
Submitted:
29 December 2024
Posted:
03 January 2025
You are already at the latest version
Abstract
Many nitrogen-fixing bacteria (NFB) can produce siderophores for iron acquisition in soil, but the impact of siderophore producing capabilities of NFB on the rhizosphere soil microecology is not well understood. To explore the effects of root inoculation with NFB strains with different siderophore producing capabilities on the rhizosphere soil microecology, the wild-type (WT) NFB strain Kosakonia radicincitans GXGL-4A and its Tn5 mutants M107 (high siderophore-producing ability) and M246-2 (deficient in siderophore production) were used as biofertilizers in cucumber rhizosphere soil. Iron is important for the growth of NFB bacterial cells, and the mutant M246-2 showed the slowest growth rate compared to the other strains when incubated in A15 nitrogen-free medium supplied with different levels of iron. The mutant M107 had the strongest chelating ability for iron, with the largest yellow halo on the CAS detection plate. There were statistically significant differences in the halo diameters among the three NFB groups. Compared with the control group, the application of NFB significantly increased the activities of soil peroxidase (S-POD) and dehydrogenase (S-DHA) and altered the soil nitrogen contents. Fertilization with the mutant M107 significantly improved cucumber biomass and reduced the abundance and diversity of bacterial communities in the rhizosphere soil compared to the other groups. The contents of soil ammonium nitrogen (NH4+−N) and total nitrogen (TN), and soil dehydrogenase (S-DHA) showed significant correlations with the abundance of most Top 50 dominant genera in the soil. Soil TN content was the essential factor affecting the abundance of Kosakonia bacteria in cucumber rhizosphere.
Keywords:
Siderophore-producing capability
; microbial diversity
; soil enzyme
; soil nitrogen content
; nitrogen-fixing bacterium GXGL-4A
; Tn5 mutants
1. Introduction
Iron is an essential nutritional element for the growth of plants and microorganisms, and it plays an important role in environmental biogeochemistry. However, trivalent iron often exists in the form of insoluble oxides or hydroxides in the environment, which greatly limits the biological absorption and utilization of iron. Siderophores are low molecular weight organic compounds produced by microorganisms and some crops under low iron stress conditions, which can efficiently bind trivalent iron ions to form Fe-chelating complexes. Root plasma membrane transporter proteins help absorb Fe siderophore complexes, and then the complexes travel to the flowering organs of plants [1]. The application of bacterial siderophores has been proven to improve nutrient absorption, leaf physiochemical characteristics and crop yield, and a variety of siderophore-producing rhizobacteria have been used for microbial biofortification of iron in plants under adverse conditions including saline-alkali and drought [2,3,4,5]. A variety of siderophore producing bacteria have been isolated and their effects on plant iron absorption and growth have been explored [6,7,8]. Recent study has shown that plants inoculated with siderophore producing bacteria have enhanced the uptake of nitrogen, phosphorous, Na+ ions, and chlorophyll content of crop plants. In addition, the production of 1-aminocyclopropane-1-carboxylate (ACC) deaminase in soil and the soil respiration activity such as dehydrogenase and alkaline phosphatase enzymes were also increased [9]. Currently, the purification, characterization and evaluation of bacterial siderophores and their possible application as Fe biofertilizers are receiving increasing attention from researchers [10,11,12]. Moreover, bacterial siderophores exhibit direct antifungal properties, which contributes to the suppression of pathogen development by increasing competition for Fe3+ compared to fungal siderophores. The structures of major siderophores produced by plant growth-promoting bacteria (PGPB) have been characterized [13]. These studies greatly facilitate the application of bacterial siderophores in sustainable agricultural production.
Nitrogen-fixing bacteria (NFB) are a group of functional bacteria that can convert nitrogen in the air into ammonia and are widely used as biostimulant agents to increase soil’s nitrogen availability [14]. In addition to biological nitrogen fixation (BNF), many nitrogen-fixing bacteria also produce siderophores to help plants absorb iron, and further promote plant growth and change the microbial community composition of rhizosphere soil [15,16,17]. In the past decades, the scientific research community has conducted in-depth studies on the isolation of NFB, the modification of nitrogen fixing activity, and the verification of their promoting effects on plant growth. However, there are few reports on the role of nitrogen-fixing bacterial siderophores in promoting plant growth and regulating plant rhizosphere microecology. Sequencing of bacterial genomes and rhizosphere microbiota diversity has become a powerful tool in recent years for exploring the biological function and agricultural application of NFB [18,19].
The NFB strain Kosakonia radicincitans GXGL-4A isolated from the rhizosphere of maize plant can produce siderophore [20]. In our early work, a mutant library of GXGL-4A was constructed based on insertional transposon mutagenesis by electroporation of released Tn5 transposition complexes [21]. To assess the impact of mutants with significantly different siderophore-producing capabilities compared to the wild-type (WT) GXGL-4A strain on the plant growth and the diversity and abundance of rhizosphere microbial communities, in this study, the siderophore synthesis-deficient mutant M246-2 were screened out from the mutant library and subjected to the inoculation of cucumber seedlings. The seedling biomass was measured, and the dynamic changes in the rhizosphere bacterial communities of cucumber plants were monitored through 16S rRNA high-throughput sequencing. Meanwhile, the effects of iron supply level on the propagation of bacterial cells, and the influence of varied siderophore synthesis capabilities of NFB strains on the enzyme activity and nitrogen content in the rhizosphere soil, and the growth of cucumber seedlings were explored.
2. Materials and Methods
2.1. Nitrogen-Fixing Bacterium (NFB), NFB Mutants and Cucumber Variety
The associative NFB Kosakonia radicincitans GXGL-4A with high capability of biological nitrogen fixation (BNF) was isolated from maize root [20]. The high-siderophore-yielding mutant M107 of GXGL-4A constructed via Tn5 insertion mutagenesis has been stored in the China General Microbiological Culture Collection Center (CGMCC) with the preservation number CGMCC 24401 [21]. The seeds of cucumber (Cucumis sativus L.) variety "Xintaimici" were from the Mici Cucumber Original Seed Farm (Xintai City, Shandong Province of China).
2.2. Detection of Siderophore Synthesis Capability of NFB Strains
The siderophore production capabilities of Tn5 mutants of GXGL-4A were determined using blue agar Chrome Azurol Sulfonate (CAS) Assay method [22]. The protocol for making the CAS agar plates was partially modified based on a previous report [23]. First, 0.073 g of HDTMA and 0.06 g of CAS were dissolved in 50 mL and 40 mL of double-distilled water, and then, the CAS solution was slowly poured into HDTMA solution, and thoroughly stirred while adding. The resulted HDTMA-CAS was subjected to a sterilization at 121 °C for 20 min, and subsequently chilled to room temperature. After a cooling, the HDTMA-CAS solution was mixed with 10 mL of 1 mmol/L ferric chloride solution (dissolved in 10 mmol/L hydrochloric acid and sterilized using a 0.22-µm-pore-size microporous filter membrane) to obtain a blue precipitate-free CAS staining agent (Recorded as Solution A, also known as Blue Dye). Solution B containing 50 mL of phosphate buffer, 60 mL of acid hydrolyzed casein, 2 mL of 1 mmol/L calcium chloride and 2 mL of 10 mmol/L magnesium sulfate (pH6.9 adjusted with 1 mol/L sodium hydroxide) per liter was prepared. To make CAS agar plates, 20 g of agar was added in 1 L of Solution B and the mixture was sterilized at 121 °C for 20 min. After sterilization, 10 mL of Solution A was added in a 100 mL of the melted Solution B (containing agar with a mass to volume ratio of 2%) and mixed well. Eventually, CAS agar plates were aseptically made on a super-clean bench. The diameters of the siderophore halos generated and conducted statistical analysis to determine the differences in siderophore synthesis ability among different strains. To test siderophore produced by the three NFB strains noted above, the diameters of siderophore halos were recorded on the 3rd, 6th, 9th and 12th days after the inoculation of NFB strains on CAS agar plates. Experiments were duplicated three times, and the capabilities of NFB strains to produce siderophores were determined based on the statistical comparison of the diameters of the siderophore halos.
2.3. Growth Curves of NFB Strains at Different Levels of Iron Supply
The bacterial growth was monitored by optical density (OD) measurements when cultured in A15 nitrogen-free medium supplied with different levels of iron. The final Fe3+concentrations were 0, 1, 2 and 4 μmol/L. The values of optical density at a wavelength of 600 nm (OD600, 12 h) were determined every 2 h. Growth curves of bacterial cell cultures were attained through repeated measures of OD600.
2.4. Germination and Sowing of Cucumber Seeds and Seedling Management
The cucumber (C. sativus L.) seeds were soaked in 75% ethanol and stirred for 30 s for surface sterilization. After rinsing with sterile water three times, they were then immersed in a 2% sodium hypochlorite solution for 10 min, intermittently stirred, and finally rinsed three times with sterile water. Two sterilized wet filter papers were put in an antiseptic petri dish, and the cucumber seeds fully presoaked with sterile water were evenly placed on the filter papers. Water in the petri dishes were changed every 2 d and these seeds were incubated at 37 ℃ in an incubator until they sprout.
The soils were taken from the Experimental Farm of School of Agriculture and Biology, Shanghai Jiao Tong University, and the nutrient substrates were from the Guangzhou Shengsheng Agriculture Limited Company (China), and then they were thoroughly mixed in a mass ratio of 1:2. The mixture was filled in the plastic flowerpots with a height of 20 cm and an inner diameter of 20 cm. The germinated seeds (0.4 to 0.6 cm of embryo in length) were sown with four seeds per pot and watered well until the seedlings emerged. After emergence, water once a day. When the cotyledons were fully flattened, water once every two days to keep the soil moist.
2.5. NFB Fertilization and the Sampling of Cucumber Rhizosphere Soils
The bacterial cells were inoculated at a rate of 1% (V/V) in 500 mL of LB medium and grown on a rotating shaker at 180 rpm, and 37 °C for 48 h. The cells were harvested by centrifugation at 8 000 rpm for 10 min, and after the supernatant was removed, these cells were resuspended in sterile water. Repeated the above operation twice, and finally the bacterial sediment was suspended in 100 mL of sterile water. After the emergence of the first true leaf of the cucumber seedling, the bacterial cells suspended in the sterile water were used as biofertilizer and applied in the cucumber rhizosphere soils. 5 mL of bacterial cell suspension (about 109 CFU/mL) was fertilized to each cucumber seedling, and an equal volume of sterile water was used as the control. The cucumber seedlings were treated every 5 d, and a total of 5 times of treatments were conducted.
On the 3rd day (Day 3), the 6th day (Day 6) and the 9th day (Day 9) after the last treatment, about 200 g of cucumber rhizosphere soils were carefully and gently collected, and the soils were placed in a sealed plastic bag. These soil samples were immediately stored at -20 ℃ for the later determinations.
2.6. Biomass Determination of the Cucumber Seedlings After NFB Fertilization
To detect the effect of NFB strains with different siderophore-producing activities on cucumber growth, the germinated cucumber seeds were planted in sterile vermiculite. The application method of bacterial cells is as described above. On the 6th day (D6) after fertilization, the biomass of cucumber seedlings including plant height, stem fresh weight, root length, and root fresh weight was measured and subjected to statistical analysis.
2.7. Evaluation of the Soil Nitrogen Content in Cucumber Rhizosphere
The total nitrogen contents in the cucumber rhizosphere soils were evaluated by the Kjeldahl method according to the procedures of the National Environmental Protection Standards of the People’s Republic of China (HJ717-2014). The 0.1 g of soil sample was added to the bottom of the Kjeldahl flask, and sequentially supplied with catalyst 1-(4-bromo-2-methyl-2H-pyrazol-3-yl)-ethanone and 12.5 mL of sulfuric acid. Each flask was placed on a digestion rack for digestion. After a complete digestion, the flasks were chilled to room temperature. Meanwhile, 30 mL of 2% boric acid solution and 20 mL of 10 mol/L sodium hydroxide solution were added in a 250-mL conical flask to start distillation. One drop of methyl red and bromocresol green solutions were successively added to the distillate, and eventually titration was carried out using standard hydrochloric acid solution (0.005 mol/L).
The determination of ammonium nitrogen, nitrite nitrogen, and nitrate nitrogen in soil was performed using potassium chloride solution extraction spectrophotometry based on the corresponding procedures of the National Environmental Protection Standards of the People’s Republic of China (HJ634-2012). 3 g of soil was put into a 15-mL centrifuge tube, and 15 mL of 1 mol/L KCl solution was added. Then, the tube was placed on a rotary shaker at 37 ℃ and 180 rpm for 1 h followed by a centrifugation at 5 000 rpm for 10 min. The supernatant was filtered through a filter paper and the filtrate was subjected to the determination of ammonium, nitrite and nitrate nitrogen contents at the Analysis and Testing Center of Shanghai Jiao Tong University.
2.8. Assessment of the Soil Enzymatic Activity
The soil samples air-dried at room temperature were ground into powder using a grinder, and then sieved through a 40-mesh sieve with a pore size of 0.45 mm. The soil powder was placed at 4 ℃ for the evaluation of soil enzymatic activity. The soils were put into a centrifuge tube according to the requirements of soil enzymatic test kits, and the reagents were added with the manual guidance. The light absorption values of the final products were determined at a specific wavelength.
In the present work, enzymatic activities of soil peroxidase (S-POD), dehydrogenase (S-DHA) and cellulase (S-CL) were determined using commercial testing kits produced by Suzhou Mengxi Biotechnology Cooperation (China) with catalogue numbers M1413B and M1412B, respectively. S-POD catalyzes the oxidation of organic matter to quinones, and the product exhibits a characteristic absorption peak at 430 nm wavelength. The soil peroxidase activity can be characterized by changes in absorbance values. The calculation formula is S-POD activity (mg/d/g soil) = 57.53 × (A+0.003), and in the formula, A represents light absorption value. The determination principle of S-DHA is that after the hydrogen receptor 2, 3, 5-chlorophenyltetrazolium (TCC) receives hydrogen in the S-DHA catalytic process, it is reduced to red triphenylmethane (TF), and the product has a maximum absorption peak at the wavelength of 485 nm. Similarly, the soil dehydrogenase activity can be characterized by the changes in absorbance values. The corresponding calculation formula is S-DHA vitality (μg/d/g soil) = 473.9 × (∆A+0.0312), and of the formula, ∆A=absorbance value of the measuring tube-absorbance value of the blank tube. Soil cellulase can catalyze the degradation of cellulose to produce reducing sugars, which further react with 3,5-dinitrosalicylic acid to produce brownish red amino compounds. The products have characteristic absorption peaks at 540 nm, and the activity of S-CL can be characterized by changes in absorbance values. The experiment was repeated three times, and all the enzymatic activities obtained were subjected to a statistical analysis.
