
Submitted:
14 May 2025
Posted:
14 May 2025
You are already at the latest version
Abstract
Wetlands are the most important natural sources of methane. Studies on the distribution and diversity of methanotrophs, especially in tropical wetlands, are limited. The studies on wetland methanotrophs help bridge the gap in the literature for understanding the community structure of methanotrophs in tropical wetlands. Our present study documents the methanotroph diversity from various wetland habitats across Western India. Samples from various sites such as freshwater ponds, lake sediments, mangroves, and small wetlands in stone quarries around multiple wetland ecosystems in Western India were collected and enriched for methanotroph isolation. As obtaining entirely pure cultures of methanotrophs is a tedious task, consuming months or years of streaking and re-streaking, methanotroph mono-cultures (a single methanotroph culture with a small number of heterotrophs) were established could be characterized using pmoA sequencing. Twenty-six mono-cultures were established, belonging to the genera Methylomonas, Methylocystis, Methylosinus, Methylocaldum, Methylocucumis, Methylomagnum, and Methylolobus genera. Eight methanotroph strains were purified in pure cultures- two Methylomonas koyamae, two Methylosinus sporium strains, one Methylolobus aquaticus, two Methylosinus trichosporium, and one strain of Methylomonas sp., which shows a close similarity with Methylomonas aureus and is possibly a novel species. A maximum number of cultures belonged to the Methylomonas and Methylocystis genera. Methylomagnum, a Type Ib methanotroph native to rice fields, was isolated from a pond in Pune. New members of Methylocucumis oryzae and Methylolobus aquaticus, two novel genera and species first reported by our lab from India, were also isolated in this study. Methanotrophs were high in most freshwater samples; in contrast, mangroves showed a relatively low abundance. Additionally, the cultivation approach helped us obtain new methanotrophs from this previously unexplored habitat, which can be used for further biotechnological and environmental applications.
Keywords:
Methanotrophs
; tropical
; wetlands
; pmoA gene
; Methylomonas
; Methylocystis
1. Introduction
Wetlands are the most important natural sources of atmospheric methane (CH4), contributing to a relatively high percentage of global emissions. Methane is produced in wetlands through the anaerobic microbial breakdown of organic matter. Methane emissions from wetlands account for 2 to 7% of net primary productivity [1]. Wetlands are essential to the terrestrial and aquatic ecosystems and contribute significantly to greenhouse gas emissions by releasing methane. These methane-rich zones shelter diverse aerobic methanotrophs, the only known biological filter for methane, thereby crucial in regulating the atmospheric methane flux. Wetlands are territories of marsh, fen, peat land, or water, either natural or manmade, permanent or temporary, with water that is static or flowing, fresh, brackish, or salty, and also includes portions of marine water whose depth at low tide does not exceed six meters [11]. As wetlands are the world’s most significant carbon sink and the primary natural source of methane, the type, area, distribution, timing, and amount of flooding of wetlands are critical factors to consider when calculating greenhouse gas emissions and carbon storage [11]. The high organic carbon concentration of the soil in wetlands makes them a significant contributor to the global carbon cycle despite making up only 5% of the planet’s land area [13]. Globally, ~20% of the methane emissions arise from wetlands. About 160 ± 40 Tg CH4 is contributed annually by natural wetlands, such as bogs, fens, flood plains, coastal zones of lakes, marshes, and swamps [19]. The majority of CH4 emissions to the atmosphere are released by tropical wetlands, which are followed by temperate and northern wetlands [19]. Oxic-anoxic conditions—typically created by the roots of wetland plants, allow aerobic and anaerobic microbial groups to coexist. Two microbial populations engaged in methane’s biogeochemical cycle in the soil are methanogens and methanotrophs. Obligate anaerobes produce methane called methanogens, which thrive in high-reduction, waterlogged conditions with low soil redox potential [8]. Aerobic and sometimes microaerophilic methanotrophs use methane, the second most significant greenhouse gas, as their only energy and carbon source [10]. Thus, methanotrophs are essential for reducing the methane produced by wetlands [5].
Methanotrophs, or methane-oxidizing bacteria, are a special class of methylotrophic bacteria that derive their energy and carbon solely from methane. They are Gram-negative, obligately aerobic, and ubiquitous to various habitats such as freshwater, sediments, and soils [14]. Most of the currently isolated methanotrophs thrive best at neutral pH at temperatures between 20 and 45◦C; however, several new methanotrophs have also lately been identified from harsh environments and continue to be discovered [7].
The limited understanding of methanotrophs from tropical wetlands increases the breadth of research on these habitats. The present study was undertaken to broaden the knowledge of methanotrophs from various wetland habitats in India, especially of the Western Indian region, some of which are included under the biodiversity hotspots of the Western Ghats.
2. Materials and Methods
All the samples were collected using gloves in sterile plastic vials or sterile plastic bags and proceeded for the enrichment of methanotrophs and oxidation of methane. Mud samples and water samples were collected in triplicates from the wetland site using sampling vials (50 ml capacity) from the littoral zones of the lakes or ponds or with about 10-15 cm water layer on the top (Figure 1).
