Submitted:
24 February 2026
Posted:
25 February 2026
You are already at the latest version
Abstract
This study aimed to isolate and characterize nitrogen-fixing bacteria from the maize rhizosphere and evaluate their plant growth-promoting potential to reduce reliance on synthetic fertilizers and enhance soil fertility. Nitrogen-free selective media were used for bacterial isolation, followed by detection of the nifH gene and nitrogenase activity. Phylogenetic identification was conducted via 16S rRNA sequencing. Growth-promoting traits, stress tolerance, and pot-based plant inoculation effects were assessed. Genetic modification of strain GN8811 was performed to improve nitrogen fixation and growth promotion. Seven isolates possessed the nifH gene and nitrogenase activity, including Azotobacter chroococcum GN2001, A. vinelandii GN1202, Azospirillum brasilense GN1004, Kosakonia sacchari GN2003, Klebsiella michiganensis GN8799 and GN8801, and K. quasivariicola GN8811. Furthermore, GN8801 and GN2001 exhibited phosphate solubilization and iron chelation, while GN1004 and GN8811 showed strong IAA production and potassium solubilization. Additionally, GN2003 and GN8811 tolerated high salinity and variable pH. Maize inoculated with GN8811 showed biomass and root enhancement comparable to nitrogen-fertilized controls. The genetically modified GN8811 strain (ΔnifL::nifA) exhibited further improvement in ni-trogen fixation and plant growth, maintaining performance even under high nitrogen conditions. Diverse nitrogen-fixing bacteria were identified from the maize rhizo-sphere, possessing multiple growth-promoting functions and stress tolerance. K. quasivariicola GN8811 demonstrated the best performance, and its genetic enhancement further improved nitrogen fixation efficiency. These findings highlight the potential of combining microbial screening with genetic engineering to develop efficient bioinocu-lants for sustainable maize cultivation. Biological nitrogen fixation by plant-associated bacteria offers a promising route to reduce synthetic nitrogen fertilizer inputs in cere-al-based agroecosystems, yet its reliability is often constrained by environmental stress and nitrogen repression. In this study, we combined systematic isolation of native maize rhizosphere diazotrophs with targeted regulatory engineering of the NifL–NifA system to generate a high-performance nitrogen-fixing strain capable of promoting maize growth even under nitrogen-replete conditions. Our results demonstrate that precise genetic rewiring of indigenous plant-associated bacteria can substantially en-hance nitrogen fixation efficiency and plant growth promotion, highlighting a viable strategy for developing next-generation biofertilizers to support sustainable maize production.
Keywords:
rhizosphere
; maize
; nitrogen-fixing bacteria
; environmental tolerance
; growth promotion
; genetic modifications
1. Introduction
Global food demand is projected to rise sharply as the world population approaches 10 billion, driving intensive agricultural production that relies heavily on synthetic fertilizers to sustain high yields [1]. This input-intensive model consumes large amounts of fossil energy and finite mineral resources, while low nutrient-use efficiency leads to substantial losses of nitrogen (N) to the environment [2]. Excessive N fertilization accelerates soil acidification, salinization, and organic matter depletion, and it contributes to water eutrophication, greenhouse gas emissions, and biodiversity decline [1,2,3,4]. These ecological and soil degradation problems threaten long-term productivity and conflict with global goals for climate-smart and sustainable agriculture [5,6]. Biological nitrogen fixation (BNF) mediated by nitrogen-fixing bacteria offers a promising strategy to partially substitute synthetic N fertilizers, reduce energy consumption and N losses, and enhance soil health, thereby supporting the green transformation of agroecosystems [4,7,8].
Nitrogen-fixing bacteria used as biofertilizers encompass several major groups, including symbiotic rhizobia, associative and endophytic diazotrophs (e.g., Azospirillum, Herbaspirillum, Gluconacetobacter), and free-living soil bacteria such as Azotobacter and some Pseudomonas and Enterobacter species [9,10,11,12,13]. Numerous field and greenhouse studies have shown that inoculation with these nitrogen-fixing bacteria can increase N acquisition, stimulate root growth, and improve yields across diverse cropping systems, while allowing partial reduction of mineral N inputs [14,15,16]. In maize (Zea mays L.), which does not form classical nodules, increasing attention has been devoted to exploiting free-living, associative and endophytic N-fixing bacteria as biofertilizers [17]. Diazotrophs such as Azospirillum spp., Herbaspirillum spp., and plant-associated Enterobacteriaceae have been shown to colonize maize rhizosphere, root mucilage, and internal tissues, contributing fixed N and emitting phytohormones that enhance root architecture and nutrient uptake [16,18,19,20]. Field trials in different agroecological zones report that maize inoculated with selected N-fixing strains can achieve comparable or higher grain yield with over 30% N input, increase biomass and grain protein content [16,21,22]. Nevertheless, considerable variability among maize genotypes, soil types, and climatic regimes often leads to inconsistent responses [23,24,25]. The effectiveness of nitrogen-fixing bacterial inoculants is strongly modulated by soil factors such as drought, salinity, and pH, which subsequently limit crop growth, productivity, and quality [23,26]. In detail, salinity and osmotic stress can impair membrane integrity, enzyme function, and colonization ability, thereby decreasing BNF and plant benefits, especially in arid and coastal regions [27]. Soil pH strongly affects diazotroph survival and nutrient availability, acidification due to excessive N fertilization often suppresses sensitive taxa and alters community composition [27,28]. Drought limits carbon flow from plants to the rhizosphere, decreases bacterial activity and reshapes bacterial community [29,30]. Critically, high levels of inorganic N in soil inhibit nitrogenase expression and activity through regulatory networks, leading to downregulation of nif genes and reduced BNF [31]. These interacting constraints explain why promising strains often perform inconsistently outside controlled conditions.
To enhance the stability and reliability of N-fixing biofertilizers, researchers are pursuing complementary strategies that integrate microbial ecology, strain selection, and genetic engineering. On the one hand, intensive screening programs and omics-guided approaches are being used to identify robust diazotrophic strains with enhanced tolerance to heat, salinity, acidity, and drought, as well as superior root colonization and plant-growth-promoting traits [32]. Consortia combining N-fixing bacteria with other plant growth-promoting rhizobacteria or mycorrhizal fungi are also being developed to increase functional redundancy and resilience under fluctuating field conditions [33,34,35]. On the other hand, molecular engineering of regulatory circuits controlling BNF aims to partially uncouple nitrogenase expression from classical ammonium- or nitrate-mediated repression [36,37]. In particular, targeted modification of the NifL–NifA regulatory system, including alterations in nifL to relieve its inhibitory control over nifA under high N, has been proposed and tested to sustain nitrogenase activity in the presence of elevated inorganic N while minimizing metabolic burden and maintaining redox balance [38,39]. These advances, together with improved formulation technologies and maize genotype–microbe matching, hold considerable promise for stabilizing the contribution of N-fixing bacteria to N supply and for enabling more predictable reductions in synthetic N fertilizer use in sustainable maize production systems [40,41].
The current study aims to systematically isolate and characterize beneficial nitrogen-fixing bacteria from the maize rhizosphere, focusing on their plant growth-promoting (PGP) capabilities, environmental resilience and stable nitrogen fixation capabilities even in the presence of higher environmental nitrogen concentrations. Our findings from this study not only identify promising indigenous nitrogen-fixing bacteria with multiple plant growth-promoting traits from the maize rhizosphere but also underscore the potential of targeted genetic modifications to optimize microbial inoculants for sustainable agriculture. This research highlights the importance of exploring microbial diversity in local agricultural ecosystems and harnessing biotechnological tools to maximize the benefits of these green helpers.
2. Material and Methods
2.1. Soil Sample Collection
Maize rhizosphere soil samples were collected from the root zone (0-20 cm depth) of healthy maize plants in an agricultural field located in Gansu province (a severely arid region in China) during the flowering stage. Soil adhering tightly to the roots after gentle shaking was considered rhizosphere soil. The soil samples were placed in sterile plastic bags, stored on ice, and transported to the laboratory for immediate processing.
2.2. Isolation and Screening of Nitrogen-Fixing Bacteria
The isolation of nitrogen-fixing bacteria was performed using a three-round enrichment culture method on nitrogen-free media. Specifically, 10 g of rhizosphere soil was added to 90 mL of sterile Ashby’s nitrogen-free broth in a 250 mL conical flask. The culture was incubated at 30 °C with shaking at 180 rpm for 7 days. After the first round of incubation, 1 mL of the culture was transferred to fresh Ashby’s broth and incubated under the same conditions for another 7 days. This subculturing process was repeated for a third round. After the third enrichment, serial dilutions of the culture were prepared and spread onto Ashby’s nitrogen-free agar plates. The plates were incubated at 30 °C for 3-5 days. Colonies exhibiting distinct morphologies and mucoid characteristics, typical of nitrogen-fixing bacteria like Azotobacter, were selected and purified by repeated streaking on fresh Ashby’s agar plates. All isolates were stored in 20% glycerol at -80 °C for subsequent studies.
