
Submitted:
20 April 2023
Posted:
20 April 2023
You are already at the latest version
Abstract
Irrigation and fertilizer are two essential factors affecting rice root traits and yield. In this respect, a pot experiment was performed at the boro (dry season irrigated) season of 2021-22 in the Department of Agronomy of Bangladesh Agricultural University, Mymensingh, Bangladesh. The variety Binadhan-10 was planted at two irrigation conditions i.e. saturation (S) and continuous flooding (CF); two potassium (K) dose e.g. 65 kg ha-1 (K65) and 98 kg ha-1 (K98) and two nitrogen (N) dose i.e. 140 kg ha-1 (N140) and 210 kg ha-1 (N210). The experiment was laid in split plot design and replicated thrice. The findings confirmed the effectiveness of irrigation, K and N on root number (RN), root length (RL), root volume (RV), leaf area index (LAI), total dry matter (TDM), yield attributes and yield. Considering interaction among irrigation, K and N, S condition with K65 and N140 showed best performance. At 80 DAT, the highest RN (373.00), RL (1700.00 cm), RV (8.90 cm3 hill-1), LAI (4.94) and TDM (25.83 g plant-1) was obtained from this combination. Grain yield (GY) and root traits except root porosity, showed a significant positive association. Grain yield (GY) was highest (27.12 g pot-1) in S condition with K65 and N140.
Keywords:
root volume
; total dry matter
; correlation matrix
; cross sectional view
; harvest index
1. Introduction
Rice (Oryza sativa L.), a significant crop in the world which is a crucial component of the daily nutrition for almost 3 billion people worldwide [1].The basic functions of the roots of plant are to absorb water and nutrient from the surrounding soil [2]. The roots play a crucial part in a variety of plant processes, such as the intake of water and ions of nutrient, the generation of plant hormones, amino acids, and organic acids, as well as providing anchorage to the plants [3]. The physiology and structure of roots are closely connected to the development and growth of aboveground parts of the plants [4]. Plant breeders are becoming more and more aware of the value of root development for preserving crop yields [5].
One of the most prevalent substances on earth, nitrogen (N), is crucial to contemporary agriculture [6]. N fertilizer has a substantial impact on root growth and Gaudin et al. [7] revealed that reduced amounts of N promoted root elongation. However, a rise in N level was linked to an increase in biomass and root length was discovered by Fan et al. [8].It is believed that a lack of N fertilizer will lead to a low N concentration in the soil which reduce N absorption by the plant, accumulation of root and shoot biomass [9] and inhibit root extension. N deficiency in the roots might prevent root development, which would therefore limit the growth of the shoots and reduce yield.
The most prevalent inorganic cation and essential for promoting healthy plant growth is potassium (K) [10]. Numerous crucial enzymes, including those involved in synthesis of protein, transportation of sugar, photosynthesis process and metabolism of C and N are activated by K. For greater yield and quality, it is essential [11].K in plants serves a number of purposes, including regulating the cell cycle, maintaining both root and shoot development, and carrying out cell death programs [12,13]. K shortage was often associated with slower root growth and a poorer ratio of root to shoot biomass [14,15], while in certain instances, the ratio of root and shoot remains stable or somewhat rises at varied K levels [16].
The formation of roots, viability and development of plants are all greatly impacted by the moisture content of the soil [17]. Root development, water and nutrient intake and plant growth are all impacted by the soil's texture, which also impacts air and water circulation in the soil. We can learn a lot about water stress from studying rice roots, including how it occurs, how it's acquired, how to respond to it, and how to tolerate it [18]. Aerobic adaptation requires an understanding of how roots respond, particularly how effectively they absorb water [19]. Higher penetration, length of roots, and root to shoot weight ratio are root-related traits that make aerobic rice agriculture more able to adapt to water shortage conditions [20].The development of crops depends on roots' capacity to uptake water as well as nutrient. Their importance is heightened in arid regions where plants must spread their roots into deeper soils to get the nutrients that are available in the wet soil because the top soil is typically dry and nutrient-deficient. The relationship between crop yield and root biomass is often demonstrated to be significant and almost invariably linear [21].
Water has a direct impact on the soil nutrient availability needed for plant development and agricultural production. The physiological mechanisms of nutrient uptake by plants are probably determined how nutrient interactions with water affect plant traits and, ultimately, crop development. Soil water deficiency can therefore reduce nutrient transfer merely by limiting the amount of water that reaches the plant. The transpiration stream is the pathway through which minerals and other nutrients are moved from root to shoot [22].Plant root development is intimately linked to soil variables such as moisture, oxygen, temperatures, and fertility, with moisture and the fertility being the two most important ones. These factors are also interdependent and interact with one another [23].When fertilizers are given to the soil or substrate, water interacts with those nutrients in a way that either positively or negatively influences plant growth [24]. The significance of soil nutrients for plant development and agricultural output is directly correlated with the availability of water. Crops' capacity to get nutrients is considerably affected by water through (i) the transformation of nutrients to usable forms, (ii) the transportation of nutrient near to roots, and (iii)loss mechanisms [25]. Therefore, a relationship exists between water and nutrients on plant and root growth and yield. The majority of earlier research, however, did not take into account how water, N, and K interact to affect root development, plant growth, and yield. In light of this, the purpose of this research was to assess variation of root traits and grain yields of Binadhan-10 in relation to their interactions under different irrigation regime with different K and N treatments.
2. Materials and Methods
2.1. Site and plant materials
In the net house of the Department of Agronomy, Bangladesh Agricultural University (latitude: 24042’55”, longitude: 90025’47”), the experiment was conducted in boro seasons of 2021-22. The experimental site is in the Old Brahmaputra floodplain (AEZ-9) [26] having subtropical monsoon climate with a humid environment. VarietyBinadhan-10 was used as study materials. From the Bangladesh Institute of Nuclear Agriculture (BINA), inbred variety Binadhan-10’s seeds were collected.
2.2. Experimental design and crop management
The split plot design was used to perform this experiment. The irrigation treatments were I1-Saturation (S), I2- Continuous flooding (CF); the K treatments were 65 kg ha-1 (K65), 98 kg ha-1 (K98) and the N treatments were 140 kg ha-1 (N140) and 210 kg ha-1 (N210). Each pot (30L plastic pots having 35 cm diameters) was put inside the net house with 25 kg of soil. The gathered soil was sun-dried, then crushed and well blended before putting into the pots. First a soil-filled pot was weighed, and then a porous pot was submerged during the night in a bowl of water to maintain saturation. Following that, the weight was taken, a computation for absorbing water was made, and it was treated as saturation. Before seedling transplanting, irrigation treatments were applied using drip irrigation method, and they were kept up until harvest while regulating various levels of saturation by following gravimetric method. Fertilizer doses (AEZ basis) for pot experiment were applied as 2.5 g, 2.81 g and 0.09 g pot-1as triple super phosphate (TSP), gypsum and zinc sulphate, respectively [27].Muriate of potash (MoP) served as the source of K and for K65 (3.25 g MoP pot-1), for K98 (4.9 g MoP pot-1) was applied during the final pot preparation as. The nitrogen supply was as urea and N for N140 (7.59 g urea pot-1) and N210 (11.39 g urea pot-1). During the final pot preparation, one-third of urea and entire amounts of all other fertilizers were added. The rest of urea was applied at 20 and 40 days after transplanting (DAT).The Binadhan-10 seedlings, which had previously been raised in the seedbed, were transplanted into the pot after they had reached forty days old. Occasionally, mostly in the early phases of development, weeds were noticed and removed. Notable insects and diseases were not found.
2.3. Determination of root morphological and physiological traits
Root morphological characteristics were noted at 20, 40, 60, 80 DAT and at harvest stage. About 3 plants were carefully removed from each pot using a deep dig to ensure that the main tap root and all lateral roots could be uprooted safely. The tested plants were kept in water-filled plastic bags for about 12 hours. The roots were extensively cleaned using 1 mm mesh sieves in order to ensure that no root was left behind and to make easy root separation possible. The estimated value of various features was then averaged.
2.3.1. Number of root (RN)
The rice plants were gently uprooted after being watered. The roots were removed and then cleaned under running water. Every plant's RN was carefully counted and averaged.
2.3.2. Root length (RL, cm)
The length of the root was determined in centimeters from base to the tip of root, and the total of the measurements was calculated.
2.3.3. Root volume (RV, cm3 hill-1)
In order to measure root volume, the root masses were placed into water filled measuring cylinder. The increase in water level was measured and expressed as cm3 hill-1[28].