Serial dilutions were set up using modified Nitrate Mineral Salts (NMS) medium as described earlier [16] from 10-1 to 10-12 by adding 1g of the sample to a 9 ml sterile (NMS) medium in 35 ml serum bottles [16]. Alternatively, microtiter plates (48 wells) were used for the dilution series for initial enrichments. All the enrichments were incubated at 28°C with methane and air in the ratio of 80:20 as headspace gas [17]. Gas Chromatography was done by injecting the headspace gas into the Chemito 8610 Gas Chromatography machine equipped with a flame ionisation detector (FID). All positive enrichments were streaked on NMS agarose medium plates to obtain cultures. The last positive dilution was noted in each sample. After growth in a methane and an air environment, colonies were obtained. Single colonies were picked up and streaked on fresh plates until a single morphotype dominated, confirmed by phase contrast microscopy (using Nikon 80i, Japan microscope with a camera) of the cells under an oil immersion lens (100x magnification). Axenic cultures were obtained by repeated streaking to eliminate heterotrophic contaminants, and the purity was confirmed by microscopy and growth on nutrient agar plates. Heterotrophic contaminants often accompany methanotrophs, and the cultures that remained non-axenic (having small numbers of heterotrophs) even after 5-6 purifications on agarose were termed methanotroph mono-cultures. Pure cultures showed no contamination on microscopic observations or growth on nutrient agar plates.
DNA was extracted from methanotroph mono-cultures (22) as well as from the four pure cultures and subjected to pmoA gene amplification and sequencing using the forward primer A189 as described [17]. Blast analyses were performed, and the cultures were classified into respective genera. An amino acid-based phylogenetic tree using the functional gene pmoA and universal 16S rRNA was constructed using MEGA XI [21]. In the case of pure cultures, the 16S rRNA gene was amplified and sequenced as described. All the sequences were submitted to NCBI, and accession numbers were obtained.
3. Results
Samples were collected from various sites representing wetland habitats such as pond sediments, stone quarries, lake sediments (freshwater), and a few samples were from mangroves (marshy sediments) (Figure 1). Enrichments that showed a decline in methane accompanied by visual growth in terms of turbidity, surface pellicle, or biofilm growth at the bottom were indicative of positive Fenrichments. The last positive serial dilution for each enrichment was noted. Methanotrophs grew in the collected samples in various abundance values, ranging from 10-2 to 10-12 (Table 1). It was seen that most of the pond types of freshwater and shallow habitats showed the presence of methanotrophs in relatively high numbers, reaching up to 10-12 cells/g of soil. However, in the case of mangrove samples, the highest dilution reached was 10-3 or 10-4, indicating a low abundance of methanotrophs.
Serial dilution enrichment followed by isolation on agarose plates in the presence of methane and air environment resulted in the cultivation of methanotrophs from various groups: Type Ia, Type Ib, and Type II. Our goal was to use a cultivation approach to document the biodiversity of methanotrophs and to expand the present knowledge of cultivable methanotrophs from this environment. In this study, 28 methanotrophs were cultured and were composed of mono-cultures of methanotrophs (single dominant methanotrophs). A clear pmoA sequence was obtained for all of the 26 cultures, and their next neighbors were determined by NCBI blast analyses (Table 1). The pmoA gene sequencing helped assign the methanotrophs to their corresponding genera and species [7,12]. Amongst these, eight axenic cultures were identified as Methylomonas fluvii, Methylomonas sp., Methylolobus aquaticus, Methylosinus sporium, and Methylosinus trichosporium respectively, after 16S rRNA sequencing (Table 2).
Most of the cultures were found to of the genera Methylocystis and Methylomonas, as seen after morphological and pmoA blast analyses. Methylocystis genus represents Type II methanotrophs, which are usually small in size (~1um diameter) and coccoid in shape (Figure 2). Out of the 26 cultures, six cultures of methanotrophs, namely: [strain name (pmoA accession number)] VLS12 (PQ821915), VLS6, VLW4 (PQ821917), MB5 (PQ821931), MG3 (PQ821909), and MMB (PQ821921) (Figure 2), isolated from freshwater, soil, mud and mangroves (Table 1) belong to the Methylocystis genera and show ~97% nucleotide pmoA gene similarity with the pmoA sequence of Methylocystis hirsuta CSC1T (Figure 3) (Table 1). Nine cultures were from the Methylomonas genera (Table 1). They were seen as thick and short rods in microscopic analyses (Figure 2). Among these, four cultures, i.e., ASQA (PQ821920), MSA (PQ821928), TM3 (PQ821925), MgM2 (PQ821911) (Figure 2), and AL2 (PQ821908), showed 96.12, 95.81, 92.47, 91.88, and 87.79 % pmoA gene similarity with Methylomonas koyamae Fw 12E-YT (Figure 3) (Table 1). The culture MgM2 shows 96.70% 16SrRNA gene similarity with Methylomonas aurea strain SURF-1T (Figure 4) (Table 2). Two cultures of Methylomonas, i.e., VUS3 (PQ821926) and TS2 (PQ821913), showed 95.19% pmoA and 99.41% 16SrRNA gene similarity with Methylomonas fluvii EbBT (Figure 3) (Figure 4) (Table 1) (Table 2). One strain, AL2B (PQ821907), showed 89.56% pmoA gene similarity with Methylomonas montana MW1T, and the strain PgA6 (PQ821927), showed 94.82% pmoA gene similarity with M. denitrificans FJG1T (Figure 3) (Table 1). Methylomonas species can be found in freshwater lake and river silt, wetland muds, activated sludge and wastewater, groundwater, and coal mine drainage water [3]. Three cultures belonged to the Type Ia methanotroph genus Methylocaldum. All Methylocaldum cultures were cultivated from mangrove regions (Figure 1). These mangroves are in saline and tropical regions from Konkan and Mumbai. Methylocaldum genus has been detected earlier in India from cow dung, compost, and biogas slurry samples [15]. Three cultures isolated from mangrove samples, i.e., MgM4 (PQ821912), MgN2 (PQ821930), and MgD2 (PQ821910) (Figure 2), show 99.18, 99.52, and 98.95% pmoA gene similarity with Methylocaldum gracile VKL-14LT (Figure 3) (Table 1). The members of the genus Methylocaldum are thermotolerant methanotrophs and are detected in diverse environments, including marine and aquatic habitats, upland soils, rice fields, and landfills [18]. Our previously published study isolated Methylocaldum gracile strain RS9 from biogas slurry [15]. Six cultures of the genera Methylosinus have been isolated in the study; four cultures belong to the species Methylosinus sporium, and two culture belongs to M. trichosporium. The cultures VLS4 (PQ821914), AS1B (PQ821919), and AW2A (PQ821929) (Figure 2) show 95.06, 99.31, and 95.05% pmoA gene similarity with M. sporium ATCC 35069T, and the culture PLW2 (PQ821916) shows 100% pmoA gene similarity with M. trichosporium OB3bT (Figure 3) (Table 1). The cultures VLS4 and AW1A show 98.96% and 98.89% 16SrRNA gene similarity with Methylosinus sporium NCIMB 11126T, and the cultures PLW2 and AW2B show 99.93% and 100% 16SrRNA gene similarity with Methylosinus trichosporium OB3bT (Figure 4) (Table 2). The cultures of the genera Methylomagnum and Methylocucumis, having a large size >5 µm, were also cultured in this study. The culture MSBM (PQ821923) (Figure 2) was isolated from a seaweed sample from a wetland patch on a hill near Mahatma Society, Pune (termed Mahatma Hill). This culture showed 99.53% pmoA gene similarity with Methylomagnum ishizawai RS11DT (Figure 4) (Table 1). Two cultures, MSBC (PQ821922) (Figure 2) and MWC (PQ821924), show 98.66% and 98.61% pmoA gene similarity with Methylocucumis oryzae Sn 10-6T (Figure 3) (Table 1). These cultures were obtained from seaweed and water samples from a wetland patch near Mahatma Society, Pune (Mahatma Hill). The culture isolated from freshwater samples from Pashan Lake Pune, i.e., PLW4 (PQ821918) (Figure 2) (Figure 3) (Figure 4), shows 97.25% pmoA and 99.35% gene similarity with Methylolobus aquaticus FWC3T (Table 1) (Table2). PLW4 would be the second culture obtained from India from this newly described genus and species, Methylolobus aquaticus. Currently, only two strains have been described from this genus; one is FWC3T, the type strain described in our study [17], and the other is from the Netherlands, earlier described as Methylotetracoccus oryzae C50C1 [9]. Thus, this study documented the cultivation of methanotrophs from two novel and newly described genera from India, Methylocucumis and Methylolobus.
4. Discussion
Varied types of wetlands were covered in this study, which include mangroves, hilltop stone quarries, small ponds with aquatic plants, etc. The geographical area covered spans Western India and mainly the Western Ghats, one of India’s two biodiversity hotspots. The Western Ghats, also known as the Sahyadris, is a mountain range that stretches 1,600 km (990 mi) along the western coast of India.
Most isolated cultures were observed to belong to the genera Methylocystis and Methylomonas. Methylomonas species can be found in freshwater lake and river silt, wetland muds, activated sludge and wastewater, groundwater, and coal mine drainage water [4]. One of the most ecologically significant methanotroph populations in terrestrial settings is the Methylocystis species. They live in various environments, including freshwater, rice paddies, peatlands, and landfills [2,4]. Groundwater and soil freshwater sediments are important ecosystems for Methylosinus species [4]. Among the unique methanotrophs, strains from the newly described genera- Methylocucumis and Methylolobus were also isolated. Methylocucumis oryzae, the newly described genus isolated from a rice field. Has been reported by our research group and has been exclusively isolated from the Western Ghat regions of India. This particular methanotroph is relatively large in size and oblong-shaped, isolated frequently from stone quarries in wetland patches of Pune city (Ref: International Microbiology). Similarly, in this study, we could also isolate the strain Methylolobus aquaticus, a newly described genus and species, isolated from a wetland in the state of Maharashtra [17]. Thus, our technique helped isolate members from novel genera. It is also noteworthy to report the isolation of another member of Methylomagnum, a genus first described in rice fields. Three of the cultures belonged to Methylocaldum, all isolated from mangrove regions. These mangroves are basically in Konkan and Mumbai’s hot and moist regions. The habitat preference of Methylocaldum has been detected in cow dung,
Wetlands are often subjected to drying and exposure to sunlight and light-related damage. Therefore, many isolates belonging to the Type I methanotrophs were found to have colors like pink and red, mostly related to the carotenoid pigments. Carotenoids are known to have a protective role in photoprotection [6]. Pink was also seen in Methylocystis cultures, as found in Methylocystis rosea, which shows pink coloration [20]. Additionally, a few of the Methylocystis cultures isolated from Indian rice landraces also show pink coloration [16]. The isolated methanotrophs can be used for various mitigation and value-adding applications, including serving as models for research on methanotroph-based methane mitigation from wetland habitats.
