
Submitted:
09 February 2026
Posted:
10 February 2026
You are already at the latest version
Abstract
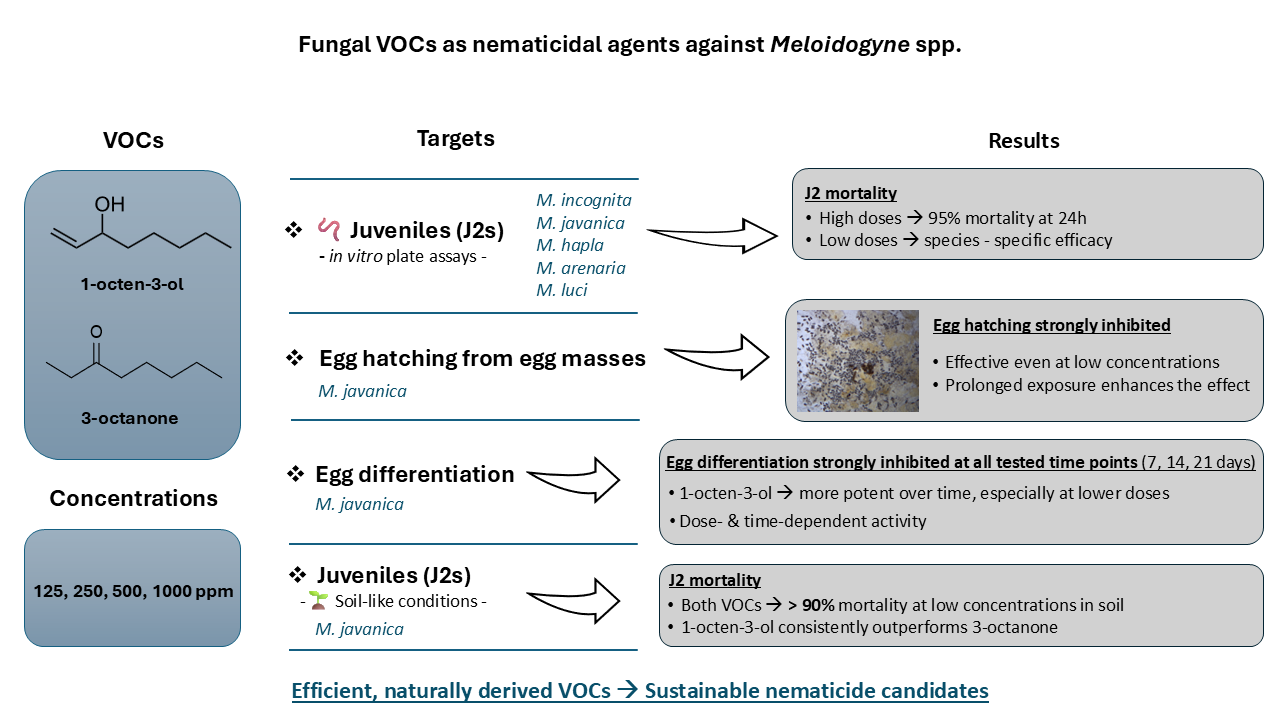
Root-knot nematodes (RKNs) of the genus Meloidogyne are major plant pests causing severe crop losses. Microbial volatile organic compounds (VOCs) have emerged as promising biopesticides. In this work, we evaluated the nematicidal activity of two fungal VOCs, 1-octen-3-ol and 3-octanone, against five Meloidogyne species (M. incognita, M. javanica, M. hapla, M. arenaria, and M. luci). Plate assays showed near-complete (95–100%) juvenile mortality at 500–1000 ppm within 24 h. At lower concentrations, mobility declined and species-specific effects appeared: 1-octen-3-ol was more effective against M. arenaria, while 3-octanone strongly affected M. hapla and moderately M. incognita and M. javanica. Egg differentiation was significantly inhibited across time points, with up to 80% reduction at 1000 ppm and effects persisting at 125 ppm. Egg hatching from egg masses was reduced up to 95% in a concentration-dependent manner, irrespective of compound type. Soil-like assays confirmed enhanced efficacy at low doses, with >90% mortality at 125 ppm after 24 h, suggesting prolonged activity. These results show that fungal VOCs are potent nematicides at low doses and short exposures, highlighting 1-octen-3-ol and 3-octanone as eco-friendly candidates for sustainable nematode control.
Keywords:
biopesticide
; Meloidogyne
; nematicide
; Volatile Organic Compounds (VOC)
1. Introduction
Plant-parasitic nematodes (PPNs) cause substantial damage to a wide range of crops, with global yield losses estimated at $157 billion annually [1,2,3]. Among them, root-knot nematodes (RKN) like Meloidogyne spp., rank among the top ten most destructive plant-parasitic nematodes [4], infecting over 3,000 plant species, including vegetables (e.g., tomato, potato), legumes (e.g., soybean), cereals (e.g., rice, maize), and fruits (e.g., banana, melon) [5,6]. Second-stage juveniles (J2s) penetrate plant roots, inducing gall formation and disrupting root function [7], with symptoms and severity varying by species [8,9,10]. Among the 98 known Meloidogyne species, M. arenaria, M. incognita, M. javanica, and M. hapla are the most widespread and devastating for agriculture [1,11]. M. luci is also considered an extremely damaging pest with a wide host range and was included in the European and Mediterranean Plant Protection Organization Alert List of harmful organisms [12,13,14].
Managing RKNs remains a significant agricultural challenge. Currently, various preventive strategies, including cover crops, crop rotations, soil solarization, and resistant cultivars, have been widely implemented to reduce nematode infestations [15,16]. Despite these efforts, synthetic nematicides remain a primary tool for managing RKN in heavily affected fields. However, the implementation of regulatory restrictions, such as Directive 91/414/EEC and Regulation 1107/2009/EC of the European Union, has led to the withdrawal of many chemical nematicides due to environmental and human health concerns [17,18,19]. In addition to regulatory restrictions, the long-term efficacy of chemical nematicides is declining due to the development of nematode resistance, further limiting control options [20,21]. Microbial biopesticides have been used as nematicides [22,23,24], but recent research efforts are increasingly focused on natural product-based nematicides as environmentally friendly alternatives [25,26,27,28].
Plant- and microorganism-derived volatile organic compounds (VOCs) have emerged as promising biopesticides [29]. These compounds are mainly fumigants and have been used in many biological control practices, exhibiting nematicidal, insecticidal, and antimicrobial properties [29,30]. Certain VOCs disrupt nematode behavior and induce plant defenses, as well as acting as fumigants, offering a viable alternative to conventional nematicides [28,31,32,33,34].
Among these, the VOCs 1-octen-3-ol and 3-octanone have shown a high effectiveness as biopesticides and biostimulants. These compounds, produced by both plants and fungi, such as the entomopathogenic fungus Metarhizium brunneum, have been shown to promote plant growth [35], activate plant defences [37], attract and kill molluscs [38], and exhibit fumigant toxicity against major soil pests such as Agriotes lineatus (wireworm), Diabrotica virgifera (corn rootworm), and Phyllopertha horticola (garden chafer), with responses varying greatly in a dose-dependent manner [35,39]. Studies suggest that high concentrations of these VOCs influence the behavior and survival of entomopathogenic nematodes (EPNs), potentially reducing their efficacy in biological control [40]. Furthermore, M. hapla has been shown to alter its behavior in response to these compounds, exhibiting attraction at low doses and repellency at higher doses [41].
Τhe potent nature and broad effectiveness of these VOCs make them promising candidates for new bionematicides. However, comprehensive studies on the efficacy of 1-octen-3-ol and 3-octanone against multiple Meloidogyne species are lacking. This study investigates their nematicidal activity across five major Meloidogyne species, focusing on their impact on survival, behavior, and potential as biopesticides.
2. Results
2.1. Nematode Mortality Bioassays
Statistical analysis revealed that compound type, nematode species, and VOC concentration each had a significant effect on lethality (p < 0.001). All two-way interactions—compound × species, compound × concentration, and species × concentration—were also significant (p < 0.001), as was the three-way interaction between these factors (p < 0.001). Among the main effects, concentration explained the largest portion of variance (F = 18,277.21), followed by species (F = 71.27) and compound (F = 46.77) (Table 1).
Both 1-octen-3-ol and 3-octanone significantly reduced nematode survival in plate assays at higher concentrations (500 and 1000 ppm), resulting in near-complete mortality within 24 hours across all five Meloidogyne species (Figure 1a, b). Even at lower concentrations, although mortality was reduced, nematodes exposed to either compound appeared visibly less active compared to untreated controls. Clear species-specific differences in sensitivity and compound performance were observed (Figure 1a, b, Table 1).
In M. arenaria, 1-octen-3-ol was significantly more effective than 3-octanone at 250 ppm, producing an average mortality of 37% versus 16% for 3-octanone (p < 0.001). At 125 ppm, 1-octen-3-ol still outperformed 3-octanone by approximately 8%, while 3-octanone did not differ significantly from the control (Figure 1). In contrast, M. hapla responded much more strongly to 3-octanone. At 250 ppm, 3-octanone induced 85% mortality, while 1-octen-3-ol had almost no effect (2%)—a highly significant difference (p < 0.001). At 125 ppm, neither compound caused notable mortality, but impaired nematode movement was evident within 24 hours. Similar but less pronounced patterns were observed in M. javanica and M. incognita. For M. javanica, 3-octanone at 250 ppm caused 25% average mortality—significantly higher than 1-octen-3-ol (14%, p < 0.001). At 125 ppm, only 1-octen-3-ol showed minor lethality (~5%). In M. incognita, 3-octanone led to 20% mortality at 250 ppm, roughly twice that of 1-octen-3-ol (11%, p < 0.001). However, responses to 3-octanone were variable across replicates (e.g., 21% vs. 4.87%), while 1-octen-3-ol maintained more consistent but lower effects. Finally, in M. luci, both compounds caused low mortality at 250 ppm, with no significant difference between treatments or compared to control (p = 0.811). At 125 ppm, neither compound induced significant lethality, although treated nematodes again displayed reduced activity (Figure 1).