2.9. Isolation of the Cucumber Rhizosphere Soil DNA
The soil metagenomic DNAs were isolated according to the product manual of an E.Z.N.A.® Soil DNA Kit (catalogue No: D5625-01) from Omega Bio-Tek (USA). The isolated DNAs were subjected to a quantification on a NanoDrop 2000 UV-vis spectrophotometer (Thermo Scientific, USA), and DNA quality was checked using agarose gel electrophoresis.
2.10. Amplification of 16S rRNA Genes
The specific primer pair of 338-F (5′-ACTCCTACGGGAGGCAGCAG-3′) and 806-R (5′-GGACTACHVGGGTWTCTAATT-3′) was used to amplify the V3–V4 hypervariable regions of 16S rRNA genes. Amplification reactions were completed on a thermocycler PCR system (GeneAmp® 9700, ABI, USA) using a program consisted of 3 min of denaturation at 95 ℃, 27 cycles of 30 s at 95 ℃, 30 s of annealing at 55 ℃, and 45 s of elongation at 72 ℃ followed by an eventual extension at 72 ℃ for 10 min. Reaction solution contained 4 μL of 5× FastPfu buffer, 2 μL of 2.5 mM dNTPs, 0.8 μL of each primer (5 μM), 0.4 μL of FastPfu polymerase, 0.2 μL of BSA and 10 ng of soil metagenomic DNAs. PCR amplicons were purified using an AxyPrep DNA Gel Extraction Kit (Axygen Biosciences, USA), and quantified using the QuantiFluor™-ST system (Promega, USA).
2.11. Illumina MiSeq Sequencing of the Soil Microbial 16S rRNA Genes
The 16S rRNA amplicons were purified and pooled in equimolar amounts, and then subjected to a paired-end sequencing (PE 2×300) on an Illumina MiSeq platform (Illumina, San Diego, USA). The sequencing procedures were done according to the protocols from the Majorbio Bio-Pharm Technology Corporation, Ltd. (Shanghai, China). Clean reads of the 16S rRNA sequencing have been stored in the Sequence Read Archive (SRA) database of the National Center for Biotechnology Information (NCBI) with an accession number PRJNA1072636.
2.12. Processing of 16S rRNA Sequencing Data
After a quality filtration using a software Trimmomatic, the raw FASTQ files were merged with software FLASH. Operational taxonomic units (OTUs) were clustered with a 97% similarity cut-off using UPARSE (version 11). All the 16S rRNA gene sequences were categorized using a RDP classifier algorithm (version 11.5) against the Silva (SSU123) 16S rRNA database with a confidence threshold of 70%.
3. Results
3.1. Detection of the Siderophile Synthesis Ability of GXGL-4A
A mutant M246-2 which were deficient in producing the siderophore was identified from the established Tn5 insertion mutant library of the NFB strain GXGL-4A by CAS method. The siderophore synthesis ability of the wild-type (WT) strain GXGL-4A and all mutants were verified by measuring the diameters of orange siderophore halos. On the CAS indicator plates, the secretion of siderophore generated a distinct yellow halo around colonies of the WT GXGL-4A and the high-siderophore-yielding mutant M107, while the mutant M246-2 exhibited no yellow halo. With the extension of culture time, the halo diameters gradually expanded. The result of one-way analysis of variance showed that there was a significant difference between the groups, GXGL-4A and M107 (P<0.05) since Day 6 (D6) (Figure 1).
3.2. Relative Siderophore-Producing Activities of GXGL-4A and the Mutant M107 Under Different Fe Supply Levels
The relative siderophore-producing activities of GXGL-4A and the mutant M107 were analyzed by measuring the optical density at wave lengths of 405 and 600 nm under the incubation conditions with iron supply levels of 0, 1, 2, and 4 µmol/L. The results showed that with the increase of Fe supply, the siderophore-producing activities of GXGL-4A and M107 in SM liquid medium decreased gradually. In addition, with the extension of incubation time, the siderophore synthesis activities of two strains gradually decreased (Figure 2). Adding a certain amount of iron during the 12-h growth phase is beneficial for the growth of NFB strains. Overall, there was no significant difference in the growth curves between the strains GXGL-4A and M107, while the growth rate of M246-2 was significantly slower than the other strains (Figure 3).
3.3. Biomass Evaluation of the Cucumber Seedlings Treated with Different NFB Bacterial Cells
The root fresh weight in M107 treatment group was the highest, with significant differences compared to the WT GXGL-4A treatment group (P<0.05), and extremely significant differences compared to the M24-2 and CK treatment groups (P<0.001). Compared with the CK group, the WT GXGL-4A treatment group showed a significant increase in root fresh weight (P<0.05), while there was no significant difference in this biomass between the M246-2 and CK groups (P>0.05) (Figure 4A). In terms of root length, the NFB treatment groups showed a significant or extremely significant increase compared to the CK group (M246-2-treated, P<0.05; GXGL-4A-treated, P<0.01; M107-treated, P<0.001). There was a significant difference between the M107 treatment group and the GXGL-4A treatment group (P<0.05), and a highly significant difference between the M107-treated group and the M246-2-treated group (P<0.01). (Figure 4B). The treatment with NFB can significantly enhance the height of cucumber seedlings, and there were significant (P<0.01) or extremely significant differences (P<0.001) between groups (Figure 4C). Similarly, NFB fertilization can greatly increase the stem fresh weight of cucumber seedlings (P<0.001). The M107 treatment group showed a significant improvement compared to the GXGL-4A treatment group (P<0.01), and an extremely significant enhancement in comparison with the M246-2-treated group (P<0.001) (Figure 4D).
3.4. Nitrogen Contents of the Cucumber Rhizosphere Soil
The contents of total nitrogen (TN), ammonium nitrogen (NH4+−N) and nitrite nitrogen (NO2-−N) in the cucumber rhizosphere soil had a significant difference between groups at three sampling time points after NFB fertilization, while the contents of nitrate nitrogen (NO3-−N) had no significant difference. The results indicated that the application of the NFB strains as biofertilizers improved the soil nitrogen contents. The TN contents of soil in the control group (CK group) gradually decreased, while in the GXGL-4A treatment group (4A group), it increased. The TN contents of soil in the M107 treatment group (M107 group) rose continually during the determination period, and eventually were significantly higher than those of the CK and 4A groups on Day 9. Meanwhile, the TN content of soil in the M246-2 treatment group (M246 group) was significantly lower than those of the other three groups (P<0.05) (Figure 5A). The NH4+−N contents of soil in the groups CK, 4A and M246-2 exhibited a trend of first increasing and then decreasing (Figure 5B). The NO3-−N contents in M246-2 group were significantly higher than those of the other three groups on Day 3 and Day 9 (P<0.05). On Day 3, the soil NO3-−N content in the CK group was significantly lower than those of the 4A, M107 and M246-2 groups (P<0.05), suggesting that NFB fertilization can rapidly increase the NO3-−N contents in the soils of cucumber rhizosphere (Figure 5C). There were no significant differences in the contents of soil nitrate nitrogen between groups (Figure 5D).
3.5. Enzymatic Activity in the Cucumber Rhizosphere Soil After NFB Fertilization
The enzymatic activities of soil dehydrogenase (S-DHA) and peroxidase (S-POD) were determined. As for S-DHA, on Day 3 after the NFB treatment, the enzymatic activity of M107 treatment group was the lowest, and the differences compared with other groups reached a highly significant level (P<0.01). Subsequently, the enzyme activity in this group continued to increase, and on Day 6, there was no significant difference compared to the other groups except for still being significantly lower than the GXGL-4A treatment group. On the 9th day after treatment (i.e. Day 9), the enzyme activity in the M107 treatment group was the highest, significantly or extremely significantly higher than other groups (P<0.01 or P<0.001) (Figure 6A). NFB treatment greatly improved the activity of S-POD. On Day 3 after NFB fertilization, the S-POD activity of each NFB treatment group was significantly higher than that of the control group (i.e. CK group) (P<0.001). Although the M246-2 treatment group had the highest enzyme activity, there was no significant difference compared to the GXGL-4A treatment group and the M107 treatment group (P>0.05). On Day 6, The difference in S-POD enzyme activity between different NFB treatment groups reached a significant (P<0.05) or extremely significant level (P<0.001) (Figure 6B). On Day 3, the S-CL activities of all groups had no significant difference. On Day 6, the S-CL activities in the GXGL-4A and M107 treatment groups were significantly higher than that in the CK group (P<0.05). On Day 9, although there was an increase in the M246-2 treatment group, it was still significantly lower than that of the M107 treatment group and maintained a highly significant difference compared to the CK group (P<0.001). The S-CL activity of CK group exhibited a trend of first decreasing and then increasing throughout the entire experimental period, but the NFB treatment groups showed an opposite trend. Overall, NFB fertilization has a promoting effect on the S-CL activity of cucumber rhizosphere soil, and this effect is particularly distinct on Day 6 (Figure 6C).
3.6. Sequencing Data Summary and the α-Diversity Analysis
A total of 2 968 900 optimized sequences were obtained from the deep sequencing of 16S rRNA genes in 48 cucumber rhizosphere soil samples. The average read length of 16S rRNA gene was 414 bp. Eventually, 18 181 operational taxonomic units (OTUs) in the soil bacterial communities were identified based on the clean reads. These OTUs belong to 48 Phyla, 158 Classes, 396 Orders, 647 Families, 1330 Genera, and 3230 Species.
3.7. α-Diversity Analysis
As shown in Table 1, among all treatments on Day 3, the bacterial community richness indices (Sobs, Chao and Ace) and the community diversity index (Shannon) of the three treatment groups (4A, M107 and M246-2) were significantly higher than those of the CK group, indicating that the bacterial community abundance and diversity significantly increased after NFB fertilization (P<0.05). On Day 6, the bacterial community richness in the M107 treatment group was significantly higher than that in the 4A treatment group (P<0.05), and there was no significant difference in bacterial community diversity among groups (P>0.05). On Day 9, the bacterial community richness and community diversity in M107 treatment group were significantly lower than those in the CK and M246-2 groups (P<0.05), but had no significant difference compared with the 4A-treated group (P>0.05). With the extension of sampling time, the Sobs, Chao and Ace indices of the CK group gradually increased, suggesting that the richness of the bacterial communities in this group continually enhanced. The community richness and diversity of the 4A and M246-2 groups decreased first, and then increased, while that of the M107 group decreased gradually. Fertilization with the M107 bacterial cells could significantly reduce the abundance and diversity of bacterial communities in the cucumber rhizosphere soil.
3.8. Analysis of Species Composition at Different Taxonomic Levels
The relative abundance of dominant genera modified after NFB fertilization. The main genera were Devosia (3.0%~6.9%), norank_f__norank_o__SBR1031 (1.6~8.0%), norank_f__norank_o__Vicinamibacterales (2.2 ~ 5.1%), Streptomyces (1.5 ~ 4.4%) and norank_f__A4b (1.4 ~ 4.1%) (Figure 7).
The microbial community composition at the phylum level was detected. The main phyla were Proteobacteria (34.0-44.1%), Actinobacteriota, (14.7-21.7%), Chloroflexi (10.5-16.4%) and Acidobacteriota (4.8-12.0%). They account for more than 70% of the total community, so they are considered as dominant phyla in cucumber rhizosphere soils. The relative abundance of the phylum Proteobacteria increased firstly, and then decreased in CK, 4A and M246-2 treatment groups. On Day 9, the abundance of the phylum Actinobacteriota decreased in 4A, M107 and M246-2 treatment groups, while in the CK group, it showed an increasing trend. After 9-d of the NFB application, the relative abundance of the phylum Chloroflexi in the M107 treatment group was lower than that in CK, 4A and M246-2 treatment groups. The relative abundances of the phylum Acidobacteriota decreased firstly, and subsequently increased in the CK, M107 and M246-2 treatment groups (Figure 8).
3.9. Difference Analysis of the Bacterial Community Structure in Cucumber Rhizosphere Soil
The difference in the soil bacterial community structure in cucumber rhizosphere was conducted by non-metric multidimensional scale (NMDS) analysis at the OTU level. The results revealed that the bacterial community structure in cucumber rhizosphere soil significantly altered after the application of NFB strains. Grouping of all the tested soil samples based on sampling time points was correct and reliable (stress: 0.101, R=0.7357 and P=0.001). The compositions of bacterial communities in the soils on Day 9 after GXGL-4A application varied significantly compared with that on Day 6 and Day 3. In the M107 group, the bacterial population structure modified on Day 3 after fertilization compared to that on Day 6 and Day 9 after fertilization (Figure 9).
There were significant differences in the composition and structure of microbiota among different soil groups. The results revealed that Actinobacteriota, Bacteroidota, Patescibacteria and Myxococcota were the dominant phyla of soil samples. Streptomyces, norank_f__A4b, Arthrobacter, unclassified_f__Chitinophagaceae and norank_f__Vicinamibacteraceae were the dominant genera in soil samples (Figure 10A).
Kruskal-Wallis H test was used to evaluate the relative abundance difference of the Kosakonia genus in the GXGL-4A, M107 and M246-2 treatment groups through multi-species difference analysis (Figure 10B). The relative abundance of Kosakonia in GXGL-4A- and M107-treated groups was statistically extremely significant (P<0.01), while no significant difference was found between GXGL-4A and M246-2 treated groups on Day 3 and Day 6 after fertilization. On Day 9, the relative abundance of Kosakonia in GXGL-4A and M246-2 treatment groups was statistically significant (P<0.01), but there was no significant difference between the GXGL-4A and M107 treatment groups.
The abundance of Kosakonia in each group was analyzed. The result indicated that the abundance of the genus Kosakonia varied in three sampling time points after the application of NFB bacterial cells. On Day 3, the abundance of Kosakonia in the M107 treatment group was always higher than that in the GXGL-4A treatment group, and the relative abundance of the M246-2 treatment group was the lowest. The abundance ratio of GXGL-4A treatment group was 2.359%, while that of M107 treatment group was 8.123% (Figure S1A). Before Day 6, the abundance of Kosakonia in the M107 treatment group was significantly higher than that in 4A and M246-2 groups (P<0.05) (Figure S1B). On Day 9, the abundance of Kosakonia in M246-2 group was significantly lower than that in GXGL-4A treatment group (P<0.05), and in the GXGL-4A treatment group, the abundance of Kosakonia increased first, and then decreased (Figure S1C).
