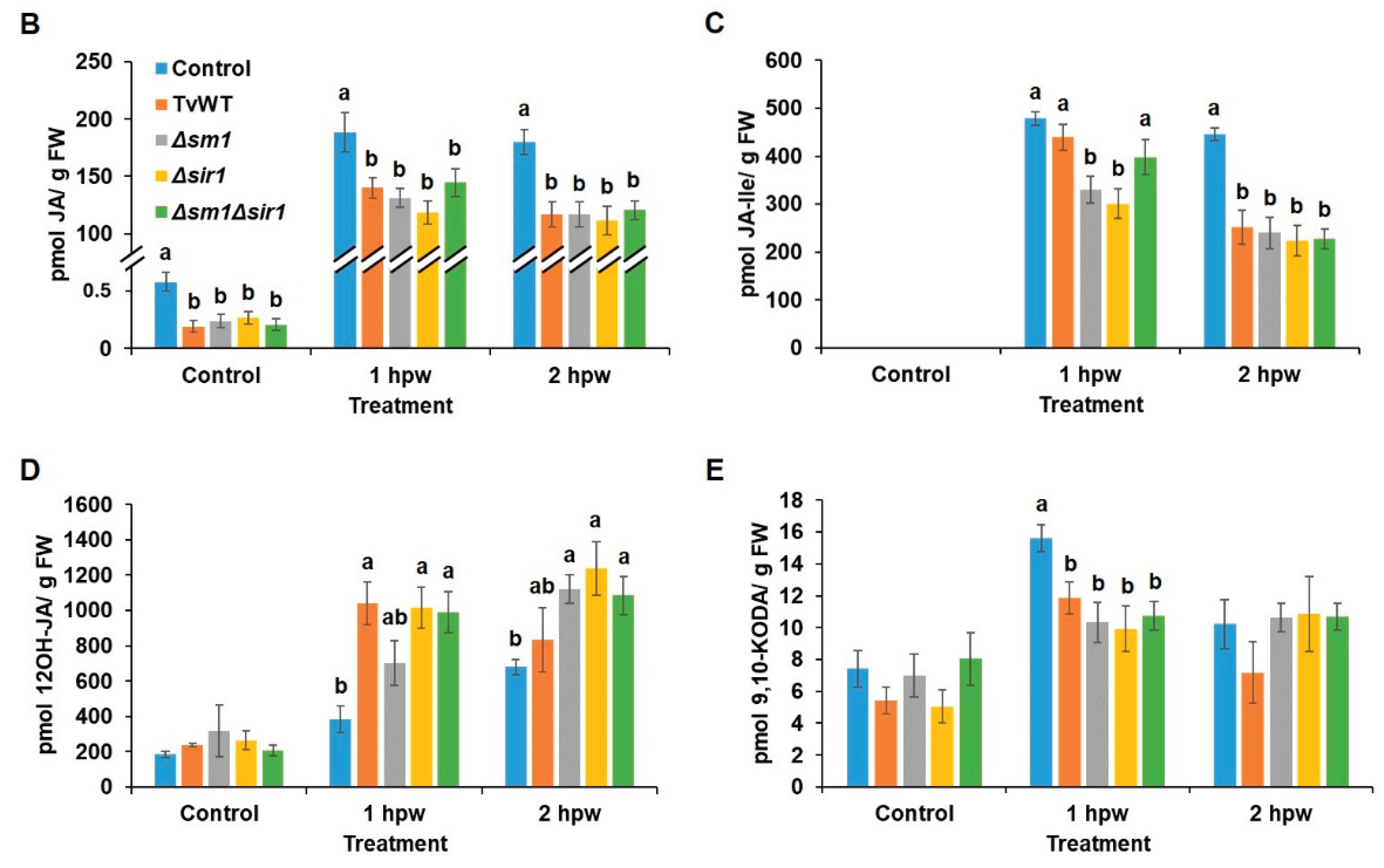
Figure 4.