2.2. Effect on Egg Development
The inhibitory effects of the two VOCs across different concentrations and time points are shown in Figure 2a-c.
Although the compound type did not have a significant overall impact on egg differentiation, both the duration of exposure and VOC concentration showed strong effects (p < 0.001). Moreover, significant interactions between exposure time (days) and concentration, as well as between compound and concentration, suggest that each compound’s effectiveness varies depending on both dose and time (Table 2).
By Day 7, exposure to the highest tested concentration (1,000 ppm) of either compound resulted in approximately 78% fewer differentiated eggs compared to the control. Specifically, diferrentiation dropped from 30.5% to 6.5% for 1-octen-3-ol and from 32.0% to 7.3% for 3-octanone. By Day 14, both compounds displayed a clear dose-dependent inhibitory effect. For 1-octen-3-ol, significant suppression began at concentrations of 250 ppm and above, culminating in a ~72% reduction at 1000 ppm. A similar pattern was observed with 3-octanone, though the overall effect was slightly less intense.
By Day 21, inhibition of differentiation remained strong for both VOCs, but 1-octen-3-ol showed higher efficacy at lower doses. A significant reduction (~26%) was already evident at just 125 ppm, highlighting its greater potency over time.
2.3. Effect on Egg Hatching from Egg Masses
The percentage of egg hatching from egg masses was significantly affected by the concentration (p < 0.001), while neither the compound itself (p = 0.503) nor the interaction between compound and concentration (p = 0.603) had a significant effect. Both compounds significantly reduced the percentage of hatched eggs from egg masses compared to the control, starting with reductions of 84% for 1-octen-3-ol and 75% for 3-octanone at the lowest tested dose (125 ppm), and reaching near-complete inhibition (~95% and 91%, respectively) at higher concentrations (500 and 1000 ppm) (Figure 3). 1-octen-3-ol consistently caused strong suppression across all tested concentrations (125-1000 ppm), with no statistically significant differences among doses (125 to 1000ppm). In contrast, 3-octanone exhibited a clear dose-dependent effect: lower concentrations were statistically less effective than higher concentrations, and higher concentrations were required to achieve comparable suppression levels to the respective ones of 1-octen-3-ol (500-1000ppm).
2.4. Soil Survival Assays
Treated nematodes were collected 24 h after exposure to each VOC in soil and were counted after 48 h. Both compounds achieved 100% mortality at the highest concentrations tested (1,000 and 500 ppm) (Figure 4). At the lower concentrations of 125 and 250 ppm, mortality remained comparably high, indicating that even the lowest concentrations were highly effective and that further increases in dose did not provide substantial additional benefits. Specifically, nematode recovery numbers for 3-octanone averaged 32 (93.6% reduction) and 36 (92.8% reduction), compared with ~500 in the control group (Figure 4). At both 125 and 250 ppm, 1-octen-3-ol treatments resulted in even lower recovery counts (18.6 and 23.4 on average, respectively) than 3-octanone (Figure 4). The nematodes that were not retrieved were either dead or unable to move through the sand, being immobilized by the VOC activity.
3. Discussion
This study evaluated the effects of two volatile compounds, 1-octen-3-ol and 3-octanone, on multiple life stages of Meloidogyne spp. using various experimental setups, demonstrating their potential as effective eco-friendly nematicidal agents. While these VOCs have already been successfully applied against various invertebrates, including mosquitoes [42,43] and wireworms [36], as well as pathogenic microbes [44], their nematicidal properties have only recently been described [41,45].
Our findings expand this body of work by demonstrating their efficacy across five species and multiple developmental stages. Highest concentrations of 500 and 1,000 ppm of both VOCs caused over 95% of J2 mortality of all five species, within 24 hours. Lower concentrations of 250 ppm and 125 ppm were also effective, primarily impairing nematode mobility, though mortality rates differed among species (Figure 1, Table 1). These results emphasize that overall potency is high and that compound-specific nematicidal activity is distinctly species-dependent. These findings align with previous research demonstrating species-dependent nematicidal effects, highlighting the importance of understanding interspecies variability when developing new formulations and their application [41,45,46,47]. Jang et al. [48] found that alkaloids from Waltheria indica exhibited differential effects on M. arenaria and M. incognita, leading to species-specific survival rates. Similarly, fluensulfone showed distinct effects on M. arenaria juveniles compared to M. incognita, eliciting different behavioral responses [49]. The species-specific variations observed in our study further support the notion that nematode susceptibility is influenced by genetic, morphological, and physiological factors, reinforcing the need for species-adapted biocontrol solutions [5,50,51,52]. Notably, M. hapla differs from other species in its ability to undergo both meiotic and mitotic parthenogenesis, which may enhance its adaptability to environmental pressures and contribute to its distinct susceptibility to 3-octanone. Further investigation is also needed to clarify why M. arenaria is particularly susceptible to 1-octen-3-ol compared to other species.
Egg hatching from egg masses was significantly inhibited by both VOCs in M. javanica, since percentages dropped sharply (over 90%) even at the lowest tested concentration (125 ppm). Several fungal and bacterial VOCs have been shown to negatively affect egg hatching on nematodes [53,54,55]. Ovicidal activity is considered a desirable trait for a successful commercial nematicide, since eggs are the main survival mode of Meloidogyne species in the soil [56]. Our study showed that the decrease in egg hatching as well as egg differentiation was generally highly concentration- and time-dependent, with effects lasting throughout the exposure period. While 1-octen-3-ol causes a higher reduction (84%) of egg hatching at 250 ppm compared to 3-octanone (75%), the differences were not significant and became marginal at the highest concentration (95 vs 91%, respectively). This contradicts previous studies that showed a substantial superior effect of 1-octen-3-ol on egg hatching in M. incognita, at the lowest concentration tested [45]. These findings may indicate that prolonged exposure or higher doses of applied nematicides lead to converging results, due to the accumulated toxicity, potentially masking compound-specific differential activities, an effect also observed by mortality assays of J2s across the five species. This observation suggests that careful selection of test conditions is essential when evaluating compound-specific or species-specific responses by nematicidal activity.
In soil-based assays, both VOCs achieved over 90% J2 mortality at 125 and 250 ppm within 24 hours (Figure 2). Compared to in vitro plate assays, soil application resulted in prolonged retention and enhanced effectiveness, likely due to the sand medium acting as a carrier for slow release. In contrast, in liquid-based systems, rapid evaporation and diffusion may have reduced prolonged exposure, leading to lower efficacy. While it remains unclear whether the observed reduction in extracted nematodes was due to actual mortality or impaired mobility preventing their migration during extraction, both outcomes are biologically significant. Reduced mobility would limit nematode root infection, while prolonged exposure to soil could eventually lead to higher mortality. These findings emphasize the potential of VOC-based treatments for effective nematode suppression and highlight the need for further soil-based investigations to assess long-term efficacy. Moreover, the higher effectiveness of 1-octen-3-ol against M. javanica in soil-based assays contradicts our initial findings in the in vitro plate assays, where 3-octanone caused higher mortality. Similarly, while our results for M. hapla align with previous observations by Khoja et al. [38], Veronico et al. [45] reported greater effectiveness of 1-octen-3-ol against M. incognita. These discrepancies in compound effectiveness across different environments suggest that their impact on nematodes is highly application-dependent, a trend also observed in other studies [41,58].
However, the results of this study indicate that these compounds are highly effective nematicides compared with known microbial VOCs [29]. Bacterial VOCs, (e.g. from Pseudomonas and Bacillus spp.), have been studied for their nematicidal properties against M. incognita and M. javanica [59,60,61,62]. Despite being less studied, recent research has identified fungal VOCs with nematicidal activity. For instance, Mei et al. [55] found that cyclohexanol, cyclohexanone, and cyclohexanamine, produced by Duddingtonia flagrans, were effective against M. incognita at specific concentrations. Similarly, Fusarium oxysporum was reported to emit 2-methylbutyl acetate, 3-methylbutyl acetate, ethyl acetate, and 2-methylpropyl acetate, which were toxic to M. incognita, causing 88 to 96% J2 mortality [56], as well as 70% and 65% reduction of infectivity and reproduction [53,62]. Moreover, VOCs from Daldinia spp. reduced viability of M. javanica J2s by 67%, while the synthetic mixture of VOCs showed a 99% reduction of J2 viability and 87% inhibition of egg hatching [61]. Our observed mortality rates exceeding 95% across five Meloidogyne species at concentrations of 500-1000 ppm, along with over 90% reductions in J2 survival under soil-like conditions, and over 90% inhibition of egg differentiation and egg hatching from egg masses at lower concentrations (125-250 ppm), highlight 1-octen-3-ol and 3-octanone as highly promising candidates for the development of multi-purpose nematicides targeting multiple life stages of the Meloidogyne life cycle. Their effectiveness is further complemented by their widespread occurrence in nature, low cost, and transient environmental persistence. Notably, both compounds are already approved for use in the food industry as flavoring agents (e.g., FEMA GRAS status, 21 CFR 172.515), which may facilitate their regulatory approval and integration into crop protection strategies.
