
Submitted:
22 May 2025
Posted:
22 May 2025
You are already at the latest version
Abstract
A methanotroph, Methylomonas strain Kb3, isolated from an Indian rice field, was found to enhance rice growth yield in a small-scale field experiment. Two methanotrophs, Methylomonas strain Kb3 and Methylomagnum ishizawai strain KRF4, and their combination, termed mixed MOB (methane-oxidizing bacteria), were used for this experiment. Methylomonas Kb3 was helpful in enhancing the plant growth height and yield in the field experiment, compared to the control plants. Methylomagnum ishizawai KRF4 alone did not show an increase in grain yield, thus indicating that there could be variability in the effects of methanotrophs on rice growth. However, the mixed MOB showed considerable positive effects, but less compared to that of Methylomonas sp. strain Kb3. The nitrogen input in this field has been considerably low (50kg N/ha) compared to the normally used fertilizer input (100 or 150 kg N/ha), and hence, this field was chosen. Methylomonas strain Kb3 was isolated by our group in 2014, from a field ~25 km away from the region where trials were taken, and the genome analysis has been done in our prior studies, indicating that it has a nitrogen fixation pathway. The strain also induced early flowering in the rice plants, followed by early grain formation and early maturation. Kb3 showed an increase in plant height of ~ 15% and an increase in grain yield of ~17% compared to the control plants, which did not receive any methanotroph inoculant. Mixed MOB showed ~15% height increase and ~15% yield increase compared to the control, whereas Methylomagnum ishizawai KRF4 showed a lesser total growth yield compared to the control plants.
Keywords:
Methanotrophs
; Methane
; rice fields
; Methylomonas
; Methylomagnum
; bio-inoculants
; yield
1. Introduction
Approximately 20% of human calories come from rice, one of the three major staple cereals consumed worldwide (8, 2). Given that the world’s population is expected to surpass 10 billion people during the next 30 years, there will likely be a major increase in demand for paddy farming [10]. Asia produces 90% of the world’s rice, and India is the world’s second-biggest producer behind China, with the largest area under cultivation (about 43.7 million hectares) (Agricultural-statistics-at-a-Glance-2020.pdf). Traditional rice farming practices follow growing the paddy in a waterlogged state, mainly due to the belief that rice is an aquatic crop, but this practice also helps eliminate the growth of weeds and pests (8, 1). However, the flooding reduces the diffusion of oxygen into water, creating an anoxic environment, where organic soil substrates are reduced by the anaerobic microbes, resulting in methane. Methane is the second most important greenhouse gas after carbon dioxide, with the potency to trap 26 times more heat than CO2, and has a relatively short half-life of ~12 years [4]. Thus, reducing methane generation from these sites can largely aid in controlling the emissions. As per the latest IPCC report [3], about 335-383 Tg methane is produced per year, of which rice fields contribute up to 10% globally, i.e., ~25-37 Tg methane/ year. The latest available data on the current status of methane regulation from India suggests that the rice fields emit ~3.9 Tg of methane per year [6]. And the yearly highest peak in the methane emission is observed between August and September, the season when maximum rice is cultivated in the country [6]. The balance between the opposite processes of methane production and oxidation determines the net amount of methane released into the atmosphere from rice fields. Methanotrophs, or methane-oxidizing bacteria, oxidize methane and are nature’s natural methane mitigation agents [9]. Methanotrophs are majorly alpha and gamma proteobacteria groups of bacteria that dwell in the oxic-anoxic interface of rice fields as well as in rice roots and play a role in the carbon-methane cycle by oxidizing methane to carbon dioxide, thus minimizing the methane emissions from the soil into the atmosphere [7,10,27]. Methane oxidation significantly reduces methane’s ability to diffuse into the atmosphere in rice fields. The presence and methanotrophic oxidizing activity of methanotrophs in rice rhizospheres oxidize up to 20% of the produced methane, thereby reducing the net flux [4]. The widely varied levels of methane and oxygen generated by roots in the rice rhizosphere suggest that the habitat of methanotrophs is highly heterogeneous [7]. Rice roots mainly harbor Type I methanotrophs, though both Type I and Type II methanotrophs are present in the soil and rhizosphere region [5,7,27]. However, methanotrophs only make up a very small portion (~1%) of the total soil flora in nature [17], thus it would be interesting to investigate whether adding more methanotrophs through inoculation or spraying can improve methane mitigation and benefit plant growth. Since higher growth output would be cost-effective for farmers, our team has been conducting experiments to see if methanotrophs can impact rice plant growth. Therefore, the goal of this study was to find an appropriate and simple way to administer methanotrophs to rice plants at the small field level. We have conducted such experiments in pots in recent years and discovered that the natively isolated methanotrophs can stimulate plant growth when administered to rice plants [11]. It would be intriguing to investigate whether more methanotroph supplementation through inoculation or spraying would improve methane mitigation and have beneficial impacts on plant growth, as methanotrophs only make up a very small portion (~1%) of the overall soil flora in nature [17]. Our team has been conducting experiments to see if methanotrophs can boost rice plant growth because it would be cost-effective for farmers to have higher growth yields. In order to find a viable and simple approach for applying methanotrophs to rice plants at the small field level, we focused on this study. In recent years, we have conducted these experiments in pots and discovered that the natively isolated methanotrophs can stimulate plant growth when used on rice plants [11].