5. Conclusion
The current study reports the diversity, abundance, and community structure of methanotrophs from tropical wetlands spanning the strip of Western India, which falls under the Western Ghats and is categorized as one of the two biodiversity hotspots in the country. Cultures from seven major genera of methanotrophs were isolated in the study, amongst which Methylomonas and Methylocystis, the two prominent genera representing Type I and Type II methanotrophs respectively, were seen to dominate the wetland community structure of various habitats. Methylocaldum, Methylosinus, Methylocucumis, Methylomagnum and Methylolobus were a few other genera isolated in the study. The abundance of methanotrophs was relatively low in mangrove soils, suggesting that vegetation, salinity, and other abiotic factors influence the growth of methanotrophs. The presented research attempts to bridge the gap in knowledge on cultivable methanotrophs from tropical wetlands, which are reported to be high-affinity methanotrophs. Further investigations can reveal the range of affinity of these methanotrophs to methane to be applied for methane mitigation strategies for methane emitted from wetlands.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, Rahul Bahulikar and Monali Rahalkar; Methodology, Kajal Pardhi, Rahul Bahulikar, Yukta Patil, and Yash Kadam; Writing – original draft, Kajal Pardhi, Shubha Manvi, and Monali Rahalkar; Writing – review & editing, Monali Rahalkar. MCR designed the study, collected samples, procured funds, edited the manuscript, and monitored the study. KP and SM performed the enrichments and cultivation of methanotrophs. SK, CS, YK, and YP assisted in enriching and culturing methanotrophs. RB performed the sample collection and prepared the phylogenetic tree. KP, SM, and MCR wrote the manuscript, and all the authors edited and approved the final version.
Funding
MCR acknowledges ANRF (SPF/2022/000045)] for providing the funds. KP acknowledges UGC for providing her with a junior research fellowship.
Data Availability
All the sequence data are available in the NCBI database. There was no involvement of humans and/or animals in this study. Hence, the following declarations are not applicable.
Acknowledgments
We thank Dr. Mahesh Shindikar, COEP, and Dr. Sagar Pandit, IISER, for collecting the mangrove samples from Mumbai.
Competing Interests
The authors have no relevant financial or non-financial interests to disclose.
Ethics approval:
Not applicable
Consent to participate:
Not applicable
Consent to publish:
Not applicable
References
- Aselmann I, Crutzen PJ (1989) Global distribution of natural freshwater wetlands and rice paddies, their net primary productivity, seasonality and possible methane emissions. Journal of Atmospheric Chemistry 8:307-358.
- Belova SE, Kulichevskaya IS, Bodelier PL, Dedysh SN (2013) Methylocystis bryophila sp. nov., a facultatively methanotrophic bacterium from acidic Sphagnum peat, and emended description of the genus Methylocystis (ex Whittenbury et al. 1970) Bowman et al. 1993. International journal of systematic and evolutionary microbiology 63:1096-1104.
- Bowman J (2016) Methylococcaceae. In: Whitman W (ed) Bergey’s Manual of Systematics of Archaea and Bacteria. John Wliey and Sons, USA.
- Bowman JS, Ducklow HW (2015) Microbial communities can be described by metabolic structure: a general framework and application to a seasonally variable, depth-stratified microbial community from the coastal West Antarctic Peninsula. PloS One 10.
- Conrad R (2009) The global methane cycle: recent advances in understanding the microbial processes involved. Environmental Microbiology Reports 1:285-292. [CrossRef]
- Dall’Osto L, Bassi R, Ruban A (2014) Photoprotective Mechanisms: Carotenoids. In: S.M. Theg, Wollman FA (eds) Plastid Biology, Advances in Plant Biology. Springer, New York.
- Dedysh SN, Knief C (2018) Diversity and phylogeny of described aerobic methanotrophs. In: Kalyuzhnaya MG, Xing X-H (eds) Methane Biocatalysis: Paving the way to sustainability. Springer, pp 17-42.
- Fazli P, Man HC, Shah UKM, Idris A (2013) Characteristics of methanogens and methanotrophs in rice fields: a review. Asia-Pacific Journal of Molecular Biology and Biotechnology 21:3-17.
- Ghashghavi M, Belova SE, Bodelier PLE, Dedysh SN, Kox MAR, Speth DR, Frenzel P, Jetten MSM, Lücker S, Lüke C (2019) Methylotetracoccus oryzae Strain C50C1 Is a Novel Type Ib Gammaproteobacterial Methanotroph Adapted to Freshwater Environments. mSphere 4:e00631-00618. [CrossRef]
- Hanson RS, Hanson TE (1996) Methanotrophic bacteria. Microbiology and Molecular Biology Reviews 60:439-471.
- Hu S, Niu Z, Chen Y (2017) Global Wetland Datasets: a Review. Wetlands 37:807-817.
- Knief C (2015) Diversity and habitat preferences of cultivated and uncultivated aerobic methanotrophic bacteria evaluated based on pmoA as molecular marker. Frontiers in Microbiology 6. [CrossRef]
- Lugo AE, Brown S, Brinson MM (1990) Concepts in wetland ecology. Ecosystems of the world, pp 53-85.
- McDonald IR, Bodrossy L, Chen Y, Murell CJ (2008) Molecular ecology techniques for the study of aerobic methanotrophs. Appl Environ Microbiol 74:1305-1315.
- Mohite JA, Manvi SS, Pardhi K, Khatri K, Bahulikar RA, Rahalkar MC (2023) Thermotolerant methanotrophs belonging to the Methylocaldum genus dominate the methanotroph communities in biogas slurry and cattle dung: A culture-based study from India. Environmental Research 228:1-6. [CrossRef]
- Rahalkar MC, Patil S, Dhakephalkar PK, Bahulikar RA (2018) Cultivated methanotrophs associated with rhizospheres of traditional rice landraces from Western India belong to Methylocaldum and Methylocystis. 3 Biotech 8. [CrossRef]
- Rahalkar MC, Khatri K, Mohite J, Pandit P, Bahulikar R (2020) A novel Type I methanotroph Methylolobus aquaticus gen. nov. sp. nov. isolated from a tropical wetland. Antonie van Leeuwenhoek 113:959-971.
