
Submitted:
11 February 2026
Posted:
11 February 2026
You are already at the latest version
Abstract
Colorectal cancer is a heterogeneous malignancy characterized by alterations in onco-genic signaling pathways and epigenetic mechanisms involved in gene regulation. Ab-errant activation of the Wnt/β-catenin pathway represents a central molecular event in colorectal tumorigenesis, while histone-associated epigenetic modifications may contribute to tumor progression and variability. This study aimed to investigate the relationship between Wnt pathway activation and histone H3 lysine 27 trimethylation in colorectal cancer and to examine their associations with clinicopathological and molecular characteristics. A retrospective observational study was performed on 83 colorectal adenocarcinoma cases using immunohistochemical evaluation of nuclear β-catenin and H3K27me3 expression in formalin-fixed, paraffin-embedded tumor samples, together with molecular analysis of KRAS, NRAS, and BRAF mutations and microsatellite instability status. Nuclear β-catenin expression was observed in 39.8% of cases, while H3K27me3 exhibited negative, mosaic, or diffuse nuclear staining pat-terns. Nuclear β-catenin expression was significantly associated with patient sex and age, whereas H3K27me3 expression patterns were significantly associated with tumor location, histological grade, disease stage, and metastatic status. These results indicate that Wnt pathway activation and H3K27me3-associated epigenetic alterations fre-quently coexist in colorectal cancer and support the value of integrated molecular and epigenetic assessment.
Keywords:
colorectal cancer
; Wnt/β-catenin pathway
; H3K27me3
; epigenetic
1. Introduction
Colorectal cancer represents the third most frequently diagnosed malignancy worldwide as well as in Romania, with nearly two million new cases reported globally and approximately 13,000 new cases diagnosed annually in Romania. Colorectal cancer accounts, in Romania for almost 6,800 deaths each year, ranking as the second leading cause of cancer-related mortality [1,2,3]. Research have focused on identifying the genetic and epigenetic changes associated with colorectal carcinogenesis.
Among the best studied oncogenic drivers are mutations that hyperactivate two central signaling cascades: the Ras–Raf–MEK–ERK and the PI3K/AKT/mTOR pathways [4,5,6,7]. These pathways regulate essential processes such as proliferation, apoptosis evasion, and metabolic adaptation. Therefore, genomic studies have reported high mutation frequencies in critical components of these pathways, with KRAS mutations identified in approximately 30–50% of colorectal cancers, BRAF mutations in 10–15%, and PIK3CA mutations in 10–20% of cases [8,9,10,11].
Moreover, aberrant activation of the Wnt/β-catenin pathway is a key molecular feature of colorectal cancer. The Wnt signaling pathway is divided into two major categories based on the involvement of β-catenin: the non-canonical pathway, which regulates cell polarity and cell migration through the planar cell polarity (PCP) and Wnt/Ca2+ pathways, and the canonical Wnt/β-catenin pathway. Under physiological conditions, β-catenin is predominantly localized in the cytoplasm, where its levels are tightly regulated by a multiprotein destruction complex composed of Axin, adenomatous polyposis coli (APC), glycogen synthase kinase-3 (GSK-3), and casein kinase 1 (CK1). Upon binding of Wnt ligands to Frizzled receptors and the low-density lipoprotein receptor–related protein 5/6 (LRP5/6) co-receptors, canonical Wnt signaling is activated. This leads to Dishevelled-mediated signaling, phosphorylation of LRP5/6, recruitment of Axin, and inhibition of the β-catenin destruction complex. As a result, β-catenin becomes stabilized, accumulates in the cytoplasm, and moves into the nucleus, where it interacts with TCF/LEF transcription factors to activate Wnt target genes. Persistent activation of this pathway supports cancer stem cell self-renewal and promotes invasion, metastasis, and resistance to therapy, thereby contributing to colorectal cancer progression [12,13,14].
Epigenetic regulation involves DNA methylation and histone modifications as the main mechanisms controlling gene expression. Histones are fundamental nuclear proteins that mediate DNA compaction, spatial organization, and functional regulation within chromatin. The nucleosome core particle is composed of histones H2A, H2B, H3, and H4, while histone H1 contributes to the stabilization of higher-order chromatin structures. These histone proteins undergo various post-translational modifications such as methylation, acetylation, and phosphorylation, which dynamically modulate chromatin architecture and determine transcriptional activity or repression [15,16].
Trimethylation of lysine 27 on histone H3 (H3K27me3) is a well-characterized epigenetic mark strongly associated with transcriptional silencing. The establishment of H3K27me3 is mediated by the PRC2, whose catalytic core contains the histone methyltransferase EZH2. EZH2 utilizes S-adenosylmethionine as a methyl donor to catalyze mono-, di-, and trimethylation of H3K27. Aberrant expression or enzymatic activity of EZH2 has been associated with pathological epigenetic repression of tumor suppressor genes and is frequently implicated in oncogenic processes, including colorectal cancer development and progression [17,18,19].
Aim and Scope
The main scope of the study is to investigate the coexistence of aberrant Wnt signaling activation and H3K27me3 associated epigenetic regulation in colorectal cancer on a population-based cohort. Additionally, the study aims to assess the association between nuclear β-catenin expression and key clinicopathological features, including age, sex, tumor location, histological subtype, grade, stage, and metastatic pattern. It also explores the relationship between H3K27me3 expression patterns and indicators of tumor aggressiveness in relation to disease stage, metastatic behavior, and molecular background, including KRAS, NRAS, BRAF mutational status, and MSI status. Furthermore, the study tries to identify independent predictors of nuclear β-catenin expression and H3K27me3 positivity.
2. Materials and Methods
2.1. Study Design and Patient Selection
The retrospective observational study included 83 patients diagnosed with colorectal adenocarcinoma. Cases were collected consecutively from the pathology archives of the Resident Laboratory, Oradea, Romania, during a time interval between 1st of January 2025 and 31 of October 2025. All cases had available FFPE tumor tissue suitable for immunohistochemical analysis. Clinicopathological data, including age, sex, tumor location, histological subtype, tumor grade, stage, metastatic pattern, MSI status, and mutational status (KRAS, NRAS, BRAF), were analyzed from pathology reports an medical records.
This study was conducted in full accordance with the ethical principles outlined in the Declaration of Helsinki and was approved by the Local Ethics Commission for Clinical and Research Developmental Studies, Resident Laboratory, Oradea, Romania (approval number 11, issued on December 20, 2024).
2.1.1. Inclusion Criteria:
Patients were included if they had a histopathological diagnosis of colorectal adenocarcinoma and available formalin-fixed, paraffin-embedded tumor tissue suitable for immunohistochemical analysis, along with complete molecular characterization, including MSI status and KRAS, NRAS mutational analysis. Only cases with successful immunohistochemical staining for β-catenin and H3K27me3, supported by appropriate internal and external controls, and with adequate tissue preservation allowing reliable evaluation of staining patterns and H-score assessment were included.
2.1.2. Exclusion Criteria:
Cases were excluded if tissue fixation or paraffin embedding was inadequate or if technical failure or suboptimal immunohistochemical staining occurred, including the absence of appropriate controls. Additional exclusion criteria included tissue processing artifacts or insufficient tumor material that interfered with accurate interpretation of β-catenin or H3K27me3 expression.
2.2. Immunohistochemical Staining;
Immunohistochemical staining for β-catenin and H3K27me3 was performed on formalin-fixed, paraffin-embedded tumor tissue sections using monoclonal antibodies. β-catenin expression was evaluated using a mouse monoclonal anti–β-catenin antibody (β-catenin-1, Dako), a well-established marker for assessing Wnt pathway activation through nuclear localization of β-catenin [20,21]. Wnt signaling was considered active when nuclear hot spot β-catenin staining was identified in more than 5% of tumor cell nuclei. H3K27me3 expression was assessed using a rabbit monoclonal antibody (RBT-H3K27me3, Bio SB) and interpreted based on nuclear staining patterns, classified as negative, mosaic, or diffuse positive.
Staining procedures were performed on the Dako Omnis platform in accordance with the manufacturer’s instructions [22]. Appropriate internal and external controls were included in each staining run to ensure technical validity and reproducibility. For H3K27me3, normal colonic mucosa served as an internal negative control, and external control slides were processed in parallel; in our setting, both internal and external controls for normal colonic mucosa consistently showed negative staining, rather than weak diffuse positivity as reported in some studies. All immunohistochemical slides were independently evaluated by two experienced pathologists who were blinded to the clinical and molecular data and stratified the slides based on hot spot region in negative, mosaic and diffuse pattern.