2. Results
2.1. Overall Health of the Plants and Early Flowering Seen in Methylomonas Kb3-Treated Plants
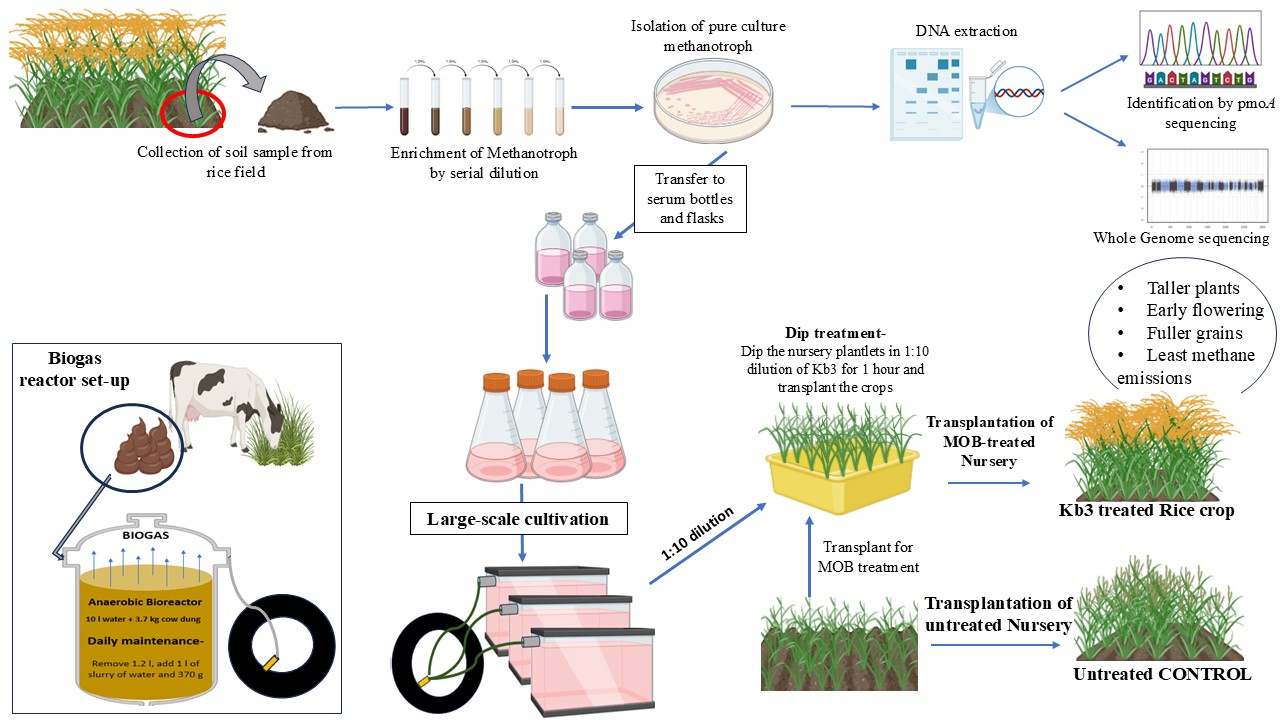
In 1L or 2L flasks, methanotrophs, Methylomonas Kb3, and Methylomagnum ishizawai KRF4 were grown in 7–10 days, and on the day of transplantation, the inoculum was transported to the field. When compared to the control plants (plants which received no inoculation), all of the methanotroph-inoculated plants were generally green and healthy at every stage and displayed no discernible variations or adverse effects (Figure 1a). During the transplantation, the field was totally flooded (rainfed), and no additional binding agent was used to allow the methanotrophs to adhere to the rice roots (Figure 1b).
Early blooming or flowering (about 10–12 days earlier than in the control and other treatments) was noted in cases where Methylomonas Kb3 was treated. Early grain formation and maturation in Methylomonas Kb3-treated plants further demonstrated this; on September 19, 2024, the day of the observations, 66% of the plants displayed grain formation (4/6 plants). Conversely, only the Methylomagnum KRF4 inoculation plants and the control plants displayed (Figure 2). About 50% of the plants (3/6 plants) that were inoculated with a mixture of MOB, specifically Methylomonas Kb3 and Methylomagnum ishizawai KRF4, showed variable outcomes. While the remaining plants in the plot only displayed flowering or early grain production (early stages), it is evident that Methylomonas Kb3-treated plants and a few mixed MOB-treated plants displayed the grain maturation stage (which drooped owing to weight) (Figure 2).
2.2. Enhanced Growth Yield, Plant Height in Methylomonas Kb3-Treated Plants
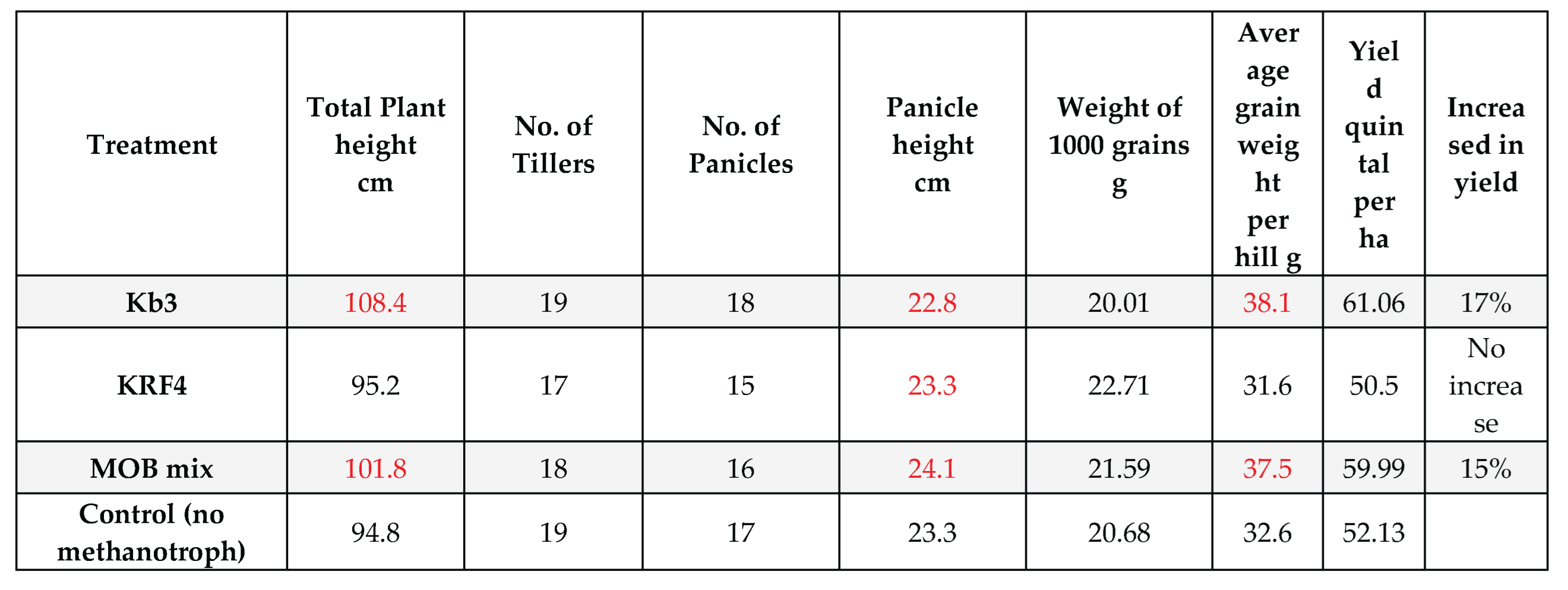
Methylomonas strain Kb3 and MOB mix outperformed the others in terms of plant height, panicle height, and tiller development height (Table 1, Figure 3a, 3b). Grain yield was also higher in plants treated with Methylomonas Kb3 after receiving combined MOB therapy.