3.10. Environmental Factor Correlation Analysis
The effect of NFB fertilization on the rhizosphere soil bacterial flora was evaluated using 16S rRNA gene high-throughput sequencing. The results indicated that the application of NFB not only affected the content of nitrogen and enzyme activity in the cucumber rhizosphere soil, but also significantly reshaped the microbial community composition and diversity. Through environmental factor correlation analysis, at the genus level, it was found that GXGL-4A and mutant strains M107 and M246-2 affect the abundance and composition of microbial communities by altering the total nitrogen (TN) level of the soil. The alteration in TN content is considered a primary and important factor. Among the top fifty dominant genera in terms of abundance, only 9 genera such as Hephaestia, Hyphomicrobium, and Steridobacterium showed no correlation with all the tested environmental factors, while the abundance of the remaining 41 genera was significantly correlated with multiple environmental factors. Total nitrogen (TN), soil dehydrogenase (S-DHA), and soil ammonium nitrogen (NH4+−N) are the three key environmental factors, which show significant or extremely significant correlations with the abundance of most dominant genera in the rhizosphere soil. Secondly, nitrate nitrogen (NO3-−N) and nitrite nitrogen (NO2-−N) are important environmental factors that are significantly or extremely significantly correlated with the abundance of many dominant genera, and are mostly negatively correlated. The activities of soil peroxidase (S-POD) and cellulase (S-CL) had negligible effect on the abundance of soil dominant microbial flora. In particular, the activity of S-CL was significantly positively correlated with the species abundance of only three genera Pseudolabrys, Luteimonas and Nocardioides in the top fifty genera (Figure S2).
3.11. Two-Way Correlation Network Analysis
The results of two-way correlation network analysis indicated that at the genus level, the abundance of bacteria was influenced by multiple environmental factors. For instance, the abundance of species belonging to the genera Asccacaulis, Sphingomonas, Devonia, Marmoricola and Bacillus was significantly influenced by the soil environmental factors such as TN, S-DHA, NH4+−N and NO3-−N. There is a networked interaction between the environmental factors evaluated and the abundance of bacterial species in the cucumber rhizosphere soil. Alterations in the nitrogen content and dehydrogenase activity of soil would reshape the structure and abundance of soil microbiota (Figure S3).
3.12. Discussion
A siderophore-deficient mutant M246-2 was obtained from the Tn5 mutant library of K. radicincitans GXGL-4A using CAS assay method in this study, and except for it, the WT GXGL-4A and high siderophore-yielding mutant M107 showed strong CAS reactivity, indicative of siderophore production. Siderophores produced by these NFB strains are essential for iron acquisition, and consequently modulate the relative growth rates of bacterial cells. In general, the siderophore-deficient mutant M246-2 grew the slowest in media supplied with limited iron (0-4 µmol/L) compared to other strains. Biofertilizers have been widely used in agricultural production because they can effectively promote plant growth. Reasonable addition of plant growth-promoting rhizobacteria (PGPR) can also regulate soil nutrients and prevent plant diseases by hindering the proliferation of pathogenic bacteria [23]. It has been confirmed that biological fertilizer can affect the rhizosphere microbial community and plant growth, but the effect of NFB as biofertilizer on the bacterial community of cucumber rhizosphere soil is unclear [24]. Currently, the development of high-throughput sequencing technology and bioinformatics technology makes it possible to study the bacterial community structure quickly and easily in different habitats. The 16S rRNA gene sequencing is a common tool in microbiome investigation and it is still being improved to increase the sensitivity and applicability in environmental bacterial population diversity analysis, which is driving the rapid progress of modern agricultural research [25].
Previous study reported that Bacillus subtilis and Bacillus licheniformis treatments increased the α-diversity of maize rhizosphere soil bacterial community, and different fertilization treatments affected the abundance and diversity of soil bacterial community [26]. Our study results were consistent with this report. The application of NFB strains significantly modified the bacterial community in the rhizosphere soil of cucumber. There was a significant difference in α-diversity between the GXGL-4A and M107 treatment groups (P<0.05), indicating that the synthesis capability of siderophore by the NFB GXGL-4A may affect the reshaping of the bacterial community in cucumber rhizosphere soils by a short-term fertilization. However, there was no significant difference in soil bacterial community between the GXGL-4A and M246-2 treatment groups. Fertilization with the mutant M107 significantly reduced the abundance and diversity of bacterial communities in the rhizosphere soil compared to the other groups.
There were a higher abundance in the genera Arthrobacter, Bacillus and Kosakonia in the rhizosphere soil after the applications of GXGL-4A、M107 and M246-2 bacterial cells. Many bacterial strains in the genus Arthrobacter can degrade nitroglycerin, benzene derivatives, polycyclic aromatic compounds, halogenated alcohols, halogenated hydrocarbons, N-heterocyclic compounds, pesticides and herbicides [27,28,29,30,31,32]. Thus, NFB fertilization theoretically could promote plant growth through the way of degradation of toxic compounds in soil.
A variety of strains in the genus of Kosakonia can increase plant growth activity by producing auxin, siderophore and increasing phosphorus [33]. In our study, the relative abundance of Kosakonia was different in diverse groups. At the same sampling time point, the relative abundance of Kosakonia in M07 group was significantly higher than that in GXGL-4A-treated group, and similarly the abundance of Kosakonia in GXGL-4A treatment group was significantly higher than that in M246-2 fertilization group (P<0.05). Therefore, we speculated that the activity of siderophore production is closely related to the survival status of a certain bacterium in soil.
Soil enzymatic activity reflects the strength of soil substance transformation (e.g., C, N and P), and is suitable for evaluating the effect of fertilization. S-CL and S-DHA enzymatic activities can well tag soil fertility [34]. In this work, the enzymatic activities of S-CL and S-DHA in cucumber rhizosphere soil were measured at three sampling time points (i.e., Day 3, Day 6, and Day 9 after NFB fertilization). Soil-cellulase (S-CL) is an important enzyme in the carbon cycle of soil [35,36]. In general, the S-CL enzyme activity of the three NFB treatment groups showed a trend of first increasing and then decreasing. On Day 6 after NFB treatment, the S-CL enzyme activity was the highest, and the three S-CL enzyme activities of NFB treatment groups were significantly higher than that of the CK group (P>0.05).These results indicated that after the application of NFB, the S-CL enzyme activity in the rhizosphere soil was greatly increased, which accelerated the decomposition of cellulose in the soil to provide sufficient carbon source for the absorption and utilization of cucumber seedlings, which was conducive to promoting their vegetative growth. S-DHA activity has been used as a sensitive indicator to monitor respiratory and biochemical processes in soil microbial communities [37]. In the present study, the enzymatic activities of GXGL-4A and M107 treatment groups increased sharply on Day 3 and Day 9 after treatment of NFB bacteria respectively, which was significantly higher than that of the other two groups, suggesting that soil respiration strength was significantly enhanced, and the nitrogen cycling in soil was improved. After the treatment by the bacterial cells of siderophore-producing mutants, the tested soil enzyme activities significantly varied, which affecting the nitrogen cycle of cucumber rhizosphere soil. These modifications in soil enzyme activity are a short-term trend, and we consider that this is a vital component of complex regulation of soil microenvironment.
Soil inorganic nitrogen content has a direct impact on crop growth yield and quality, so it is regarded as an evaluation index for nutrient management in the field [38]. In this study, the three NFB biofertilizers could significantly increase the contents of TN, NH4+−N, and NO2-−N in cucumber rhizosphere soils in a short term. One mechanism is that NFB bacteria may help host plants produce root secretions that, in turn, absorb beneficial bacterial communities, thereby increasing the levels of dissolved nutrients in the soil. In addition, a growing body of research indicates that the microbial composition is largely determined by environmental factors [39,40]. The differences in siderophore-producing abilities of the NFB mutants affect the iron absorption capabilities of bacteria themselves and the rhizosphere soil microbiota, modulate the utilization efficiency of soil microorganisms for nitrogen and iron nutrients, and thus reshape the rhizosphere soil bacterial communities.
Supplementary Materials
Figure S1: Difference test analysis of samples at Kosakonia genus level. Kruskal–Wallis H test was used for significance testing among soil sample groups. Figure S2: Spearman Correlation Heatmap at the genus level. An asterisk indicates a significant correlation between an environmental factor and the abundance of species in the genus (P<0.05). Similarly, two or three asterisks indicate a highly significant correlation between an environmental factor and the abundance of species in that genus (P<0.01 and P<0.001, respectively). The top 50 genera in terms of abundance are displayed in descending order in the graph according to their abundance size. Figure S3: Network diagram of the correlation between bacterial species and environmental factors at the genus level. The red and green lines respectively represent the positive or negative correlation between environmental factors and the abundance of a certain genus species. The thicker the line, the stronger the correlation.
Author Contributions
Conceptualization, Y.C; methodology and investigation, Y.Z. and E.W.; data curation, B.F. and L.X.; formal analysis, Y.Z. and Y.X.; writing-original draft preparation, Y.Z.; writing-reviewing and editing, Y.C; software, Y.Z. and E.W.; supervision, Y.C.; project administration, Y.C.; funding acquisition, Y.C. All authors have read and agree to the published version of the manuscript.
Funding
This work was financially supported by the National Natural Science Foundation of China (No. 31870496).
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Materials, further inquires can be directed to the corresponding author. The 16S rRNA gene amplicon sequencing data of the cucumber rhizosphere soils respectively treated with the wild-type strain GXGL-4A and the mutants M246-2 and M107 have been deposited in Sequence Read Archive (SRA) database with a BioProject number PRJNA1072636.
Acknowledgments
The authors thank Mengting Zhang (Department of Resources and Environment, School of Agriculture and Biology, Shanghai Jiao Tong University) and Yuqing Bao (School of Health Science and Engineering, University of Shanghai for Science and Technology) for helping to construct and preserve Tn5 mutants of the nitrogen-fixing bacterium GXGL-4A.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Mushtaq.; Z, Al-Ashkar. I; Habib-ur-Rahman, M.; Sabagh, A.E.; Ilić, P. Biofortification of iron in potato through rhizobacteria and plant growth regulator. Potato Res 2023, 1–9. [CrossRef]
- Ahmad, F.; Mushtaq, Z.; Anwar, W.; Nazir, A.; Akhtar, A.; Liaquat, M.; Jaffar, M.T.; Chaudhry, A.; Saeed, I.; Khan, H.A.A. Impact of siderophore producing rhizobacteria on growth and iron content in potato. Pak J Sci 2023, 75(2): 338-344.
- Ghehestani, M.M.H.; Azari, A.; Rahimi. A.; Maddah-Hosseini, S.; Ahmadi-Lahijani, M.J. Bacterial siderophore improves nutrient uptake, leaf physiochemical characteristics, and grain yield of cumin (Cuminum cyminum L.) ecotypes. J Plant Nutr 2021, 44(12): 1794-1806. [CrossRef]
- Mushtaq, Z.; Liaquat, M.; Jehan, S.; Akram, MT.; Jaffar, M.T.; Alasmari, A. Microbial biofortification of iron Solanum tuberosum L. through siderophore-producing rhizobacteria along with L-tryptophan and iron sulphate supply. Potato Res 2024,. [CrossRef]
- Liu, J.; Tang, L.; Gao, H.; Zhang, M.; Guo, C. Enhancement of alfalfa yield and quality by plant growth-promoting rhizobacteria under saline-alkali conditions. J Sci Food Agric 2019, 99: 281-289. [CrossRef]
- Rajkumar, M.; Narayanasamy, S.; Uthandi, S. A root-associated Bacillus albus LRS2 and its metabolites for plant growth promotion and drought stress tolerance in little millet (Panicum sumatrense L.). Plant Stress 2024, 12: 100446. [CrossRef]
- Sarwar, S; Khaliq, A; Yousra, M; Sultan, T. Iron biofortification potential of siderophore producing rhizobacterial strains for improving growth, yield and iron contents of groundnut. J Plant Nutr 2022, 45(15): 2332-2347. [CrossRef]
- Liu, D.; Yang, Q.; Ge, K.; Hu, X.; Qi, G.; Du, B.; Liu, K. Promotion of iron nutrition and growth on peanut by Paenibacillus illinoisensis and Bacillus sp. strains in calcareous soil. Braz J Microbiol 2017, 48: 656-670. [CrossRef]
- Chandwani, S.; Amaresan, N. Siderophore and ACC deaminase producing bacteria enhance the growth of Vigna spp under iron limited saline soils. J Soil Sci Plant Nutr 2024, 24: 3734-3748. [CrossRef]
- Nithyapriya, S.; Sundaram, L.; Eswaran, S.U.D.; Perveen, K.; Alshaikh, N.A.; Sayyed, R.Z.; Mastinu, A. Purification and characterization of desferrioxamine B of Pseudomonas fluorescens and its application to improve oil content, nutrient uptake, and plant growth in peanuts. Microb Ecol 2024, 87: 60. [CrossRef]
- Lozano-González, J.M.; Valverde, S.; Montoya, M.; Martín, M.; Rivilla, R.; Lucena, J.J.; López-Rayo, S. Plants 2023, 12: 4054. [CrossRef]
- Gao, B.; Chai, X.; Huang, Y.; Wang, X.; Han, Z.; Xu, X.; Wu, Ting.; Zhang, X.; Wang, Y. Siderophore production in Pseudomonas SP. strain SP3 enhances iron acquisition in apple rootstock. J Appl Microbiol 2022, 133:720-732. [CrossRef]
- Timofeeva, A.M.; Galyamova, M.R.; Sedykh, S.E. Bacterial siderophores: classification, biosynthesis, perspectives of use in agriculture. Plants 2022, 11: 3065. [CrossRef]
- Wu, Q.; Yang, L.; Liang, H.; Liu, M.; Chen, Y.; Chen, D.; Shen, P. Impacts of soil compaction and phosphorus levels on the dynamics of phosphate-solubilizing and nitrogen-fixing bacteria in the peanut rhizosphere. Agronomy 2024, 14(9): 1971. [CrossRef]
- Adediji, A.O.; Ojo, J.A.; Olowoake, A.A.; Alabi, K.O.; Atiri, G.I. Complete genome of Achromobacter xylosoxidans, a nitrogen-fixing bacterium from the rhizosphere of cowpea (Vigna unguiculata [L.] Walp) tolerant to cucumber mosaic virus infection. Curr Microbiol 2024, 81(11): 356. [CrossRef]
- Widawati, S.; Suliasih; Susilowati; D.N.; Sumardi. The population and potential of nitrogen-fixing bacteria from sandalwood (Santalum album L.) rhizosphere as a producer of phytohormones and stress resistance indicators. AIP Conf Proc 2024, 2970(1): 050006. [CrossRef]
- Luo, D.; Shi, J.; Li, M.; Chen, J.; Wang, T.; Zhang, Q.; Yang, L.; Zhu, N.; Wang, Y. Consortium of phosphorus-solubilizing bacteria promotes maize growth and changes the microbial community composition of rhizosphere soil. Agronomy 2024, 14(7): 1535. [CrossRef]
- Adediji, A.O.; Ojo, J.A.; Olowoake, A.A.; Alabi, K.O.; Atiri, G.I. Complete genome of Achromobacter xylosoxidans, a nitrogen-fixing bacteria from the rhizosphere of cowpea (Vigna unguiculata [L.] Walp) tolerant to cucumber mosaic virus infection. Curr Microbiol 2024, 81: 356. [CrossRef]
- Peng, T.; Meng, L.; Wang, Y.; Jin, L.; Jin, H.; Yang, T.; Yao, Y. Alterations of the rhizosphere soil microbial community composition and metabolite profiles of Angelica sinensis seedlings by co-application of nitrogen fixing bacteria and amino acids. Plant soil 2023, 493(1-2): 535-554. [CrossRef]
- Sun, S.; Chen, Y.; Cheng, J.; Li, Q.; Zhang, Z.; Lan, Z. Isolation, characterization, genomic sequencing, and GFP-marked insertional mutagenesis of a high-performance nitrogen-fixing bacterium, Kosakonia radicincitans GXGL-4A and visualization of bacterial colonization on cucumber roots. Folia Microbiol 2018, 63: 789-802. [CrossRef]
Figure 1.