Nuclear β-catenin immunoreactivity was assessed by estimating the percentage of tumor cells exhibiting unequivocal nuclear staining. In accordance with criteria commonly reported in the literature, a 5% cut-off was applied to define nuclear positivity, such that cases with ≥5% (or >5%) of tumor cell nuclei stained were classified as positive. This threshold has been widely used to distinguish tumors with biologically relevant activation of the Wnt/β-catenin signaling pathway from cases showing only occasional or focal nuclear staining. This cut-off ensures methodological consistency and facilitates comparability with previously published studies [23,24].
2.3. KRAS, NRAS, BRAF Genotyping:
KRAS and NRAS genotyping was performed using a real-time polymerase chain reaction (PCR) assay, the AmoyDx® KRAS/NRAS Mutations Detection Kit (Amoy Diagnostics, Xiamen, China; Cat. No. ADx KRAS/NRAS-32), which detects 32 hotspot mutations across codons 12, 13, 59, 61, 117, and 146. DNA was extracted from FFPE tumor samples using the QIAamp DNA FFPE Tissue Kit (Qiagen, Hilden, Germany; Cat. No. 56404), and amplification was carried out on an ABI 7500 Fast Real-Time PCR System (Applied Biosystems) according to the manufacturer’s protocol. Mutation calls were automatically generated by the AmoyDx proprietary software using predefined Ct thresholds [25].
2.4. Microsatellite Instability Analysis:
MSI status was determined using a five-marker mononucleotide PCR panel (BAT-25, BAT-26, NR-21, NR-24, MONO-27; Promega MSI Analysis System v1.2) [26]. DNA was extracted from FFPE tumor tissue using the QIAamp DNA FFPE Tissue Kit (Qiagen, Cat. No. 56404), according to the manufacturer’s protocol. PCR reactions were prepared in a 25-μL volume with standard reagent concentrations. PCR amplification was performed with an initial denaturation at 95°C for 10 min, followed by 35 cycles of denaturation at 95°C for 30 s, annealing at 55°C for 30 s, and extension at 72°C for 30 s, with a final extension at 72°C for 10 min. Amplicons were analyzed by capillary electrophoresis on an ABI Prism 3130xl Genetic Analyzer, and fragment analysis was performed using GeneMapper software v5.0. Tumors were classified as MSI-H if instability was present in ≥2 loci, MSI-L if present in one locus, and MSS if no instability was observed, according to established guidelines [26,27] .
2.5. Statistical Analysis:
Statistical analyses were performed using Statistical Package of Social Science (SPSS), version 26. Categorical variables were summarized as frequencies and percentages. Associations between β-catenin expression, H3K27me3 patterns, and clinicopathological or molecular variables were evaluated using chi-square or Fisher’s exact tests, as appropriate.
Multivariate binary logistic regression models were constructed to identify independent predictors of nuclear β-catenin expression and H3K27me3 positivity (mosaic/diffuse vs. negative). Odds ratios (ORs) with 95% confidence intervals (CIs) were calculated. A p-value < 0.05 was considered statistically significant.
3. Results
The study cohort included 83 patients with colorectal adenocarcinoma, predominantly male (61.4%), with most patients aged between 51 and 70 years. Tumors were more frequently located in the left colon (71.1%) and were mainly conventional adenocarcinomas (84.3), with a predominance of moderately differentiated tumors (G2, 85.5%). Advanced-stage disease was common, with stage IV tumors representing 67.5% of cases and hepatic involvement being the most frequent metastatic site. Nuclear β-catenin expression, indicative of active Wnt signaling, was observed in 39.8% of cases, while H3K27me3 expression showed negative or mosaic patterns in the majority of tumors.

Table 1.
Clinicopathological, Molecular, and Immunohistochemical Characteristics of the Study Cohort (n = 83).
Table 1.
Clinicopathological, Molecular, and Immunohistochemical Characteristics of the Study Cohort (n = 83).
| Variable | Category | n (%) |
| Sex | Male | 51 (61.4) |
| Female | 32 (38.6) | |
| Age (years) | 30–40 | 8 (9.6) |
| 41–50 | 8 (9.6) | |
| 51–60 | 23 (27.7) | |
| 61–70 | 24 (28.9) | |
| 71–80 | 20 (24.1) | |
| Tumor location | Right colon | 24 (28.9) |
| Left colon | 59 (71.1) | |
| Histological type | Adenocarcinoma | 70 (84.3) |
| Mucinous adenocarcinoma | 13 (15.7) | |
| Tumor grade | G1 | 8 (9.6) |
| G2 | 71 (85.5) | |
| G3 | 4 (4.8) | |
| Tumor stage | II | 4 (4.8) |
| III | 23 (27.7) | |
| IV | 56 (67.5) | |
| Metastatic status | None | 27 (32.5) |
| Hepatic | 38 (45.8) | |
| Pulmonary | 10 (12.0) | |
| Other or multiple sites | 8 (9.7) | |
| MSI status | MSS | 75 (90.4) |
| MSI-H | 8 (9.6) | |
| Gene mutation status | Wild-type | 21 (25.3) |
| KRAS | 46 (55.4) | |
| NRAS | 8 (9.6) | |
| BRAF | 8 (9.6) | |
| β-catenin expression | Membranous | 50 (60.2) |
| Nuclear (active Wnt) | 33 (39.8) | |
| H3K27me3 expression pattern | Negative | 42 (50.6) |
| Mosaic | 33 (39.8) | |
| Diffuse positive | 8 (9.6) |
Data are presented as number (percentage). β-catenin nuclear expression was considered indicative of active Wnt signaling when more than 5% of tumor cell nuclei showed immunoreactivity. H3K27me3 expression was classified based on nuclear staining patterns as negative, mosaic, or diffuse positive. MSI- microsatellite instability.
Significant associations were observed between clinicopathological variables and both β-catenin expression and H3K27me3 patterns. Nuclear β-catenin expression showed significant associations with sex and age group, being more frequent in female and younger patients. H3K27me3 expression patterns were significantly associated with tumor location, grade, stage, and metastatic status, indicating a relationship between epigenetic alterations and advanced disease characteristics. No significant associations were observed between β-catenin expression and tumor location, grade, stage, or metastatic status.