Eight rice hills, each measuring around 100 cm by 50 cm (5000 cm2=0.5 square meter), were harvested for each treatment. The data were then extrapolated from grain yield in g/m2 to quintals/ha or tons/ha by multiplying by the factor. Treating plantlets with Methylomonas Kb3 at the transplantation stage was beneficial because the plant height increased by approximately 15% and the total grain yield increased by 17%, 61.06 quintals/ha total yield, which is approximately nine quintals/ha more than the control plants (Table 1, Figure 3b). With 59.99 quintals of grains/ha, which was almost 8 quintals/ha more than the control plant yields, the mixed MOB, which combined Methylomonas and Methylomagnum, also shown similar yield differences, but slightly less than the pure culture employed (Table 1, Figure 3b). The control showed about 52.13 grains in quintals/ha. With a 17% increase in grain output, Kb3 demonstrated the highest yield, followed by the MOB mix with a 15% increase. The KRF4 treatment alone did not appear to be helpful because the total grain production was only slightly less than the control grain output (50.5 quintals/ha). In terms of 1000-grain weight, KRF4 showed a slightly higher weight compared to the other treatments; however, due to fewer panicles, the total grain yield was probably lower in the case of the KRF4 treatment (Table 1). The number of tillers, panicles, and panicle height showed no significant difference in all the plants, control as well as inoculated (Figure 3a). The total plant height was significantly higher in Methylomonas Kb3 inoculated plants and mixed MOB inoculated plants compared to the control (Figure 3 b). In the same trial, plants inoculated with Methylomonas strain Kb3 also showed early flowering, grain development, and grain maturation roughly 10-12 days before the other plants, reducing the rice season by 2 weeks (Fig.S2). This was also evident from the more dried grains in Methylmonas Kb3 inoculated plants at the harvest. The panicle number and height were higher in the inoculated methanotrophs, except in KRF4 inoculated plants, where the panicle number was a little less. Principal Component Analysis (PCA) was performed using all plant growth parameters listed in Table 2. The analysis revealed a clear separation between the plants inoculated with Kb3 and the Mix MOB, as compared to the control and KRF4 treatments. The first two principal components accounted for 90.6% of the total variance (Figure 4). Vectors associated with plant height, grain yield per hill, and estimated yield were closely aligned with Kb3 and MOB mix. In contrast, the control and KRF4 treatments clustered together in the left lower quadrant. Overall, the PCA effectively distinguished the different inoculation treatments based on plant growth parameters.
2.3. Re-Isolation of Methanotrophs
The serial endpoint dilution method was used for re-isolation of methanotrophs from the applied plants as described before [18]. Rice root samples with attached soil after about 27 days of transplantation at the tillering stage were used for the serial dilution in microtiter plates and isolation from the last dilutions. It was seen that methanotrophs grew till 10-8 dilution in the case of Methylomonas Kb3 and mixed MOB, showing characteristic pink color growth. In case of Methylomagnum KRF4, white growth was observed in the last positive dilutions, and microscopic examination revealed the presence of Methylomagnum ishizawai KRF4 cells Figure S1. Methylomonas Kb3 could be re-isolated from the enrichment culture done in an endpoint dilution series in liquid, followed by streaking on solid agarose plates. The colony morphology and cell morphology confirmed that Methylomonas-like cells were observed. Further, pmoA PCR amplification and sequencing of the isolate confirmed that the culture was indeed Methylomonas Kb3, which was associated with the rhizosphere soil. Similarly, in the case where KRF4 was applied, KRF4 was isolated along with some other methanotrophs. The morphology of KRF4 was again confirmed by microscopy, where characteristic large, elliptical cells of Methylomagnum could be observed (Figure S1). As Methylomagnum ishizawai KRF4 has more than five μm in size and a distinct shape (Figure S1B), [18] it could be quickly confirmed using microscopic observation. In the control soil, the abundance of methanotrophs was less, i.e., growth was obtained only up to 10-3 or 10-4 dilution, and Type II-like methanotrophs (of the genera Methylosinus and Methylocystis were cultured (as depicted by microscopic observations and colony morphology, half-moon-shaped or pear-shaped: Methylosinus with cream-colored colonies and coccoid-shaped with white colonies: Methylocystis), Figure S1.
3. Discussion
3.1. Need for Novel Bio-Inoculants in Rice
Since rice is a staple in most countries, efforts must be made to cultivate paddy in a way that produces fuller grains with minimal requirements of N-based fertilizers like urea, which are costly for both the environment and farmers, and emit less methane in order to meet the growing demand for nutritional needs. Utilizing urea and other nitrogenous fertilizers degrades the land’s fertility and contaminates the surrounding waterways [23]. Therefore, it is imperative to move towards sustainable practices to continue supporting life on Earth. Rice cultivation has a higher requirement of nitrogenous fertilizers, usually supplied as Urea or DAP (diammonium phosphate). The consumption of Urea in India in the year 2022-2023 was 35.7 million tons. The Indian government is working on a national policy to boost local fertilizer manufacturing and reduce dependency on imports. Excessive and inefficient use of fertilizers leads to nutrient losses to the environment and could also result in drinking water contamination and impact human lives as a result of unsafe storage practices. Fertilizers are also hazardous to the environment and contribute largely to land, water, and air pollution [28]. As a result of the increasing population needs in the future, India will have a large requirement of food grains by 2050, 400 million tonnes, against the current production of 285 million tonnes [25]. Given the massive requirement for nutrients, the total nutrient needs of Indian soils cannot be met only through organic and bio sources. The efficiency of chemical fertilizers increases when used in combination with organic manure. Chemical fertilizers are expensive to farmers, thus reducing their profit margins. Using fertilizers each consecutive year destroys the natural flora and fertility of the land. The run-offs from the fields enter water bodies, and the rich nitrogen and phosphate components encourage eutrophication-like conditions. Excess nitrogenous and sulfur emissions escape the atmosphere, contributing to the greenhouse effect [23]. Hence, a good alternative to chemical-based fertilizers is to use bacteria with plant growth-promoting abilities, which have been considered essential in enhancing plant health without being hazardous to the environment [24].