2.3. Molecular Identification and Phylogenetic Analysis
Genomic DNA from the seven isolates was extracted using a bacterial genomic DNA extraction kit. The 16S rRNA gene was amplified via PCR using universal primers 27F (5’-AGAGTTTGATCMTGGCTCAG-3’) and 1492R (5’-GGTTACCTTGTTACGACTT-3’)[42]. A phylogenetic tree was constructed using the Maximum Likelihood method in MEGA X software with bootstrap analysis based on 1000 replications [43].
2.4. Detection of the Nifh Gene and Assay of Nitrogenase Activity
The presence of the nifH gene, a key marker for nitrogen fixation, was detected by PCR amplification using specific primers Nh21F (5’-GCIWTYTAYGGNAARGG-3’) and Cy55Nh428R (5’-CCRCCRCANACMACGTC-3’) [44,45]. The acetylene reduction assay (ARA) was employed to measure nitrogenase activity [46]. Briefly, bacterial strains were cultured in nitrogen-free Ashby’s broth to the logarithmic growth phase. Then, 10 mL of each bacterial suspension was transferred to a 50 mL glass vial, sealed with a rubber stopper, and 10% of the headspace air was replaced with acetylene (C2H2). The vials were incubated at 30 °C for 2 hours. Gas samples (200 µL) from the headspace were analyzed using a gas chromatograph equipped with a flame ionization detector to quantify the amount of ethylene (C2H4) produced. Nitrogenase activity was expressed as nmol C2H4 produced per mg of bacterial protein per hour.
2.5. In Vitro Evaluation of Plant Growth-Promoting (pgp) Traits
The plant growth-promoting traits of the bacterial isolates were qualitatively evaluated using standard plate-based assays. The value of OD600 of all tested trains were adjusted to 0.05. IAA production was assessed by spot inoculating bacterial strains on Ashby’s agar supplemented with 5 mM L-tryptophan and incubating at 30 °C for 72 h. The appearance of a pink-to-red halo after treatment with Salkowski’s reagent was considered indicative of IAA production [47]. Phosphate-solubilizing ability was determined on Pikovskaya’s agar containing insoluble tricalcium phosphate. Plates were incubated at 30 °C for 5 days, and the formation of a clear halo around the colonies indicated phosphate solubilization [48]. Siderophore production was evaluated using Chrome Azurol S (CAS) agar. Isolates were incubated at 30 °C for 3–5 days, and the development of an orange halo surrounding the colonies indicated siderophore secretion [49]. Potassium solubilization was examined by spot inoculation on plate containing potassium-bearing minerals, followed by incubation at 30 °C for 7 days. The presence of a clear halo zone around colonies was considered positive for potassium solubilization [50].
2.6. Evaluation of Environmental Stress Tolerance
The tolerance of the bacterial isolates to abiotic stresses was evaluated by monitoring their growth under different salinity and pH conditions. Bacterial strains (5% v/v) were inoculated into Ashby’s liquid medium supplemented with different concentrations of NaCl (1%, 5%, and 10% w/v). The cultures were incubated at 30 °C with shaking at 180 rpm. Similarly, Ashby’s liquid medium was adjusted to different pH levels (5, 8, and 10) using sterile HCl or NaOH solutions. Bacterial strains were inoculated and incubated as described above. Bacterial growth was monitored by measuring the optical density at 600 nm (OD600) at 24-hour intervals for 3 days.
2.7. Genetic Modification
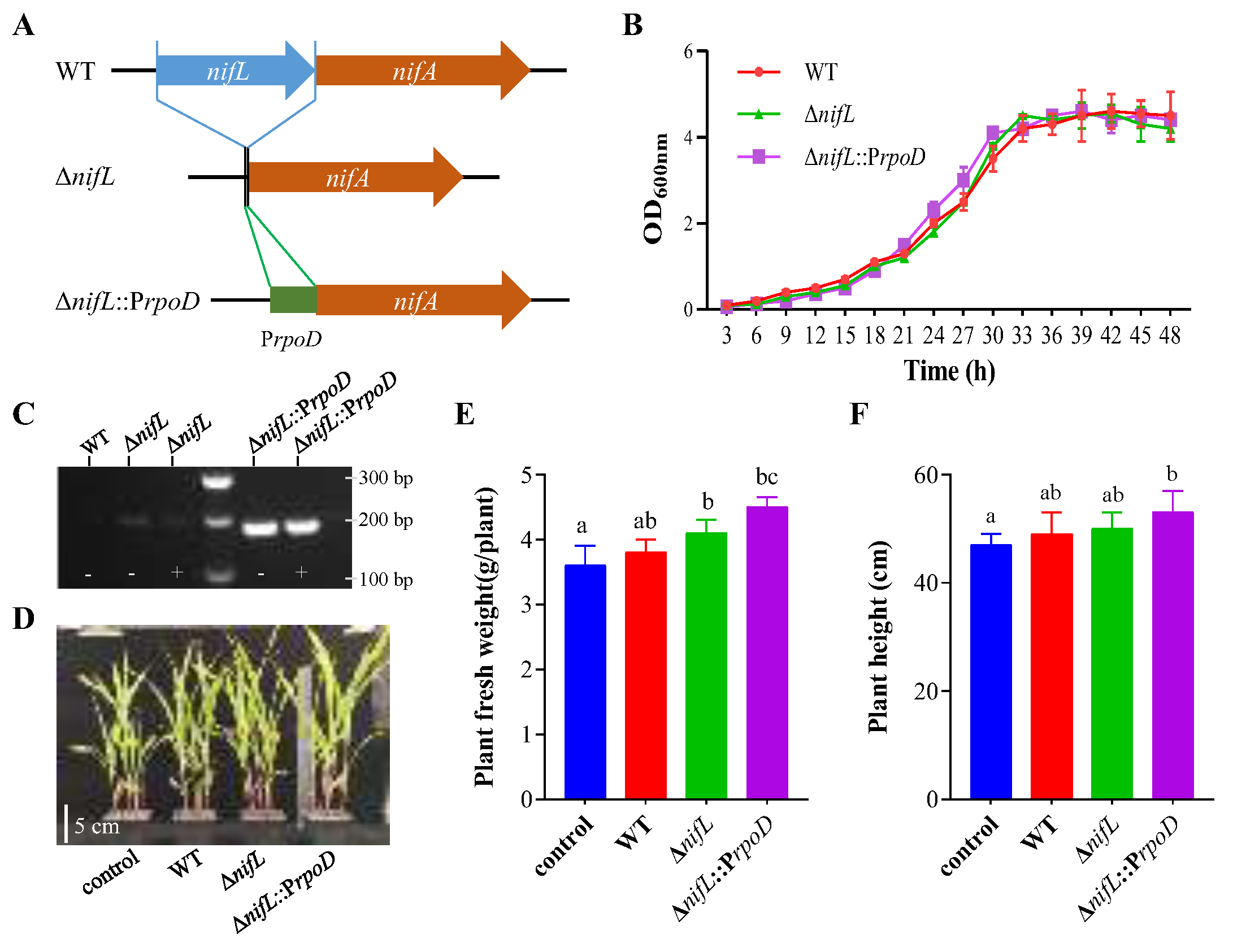
Strains, plasmids, and primers used in this work are listed in Table S1 in the supplemental material. The genetic modification procedure was performed as described previously [51]. The pJQ200SK was digested by a SmaI. The upstream and downstream homologous arms (600-800 bp) of the nifL gene were amplified by PCR and sequentially inserted into the sucrose-sensitive suicide plasmid pJQ200SK-Km using the In-Fusion cloning method, constructing the knockout vector pJQ200SK-ΔnifL. This vector was introduced into GN8811 via conjugation. Single-crossover integrants were selected with kanamycin resistance. Subsequently, sucrose counter-selection was performed to induce a second crossover event, yielding the nifL deletion mutant (ΔnifL). The mutant was verified by PCR and sequencing. Furthermore, to constitutively express the nifA gene (a positive regulator), a DNA fragment containing the constitutive rpoD promoter was synthesized and used to replace the native promoter of nifA in the ΔnifL mutant background via a similar homologous recombination strategy, generating the engineered strain ΔnifL::PrpoD.
The transcript levels of nifA in the wild-type and mutant strains under low-nitrogen (0 mM NH4NO3) and high-nitrogen (10 mM NH4NO3) conditions were analyzed by semi-quantitative RT-PCR.