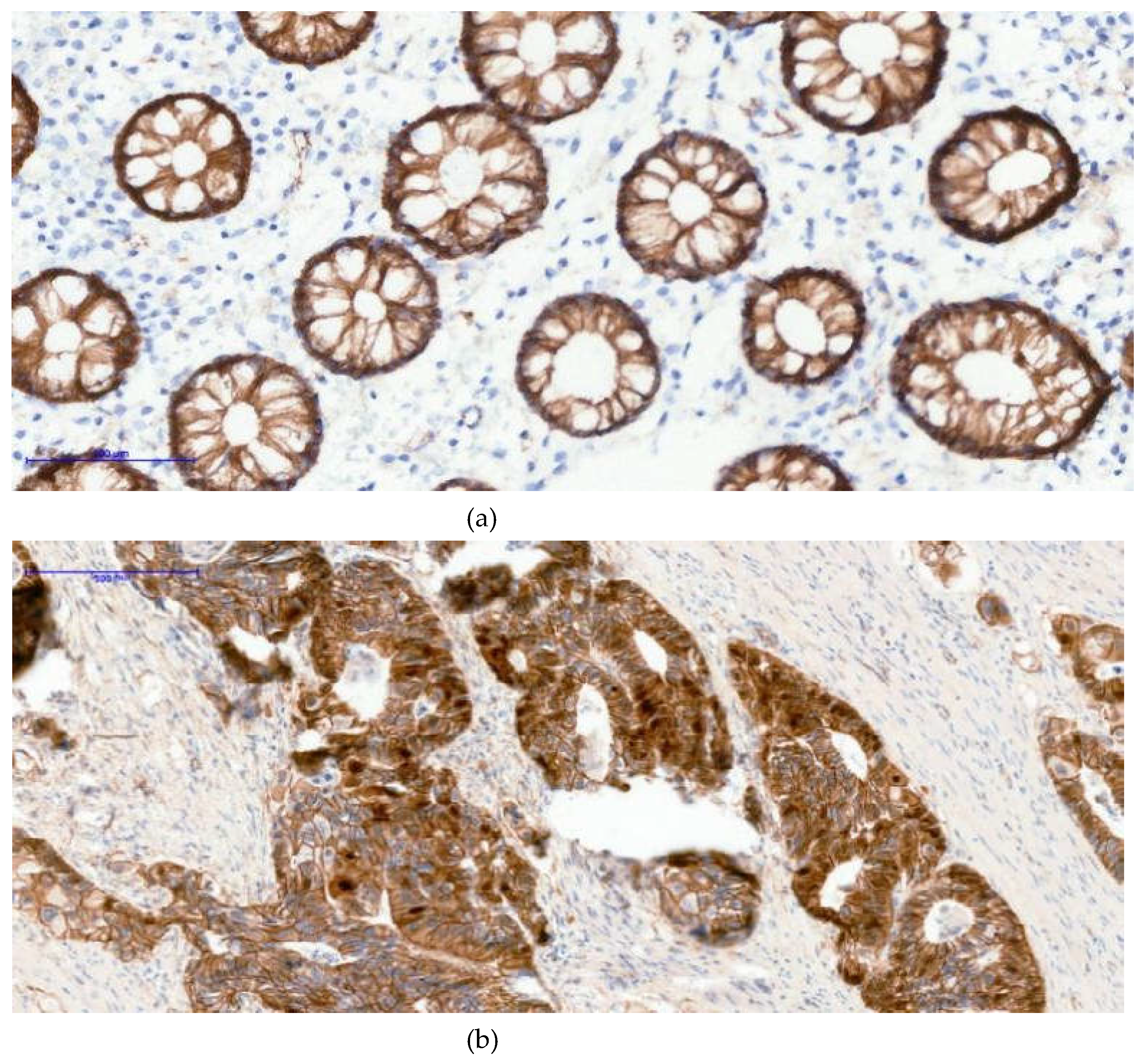
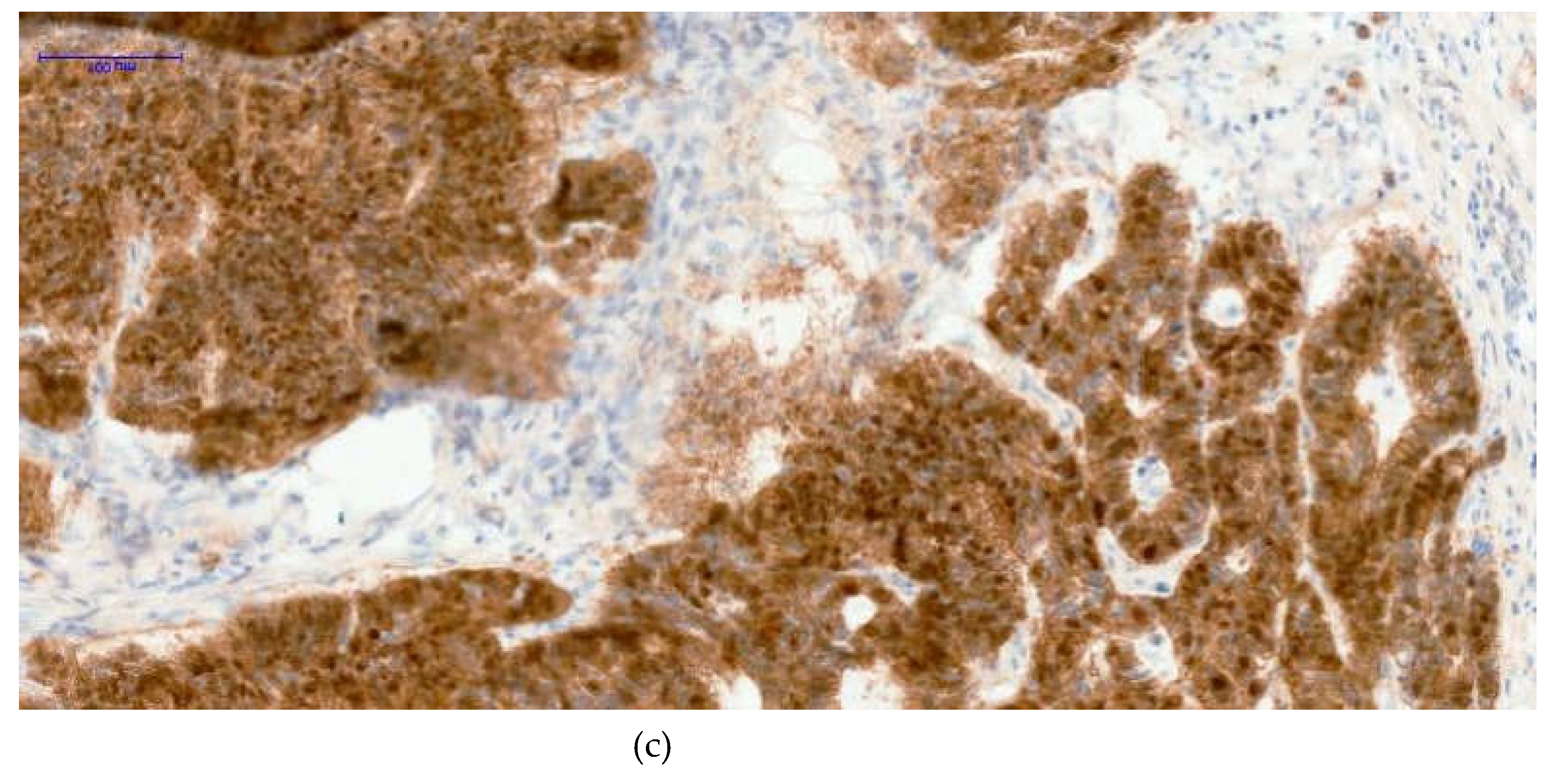
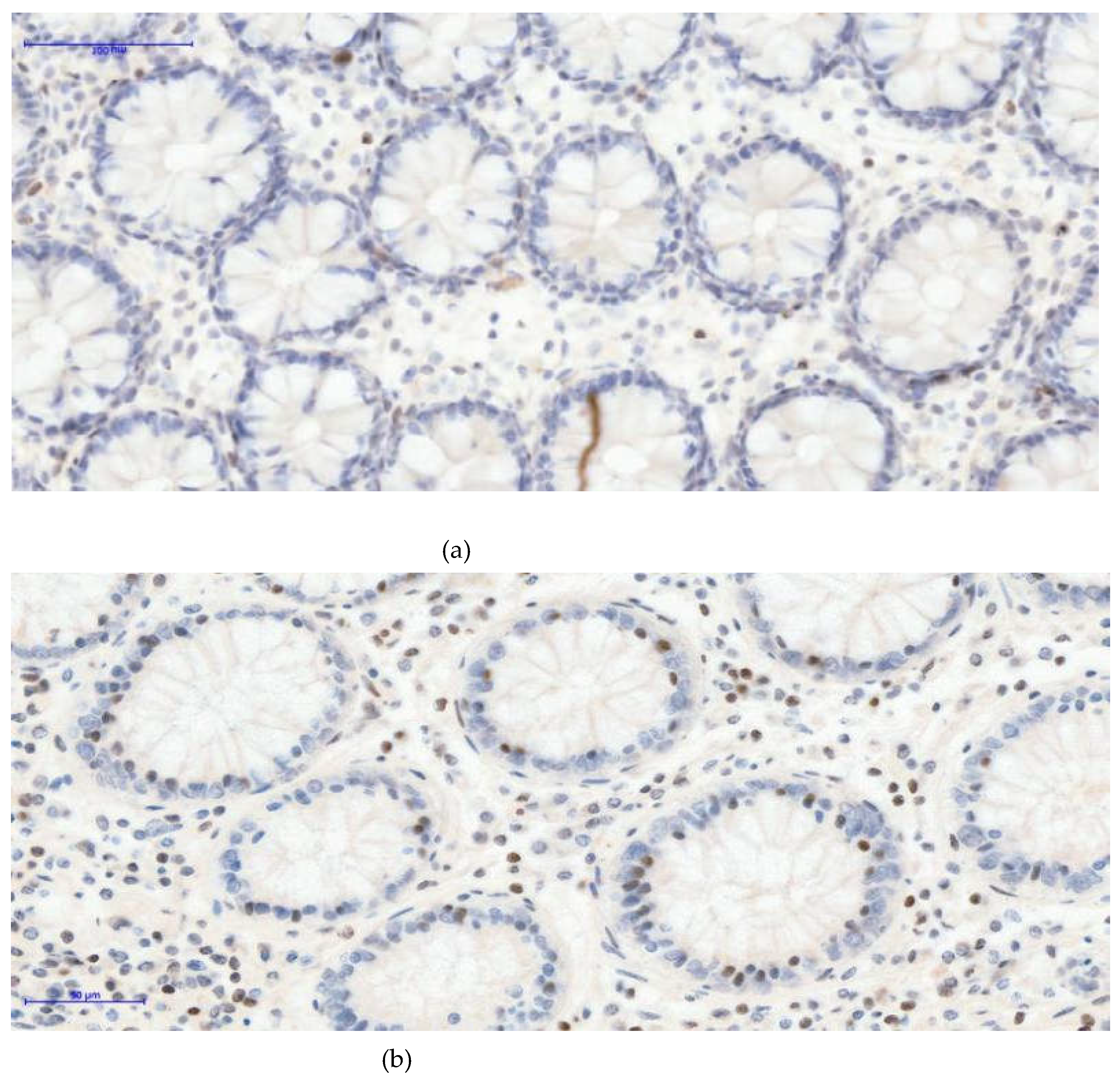
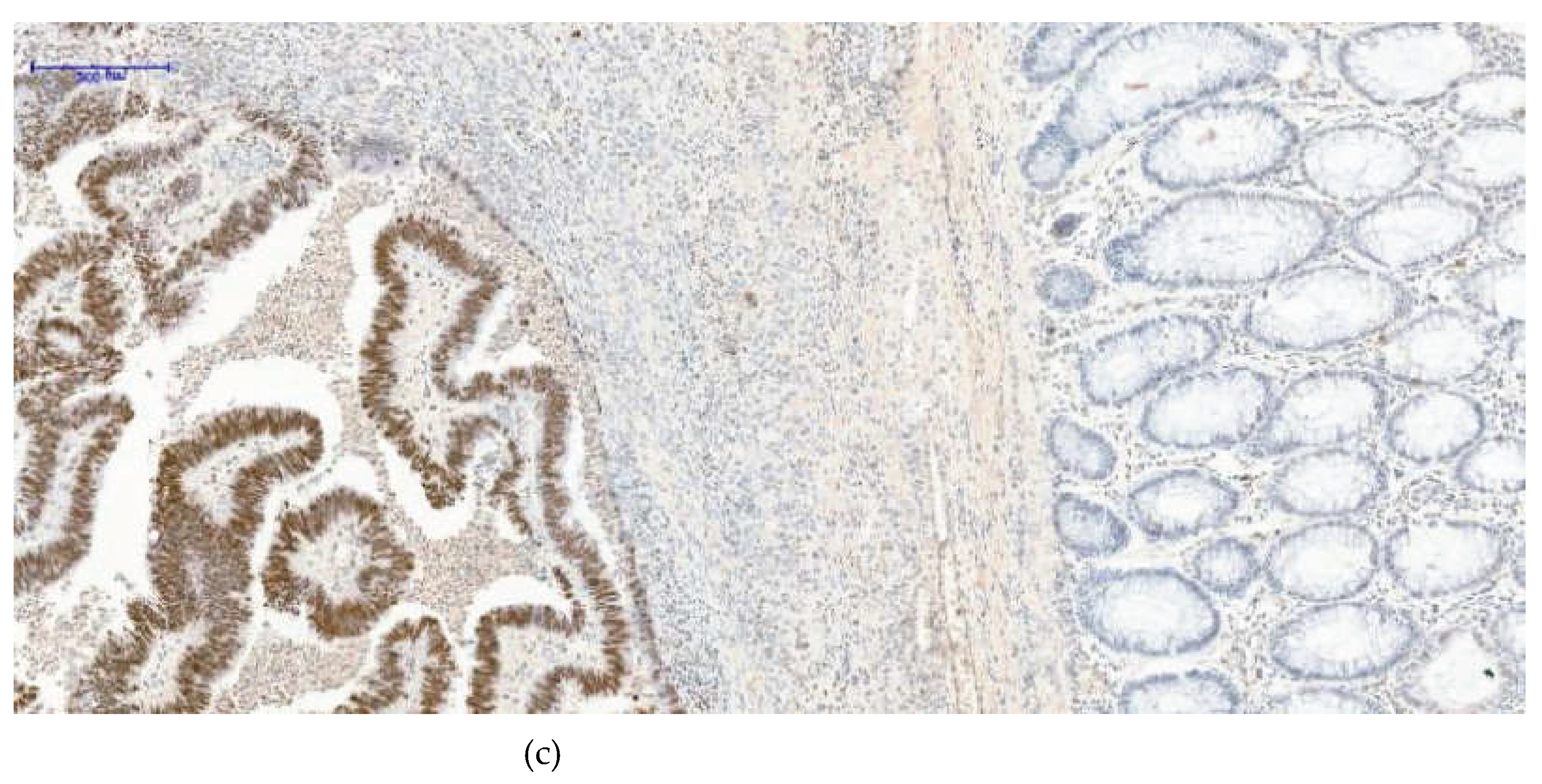
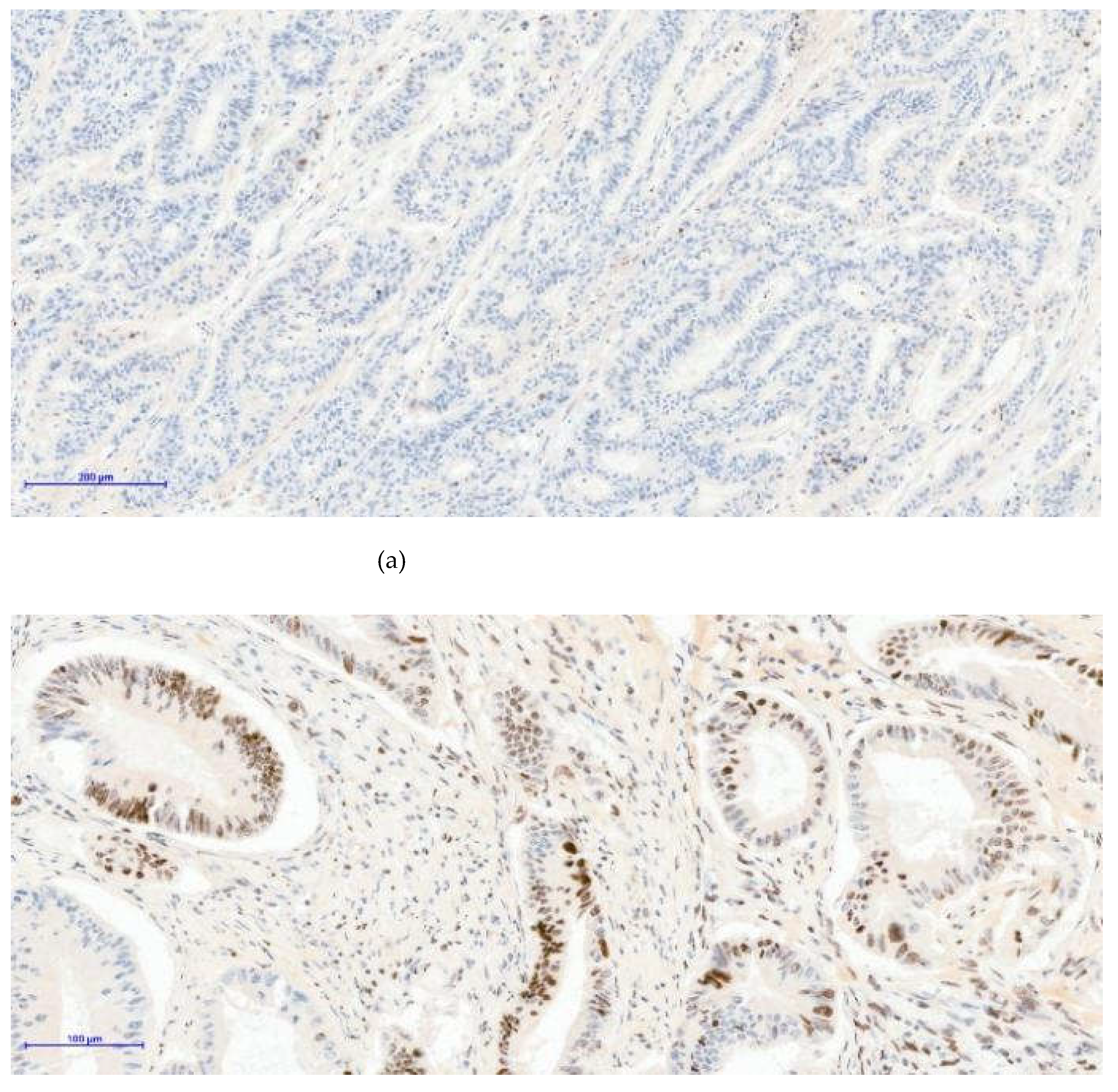
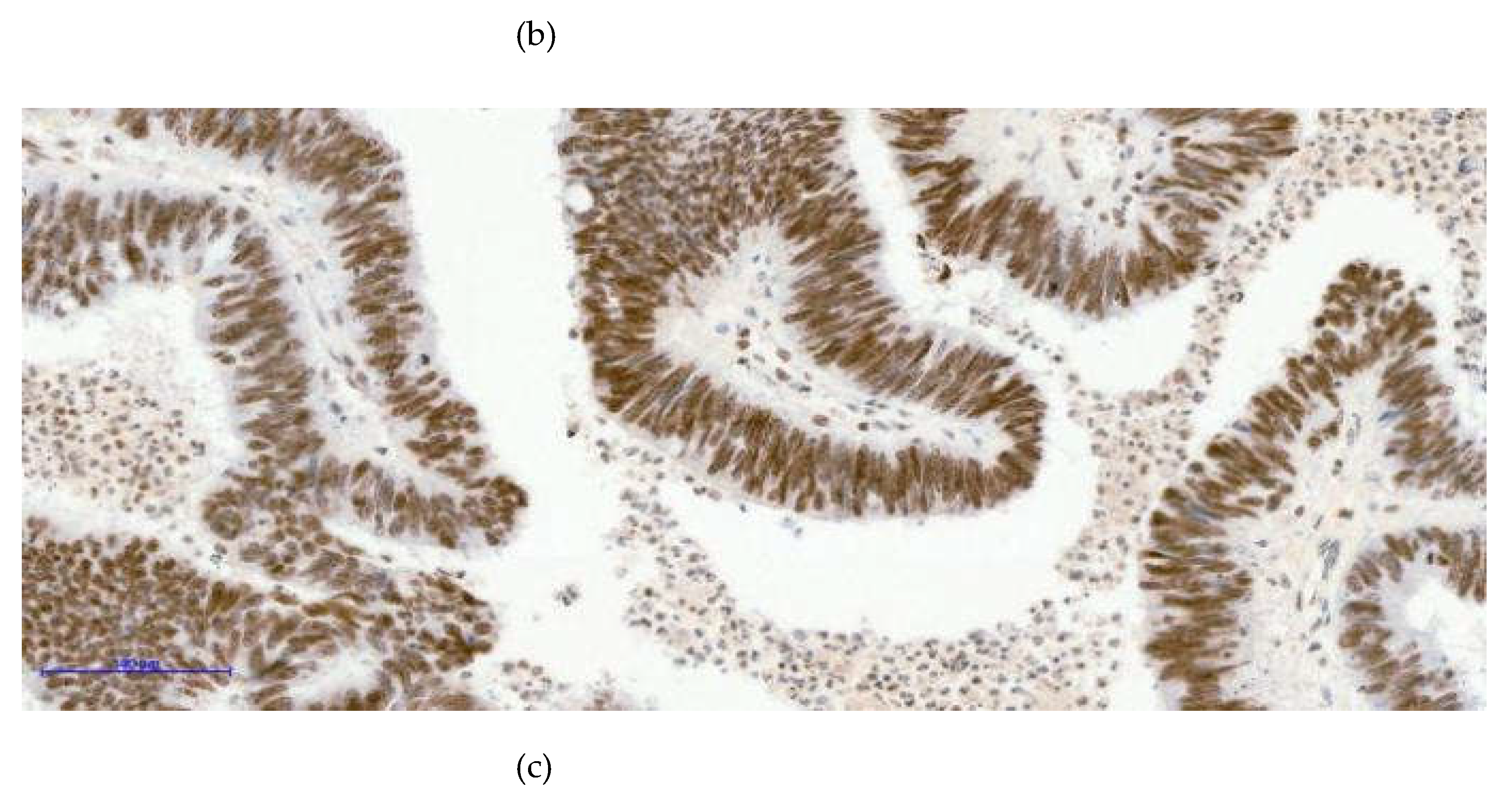
Table 2.
Associations between β-catenin Expression, H3K27me3 Patterns, and Clinicopathological Variables.
Table 2.
Associations between β-catenin Expression, H3K27me3 Patterns, and Clinicopathological Variables.
| Clinicopathological variable | Comparison (Chi-square test) | p-value |
| Sex | β-catenin (membranous vs. nuclear) | 0.001 |
| Sex | H3K27me3 pattern | 0.038 |
| Age group | β-catenin (membranous vs. nuclear) | <0.001 |
| Age group | H3K27me3 pattern | 0.007 |
| Tumor location | β-catenin (membranous vs. nuclear) | 0.446 |
| Tumor location | H3K27me3 pattern | 0.003 |
| Tumor grade | β-catenin (membranous vs. nuclear) | 0.220 |
| Tumor grade | H3K27me3 pattern | <0.001 |
| Tumor stage | β-catenin (membranous vs. nuclear) | 0.238 |
| Tumor stage | H3K27me3 pattern | <0.001 |
| Metastatic status | β-catenin (membranous vs. nuclear) | 0.105 |
| Metastatic status | H3K27me3 pattern | <0.001 |
| Histological type | β-catenin (membranous vs. nuclear) | 0.258 |
| Histological type | H3K27me3 pattern | 0.049 |
Bold values indicate statistically significant associations (p < 0.05). Associations were assessed using the Pearson chi-square test; Fisher’s exact test was applied when expected cell counts were <5. β-catenin nuclear expression was considered indicative of active Wnt signaling. H3K27me3 expression was classified as negative, mosaic, or diffuse positive.
Figure 1.