The mechanism of action of these VOCs has been recently explored by Veronico et al. [45], who showed that exposure of M. incognita J2 to 1-octen-3-ol and 3-octanone as fumigants in a closed environment resulted in elevated levels of ROS, causing oxidative stress, inducing damage to internal structures, and leading to death. This mechanism is similar to the mode of action of other nematicides [45]. If this mechanism is consistent across species, the species-specific responses observed in this study could be attributed to differences in oxidative stress tolerance or detoxification mechanisms. Supporting the idea of conserved toxicity, studies in Drosophila have shown that 1-octen-3-ol can also cause neurological damage by disrupting dopamine homeostasis and interacting with genetic variants in dopamine biosynthesis to enhance dopaminergic neurodegeneration [63]. In addition to their nematicidal properties, 1-octen-3-ol and 3-octanone are already known to promote plant growth in various angiosperms, many of which are also susceptible to nematode infestations [35]. By simultaneously reducing nematode populations and enhancing plant vigor, these compounds could contribute to an integrated approach to crop protection, improving overall plant health and resilience in agricultural systems.
Overall, this study provides compelling evidence that 1-octen-3-ol and 3-octanone are highly effective nematicidal VOCs with species-specific effects on major Meloidogyne species. Their efficacy, in both in vitro and in soil-based assays, highlights their potential as sustainable alternatives to conventional nematicides. The observed variations in species susceptibility highlight the importance of further investigating the physiological and genetic factors underlying nematode responses to VOCs. Careful selection of test concentrations and exposure durations is essential when evaluating nematicidal activity, as high doses or prolonged exposure may obscure compound- or species-specific differences due to cumulative toxicity. Additionally, the dual functionality of these compounds in promoting plant growth, while suppressing nematode populations and nematode mobility, suggests their applicability in integrated pest management strategies. Future research should focus on optimizing formulation strategies, assessing long-term environmental impacts, and exploring the potential synergistic effects of VOCs combinations to enhance their efficacy in field applications [19]. By advancing our understanding of fungal VOCs as nematicidal agents, this study contributes to the development of innovative and eco-friendly solutions for nematode management in agriculture.
4. Materials and Methods
4.1. Nematode Cultures
Five species of Meloidogyne spp. were used in these experiments, i.e. M. incognita, M. luci, M. arenaria, M. javanica and M. hapla. All Meloidogyne species used in the present study were originally maintained in a greenhouse at 25-30°C, and 16 h photoperiod at Crete, Greece for many years. For this work, these populations were multiplied on tomato plants (Lycopersicon esculentum Mill. cv. Belladona) at the five-leaf stage, grown in 18cm diameter 1 L plastic pots containing a 50:50 mixture of peat and perlite. The plants were kept in a growth chamber at 25±2°C, with 65% RH and 16 h photoperiod at the Agricultural University of Athens, Greece. After 40 days, the plants were removed from the pots, and the roots were carefully washed to remove soil residues. Egg extraction was performed using the hypochlorite method [64], with a 1% sodium hypochlorite solution, and second-stage juveniles (J2s) were obtained by placing eggs on a Baermann funnel at ambient temperature (27 ± 1 °C). J2s collected during the first three days were discarded, while newly hatched juveniles were collected every two days and used in the experiments.
4.2. Nematode Mortality Bioassays
In vitro assays were conducted to assess the nematicidal effect of the two VOCs on the five Meloidogyne species. The experiments were conducted using 24-well polystyrene plates (Nunc, Germany). The VOCs, 1-octen-3-ol (CAS number 3391-86-4, Sigma-Aldrich) and 3-octanone (CAS number 106-68-3, Sigma-Aldrich), were separately mixed with ethanol (20μl of compound to 100 μl ethanol) and brought to 1 ml with 0.3% v/v Tween-20 aqueous solution, to create the stock solution. Nematodes exposed at these concentrations of ethanol and Tween-20 were not affected, as preliminary tests and previous work indicate [28]. For each nematode species and VOC concentration, five replicate wells were used, with each well containing approximately 50 nematodes exposed to final compound concentration of 1000, 500, 250 or 125 ppm. The final compound solutions and nematode suspensions were mixed in each well at a 1:1 (v/v) ratio. Negative control treatments contained less than 0.1% ethanol and 0.3% Tween-20 aqueous solution. To minimize VOC evaporation, all plates were sealed with parafilm and covered with lids. The entire assay was repeated twice to ensure reproducibility.
Nematode viability and mobility were assessed and recorded at 24 hours. Observations were performed under an inverted microscope (Zeiss, Oberkochen, Germany) at 40x and 100x magnifications. Individuals were classified as either mobile or dead in a window of 10 seconds. The shape of the juveniles was determined to be straight (I-shaped) or bent (banana-shaped or L-shaped) which indicated the difference between paralyzed (dead) or motile (alive) nematodes, respectively. Lack of movement was considered a strong indication of paralysis.
A General Linear Model (GLM) was fitted to evaluate the main effects and interactions of compound type and concentration across different species on recorded J2 nematode mortality. Pairwise comparisons of estimated marginal means (EMMeans) were conducted to identify specific differences between groups within each interaction level, with appropriate adjustments for multiple testing.
4.3. Effect on Egg Development
Meloidogyne javanica eggs were extracted from infected tomato roots (Solanum lycopersicum cv. Belladona) using the sodium hypochlorite method described by Hussey and Barker [64]. The egg suspension was passed through 53 μm and 38 μm sieves, thoroughly rinsed with tap water, and collected in a 100 ml beaker. Egg concentration was estimated under an inverted microscope (100x), and the suspension was adjusted to 50 eggs/ml for use in the bioassay.
The effect of 1-octen-3-ol and 3-octanone on egg differentiation of M. javanica was evaluated at four concentrations (125, 250, 500 and 1000 ppm). Both compounds were prepared following previously described procedures and and mixed to achieve a 1:1 (v/v) ratio with egg suspensions in Cellstar® 24-well plates. Each well received 0.5 ml of egg suspension (approximately 50 eggs, of which 80-90% were undifferentiated-day 0), followed by 0.5 ml of the test solution. Controls consisted of either distilled water or water with ethanol and Tween-20 at concentrations equivalent to those used in treatments. All plates were sealed with lids to prevent evaporation and incubated at 26 ± 1 °C.
Egg hatching was monitored at 7, 14, and 21 days post-exposure under an inverted microscope (100×). The number of differentiated J2s was recorded at each time point. To account for natural hatching variation, differentiation rates (%) were calculated for each replicate based on the number of eggs that differentiated relative to day 0. The experiment was conducted four times with 5 replicates each.
A GLM was fitted to evaluate the effects of the applied compound, the compound’s concentration and the day post-treatment, as well as their interactions on the percentage of differentiated eggs. To further probe these higher-order interactions, we conducted EMMeans were compared to provide adjusted group means while controlling for the factors in the model. Pairwise comparisons of EMMeans were used to identify specific differences between groups within each interaction level, with appropriate adjustments for multiple testing.
4.4. Effect on Egg Hatching from Egg Masses
Mature egg masses of M. javanica were carefully collected from washed tomato roots and placed individually in small extraction trays (6 cm Petri dishes), with one egg mass per tray. Each egg mass was exposed to 10 ml of either 1-octen-3-ol or 3-octanone at one of four concentrations (125, 250, 500, and 1,000 ppm). Test solutions of 1-octen-3-ol and 3-octanone were prepared as described previously by dissolving each compound in ethanol and bringing them to volume with a Tween-20 aqueous solution.
Egg masses remained submerged in the treatment solutions for seven days at 26 ± 1 °C. After this period, solutions were discarded, and the egg masses were gently rinsed twice with clean water to remove residual VOC before being transferred to fresh trays containing 10 ml of distilled water. The trays were covered to prevent evaporation and maintained at 26 ± 1 °C. Juveniles (J2) that hatched were counted and removed at weekly intervals. After four weeks, the experiment was terminated (hatching was not observed in control samples). Each egg mass was transferred to a microscope slide, gently crushed under a coverslip and the number of unhatched eggs was recorded using an inverted microscope. Each treatment was replicated four times, and the experiment was independently repeated twice, for a total of 10 replicates per treatment.
A GLM was fitted to evaluate the effect of the two compounds and applied concentrations, as well as their interactions on percent J2 hatching from egg masses. Pairwise comparisons of EMMeans were used to identify specific differences between groups within each interaction level, with appropriate adjustments for multiple testing.
4.5. Soil Assays
The third assay evaluated the effects of 1-octen-3-ol and 3-octanone under soil conditions using sand-filled falcon tubes. An aqueous suspension containing >500 M. javanica nematodes was injected into tubes filled with 80 g of wet sand after the addition of the compound solution (equivalent to 60 g of dry sand). Compound solutions had been pre-mixed with the test compound diluted in ethanol (as previously described) and serially diluted in a 0.3% Tween-20 aqueous solution to reach final concentrations. Negative controls contained less than 0.1% ethanol and 0.3% Tween-20 aqueous solution.
After 24 hours, nematodes were extracted using the Baermann funnel technique, and their numbers were recorded under a microscope after a total of 48 hours. Concentrations of 1000, 500, 250, and 125 ppm were tested, each with five replicates. The experiment was repeated twice to ensure reproducibility.