2.8. Pot Experiment to Evaluate Plant Growth Promotion
Maize seeds (maize varieties Zhengdan 958, ZD958) were surface-sterilized with 75% ethanol for 3 minutes and 2% sodium hypochlorite for 5 minutes, followed by rinsing thoroughly with sterile distilled water. The seeds were then coated with bacterial suspensions (OD600 ≈ 1.0) of the wild-type or engineered strains for 2 hours. Seeds coated with sterile water served as the control. The treated seeds were sown in pots (one plant per pot) filled with a sterile mixture of sand and vermiculite (2:1, v/v). The plants were grown in a greenhouse under a 16/8 h light/dark cycle at 25-28 °C. The following fertilizer treatments were applied: 1) 100% nitrogen (full N, 200 kg N/ha equivalent), 2) 80% nitrogen, 3) 0% nitrogen, and 4) 0% nitrogen + bacterial inoculation. Each treatment had at least three replicates. Plants were watered as needed with a nitrogen-free Hoagland nutrient solution. After 45 days of growth, plant height, root length, and the fresh weight of shoots and roots were measured to assess the plant growth promotion effects of the bacterial inoculations.
2.9. Data Visualization and Statistical Analysis
Data visualization was performed using GraphPad Prism 10 software. Statistical analysis of phenotypic data from bacterial strain inoculation, based on three independent biological replicates, was conducted using one-way ANOVA followed by Tukey’s test (α= 0.05) in SPSS 25. Significant differences are indicated by lowercase letters.
3. Results
3.1. Isolation and Nitrogen-Fixing Activity Assessment of Nitrogen-Fixing Bacteria from Maize Rhizosphere
Using a multi-step enrichment process on nitrogen-free medium, we isolated a total of seven bacterial strains from maize rhizosphere soil that were capable of stable growth under nitrogen-free conditions (Figure 1). All strains formed viscous bacterial lawns on the nitrogen-free medium and were designated as GN1004, GN8799, GN2003, GN8811, GN2001, GN8801, and GN1202 (Figure 1 A). Further detection of the nifH gene, a key marker for nitrogen fixation, confirmed that all seven isolates possessed the nifH gene (Supplementary Table 1), with a core conserved fragment length of approximately 250 bp (Figure 1 B). Moreover, nitrogenase activity was highest in GN8811, followed by GN2003, GN2001, GN8799 and GN1202 (which showed similar activities), and then GN1004 and GN8801 (which showed the lowest and similar activities) (Figure 1 C). Additionally, A1501, a well-characterized model strain, exhibited moderate nitrogenase activity (Figure 1 C).
3.2. Phylogenetic Analysis of Isolated Nitrogen-Fixing Bacteria
To elucidate the taxonomic status of the seven isolated strains, a phylogenetic analysis based on 16S rRNA gene sequences was conducted. The results revealed that the seven strains were distributed across four genera and six species (Figure 2), demonstrating a high level of microbial diversity. Specifically, strain GN2001 was identified as Azotobacter chroococcum, GN1202 as Azotobacter vinelandii, GN1004 as Azospirillum brasilense, GN2003 as Kosakonia sacchari, and GN8799 as Klebsiella michiganensis. Both GN8801 and GN8811 were classified as Klebsiella quasivariicola (Figure 2).
3.3. Evaluation of Plant Growth-Promoting Traits of the Isolated Strains
To comprehensively evaluate the plant growth-promoting potential of the seven isolated strains, their abilities to produce indole-3-acetic acid (IAA), solubilize phosphate, produce siderophores, and solubilize potassium were qualitatively assessed. The results indicated that strain GN1004 exhibited the strongest IAA production, as evidenced by the deepest pink coloration on the white porcelain plate assay, followed by GN8811, while GN2003 demonstrated the weakest activity (Figure 3 A). In terms of phosphate solubilization, only GN8801 formed a visible dissolution zone on the agar plate (Figure 3 B). For siderophore production, solely GN2001 demonstrated significant iron-chelating ability, forming a distinct halo on the assay plate (Figure 3 C). Regarding potassium solubilization, five strains—GN2001, GN8799, GN8801, GN8811, and GN2003—exhibited varying abilities to solubilize potassium, with GN2003 showing the weakest activity, indicated by a faint dissolution zone (Figure 3 D).
3.4. Physiological Characterization of Acid, Alkali, and Salt Tolerance in the Test Isolates
Furthermore, to assess the tolerance of the tested strains to adverse environmental conditions, their growth status was evaluated under different salt concentrations and pH levels. The results demonstrated that higher salinity significantly inhibited bacterial growth (Figure 4 A). Specifically, at 10% NaCl concentration, biomass was markedly suppressed and showed negligible increase over time. In contrast, at lower salt levels (1% and 5%), the biomass of all strains gradually increased with prolonged incubation, with greater biomass accumulation observed at lower salinity (Figure 4 A). Notably, strains GN2001 and GN1202 exhibited the lowest salt tolerance, while GN2003 and GN8811 showed the highest. The remaining strains displayed intermediate tolerance without significant differences. Similarly, alkaline conditions (pH 10) strongly inhibited the growth of most strains, resulting in a significant reduction in biomass (Figure 4 B). In comparison, at pH 5 and pH 8, bacterial biomass increased substantially (Figure 4 B). Among the tested strains, GN1004, GN8799, GN2003, GN8811, and GN8801 demonstrated better adaptation to varying pH conditions, whereas GN2001 and GN1202 were the most sensitive to acidic and alkaline stress (Figure 4 B).
3.5. Inoculation Effects of Nitrogen-Fixing Bacterial Strains on Maize Growth
The plant growth-promoting effects of the nitrogen-fixing bacteria were evaluated through inoculation experiments. Higher nitrogen application levels generally resulted in improved plant growth, as reflected by increased plant height, as well as shoot fresh weights and root fresh weight (Figure 5). Overall, the growth performance of maize inoculated with strain GN8811 was comparable to that of the 100% nitrogen treatment, and even exceeded it in terms of root length and root weight (Figure 5 C, E). Regarding plant height, all inoculation treatments performed similarly to the 80% N and 0% N treatments, while GN8811 inoculation achieved results equivalent to the 100% N treatment (Figure 5 B). In terms of root length, both GN2003 and GN8811 significantly outperformed all nitrogen treatment groups and other inoculated groups (Figure 5 C). For root weight, GN8799 and GN8811 reached levels comparable to the 100% N treatment, and GN8799, GN8811, and GN1201 all exceeded the 80% N treatment level, whereas the remaining inoculation treatments showed no significant difference from the 80% N treatment (Figure 5 E). Regarding shoot fresh weight, all bacterial inoculations significantly outperformed the 0% N treatment. Among them, GN8801 and GN8811 reached the level of the 80% N treatment, though slightly lower than that of the 100% N treatment (Figure 5 D).
3.6. Genetic Modification of GN8811 Enhanced Plant-Growth Promotion
To enhance the nitrogen-fixing capability of the outstanding strain GN8811, genetic modifications were performed to eliminate nitrogen fixation repression and strengthen nifA expression (Figure 6A). The wild-type (WT), ΔnifL mutant, and ΔnifL::PrpoD engineered strain showed comparable growth rates (Figure 6 B). Under high-nitrogen conditions, nifA expression was undetectable in the WT, while the ΔnifL mutant exhibited weak nifA expression under both low- and high-nitrogen conditions. In contrast, the ΔnifL::PrpoD strain demonstrated strong nifA expression regardless of nitrogen availability (Figure 6 C). Further pot experiment results confirmed that inoculation with GN8811 WT significantly promoted maize growth (Figure 6 D), reflected by notable increases in fresh weight and plant height (Figure 6 E, F). Moreover, maize inoculated with the ΔnifL and ΔnifL::PrpoD strains showed further improvement in growth compared to the WT, with ΔnifL::PrpoD exhibiting the most pronounced enhancing effect (Figure 6 E, F).
4. Discussion
This study isolated seven indigenous nitrogen-fixing rhizobacteria from the maize rhizosphere, revealed considerable taxonomic and functional diversity among them, and identified GN8811 as an outstanding strain with strong nitrogenase activity, multiple plant growth-promoting traits, and broad environmental tolerance. Furthermore, targeted genetic engineering of GN8811 by deleting nifL and enhancing nifA expression markedly improved nitrogen fixation and maize growth promotion, even under high-N conditions. Together, these results demonstrate a dual strategy—ecological screening plus regulatory-circuit engineering—for developing robust microbial inoculants that can support maize production with reduced dependence on synthetic N fertilizers.