2.3.4. Root porosity (RP, %)
The stored roots were kept in water with an airtight polybags to maintain their original temperature. Measurements were made of the pycnometer vials' weights with and without water. The sample roots were gently blotted using tissue paper. An analytical balance was used to calculate the root weight. The roots were inserted into a water-filled vial. In the event that air bubbles were found, they were expelled by gently moving the immersed roots with a clean needle within the pycnometer vial. An analytical balance was used to determine the weight of water filled pycnometer and fresh roots. After that, the roots were removed from the vial and blended in a glass mortar and pestle. The homogenate was transferred completely and filled the pycnometer to capacity. Weight was obtained after bringing the homogenate and pycnometer to room temperature. The following formula was used to calculate porosity [29]:
Here, Whr+w = weight of homogenized roots and water filled pycnometer vial, Wfr+w= weight of fresh roots and water filled pycnometer vial, Ww = weight of water filled pycnometer vial, Wfr = weight of fresh roots
2.3.5. Leaf area Index (LAI)
The leaf area (LA) was calculated using a leaf area meter (LI 3100, Licor, Inc., LincoIn, NE, USA) after the leaf blades and sheaths had been separated. The ratio of LA to ground area was calculated to be the leaf area index (LAI).The usual formulas were used to compute the LAI, crop growth rate (CGR), relative growth rate (RGR), and net assimilation rate (NAR) [30,31].
2.3.6. Total dry matter (TDM)
Three hills (plants) from each pot were uprooted at each development stage. Collected leaves, culms, and panicles of the samples were oven dried in brown paper bags for 72 hours at 650C before being weighed by an electronic balance to get the average data on their dry weights(ghill-1).Summarizing the weights of dry plant parts yielded total dry matter (TDM).
2.3.7. Yield parameters and yield
At maturity (90% ripened grain), the entire plant was cut with a sickle at the ground level. The weight of the rice grains was determined and reported as g pot-1 after adjusting for the 14% moisture content. Data for plant height (PH), no. of effective tiller plant-1 (ET), length of panicle (PL), no. of grains panicle-1 (GP), weight of 1000 grains (TGW), grain yield (GY) and straw yield (SY) for each plant were noted. The harvest index (HI, %) was calculated by dividing the grain biomass into the biological biomass of plant [32].
2.4. Root cross section
The cross section of root was seen by cutting 2 cm of the root from the tip. The root was then preserved in water to keep it fresh and it was examined under microscope.
2.5. Statistical analysis of data
The statistical program JMP Pro 16(SAS Institute Inc., Cary, NC, USA), was used to conduct the two-way analysis of variance (ANOVA) test. Tukey's honestly significant difference (HSD) post hoc test was used to examine the mean differences at the 0.05 and 0.01 probability levels. The data visualization and correlation matrix were developed using R (R for Windows 4.1.2) and Sigma Plot v14 (Systat Software, Inc., San Jose, CA, USA, http:// www.systatsoftware.com) (accessed on 20 March 2023) [33].
3. Results
3.1. Morphological traits of root, total dry matter and leaf area index
The RN and RL were significantly affected by irrigation, K and N at 20, 40, 60, 80 DAT and at harvest stage. At every observation RN was highest in S condition, at K65 and at N140 level while lowest value was found in CF, at K98 and at N210 level. In case of RL similar trend was noticed. At 80 DAT higher RN in S condition (368.7), at K65 (365.50) and at N140 (364) was observed. RL was also maximum in S condition (1690.50 cm), at K65 (1683.67 cm) and at N140 (1679.58 cm) at 80 DAT, while minimum was recorded in CF (1663.48 cm), at K98 (1670.31 cm) and at N210 (1674.40 cm) (data not shown). The RN and RL were also greatly influenced by the interaction among irrigation, K and N at all observations (Figure 1). At 80 DAT in case of RN the value ranged from 373.00 to 351.67. The highest value was found in interaction among S condition, K65 and N140 (373.00) while lowest was observed in interaction among CF, K98 and N210 (351.67) at 80 DAT. At the same time the highest value of RL was noticed in interaction among S condition, K65 and N140 (1700.00 cm) and lowest was registered in interaction among CF, K98 and N210 (1655.75 cm) at 80 DAT.
RV and RP were significantly affected by irrigation, K and N at all observations. RV increased sharply up to 80 DAT and at this, highest RV was noticed in S condition (8.86 cm3 hill-1), at K65 (8.83 cm3 hill-1) and at N140 (8.81 cm3 hill-1). In case of RP at 80 DAT highest value was observed in CF (27.47 %), at K98 (27.04 %) and at N210 (26.94 %) and lowest value of RP was found in S condition (25.89 %), at K65 (26.33 %) and at N140 (26.43 %) (data not shown).
The interaction among irrigation, K, and N had a substantial impact on RV and RP throughout all observations (Figure 1). In case of RV the value ranged from 8.90 to 8.67 (cm3 hill-1) at 80 DAT. At 80 DAT the highest value was found in interaction among S condition, K65 and N140 (8.90 cm3 hill-1) while lowest was observed in interaction among CF, K98 and N210 (8.67 cm3 hill-1). At the same time the maximum RP value was noticed in interaction among CF, K98 and N210 (28.09 %) and lowest was registered in interaction among S condition, K65 and N140 (25.52 %) at 80 DAT.
The impact of irrigation, K and N on the LAI was significant. The value of LAI reached peak at 80 DAT and then it was decreased. In irrigation, the value of LAI at 80 DAT was higher (4.91) in S condition than CF (4.86). In case of K, the value was higher (4.90) at K65 than at K98 (4.87). Under N, the value was higher (4.90) at N140 than at N210 (4.88) (data not shown). The interaction among irrigation, K, and N had also a substantial impact on the LAI as well.At 80 DAT the highest (4.94) value of LAI was recorded in S condition along with K65 and N120 (Figure 2).
The effect of irrigation, K and N on TDM was significant at 20, 40, 60 and 80 DAT respectively. The value of TDM increased progressively and attained peak at 80 DAT. Under irrigation treatment at 80 DAT the higher (25.67 g plant-1) value of TDM was registered in S condition than CF (25.25 g plant-1). In case of K, the higher value was found at K65 (25.56 g plant-1) than K98 (25.36 g plant-1) at 80 DAT. On the other hand, in N at 80 DAT the value of TDM was higher at N140 (25.51 g plant-1) than N210 (25.41 g plant-1) (data not shown). The combination among irrigation, K, and N had a considerable impact on TDM. In this situation the highest (25.83 g plant-1) value of TDM was found in S condition along with K65 and N140 whereas lowest (25.11 g plant-1) value was noticed in CF along with K98 and at N210 at 80 DAT (Figure 2).
3.2. Growth parameters
CGR was greatly influenced by irrigation, K and N at 20-40 DAT (1st), 40-60 DAT (2nd) and at 60-80 DAT (3rd). In case of irrigation at 60-80 DAT the CGR was higher (9.26 g m-2 day-1) in S condition than in CF (9.20 g m-2 day-1). Under K, the value of CGR was higher (9.25 g m-2 day-1) at K65 than at K98 (9.21 g m-2 day-1) at 60-80 DAT. In N, the value was higher (9.24 g m-2 day-1) at N140 than at N210 (9.22 g m-2day-1) (data not shown). At all observation, the interaction of irrigation, K and N had a substantial impact on CGR, and the trend of CGR is linear with leaf area. At 60-80 DAT (3rd) the maximum (9.28 g m-2 day-1) CGR was registered with S condition along with K65 and N98 (Figure 3).
Irrigation, K and N had a substantial impact on RGR at 20-40 DAT (1st), 40-60 DAT (2nd) and at 60-80 DAT (3rd). The value of RGR was higher at initial growth stage and then it was diminishing trend. In case of irrigation, RGR was higher (9.08 mg g-1 day-1) in CF than S condition (8.96 mg g-1day-1) at 60-80 DAT (3rd). At same DAT, under K, the RGR was higher (9.04 mg g-1day-1) at K98 than at K65 (9.00 mg g-1 day-1). In N, the value of RGR was higher (9.03 mg g-1 day-1) at N210 than at N140 (9.01 mg g-1day-1) at 60-80 DAT (data not shown). The interaction among irrigation, K and N had a substantial impact on RGR. At 60-80 (3rd) DAT the highest (9.11 mg g-1 day-1) RGR value was noticed in CF with K98 and N210 and lowest (8.92 mg g-1 day-1) value was registered in S condition with K65 and N140 (Figure 3).
At every observation, the interaction among irrigation, K and N had a considerable impact on NAR. The figure revealed that when LAI increased, NAR decreased in all interactions, which may be related to increased tillering and leaf area development. At 40-60 (2nd) DAT the highest (0.39 mg cm-2 day-1) NAR value was found in CF with K98 and N210 and lowest (0.36 mg cm-2 day-1) value was registered in S condition with K65 and N140 (Figure 3).