The detection and relative quantification of siderophores produced by the NFB strains GXGL-4A, M107 and M246-2 using the chrome azurol sulfonate (CAS) assay. A: clear yellow haloes exhibited on Day 3 (D3), Day 6 (D6), Day 9 (D9), and Day 12 (D12); B: significant differences in the halo diameters between groups. An asterisk indicates a significant difference at the p=0.05 level. Similarly, double and three asterisks represent a significant difference at the P=0.01 and P=0.001 levels, respectively.
Figure 1.
The detection and relative quantification of siderophores produced by the NFB strains GXGL-4A, M107 and M246-2 using the chrome azurol sulfonate (CAS) assay. A: clear yellow haloes exhibited on Day 3 (D3), Day 6 (D6), Day 9 (D9), and Day 12 (D12); B: significant differences in the halo diameters between groups. An asterisk indicates a significant difference at the p=0.05 level. Similarly, double and three asterisks represent a significant difference at the P=0.01 and P=0.001 levels, respectively.
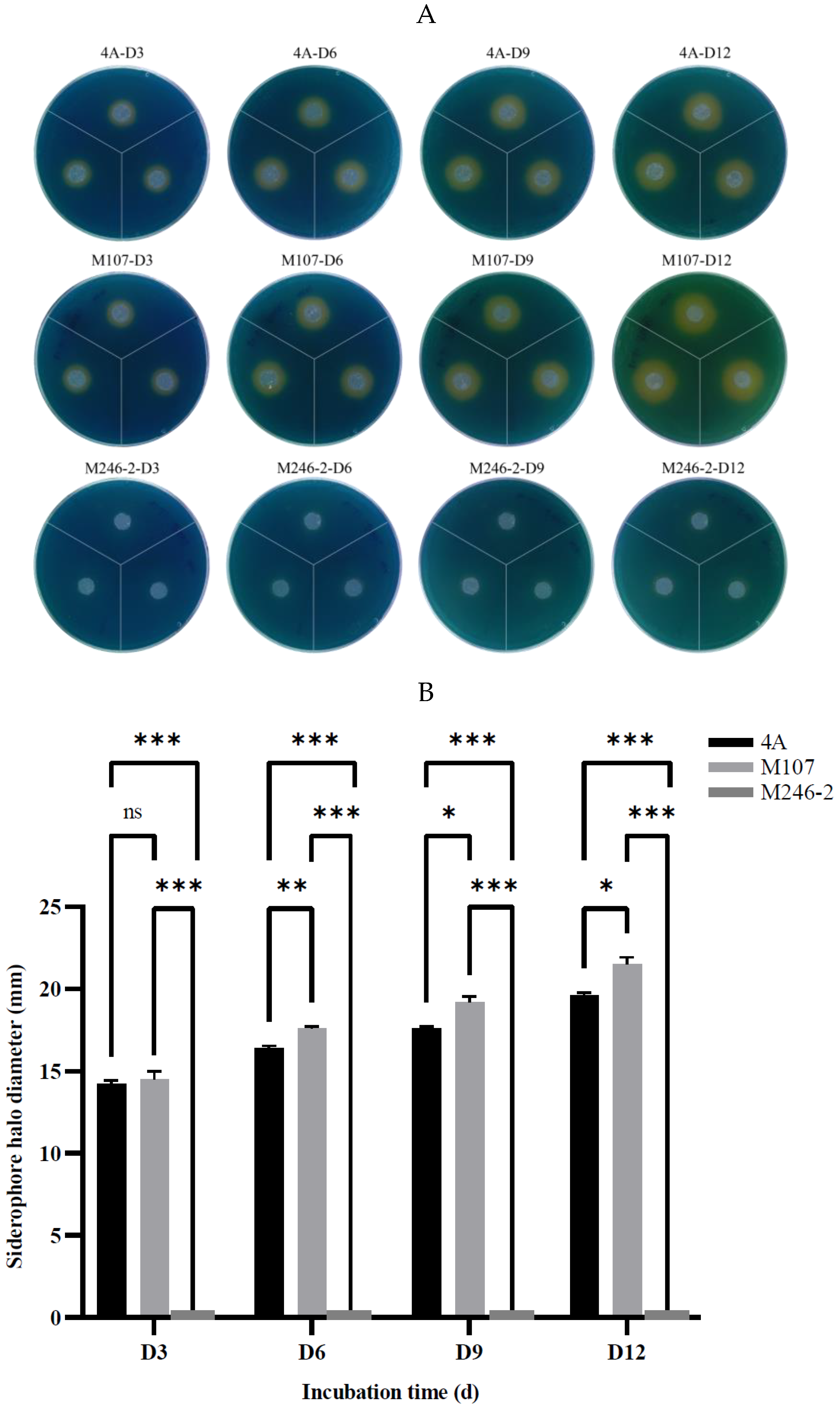
Figure 2.
Relative siderophore-producing capabilities of the NFB strains GXGL-4A and M107 at different ferric supply levels (A: the wild-type strain GXGL-4A; B: the mutant M107).
Figure 2.
Relative siderophore-producing capabilities of the NFB strains GXGL-4A and M107 at different ferric supply levels (A: the wild-type strain GXGL-4A; B: the mutant M107).
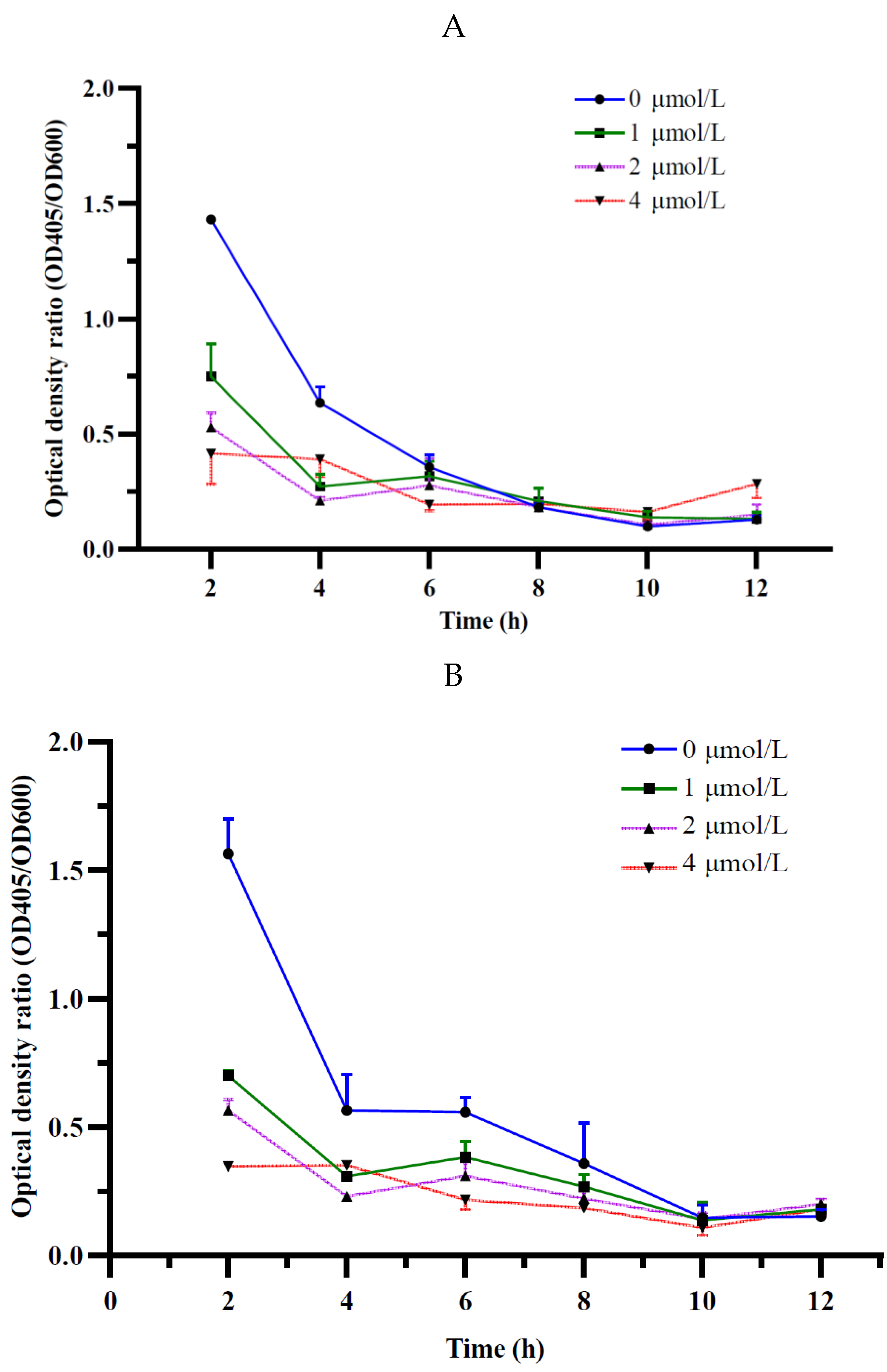
Figure 3.
Bacterial growth curves of the NFB strains GXGL-4A, M107 and M246-2.
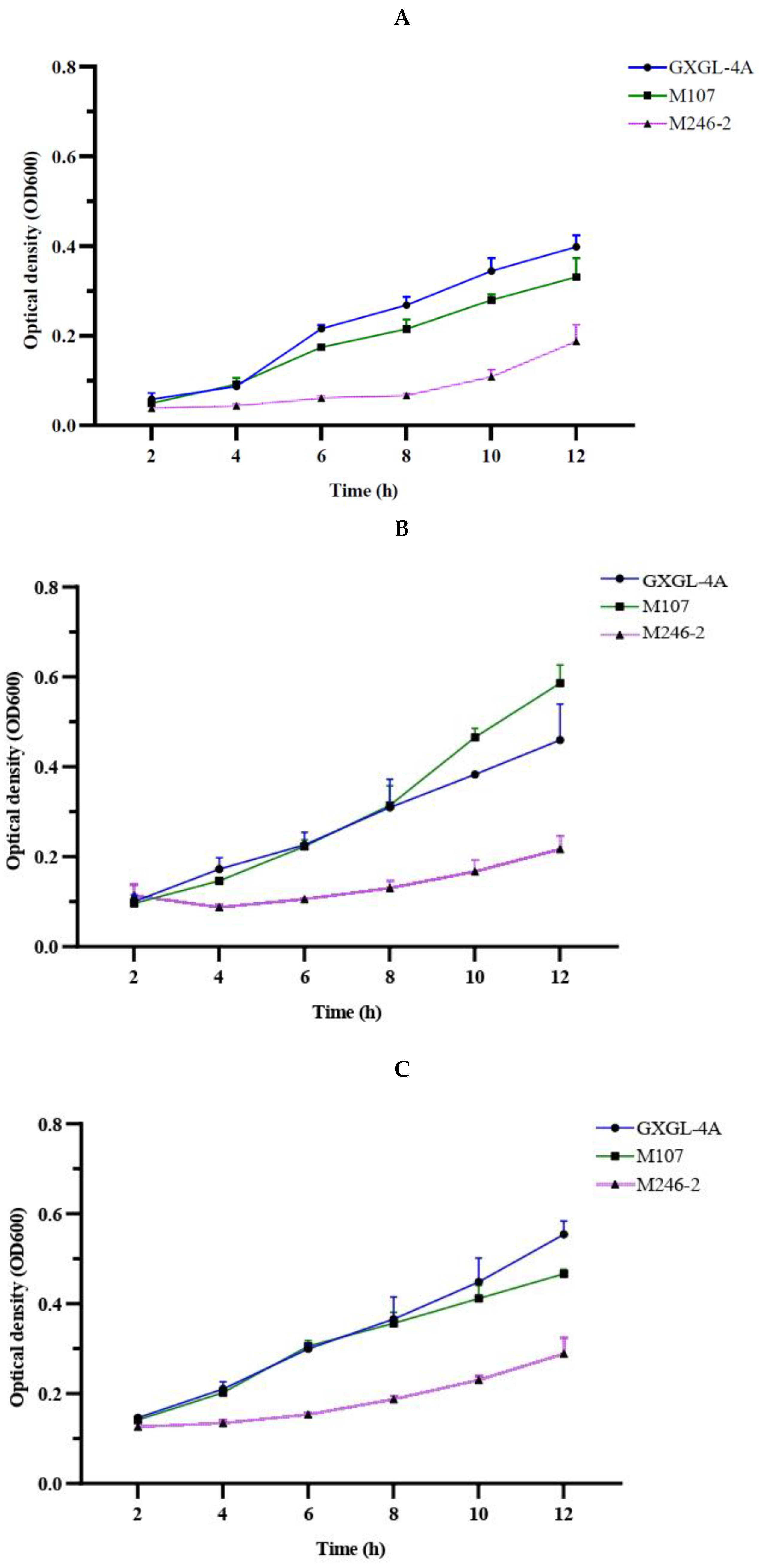
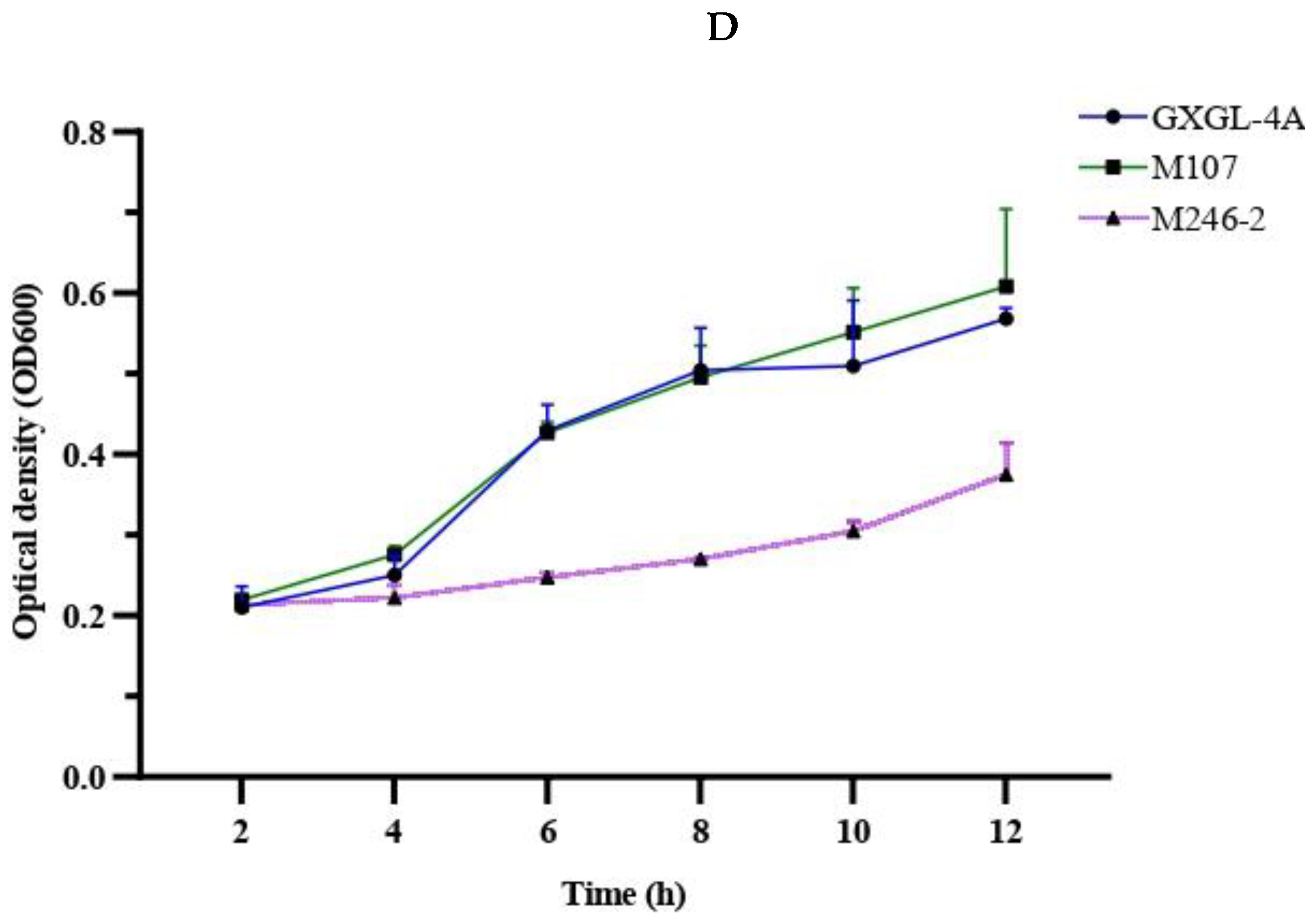
Figure 4.