Our findings are broadly consistent with previous reports showing that maize rhizospheres harbor diverse diazotrophs, including Azotobacter, Azospirillum, Klebsiella and Kosakonia, which contribute to N supply and plant growth promotion through BNF and additional PGP traits such as phytohormone production and nutrient solubilization [52,53,54,55]. The identification of Azotobacter chroococcum, Azotobacter vinelandii and Azotobacter brasilense among our isolates agrees with studies showing these genera as common maize-associated diazotrophs with IAA production and nutrient-solubilizing capacities [56,57]. Our discovery of Klebsiella quasivariicola and Klebsiella michiganensis, together with Kosakonia sacchari, corroborates recent work highlighting Enterobacteriaceae as important plant-associated diazotrophs capable of colonizing maize roots and mucilage and promoting grain yield under reduced N fertilization [58,59]. Compared with previous studies, our work extends the characterization of maize-associated diazotrophs in several ways. First, we systematically evaluated multiple PGP traits across all isolates, showing clear functional differentiation: GN1004 had the strongest IAA production, GN8801 was the only strain with detectable phosphate solubilization, GN2001 uniquely produced siderophores, and several strains (including GN8801 and GN8811) solubilized potassium. Similar trait partitioning has been described in other PGPR consortia where different strains contribute complementary functions [60,61], suggesting that assembling GN2001, GN8801, GN8811 and others into tailored consortia may further enhance maize performance. Second, our evaluation of salt and pH tolerance revealed that GN2003 and GN8811 maintained relatively robust growth under 10% NaCl and across pH 5–10, which is comparable to or higher than tolerance levels reported for many diazotrophs used in saline or acidic soils [62,63]. Such broad tolerance is particularly relevant for regions facing soil salinization and acidification due to intensive N fertilization [64,65].
The inoculation experiments showed that GN8811, and to a lesser extent GN2003 and GN8799, substantially improved maize biomass, root length and root weight, achieving performance similar to or exceeding 100% N-fertilized controls for specific traits. These results align with previous field and pot studies where inoculation with Azospirillum, Herbaspirillum and Enterobacteriaceae enabled 20–30% reductions in mineral N without yield penalties [66,67], indicating a realistic potential for partial replacement (20–30%) of synthetic N inputs under controlled conditions. However, we also observed that not all isolates with nifH and nitrogenase activity produced equally strong growth responses, and that different strains preferentially enhanced specific components (e.g., root length versus shoot biomass). Such variability echoes earlier reports of inconsistent field responses driven by strain × genotype × environment interactions [68,69] and underscores the importance of matching inoculant strains to target environments and crop varieties. A key novelty of this study lies in the genetic enhancement of GN8811 via modification of the NifL–NifA regulatory system. While previous work has proposed and demonstrated that altering nifL or engineering nifA expression can partially uncouple nitrogenase from classic ammonium/nitrate repression in diazotrophs [38,39,70], limiting the contribution of inoculants in fertilized fields. Our results also confirmed that a ΔnifL::PrpoD construct in a maize-associated Klebsiella strain maintains strong nifA expression and enhances plant growth under high nitrogen concentration. By showing that engineered GN8811 can still promote maize growth when mineral nitrogen is abundant, indicating rational modification of regulatory circuits can improve the reliability of N-fixing inoculants under realistic management regimes.
Despite these strengths, several limitations must be acknowledged. Our inoculation experiments were conducted under controlled pot conditions; although they simulate different N levels, they do not fully capture the complexity and variability of field environments, including competition with native microbiota, fluctuating temperature and moisture, and multi-year crop rotations [71]. Additionally, we primarily used culture-based methods and 16S rRNA phylogeny, which may underestimate the diversity of uncultured diazotrophs and provide limited resolution of strain-level genomic differences that shape PGP traits and environmental adaptation [72]. Finally, the long-term ecological and biosafety implications of releasing genetically modified Klebsiella strains into agricultural soils—particularly with enhanced N-fixation under high N—require rigorous assessment, including horizontal gene transfer risks and effects on native microbial communities and greenhouse gas fluxes [73,74,75].
Future research should therefore prioritize multi-location field trials to validate the performance and stability of GN8811 and its engineered derivatives across contrasting soil types, climates and management systems, ideally coupled with 15N-based quantification of BNF contributions. Comparative genomics and transcriptomics of GN8811, GN2003 and other promising isolates could reveal the genetic determinants underlying their high nitrogenase activity, stress tolerance and PGP traits, informing further strain improvement and the design of synthetic consortia. In addition, stacking regulatory modifications with traits that enhance rhizosphere competitiveness, root colonization and formulation stability—such as biofilm formation, exopolysaccharide production, or compatibility with mycorrhizal fungi—may produce next-generation inoculants with greater robustness [76,77,78]. At a broader scale, integrating engineered diazotrophs with precision N management and climate-smart agronomic practices could enable more predictable reductions in synthetic N use while maintaining or enhancing maize yields [79,80,81]. Ultimately, by combining exploration of local microbial diversity with targeted regulatory engineering, this work contributes to the development of reliable, high-performance biofertilizers that support sustainable intensification of maize-based agroecosystems.
5. Conclusion
In summary, this study isolated and characterized multiple nitrogen-fixing bacteria from the maize rhizosphere, which possessed various PGP traits and exhibited broad environmental tolerance. Among these, Klebsiella quasivariicola GN8811 was the most outstanding. More importantly, through precise genetic manipulation (deleting nifL and enhancing nifA expression), we successfully constructed engineered strains with higher nitrogen fixation efficiency and enhanced plant growth promotion, providing excellent candidate strains and a novel technological pathway for reducing reliance on chemical nitrogen fertilizers.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Acknowledgments
This work was supported by Scientifc Research Project of Yibin University (No. 2017RC03), Solidstate Fermentation Resource Utilization Key Laboratory of Sichuan Province (No.2018GTY001), Sichuan Science and Technology Program (No. 2021YJ0294).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Shahwar, D., et al., Role of microbial inoculants as bio fertilizers for improving crop productivity: A review. Heliyon, 2023. 9(6). [CrossRef]
- Dikir, W., Effect of long-term chemical fertilizer application on soil chemical properties: a review. J Biol Agric Healthc, 2023. 13: p. 11-18.
- Kim, H.-W., et al., Screening and isolated of plant growth promoting rhizobacteria Azotobacter sp. from maize rhizosphere. Probe - Plant & Animal Sciences, 2025. [CrossRef]
- Barron, S., F. Mus, and J.W. Peters, Nitrogen-Fixing Gamma Proteobacteria Azotobacter vinelandii—A Blueprint for Nitrogen-Fixing Plants? Microorganisms, 2024. 12(10): p. 2087. [CrossRef]
- Hossain, M.E., S. Shahrukh, and S.A. Hossain, Chemical fertilizers and pesticides: impacts on soil degradation, groundwater, and human health in Bangladesh, in Environmental degradation: challenges and strategies for mitigation. 2022, Springer. p. 63-92.
- Penuelas, J., F. Coello, and J. Sardans, A better use of fertilizers is needed for global food security and environmental sustainability. Agriculture & Food Security, 2023. 12(1): p. 1-9. [CrossRef]
- Sahoo, R.K., et al., Phenotypic and molecular characterisation of efficient nitrogen-fixing Azotobacter strains from rice fields for crop improvement. Protoplasma, 2013. 251(3): p. 511-523. [CrossRef]
- Macias-Bobadilla, I., et al., Hormetic and xenohormetic potential in the phytobiome of the center of origin. Genetic Resources and Crop Evolution, 2020. 67(5): p. 1331-1344. [CrossRef]
- Yang, J., et al., Mechanisms underlying legume–rhizobium symbioses. Journal of Integrative Plant Biology, 2022. 64(2): p. 244-267. [CrossRef]
- Pedraza, R.O., Recent advances in nitrogen-fixing acetic acid bacteria. International Journal of Food Microbiology, 2008. 125(1): p. 25-35. [CrossRef]
- Saranraj, P., et al. Isolation and nitrogen fixing efficiency of Gluconacetobacter diazotrophicus associated with sugarcane: A review. in IOP Conference Series: Earth and Environmental Science. 2021. IOP Publishing.
- Madhaiyan, M., et al., Improvement of plant growth and seed yield in Jatropha curcas by a novel nitrogen-fixing root associated Enterobacter species. Biotechnology for biofuels, 2013. 6(1): p. 140. [CrossRef]
- Li, H.-B., et al., Genetic diversity of nitrogen-fixing and plant growth promoting Pseudomonas species isolated from sugarcane rhizosphere. Frontiers in Microbiology, 2017. 8: p. 1268. [CrossRef]
- Jalal, A., et al., Plant growth-promoting bacteria and nitrogen fixing bacteria: sustainability of non-legume crops, in Nitrogen fixing bacteria: sustainable growth of non-legumes. 2022, Springer. p. 233-275.