3.3. Yield contributing parameters and Yield
Treatments with irrigation, K and N had a significant effect on rice yield components and output (Table 1). In irrigation, ET was higher (16.17) in S condition than in CF (13.67). K revealed that at K65 the value was higher (15.67) than at K98 (14.17). In case of N, the value was higher (15.33) at N140 than at N210 (14.50).The higher PL (24.29 cm) was found in S condition than in CF (18.67 cm) in case of irrigation. Under K, at K65 level higher value (22.50 cm) of PL was noticed than at K98 (20.46 cm). On the other hand, at N140 higher PL (21.92 cm) was observed than at N210 (21.04). In case of irrigation, the value of GP was higher (124.33) in S condition than in CF (117.67). Similarly, in case of K, the higher value (122.83) was found at K65 than at K98 (119.17). Under N, the higher (121.75) value of GP was found at N140 than at N210(120.25). In case of irrigation, the higher TGW (26.90 g)was noticed in S condition than in CF (22.81 g). In case of K, the higher (25.87 g) value was observed at K65 than at K98 (23.85 g). In N at N140 produced higher (121.75 g) value than at N210 (120.25 g). In case of GY, under irrigation treatment the value of grain yield was higher (25.70 g pot-1) in S condition than in CF (20.65 g pot-1). In K, the value was higher (24.25 g pot-1) at K65 than at K98 (22.10 g pot-1). On the other hand, higher (23.83 g pot-1) value of GY was found at N140 than at N210 (22.52 g pot-1).
The influence of interactions among irrigation, K and N treatments on yield and yield characteristics is shown in Figure 4. S condition with K65 and N140 produced highest (17.00) ET while lowest (12.00) ET was found at CF with K98 and N210. The highest value of PL (25.50 cm) was found in S condition along with K65 and N140 whereas lowest (17.00 cm) was registered in CF with K98 and N210. The highest GP (126.67) value was found in interaction among S condition, K65 and N140 whereas lowest value was found in interaction among CF, K98 and N210 (115.00). In case of TGW, the highest (28.00 g) value was registered in S condition with K65 and N140 while lowest value (20.75 g) was observed in CF with K98 and N210. Finally, the highest (27.12 g pot-1) value of GY was registered in S condition along with K65 and N140 while lowest (18.26 g pot-1) value was noticed in CF along with K98 and N210.
3.4. Relationship among root traits, growth indices, yield and yield attributes
The correlation matrix of root traits, growth indices, yield, and yield parametersis shown in Figure 5 to examine their relationship. LAI had significant and positive relationship with all root traits except RP. GY, SY and BY is significantly and positively correlated with RN, RL and RV while negative relationship withRP. Again, PH, ET, PL, GP, and TGW had significant relationships with RN, RL, and RV, while RP had relationships that were negative.
3.5. Root cross sectional view
The root cross section of Binadhan-10 at 80 DAT under different combination of treatments is shown in Figure 6. In case of S condition with K and N treatments there was no aerenchymatous tissue found. At combination of CF with K and N treatments developed aerenchymatous tissue was developed. From the cross section it was found that under CF at higher dose of K and N that is at K98 and at N210 the formation of aerenchyma was more than at K65 and at N140 level.
4. Discussion
There are research contains investigation on the solo impact of irrigation, potassium and nitrogen on rice roots. But interaction among irrigation, potassium and nitrogen on rice root is still limited. In this experiment, root traits, yield and yield contributing features of Binadhan-10 varied under interaction among different treatment of irrigation, K and N. This study set out to change in root traits and how they related to GY while subjected to various irrigation, K and N.
An essential agronomic indicator, the LAI, measures crop growth and forecasts crop production. Crop yields are significantly influenced by leaf area [34]. A proper LAI is a key indicator of great crop output, balancing the growth of each organ in crops and regulating source-sink relationship of crops. The growth factors that have contribution to the greater yield of rice include CGR, RGR and NAR. The variety and growth stage have a significant impact on these growth factors. These growth metrics were significantly impacted by irrigation, K and N in this research. The LAI, CGR, RGR and NAR increased markedly in S condition, at K65 and at N140 over CF, K98 and N210. The term "interactions between water and fertilizers" describes how water and the nutrients added to the soil interact with each other, having a favorable impact on plant growth [24,35,36,37]. The interaction between water and nutrients had a considerable and advantageous impact on the growth indices of the rice plant, according to our findings.Our results corroborated the claims made by Fageria et al. [38] that rice growth can be accelerated by the N and K interaction. The availability of one nutrient affects how well another is absorbed, interactions between nutrients are governed. Since K+ functions as an electrochemical balance for NO3-, applying K can increase plants' ability to absorb N [39]. N availability is affected by the interactions between K+ and NH4+ during exchange [40]. In appropriate soil moisture condition, N and K interaction accelerated growth indices in this research. Nutritionally, K+ and NH4+ have an antagonistic connection; nevertheless, K+ and NO3- acquisition rates are frequently observed to be favorably associated [41], and anadequate K supply can boost amino acid and protein synthesis, stimulate N metabolism, and enhance rice plant growth and production [42].
In this study, the value of RN, RL and RV was higher in S condition than CF which followed the theme that irrigation that conserves water is more successful than flooding irrigation at enhancing root activity and establishing healthy root morphology [43,44]. K has a variety of activities in plants, including controlling the cell cycle [12] and carrying out cell death programs, which both contribute to supporting root development [13] and that’s why in our research at K65 (standard dose) level highest value of root traits was found than at K98 level. Besides, over dose of N in detrimental to root growth which is connected the statement of Britto and Kronzucker [45], that excess N fertilizer might have a detrimental impact on root development due to ammonium toxicity and at the best N fertilization increases root length and diameter [46]. The consequence of an optimal supply of moisture and N enhanced root growth, as previously documented by Mahajan et al. [47]. Despite the fact that water and N interact and have a linked influence on root development [48]. Again, root development and growth of plants can be influenced by the input of N and K fertilizers or the combination of these fertilizers [49,50,51,52]. Combining K and N applications produced a notable beneficial reciprocal effect on crops and was a key strategy for increasing K usage efficiency [53]. According to studies, increased root length and root biomass were related to better N absorption through nutrient interactions like N and K [54,55]. The findings of this study showed that root properties were enhanced by interactions among irrigation, K and N. As a result, we hypothesized that balanced fertilization and optimal irrigation supply might enhance root development, growth indices, and ultimately crop production. This research moved forward our knowledge of the relevance of balanced fertilization and ideal watering for enhanced root characteristics and offers practical advice for optimization of fertilizer management based on the root development response to nutrient supply. In this research, high RP was observed in CF, anaerobic conditions as a result of aerenchyma development (Figure 5), which followed the statement of Lynch et al. [59] that is increased aerenchyma production was a root response to low oxygen levels. The aerenchyma ability to transfer oxygen helps rice roots in submerged soils meet their oxygen requirements. Nutritional imbalance in rhizosphere results in the induction of aerenchymatous tissue [60,61].
There have been several publications on the connections between high yield and the accumulation and translocation of dry materials.The quantity of carbohydrates that plants store before to heading and those that plants create through photosynthesis after heading are the primary determinants of the GY, which is a byproduct of dry matter production [56]. One of the most crucial plant nutrients, N is essential for the creation of biomass and photosynthesis in plants.The primary contributing component to the increase in yield after N incorporation is an increase in panicle numbers [57,58]. Moreover, K, in particular, can enhance chlorophyll, shield the photosynthetic organs from dryness, and enable them to properly perform their function, which will boost the photosynthesis. These factors together have enhanced the output of dry land crops [53]. Therefore, optimum supply of N and K under optimum irrigation can increase TDM, ET and GP.
Rice output is thought to be increased by a synergistic relationship between soil moisture and nitrogenous fertilizer during rice growth [62,63]. According to prior research by Mandal et al.[64], this may be attributed to a constant and optimal supply of N present together with optimal soil moisture in the root zone during the crop growth cycle. Application of K and N enhanced plant K content, GP, and straw production [65]. The yield of grains may be impacted by the interactions between potassium (K) and nitrogen (N), according to studies of Duncan et al. [55]. As per Metho et al.[66], with the combine application of N and K fertilizers, the yield was higher than when the nutrients were added separately. In correlation matrix, the value of RL, RN and RV was significantly correlated with the GY while interacting between K and N. According to prior research, N and K showed significant influence of RL, RV, and RN in relation to GY [51,55,67], which may be occurred due to optimum supply of irrigation that can promote the activity of nutrients.
5. Conclusions
Based on this study, it is found that optimum supply of water and dose of potassium and nitrogen influences root traits, growth indices and yield positively. At S with K65 and N140level the value of RN, RL, RV, LAI TDM, yield attributes and yield were reached at peak than continuous flooding, K98 and N210 level. The highest value of RN, RL and RV were found at 80 DAT then these values were decreased at harvest stage in most cases. There were positive correlation found between root traits and yield except root porosity. It can be concluded that, interactions among irrigation, potassium and nitrogen significantly benefited for rice root traits as well as growth indices and yield.
Author Contributions
Conceptualization: M.S.K., U.K.S., M.R.U., A.K.H. and M.A.H.; methodology, data collection and original data analysis: M.S.K., S.A.K., and U.S.; data presentation, writing: M.S.K, A.H. and E.F.A.; reviewing and editing: U.K.S., M.R.U, M.A.H., E.F.A., G.S. and A.K.C.; funding acquisition: M.R.U and E.F.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Bangladesh Agricultural University Research Systems (BAURES) Project number (2019/15/BAU),Bangladesh. The authors also would like to extend their sincere appreciation to the Researchers Supporting Project Number (RSP2023R134), King Saud University, Riyadh, Saudi Arabia.