- Takeuchi M, Kamagata Y, Oshima K, Hanada S, Tamaki H, Marumo K, Sakata S (2014) Methylocaldum marinum sp. nov., a thermotolerant, methane-oxidizing bacterium isolated from marine sediments, and emended description of the genus Methylocaldum. International Journal of systematic and evolutionary microbiology 64:3240-3246.
- Wang Z, Zeng D, Patrick WH (1996) Methane emissions from natural wetlands. Environmental Monitoring and Assessment 42:143-161.
- Wartiainen I, Hestnes AG, McDonald IR, Svenning MM (2006) Methylocystis rosea sp. nov., a novel methanotrophic bacterium from Arctic wetland soil, Svalbard, Norway (78° N) Free. International Journal of systematic and evolutionary microbiology 56:541-547.
Figure 1.
Sampling sites of different wetland patches visited for the study. A. Mahatma Hill in winter B. Mahatma Hill in Summer C. Venna Lake D. Vettal Hills E. Diveagar mangroves F. Pashan Lake.
Figure 1.
Sampling sites of different wetland patches visited for the study. A. Mahatma Hill in winter B. Mahatma Hill in Summer C. Venna Lake D. Vettal Hills E. Diveagar mangroves F. Pashan Lake.

Figure 2.
Morphology of the isolated cultures as seen under a phase-contrast microscope (Nikon 80i, Japan microscope with a camera) under 100X magnification with oil emulsion A, Methylocystis hirsuta culture VLS12; B, Methylocystis hirsuta VLS6; C, Methylosinus sporium strain VLS4; D, Methylocystis hirsuta MG3; E, Methylomonas fluvii strain TS2; F, Methylomonas koyamae culture MgM2; G, Methylocucumis oryzae MSBC; H, Methylomangnum ishizawai culture MSBM; I, Methylolobus aquaticus culture PLW4; J, Methylocaldum gracile culture MgM4; K, Methyloaldum gracile culture MgD2; L, Methylocaldum gracile culture MgN2.
Figure 2.
Morphology of the isolated cultures as seen under a phase-contrast microscope (Nikon 80i, Japan microscope with a camera) under 100X magnification with oil emulsion A, Methylocystis hirsuta culture VLS12; B, Methylocystis hirsuta VLS6; C, Methylosinus sporium strain VLS4; D, Methylocystis hirsuta MG3; E, Methylomonas fluvii strain TS2; F, Methylomonas koyamae culture MgM2; G, Methylocucumis oryzae MSBC; H, Methylomangnum ishizawai culture MSBM; I, Methylolobus aquaticus culture PLW4; J, Methylocaldum gracile culture MgM4; K, Methyloaldum gracile culture MgD2; L, Methylocaldum gracile culture MgN2.
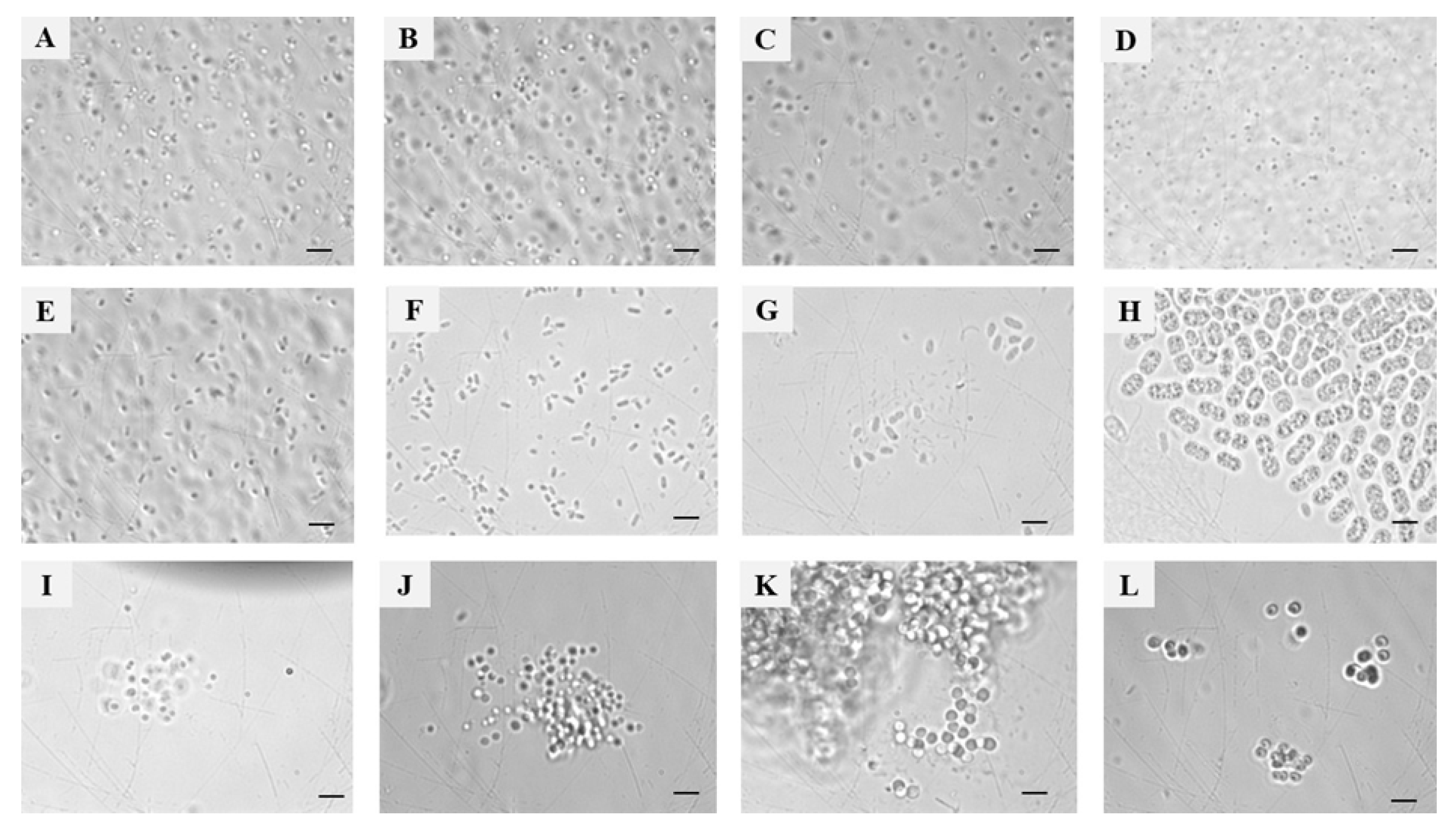
Figure 3.
