
Submitted:
09 May 2023
Posted:
12 May 2023
You are already at the latest version
Abstract
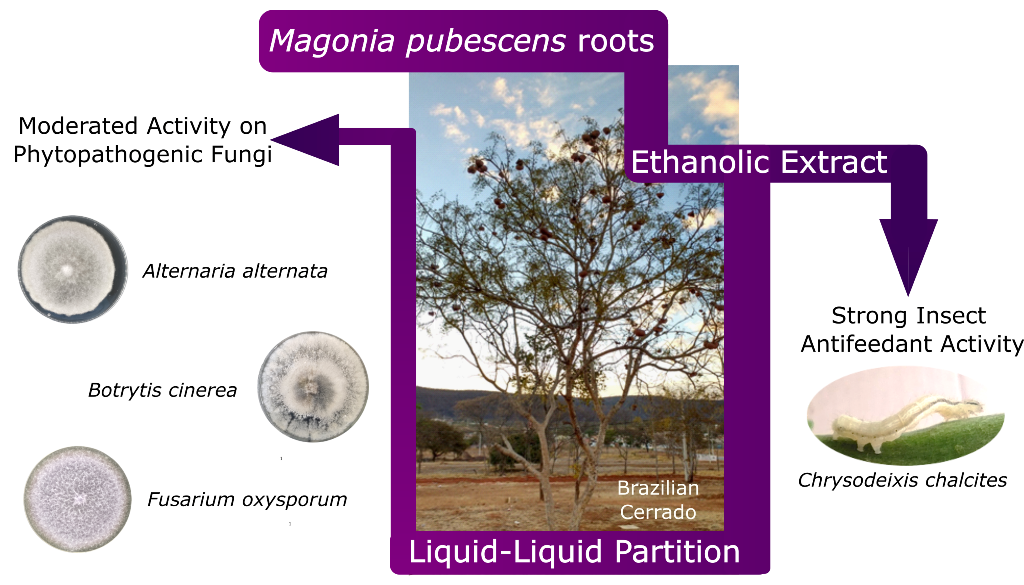
Botanical biopesticides have emerged as sustainable and eco-friendly alternatives to synthetic pesticides, whose indiscriminate use leads to several drawbacks to human and environment health. The present study reports on the evaluation of the roots extract from Magonia pubescens, species from the Cerrado (Brazilian savannah), on the phytopathogenic fungi, Alternaria alternata, Botrytis cinerea, and Fusarium oxysporum. In addition, its insect antifeedant effect was assayed against Chrysodeixis chalcites. Thus, an in vitro test-assay was used to determine the fungicide potential (percentage growth inhibition, % GI) of the ethanolic extract of this plant species, whereas a leaf-disk bioassay on the 5th instar larvae of C. chalcites was performed to evaluate the insecticide potential. The ethanolic extract was further fractionated by liquid-liquid extraction, using organic solvents of increasing polarity. The hexane/dichloromethane fraction exhibited a moderated potency, and similar to the ethanolic extract on the three assayed fungi (around % GI 30 at 1 mg/mL), whereas the n-butanol fraction showed a slight improvement of the fungicide effect against B. cinerea (% GI 39.18 at 1 mg/mL). Moreover, the etanolic extract exhibited a strong antifeedant activity, with a refusal rate (FR) higher than 90% in both choice and non-choice assays against C. chalcites, while the ethyl acetate and n-butanol fractions behaved as appetite suppressors. These results highlight M. pubescens as a promising source of biopesticides and deserve further investigations to optimize extraction procedures.
Keywords:
Magonia pubescens
; biopesticides
; phytopathogenic fungi
; Chrysodeixis chalcites
1. Introduction
The Food and Agriculture Organization of the United Nations (FAO) estimates that the world population will increase by over 32% by 2050, which will mean an increase in the demand for food [1]. This fact highlights the need to adopt strategies that ensure the efficiency, productivity, sustainability, and security of our food supply. One of the challenges to achieve these goals is the control of pests affecting crops, which, in recent times. have increased due to globalization, climate change and the emergence of resistance to conventional pesticides [2]. Among the pests that most severely affect crops, fungal infections via ascomycetes, such as Botrytis cinerea [3], Fusarium oxysporum [4], and Alternaria alternata [5], have a great impact worldwide. In Brazil, B. cinerea affects several crops, mainly eucalyptus in the southwest region [6], and grapes [7] and strawberries [8] in the south. Recent studies show that some strains of this fungus are resistant to some commonly used fungicides such as external quinone inhibitors (QoI) [9]. Moreover, F. oxysporum causes banana Fusarium wilt, a pandemic that has affected crops and the export of this fruit in Brazil, which is the 4th largest banana producer in the world [10]. Furthermore, crops in this country have also been affected by different lineages of the fungus, which seem to differ between regions [11]. Regarding Alternaria brown spot caused by A. alternata, this was first reported in 2001 and spread quickly in the country, affecting mainly tangerine crops, in which Brazil is the 5th largest producer [12]. Moreover, there are few fungicides registered in the country able to control the spread of this fungus, and there is evidence of the emergence of resistant strains of this pathogen [9].