Data availability statement
Data sets analyzed during the present study are accessible from the current author on reasonable request.
Acknowledgments
The authors extend their appreciation to BAURES Project number (2019/15/BAU),Bangladesh Agricultural University, Mymensingh, The authors also would like to extend their heartfelt gratitude to the Researchers Supporting Project Number (RSP2023R134), King Saud University, Riyadh, Saudi Arabia.
Conflicts of Interest
The authors state no conflict of interest.
References
- Zhao, C.; Chen, M.; Li, X.; Dai, Q.; Xu, K.; Guo, B.; Hu, Y.; Wang, W.; Huo, Z. Effects of soil types and irrigation modes on rice root morphophysiological traits and grain quality. Agronomy 2021, 11, 120. [Google Scholar] [CrossRef]
- Kreszies, T.; Schreiber, L.; Ranathunge, K. Suberized transport barriers in Arabidopsis, barley and rice roots: From the model plant to crop species [J]. J. Plant Physiol. 2018, 227, 5–83. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.M.; Yang, L.Z.; Yang, Y.X.; Ouyang, Z. Rice root growth and nutrient uptake as influenced by organic manure in continuously and alternately flooded paddy soils. Agric. Water Manag. 2004, 70, 67–81. [Google Scholar] [CrossRef]
- Waines, J.G.; Ehdaie, B. Domestication and crop physiology: Roots of green-revolution wheat. Ann. Bot. 2007, 100, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Gewin, V. Food: An underground revolution. Nature 2010, 466, 552–553. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Wang, L.; Fabrice, M.R.; Tian, Y.; Qi, K.; Chen, Q.; Cao, P.; Wang, P.; Zhang, S.; Wu, J.; Tao, S. Physiological and nutritional responses of pear seedlings to nitrate concentrations. Front. Plant Sci. 2018, 9, 1679. [Google Scholar] [CrossRef] [PubMed]
- Gaudin, A.C.M.; Mcclymont, S.A.; Holmes, B.M.; Lyons, E.; Raizada, M.N. Novel temporal, fine-scale and growth variation phenotypes in roots of adult-stage maize (Zea mays L.) in response to low nitrogen stress. Plant Cell Environ. 2011, 34, 2122–2137. [Google Scholar] [CrossRef]
- Fan, J.B.; Zhang, Y.L.; Turner, D.; Duan, Y.H.; Wang, D.S.; Shen, Q.R. 2010. Root physiological and morphological characteristics of two rice cultivars with different nitrogen-use efficiency. Pedosphere 2010, 20, 446–455. [Google Scholar] [CrossRef]
- Yang, G.Z.; Chu, K.Y.; Tang, H.Y.; Nie, Y.C.; Zhang, X.L. Fertilizer 15N accumulation, recovery and distribution in cotton plant as affected by N rate and split. J. Integr. Agr. 2013, 12, 999–1007. [Google Scholar] [CrossRef]
- White, P.J.; Karley, A.J. Potassium Cell Biology of Metals and Nutrients; Springer: Berlin, Germany, 2010; pp. 199–224. [Google Scholar]
- Oosterhuis, D.; Loka, D.; Kawakami, E.; Pettigrew, W. The physiology of potassium in crop production. Adv. Agron. 2014, 126, 203–234. [Google Scholar] [CrossRef]
- Sano, T.; Becker, D.; Ivashikina, N.; Wegner, L.H.; Zimmermann, U.; Roelfsema, M.R.; Nagata, T.; Hedrich, R. Plant cells must pass a K+ threshold to re-enter the cell cycle. Plant J. 2007, 50, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.; Chin, C.K. Potassium loss is involved in tobacco cell death induced by palmitoleic acid and ceramide. Arch. Biochem. Biophys. 2007, 465, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Hermans, C.; Hammond, J.P.; White, P.J.; Verbruggen, N. How do plants respond to nutrient shortage by biomass allocation? Trends Plant Sci. 2006, 11, 610–617. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Chen, L.; Qu, H.; Lian, J.; Liu, W.; Hu, Y.; Xu, G. Alteration of nutrient allocation and transporter genes expression in rice under N, P, K, and Mg deficiencies. Acta Physiol. Plant 2012, 34, 939–946. [Google Scholar] [CrossRef]
- Moriconi, J.I.; Buet, A.; Simontacchi, M.; Santa-Marıa, G.E. Nearisogenic wheat lines carrying altered function alleles of the Rht-1 genes exhibit differential responses to potassium deprivation. Plant Sci. 2012, 185, 199–207. [Google Scholar] [CrossRef]
- Huang, Y.D.; Zhang, Z.L.; Wei, F.Z.; Li, J.C. Ecophysiological effect of dry cultivated and plastic film-mulched rice planting. Chin. J. Appl. Ecol. 1999, 10, 305–308. [Google Scholar]
- Geng, D.; Chen, P.; Shen, X.; Zhang, Y.; Li, X.; Jiang, L.; Xie, Y.; Niu, C.; Zhang, J.; Huang, X.; Ma, F.; Guan, Q. Mdmyb88 and Mdmyb124 enhance drought tolerance by modulating root vessels and cell walls in apple. Plant Physiol. 2018, 178, 1296–1309. [Google Scholar] [CrossRef]
- Bengough, A.G.; McKenzie, B.M.; Hallet, P.D.; Valentine, T.A. Root elongation, water stress, and mechanical impedance: A review of limiting stresses and beneficial root tip traits. J. Exp. Bot. 2011, 62, 59–68. [Google Scholar] [CrossRef]
- Ali, M.H.; Khatun, M.M.; Mateo, L.G. Influences of various level of water depth on rice growth in rice-fish culture under wetland rice ecosystems. J. Geo. Environ. 2000, 4, 23–30. [Google Scholar]
- Li, S.; Wang, Z.; Malhi, S.S.; Li, S.; Gao, Y.; Tian, X. Nutrient and Water Management Effects on Crop Production, and Nutrient and Water Use Efficiency in Dryland Areas of China. Adv. Agron. 2009, 102, 223–265. [Google Scholar] [CrossRef]
- Lower, S.S.; Orians, C.M. Soil nutrients and water availability interact to influence willow growth and chemistry but not leaf beetle performance. Entomol. Exp. Appl. 2003, 107, 69–79. [Google Scholar] [CrossRef]
- Wang, X.B.; Zhou, B.Y.; Sun, X.F.; Yue, Y.; Ma, W.; Zhao, M. Soil tillage management affects maize grain yield by regulating spatial distribution coordination of roots, soil moisture and nitrogen status. PLoS ONE 2015, 10, e0129231. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wang, C.S.; Dell, B.; Zhao, Z.G.; Guo, J.J.; Xu, D.P.; Zeng, J. Growth and nutrient dynamics of Betulaalnoides seedlings under exponential fertilization. J. For. Res. 2018, 29, 111–119. [Google Scholar] [CrossRef]
- Gregory, P.J.; Simmonds, L.P.; Warren, G.P. Interactions between plant nutrients, water and carbon dioxide as factors limiting crop yields. Phil. Trans. R. Soc. Lond. B. 1997, 352, 987–996. [Google Scholar] [CrossRef]
- UNDP and FAO (United Nations Development Program and Food and Agriculture Organization). Land Resources Appraisal of Bangladesh for Agricultural Development. Report 2. Agro-Ecological Region of Bangladesh; United Nations Development Program andFood and Agricultural Organization: Dhaka, Bangladesh, 1988; pp. 212–221. [Google Scholar]
- FRG. Fertilizer Recommendation Guide; Bangladesh Agricultural Research Council (BARC): Dhaka, Bangladesh, 2018; p. 274. [Google Scholar]
- Bridgit, A.J.; Potty, N.N. Influence of root characters on rice productivity in iron soils of Kerala. Int. Rice Res. News 2002, 27, 45–46. [Google Scholar]
- Jensen, C.R.; Luxmoore, R.J.; Van Gundy, S.D.; Stolzy, L.H. Root air space measurements by a pycnometer method. Agron. J. 1969, 61, 474–475. [Google Scholar] [CrossRef]
- Radford, P.J. Growth analysis formulae-their use and abuse. Crop Sci. 1967, 7, 171–175. [Google Scholar] [CrossRef]
- Hunt, R. Plant Growth Analysis. Studies in Biology; Edward Arnold: London, UK, 1978. [Google Scholar]
- Yoshida, S.; Foron, D.A.; Cock, J.H. Laboratory Manual for Physiological Studies of Rice; International Rice Research Institute: LosBaños, Philippines, 1971; p. 70. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation Statistical Computation: Vienna, Austria, 2013; Available online: http://www.R-project.org/ (accessed on 20march 2023).