Amino acid-based phylogenetic tree based on pmoA gene of isolated methanotrophs with its closest members. The phylogenetic tree was constructed using the partial pmoA sequence of the isolated methanotrophs in comparison with type cultures using MEGA XI software. It was inferred by the Maximum Likelihood method and the Tamura-Nei model. The bar shows a 5% divergence.
Figure 3.
Amino acid-based phylogenetic tree based on pmoA gene of isolated methanotrophs with its closest members. The phylogenetic tree was constructed using the partial pmoA sequence of the isolated methanotrophs in comparison with type cultures using MEGA XI software. It was inferred by the Maximum Likelihood method and the Tamura-Nei model. The bar shows a 5% divergence.
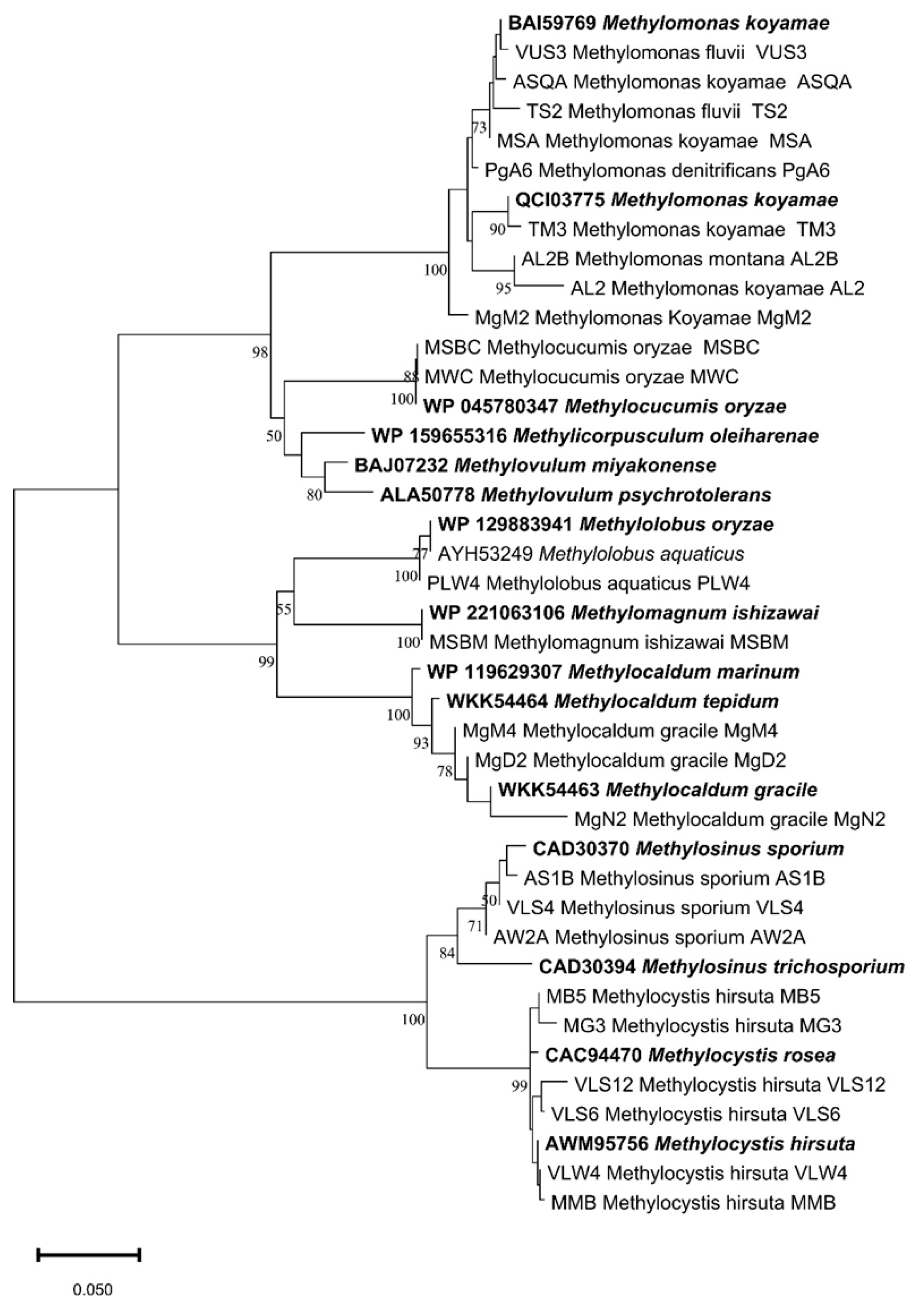
Figure 4.
Maximum-likelihood 1,000 bootstrap tree of 16S rRNA-based phylogenetic tree of pure methanotrophic strains (showed in bold) with their closest members. The evolutionary history was inferred by using the maximum-likelihood method and the Tamura–Nei model. Evolutionary analyses were conducted in MEGA XI. The bar represents 5% divergence.