- Bhattacharjee, R.B., A. Singh, and S. Mukhopadhyay, Use of nitrogen-fixing bacteria as biofertiliser for non-legumes: prospects and challenges. Applied microbiology and biotechnology, 2008. 80(2): p. 199-209. [CrossRef]
- Kuan, K.B., et al., Plant growth-promoting rhizobacteria inoculation to enhance vegetative growth, nitrogen fixation and nitrogen remobilisation of maize under greenhouse conditions. PloS one, 2016. 11(3): p. e0152478. [CrossRef]
- Abadi, V., et al., Diversity and abundance of culturable nitrogen-fixing bacteria in the phyllosphere of maize. Journal of Applied Microbiology, 2021. 131(2): p. 898-912. [CrossRef]
- Yuan, L., C. San-Feng, and L. Ji-Lun, Colonization pattern of Azospirillum brasilense Yu62 on maize roots. Journal of Integrative Plant Biology, 2003. 45(6): p. 748.
- Takahashi, W.Y., et al., Tracking maize colonization and growth promotion by Azospirillum reveals strain-specific behavior and the influence of inoculation method. Plant Physiology and Biochemistry, 2024. 215: p. 108979. [CrossRef]
- Monteiro, R.A., et al., Herbaspirillum-plant interactions: microscopical, histological and molecular aspects. Plant and Soil, 2012. 356(1): p. 175-196. [CrossRef]
- Di Salvo, L.P., et al., Plant growth-promoting rhizobacteria inoculation and nitrogen fertilization increase maize (Zea mays L.) grain yield and modified rhizosphere microbial communities. Applied Soil Ecology, 2018. 126: p. 113-120.
- Van Deynze, A., et al., Nitrogen fixation in a landrace of maize is supported by a mucilage-associated diazotrophic microbiota. PLoS biology, 2018. 16(8): p. e2006352.
- Sun, W., M.H. Shahrajabian, and Q. Cheng, Nitrogen fixation and diazotrophs–a review. Rom Biotechnol Lett, 2021. 26(4): p. 2834-2845. [CrossRef]
- Aira, M., et al., Plant genotype strongly modifies the structure and growth of maize rhizosphere microbial communities. Soil Biology and Biochemistry, 2010. 42(12): p. 2276-2281. [CrossRef]
- Zhao, M., et al., Zonal soil type determines soil microbial responses to maize cropping and fertilization. MSystems, 2016. 1(4): p. 10.1128/msystems. 00075-16. [CrossRef]
- Pittol, M., et al., Agronomic and environmental aspects of diazotrophic bacteria in rice fields. Annals of Microbiology, 2016. 66(2): p. 511-527. [CrossRef]
- Namsaraev, Z., et al., Effect of salinity on diazotrophic activity and microbial composition of phototrophic communities from Bitter-1 soda lake (Kulunda Steppe, Russia). Extremophiles, 2018. 22(4): p. 651-663. [CrossRef]
- Wang, Y., et al., Soil pH is a major driver of soil diazotrophic community assembly in Qinghai-Tibet alpine meadows. Soil Biology and Biochemistry, 2017. 115: p. 547-555. [CrossRef]
- Chukwudi, U.P., et al., Field application of beneficial microbes to ameliorate drought stress in maize. Plant and Soil, 2025: p. 1-20. [CrossRef]
- Wang, R., et al., Carbon allocation to the rhizosphere is affected by drought and nitrogen addition. Journal of Ecology, 2021. 109(10): p. 3699-3709. [CrossRef]
- Dixon, R. and D. Kahn, Genetic regulation of biological nitrogen fixation. Nature Reviews Microbiology, 2004. 2(8): p. 621-631. [CrossRef]
- Wu, S.C., et al., Effects of biofertilizer containing N-fixer, P and K solubilizers and AM fungi on maize growth: a greenhouse trial. Geoderma, 2005. 125(1-2): p. 155-166. [CrossRef]
- Wu, Z.H., et al., A synthetic bacterial consortium improved the phytoremediation efficiency of ryegrass on polymetallic contaminated soil. Ecotoxicol Environ Saf, 2024. 282: p. 116691. [CrossRef]
- Brtnicky, M., et al., Biodegradation of poly-3-hydroxybutyrate after soil inoculation with microbial consortium: Soil microbiome and plant responses to the changed environment. Sci Total Environ, 2024. 946: p. 174328. [CrossRef]
- Akashdeep, S. Kumari, and N. Rani, Novel cereal bran based low-cost liquid medium for enhanced growth, multifunctional traits and shelf life of consortium biofertilizer containing Azotobacter chroococcum, Bacillus subtilis and Pseudomonas sp. J Microbiol Methods, 2024. 222: p. 106952. [CrossRef]
- Rosenblueth, M., et al., Nitrogen Fixation in Cereals. Front Microbiol, 2018. 9: p. 1794.
- Zhao, Z., A.R. Fernie, and Y. Zhang, Engineering nitrogen and carbon fixation for next-generation plants. Curr Opin Plant Biol, 2025. 85: p. 102699. [CrossRef]
- Mus, F., et al., Genetic Determinants of Ammonium Excretion in nifL Mutants of Azotobacter vinelandii. Appl Environ Microbiol, 2022. 88(6): p. e0187621. [CrossRef]
- Lan, P., et al., Modification of nifL-nifA operon of Klebsiella oxytoca improves N(2)-fixation efficiency, ammonium-excretion and plant-growth promotion ability. Arch Microbiol, 2025. 207(10): p. 253. [CrossRef]
- Wu, W., et al., Harnessing biological nitrogen fixation: Multi-scale engineering for self-sustaining agroecosystems. Biotechnol Adv, 2025. 84: p. 108687. [CrossRef]
- Soumare, A., et al., Exploiting Biological Nitrogen Fixation: A Route Towards a Sustainable Agriculture. Plants (Basel), 2020. 9(8). [CrossRef]
- Kim, M., M. Morrison, and Z. Yu, Evaluation of different partial 16S rRNA gene sequence regions for phylogenetic analysis of microbiomes. Journal of microbiological methods, 2011. 84(1): p. 81-87. [CrossRef]
- Kumar, S., et al., MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol Biol Evol, 2018. 35(6): p. 1547-1549. [CrossRef]
- Deslippe, J.R., K.N. Egger, and G.H. Henry, Impacts of warming and fertilization on nitrogen-fixing microbial communities in the Canadian High Arctic. FEMS microbiology ecology, 2005. 53(1): p. 41-50. [CrossRef]
- Widmer, F., et al., Analysis of nifH gene pool complexity in soil and litter at a Douglas fir forest site in the Oregon Cascade Mountain Range. Applied and Environmental Microbiology, 1999. 65(2): p. 374-380. [CrossRef]
- Lv, F., et al., Regulation of hierarchical carbon substrate utilization, nitrogen fixation, and root colonization by the Hfq/Crc/CrcZY genes in Pseudomonas stutzeri. Iscience, 2022. 25(12). [CrossRef]
- Patel, D., et al., A resourceful methodology to profile indolic auxins produced by rhizo-fungi using spectrophotometry and HPTLC. 3 Biotech, 2018. 8(10): p. 413. [CrossRef]
- Muleta, A., et al., Phosphate solubilization and multiple plant growth promoting properties of Mesorhizobium species nodulating chickpea from acidic soils of Ethiopia. Arch Microbiol, 2021. 203(5): p. 2129-2137. [CrossRef]
- Himpsl, S.D. and H.L.T. Mobley, Siderophore Detection Using Chrome Azurol S and Cross-Feeding Assays. Methods Mol Biol, 2019. 2021: p. 97-108.