The images below illustrate external control staining for β-catenin in normal colonic mucosa, in which all epithelial gland nuclei remain negative (Image A), while the cell membranes and cytoplasm show physiological β-catenin immunoreactivity. The other two images (image B and C) demonstrate nuclear β-catenin positivity in tumor tissue; in case C, nearly all adenocarcinoma cell nuclei exhibit strong nuclear staining. (a)-β-catenin 20xOB- Control- negative nuclei; (b)- β-catenin 20xOB- Positive 30% nuclei; (c)- β-catenin 20xOB- Positive 90% nuclei.
Figure 1.
The images below illustrate external control staining for β-catenin in normal colonic mucosa, in which all epithelial gland nuclei remain negative (Image A), while the cell membranes and cytoplasm show physiological β-catenin immunoreactivity. The other two images (image B and C) demonstrate nuclear β-catenin positivity in tumor tissue; in case C, nearly all adenocarcinoma cell nuclei exhibit strong nuclear staining. (a)-β-catenin 20xOB- Control- negative nuclei; (b)- β-catenin 20xOB- Positive 30% nuclei; (c)- β-catenin 20xOB- Positive 90% nuclei.
Figure 2.
(a)- H3K27me3 External control 20XOB- negative glandular nuclei, some positive nuclear cells of lamina propria -Based on internal and external controls for H3K27me3, we observed that the nuclei of normal glandular epithelial cells remain negative, while nuclear staining is present in cells of the lamina propria. (b)- H3K27me3 External control 20XOB- negative glandular nuclei, more positive nuclear cells of lamina propria (inflammatory condition)- A higher density of inflammatory cells with nuclear positivity is evident, an inflammatory condition of the colon. In the (c)- H3K27me3 Internal control 10XOB- Positive tumoral nuclei (left side), negative epithelial nuclei normal glands (right