Furthermore, Chrysodeixis chalcites (Esper) (Lepidoptera: Noctuidae), known as the golden twin-spot moth, is a highly polyphagous species that causes serious damage to crop in many regions of Europe, the Mediterranean and the Middle East and Africa [13]. The larvae of this insect feed on several fruit and horticultural crops, such as bananas (Musa sp. L.), cabbages (Brassica oleracea L.), beans (Phaseolus vulgaris L.), corn (Zea mays L.), potatoes (Solanum tuberosum L.), cucumbers (Cucumis sativus L.), peppers (Piper nigrum L.), soybeans (Glycine max L.) or tomatoes (Solanum lycopersicum L.) [14,15]. The control of C. chalcites is mainly based on multiple applications of a few active substances authorized by the European Union throughout the crop cycle [16]. Moreover, Chrysodeixis includens is a pest that affects crops in North and South America, especially in the USA, Argentina, and Brazil. This insect causes economic losses in many cultivars, like soybeans, cotton, and tomatoes. In Brazil, the insect is also one of the major defoliators of soybean crops, especially relevant as the country is the world’s leading soybean producer and exporter [17]. Usually, C. includens is seen as a secondary pest, naturally controlled by parasitoids and entomopathogenic fungi, but the increased use of fungicides has allowed the growth of larvae population, triggering the need for new control agents [18].
Currently, crop protection relies almost exclusively on the use of synthetic pesticides [2]. However, their indiscriminate use has aroused social concerns due to overwhelming evidence of their detrimental effects on the environment and human health, increasing the risks for development resistance strains, and the erratic supply of beneficial organisms [19]. Therefore, there is a need for research into eco-friendly pesticides for use in agricultural pest management programs. Botanical biopesticides are considered as safe, ecologically sound, and sustainable alternatives to synthetic pesticides [20,21]. They have key advantages over traditional pesticides, including, lower toxicity, easily biodegradable (less persistent in the environment), exclusively affecting the pest in question and closely related species, new modes of action, and can be used in organic agriculture [22]. Although biopesticides have less effectiveness and take more time for disease control compared to synthetic pesticides, which give quick results, they are gaining popularity by ensuring environmental health versus harmful indiscriminate usage of synthetic pesticides [2]. In fact, several plants and plant-derived products have been reported as botanical biopesticides [23,24]. In addition, metabolites with insecticidal effects have been reported from several plants, such as Azadirachtin, isolated from Azadirachta indica, and pyrethrins from Tanacetum cinerariifolium. These are two examples of successful natural insecticides, which are the basis of most current, commercial botanical insecticides [20].
Sapindaceae family comprises about 140 genera and 1,900 species, distributed mainly in tropical regions of the world. In Brazil, this botanical family is represented by 28 genera and 417 species [25]. Species of this family have high economic impact as they are used as wood for civil construction and handicrafts, as a food source, and from an ethnopharmacology point of view, some species are used in the treatment of fever, ulcers, pain, dermatological problems, and dysentery, among others [26]. Magonia pubescens A. St.-Hil. (Sapindaceae) is an endemic species from Brazil popularly known as ‘tingui’ or ‘timbó’. It is the only species of the Magonia genus and is widely distributed in the states of Goiás, Mato Grosso, Mato Grosso do Sul and Minas Gerais [27] in the Brazilian savannah (the Cerrado), a biodiversity hotspot in the center of the country [28]. The fruits and seeds of the plant are used to prepare a soap to treat dermatitis, lice infestations and as an insecticide and larvicide [29]. Larvicidal activity of ethanolic extracts from the M. pubescens stem have been reported against different species of the Aedes genera, the main vector of the dengue disease [30]. There is also a report on the leishmanicidal activity of the stem bark ethanolic extract [31]. The volatile and flavonoid profile in flowers and leaves from M. pubescens have also been investigated with results suggesting it could be effective against free radicals [32]. In addition, essential oils from M. pubescens inflorescences and their cytotoxic activity has been studied [33]. Root infusions are employed in fishing, through the usual technique of poisoning water with extracts. Infusions of the roots are also used as a tranquilizer, and its ethanolic extracts are used to treat wounds and pain [34]. This extract shows inhibitory activity against insects, fungi, and bacteria. In addition, the ethanolic extract of M. pubescens roots showed strong cytotoxic activity against MDA-MB-435 breast cancer cell lines (IC50 7.9 μg/mL) [29]. Furthermore, a study on the insecticide irritability effect of the ethanol extract from the roots of M. pubescens demonstrated that this induced significantly greater insecticide-irritability behavior than the control, permethrin, against Sitophilus zeamais in stored maize [35]. Analysis of the external coat mucilage from M. pubescens seeds has also been reported [36].