- Sen, A., et al., Assessing the effectiveness of indigenous phosphate-solubilizing bacteria in mitigating phosphorus fixation in acid soils. 3 Biotech, 2024. 14(9): p. 197. [CrossRef]
- Cui, W.J., et al., Lineage-Specific Rewiring of Core Pathways Predating Innovation of Legume Nodules Shapes Symbiotic Efficiency. mSystems, 2021. 6(2). [CrossRef]
- Sumbul, A., et al., Azotobacter: A potential bio-fertilizer for soil and plant health management. Saudi J Biol Sci, 2020. 27(12): p. 3634-3640. [CrossRef]
- Fukami, J., P. Cerezini, and M. Hungria, Azospirillum: benefits that go far beyond biological nitrogen fixation. AMB Express, 2018. 8(1): p. 73. [CrossRef]
- Bao, Y.Q., et al., Construction, Characterization, and Application of an Ammonium Transporter (AmtB) Deletion Mutant of the Nitrogen-Fixing Bacterium Kosakonia radicincitans GXGL-4A in Cucumis sativus L. Seedlings. Curr Microbiol, 2023. 80(2): p. 58. [CrossRef]
- Yoshidome, D., et al., Glutamate production from aerial nitrogen using the nitrogen-fixing bacterium Klebsiella oxytoca. Commun Biol, 2024. 7(1): p. 443. [CrossRef]
- Wang, J., et al., Beneficial bacteria activate nutrients and promote wheat growth under conditions of reduced fertilizer application. BMC Microbiol, 2020. 20(1): p. 38. [CrossRef]
- Ayyaz, K., et al., Isolation and identification by 16S rRNA sequence analysis of plant growth-promoting azospirilla from the rhizosphere of wheat. Braz J Microbiol, 2016. 47(3): p. 542-50. [CrossRef]
- Liang, P., et al., Klebsiella michiganensis: a nitrogen-fixing endohyphal bacterium from Ustilago maydis. AMB Express, 2023. 13(1): p. 146. [CrossRef]
- Bloch, S.E., et al., Biological nitrogen fixation in maize: optimizing nitrogenase expression in a root-associated diazotroph. J Exp Bot, 2020. 71(15): p. 4591-4603.
- Ma, H., et al., Synthetic Microbial Community Isolated from Intercropping System Enhances P Uptake in Rice. Int J Mol Sci, 2024. 25(23). [CrossRef]
- Wang, Z., et al., A Synthetic Microbial Community of Plant Core Microbiome Can Be a Potential Biocontrol Tool. J Agric Food Chem, 2023. 71(13): p. 5030-5041. [CrossRef]
- Sharma, S., J. Kulkarni, and B. Jha, Halotolerant Rhizobacteria Promote Growth and Enhance Salinity Tolerance in Peanut. Front Microbiol, 2016. 7: p. 1600. [CrossRef]
- Ishaq, S., et al., Plant growth-promoting bacteria from Uzungöl forest stimulate rice growth via seed biopriming and root inoculation: isolation and functional characterization of potent PGPR strains from rhizosphere soils of different trees. Front Plant Sci, 2025. 16: p. 1622951. [CrossRef]
- Zheng, Y., et al., Effect of planting salt-tolerant legumes on coastal saline soil nutrient availability and microbial communities. J Environ Manage, 2023. 345: p. 118574. [CrossRef]
- Huang, K., et al., Soil acidification and salinity: the importance of biochar application to agricultural soils. Front Plant Sci, 2023. 14: p. 1206820. [CrossRef]
- Hungria, M., et al., Improving maize sustainability with partial replacement of N fertilizers by inoculation with Azospirillum brasilense. Agronomy Journal, 2022. 114(5): p. 2969-2980. [CrossRef]
- dos Santos, C.L.R., et al., Contribution of a mixed inoculant containing strains of Burkholderia spp. and Herbaspirillum ssp. to the growth of three sorghum genotypes under increased nitrogen fertilization levels. Applied Soil Ecology, 2017. 113: p. 96-106.
- Schmidlin, K., et al., Environment-by-Environment Interactions Differ Across Genetic Backgrounds. Mol Biol Evol, 2025. 42(11). [CrossRef]
- Gimmi, E. and C. Vorburger, Strong genotype-by-genotype interactions between aphid-defensive symbionts and parasitoids persist across different biotic environments. J Evol Biol, 2021. 34(12): p. 1944-1953. [CrossRef]
- Gerhardt, E.C.M. and K.A. Selim, Unlocking nitrogen regulation: structural insights into the NifL-NifA complex and prospects for engineered diazotrophs. Febs j, 2025. [CrossRef]
- Poorter, H., et al., Pot size matters: a meta-analysis of the effects of rooting volume on plant growth. Funct Plant Biol, 2012. 39(11): p. 839-850. [CrossRef]
- Leontidou, K., et al., Plant growth promoting rhizobacteria isolated from halophytes and drought-tolerant plants: genomic characterisation and exploration of phyto-beneficial traits. Sci Rep, 2020. 10(1): p. 14857. [CrossRef]
- Smets, B.F. and T. Barkay, Horizontal gene transfer: perspectives at a crossroads of scientific disciplines. Nat Rev Microbiol, 2005. 3(9): p. 675-8. [CrossRef]
- Atieno, M., et al., Assessment of biofertilizer use for sustainable agriculture in the Great Mekong Region. J Environ Manage, 2020. 275: p. 111300. [CrossRef]
- Vejan, P., et al., Role of Plant Growth Promoting Rhizobacteria in Agricultural Sustainability-A Review. Molecules, 2016. 21(5). [CrossRef]
- Singh, R.P. and K. Kumari, Bacterial type VI secretion system (T6SS): an evolved molecular weapon with diverse functionality. Biotechnol Lett, 2023. 45(3): p. 309-331. [CrossRef]
- Bhagwat, A., et al., Bacterial metabolism in the host and its association with virulence. Virulence, 2025. 16(1): p. 2459336. [CrossRef]
- do Amaral, F.P., et al., Diverse Bacterial Genes Modulate Plant Root Association by Beneficial Bacteria. mBio, 2020. 11(6). [CrossRef]
- Das, H.K., Azotobacters as biofertilizer. Adv Appl Microbiol, 2019. 108: p. 1-43.
- Gómez-Godínez, L.J., et al., A Look at Plant-Growth-Promoting Bacteria. Plants (Basel), 2023. 12(8).
- Ercole, T.G., et al., The role of endophytic bacteria in enhancing plant growth and health for sustainable agriculture. Antonie Van Leeuwenhoek, 2025. 118(7): p. 88. [CrossRef]
Figure 1.
Isolation, characterization, and nitrogenase activity assessment of isolated nitrogen-fixing strains. (A) The colony morphology of the isolated strains on Ashby’s medium; (B) The nifH amplification of isolated strains; (C) Nitrogenase activity determination of isolated strains. A1501, a well-studied nitrogen-fixing bacteria, was used as a positive control. Error bars represent SD. Significant differences between means were indicated by different letters based on ANOVA followed by Tukey’s test (α= 0.05).
Figure 1.
Isolation, characterization, and nitrogenase activity assessment of isolated nitrogen-fixing strains. (A) The colony morphology of the isolated strains on Ashby’s medium; (B) The nifH amplification of isolated strains; (C) Nitrogenase activity determination of isolated strains. A1501, a well-studied nitrogen-fixing bacteria, was used as a positive control. Error bars represent SD. Significant differences between means were indicated by different letters based on ANOVA followed by Tukey’s test (α= 0.05).
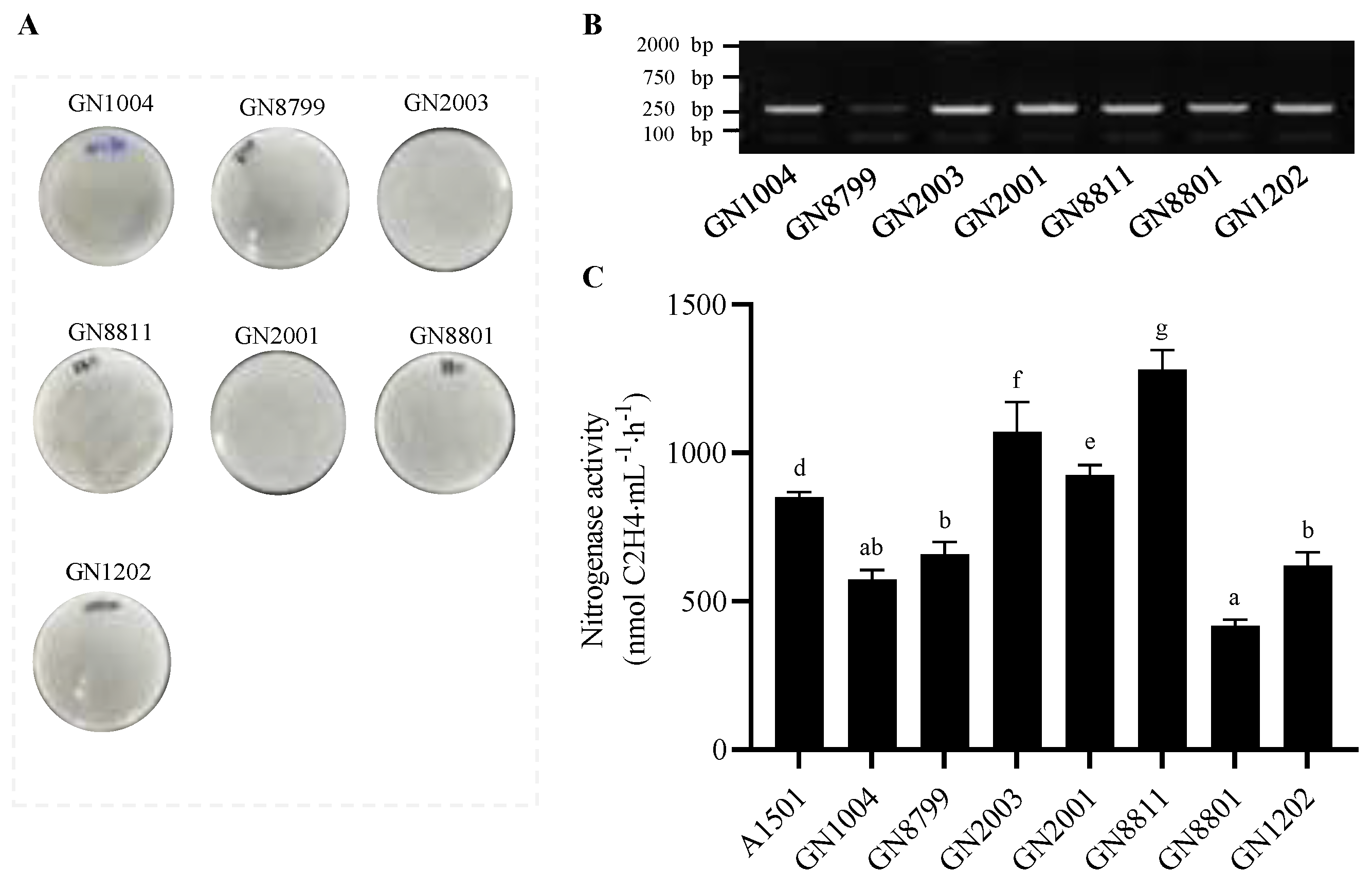
Figure 2.