- Fageria, N.K.; Baligar, V.C.; Clark, R.B. Physiology of crop production; Haworth Press Inc., 2006; pp. 1–356. [Google Scholar]
- Elmi, A.A.; Madramootoo, C.; Egeh, M.; Hamel, C. Water and fertilizer nitrogen management to minimize nitrate pollution from a cropped soil in southwestern Quebec, Canada. Water Air Soil Pollut. 2004, 151, 117–134. [Google Scholar] [CrossRef]
- Guttieri, M.J.; R. McLean, J.C.; Stark, Souza, E. Managing irrigation and nitrogen fertility of hard spring wheats for optimum and noodle quality. Crop Sci. 2005, 45, 2049–2059. [Google Scholar] [CrossRef]
- Islam, S.M.M.; Gaihre, Y.K.; Biswas, J.C.; Jahan, M.S.; Singh, U.; Adhikary, S.K.; Satter, M.A.; Saleque, M.A. Different nitrogen rates and methods of application for dry season rice cultivation with alternate wetting and drying irrigation: Fate of nitrogen and grain yield. Agr. Water Mgt. 2018, 196, 144–153. [Google Scholar] [CrossRef]
- Fageria, N.K.; Moreira, A.; Moraes, L.A.C.; Moraes, M.F. Root Growth, Nutrient Uptake, and Nutrient-Use Efficiency by Roots of Tropical Legume Cover Crops as Influenced by Phosphorus Fertilization. Commun. Soil Sci. Plant Anal. 2014, 45, 555–569. [Google Scholar] [CrossRef]
- Blevins, D.G.; Barnett, N.M.; Frost, W.B. Role of Potassium and Malate in Nitrate Uptake and Translocation by Wheat Seedlings. Plant Physiol. 1978, 62, 784–788. [Google Scholar] [CrossRef]
- Miller, A.J.; Cramer, M.D. Root Nitrogen Acquisition and Assimilation. In Root Physiology: From Gene to Function; Plant, Ecophysiology, Lambers, H., Colmer, T.D., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 1–36. [Google Scholar] [CrossRef]
- Coskun, D.; Britto, D.T.; Kronzucker, H.J. The nitrogen–potassium intersection: Membranes, metabolism, and mechanism. Plant Cell Environ. 2016, 10, 2029–2041. [Google Scholar] [CrossRef]
- Ruiz, J.; Romero, L. Relationship between potassium fertilisation and nitrate assimilation in leaves and fruits of cucumber (Cucumissativus) plants. Ann. Appl. Biol. 2002, 140, 241–245. [Google Scholar] [CrossRef]
- Alou, I.N.; Steyn, J.M.; Annandale, J.G.; van der Laan, M. Growth, phenological, and yield response of upland rice (Oryza sativa L. cv. Nerica 4 R ) to water stress during different growth stages. Agr. Water. Manage. 2018, 198, 39–52. [Google Scholar] [CrossRef]
- Song, T.; Xu, F.; Yuan, W.; Zhang, Y.; Liu, T.; Chen, M.; Hu, Q.; Tian, Y.; Xu, W.; Zhanh, J. Comparison on physiological adaptation and phosphorus use efficiency of upland rice and lowland rice under alternate wetting and drying irrigation. Plant Growth Regul. 2018, 86, 195–210. [Google Scholar] [CrossRef]
- Britto, D.T.; Kronzucker, H.J. NH4+ toxicity in higher plants: A critical review. J Plant Physiol. 2002, 159, 567–584. [Google Scholar] [CrossRef]
- Costa, C.; Dwyer, L.M.; Zhou, X.; Dutilleul, P.; Hamel, C.; Reid, L.M.; Smith, D.L. Root morphology of contrasting maize genotypes. Agronomy Journal 2002, 94, 96–101. [Google Scholar] [CrossRef]
- Mahajan, G.; Chauhan, B.S.; Timsina, J.; Singh, P.P.; Singh, K. Crop Performance and Water and Nitrogen-Use Efficiencies in Dry-Seeded Rice in Response to Irrigation and Fertilizer Amounts in Northwest India. Field Crops Res. 2012, 134, 59–70. [Google Scholar] [CrossRef]
- Reis, A. Aerobic rice system improves water productivity, nitrogen recovery and crop performance in brazilian weathered lowland soil. Field Crop Res. 2018, 218, 59–68. [Google Scholar] [CrossRef]
- McMurtrie, R.E.; Iversen, C.M.; Dewar, R.C.; Medlyn, B.E.; Näsholm, T.; Pepper, D.A.; Norby, R.J. Plant Root Distributions and Nitrogen Uptake Predicted by a Hypothesis of Optimal Root Foraging. Ecol. Evol. 2012, 2, 1235–1250. [Google Scholar] [CrossRef]
- Teng, W.; Deng, Y.; Chen, X.; Xu, X.; Chen, R.; Lv, Y.; Zhao, Y.; Zhao, X.; He, X.; Li, B.; et al. Characterization of root response to phosphorus supply from morphology to gene analysis in field-grown wheat. J. Exp. Bot. 2013, 64, 1403–1411. [Google Scholar] [CrossRef]
- Fageria, N.K.; Oliveira, J.P. Nitrogen, Phosphorus and Potassium Interactions in Upland Rice. J. Plant Nutr. 2014, 37, 1586–1600. [Google Scholar] [CrossRef]
- Deng, Y.; Teng, W.; Tong, Y.; Chen, X.; Zou, C. Phosphorus Efficiency Mechanisms of Two Wheat Cultivars as Affected by a Range of Phosphorus Levels in the Field. Front. Plant Sci. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Li, X. , Lu, J.; Wu, L.; Chen, F. The difference of potassium dynamics between yellowish red soil and yellow cinnamon soil under rapeseed (Brassica napus L.)–rice (Oryza sativa L.) rotation. Plant Soil 2009, 320, 141–151. [Google Scholar] [CrossRef]
- Duan, Y.; Shi, X.; Li, S.; Sun, X.; He, X. Nitrogen Use Efficiency as Affected by Phosphorus and Potassium in Long-Term Rice and Wheat Experiments. J. Integr. Agric. 2014, 13, 588–596. [Google Scholar] [CrossRef]
- Duncan, E.G.; O’Sullivan, C.A.; Roper, M.M.; Biggs, J.S.; Peoples, M.B. Influence of Co-Application of Nitrogen with Phosphorus, Potassium and Sulphur on the Apparent Efficiency of Nitrogen Fertiliser Use, Grain Yield and Protein Content of Wheat: Review. Field Crops Res. 2018, 226, 56–65. [Google Scholar] [CrossRef]
- Weng, J.H.; Takeda, T.; Agata, W.; Hakoyama, S. Studies on dry matter and grainproduction of rice plants: I. Influence of the reserved carbohydrate until heading stage and the assimilation products during the ripening period on grain production. Japanese J. Crop Sci. 1982, 51, 500–509. [Google Scholar] [CrossRef]
- Bindra, A.D.; Kalia, B.D.; Kumar, S. 2000. Effect of nitrogen levels and dates of transplanting on growth, yield and yield attributes of scented rice. Adv. Agric. Res. 2000, 10, 45–48. [Google Scholar]
- Laroo, N.; Shivay, Y.S. 2011. Effect of nitrogen and sulphur levels on growth and productivity of scented rice. Current Advances in Agric. Sci. 2011, 3, 45–48. [Google Scholar]
- Lynch, J.; Marschner, P.; Rengel, Z. Effect of internal and external factors on root growth and development. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: London, UK, 2012; pp. 331–346. [Google Scholar] [CrossRef]
- Yamauchi, Y.; Colmer, T.D.; Pedersen, O.; Nakazono, M. Regulation of root traits for internal aeration and tolerance to soil waterlogging-flooding stress. J. Plant Physiol. 2018, 176, 1118–1130. [Google Scholar] [CrossRef]
- Lynch, J.P. Root phenotypes for improved nutrient capture: An underexploited opportunity for global agriculture. New Phytol. 2019, 223, 548–564. [Google Scholar] [CrossRef]
- Liu, L.; Chen, T.; Wang, Z.; Zhang, H.; Yang, J.; Zhang, J. Combination of site-specific nitrogen management and alternate wetting and drying irrigation increases grain yield and nitrogen and water use efficiency in super rice. Field Crops Res. 2013, 154, 226–235. [Google Scholar] [CrossRef]
- Yang, J.C. Approaches to achieve high grain yield and high resource use efficiency in rice. Front AgricSciEng. 2015, 2, 115–123. [Google Scholar] [CrossRef]
- Mandal, K.G.; Kundu, D.K.; Thakur, A.K.; Kannan, K.; Brahmanand, P.S.; Kumar, A. Aerobic Rice Response to Irrigation Regimes and Fertilizer Nitrogen Rates. J Food Agric Environ. 2013, 11, 1148–1153. [Google Scholar]
- Rajeev, K.; Sandeep, K.; Anil, K.; Harnam, S.; K. Sanjeev, K. 2013. Effect of top dressing nitrogen and potassium on yield and yield component of rice (Oryza sativa L). Agriways 2013, 1, 90–94. [Google Scholar]
- Metho, L.A.; Hammes, P.S.; de Beer, J.M.; Groeneveld, H.T. Interaction between Cultivar and Soil Fertility on Grain Yield, Yield Components and Grain Nitrogen Content of Wheat. S. Afr. J. Plant Soil 1997, 14, 158–164. [Google Scholar] [CrossRef]
- Epie, K.E.; Maral, E. Shoot and Root Biomass, Phosphorus and Nitrogen Uptake of Spring Wheat Grown in Low Phosphorus and Moisture Content Conditions in a Pot Experiment. J. Plant Nutr. 2018, 41, 2273–2280. [Google Scholar] [CrossRef]
Figure 1.