Biomass of cucumber seedlings fertilized by NFB strains with different siderophore-producing abilities. A: root fresh weight; B: root length; C: seedling height; D: stem fresh weight.
Figure 4.
Biomass of cucumber seedlings fertilized by NFB strains with different siderophore-producing abilities. A: root fresh weight; B: root length; C: seedling height; D: stem fresh weight.
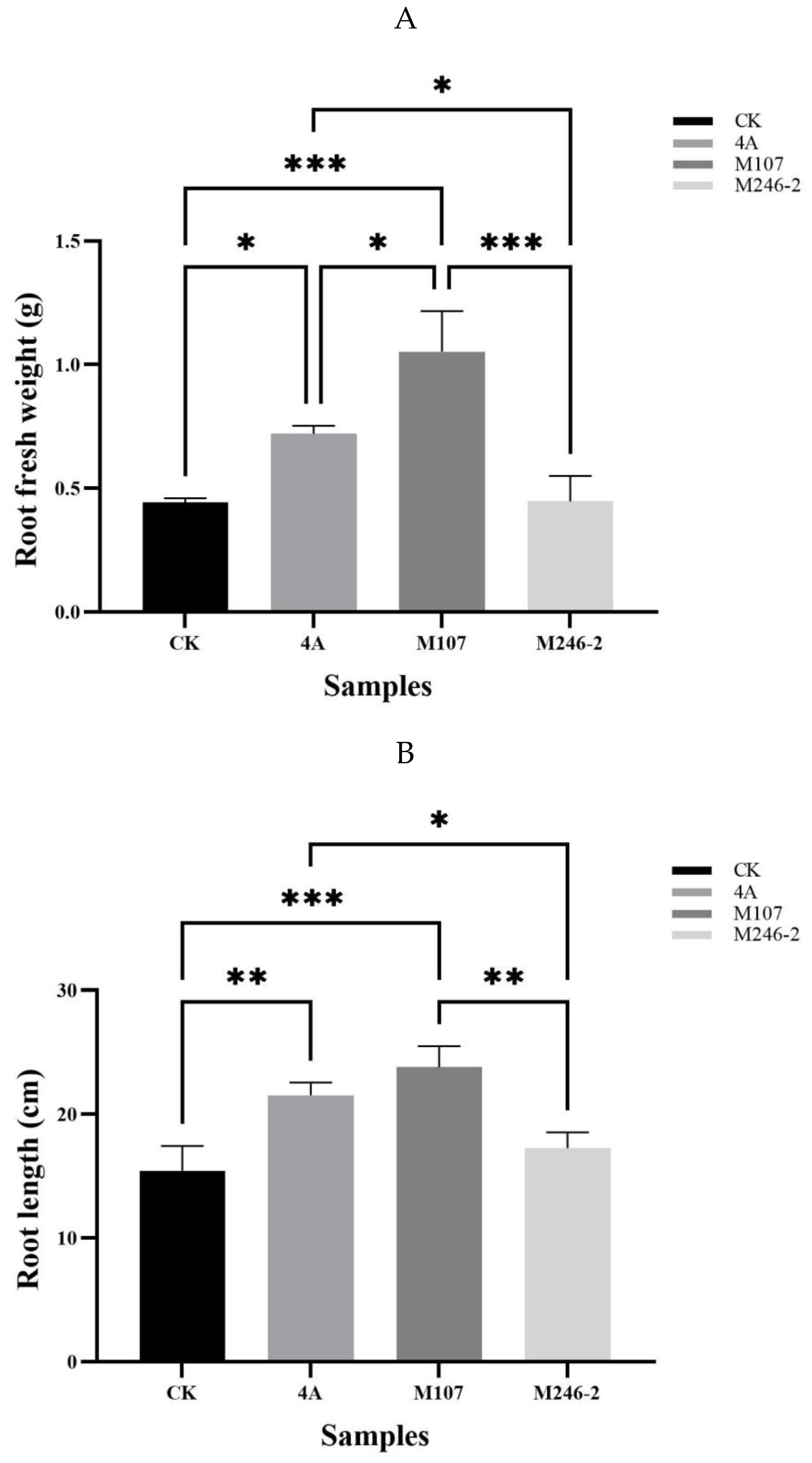
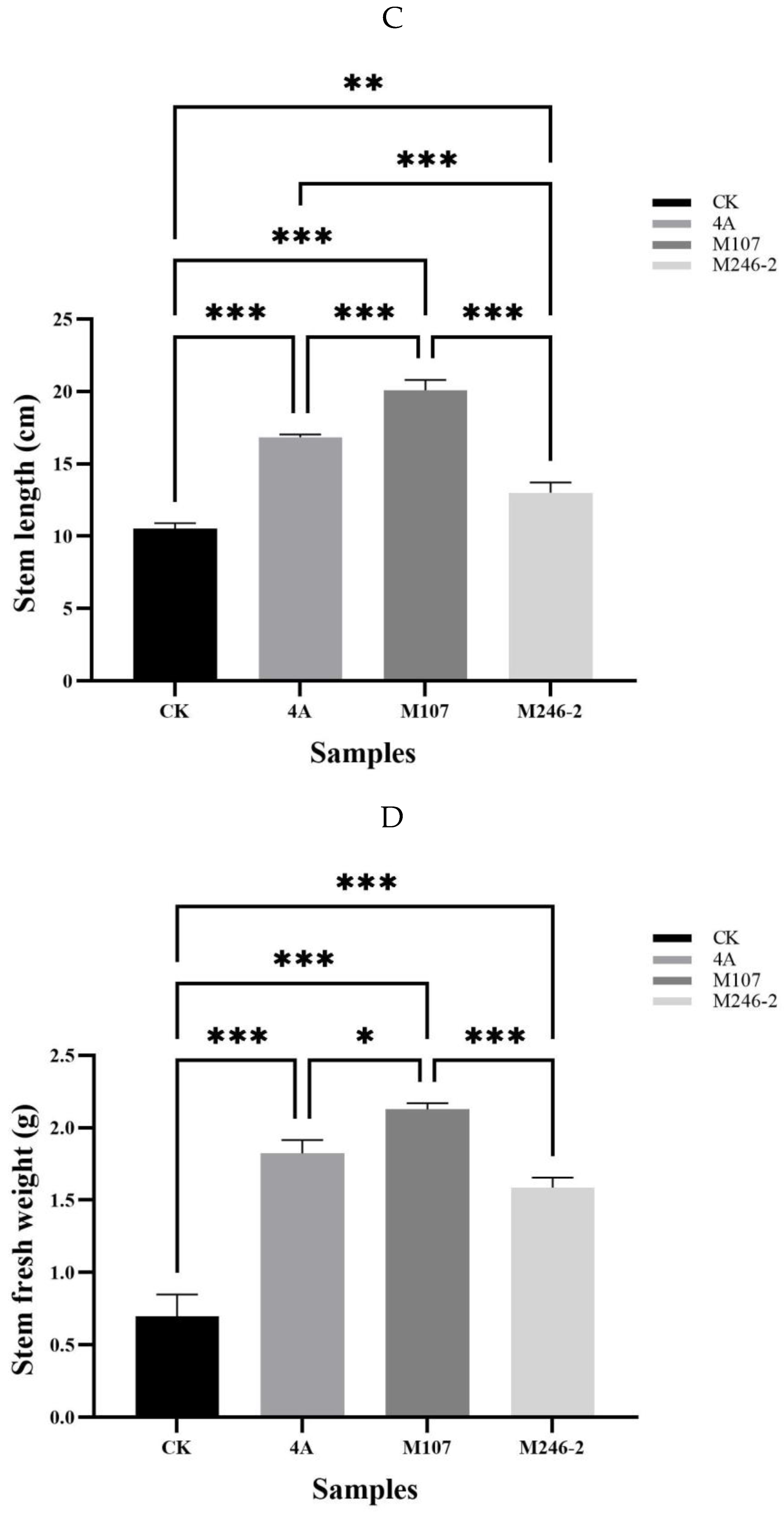
Figure 5.
Nitrogen contents of the cucumber rhizosphere soils treated with NFB strains with different siderophore-producing abilities. A: Total nitrogen (TN); B: ammonium nitrogen (NH4+−N); C: nitrite nitrogen (NO2-−N); D: nitrate nitrogen (NO3-−N).
Figure 5.
Nitrogen contents of the cucumber rhizosphere soils treated with NFB strains with different siderophore-producing abilities. A: Total nitrogen (TN); B: ammonium nitrogen (NH4+−N); C: nitrite nitrogen (NO2-−N); D: nitrate nitrogen (NO3-−N).