3.2. Nitrogen Fixation by Methanotrophs
Methanotrophs also often possess nitrogen-fixation pathways and are active players in nitrogen fixation in rice roots [1]. As per an estimate, methanotrophs are capable of fixing 1.2 to 1014 g N/year [29] globally. The ability of methanotrophs to fix nitrogen has been demonstrated earlier in pure cultures [13] and by studying the nitrogen fixation genes/pathways [1]. Our recent studies, where we sequenced the draft genomes of methanotrophs that were isolated from Indian rice fields, all showed complete nitrogen fixation pathways, e.g., in Methylomonas sp., Methylococcus sp., Methylobacter sp., Methylolobus sp., etc. [12]. Stable isotope probing (SIP) experiments with 13CH4 and 15N2 [4] have recently verified that methanotroph-mediated nitrogen fixation occurs in rice roots. The nitrogen fixation in methane-consuming roots was found to be dominated by Methylomonas. The authors also noted that the microbial N2-fixing activity was stimulated by low nitrogen conditions [4].
3.3. Methanotrophs as a New Class of Bio-Inoculants
Methanotrophs, a unique class of prokaryotic bacteria, seem to have a solution to this problem as they are one of the only known biological filters of methane [9]. Our research group has been a pioneer in isolating and cultivating methanotrophs in India and has been exploring the potential of methanotrophs in plant growth promotion for a few years [15,16,17,18,19,20,21,22]. Using a distinctive cultivation method to isolate methanotrophs, methanotrophs belonging to seven distinctive genera of three classes (Type Ia, Type Ib, and Type II) were cultured from rice fields in India [18]. As methanotrophs make up the natural rhizosphere flora of rice fields, it has been hypothesized that they may also contribute to the growth promotion in terms of increased biomass and grain yield by means such as nitrogen fixation [12]. To cover the demands of the growing population, there is a need for the use of fertilizers to ensure healthy crops with higher yields. Bioinoculants act as natural fertilizers for crops, promoting their growth while caring for their nitrogen requirements. Only a few bacteria that primarily fix atmospheric nitrogen are used as biofertilizers for rice cultivation. Both free-living diazotrophs, such as Azospirillum and Azotobacter, and Anabaena-Azolla-based fertilizers are being used for rice [26]. So far, bioinoculants have been successfully prepared using microbes like Trichoderma, Azospirillum, Azotobacter, Bacillus, Pseudomonas, etc., in the plant growth promotion of various crops in the form of solid encapsulation, liquid sprays, or emulsifiers [14]. Certain substitute agricultural practices like alternate wetting and drying, direct seeded rice, and improved rice cultivars have been noted and published for their efficiency in curbing methane emissions. However, there is a requirement for specific infrastructures, and educating the farmers about the standard methodologies is necessary.
In our first studies done in pots, we found that the use of selected methanotroph cultures as bioinoculants in rice agriculture promoted increased plant height, early flowering, and enhanced grain yield, as well as negligible methane emissions from the sites due to the high methane-oxidizing activity of the inoculated methanotrophs [12]. In these trials, Type I methanotrophs were observed to be better at increasing grain yield both individually and in consortia (~35%). In contrast, Type II methanotrophs increased plant height with no significant role in increasing grain yield when used individually, but increased grain yield and height when used in consortia (~12%). In contrast to the standard dosage used in India, which is approximately 150 kg N per ha or 135 kg Urea per acre, low nitrogen was applied in the trials, at about 10 or 47.5 kg of Urea per acre, or roughly 1/10th or 1/3rd of the amount of nitrogen, respectively [11]. Although it was conducted on a limited scale and with the farmer’s consent and assistance during the experiment, this field study is among the first to use pure cultures of methanotrophs in the field. In this case, the farmer had been applying 50 kg N/ha annually, which is roughly one-third to one-fourth of the dosage applied in a typical setting. (150 kg N/ha-200 kg N/ha). This land was perfect for our experimentation because the farmer had been cultivating with low nitrogen inputs for a long time. Usually, if the inorganic N fertilizer input is low, there is a chance that biological nitrogen fixation can take place in bacteria, as demonstrated in switchgrass [2]. Since Methylomonas Kb3 exhibits the genes for the nitrogen fixation pathway and is capable of growing in nitrogen-free environments, nitrogen fixation may be one of the factors contributing to increased plant growth output, even if we have not shown this to be the case [11,18]. To determine the causes of the increased yields and the role of nitrogen fixation, more research is required. Due to the early grain formation and maturity observed in Methylomonas Kb3 inoculations, rice can mature faster, allowing farmers to harvest earlier. Recently, a rice variety has been developed in India, with early maturation and the advantages of 10-15 days early maturation, low nitrogen fertilizer requirement: Recipe with science, sentiment, and rice, and thus, our results showing early maturation with Methylomonas Kb3 are important. It was also observed that, in contrast to spraying or directly applying the methanotroph inoculant on the field, dipping rice roots during the transplantation stage was a simpler technique that farmers could readily adopt. The directly administered inoculum may wash out because the rice crop is frequently inundated. Similarly, because of the frequent rains during this time, it might not be feasible to spray the inoculum.
3.4. A Methanotroph, Methylococcus Capsulatus, Has Been Shown to Act as a Bio-Stimulant on Multiple Levels
Recently, a company in India (Stringbio, Bangalore, India) used a similar approach, and a methanotroph biostimulant (consisting of Methylococcus capsulatus termed CleanRise R) was used over three rice seasons in open-field studies, where up to 39% extra grain yield was obtained (under 100% fertilizer condition) and ~34% extra grain yield with 75% fertilizer application [11]. In the same study, it has also been claimed that there was a 30-60% reduction in methane emissions as the methanotroph bioinoculant was applied. The potential mechanisms for the microbial bio-stimulant effect on rice were due to indole acetic acid production, modulation of photosynthesis, tillering, and panicle development, which together resulted in a superior yield [11]. As this study used plant transcript analysis, there is now a stronger basis for the hypothesis that methanotrophs can be used for rice plant growth promotion and methane mitigation. Though this study used a single methanotroph (Methylococcus capsulatus), similar mechanisms could exist in the interactions of methanotrophs and rice plants, as similar basic biochemical pathways exist in most methanotrophs [9].
We believe that a similar mechanism may occur in the case of Methylomonas Kb3 interaction with rice, as observed with Methylococcus capsulatus [10], even if we have not examined the specific cause of the increased rice yield in our experiments by examining the rice transcriptome. Additionally, we found that the field trial yielded a 17% higher growth output than the control and that the use of low nitrogen fertilizer (1/3rd or 1/4th of the usual dose) increased productivity while also improving the sustainability of rice agriculture. Additionally, the reported yield for the Indrayani variety, which is approximately 70 Quintals/ha or 7 tons/ha if the full fertilizer dose and irrigation are used, was comparable to the total rice yield obtained with Methylomonas Kb3 inoculation (61.06 Quintals/ha or 6.11 tons/ha) with minimal inorganic fertilizer input New Paddy Varieties in India: High-Yielding, Disease-Resistant, and Aromatic Rice Cultivars.