Reduced defense against FAW by T. virens colonization is modulated by JA signaling. Maize B73 wild-type or opr7opr8 double mutant seedlings were colonized by TvWT for 14 days or left as untreated control (CK) before FAW infestation. FAW larval weight was determined after neonates feeding on maize plants for 7 days. Bars are means ± SE. Different letters indicate statistically significant differences on log-transformed data (Two-way ANOVA with Tukey’s HSD test, P < 0.05).
Figure 4.
Reduced defense against FAW by T. virens colonization is modulated by JA signaling. Maize B73 wild-type or opr7opr8 double mutant seedlings were colonized by TvWT for 14 days or left as untreated control (CK) before FAW infestation. FAW larval weight was determined after neonates feeding on maize plants for 7 days. Bars are means ± SE. Different letters indicate statistically significant differences on log-transformed data (Two-way ANOVA with Tukey’s HSD test, P < 0.05).
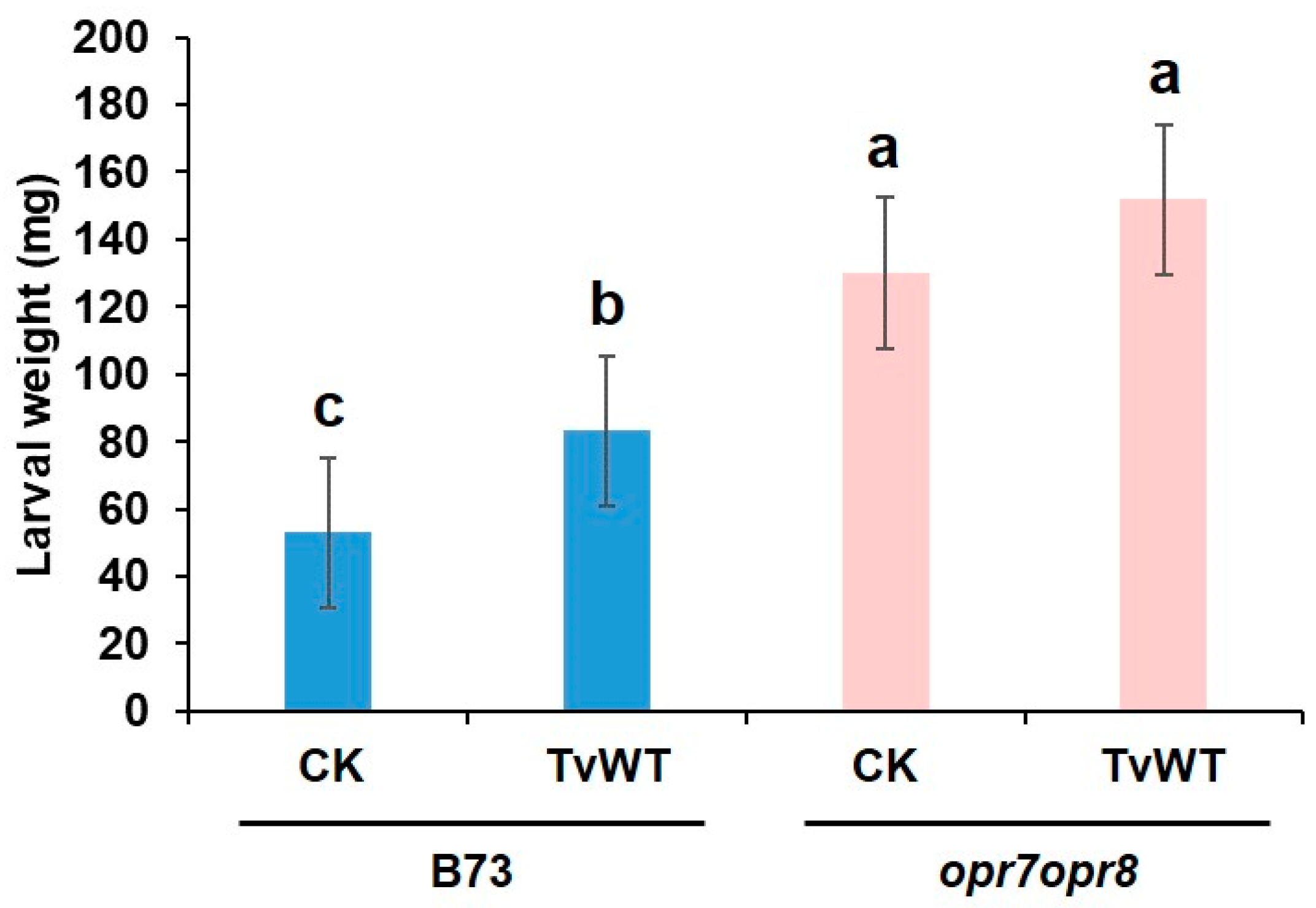
Figure 5.