side)- image, serving as an internal control, a clear contrast is observed: the tumor tissue on the left side shows strong diffuse nuclear positivity, whereas the adjacent normal colonic mucosa on the right side demonstrates complete absence of nuclear staining in the glandular epithelium.
Figure 2.
(a)- H3K27me3 External control 20XOB- negative glandular nuclei, some positive nuclear cells of lamina propria -Based on internal and external controls for H3K27me3, we observed that the nuclei of normal glandular epithelial cells remain negative, while nuclear staining is present in cells of the lamina propria. (b)- H3K27me3 External control 20XOB- negative glandular nuclei, more positive nuclear cells of lamina propria (inflammatory condition)- A higher density of inflammatory cells with nuclear positivity is evident, an inflammatory condition of the colon. In the (c)- H3K27me3 Internal control 10XOB- Positive tumoral nuclei (left side), negative epithelial nuclei normal glands (right side)- image, serving as an internal control, a clear contrast is observed: the tumor tissue on the left side shows strong diffuse nuclear positivity, whereas the adjacent normal colonic mucosa on the right side demonstrates complete absence of nuclear staining in the glandular epithelium.
Figure 3.
Three distinct nuclear staining patterns were observed in colorectal tumor samples: negative expression [image (a)- H3K27me3 10XOB- negative tumoral nuclei], a mosaic pattern characterized by heterogeneous nuclear positivity [image (b)- H3K27me3 20XOB- positive tumoral nuclei, mosaic pattern], and a strong diffuse pattern with uniform nuclear staining across tumor cells [image (c)- H3K27me3 20XOB- positive tumoral nuclei, diffuse pattern (100%)].