The effectiveness of the methanotroph inoculation in reducing methane emissions from this significant niche would also be determined by additional multi-location studies in the Western India region using Methylomonas Kb3 and measurements of methane emissions from the rice fields.
4. Materials and Methods
4.1. Selection of a Rice Field for the Field Experiment
A rice farm in Malegaon village near Lavasa was selected to conduct the experiments after talking to the farmer about the farming practices and getting his permission. The farmer (Mr. Shivajirao Gund, farm owner) agreed and allowed us to experiment on his rice field. All the experiments were performed in his presence. The field is located at 18.4097° N and 73.5066° E on the Lavasa road in the interiors of the village, Malegaon. This area has been traditionally used for rice farming, and the rice farmer practiced traditional rice farming by transplanting rice and using relatively low nitrogen fertilizer inputs, i.e., with low Urea inputs (50 kg N/ha), which were ideal for our experiments. We visited the field initially in May 2024 to get an idea of the area, and then on 20th June 2024 (around ~15 days after he had sown the grains), when the plants were in the early nursery stage. The entire nursery was prepared under natural conditions with no Urea additions, as per our instructions. Approximately 2 L of each fully grown and fresh culture was carried to the field in plastic cans, which consisted of ~1-1.5 OD of the culture. The following methanotrophs were used: Methylomonas strain Kb3 (Type Ia, methanotroph, this methanotroph has been one of the promising candidates for the application) and Methylomagnum ishizawai strain KRF4 (second candidate for the application) were used for the field application, and a combination of the same, termed as mixed MOB (methane-oxidizing bacteria). The experiment was continued till the end of the rice season, and the mature grains were harvested. The overall experiment (Figure 1a) shows the different stages of the rice field experiment.
4.2. Treatment of Rice Plantlets with Selected Methanotroph Strains
The field experiment was started at the transplantation stage (3rd July 2024). Plots measuring 4m x 3m were marked using wooden sticks (Figure 1b), and a randomized plot design was used. The ~25-day-old nursery plantlets were briefly washed by dipping in a tub of water and then treated with the selected methanotroph cultures by keeping them dipped in the diluted inoculum, and a mixed consortium was prepared with both strains mixed in the field, before application. Thus, the following four treatments were given: Kb3, KRF4, Kb3+KRF4 (MOB) mix, and control (NMS medium) in the ratio of 1:20 (1L of inoculum and 19L of water) for 1 hour, and the treated plants were carefully transplanted to the field. The plants were planted in a pre-marked rectangle of 4m x 3m in rows using a rope with a 25cm distance between each plant hill, resulting in roughly 16 plant hills per square meter and ~192-200 plant hills per treatment. After transplantation, the plants were allowed to stand in water, and no special treatment was given for the microbes to stick to the roots, thus allowing only tightly attached bacteria within the one-hour treatment to get carried over further.
4.3. Data Collection, Farm Visits and Soil Sampling
The field was visited regularly, and data were collected at the tillering, flowering-early grain filling, and grain maturation stages on 31st July 2024 (tillering stage), 19th September 2024 (flowering stage), and 28th October 2024 (grain maturation stage and harvest). The flowering stage and early grain formation stage were noted on 19th September 2024, and plants were photographed using a mobile camera. The treated rice crops were harvested after 4 months of transplantation and assessed for their growth in height, number of panicles, number of tillers, height of the panicle, weight of 1000 grains, and total weight of grains per hill.
4.4. Soil Sampling for Enrichment of Methanotrophs
Rhizosphere soil from the field was sampled at the tillering stage by uprooting three random plants from the treated and from the control plants. Three plants from each treatment were uprooted, and the roots with attached soil were further used. These were diluted in serial dilutions in microtiter plates and incubated in a methane and air environment in a desiccator as described (Rahalkar et al. 2021). After 3-4 weeks, the grown dilutions were streaked on NMS agarose plates. Colonies were purified, and the cultures were microscopically observed. Pure cultures were further subjected to pmoA PCR and sequencing analysis.
4.5. Data Analysis
MS Excel was used to store the date and also for the basic analysis like averages, standard deviation etc. PCA analysis was carried out using PAST software (version 4.17) [8].
5. Conclusions
Two indigenously isolated methanotrophs, Methylomonas sp. and Methylomagnum sp., were used individually and, in combination, in a farmer’s field to check for plant growth promotion in rice. The farmer used a low amount of nitrogen fertilizer in the form of Urea (~50kg N/ha). It was seen that Methylomonas strain Kb3, which was indigenously isolated from Western India, was able to enhance rice plant growth yield by 17%, and in combination with Methylomagnum, the growth yield was enhanced by ~15%. Other parameters, such as plant height and panicle numbers, were also enhanced in these treatments. Although this was a single field experiment, the results show promising data with respect to the use of methanotrophs as plant growth-promoting agents in rice, and prompt us to do multilocation trials using this methanotroph for the region. As the world population increases and demand for rice is growing, the need for sustainable solutions is rising. Methanotrophs may offer a solution in terms of novel bio-inoculants, which may also enable enhanced oxidation of methane emitted from rice agriculture.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Colony morphology and phase contrast microscopic images of methanotrophs under 100X oil immersion : (A) Type II methanotrophs isolated from Malegaon control soil, and microscopic image; (B) Methylomagnum ishizawai strain KRF4 re-isolated from KRF4-treated rice plants, and microscopic image; (C) Methylomonas strain Kb3 re-isolated from treated Kb3 rice plants and microscopic image. Figure S2: Grain harvest stage: Kb3 inoculated plants can be seen, which have already lodged down (in the front). In the back, control plants can be seen.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used: “Conceptualization, M.C.R. and R.B.; methodology, R.B., S.M., K.P., S.K., Y.K., Y.P.; validation, S.M., K.P., S.K.; formal analysis, S.M., K.P., S.K.; writing—original draft preparation, S.M. and M.C.R.; writing—review and editing, M.C.R. and S.M.; visualization, R.B. and M.C.R.; supervision, M.C.R.; project administration, M.C.R.; funding acquisition, M.C.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by SERB (ANRF), grant number SPF/2022/000045.
Acknowledgments
We thank and express our gratitude towards the rice farmer Mr. Shivajirao Gund (Malegaon village, Lavasa Road) for allowing us to conduct the experiments on his field. We are also thankful to his family, workers in the field, for their kind cooperation and help. We thank Ms. Jyoti Mohite for her assistance during the cultivation of methanotroph cultures. K.P. thanks UGC for her junior research fellowship.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| MOB | Methane-oxidizing bacteria |
References
- Auman AJ, Speake CC, Lidstrom ME (2001) nifH Sequences and Nitrogen Fixation in Type I and Type II Methanotrophs. Applied and Environmental Microbiology 67:4009-4016.