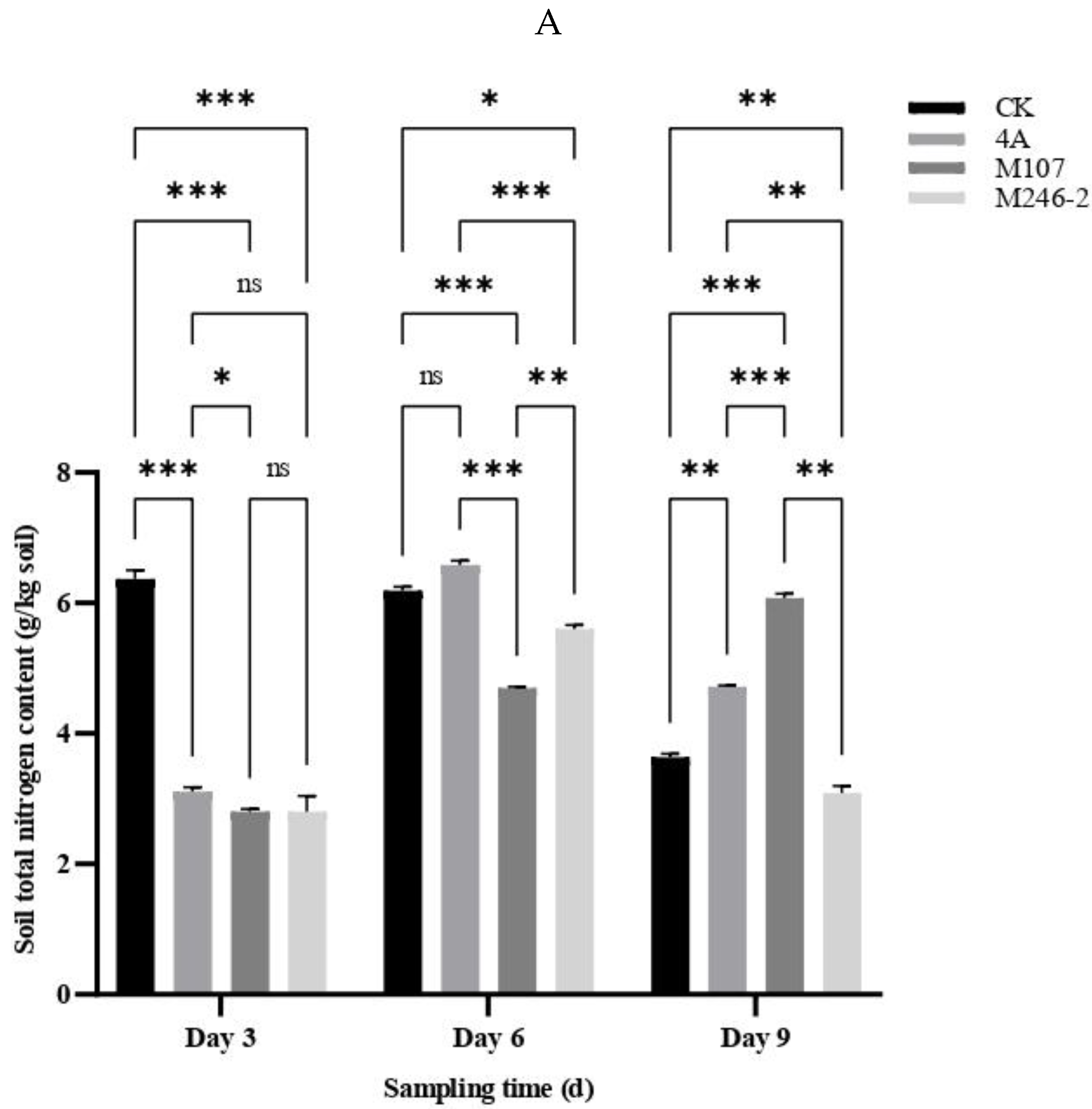
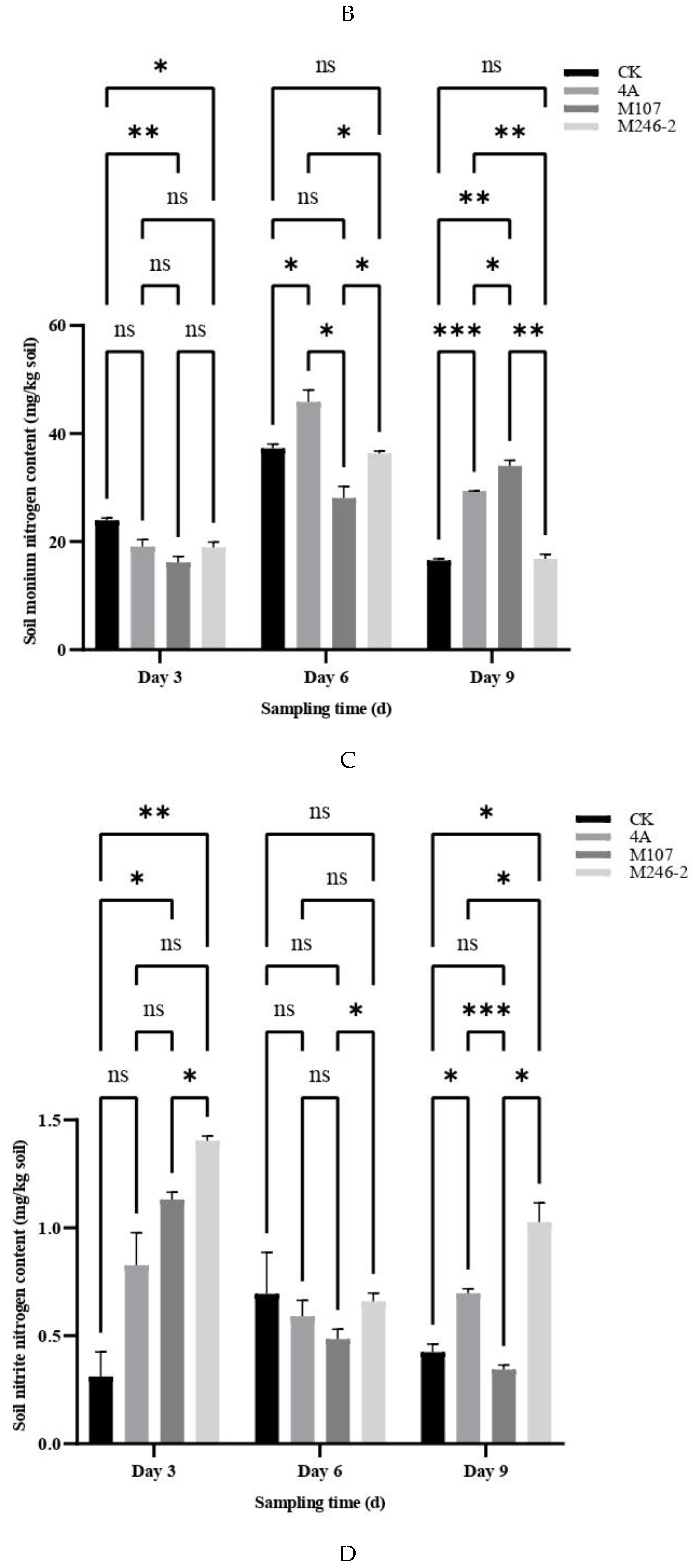
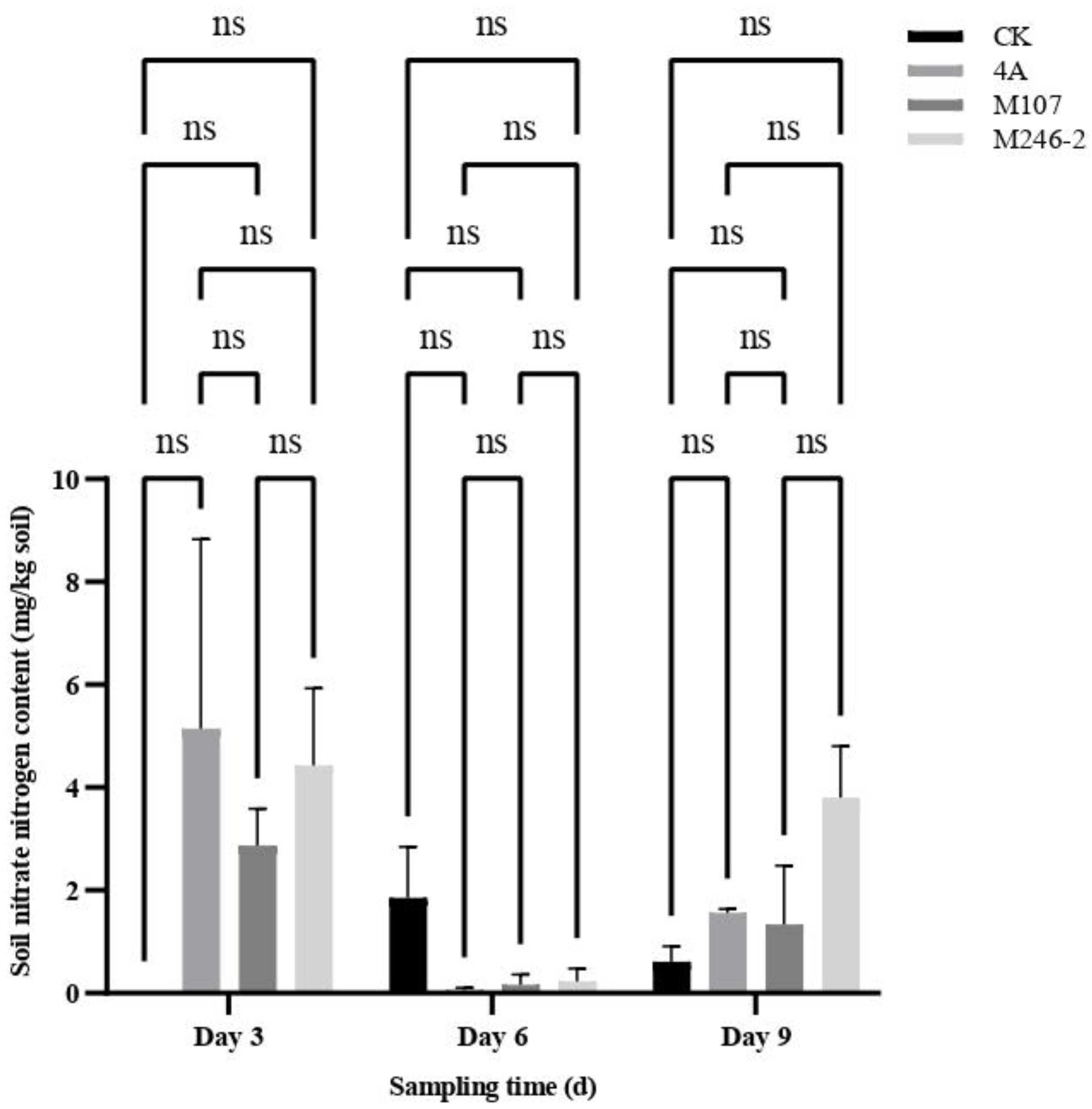
Figure 6.
Soil enzymatic activity in cucumber rhizosphere after NFB application. The figures 6A, 6B and 6C respectively show the dynamic changes in the enzymatic activities of soil dehydrogenase (S-DHA), peroxidase (S-POD) and cellulase (S-CL) in the rhizosphere soil of cucumber sampled on the 3rd, 6th, and 9th days after NFB treatment.
Figure 6.
Soil enzymatic activity in cucumber rhizosphere after NFB application. The figures 6A, 6B and 6C respectively show the dynamic changes in the enzymatic activities of soil dehydrogenase (S-DHA), peroxidase (S-POD) and cellulase (S-CL) in the rhizosphere soil of cucumber sampled on the 3rd, 6th, and 9th days after NFB treatment.
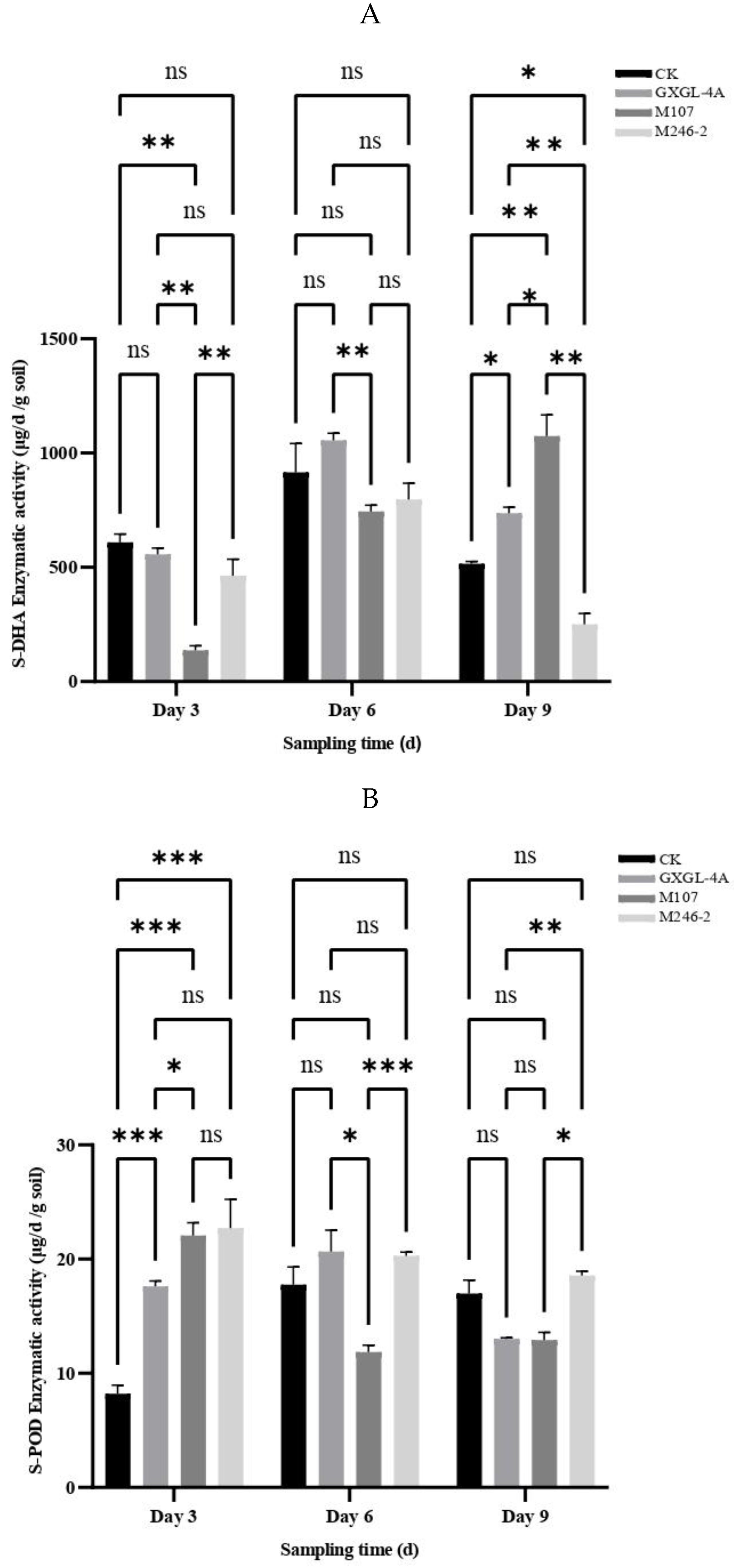
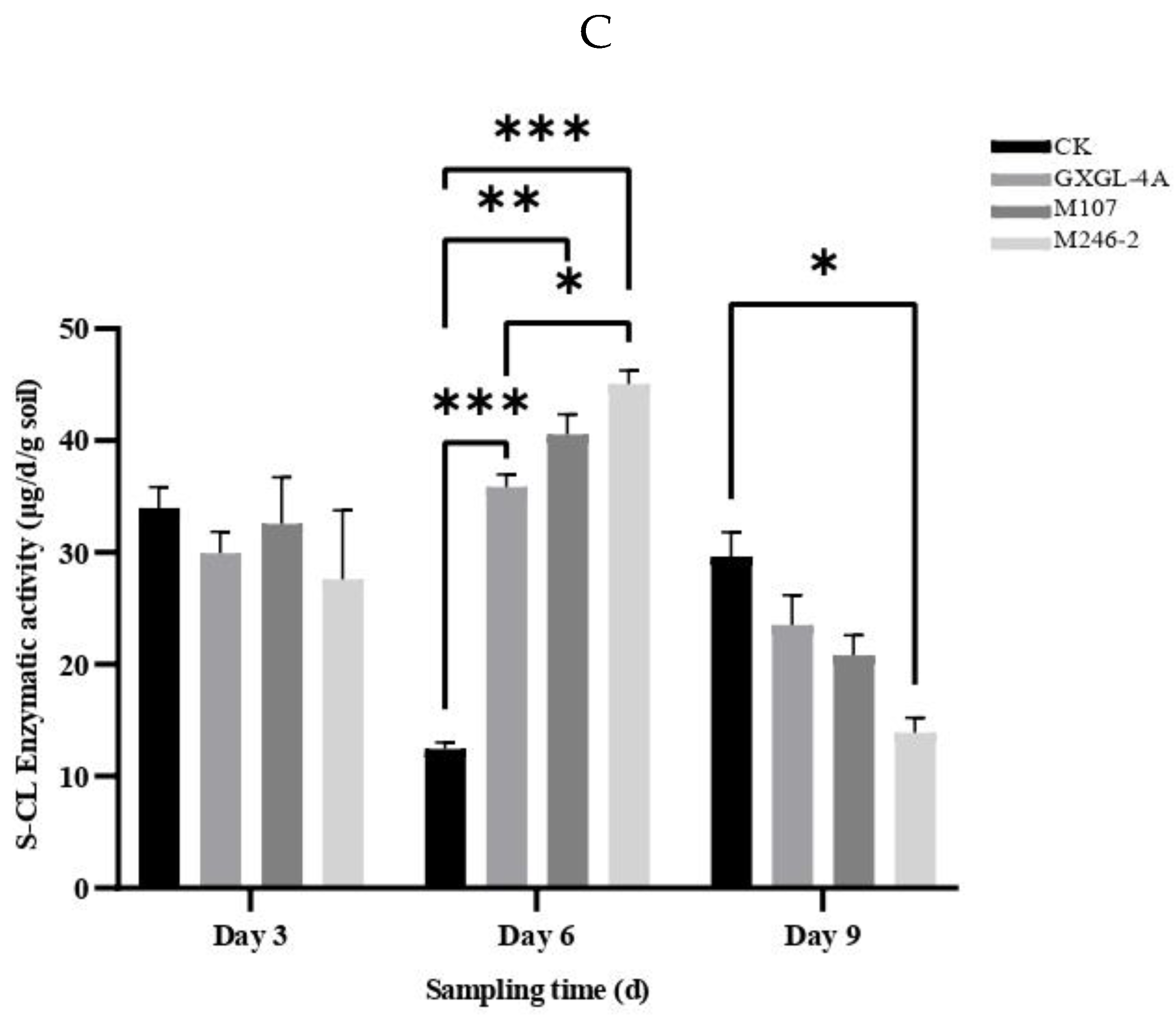
Figure 7.