The phylogenetic tree based on 16S rRNA genes of isolated strains. Maximum likelihood phylogenetic trees of 21 (7 candidates, 14 reference strains) 16S rRNA sequences was constructed by Mega X software. Bootstrap values above 60% were shown at the branch points. Bar, 0.02 substitutions per nucleotide position. Accession numbers of the reference sequences from GenBank were given in brackets.
Figure 2.
The phylogenetic tree based on 16S rRNA genes of isolated strains. Maximum likelihood phylogenetic trees of 21 (7 candidates, 14 reference strains) 16S rRNA sequences was constructed by Mega X software. Bootstrap values above 60% were shown at the branch points. Bar, 0.02 substitutions per nucleotide position. Accession numbers of the reference sequences from GenBank were given in brackets.

Figure 3.
Quantitative assessment of plant growth-promoting characteristics of isolated strains under in vitro conditions. IAA biosynthesis (A), P solubilization(B), Siderophore production(C) and K solubilization(D) were tested, respectively.
Figure 3.
Quantitative assessment of plant growth-promoting characteristics of isolated strains under in vitro conditions. IAA biosynthesis (A), P solubilization(B), Siderophore production(C) and K solubilization(D) were tested, respectively.
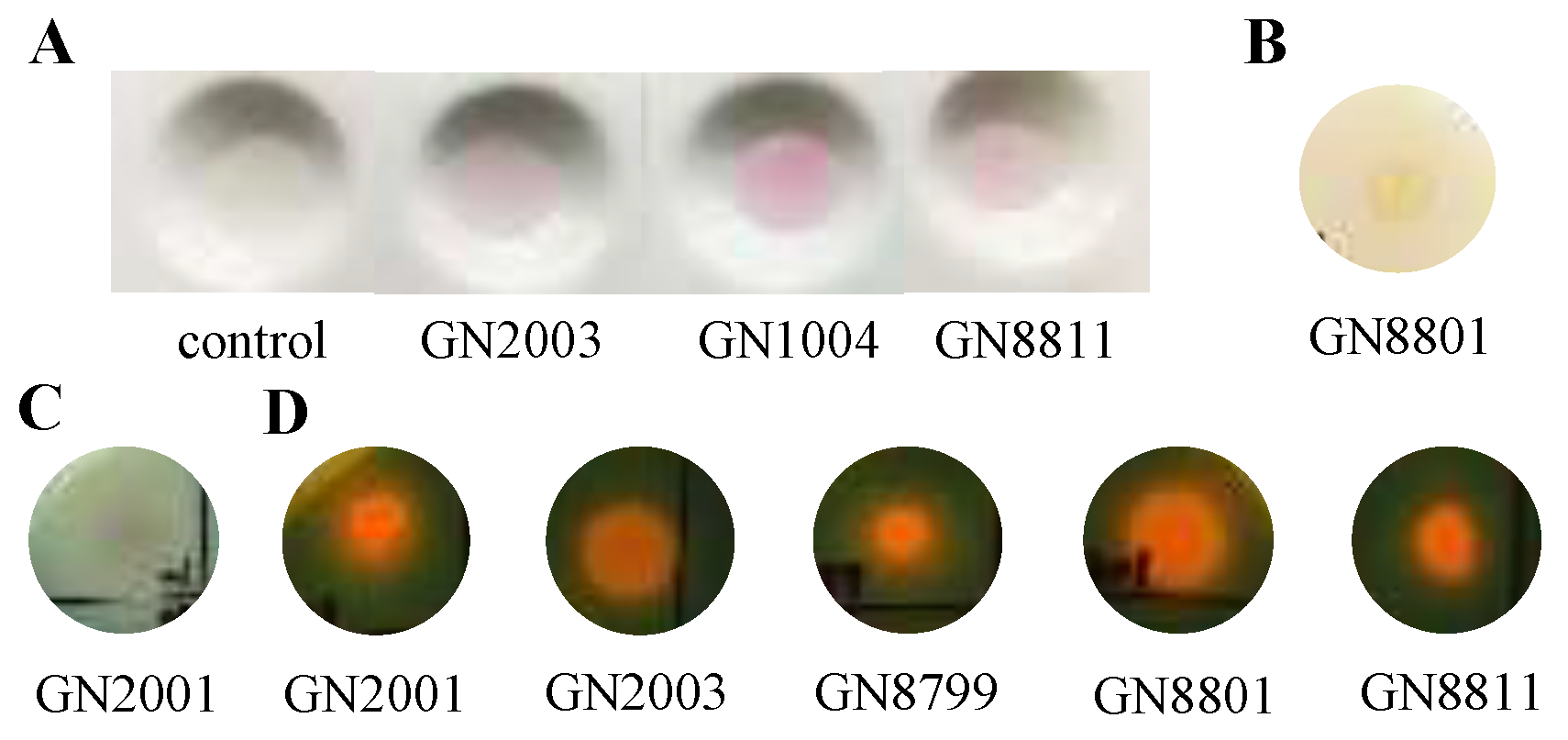
Figure 4.
Effect of varying NaCl concentrations (A) and pH levels (B) on the growth of the tested strains. DPI, days post inoculation. Error bars represent SD. At least three replications of the experiment were carried out.
Figure 4.
Effect of varying NaCl concentrations (A) and pH levels (B) on the growth of the tested strains. DPI, days post inoculation. Error bars represent SD. At least three replications of the experiment were carried out.