- Bahulikar RA, Chaluvadi SR, Torres-Jerez I, Mosali J, Bennetzen JL, Udvardi M (2021) Nitrogen Fertilization Reduces Nitrogen Fixation Activity of Diverse Diazotrophs in Switchgrass Roots. Phytobiomes . [CrossRef]
- Canadell JG, Monteiro PMS (2021) Global Carbon and Other Biogeochemical Cycles and Feedback. In: IPCC (ed) Contribution of Working Group I to the Sixth Assessment Report of the IPCC, pp 673-779.
- Conrad R (2009) The global methane cycle: recent advances in understanding the microbial processes involved. Environmental Microbiology Reports 1:285-292.
- Ferrando L, Tarlera S (2009) Activity and diversity of methanotrophs in the soil-water interface and rhizospheric soil from a flooded temperate rice field. J Appl Microbiol 106:306-316.
- Ganesan A, Rigby M, Lunt MF, Parker RJ, Boesch H, Goulding N, Umezawa T, Zahn A, Chatterjee A, Prinn RG, Tiwari YK, Schoot Mvd, Krummel PB (2017) Atmospheric observations show accurate reporting and little growth in India’s methane emissions. Nature Communications 8:836.
- Gilbert B, Frenzel P (1998) Rice roots and CH4 oxidation: the activity of bacteria, their distribution and the microenvironment. Soil Biol Biochem 30:1903-1916.
- Hammer O, Harper DAT, Ryan PD (2001) PAST: Paleontological statistics software package for education and data analysis. Paleontologica Electronica 4:9.
- Hanson RS, Hanson TE (1996) Methanotrophic bacteria. Microbiology and Molecular Biology Reviews 60:439-471.
- Hardoim PR, Andreote FD, Reinhold-Hurek B, Sessitsch A, van Overbeek LS, van Elsas JD (2011) Rice root-associated bacteria: insights into community structures across 10 cultivars. FEMS Microbiology Ecology 77:154-164.
- Kumar SR, David EM, Pavithra GJ, Kumar GS, Subbian E (2024) Methane-derived microbial biostimulant reduces greenhouse gas emissions and improves rice yield. Frontiers in Plant Science 15.
- Mohite J, Khatri K, Pardhi K, Manvi SS, Jadhav R, Rathod S, Rahalkar MC (2023) Exploring the potential of methanotrophs for plant growth promotion in rice agriculture. Methane 2:361-371.
- Murrell JC, Dalton H (1983) Nitrogen-fixation in obligate methanotrophs. Journal of General Microbiology 129:3481-3486.
- Orozco-Mosqueda MDC, Rocha-Granados MDC, Glick BR, Santoyo G (2018) Microbiome engineering to improve biocontrol and plant growth-promoting mechanisms. Microbiological Research 208:25-31.
- Pandit PS, Rahalkar M, Dhakephalkar P, Ranade DR, Pore S, Arora P, Kapse N (2016) Deciphering community structure of methanotrophs dwelling in rice rhizospheres of an Indian rice field using cultivation and cultivation independent approaches. Microbial Ecology 71:634-644.
- Pandit PS, Ranade DR, Dhakephalkar PK, Rahalkar MC (2016) A pmoA-based study reveals dominance of yet uncultured Type I methanotrophs in rhizospheres of an organically fertilized rice field in India. 3 Biotech 6:135.
- Pandit PS, Rahalkar MC (2019) Renaming of ‘Candidatus Methylocucumis oryzae’ as Methylocucumis oryzae gen. nov., sp. nov., a novel Type I methanotroph isolated from India. Antonie van Leeuwenhoek 112:955-959.
- Rahalkar M, Khatri K, Pandit PS, Bahulikar R, Mohite JA (2021) Cultivation of Important Methanotrophs From Indian Rice Fields. Frontiers in Microbiology:1-15.
- Rahalkar MC, Pandit P (2018) Genome-based insights into a putative novel Methylomonas species (strain Kb3), isolated from an Indian rice field. Gene Reports 13:9-13.
- Rahalkar MC, Khatri K, Pandit PS, Dhakephalkar PK (2019) A putative novel Methylobacter member (KRF1) from the globally important Methylobacter clade 2: cultivation and salient draft genome features. Antonie van Leeuwenhoek 112:1399-1408.
- Rahalkar MC, Khatri K, Mohite J, Pandit P, Bahulikar R (2020) A novel Type I methanotroph Methylolobus aquaticus gen. nov. sp. nov. isolated from a tropical wetland. Antonie van Leeuwenhoek 113:959-971.
- Rahalkar MC, Khatri K, Pandit P, Mohite J (2024) Polyphasic Characterization of Ca. Methylomicrobium oryzae: A Methanotroph Isolated from Rice Fields. Indian Journal of Microbiology.
- Scagliola M, Valentinuzzi F, Mimmo T, Cesco S, C C, Y P (2021) Bioinoculants as Promising Complement of Chemical Fertilizers for a More Sustainable Agricultural Practice. Front Sustain Food Syst 4:622169:1-12.
- Tallapragada P, Seshagiri S (2017) Application of Bioinoculants for Sustainable Agriculture. In: Kumar V, Kumar M, Sharma S, Prasad R (eds) Probiotics and Plant Health. Springer, Singapore, pp 473–495.
- The Fertilizer Association of India ND (2019) FAI Annual Seminar – 2019 on New Approach to Fertilizer Sector.
- Thomas L, Singh I (2019) Microbial Biofertilizers: Types and Applications. In: Giri B, Prasad R, Wu Q, Varma A (eds) Biofertilizers for Sustainable Agriculture and Environment Springer, pp 1-19.
- Wu L, Ma K, Lu Y (2009) Rice roots select for type I methanotrophs in rice field soil. Systematic and applied microbiology 32:421-428.
- Yuan J, Yuan Y, Zhu Y, Linkui Cao (2018) Effects of different fertilizers on methane emissions and methanogenic community structures in paddy rhizosphere soil. Science of The Total Environment 627:770-781.
- Yuanfeng Cai, Yan Zheng, Paul L. E. Bodelier, Conrad R, Jia Z (2016) Conventional methanotrophs are responsible for atmospheric methane oxidation in paddy soils. Nature Communications 11728.
Figure 1.
(a) Various stages of the field experiment conducted in Malegaon, off Lavasa road near Pune, in a farmer’s field (b) Transplantation of the treated plants in 4m x 3m plots in a flooded rice field in a randomized design was done (3rd July 2024).