Figure 4.
Maximum-likelihood 1,000 bootstrap tree of 16S rRNA-based phylogenetic tree of pure methanotrophic strains (showed in bold) with their closest members. The evolutionary history was inferred by using the maximum-likelihood method and the Tamura–Nei model. Evolutionary analyses were conducted in MEGA XI. The bar represents 5% divergence.
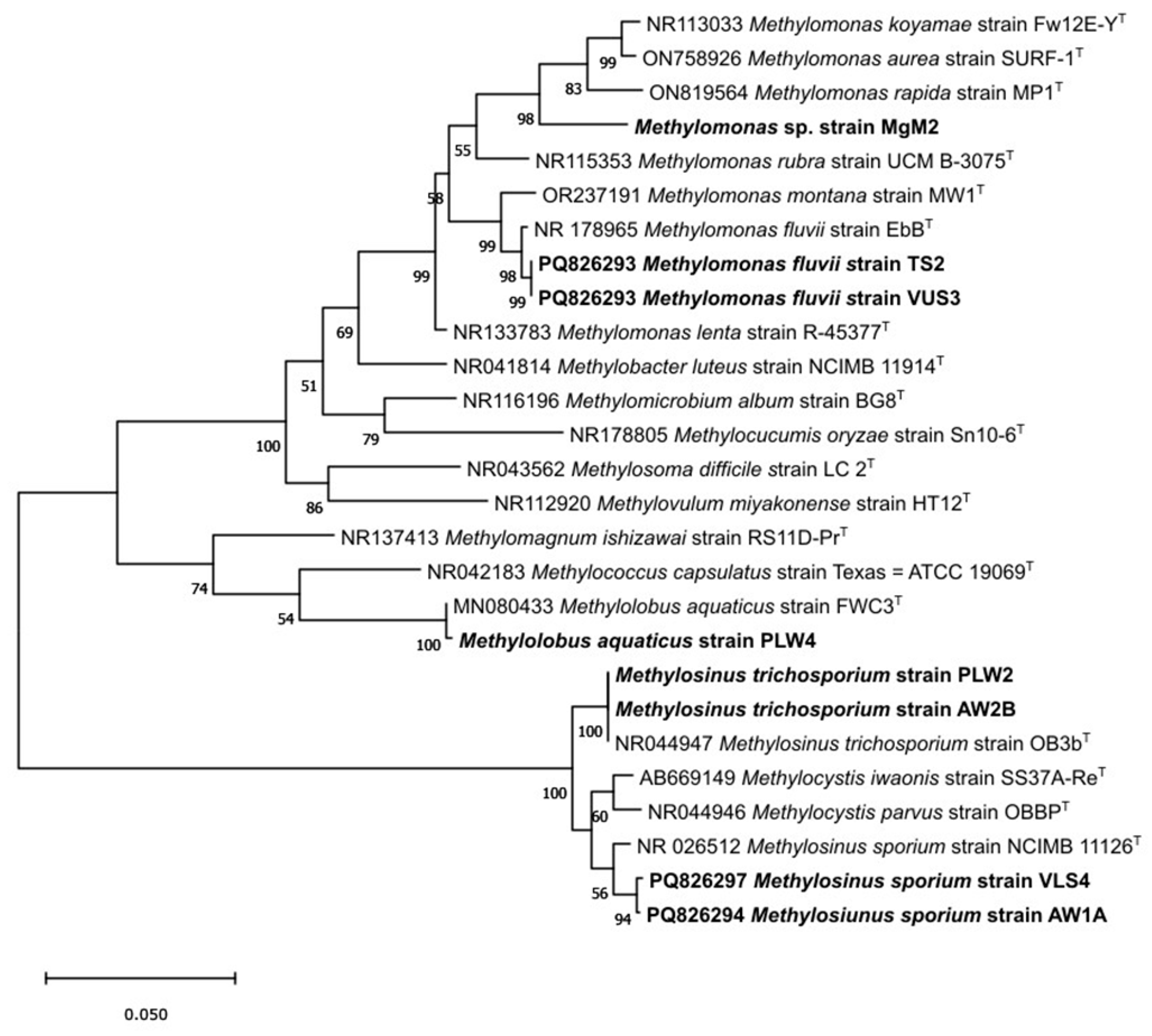

Table 1.
Tabular summary describing the pmoA identification of the representative sample isolated from the respective collection site with their names, dates and dilution.
Table 1.
Tabular summary describing the pmoA identification of the representative sample isolated from the respective collection site with their names, dates and dilution.