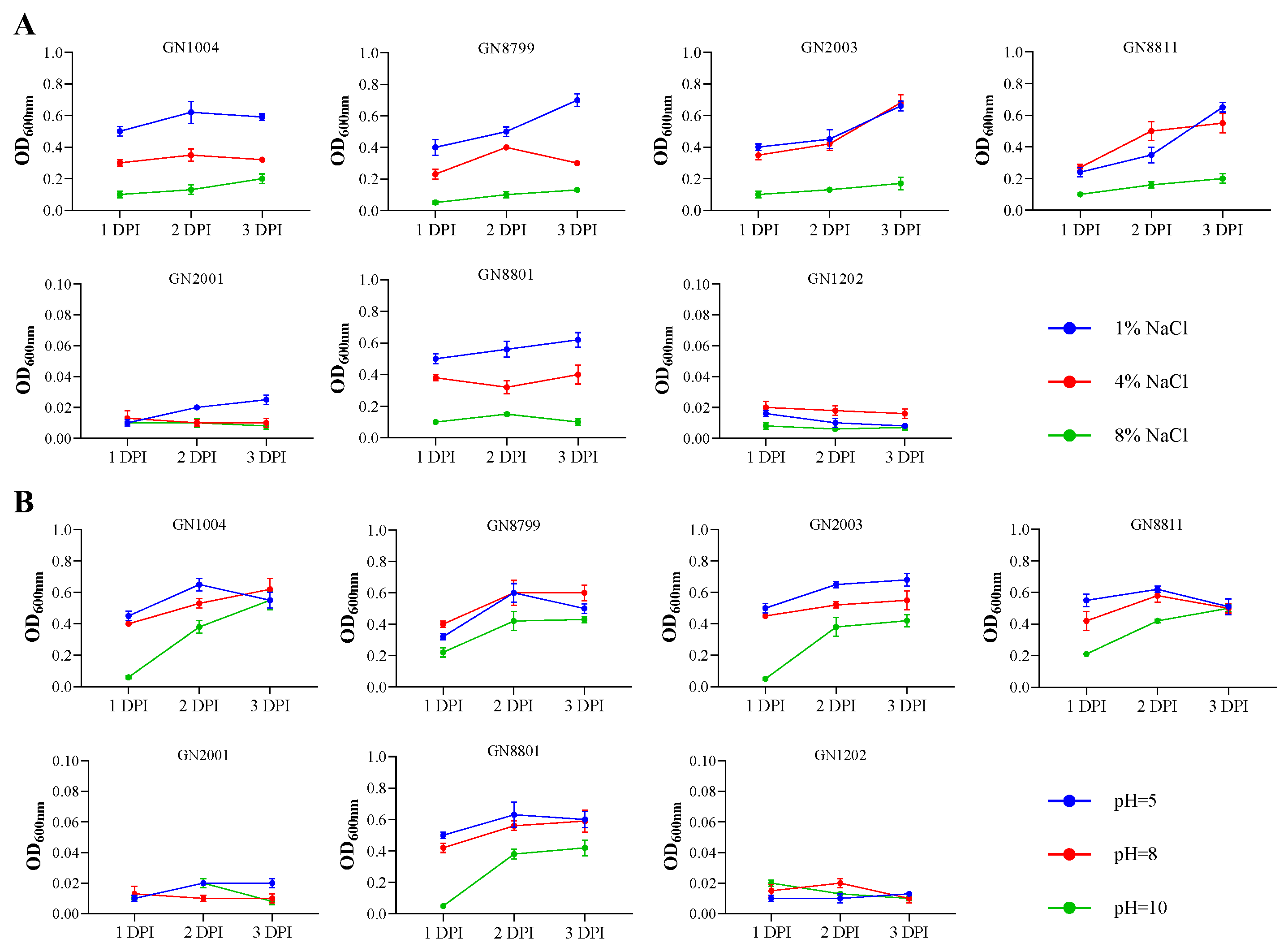
Figure 5.
The inoculation effect of tested strains on maize. (A) Growth and morphological phenotypes of maize plants treated with isolated strains. Shoot fresh weight (B), root fresh weight (C), shoot height (D) and root length (E) were measured to evaluate the effect of the inoculation performance of individual isolates on maize. Error bars represent SD. Significant differences between means are indicated by different letters based on ANOVA followed by Tukey’s test (alpha = 0.05). At least two additional replications of the experiment were carried out.
Figure 5.
The inoculation effect of tested strains on maize. (A) Growth and morphological phenotypes of maize plants treated with isolated strains. Shoot fresh weight (B), root fresh weight (C), shoot height (D) and root length (E) were measured to evaluate the effect of the inoculation performance of individual isolates on maize. Error bars represent SD. Significant differences between means are indicated by different letters based on ANOVA followed by Tukey’s test (alpha = 0.05). At least two additional replications of the experiment were carried out.
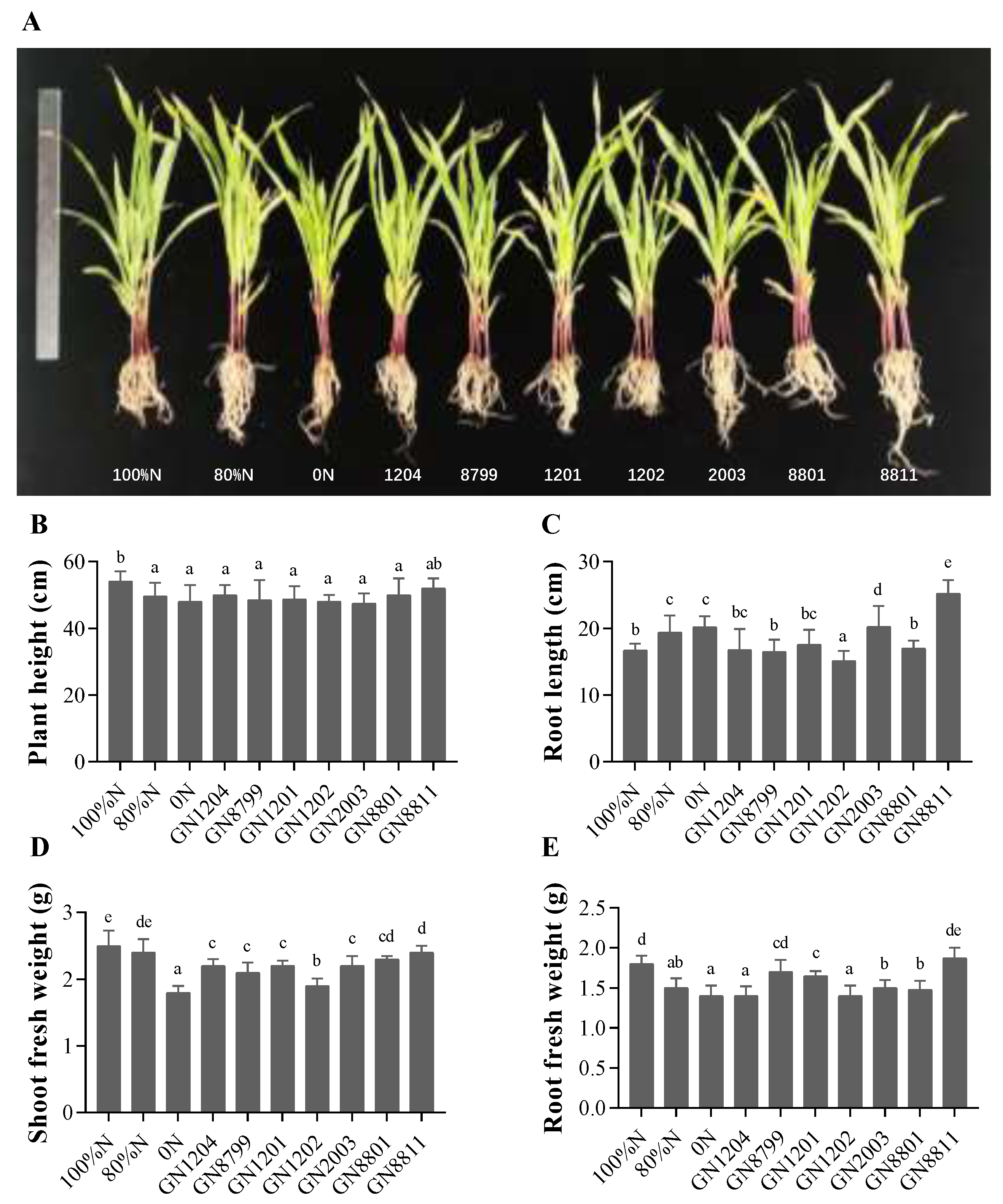
Figure 6.
The GN8811 genetic modification enhance nitrogenase activity and plant growth.(A) Genetic modification model. WT, wild type; ΔnifL,nifL mutant; ΔnifL::PrpoD,nifL mutant compensate with rpoD promoter. (B), Growth curve of GN8811 and its deritive mutants. (C), The nifA transcript activity was detected through semi-quantitative PCR. The symbols - and + represent nitrate ammonium concentrations of 0 mM and 10 mM, respectively. Inoculation performance (D), plant fresh weight (E) and plant height (F) were used to evaluate the effect of the individual isolates on maize growth. Error bars represent SD. Significant differences between means are indicated by different letters based on ANOVA followed by Tukey’s test (alpha = 0.05). At least two additional replications of the experiment were carried out.
Figure 6.
The GN8811 genetic modification enhance nitrogenase activity and plant growth.(A) Genetic modification model. WT, wild type; ΔnifL,nifL mutant; ΔnifL::PrpoD,nifL mutant compensate with rpoD promoter. (B), Growth curve of GN8811 and its deritive mutants. (C), The nifA transcript activity was detected through semi-quantitative PCR. The symbols - and + represent nitrate ammonium concentrations of 0 mM and 10 mM, respectively. Inoculation performance (D), plant fresh weight (E) and plant height (F) were used to evaluate the effect of the individual isolates on maize growth. Error bars represent SD. Significant differences between means are indicated by different letters based on ANOVA followed by Tukey’s test (alpha = 0.05). At least two additional replications of the experiment were carried out.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