Despite the potential of M. pubescens, limited studies have been performed on this plant. The aim of the present study is to investigate the potential of M. pubescens roots extract as a biopesticide. Herein, the evaluation of the ethanolic extract of the roots from this species on the phytopathogenic fungi, Alternaria alternata, Botritis cinerea, and Fusarium oxysporum is reported. In addition, the insecticide effect of the plant was evaluated on Chrysodeixis chalcites, insect causing one of the most serious lepidopteran pests in economically important crops. The results of this study highlight the potential of M. pubescens as a source of eco-friendly biopesticides to suppress highly phytopathogenic species that cause crop losses worldwide.
2. Materials and Methods
2.1. Chemicals and Reagents
All solvents were analytical grade (Panreac, Barcelona, Spain). Polygram Sil G/UV254 foils used for analytical thin-layer chromatography (TLC) were purchased from Macherey-Nagel. PGA culture medium (Potato glucose agar, Sigma Aldrich, St Louis, MO, USA) and Petri dishes (9 cm diameter) from Sarstedt (Nümbrecht, Germany) were used for the maintenance of fungal colonies and performance of bioassays. Tetracycline (50 mg/L, Sigma Aldrich, St Louis, MO, USA) was added to avoid bacterial growth on Petri dishes. Agarose (Sigma Aldrich, St. Louis, MO, USA, 30 g/L) was used for insect bioassays. Stock solutions of plant extract/fractions were prepared with absolute ethanol (Sigma Aldrich, St Louis, MO, USA).
2.2. Plant Material
Roots of M. pubescens A.St.-Hil. were collected in the municipality of Montes Claros (geographical coordinates 16º 25 ’26’’S, 43º 32’ 10’’O) in the north of Minas Gerais - Brazil in October 2019 by Maria Olívia Mercadante-Simões, PhD (UNIMONTES). A voucher specimen was deposited at the University Federal Minas Gerais (UFMG) herbarium under No. 106750. The plant material was registered at Conselho de Gestão do Patrimônio Genético (CGEN/SisGen) under voucher number AAC6C0E.
2.3. Plant Extract Preparation
The air-dried, powdered roots of Magonia pubescens (1.0 Kg) were extracted by exhaustive maceration (four times) with 98% ethanol (10 mL of ethanol/g of plant material) at room temperature for 94 h for each maceration process. The extractive solvent was removed under reduced pressure at 40 °C using a rotary evaporator, yielding 123 g of residue extract. Aliquots of this extract were kept for further antifungal (30 mg) and antifeedant (150 mg) evaluation.
2.4. Liquid–liquid Partition Procedure
The ethanolic plant extract was sequentially fractionated by liquid–liquid partition using a series of solvents of increasing polarity by a modification of the Kupchan method [37]. Thus, the crude extract was dissolved in a mixture of methanol/water (2:8), and fractionated with hexanes (500 mL), dichloromethane (500 mL), ethyl acetate (4 x 500 mL), n-butanol (4 x 500 mL) and water (4 x 500 mL). The organic phases were dried with Na2SO4, filtered, and solvents removed under reduced pressure to afford the corresponding organic fractions. The aqueous phase was lyophilized, thereby yielding the aqueous fraction. A preliminary Thin Layer Chromatography (TLC) analysis of the fractions revealed a similar phytochemical profile of the hexane and dichloromethane fractions, and these were combined. Thus, fractions extracted with hexane/ dichloromethane (8.7 g), ethyl acetate (46.0 g), n-butanol (47.6 g) and water (18.4 g) were obtained, and an aliquot of each was used for biological evaluation.