| Sampling details | Dilution used for isolation of methanotrophs | Representative cultures | Identification using pmoA gene | |||||
| Sampling site | Sample name | Sampling date | Strain name | GeneBank accession number | Nearest match (with type cultures) | % similarity (nucleotide) | % similarity (protein) |
|
|
Venna Lake Sediments |
soil sample | 28.12.2022 | 10-12 | VLS12 | PQ821915 |
Methylocystis hirsuta CSC1 |
97.25 | 98.60 |
| soil sample | 28.12.2022 | 10-6 | VLS6 | - | Methylocystis hirsuta CSC1 | 97.70 | 98.56 | |
| soil sample |
28.12.2022 | 10-4 | VLS4 | PQ821914 |
Methylosinus sporium ATCC |
95.06 | 98.55 | |
| soil sample | 28.12.2022 | 10-3 | VUS3 | PQ821926 | Methylomonas fluvii EbB | 95.19 | 100 | |
| water sample | 28.12.2022 | 10-4 | VLW4 | PQ821917 | Methylocystis hirsuta strain CSC1 | 97.35 | 97.93 | |
| soil sample | 28.12.2022 | 10-5 | MB5 | PQ821931 | Methylocystis hirsuta strain CSC1 | 97.12 | 98.66 | |
| Vetal hill Pond | Stone Sample |
15-01-2024 | 10-2 | AS1B | PQ821919 | Methylosinus sporium strain ATCC 35069 | 99.31 | 98.61 |
| water sample | 15-01-2024 | 10-8 | AW2A | PQ821929 | Methylosinus sporium strain ATCC 35069 | 95.05 | 97.93 | |
| water sample | 04-02-2024 | 10-2 | ASQA | PQ821920 | Methylomonas koyamae strain Fw12E-Y | 96.12 | 99.31 | |
| Mahatma hill pond | Seaweed Sample |
21-04-2024 | 10-2 | MSBM | PQ821923 | Methylomagnum ishizawai strain RS11D | 99.53 | 99.29 |
| Seaweed sample |
21-04-2024 | 10-6 | MSA | PQ821928 | Methylomonas koyamae strain Fw12E-Y | 95.81 | 100 | |
| Seaweed sample |
21-04-2024 | 10-8 | MSBC | PQ821922 | Methylocucumis oryzae strain Sn 10-6 | 98.66 | 100 | |
| water sample | 21-04-2024 | 10-7 | MWC | PQ821924 | Methylocucumis oryzae strain Sn 10-6 | 98.61 | 100 | |
| Mud sample |
18-12-2023 | 10-3 | TM3 | PQ821925 | Methylomonas koyamae strain Fw12E-Y | 92.47 | 97.18 | |
| Mud sample |
21-04-2024 | 10-5 | MMB | PQ821921 | Methylocystis hirsuta strain CSC1 | 97.46 | 98.62 | |
| Pashan Lake | water sample | 08.07.2023 | 10-2 | PLW2 | PQ821916 | Methylosinus trichosporium strain OB3b | 100 | 99.31 |
| water sample | 08.07.2023 | 10-4 | PLW4 | PQ821918 | Methylolobus aquaticus strain FWC3 | 97.25 | 100 | |
| Tamhini river | sediment | 16-06-2023 | 10-2 | TS2 | PQ821913 | Methylomonas fluvii strain EbB | 95.19 | 98.53 |
| ARI pond | Lotus root sample |
02-01-2024 | 10-2 | AL2 | PQ821908 | Methylomonas koyamae strain Fw12E-Y | 87.79 | 95.16 |
| Lotus root sample |
02-01-2024 | 10-2 | AL2B | PQ821907 | Methylomonas montana strain MW1 | 89.56 | 97.90 | |
|
Paragrass BAIF pond |
Root sample | 09-01-2024 | 10-6 | PgA6 | PQ821927 | Methylomonas denitrificans strain FJG1 | 94.82 | 98.03 |
|
Mumbai Mangroves |
soil sample | 21-09-2023 | 10-2 | MgM2 | PQ821911 | Methylomonas koyamae strain Fw12E-Y | 91.88 | 97.14 |
| soil sample | 21-09-2023 | 10-4 | MgM4 | PQ821912 | Methylocaldum gracile strain VKM-14L | 99.18 | 100 | |
| Alibag mangroves | soil sample | 23-03-2023 | 10-3 | MG3 | PQ821909 | Methylocystis hirsuta strain CSC1 | 97.50 | 99.29 |
| soil sample | 23-03-2023 | 10-3 | MgN2 | PQ821930 | Methylocaldum gracile strain VKM-14L | 99.52 | 100 | |
| Diveagar mangroves | soil sample | 29-12-2023 | 10-2 | MgD2 | PQ821910 | Methylocaldum gracile strain VKM-14L | 98.95 | 100 |
Table 2.
Taxonomy of the methanotroph cultures identified using 16S rRNA gene sequencing with details on their sampling sites, dilution and representative strain.
Table 2.
Taxonomy of the methanotroph cultures identified using 16S rRNA gene sequencing with details on their sampling sites, dilution and representative strain.
| Sampling details | Dilution used for isolation of methanotrophs | Representative strain | Identification using 16SrRNA gene | ||||
| Sampling site | Sample name | Sampling date | Strain name | Gene accession number | Nearest match with type strain | % similarity | |
| Vetal hill pond | Water sample | 16-01-2023 | 10-8 | AW1A | PQ826297 |
Methylomonas sporium strain NCIMB 11126 | 98.89 |
| Tamhini river | sediment | 16-06-2023 | 10-2 | TS2 | PQ826293 |
Methylomonas fluvii strain EbB | 99.41 |
| Venna lake | Soil sample | 03-01-23 | 10-3 | VUS3 | PQ826293 |
Methylomonas fluvii strain EbB | 99.41 |
| Venna Lake | Soil sample | 03-01-2023 | 10-4 | VLS4 | PQ826294 |
Methylomonas sporium strain NCIMB 11126 | 98.96 |
| Pashan Lake | water sample | 08.07.2023 | 10-2 | PLW2 | - | Methylosinus trichosporium strain OB3b | 99.93 |
| Pashan Lake | water sample | 08.07.2023 | 10-4 | PLW4 | - | Methylolobus aquaticus strain FWC3 | 99.43 |
|
Mumbai Mangroves |
soil sample | 21-09-2023 | 10-2 | MgM2 | PV630802 | Methylomonas aurea strain SURF-1 | 96.70 |
| Vetal hill Pond | Water Sample |
15-01-2024 | 10-8 | AW2B | - | Methylosinus trichosporium strain OB3b | 100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