Figure 3.
Three distinct nuclear staining patterns were observed in colorectal tumor samples: negative expression [image (a)- H3K27me3 10XOB- negative tumoral nuclei], a mosaic pattern characterized by heterogeneous nuclear positivity [image (b)- H3K27me3 20XOB- positive tumoral nuclei, mosaic pattern], and a strong diffuse pattern with uniform nuclear staining across tumor cells [image (c)- H3K27me3 20XOB- positive tumoral nuclei, diffuse pattern (100%)].
As presented below, in Table 3, a significant association was identified between nuclear β-catenin expression and H3K27me3 expression patterns, supporting a relationship between Wnt pathway activation and epigenetic regulation. Nuclear β-catenin expression was also significantly associated with MSI status and with KRAS/NRAS/BRAF mutational status. H3K27me3 expression patterns showed significant associations with gene mutation status, whereas no significant associations were observed between H3K27me3 expression and MSI status.
Table 3.
Associations between β-catenin Expression, H3K27me3 Patterns, and Molecular Variables.
| Molecular variable | Comparison | p-value |
| β-catenin | H3K27me3 pattern | 0.002 |
| MMR status | β-catenin (membranous vs. nuclear) | 0.016 |
| MMR status | H3K27me3 pattern | 0.581 |
| Gene mutation status (KRAS/NRAS/BRAF) | β-catenin (membranous vs. nuclear) | 0.004 |
| Gene mutation status (KRAS/NRAS/BRAF) | H3K27me3 pattern | <0.001 |
Bold values indicate statistically significant associations (p < 0.05). Associations were assessed using the Pearson chi-square test; Fisher’s exact test was applied when expected cell counts were <5. β-catenin nuclear expression was considered indicative of active Wnt signaling. H3K27me3 expression was classified as negative, mosaic, or diffuse positive.
In multivariate binary logistic regression assessing predictors of nuclear β-catenin expression, female sex and younger age remained independently associated with nuclear/active β-catenin. Female patients had higher odds of nuclear β-catenin compared to males (OR 8.83, 95% CI 2.65–29.44; p = 0.0004), whereas age ≥60 years was associated with lower odds compared to <60 years (OR 0.20, 95% CI 0.06–0.65; p = 0.0077). Stage IV disease and the presence of metastasis showed non-significant trends toward increased odds of nuclear β-catenin; however, confidence intervals could not be reliably estimated due to sparse data in some categories.
Table 4.
Multivariate logistic regression for nuclear β-catenin expression.
| Predictor | OR | 95% CI | p-Value |
| Female sex | 8.83 | 2.65–29.44 | 0.0004 |
| Age ≥60 years | 0.20 | 0.06–0.65 | 0.0077 |
| Metastasis (any vs. none) | 2.07 | CI not reliably estimated* | n.s. |
| Stage (IV vs. II–III) | 2.53 | CI not reliably estimated* | n.s. |
Odds ratios (ORs) were estimated using logistic regression. CI denotes the 95% confidence interval. n.s. indicates lack of statistical significance (p ≥ 0.05). Confidence intervals for some predictors were not reliably estimable owing to small sample size and model instability.
In the second multivariate model evaluating predictors of H3K27me3 positivity (mosaic/diffuse vs. negative), nuclear β-catenin was identified as an independent predictor of H3K27me3 expression (OR 4.92, 95% CI 1.24–19.55; p = 0.024), therefore, cases with nuclear β-catenin expression had approximately five-fold higher odds of exhibiting a mosaic or diffuse H3K27me3 pattern compared with negative expression. Age ≥60 years and female sex were not significantly associated with H3K27me3 status, while MSI-H showed a non-significant positive association with wide confidence intervals, likely reflecting limited subgroup size.
Table 5.
Multivariate logistic regression for H3K27me3 positivity (mosaic/diffuse vs. negative).
| Predictor | OR | 95% CI | p-value |
| Nuclear β-catenin | 4.92 | 1.24–19.55 | 0.024 |
| Age ≥60 years | 0.43 | 0.10–1.75 | 0.237 |
| Female sex | 0.38 | 0.09–1.63 | 0.193 |
| MSI-H | 6.65 | 0.66–66.62 | 0.107 |
OR, odds ratio; CI, confidence interval; MSI-H, microsatellite instability–high. ORs were derived from multivariate logistic regression analysis. CI denotes the 95% confidence interval; p-values < 0.05 were considered statistically significant.
4. Discussion
The results of the present study should be interpreted with consideration of potential bias related to tumor grade, particularly in advanced-stage and high-grade (grade IV) colorectal carcinomas. These tumors are often underrepresented in retrospective cohorts and show increased biological and epigenetic heterogeneity, which may contribute to variability in immunohistochemical results and reduced statistical power. As a result, associations involving advanced disease stage or metastatic status may appear weaker or fail to reach statistical significance, reflecting limited subgroup size or within-group heterogeneity rather than the absence of a true biological relationship.
These limitations highlight the need for cautious interpretation of findings derived from high-grade tumors and support further validation in larger, stage-balanced cohorts. Future studies with prospective designs and more homogeneous clinical stratification may help clarify the role of Wnt/β-catenin signaling across different stages of colorectal cancer progression.
Colorectal cancer represents a biologically heterogeneous malignancy driven by intricate interactions between oncogenic signaling pathways and epigenetic regulatory mechanisms. Although aberrant activation of the Wnt/β-catenin pathway is a well-established hallmark of colorectal tumorigenesis, the contribution of histone-mediated epigenetic modifications remains comparatively underexplored. In particular, current evidence regarding histone H3 lysine 27 trimethylation (H3K27me3) in colorectal cancer is limited, with only a small number of studies examining its global distribution and intratumoral heterogeneity. This relative scarcity of data highlights the need for further systematic investigation into the crosstalk between oncogenic signaling pathways and histone-based epigenetic regulation in colorectal cancer.
Dysregulation of histone H3 lysine 27 trimethylation (H3K27me3) has been widely implicated in tumor progression and clinical prognosis across diverse malignancies. Prior studies have consistently demonstrated that reduced H3K27me3 expression is associated with adverse clinicopathological features, including increased tumor size, advanced disease stage, estrogen receptor negativity, and lymph node involvement in breast carcinoma, as well as higher tumor grade and stage in ovarian cancer and higher tumor grade in pancreatic cancer [17,28]. In concordance with these observations, the present study demonstrates a statistically significant association between advanced disease stage—reflected by increased tumor size—and altered H3K27me3 expression.
Previous studies have reported heterogeneous associations between nuclear β-catenin expression and clinicopathological characteristics in colorectal cancer. Gao et al. observed significant associations between nuclear β-catenin expression and TNM stage, lymph node involvement, and histological differentiation, while no significant correlations were identified with patient age or sex [29]. Hussein et al. reported significant relationships between nuclear β-catenin expression, patient age, lymph node involvement, and distant metastases [30]. Consistent with this variability, multivariate analysis identified female sex and younger age as independent predictors of nuclear β-catenin expression, suggesting that demographic factors may contribute to differential activation of the Wnt/β-catenin pathway in colorectal cancer. Although advanced disease stage and metastatic status were associated with increased odds of nuclear β-catenin accumulation, these associations did not reach statistical significance. This lack of statistical significance may reflect limited sample size or reduced statistical power within specific subgroups rather than the absence of a biological relationship. Accordingly, the observed patterns indicate variability in Wnt/β-catenin pathway activity across patient subgroups and disease contexts.
In the second multivariate model, nuclear β-catenin expression emerged as the strongest independent predictor of H3K27me3 positivity, supporting a close relationship between Wnt pathway activation and epigenetic regulation. The approximately five-fold increased likelihood of mosaic or diffuse H3K27me3 expression in tumors with nuclear β-catenin suggests a potential functional interplay between oncogenic signaling and chromatin-modifying mechanisms. To further contextualize these associations within a biological framework, it is relevant to consider the role of Polycomb-mediated epigenetic regulation in colorectal cancer. The PRC2 represents a major chromatin-regulatory system implicated in the maintenance of tissue homeostasis. PRC2 is composed of the core subunits EZH1/2, EED, SUZ12, and RBAP46/48, with EZH2 functioning as the catalytic lysine methyltransferase responsible for H3K27me3, a modification commonly associated with chromatin compaction and transcriptional repression [31,32,33]. Although EZH2 overexpression has been frequently reported in colorectal cancer, its functional implications remain incompletely defined and appear to be context dependent. Experimental evidence suggests that activation of Wnt/β-catenin signaling may locally attenuate PRC2-mediated H3K27me3 at Wnt target gene promoters, thereby permitting transcriptional activation. In contrast, sustained β-catenin signaling has been reported to coincide with broader alterations in the genomic distribution of H3K27me3, which may reflect epigenetic reprogramming processes occurring during tumor progression rather than direct causal mechanisms [32].