Author Contributions
Conceptualization: A.M.K., I.O.G., T.M.B.; Data curation: A.M.K., D.N.; Investigation: A.M.K, D.N.; Funding acquisition: I.O.G., T.M.B.; Resources: D.N., I.O.G.; Supervision: I.O.G., V.N.K.; Visualization: A.M.K, D.N.; Writing – original draft: A.M.K.; Writing – review & editing: A.M.K D.N., I.O.G., V.N.K., E.J.L.., D.C.E., T.M.B.
Funding
This work was supported through Swansea University's EPSRC Impact Acceleration Account 2022 – 2026 (EP/X525637/1) and ‘by UK Research and Innovation Building a Green Future strategic theme [grant number UKRI239]’
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding author(s).
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Li, J.; Zou, C.; Xu, J.; Ji, X.; Niu, X.; Yang, J.; Zhang, K.Q. Molecular mechanisms of nematode-nematophagous microbe interactions: Basis for biological control of plant-parasitic nematodes. Annu. Rev. Phytopathol. 2015, 53, 67–95. [Google Scholar] [CrossRef]
- Kayani, M.Z.; Mukhtar, T.; Hussain, M.A. Effects of southern root-knot nematode population densities and plant age on growth and yield parameters of cucumber. Crop Prot. 2017, 92, 207–213. [Google Scholar] [CrossRef]
- Sikora, R.; Molendijk, L.P.G.; Desaeger, J. Integrated nematode management and crop health: Future challenges and opportunities. In Integrated Nematode Management: State-of-the-Art and Visions for the Future; Sikora, R., Desaeger, J., Molendijk, L.P.G., Eds.; CAB International: Wallingford, UK, 2022; pp. 3–10. [Google Scholar] [CrossRef]
- Jones, J.T.; Haegeman, A.; Danchin, E.G.; Gaur, H.S.; Helder, J.; Jones, M.G.; Kikuchi, T.; Manzanilla-López, R.; Palomares-Rius, J.E.; Wesemael, W.M.; Perry, R.N. Top 10 plant-parasitic nematodes in molecular plant pathology. Mol. Plant Pathol. 2013, 14, 946–961. [Google Scholar] [CrossRef] [PubMed]
- Trudgill, D.L.; Blok, V.C. Apomictic, polyphagous root-knot nematodes: Exceptionally successful and damaging biotrophic root pathogens. Annu. Rev. Phytopathol. 2001, 39, 53–77. [Google Scholar] [CrossRef]
- Bebber, D.P.; Holmes, T.; Gurr, S.J. The global spread of crop pests and pathogens. Glob. Ecol. Biogeogr. 2014, 23, 1398–1407. [Google Scholar] [CrossRef]
- Onkendi, E.M.; Kariuki, G.M.; Marais, M.; Moleleki, L.N. The threat of root-knot nematodes (Meloidogyne spp.) in Africa: A review. Plant Pathol. 2014, 63, 727–737. [Google Scholar] [CrossRef]
- Daramola, F.; Popoola, J.; Eni, A.; Sulaiman, O. Characterization of root-knot nematodes (Meloidogyne spp.) associated with Abelmoschus esculentus, Celosia argentea and Corchorus olitorius. Asian J. Biol. Sci. 2014, 8, 42–50. [Google Scholar] [CrossRef]
- Ralmi, N.H.A.; Khandaker, M.M.; Mat, N. Occurrence and control of root-knot nematode in crops: A review. Aust. J. Crop Sci. 2016, 10, 1649–1654. [Google Scholar] [CrossRef]
- Ntinokas, D.; Roussis, I.; Mavroeidis, A.; Stavropoulos, P.; Folina, A.; Kakabouki, I.; Tzortzakakis, E.A.; Bilalis, D.; Giannakou, I.O. Virulence of five root-knot nematodes (Meloidogyne spp.) on nine industrial hemp (Cannabis sativa L.) varieties and nematicidal potential of hemp seed extracts against Meloidogyne javanica. Plants 2025, 14, 227. [Google Scholar] [CrossRef]
- Ye, L.; Wang, J.Y.; Liu, X.F. Nematicidal activity of volatile organic compounds produced by Bacillus altitudinis AMCC 1040 against Meloidogyne incognita. Arch. Microbiol. 2022, 204, 521. [Google Scholar] [CrossRef]
- Carneiro, R.M.D.G.; Gomes, A.C.M.M.; Almeida, M.R.A.; Correa, V.R.; Karssen, G.; Castagnone-Sereno, P.; Mohammad Deimi, A. Meloidogyne luci n. sp. (Nematoda: Meloidogynidae), a root-knot nematode parasitising different crops in Brazil, Chile and Iran. Nematology 2014, 16, 289–301. [Google Scholar] [CrossRef]
- Gerič Stare, B.; Strajnar, P.; Susič, N.; Urek, G.; Širca, S. Reported populations of Meloidogyne ethiopica in Europe identified as Meloidogyne luci. Plant Dis. 2017, 101, 1627–1632. [Google Scholar] [CrossRef] [PubMed]
- Bačić, J.; Pavlović, M.; Kušić-Tišma, J.; Širca, S.; Theuerschuh, M.; Gerič Stare, B. First report of the root-knot nematode Meloidogyne luci on tomato in Serbia. Plant Dis. 2023, 107, 8. [Google Scholar] [CrossRef]
- Francis, C.A.; Clegg, M.D. Crop rotations in sustainable production systems. In Sustainable Agricultural Systems; CRC Press: Boca Raton, FL, USA, 2020; pp. 107–122. [Google Scholar] [CrossRef]
- Sasanelli, N.; Konrat, A.; Migunova, V.; Toderas, I.; Iurcu-Straistaru, E.; Rusu, S.; Bivol, A.; Andoni, C.; Veronico, P. Review on control methods against plant parasitic nematodes applied in Southern Member States (C Zone) of the European Union. Agriculture 2021, 11, 602. [Google Scholar] [CrossRef]
- Martin, F.N. Development of alternative strategies for management of soilborne pathogens currently controlled with methyl bromide. Annu. Rev. Phytopathol. 2003, 41, 325–350. [Google Scholar] [CrossRef]
- Villaverde, J.J.; Sevilla-Morán, B.; Sandín-España, P.; López-Goti, C.; Alonso-Prados, J.L. Biopesticides in the framework of the European pesticide regulation (EC) No. 1107/2009. Pest Manag. Sci. 2014, 70, 2–5. [Google Scholar] [CrossRef]
- Couceiro, J.C.; Wood, M.J.; Papadopoulos, A.; Silva, J.J.; Vontas, J.; Dimopoulos, G. Development of mycoinsecticides: Advances in formulation, regulatory challenges and market trends for entomopathogenic fungi. J. Fungi 2026, 12, 7. [Google Scholar] [CrossRef]
- Meher, H.C.; Gajbhiye, V.T.; Chawla, G.; Singh, G. Virulence development and genetic polymorphism in Meloidogyne incognita after prolonged exposure to sublethal concentrations of nematicides and continuous growing of resistant tomato cultivars. Pest Manag. Sci. 2009, 65, 1201–1207. [Google Scholar] [CrossRef]
- Chen, J.; Li, Q.X.; Song, B. Chemical nematicides: Recent research progress and outlook. J. Agric. Food Chem. 2020, 68, 12175–12188. [Google Scholar] [CrossRef]
- Sharon, E.; Bar-Eyal, M.; Chet, I.; Herrera-Estrella, A.; Kleifeld, O.; Spiegel, Y. Biological control of the root-knot nematode Meloidogyne javanica by Trichoderma harzianum. Phytopathology 2001, 91, 687–693. [Google Scholar] [CrossRef]
- Siddiqui, I.A.; Shaukat, S.S.; Khan, A. Differential impact of some Aspergillus species on Meloidogyne javanica biocontrol by Pseudomonas fluorescens strain CHA0. Lett. Appl. Microbiol. 2004, 39, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Xie, G.L.; Soad, A.; Coosemans, J. Suppression of Meloidogyne javanica by antagonistic and plant growth-promoting rhizobacteria. J. Zhejiang Univ. Sci. B 2005, 6, 496–501. [Google Scholar] [CrossRef]
- Caboni, P.; Saba, M.; Tocco, G.; Casu, L.; Murgia, A.; Maxia, A.; Menkissoglu-Spiroudi, U.; Ntalli, N. Nematicidal activity of mint aqueous extracts against the root-knot nematode Meloidogyne incognita. J. Agric. Food Chem. 2013, 61, 9784–9788. [Google Scholar] [CrossRef]
- Eloh, K.; Kpegba, K.; Sasanelli, N.; Koumaglo, H.K.; Caboni, P. Nematicidal activity of some essential plant oils from tropical West Africa. Int. J. Pest Manag. 2019, 66, 131–141. [Google Scholar] [CrossRef]
- Leoni, C.; Piancone, E.; Sasanelli, N.; Bruno, G.L.; Manzari, C.; Pesole, G.; Ceci, L.R.; Volpicella, M. Plant health and rhizosphere microbiome: Effects of the bionematicide Aphanocladium album in tomato plants infested by Meloidogyne javanica. Microorganisms 2020, 8, 1922. [Google Scholar] [CrossRef]
- Nasiou, E.; Giannakou, I.O. Nematicidal potential of thymol against Meloidogyne javanica (Treub) Chitwood. Plants 2023, 12, 1851. [Google Scholar] [CrossRef]
- Bui, H.X.; Desaeger, J.A. Volatile compounds as potential bio-fumigants against plant-parasitic nematodes—A mini review. J. Nematol. 2021, 53, e2021-14. [Google Scholar] [CrossRef]
- De Clerck, C.; Genva, M.; Jijakli, M.H.; Fauconnier, M.L. Use of essential oils and volatile compounds as biological control agents. Foods 2021, 10, 1062. [Google Scholar] [CrossRef] [PubMed]
- Thomas, G.; Withall, D.; Birkett, M. Harnessing microbial volatiles to replace pesticides and fertilizers. Microb. Biotechnol. 2020, 13, 1366–1376. [Google Scholar] [CrossRef] [PubMed]