Dynamics root morphological traits of Binadhan-10 under two irrigation, potassium and nitrogen treatments from 20 DAT to harvest stage.I1: Saturation I2: Continuous flooding K65: 65 kg K ha−1 K98: 98 kg K ha−1N140: 140 kg N ha−1 N210: 210 kg N ha−1; (A), (B),(C), (D) and (E) represent root number; (F), (G), (H), (I) and (J) represent root length; (K), (L), (M), (N) and (O) represent root porosity; (P), (Q), (R), (S) and (T) represent root volume.
Figure 1.
Dynamics root morphological traits of Binadhan-10 under two irrigation, potassium and nitrogen treatments from 20 DAT to harvest stage.I1: Saturation I2: Continuous flooding K65: 65 kg K ha−1 K98: 98 kg K ha−1N140: 140 kg N ha−1 N210: 210 kg N ha−1; (A), (B),(C), (D) and (E) represent root number; (F), (G), (H), (I) and (J) represent root length; (K), (L), (M), (N) and (O) represent root porosity; (P), (Q), (R), (S) and (T) represent root volume.
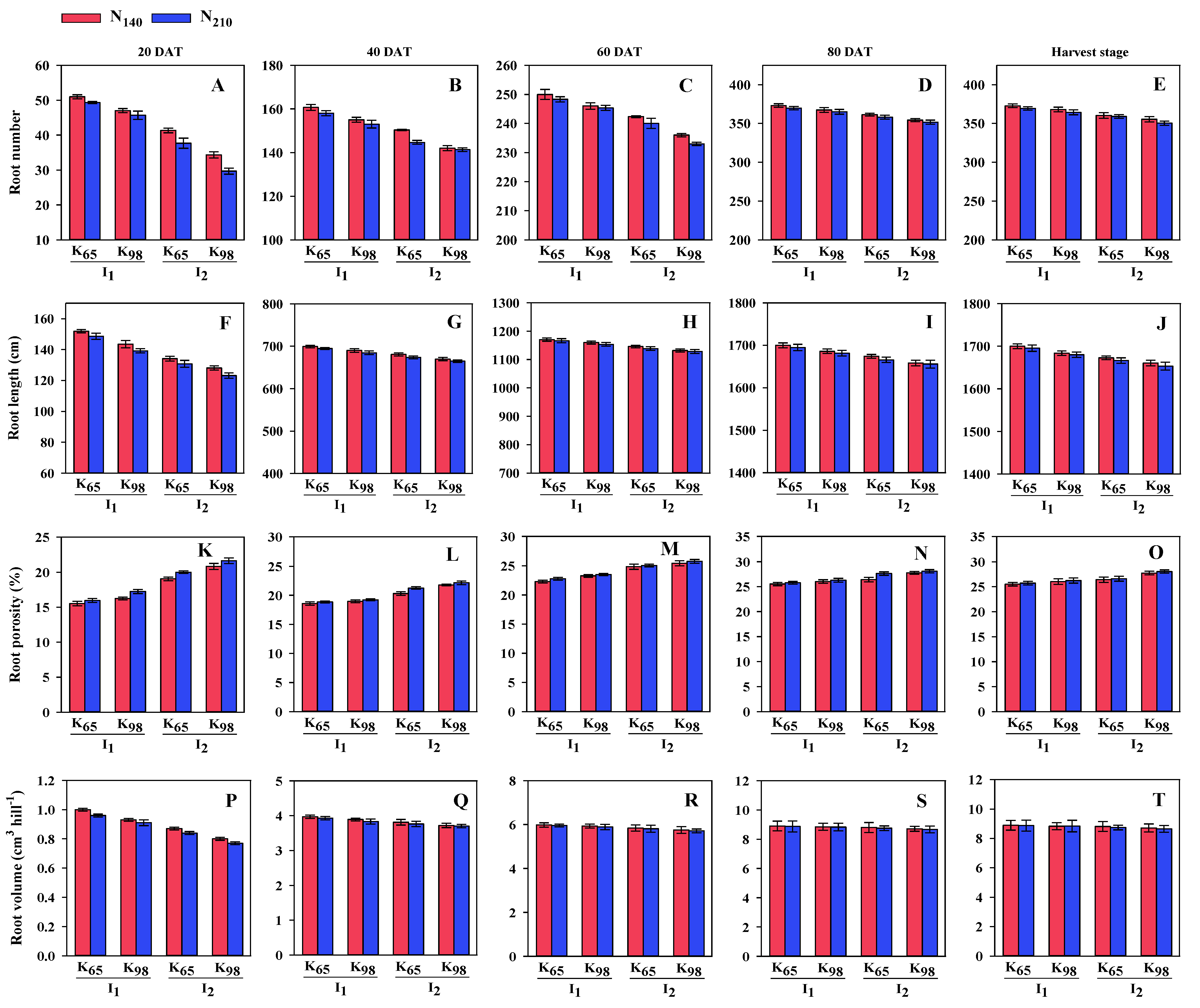
Figure 2.
Leaf area index (LAI) and total dry matter (TDM) of Bina dhan-10under two irrigation, potassium and nitrogen treatments from 20 DAT to 80 DAT. I1: Saturation I2: Continuous floodingK65: 65 kg ha−1 K98: 98 kg ha−1N140: 140 kg ha−1 N210: 210 kg ha−1; (A), (B), (C) and (D) represent LAI; (E), (F), (G) and (H) represent TDM.
Figure 2.
Leaf area index (LAI) and total dry matter (TDM) of Bina dhan-10under two irrigation, potassium and nitrogen treatments from 20 DAT to 80 DAT. I1: Saturation I2: Continuous floodingK65: 65 kg ha−1 K98: 98 kg ha−1N140: 140 kg ha−1 N210: 210 kg ha−1; (A), (B), (C) and (D) represent LAI; (E), (F), (G) and (H) represent TDM.
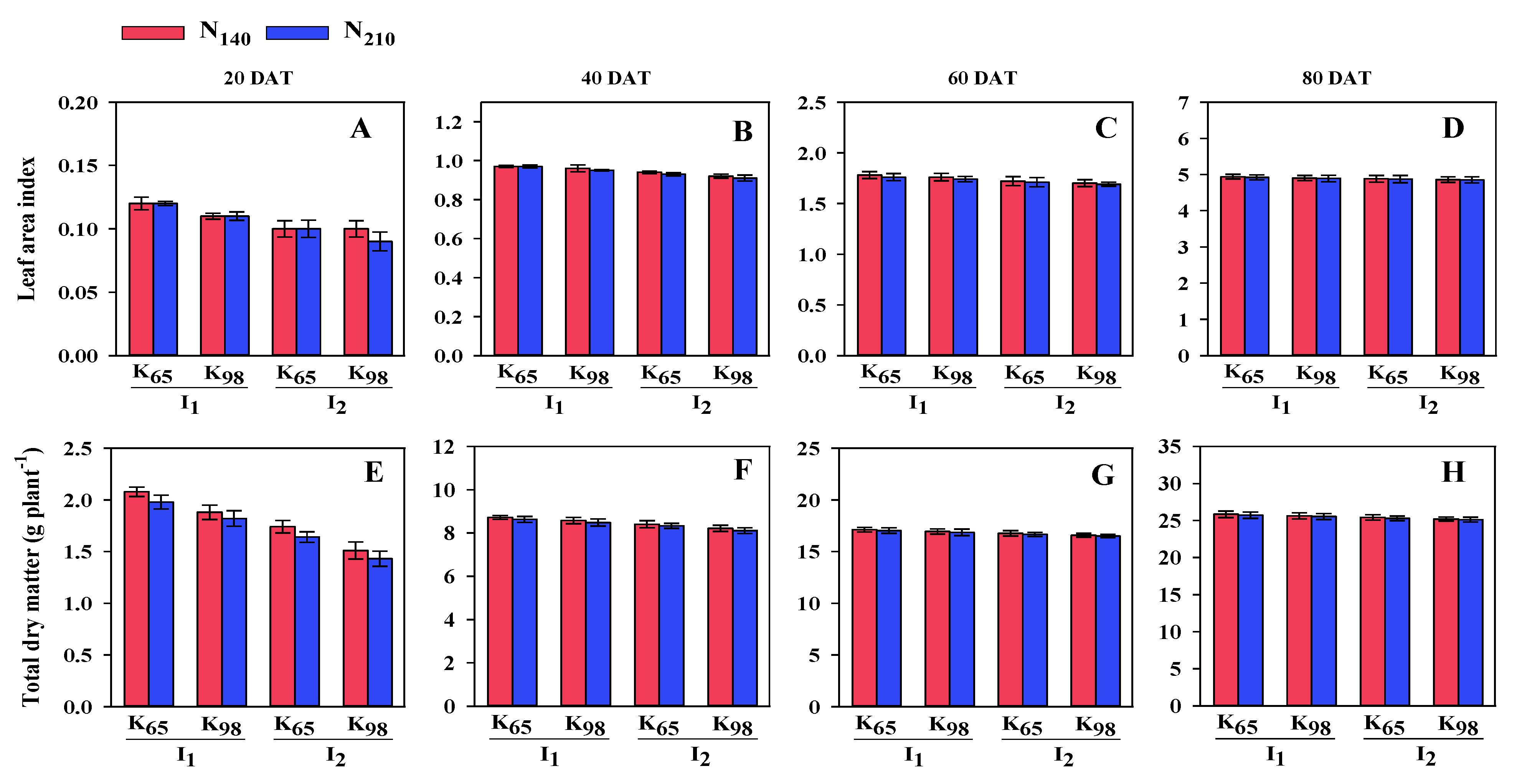
Figure 3.