T. virens and P. chlororaphis 30-84 colonization enhanced growth but suppressed defense against FAW only in sterile soil while increased resistance to C. graminicola in both soil types. Maize seedlings were colonized by TvWT, P chlororaphis 30-84, or both for 14 days before plant height measurement and C. graminicola infection or FAW infestation. (A) Plant height measured at 14 days after colonization. (B) FAW larval weight and (C) lesion areas were determined at 7 days after infection or infestation. Bars are means ± SE. Different letters indicate statistically significant differences on log-transformed data within the same treatment (Tukey’s HSD test, P < 0.05). Asterisks represent statistically significant differences between sterile and nonsterile soils (Student’s t-test, **P < 0.01, ***P < 0.001).
Figure 5.
T. virens and P. chlororaphis 30-84 colonization enhanced growth but suppressed defense against FAW only in sterile soil while increased resistance to C. graminicola in both soil types. Maize seedlings were colonized by TvWT, P chlororaphis 30-84, or both for 14 days before plant height measurement and C. graminicola infection or FAW infestation. (A) Plant height measured at 14 days after colonization. (B) FAW larval weight and (C) lesion areas were determined at 7 days after infection or infestation. Bars are means ± SE. Different letters indicate statistically significant differences on log-transformed data within the same treatment (Tukey’s HSD test, P < 0.05). Asterisks represent statistically significant differences between sterile and nonsterile soils (Student’s t-test, **P < 0.01, ***P < 0.001).