2.5. In Vitro Test-assay on Mycelium
The phytopathogenic fungi used in this bioassay belong to cosmopolitan genera that cause serious crop damage: Alternaria alternata, Botrytis cinerea, and Fusarium oxysporum. These fungi were maintained at 25 ᴼC in darkness, and periodically replicated in Petri dishes with PGA culture medium and tetracycline to avoid contamination. Strains of B. cinerea (B05.10) and A. alternata (Aa 100), isolated from Vitis vinifera and Lycopersicon esculentum, respectively, were supplied by the Universidad de La Laguna, Tenerife. F. oxysporum f. sp. lycopersici (2715) strain, isolated from Lycopersicon esculentum, was provided by the Colección Española de Cultivos Tipo (CECT) from Valencia, Spain. Antifungal activity was analyzed through mycelial growth inhibition using an agar-dilution method [38]. Once the culture medium solidified, eight 4 mm diameter discs of the target fungus per dish were placed in test and control Petri dishes. One dish per pathogen for each sample with eight sub-replicates in each dish was used. A stock solution of 50 mg/mL was prepared, using ethanol as solvent, and samples were assayed at the concentration of 1 mg/mL, and those showing a % I higher than 20% at a concentration of 1 mg/mL were then tested at lower doses (0.5 and 0.1 mg/mL). Colonies grown on Petri dishes were incubated for 48 h for B. cinerea and 72 h for A. alternata and F. oxysporum and were digitalized and measured with the application ImageJ (Image J, http://rsb.info.nih.gov/ij). Percent growth inhibition (% I) was calculated as % I = [(C – T)/C] × 100, where C is the diameter of the control colonies and T is the diameter of the test colonies. Fosbel-Plus (Probelte S.A., Murcia, Spain), used as a reference fungicide [39], is a commercial product purchased in a phytosanitary product store, whereas ethanol was used as a negative control, using one dish per pathogen and 8 discs for each control.
2.6. Leaf-Disk Bioassay
A laboratory colony of Chrysodeixis chalcites was initiated using larvae collected from banana crops in Tenerife. Larvae were reared on an artificial diet [40] at 21 ºC, 70% relative humidity and a photoperiod of 16:8 h (L:D) in a growth chamber. Adults were fed 30% w/v with diluted honey [41], and the newly fifth-instar larvae of C. chalcites was used in the antifeedant bioassays. Indoxacarb 30% (Steward 30WG, FMC Agricultural Solutions, Madrid, Spain) [42], a commercial insecticide was used as a positive control for comparative purposes.
The bioassay was based on the protocol described by Escoubas et al. [43] and González-Coloma et al. [44] with modifications. Thus, petri dishes with a banana leaf (grown in a greenhouse free of chemical treatments), and a 5 mm deep agarose layer, which delineates intake zones and prevents the leaf disk from drying out during the assay, were prepared. Each petri dishes had nine circular holes (1 cm diameter each) so that the larvae can access the leaf. Leaves were previously cleaned with distilled water and left to dry. In the choice assay, leaf disks (5 cm diameter) were alternatively treated with the tested sample solution in ethanol (5 µL per disk–0.2 mg/cm2), or solvent as control. In the non-choice assay, all disks of the same plate were treated with the test solution at the same concentration, or solvent. After complete evaporation of the solvent, three fifth-instar larvae of C. chalcites were placed in each petri dishes (6 replicates for each experiment) and allowed to feed in a growth chamber. The duration of the bioassay under these conditions was 20 h. Results were expressed in terms of refusal rate (FR) [45]. To calculate the FR in each dish, the leaf area consumed was measured with the ImageJ 1.53e program [46], and the refusal rate (FR) calculated using the following formula: FR = [1 - (% Treated disks consumed / % Control disks consumed)] x 100.
3. Results and Discussion
To our knowledge, there have been no reports on the bioactivity of M. pubescens on phytopathogens affecting crops as a potential fungicide or antifeedant. This has encouraged us to investigate the potential of the roots of this plant as a source of biopesticides.
3.1. Antifungal In Vitro Test-assay on Mycelium
First, the ethanolic extract of the roots from Magonia pubescens was evaluated against Botrytis cinerea, Fusarium oxysporum, and Alternaria alternata (Figure 1, Figure 2 and Figure 3, and Table S1 in Supplementary Materials), phytopathogenic fungi that affect crops worldwide, in particular, banana and tangerine crops, which are of economic relevance in Brazil. The evaluation of this ethanolic extract (EE) showed percentage inhibition (% GI) of 30.90, 33.63 and 31,88 against A. alternata, B. cinerea and F. oxysporum, respectively, at the concentration of 1 mg/mL. These results suggest some degree of antifungal activity against the tested fungi. Subsequently, in order to enrich this extract in bioactive components, the ethanolic crude extract was subjected to a liquid-liquid partition into five fractions, hexanes (F-H), dichloromethane (F-D), ethyl acetate (F-E), n-butanol (F-B) and water (F-W) fractions. A preliminary Thin Layer Chromatography (TLC) analysis of these fractions revealed similar phytochemical profiles of hexanes and dichloromethane fractions, and these were combined (F-H/D). These fractions were further evaluated against the three phytopathogenic fungi. The extract and fractions were first tested at a concentration of 1 mg/mL, and those exhibiting growth inhibition higher than 20% were tested at lower concentrations, namely, 0.5 and 0.1 mg/mL. Fosbel plus was used as a reference fungicide [39], whereas ethanol was used as a negative control, using one dish per pathogen and 8 disks for each control.