- Kang, B.R.; Anderson, A.J.; Kim, Y.C. Hydrogen cyanide produced by Pseudomonas chlororaphis O6 exhibits nematicidal activity against Meloidogyne hapla. Plant Pathol. J. 2018, 34, 35–43. [Google Scholar] [CrossRef]
- Yan, D.; Cao, A.; Wang, Q.; Li, Y.; Guo, M.; Guo, X. Dimethyl disulfide (DMDS) as an effective soil fumigant against nematodes in China. PLoS ONE 2019, 14, e0224456. [Google Scholar] [CrossRef]
- Sarri, K.; Mourouzidou, S.; Ntalli, N.; Monokrousos, N. Recent advances and developments in the nematicidal activity of essential oils and their components against root-knot nematodes. Agronomy 2024, 14, 213. [Google Scholar] [CrossRef]
- Wood, M.J.; Kortsinoglou, A.M.; Khoja, S.; Kouvelis, V.N.; Myrta, A.; Midthassel, A.; Loveridge, E.J.; Butt, T.M. Metarhizium brunneum (Hypocreales: Clavicipitaceae) and its derived volatile organic compounds as biostimulants of commercially valuable angiosperms and gymnosperms. J. Fungi 2022, 8, 1052. [Google Scholar] [CrossRef] [PubMed]
- Wood, M.J.; Kortsinoglou, A.M.; Bull, J.C.; Eastwood, D.C.; Kouvelis, V.N.; Bourdon, P.A.; Loveridge, E.J.; Mathias, S.; Meyrick, A.; Midthassel, A.; Myrta, A.; Butt, T. Evaluation of Metarhizium brunneum- and Metarhizium-derived VOCs as dual-active biostimulants and pest repellents in a wireworm-infested potato field. J. Fungi 2023, 9, 599. [Google Scholar] [CrossRef]
- Kishimoto, K.; Matsui, K.; Ozawa, R.; Takabayashi, J. Volatile 1-octen-3-ol induces a defensive response in Arabidopsis thaliana. J. Gen. Plant Pathol. 2007, 73, 35–37. [Google Scholar] [CrossRef]
- Khoja, S.; Eltayef, K.M.; Baxter, I.; Bull, J.C.; Loveridge, E.J.; Butt, T. Fungal volatile organic compounds show promise as potent molluscicides. Pest Manag. Sci. 2019, 75, 3392–3404. [Google Scholar] [CrossRef]
- Bourdon, P.A.; Zottele, M.; Baxter, I.; Myrta, A.; Midthassel, A.; Wechselberger, K.F.; Butt, T.M. Fumigation of three major soil pests (Agriotes lineatus, Diabrotica virgifera virgifera, Phyllopertha horticola) with 3-octanone and 1-octen-3-ol enantiomers. Biocontrol Sci. Technol. 2022, 32, 863–876. [Google Scholar] [CrossRef]
- Hummadi, E.H.; Dearden, A.; Generalovic, T.; Clunie, B.; Harrott, A.; Cetin, Y.; Demirbek, M.; Khoja, S.; Eastwood, D.; Dudley, E.; Hazir, S.; Touray, M.; Ulug, D.; Gulsen, S.H.; Cimen, H.; Butt, T. Volatile organic compounds of Metarhizium brunneum influence the efficacy of entomopathogenic nematodes in insect control. Biol. Control 2021, 155, 104527. [Google Scholar] [CrossRef] [PubMed]
- Khoja, S.; Eltayef, K.M.; Baxter, I.; Myrta, A.; Bull, J.C.; Butt, T. Volatiles of the entomopathogenic fungus Metarhizium brunneum attract and kill plant parasitic nematodes. Biol. Control 2021, 152, 104472. [Google Scholar] [CrossRef]
- Kline, D.L.; Allan, S.A.; Bernier, U.R.; Welch, C.H. Evaluation of the enantiomers of 1-octen-3-ol and 1-octyn-3-ol as attractants for mosquitoes associated with a freshwater swamp in Florida, USA. Med. Vet. Entomol. 2007, 21, 323–331. [Google Scholar] [CrossRef]
- Xu, P.; Zhu, F.; Buss, G.K.; Leal, W.S. 1-Octen-3-ol—the attractant that repels. F1000Research 2015, 4, 156. [Google Scholar] [CrossRef]
- Chitarra, G.S.; Abee, T.; Rombouts, F.M.; Posthumus, M.A.; Dijksterhuis, J. Germination of Penicillium paneum conidia is regulated by 1-octen-3-ol, a volatile self-inhibitor. Appl. Environ. Microbiol. 2004, 70, 2823–2829. [Google Scholar] [CrossRef] [PubMed]
- Veronico, P.; Sasanelli, N.; Troccoli, A.; Myrta, A.; Midthassel, A.; Butt, T. Evaluation of fungal volatile organic compounds for control of the plant-parasitic nematode Meloidogyne incognita. Plants 2023, 12, 1935. [Google Scholar] [CrossRef]
- Gourd, T.R.; Schmitt, D.P.; Barker, K.R. Differential sensitivity of Meloidogyne spp. and Heterodera glycines to selected nematicides. J. Nematol. 1993, 25, 746–751. [Google Scholar]
- Watson, T.T. Sensitivity of Meloidogyne enterolobii and Meloidogyne incognita to fluorinated nematicides. Pest Manag. Sci. 2022, 78, 1398–1406. [Google Scholar] [CrossRef]
- Jang, J.; Đăng, Q.; Choi, G.; Park, H.; Kim, J. Control of root-knot nematodes using Waltheria indica producing 4-quinolone alkaloids. Pest Manag. Sci. 2019, 75, 2264–2270. [Google Scholar] [CrossRef]
- Oka, Y. Nematicidal activity of fluensulfone against some migratory nematodes under laboratory conditions. Pest Manag. Sci. 2014, 70, 1850–1858. [Google Scholar] [CrossRef]
- Eisenback, J.D.; Hirschmann, H.; Sasser, J.N.; Triantaphyllou, A.C. A guide to the four most common species of root-knot nematodes (Meloidogyne spp.), with a pictorial key; North Carolina State University Graphics: Raleigh, NC, USA, 1981. [Google Scholar]
- Adam, M.A.M.; Phillips, M.S.; Blok, V.C. Molecular diagnostic key for identification of single juveniles of seven common and economically important species of root-knot nematode (Meloidogyne spp.). Plant Pathol. 2007, 56, 190–197. [Google Scholar] [CrossRef]
- Castagnone-Sereno, P.; Danchin, E.G.; Perfus-Barbeoch, L.; Abad, P. Diversity and evolution of root-knot nematodes, genus Meloidogyne: New insights from the genomic era. Annu. Rev. Phytopathol. 2013, 51, 203–220. [Google Scholar] [CrossRef] [PubMed]
- Terra, W.C.; Campos, V.P.; Martins, S.J.; Costa, L.S.A.S.; da Silva, J.C.P.; Barros, A.F.; Lopez, L.E.; Santos, T.C.N.; Smant, G.; Oliveira, D.F. Volatile organic molecules from Fusarium oxysporum strain 21 with nematicidal activity against Meloidogyne incognita. Crop Prot. 2018, 106, 125–131. [Google Scholar] [CrossRef]
- Huang, D.; Yu, C.; Shao, Z.; Cai, M.; Li, G.; Zheng, L.; Yu, Z.; Zhang, J. Identification and characterization of nematicidal volatile organic compounds from deep-sea Virgibacillus dokdonensis MCCC 1A00493. Molecules 2020, 25, 744. [Google Scholar] [CrossRef]
- Mei, X.; Wang, X.; Li, G. Pathogenicity and volatile nematicidal metabolites from Duddingtonia flagrans against Meloidogyne incognita. Microorganisms 2021, 9, 2268. [Google Scholar] [CrossRef] [PubMed]
- Freire, E.S.; Campos, V.P.; Pinho, R.S.; Oliveira, D.F.; Faria, M.R.; Pohlit, A.M.; Noberto, N.P.; Rezende, E.L.; Pfenning, L.H.; Silva, J.R. Volatile substances produced by Fusarium oxysporum from coffee rhizosphere and other microbes affect Meloidogyne incognita and Arthrobotrys conoides. J. Nematol. 2012, 44, 321–328. [Google Scholar]
- Eisenback, J.D.; Hunt, D.J. General morphology. In Root-knot Nematodes; Perry, R.N., Moens, M., Starr, F.J., Eds.; CAB International: Wallingford, UK, 2009; p. 23. [Google Scholar]
- Chen, W.; Wang, J.; Huang, D.; Cheng, W.; Shao, Z.; Cai, M.; Zheng, L.; Yu, Z.; Zhang, J. Volatile organic compounds from Bacillus aryabhattai MCCC 1K02966 with multiple modes against Meloidogyne incognita. Molecules 2022, 27, 103. [Google Scholar] [CrossRef]
- Deng, X.; Wang, X.; Li, G. Nematicidal effects of volatile organic compounds from microorganisms and plants on plant-parasitic nematodes. Microorganisms 2022, 10, 1201. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.-Q.; Mo, M.-H.; Zhou, J.-P.; Zou, C.-S.; Zhang, K.-Q. Evaluation and identification of potential organic nematicidal volatiles from soil bacteria. Soil Biol. Biochem. 2007, 39, 2567–2575. [Google Scholar] [CrossRef]
- Liarzi, O.; Bucki, P.; Braun Miyara, S.; Ezra, D. Bioactive volatiles from an endophytic Daldinia cf. concentrica isolate affect the viability of the plant parasitic nematode Meloidogyne javanica. PLoS ONE 2016, 11, e0168437. [Google Scholar] [CrossRef]
- Terra, W.C.; Campos, V.P.; Pedroso, M.P.; da Costa, A.L.; Freire, E.S.; de Pinto, I.P.; da Silva, J.C.P.; Lopez, L.E.; Santos, T.C.N. Volatile molecules of Fusarium oxysporum strain 21 are retained in water and control Meloidogyne incognita. Biol. Control 2017, 112, 34–40. [Google Scholar] [CrossRef]
- Inamdar, A.A.; Hossain, M.M.; Bernstein, A.I.; Miller, G.W.; Richardson, J.R.; Bennett, J.W. Fungal-derived semiochemical 1-octen-3-ol disrupts dopamine packaging and causes neurodegeneration. Proc. Natl. Acad. Sci. USA 2013, 110, 19561–19566. [Google Scholar] [CrossRef]
- Hussey, R.S.; Barker, K.R. A comparison of methods of collecting inocula of Meloidogyne spp., including a new technique. Plant Dis. Rep. 1973, 57, 1025–1028. [Google Scholar]
Figure 1.