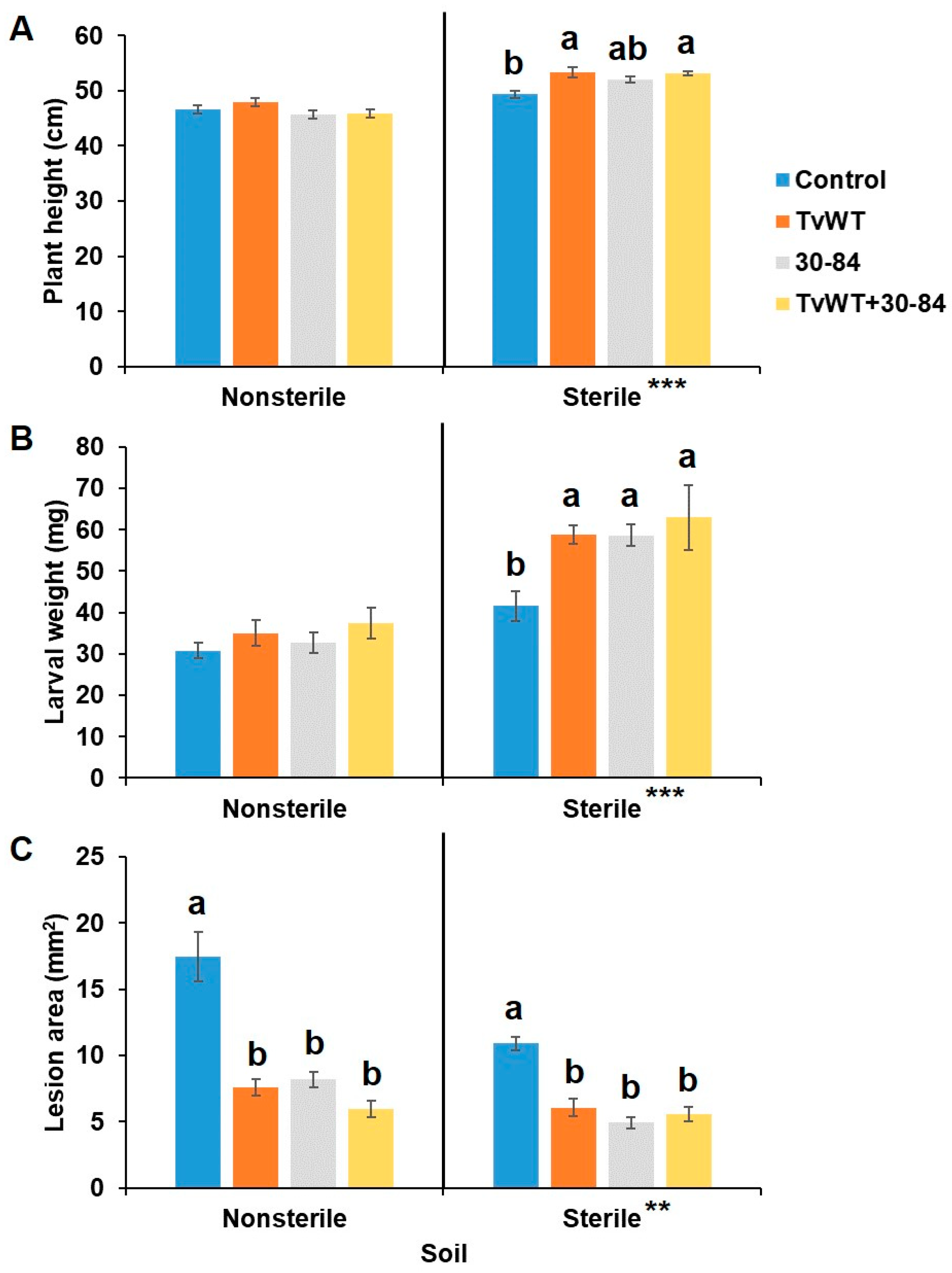
Figure 6.
T. virens and P. chlororaphis 30-84 colonization suppressed wound-induced JA biosynthesis and enhanced JA catabolism in sterile soil. (A) Heatmap showing relative accumulation of oxylipins and phytohormones. Contents of (B) JA, (C) JA-Ile, (D) 12OH-JA, and (E) 9,10-KODA in plants colonized by T. virens, P. chlororaphis 30-84, and both or control in response to wounding at 0 and 1 hpw. Abbreviations for samples: NS, nonsterile soil; SS, sterile soil. Abbreviation for compounds were listed in Supplemental Table 1. Bars are means ± SE. Different letters indicate statistically significant differences within the same treatment per time point (Tukey’s HSD test, P < 0.05).
Figure 6.
T. virens and P. chlororaphis 30-84 colonization suppressed wound-induced JA biosynthesis and enhanced JA catabolism in sterile soil. (A) Heatmap showing relative accumulation of oxylipins and phytohormones. Contents of (B) JA, (C) JA-Ile, (D) 12OH-JA, and (E) 9,10-KODA in plants colonized by T. virens, P. chlororaphis 30-84, and both or control in response to wounding at 0 and 1 hpw. Abbreviations for samples: NS, nonsterile soil; SS, sterile soil. Abbreviation for compounds were listed in Supplemental Table 1. Bars are means ± SE. Different letters indicate statistically significant differences within the same treatment per time point (Tukey’s HSD test, P < 0.05).