Figure 1.
(a) Various stages of the field experiment conducted in Malegaon, off Lavasa road near Pune, in a farmer’s field (b) Transplantation of the treated plants in 4m x 3m plots in a flooded rice field in a randomized design was done (3rd July 2024).

Figure 2.
Early flowering and grain formation were observed in rice plants inoculated with Methylomonas Kb3. In the background, control plants can be seen (Observations taken on 19th September 2024, ~ 100 days after sowing).
Figure 2.
Early flowering and grain formation were observed in rice plants inoculated with Methylomonas Kb3. In the background, control plants can be seen (Observations taken on 19th September 2024, ~ 100 days after sowing).

Figure 3.
(a) Comparison of plant height, tiller number, panicle height and panicle number within treatments. (b) Comparison of grain yield in quintals/ha within treatments.
Figure 3.
(a) Comparison of plant height, tiller number, panicle height and panicle number within treatments. (b) Comparison of grain yield in quintals/ha within treatments.
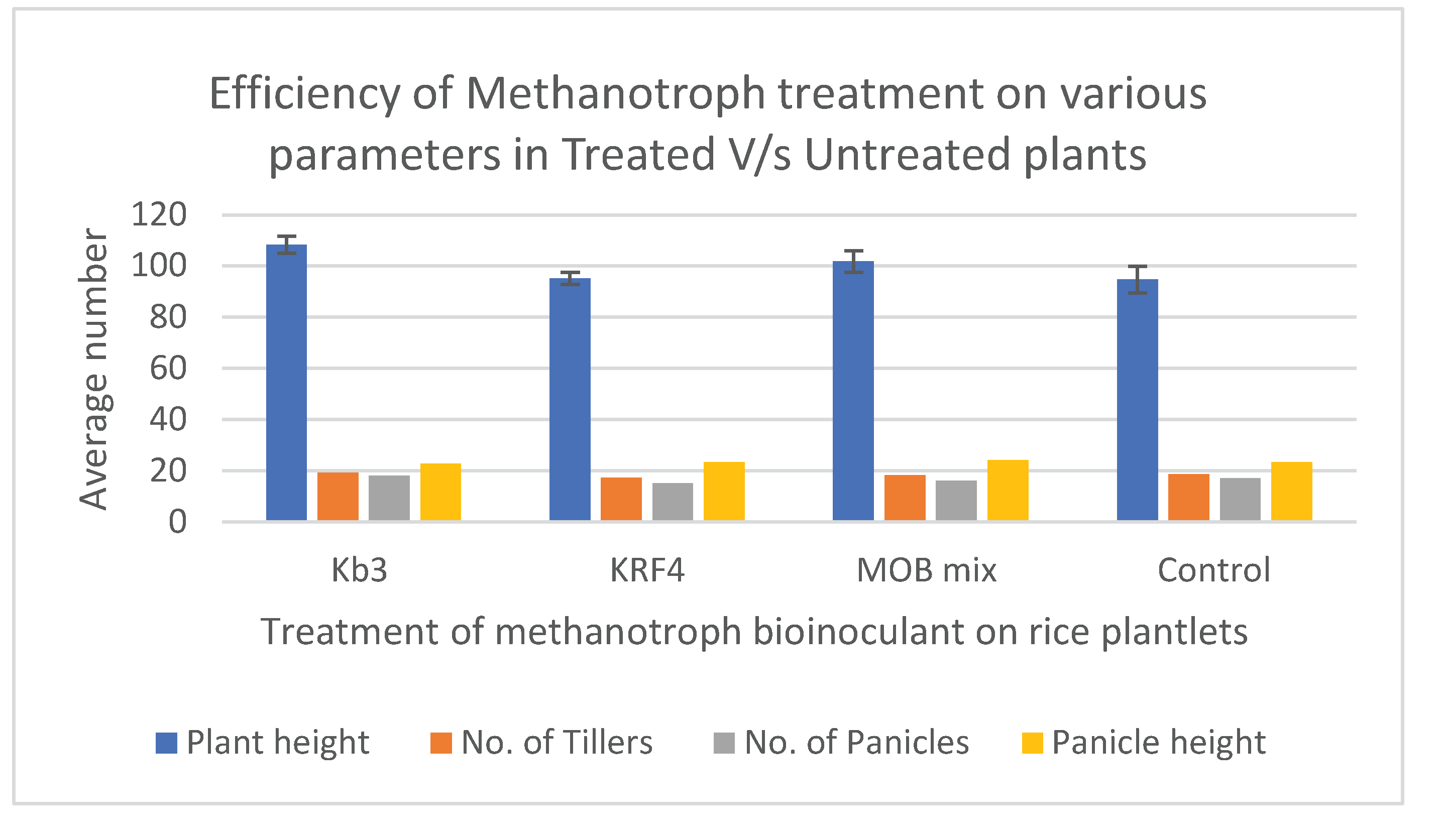
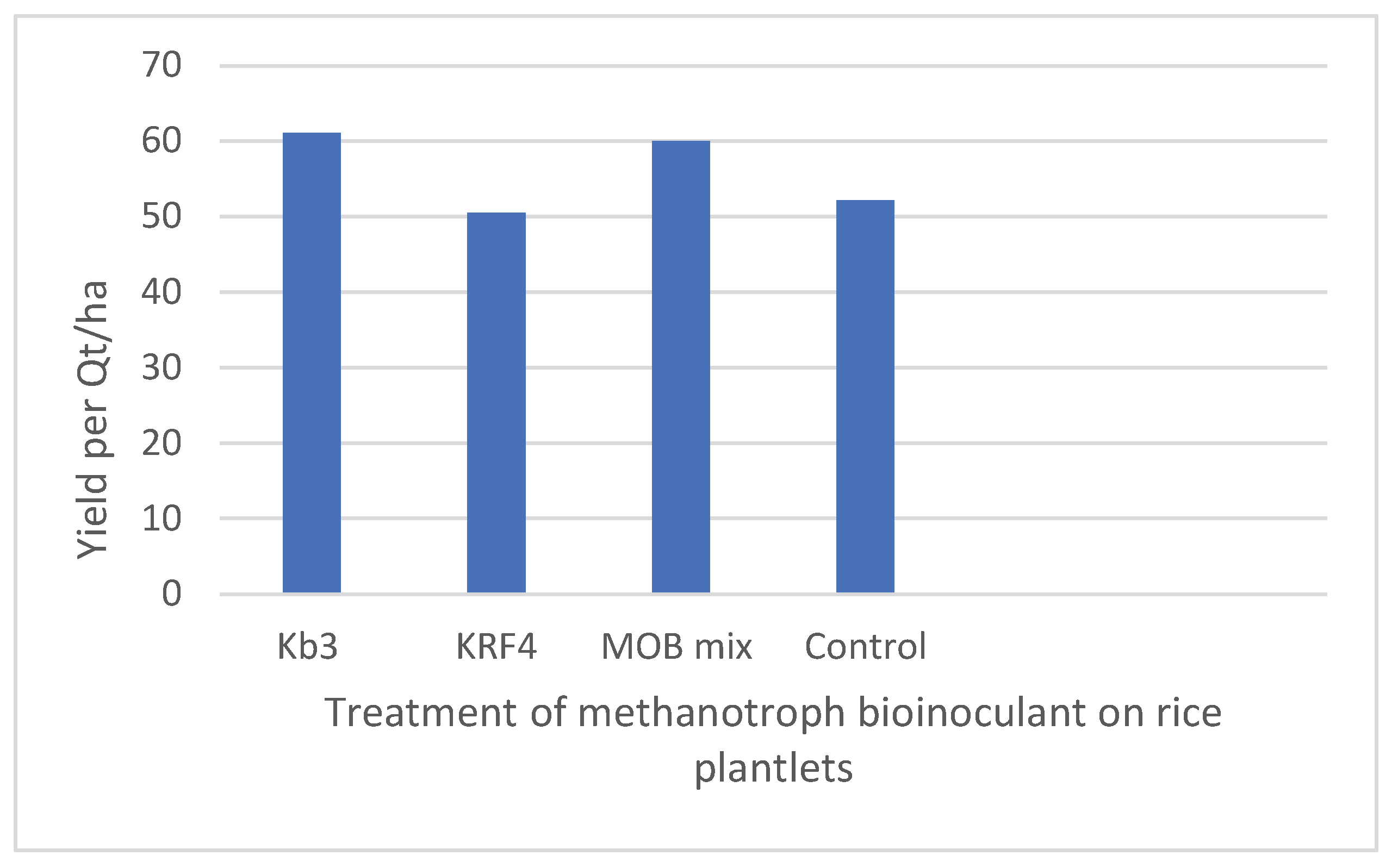
Figure 4.
Principal component analysis (PCA) based on the effect of inoculation of methanotrophs on plant growth parameters. The first two coordinates contribute to 90.6% variance.
Figure 4.
Principal component analysis (PCA) based on the effect of inoculation of methanotrophs on plant growth parameters. The first two coordinates contribute to 90.6% variance.
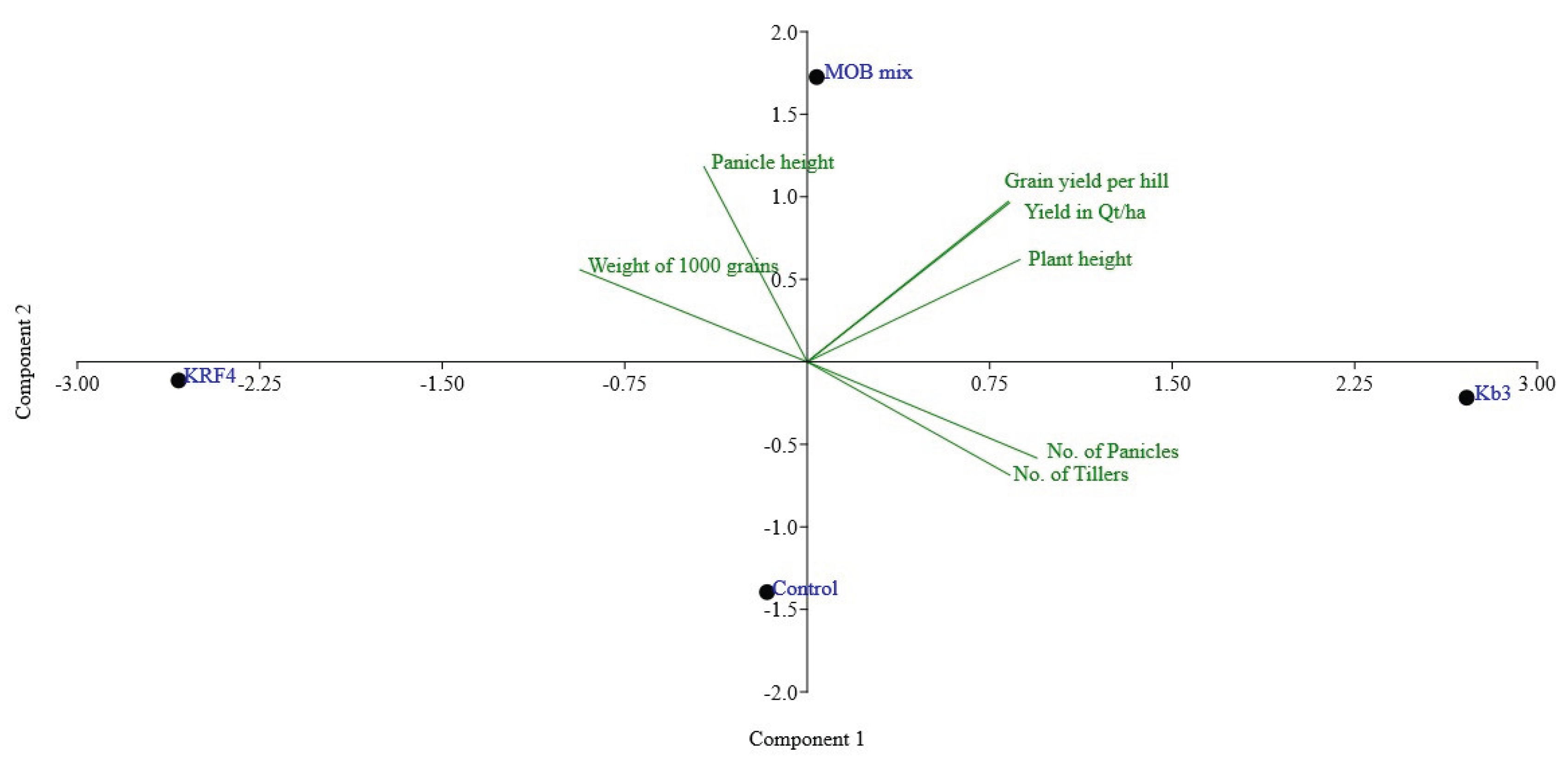

Table 1.
Enlisting of observed values in treated rice plants after harvest.
 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