a, b. Mean percent mortality for five Meloidogyne species: M. arenaria, M. incognita, M. luci, M. javanica, and M. hapla, 24 hours after exposure to varying concentrations of 1-octen-3-ol (a) and 3-octanone (b). Each line represents the average lethality for a given species across concentrations, with error bars indicating 95% confidence intervals.
Figure 1.
a, b. Mean percent mortality for five Meloidogyne species: M. arenaria, M. incognita, M. luci, M. javanica, and M. hapla, 24 hours after exposure to varying concentrations of 1-octen-3-ol (a) and 3-octanone (b). Each line represents the average lethality for a given species across concentrations, with error bars indicating 95% confidence intervals.
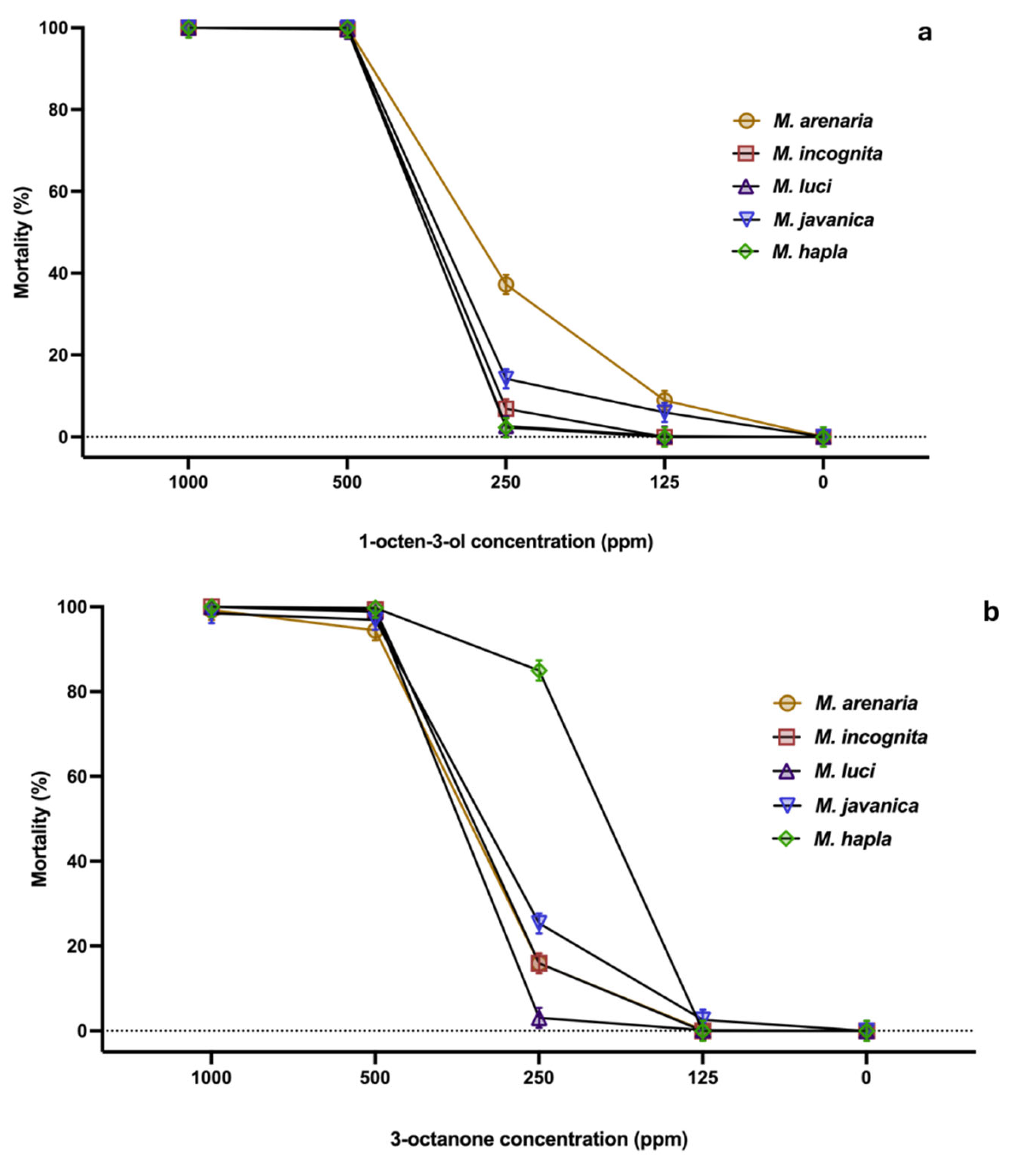
Figure 2.
a-c. Mean percentage of differentiated M. incognita eggs after exposure to increasing concentrations of 1-octen-3-ol and 3-octanone across three time points 7 days (a), 14 days (b) and 21 days (c). Egg differentiation rates were calculated per replicate as the percentage of eggs hatched relative to day 0, and the graph shows the mean of these relative percentages. Error bars indicate 95% confidence intervals. 5 replicates were used per treatment, and the experiment was repeated 4 times (n = 20 per treatment). Hatching rates generally declined with increasing compound concentration and exposure time. Different lowercase letters indicate statistically significant differences among concentrations within each compound. Treatments that do not share a letter (within each compound) differ significantly.
Figure 2.
a-c. Mean percentage of differentiated M. incognita eggs after exposure to increasing concentrations of 1-octen-3-ol and 3-octanone across three time points 7 days (a), 14 days (b) and 21 days (c). Egg differentiation rates were calculated per replicate as the percentage of eggs hatched relative to day 0, and the graph shows the mean of these relative percentages. Error bars indicate 95% confidence intervals. 5 replicates were used per treatment, and the experiment was repeated 4 times (n = 20 per treatment). Hatching rates generally declined with increasing compound concentration and exposure time. Different lowercase letters indicate statistically significant differences among concentrations within each compound. Treatments that do not share a letter (within each compound) differ significantly.