CGR, RGR and NAR of Binadhan-10 under two irrigation, potassium and nitrogen treatments at 20-40 (1st), 40-60 DAT (2nd) and 60-80 DAT (3rd). I1: Saturation I2: Continuous flooding K65: 65 kg ha−1 K8: 98 kg ha−1N140: 140 kg ha−1 N210: 210 kg ha−1; (A), (B) and (C) represent CGR; (D), (E) and (F) represents RGR; (G), (H) and (I) represent NAR.
Figure 3.
CGR, RGR and NAR of Binadhan-10 under two irrigation, potassium and nitrogen treatments at 20-40 (1st), 40-60 DAT (2nd) and 60-80 DAT (3rd). I1: Saturation I2: Continuous flooding K65: 65 kg ha−1 K8: 98 kg ha−1N140: 140 kg ha−1 N210: 210 kg ha−1; (A), (B) and (C) represent CGR; (D), (E) and (F) represents RGR; (G), (H) and (I) represent NAR.
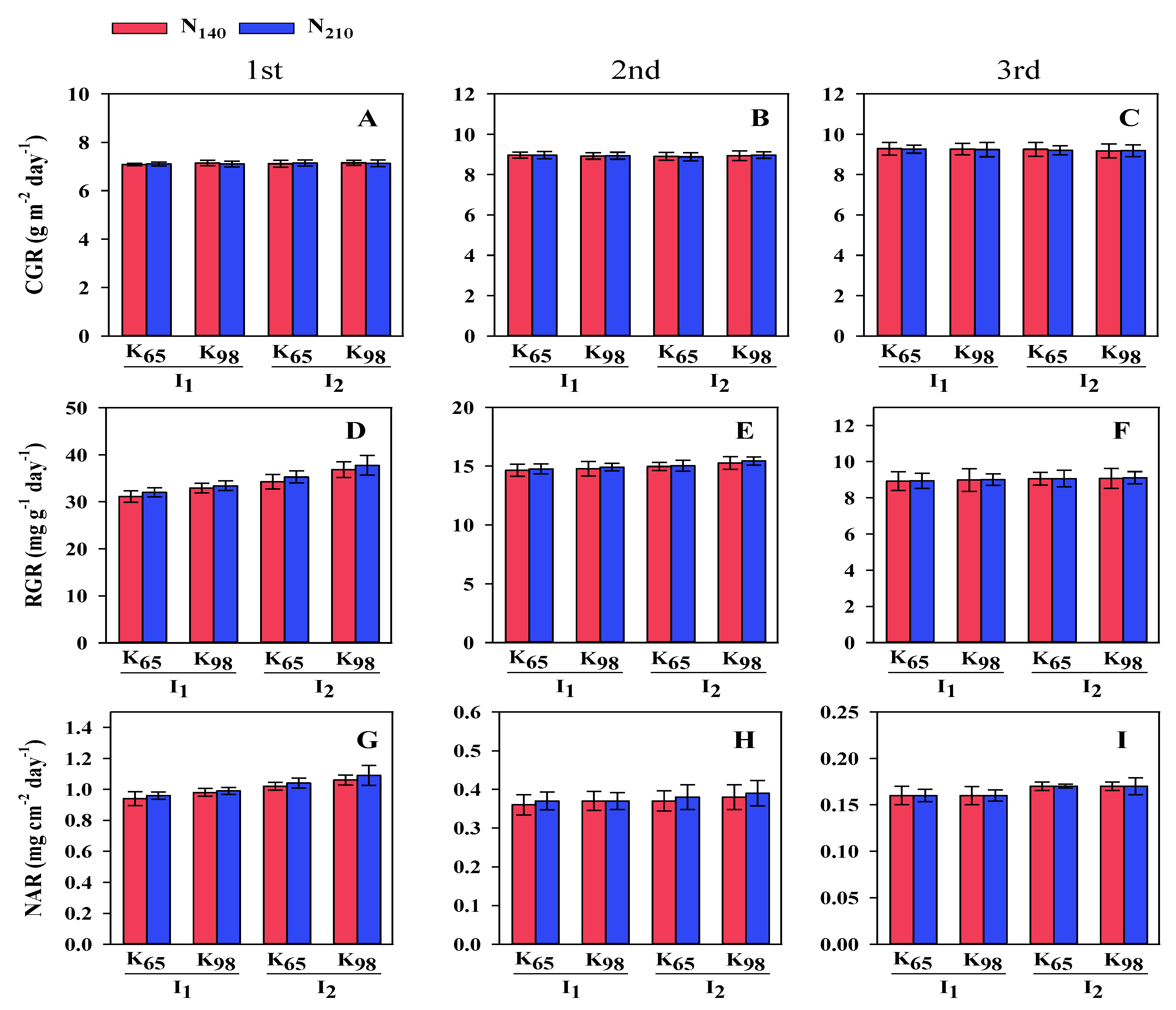
Figure 4.
Yield and yield contributing parameters ofBina dhan-10 under two nitrogen, potassium and irrigation treatments. I1: Saturation I2: Continuous floodingK65: 65 kg ha−1 K8: 98 kg ha−1N140: 140 kg ha−1 N210: 210 kg ha−1.
Figure 4.
Yield and yield contributing parameters ofBina dhan-10 under two nitrogen, potassium and irrigation treatments. I1: Saturation I2: Continuous floodingK65: 65 kg ha−1 K8: 98 kg ha−1N140: 140 kg ha−1 N210: 210 kg ha−1.
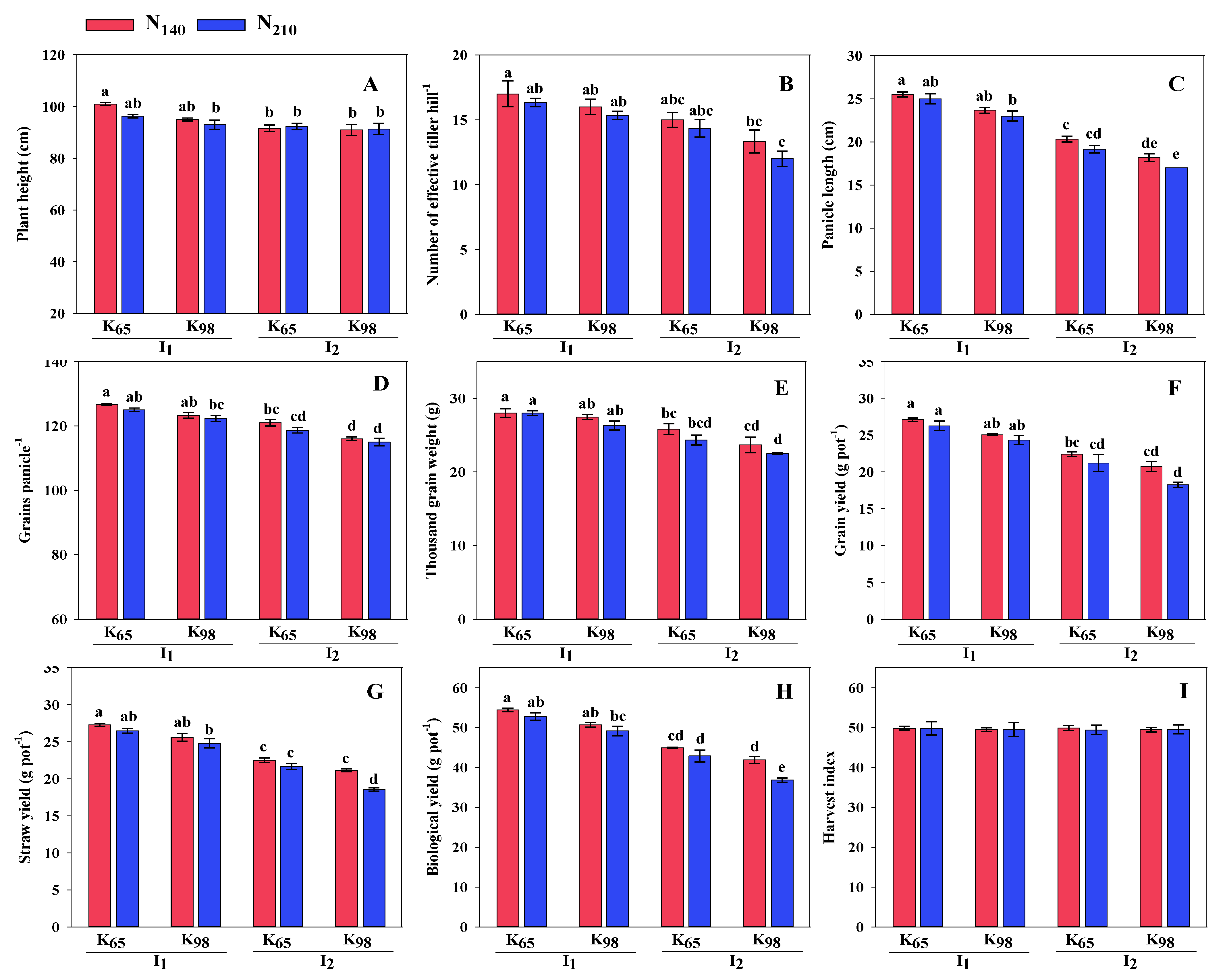
Figure 5.