Although MSI-H status was associated with a non-significant increase in H3K27me3 expression, the wide confidence intervals necessitate cautious interpretation of this observation. This finding may reflect limited statistical power and highlights the need for further evaluation in larger, well-characterized cohorts, particularly given the scarcity of existing data addressing the relationship between microsatellite instability and H3K27me3-associated epigenetic regulation.
Data regarding global H3K27me3 expression in colorectal cancer remain limited in the literature, with only a small number of studies addressing this epigenetic mark[28,34]. An early investigation by Nakazawa et al. reported no significant differences in global nuclear H3K27me3 expression among normal colorectal mucosa, adenomas, and adenocarcinomas, nor across tumor differentiation grades or histological subtypes, suggesting relative stability of this epigenetic mark at a global level [34]. In contrast, the present findings describe distinct negative, mosaic, and diffuse nuclear H3K27me3 staining patterns, indicating a degree of epigenetic heterogeneity that may not be captured by global expression assessments alone. Notably, the mosaic pattern reflects intratumoral variability in chromatin regulation and supports the concept of dynamic epigenetic remodeling during tumor progression rather than a fixed epigenetic phenotype, with diffuse positivity potentially representing a more uniform and stabilized epigenetic state across tumor cells.
The strong association between nuclear β-catenin expression and mosaic or diffuse H3K27me3 patterns observed in multivariate analysis supports the concept of a coordinated relationship between Wnt pathway activation and epigenetic regulation in colorectal cancer. Therefore, the presence of heterogeneous H3K27me3 expression patterns may contribute to variability in tumor behavior and therapeutic response, although further studies are required to clarify their prognostic or predictive significance.
Accumulating evidence supports a complex and context-dependent relationship between Wnt/β-catenin signaling and PRC2-mediated epigenetic regulation. Activation of Wnt signaling and nuclear accumulation of β-catenin have been reported to locally antagonize PRC2 activity and reduce H3K27me3 deposition at specific Wnt target gene loci; however, this effect appears to be largely locus specific rather than global in nature [35]. In parallel, several studies have suggested that sustained β-catenin activation may be associated with broader epigenetic reprogramming, including alterations in PRC2 activity and redistribution of H3K27me3 toward alternative genomic regions, such as genes involved in differentiation and tumor suppression. In line with this concept, the present findings indicate that nuclear β-catenin and H3K27me3 can coexist within the same tumor samples, and importantly, that nuclear β-catenin acts as a significant predictive factor for H3K27me3 expression [16,36]. These observations are compatible with a model in which nuclear β-catenin localization is associated with a tumor context characterized by increased epigenetic plasticity, whereby β-catenin–driven transcriptional activity may coexist with compensatory or adaptive PRC2-mediated repression at other genomic loci. In this setting, β-catenin signaling and H3K27me3-dependent chromatin regulation appear to operate in a coordinated, though not necessarily mutually exclusive, manner during colorectal cancer progression.
Limitations of the Study
The presented study has several limitations. First, it has a retrospective design and relatively small sample size may have limited statistical power, particularly in subgroup analyses, resulting in wide confidence intervals in multivariate models. The single-population setting may also limit the generalizability of the findings. The observed associations between Wnt pathway activation and H3K27me3-associated epigenetic regulation suggest the existence of potential biological links between these processes. However, these results should be interpreted with caution, as the analyses were performed in a single laboratory, which may introduce methodological and technical limitations and affect the generalizability of the findings. This study may be limited by the underrepresentation and increased heterogeneity of advanced-stage, high-grade colorectal carcinomas, which could influence the strength of observed associations. Consequently, further validation through independent, multicenter cohorts, as well as functional and prospective studies, is required to confirm these observations and to establish a causal relationship.
5. Conclusions
In this study, activation of the Wnt/β-catenin pathway was frequently observed together with H3K27me3 expression in colorectal cancer. Tumors showed different H3K27me3 staining patterns, indicating that epigenetic regulation is not uniform across all tumor cells.
Nuclear β-catenin was independently associated with mosaic and diffuse H3K27me3 expression, suggesting that epigenetic changes may occur alongside Wnt pathway activation in colorectal tumors. This association may help explain differences observed between tumors and within individual tumors.
Although causal relationships cannot be established, these findings highlight the importance of evaluating epigenetic features together with key signaling pathways in colorectal cancer. Further studies are needed to better understand the biological and clinical relevance of these interactions.
Author Contributions
Conceptualization P.O, A.R and C.A.; methodology, A.R, A.L, C.A; formal analysis, C.A and C.O, I.C.F, V.L writing—original draft preparation; writing—review and editing, P.O., C.A..; supervision P.O. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the University of Oradea, Oradea, Romania.
Institutional Review Board Statement
This study was conducted in accordance with the Declaration of Helsinki and was approved by the Institutional Review Board (or Ethics Committee) of Resident Laboratory (nr. 11/20.12.2024)
Informed Consent Statement
The study was retrospective, used archived material, and required no further procedures.
Data Availability Statement
The data used in this study can be made available upon reasonable request.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| AKT | Protein kinase B |
| APC | Adenomatous polyposis coli |
| BRAF | B-Raf proto-oncogene, serine/threonine kinase |
| CI CK1 CRC Ct DNA EED ERK EZH1/2 EZH2 FFPE FIGO GSK-3 H3K27me3 H-score IHC KRAS LEF LRP5/6 MEK MMR MSS MSI MSI-H MSI-L mTOR NRAS OB OR PCP PCR PD-L1 PI3K PRC2 RBAP46/48 RAS SPSS SUZ12 TCF TNM Wnt |
Confidence interval Casein kinase 1 Colorectal cancer Cycle threshold Deoxyribonucleic acid Embryonic ectoderm development Extracellular signal-regulated kinase Enhancer of zeste homolog 1/2 Enhancer of zeste homolog 2 Formalin-fixed, paraffin-embedded International Federation of Gynecology and Obstetrics Glycogen synthase kinase 3 Trimethylation of lysine 27 on histone H3 Histological score Immunohistochemical Kirsten rat sarcoma viral oncogene homolog Lymphoid enhancer-binding factor Low-density lipoprotein receptor–related protein 5/6 Mitogen-activated protein kinase kinase Mismatch repair Microsatellite stable Microsatellite instability Microsatellite instability–high Microsatellite instability–low Mechanistic target of rapamycin Neuroblastoma RAS viral oncogene homolog Objective (microscope objective) Odds ratio Planar cell polarity Polymerase chain reaction Programmed cell death protein 1 Phosphoinositide 3-kinase Polycomb repressive complex 2 Retinoblastoma-associated proteins 46/48 Rat sarcoma virus oncogene family Statistical Package for the Social Sciences Suppressor of zeste 12 T-cell factor Tumor–node–metastasis staging system Wingless-related integration site |
References
- World Health Organization. Available online: https://www.who.int/news-room/fact-sheets/detail/colorectal-cancer (accessed on 13 Jan 2025).
- GLOBOCAN. Available online: https://gco.iarc.who.int/media/globocan/factsheets/cancers/41-colorectum-fact-sheet.pdf (accessed on 13 Jan 2025).
- Cancer-plan.ro. Available online: https://cancer-plan.ro/copac-cancerul-colorectal-peste-13-din-totalul-cancerelor-in-romania/ (accessed on 13 Jan 2024).