Relative abundance of the dominant genus in cucumber rhizosphere soils on the 3rd, 6th, and 9th days after the application of NFB strains. The soil samples sampled on the 3rd, 6th, and 9th days after treatment with sterile water in the control groups were represented by CK_T1, CK_T2 and CK_T3, respectively. The soil samples collected in the GXGL-4A treatment groups on the 3rd, 6th and 9th days after fertilization were referred to as 4A_T1, 4A_T2 and 4A_T3, respectively. Similarly, the soil samples sampled in the M107, and M246-2 treatment groups are recorded as M107_T1, M107_T2, M107_T3, M246-2_T1, M246-2_T2 and M246-2_T3, respectively.
Figure 7.
Relative abundance of the dominant genus in cucumber rhizosphere soils on the 3rd, 6th, and 9th days after the application of NFB strains. The soil samples sampled on the 3rd, 6th, and 9th days after treatment with sterile water in the control groups were represented by CK_T1, CK_T2 and CK_T3, respectively. The soil samples collected in the GXGL-4A treatment groups on the 3rd, 6th and 9th days after fertilization were referred to as 4A_T1, 4A_T2 and 4A_T3, respectively. Similarly, the soil samples sampled in the M107, and M246-2 treatment groups are recorded as M107_T1, M107_T2, M107_T3, M246-2_T1, M246-2_T2 and M246-2_T3, respectively.
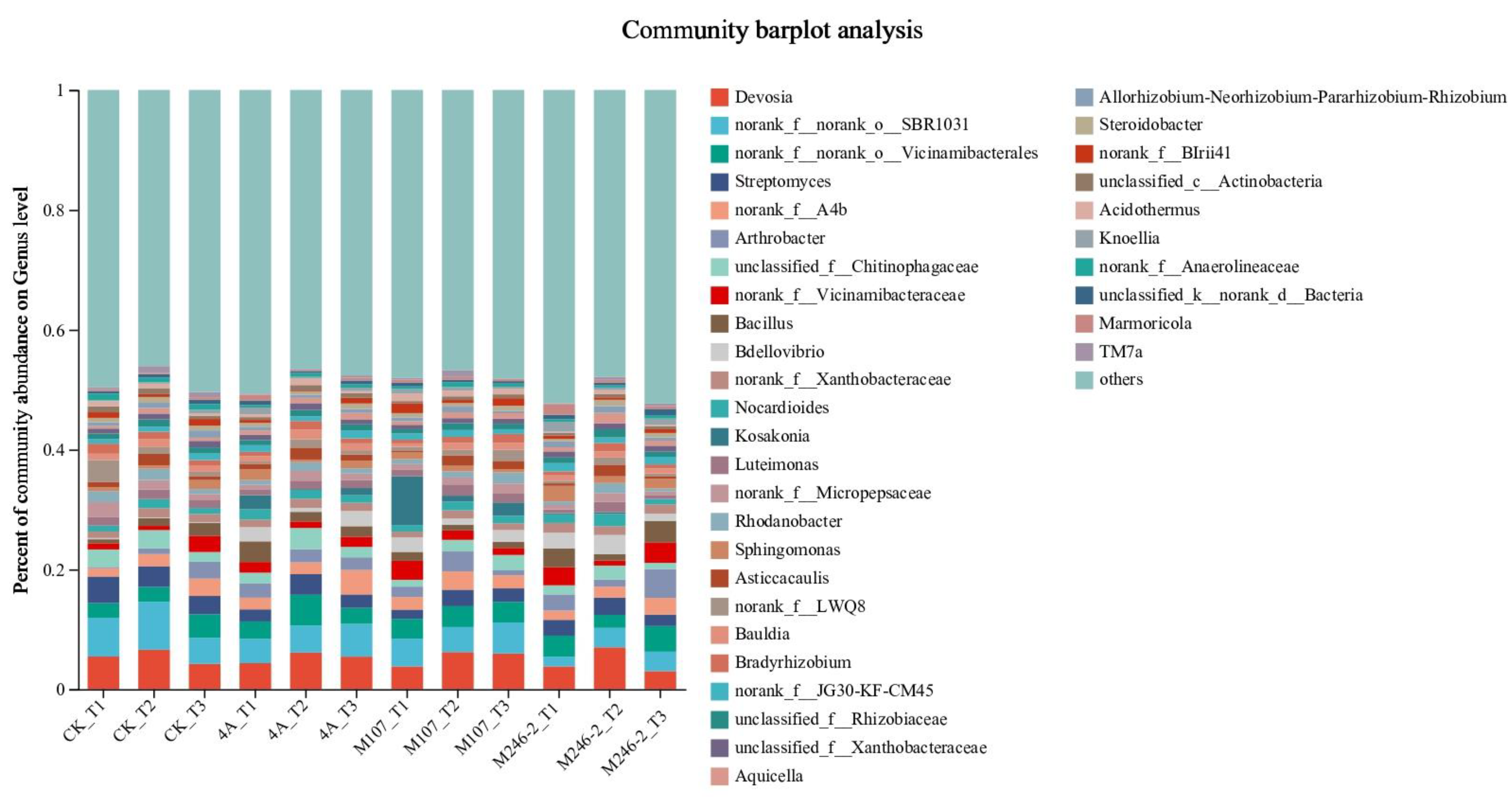
Figure 8.
Relative abundance of the dominant phyla in cucumber rhizosphere soils on the 3rd, 6th, and 9th days after the application of NFB strains. The soil samples sampled on the 3rd, 6th, and 9th days after treatment with sterile water in the control groups were represented by CK_T1, CK_T2 and CK_T3, respectively. The soil samples collected in the GXGL-4A treatment groups on the 3rd, 6th, and 9th days after fertilization were referred as 4A_T1, 4A_T2 and 4A_T3, respectively. Similarly, the soil samples sampled in the M107, and M246-2 treatment groups are recorded as M107_T1, M107_T2, M107_T3, M246-2_T1, M246-2_T2 and M246-2_T3, respectively.
Figure 8.
Relative abundance of the dominant phyla in cucumber rhizosphere soils on the 3rd, 6th, and 9th days after the application of NFB strains. The soil samples sampled on the 3rd, 6th, and 9th days after treatment with sterile water in the control groups were represented by CK_T1, CK_T2 and CK_T3, respectively. The soil samples collected in the GXGL-4A treatment groups on the 3rd, 6th, and 9th days after fertilization were referred as 4A_T1, 4A_T2 and 4A_T3, respectively. Similarly, the soil samples sampled in the M107, and M246-2 treatment groups are recorded as M107_T1, M107_T2, M107_T3, M246-2_T1, M246-2_T2 and M246-2_T3, respectively.
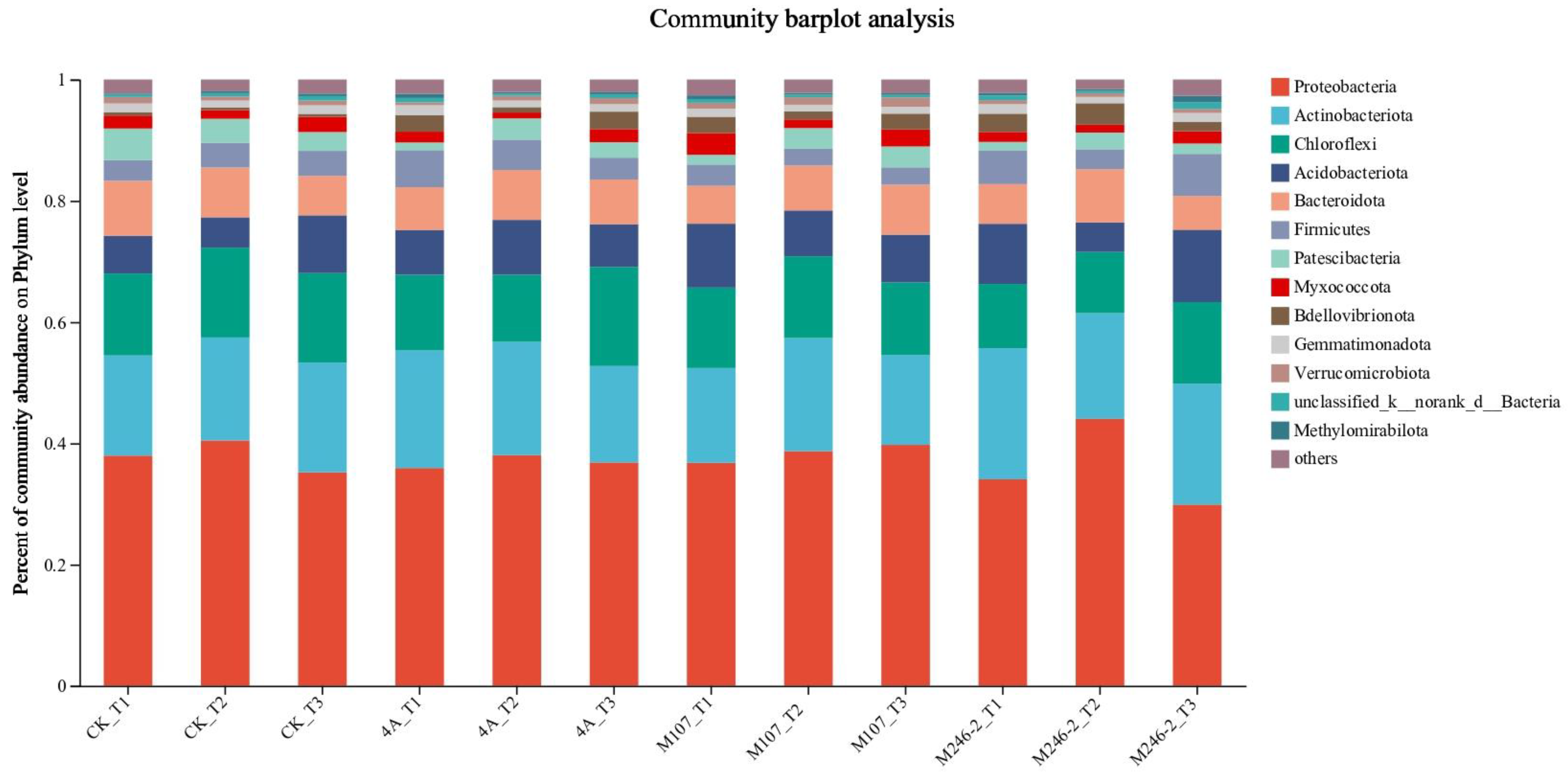
Figure 9.
The non-metric multidimensional scale (NMDS) analysis (stress: 0.101, R=0.7357, P=0.001) at the OTU level. The soil samples sampled on the 3rd, 6th, and 9th days after treatment with sterile water in the control groups were represented by CK_T1, CK_T2 and CK_T3, respectively. The soil samples collected in the GXGL-4A treatment groups on the 3rd, 6th, and 9th days after fertilization were referred as 4A_T1, 4A_T2 and 4A_T3, respectively. Similarly, the soil samples sampled in the M107, and M246-2 treatment groups are recorded as M107_T1, M107_T2, M107_T3, M246-2_T1, M246-2_T2 and M246-2_T3, respectively.
Figure 9.
The non-metric multidimensional scale (NMDS) analysis (stress: 0.101, R=0.7357, P=0.001) at the OTU level. The soil samples sampled on the 3rd, 6th, and 9th days after treatment with sterile water in the control groups were represented by CK_T1, CK_T2 and CK_T3, respectively. The soil samples collected in the GXGL-4A treatment groups on the 3rd, 6th, and 9th days after fertilization were referred as 4A_T1, 4A_T2 and 4A_T3, respectively. Similarly, the soil samples sampled in the M107, and M246-2 treatment groups are recorded as M107_T1, M107_T2, M107_T3, M246-2_T1, M246-2_T2 and M246-2_T3, respectively.