Correlation matrix and heatmap of the root traits, growth parameters, yield attributes and yield . The positive and negative correlations were indicated by blue and red ellipses. Thegreater coefficient is reflected by higher color intensity. *, ** and *** indicates level of significance at 5, 1 and 0.1% level of probability.Traitsdetails: PH—plant height; ET—number of effective tillers hill-1; RN—root number; RL—root length; RP—root porosity; RV—root volume; PL—panicle length; GP—grains per panicle; TGW—thousand grain weight; GY—grain yield; SY—straw yield; BY—biological yield; HI—harvest index.
Figure 5.
Correlation matrix and heatmap of the root traits, growth parameters, yield attributes and yield . The positive and negative correlations were indicated by blue and red ellipses. Thegreater coefficient is reflected by higher color intensity. *, ** and *** indicates level of significance at 5, 1 and 0.1% level of probability.Traitsdetails: PH—plant height; ET—number of effective tillers hill-1; RN—root number; RL—root length; RP—root porosity; RV—root volume; PL—panicle length; GP—grains per panicle; TGW—thousand grain weight; GY—grain yield; SY—straw yield; BY—biological yield; HI—harvest index.
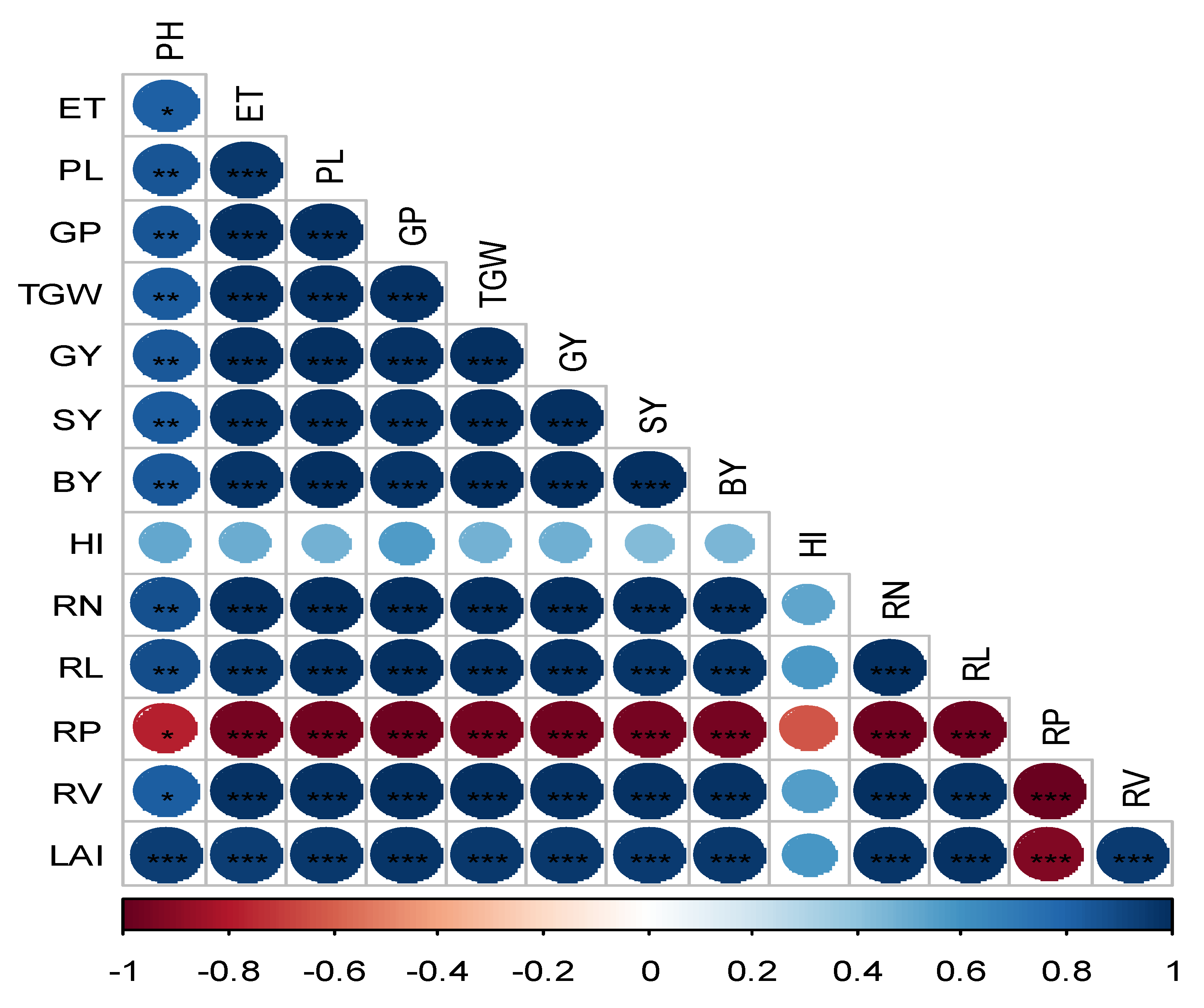
Figure 6.
The differences root crosses-section of Binadhan-10 under different combination of treatments at 80 DAT. Here, N140: 140 kg ha-1, N210: 210 kg ha-1, K65: 65 kg ha-1, K98: 96 kg ha-1; I1: Saturation I2: Continuous flooding.
Figure 6.
The differences root crosses-section of Binadhan-10 under different combination of treatments at 80 DAT. Here, N140: 140 kg ha-1, N210: 210 kg ha-1, K65: 65 kg ha-1, K98: 96 kg ha-1; I1: Saturation I2: Continuous flooding.


Table 1.
Yield components of Binadhan-10 under two irrigation, potassium and nitrogen treatments.
| Irrigation (I) | PH (cm) | ET (no.) | PL (cm) | GP (no.) | TGW (g) | GY (g pot-1) |
SY (g pot-1) |
HI (%) |
|---|---|---|---|---|---|---|---|---|
| I1 | 96.33 a | 16.17 a | 24.29 a | 124.33 a | 26.90 a | 25.70 a | 26.05 a | 49.66 |
| I2 | 91.58 b | 13.67 b | 18.67 b | 117.67 b | 22.81 b | 20.65 b | 20.98 b | 49.57 |
| CV (%) | 3.24 | 9.07 | 6.15 | 2.01 | 6.02 | 7.07 | 5.94 | 1.75 |
| Potassium (K) | ||||||||
| K65 | 95.33 a | 15.67a | 22.50 a | 122.83 a | 25.87 a | 24.25 a | 24.49 a | 49.73 |
| K98 | 92.58 b | 14.17 b | 20.46 b | 119.17 b | 23.85 b | 22.10 b | 22.54 b | 49.51 |
| CV (%) | 3.89 | 11.46 | 14.15 | 3.14 | 9.59 | 12.42 | 11.95 | 1.74 |
| Nitrogen (N) | ||||||||
| N140 | 94.67 a | 15.33 a | 21.92 a | 121.75 a | 121.75 a | 23.83 a | 24.14 a | 49.67 |
| N210 | 93.25 b | 14.50 b | 21.04 b | 120.25 b | 120.25 b | 22.52 b | 22.89 b | 49.56 |
| CV (%) | 4.10 | 12.27 | 14.84 | 3.46 | 10.33 | 13.05 | 12.41 | 1.75 |
| ANOVA | ||||||||
| I | ** | ** | ** | ** | ** | ** | ** | NS |
| K | * | ** | ** | ** | ** | ** | ** | NS |
| N | * | * | ** | * | * | ** | ** | NS |
Notes: Within every column, means indicated by the identical letters were not substantially dissimilar. **,* and NS denote significance at the 1%, 5% levels and non-significance, respectively, depending on the ANOVA.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