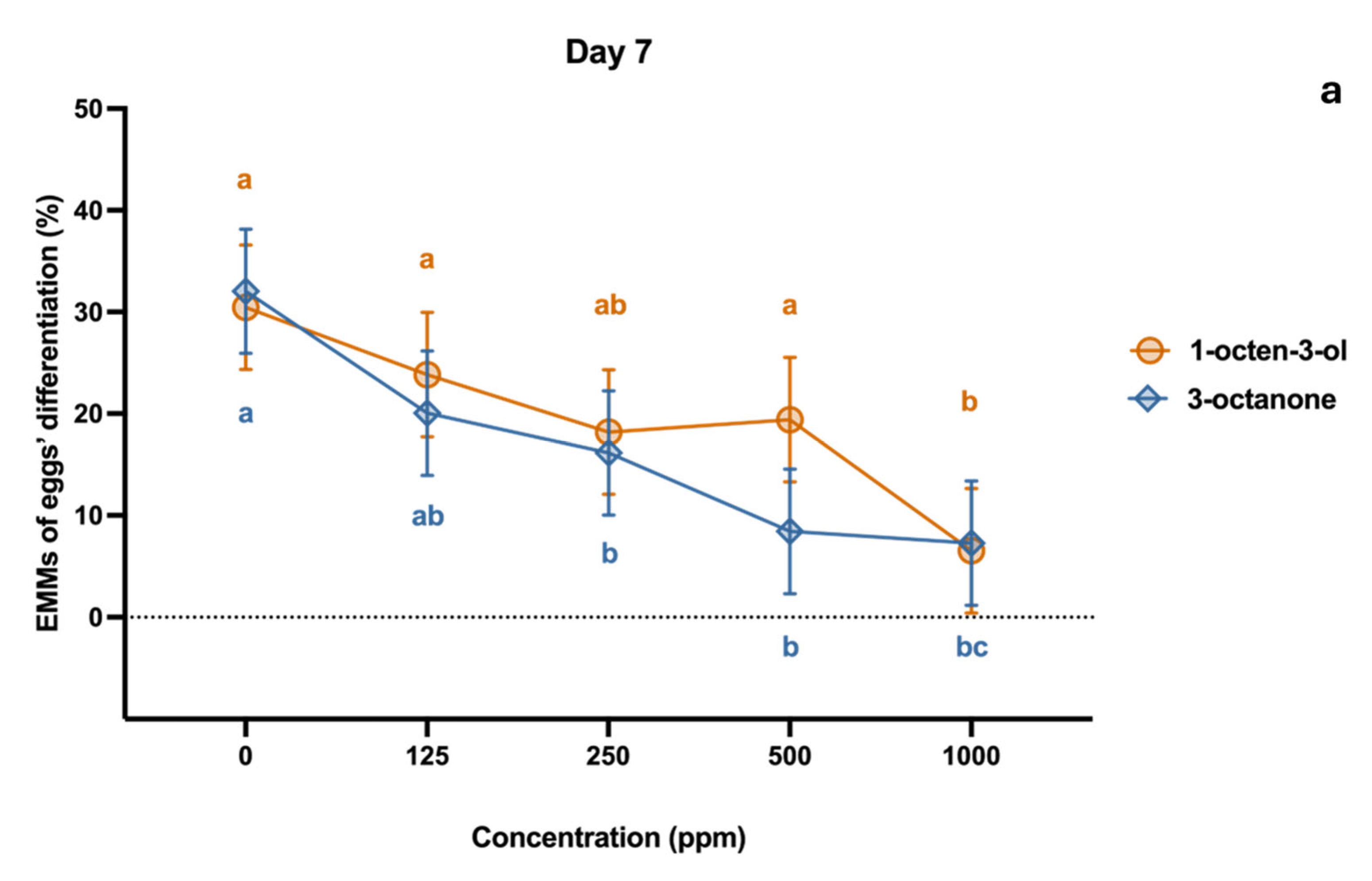
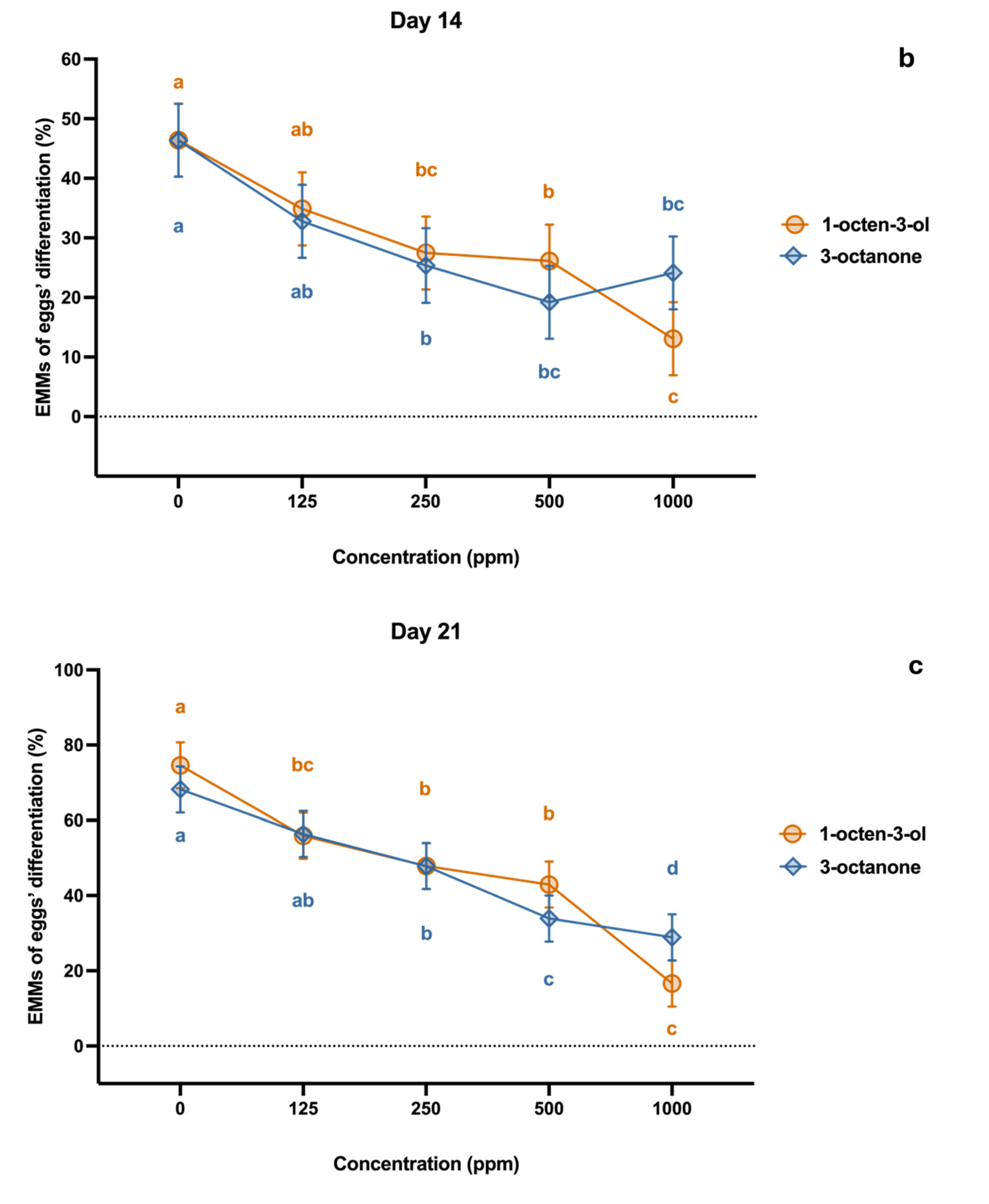
Figure 3.
Effect of 1-octen-3-ol and 3-octanone concentration on egg hatching from egg masses. Estimated marginal means (EMMs) of hatched eggs of M. javanica are shown for each compound, with error bars indicating 95% confidence intervals. Both compounds significantly reduced hatching compared to the control. 1-octen-3-ol caused consistent suppression compared to the control across all concentrations tested (125-1000 ppm), with no significant differences in efficacy among doses (125-1000ppm). In contrast, 3-octanone showed reduced efficacy at lower concentrations and greater suppression at 500 and 1000 ppm, compared to the lower doses. Letters indicate statistically distinct groups based on pairwise comparisons (p < 0.05).
Figure 3.
Effect of 1-octen-3-ol and 3-octanone concentration on egg hatching from egg masses. Estimated marginal means (EMMs) of hatched eggs of M. javanica are shown for each compound, with error bars indicating 95% confidence intervals. Both compounds significantly reduced hatching compared to the control. 1-octen-3-ol caused consistent suppression compared to the control across all concentrations tested (125-1000 ppm), with no significant differences in efficacy among doses (125-1000ppm). In contrast, 3-octanone showed reduced efficacy at lower concentrations and greater suppression at 500 and 1000 ppm, compared to the lower doses. Letters indicate statistically distinct groups based on pairwise comparisons (p < 0.05).
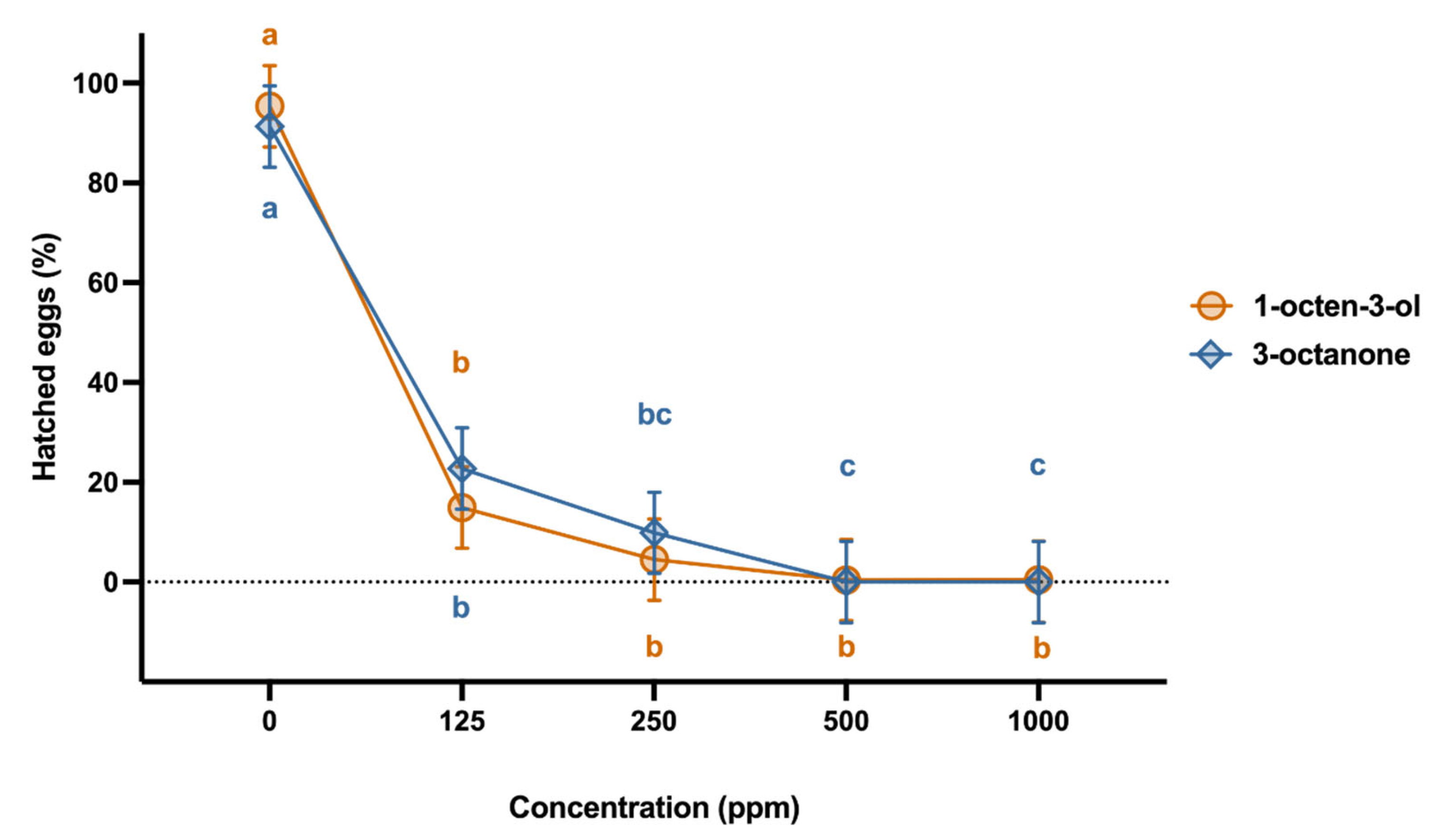
Figure 4.