- Cecchini, M.; Sokol, E.; Vasan, N.; Pavlick, D.C.; Huang, R.S.P.; Pelletier, M.; Levy, M.A.; Pusztai, L.; Lacy, J.; Eder, J.P.; et al. Molecular characteristics of advanced colorectal cancer and multi-hit PIK3CA mutations. J. Clin. Oncol. 2022, 40, 3535–3535. [Google Scholar] [CrossRef]
- Afrăsânie, V.A.; Marinca, M.V.; Gafton, B.; Alexa-Stratulat, T.; Rusu, A.; Froicu, E.M.; Sur, D.; Lungulescu, C.V.; Popovici, L.; Lefter, A.V.; et al. Clinical, Pathological and Molecular Insights on KRAS, NRAS, BRAF, PIK3CA and TP53 Mutations in Metastatic Colorectal Cancer Patients from Northeastern Romania. Int. J. Mol. Sci. 2023, 24, 1–16. [Google Scholar] [CrossRef]
- Jin, J.; Shi, Y.; Zhang, S.; Yang, S. PIK3CA mutation and clinicopathological features of colorectal cancer: a systematic review and Meta-Analysis. Acta Oncol. (Madr) 2020, 59, 66–74. [Google Scholar] [CrossRef]
- Tan, E.S.; Fan, W.; Knepper, T.C.; Schell, M.J.; Sahin, I.H.; Fleming, J.B.; Xie, H. Prognostic and Predictive Value of PIK3CA Mutations in Metastatic Colorectal Cancer. Target. Oncol. 2022, 17, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Abrudan, R.; Abrudan, L.; Camarasan, A.; Camarasan, O.; Burta, O.; Pop, O. KRAS and NRAS Mutations in Relation to Microsatellite Status in Colorectal Cancer: A Single-Center Study From Romania Case selection. 2025, 17. [Google Scholar] [CrossRef] [PubMed]
- Bellio, H.; Fumet, J.D.; Ghiringhelli, F. Targeting BRAF and RAS in colorectal cancer. Cancers (Basel) 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Lian, S.Y.; Tan, L.X.; Liu, X.Z.; Yang, L.J.; Li, N.N.; Feng, Q.; Wang, P.; Wang, Y.; Qiao, D.B.; Zhou, L.X.; et al. KRAS, NRAS, BRAF signatures, and MMR status in colorectal cancer patients in North China. Med. (United States) 2023, 102, E33115. [Google Scholar] [CrossRef]
- Chowdhury, S.; Ferdous Ara, S.J.; Mili, S.M.; Momotaz, T.; Ahmed Molla, M.M.; Anwar, S.; Saleh, A.A. Mutational profile of KRAS, NRAS, BRAF, PIK3CA, and AKT1 genes in colorectal cancer patients in a tertiary care hospital, Dhaka. Adv. Cancer Biol. - Metastasis 2022, 5, 100054. [Google Scholar] [CrossRef]
- Zhu, Y.; Li, X. Advances of Wnt Signalling Pathway in Colorectal Cancer. Cells 2023, 12, 1–22. [Google Scholar] [CrossRef]
- Hayat, R.; Manzoor, M.; Hussain, A. Cell B iology Wnt signaling pathway : A comprehensive review. 2022, 863–877. [Google Scholar] [CrossRef]
- Liu, J.; Xiao, Q.; Xiao, J.; Niu, C.; Li, Y.; Zhang, X.; Zhou, Z.; Shu, G.; Yin, G. Wnt/β-catenin signalling: function, biological mechanisms, and therapeutic opportunities. Signal Transduct. Target. Ther. 2022, 7. [Google Scholar] [CrossRef]
- Wong, L.H.; Tremethick, D.J. Multifunctional histone variants in genome function. Nat. Rev. Genet. 2025, 26, 82–104. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Chen, X.; Jiang, Y.; Liu, S.; Liu, H.; Sun, X.; Zhang, H.; Liu, Z.; Tao, Y.; Li, C.; et al. Elevating H3K27me3 level sensitizes colorectal cancer to oxaliplatin. 2020, 12, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Wen, B.; Liang, Y.; Yu, W.; Li, H. Histone Modifications and their Role in Colorectal Cancer ( Review ). 2023, 2023–2033. [Google Scholar] [CrossRef]
- Carvalho, S.; Freitas, M.; Antunes, L.; Monteiro, S.; Marcia, R.; Coimbra, V.; Tavares, A. Prognostic value of histone marks H3K27me3 and H3K9me3 and modifying enzymes EZH2, SETDB1 and LSD-1 in colorectal cancer. J. Cancer Res. Clin. Oncol. 2018, 0, 0. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Zhou, W.; Lin, C. Review Article The Heightened Importance of EZH2 in Cancer Immunotherapy. 2023, 2, 117–129. [Google Scholar] [CrossRef]
- Tissue, P.A. Cancer Diagnostics. Bio SB Tri-Methyl-Histone. Available online: https://www.cancerdiagnostics.com/core/media/media.nl?_xt=.pdf&c=3378782&h=kcqBOaoVCZ2PVQco0GkUpxmN-2o_XTcAMNpFoACL7xLHxIHw&id=8203836&utm (accessed on 12 Dec 2025).
- Dako. Monoclonal Mouse Anti-Human Beta-Catenin IR702. Available online: https://www.agilent.com/cs/library/packageinsert/public/119464001.PDF (accessed on 12 Dec 2025).
- Dako. General Instructions For Immunohistochemical Staining. Available online: https://www.agilent.com/cs/library/packageinsert/public/108007002.PDF (accessed on 12 Dec 2025).
- Sung, R.; Kang, L.; Han, J.; Choi, J.; Lee, S.H.; Lee, T.H.; Park, S.; Ja, H. Differential Expression of E-Cadherin, β-Catenin, and S100A4 in Intestinal Type and Nonintestinal Type Ampulla of Vater Cancers. 2014, 8, 94–101. [Google Scholar] [CrossRef]
- Mordhorst, L.B.; Ahlin, C.; Sorbe, B. Prognostic impact of the expression of Wnt-signaling proteins in cervical carcinoma FIGO stage I-IV treated with radiotherapy or chemoradiotherapy. 2016, 7. [Google Scholar] [CrossRef]
- Iida, T.; Mizuno, Y.; Kaizaki, Y. Real-time PCR-based method for the rapid detection of extended RAS mutations using bridged nucleic acids in colorectal cancer. Clin. Chim. Acta 2019, 489, 164–168. [Google Scholar] [CrossRef]
- Hause, R.J.; Pritchard, C.C.; Shendure, J.; Salipante, S.J. Classification and characterization of microsatellite instability across 18 cancer types. Nat. Med. 2016, 22, 1342–1350. [Google Scholar] [CrossRef]
- Luchini, C.; Bibeau, F.; Ligtenberg, M.J.L.; Singh, N.; Nottegar, A.; Bosse, T.; Miller, R.; Riaz, N.; Douillard, J.Y.; Andre, F.; et al. ESMO recommendations on microsatellite instability testing for immunotherapy in cancer, and its relationship with PD-1/PD-L1 expression and tumour mutational burden: A systematic review-based approach. Ann. Oncol. 2019, 30, 1232–1243. [Google Scholar] [CrossRef]
- Tamagawa, H.; Oshima, T.; Numata, M.; Yamamoto, N.; Shiozawa, M.; Morinaga, S. Global histone modification of H3K27 correlates with the outcomes in patients with metachronous liver metastasis of colorectal cancer. Eur. J. Surg. Oncol. 2013, 39, 655–661. [Google Scholar] [CrossRef]
- Gao, Z.; Lu, C.; Wang, M. Differential β -catenin expression levels are associated with morphological features and prognosis of colorectal cancer. 2014, 2069–2076. [Google Scholar] [CrossRef]
- Hussein, Z.H.; Hassawi, B. Al; Ibraheem, Q. Aberrant β -Catenin Expression and Its Association With Epithelial-Mesenchymal Transition and Clinical Outcomes of Colorectal Cancer. 2024, 16, 1–18. [CrossRef] [PubMed]
- Carrasco, M.E.; Thaler, R.; Nardocci, G.; Dudakovic, A.; Wijnen, A.J. Van Inhibition of Ezh2 redistributes bivalent domains within transcriptional regulators associated with WNT and Hedgehog pathways in osteoblasts. J. Biol. Chem. 2023, 299, 105155. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, H.; Truc, T.; Jane, W.; Peter, C.; Heather, M.; Hagan, O. iScience ll Activation of AKT induces EZH2-mediated b -catenin trimethylation in colorectal cancer. ISCIENCE 2023, 26, 107630. [Google Scholar] [CrossRef]
- Dong, G.; Xu, J.-L. Critical Roles of Polycomb Repressive Complexes in Transcription and Cancer. Int. J. Mol. Sci. 2022, 23. [Google Scholar] [CrossRef]
- Nakazawa, T.; Kondo, T.; Ma, D. Global histone modification of histone H3 in colorectal cancer and its precursor lesions ☆, ☆☆. Hum. Pathol. 2012, 43, 834–842. [Google Scholar] [CrossRef]
- Schuijers, J.; Mokry, M.; Hatzis, P.; Cuppen, E.; Clevers, H. Wnt-induced transcriptional activation is exclusively mediated by TCF/LEF. 2014, 33. [Google Scholar] [CrossRef]
- Verma, S.; Goyal, N.; Goyal, S.; Kaur, P.; Gupta, S. EZH2 Dysregulation and Its Oncogenic Role in Human Cancers. Cancers (Basel) 2025, 1–43. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