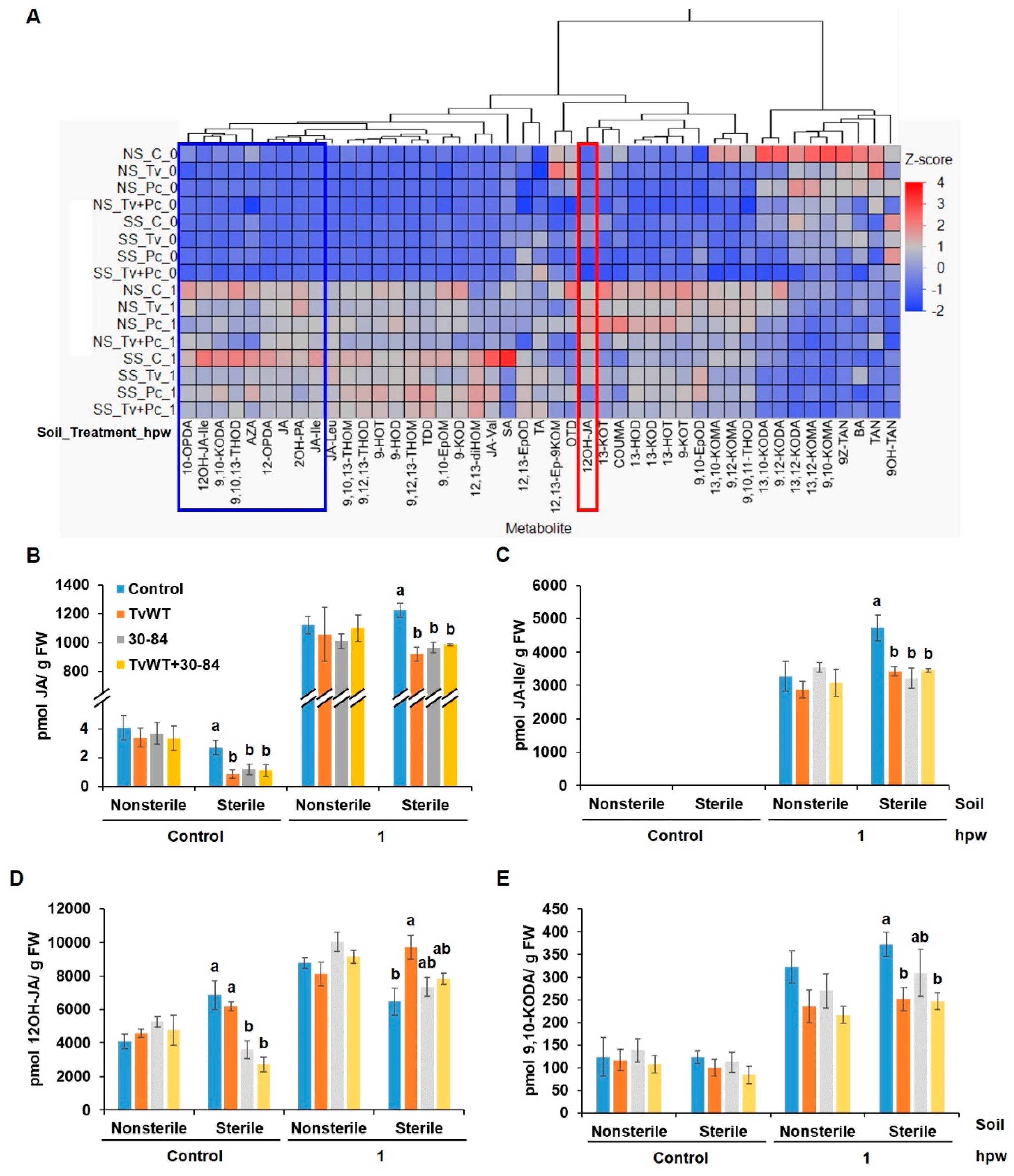
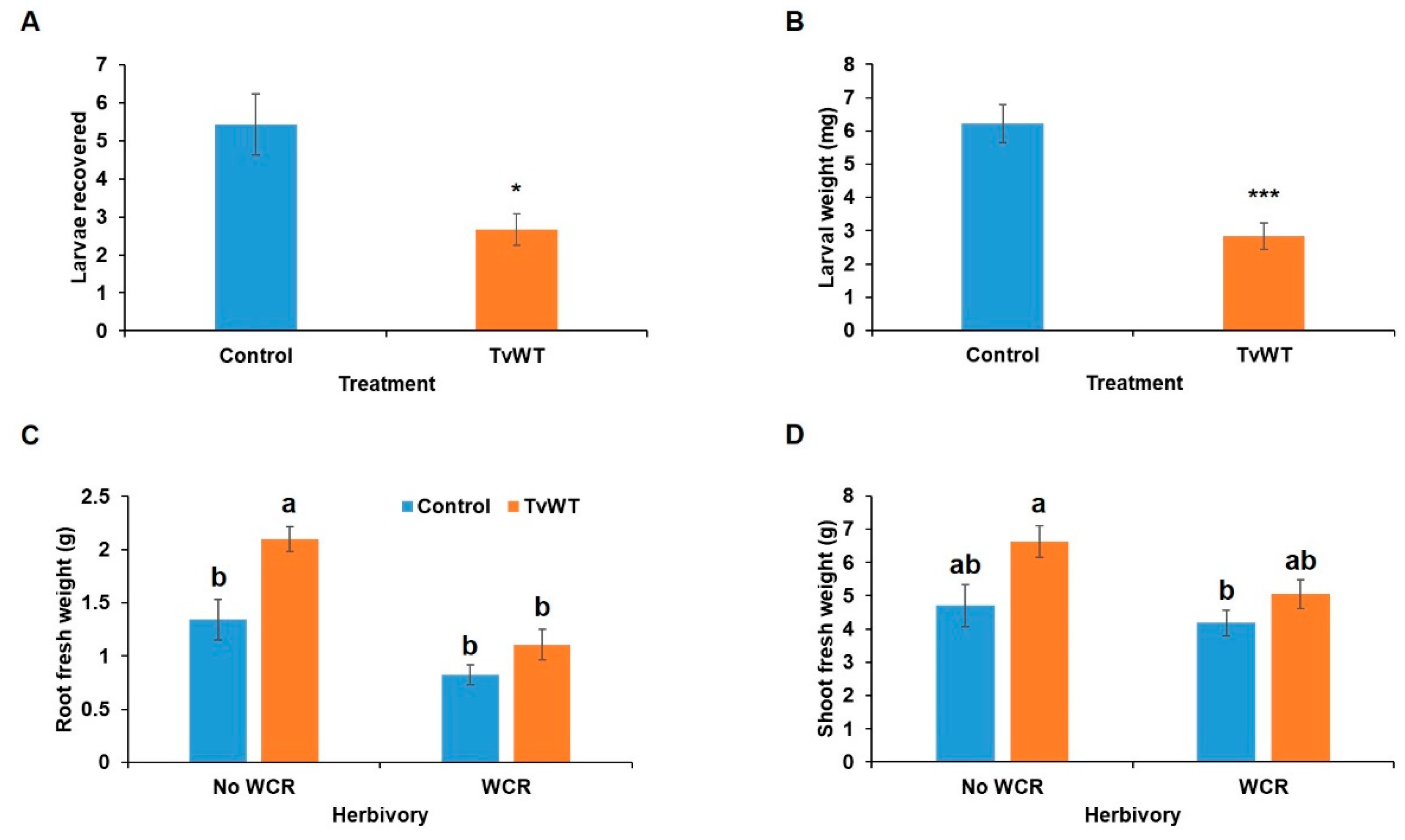
Figure 7.
T. virens colonization suppressed WCR larval survival and weight gain. (A) Larvae recovery, (B) larval mass, (C) root, and (D) shoot ratios were recorded 10 days post-infestation. Bars are means ± SE. Asterisks represent statistically significant differences (Student's t-test, * P < 0.05, *** P < 0.001). Different letters indicate statistically significant differences (Two-way ANOVA with Tukey’s HSD test, P < 0.05).
Figure 7.
T. virens colonization suppressed WCR larval survival and weight gain. (A) Larvae recovery, (B) larval mass, (C) root, and (D) shoot ratios were recorded 10 days post-infestation. Bars are means ± SE. Asterisks represent statistically significant differences (Student's t-test, * P < 0.05, *** P < 0.001). Different letters indicate statistically significant differences (Two-way ANOVA with Tukey’s HSD test, P < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