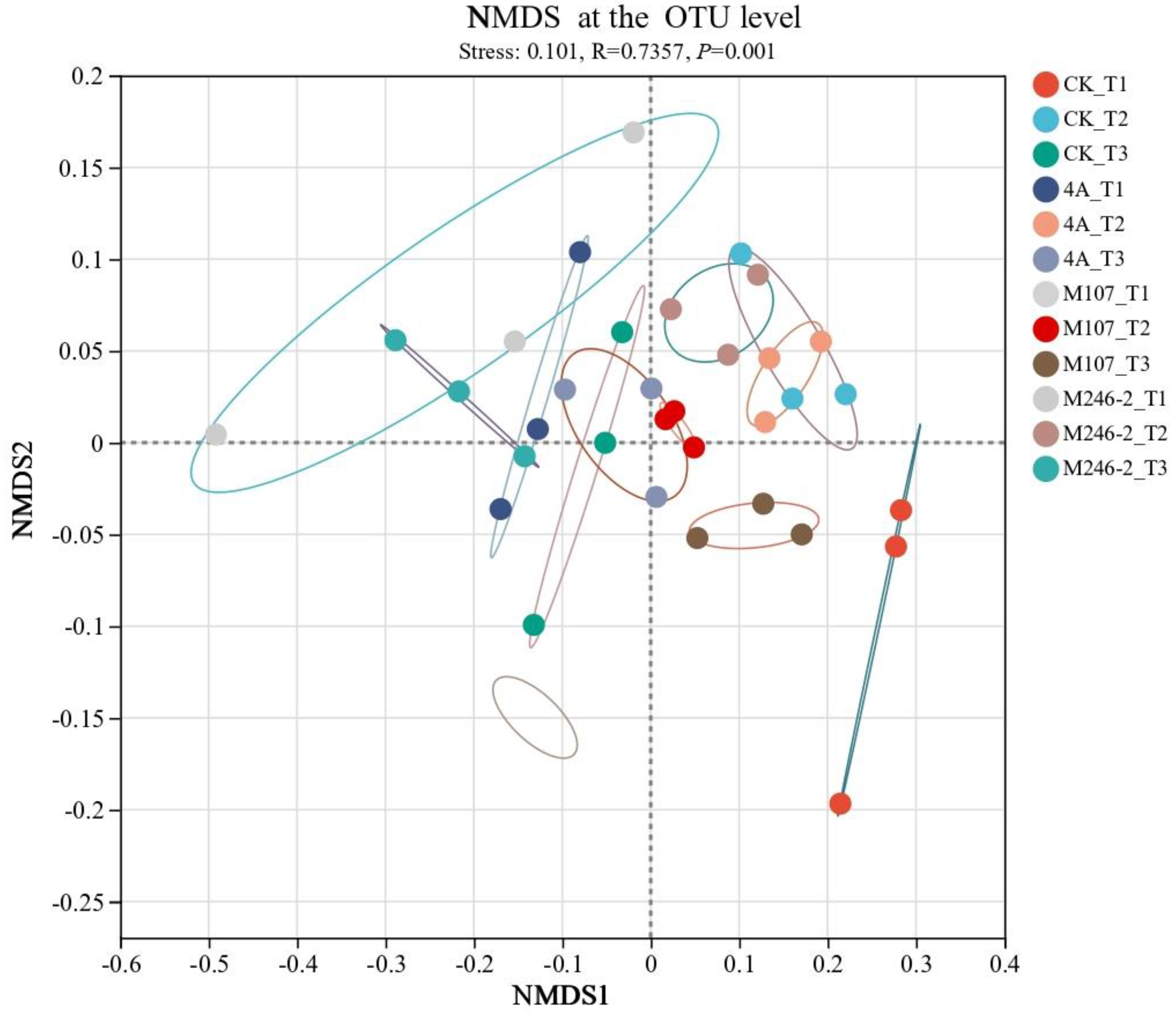
Figure 10.
Significant differences were analyzed at the phylum (A) and genus (B) levels between treatment groups. One asterisk (*) indicates a significant difference in the relative abundance of microbial species (P<0.05). The multiple testing correction method for P values was the false discovery rate (FDR).
Figure 10.
Significant differences were analyzed at the phylum (A) and genus (B) levels between treatment groups. One asterisk (*) indicates a significant difference in the relative abundance of microbial species (P<0.05). The multiple testing correction method for P values was the false discovery rate (FDR).
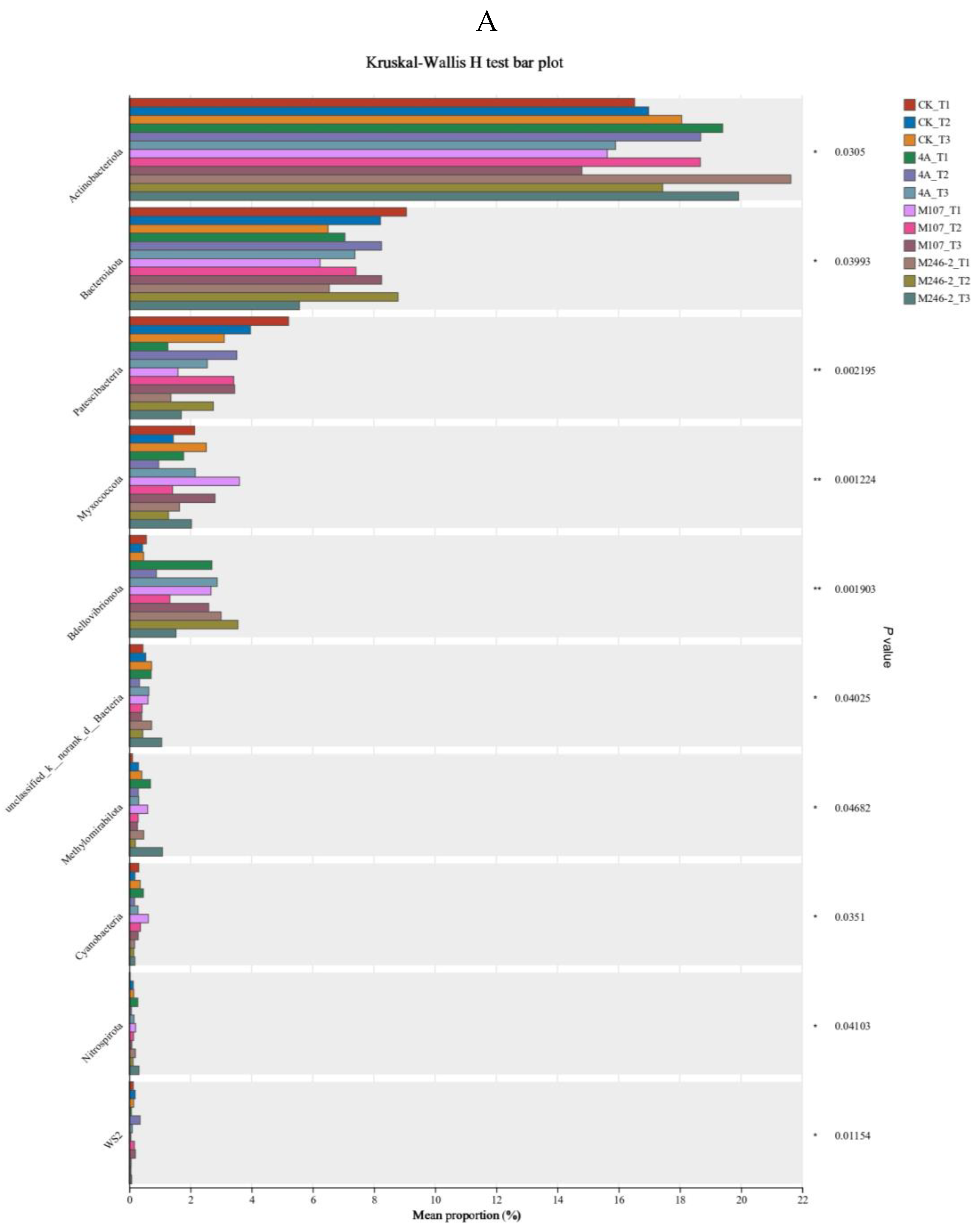
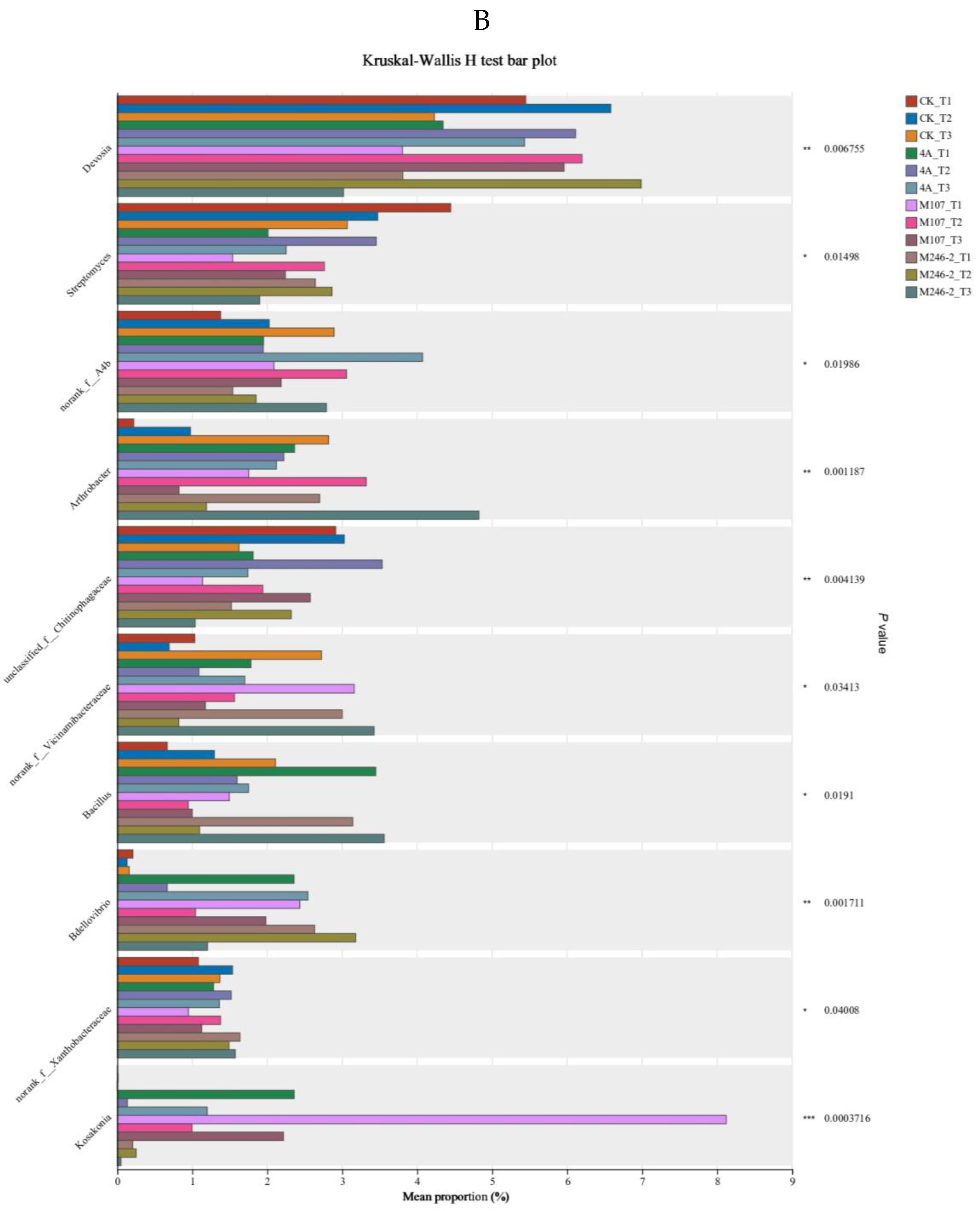

Table 1.
The abundance and diversity indices of the bacterial community in the NFB treatment cucumber rhizosphere soils.
Table 1.
The abundance and diversity indices of the bacterial community in the NFB treatment cucumber rhizosphere soils.
| Treatments | Ace index | Chao 1 index | Shannon index | Sobs | |
| Day 3 | |||||
| CK-T1 | 3607.64±524.56 b | 3492.49±439.99 b | 6.07±0.32 b | 2680.67±366.85 b | |
| 4A-T1 | 4844.78±307.74 a | 4616.06±227.80 a | 6.56±0.08 a | 3522.67±169.09 a | |
| M107-T1 | 5204.10±199.79 a | 4967.41±178.73 a | 6.50±0.20 a | 3710.67±164.81 a | |
| M246-2-T1 | 4824.56±144.89 a | 4524.58±158.61 a | 6.56±0.16 a | 3507.33±131.96 a | |
| Day 6 | |||||
| CK-T2 | 4167.03±345.49 ab | 3976.52±310.54 ab | 6.03±0.31 a | 2913.67±277.03 ab | |
| 4A-T2 | 4007.96±338.46 b | 3862.98±305.97 b | 6.05±0.09 a | 2807.00±189.61 b | |
| M107-T2 | 4760.31±6.17 a | 4514.90±53.70 a | 6.35±0.03 a | 3337.33±55.79 a | |
| M246-2-T2 | 4340.41±450.89 ab | 4067.11±357.73 ab | 6.24±0.11 a | 3081.33±276.96 ab | |
| Day 9 | |||||
| CK-T3 | 5197.03±181.39 a | 4910.15±194.28 a | 6.63±0.03 a | 3703.67±80.75 a | |
| 4A-T3 | 4887.19±249.69 ab | 4595.86±210.80 ab | 6.45±0.23 ab | 3444.00±203.74 ab | |
| M107-T3 | 4531.32±292.67 b | 4302.62±257.56 b | 6.28±0.19 b | 3193.00±229.69 b | |
| M246-2-T3 | 5179.48±125.12 a | 4884.79±45.97 a | 6.74±0.05 a | 3751.67±67.72 a |
Note: The soil samples of the CK group sampled on Day 3, Day 6, and Day 9 after sterile water treatment were represented by CK-T1, CK-T2 and CK-T3, respectively. Soil samples sampled on Day 3, Day 6 and Day 9 after GXGL-4A treatment were recorded as 4A-T1, 4A-T2 and 4A-T3, respectively. Soil samples sampled on Day 3, Day 6 and Day 9 after M107 treatment are showed as M107-T1, M107-T2 and M107-T3, respectively. Soil samples sampled on Day 3, Day 6 and Day 9 after GXGL-4A treatment were recorded as 4A-T1, 4A-T2 and 4A-T3, respectively. Sobs, Chao 1, Shannon and Ace indices based on OTUs level were shown. Lowercase letter indicate that there is a significant difference in α-diversity index between two groups by t-test (P<0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