Number of viable M. javanica J2s after treatment with different concentrations of 1-octen-3-ol and 3-octanone in soil assays. The numbers on the y-axis represent the mean numbers of viable J2s counted following the 24h exposure. Error bars represent standard error of mean values.
Figure 4.
Number of viable M. javanica J2s after treatment with different concentrations of 1-octen-3-ol and 3-octanone in soil assays. The numbers on the y-axis represent the mean numbers of viable J2s counted following the 24h exposure. Error bars represent standard error of mean values.
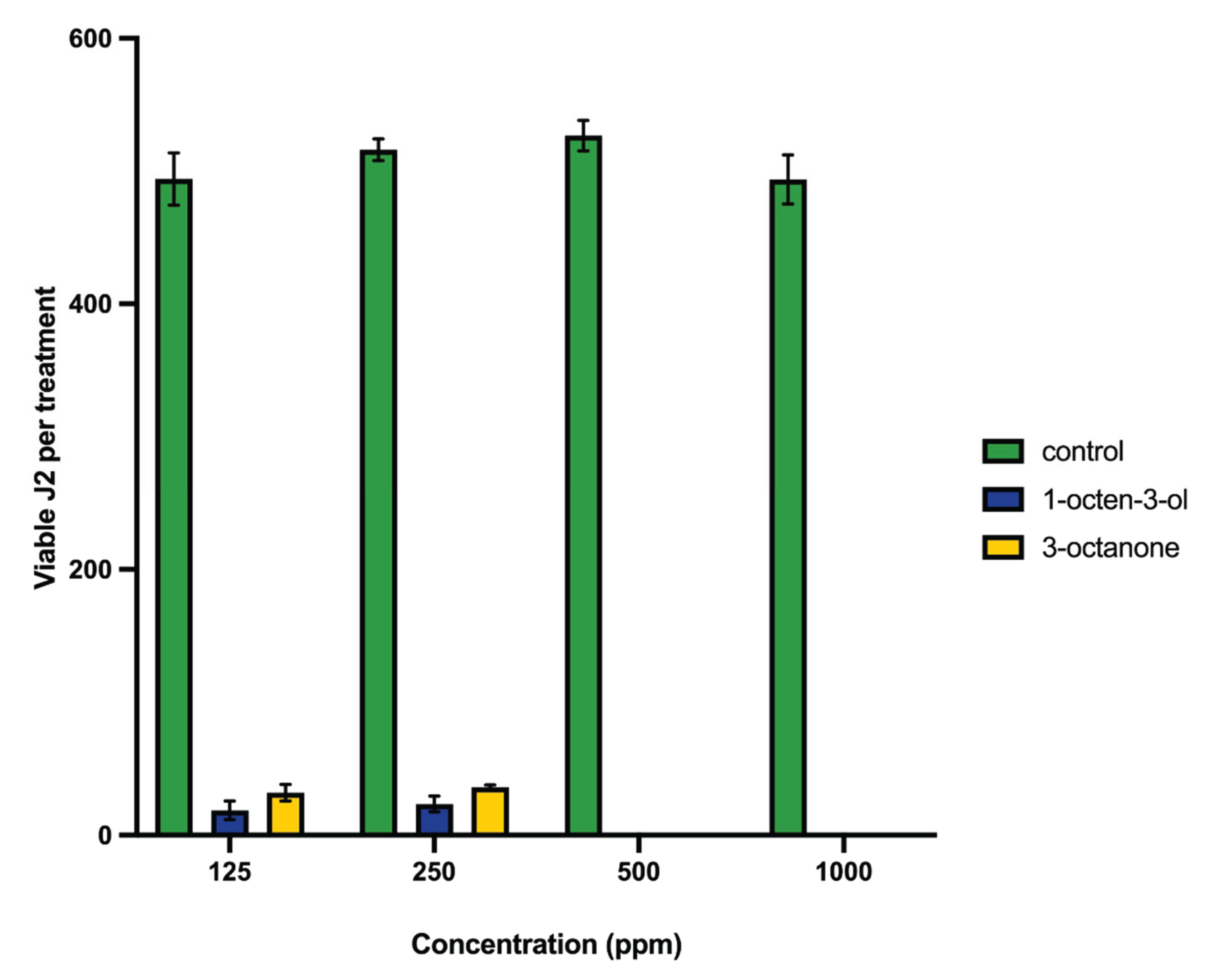

Table 1.
Summary of statistical analysis results testing the effects of compound type, nematode species, and VOC concentration on nematode percent mortality. The model includes main effects (Compound, Species, Concentration), all possible two-way interactions, and the three-way interaction. All factors and interactions were statistically significant (p < 0.001). The model explained 99.4% of the total variance (R² = 0.994; Adjusted R² = 0.994).
Table 1.
Summary of statistical analysis results testing the effects of compound type, nematode species, and VOC concentration on nematode percent mortality. The model includes main effects (Compound, Species, Concentration), all possible two-way interactions, and the three-way interaction. All factors and interactions were statistically significant (p < 0.001). The model explained 99.4% of the total variance (R² = 0.994; Adjusted R² = 0.994).
| Source | Type III Sum of Squares | df | Mean Square | F | Significance |
| Corrected Model | 1095061 | 49 | 22348.19 | 1575.517 | <0.001 |
| Intercept | 979282.6 | 1 | 979282.6 | 69038.09 | <0.001 |
| Compound | 663.46 | 1 | 663.46 | 46.773 | <0.001 |
| Species | 4043.962 | 4 | 1010.99 | 71.273 | <0.001 |
| Concentration | 1037025 | 4 | 259256.2 | 18277.21 | <0.001 |
| Compound x Species | 7537.917 | 4 | 1884.479 | 132.853 | <0.001 |
| Compound x Concentration | 6316.356 | 4 | 1579.089 | 111.324 | <0.001 |
| Species x Concentration | 15836.88 | 16 | 989.805 | 69.78 | <0.001 |
| Compound x Species x Concentration | 23638.1 | 16 | 1477.381 | 104.153 | <0.001 |
| Error | 6383.102 | 450 | 14.185 | ||
| Total | 2080727 | 500 | |||
| Corrected Total | 1101444 | 499 |
R² = 0.994 Adjusted R²= 0.994.
Table 2.
Results of the general linear model (GLM) analyzing the effects of compound, time (days), and VOC concentration on the percentage of egg differentiation in M. javanica. Significant main effects were observed for Day, Concentration, and the Compound × Concentration and Day × Concentration interaction terms (p < 0.001). However, the effect of Compound alone was insignificant (p = 0.311), nor were the Compound × Day or Compound × Day × Concentration interactions. This indicates that differential suppression was primarily driven by exposure duration and compound concentration, rather than compound identity alone.
Table 2.
Results of the general linear model (GLM) analyzing the effects of compound, time (days), and VOC concentration on the percentage of egg differentiation in M. javanica. Significant main effects were observed for Day, Concentration, and the Compound × Concentration and Day × Concentration interaction terms (p < 0.001). However, the effect of Compound alone was insignificant (p = 0.311), nor were the Compound × Day or Compound × Day × Concentration interactions. This indicates that differential suppression was primarily driven by exposure duration and compound concentration, rather than compound identity alone.
| Source | Type III Sum of Squares | df | Mean Square | F | Significance | |
| Corrected Model | 177800.038 | 29 | 6131.036 | 31.631 | <0.001 | |
| Intercept | 602241.240 | 1 | 602241.240 | 3107.101 | <0.001 | |
| Compound | 199.341 | 1 | 199.341 | 1.028 | 0.311 | |
| Day | 85883.546 | 3 | 42941.773 | 221.546 | <0.001 | |
| Concentration | 77365.078 | 4 | 19341.269 | 99.786 | <0.001 | |
| Compound × Day | 234.110 | 2 | 117.055 | 0.604 | 0.547 | |
| Compound × Concentration | 4388.443 | 4 | 1097.111 | 5.660 | <0.001 | |
| Day × Concentration | 8636.658 | 8 | 1079.582 | 5.570 | <0.001 | |
| Compound × Day × Concentration | 1121.377 | 8 | 140.172 | 0.723 | 0.671 | |
| Error | 110287.779 | 569 | 193.827 | |||
| Total | 890782.409 | 599 | ||||
| Corrected Total | 288087.817 | 598 | ||||
R² = 0.617, Adjusted R² = 0.598.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