Results obtained on A. alternata (Figure 1) revealed that fractions F-H/D and F-B (% GI 27.10 and 29.35, respectively) showed similar potency to the crude extract (% GI 30.90), whereas the F-E and F-W fractions were inactive (GI < 10% at 1 mg/mL concentration). Thus, this procedure resulted in maintaining the activity of the two organic fractions, hexanes/dichloromethane and n-butanol fractions in comparison with the ethanolic extract at all the assayed concentrations, but not an enrichment in bioactive components.
Likewise, antifungal activity of the ethanolic extract and fractions on B. cinerea (Figure 2) led to a similar effect of the F-H/D fraction (% GI 31.74 at 1 mg/mL), and a slight increase in activity of the F-B fraction (% GI 39.18 at 1 mg/mL) compared to the ethanolic extract (% GI 33.63 at 1 mg/mL). However, the F-E and F-W showed low potency to inactivity with respect to the EE.
F. oxysporum (Figure 3) showed similar behavior to the extract and fractions as the other two assayed fungi. Thus, F-H/D (% GI 28,34) and F-B (% GI 32,15) fractions exhibited similar effects as the ethanolic extract (% GI 31.88), whereas F-E and F-W fractions were almost inactive (GI around or lower than 10% at 1 mg/mL).
The F-W fraction showed low to non-activity on the three phypathogenic fungi, revealing that the components in the plant responsible for antifungal activity have medium to low polarity. Moreover, although none of the fractions exhibited higher growth inhibition than Fosbel-Plus, used as a positive control, these potential biopesticides are expected to be less harmful than synthetic pesticides, making their use more desirable.
3.2. Insect Antifeedant Activity against Chrysodeixis chalcites
The ethanolic extract and fractions from M. pubescens roots were evaluated on Chrysodeixis chalcites, an insect that causes severe damage to crop worldwide. In this assay, samples were dissolved in ethanol, and 5 µL from a stock solution of 40 mg/mL per disk 0.2 mg/cm2 were used. Indoxacarb 30% (Steward 30WG), a frequently used synthetic insecticide, was used as a positive control, whereas solvent was used as a negative control. Indoxacarb, used in the chemical control of banana crops, is moderately harmful for beneficial insects [18]. Under the experimental conditions described in the previous section in the choice assay and the non-choice assays, three five-instar larvae of C. chalcites were placed in each petri dish (six replicates for each experiment), and after a 20-h period, the ingested leaf area was measured, and the refusal rate (FR) was calculated. A value of FR > 50% in the choice or non-choice assays indicates a significant feeding inhibition [44]. The combination of FR values > 50% at a dose of 0.2 mg/cm2 in both assays indicate a strong antifeedant effect. Samples tested with a combination of FR < 50% were considered inactive (Table 1).
The results indicate that the crude extract (EE) had a FR > 90% in the choice and non-choice assays, meaning that the extract acts very effectively as an antifeedant for C. chalcites larvae. Subsequently, hexane/dichloromethane (F-H/D), ethyl acetate (F-E), n-butanol (F-B) and water (F-W) fractions obtained by liquid-liquid partition of the ethanolic crude extract were further evaluated against C. chalcites. The results revealed that the hexane/dichloromethane fraction (F-H/D) acts as a weak insect antifeedant, since in the choice assay larvae preferred to feed on untreated disks (FR = 60.52%), indicating good activity, but in the non-choice assay, the larvae fed from the treated disks (FR = -27.67%), so it does not have a complete antifeedant effect. Moreover, the ethyl acetate (F-E) and n-butanol (F-B) fractions showed positive FR values in the non-choice assay (FR 61.10% and 60.71%, respectively), though activity was not confirmed by the choice assay (low FR values). Thus, F-E and F-B fractions are appetite suppressants since larvae does not feed of treated disks in the non-choice assay. On the other hand, the water (F-W) fraction was inactive, showing FR values of -74.57 and 44.73%, respectively, in the assays.
Notably, the crude ethanolic extract showed similar effects as Steward 30WG [42], the commercial insecticide used as a positive control, exhibiting a FR of 92.14% and 95.16% in the choice and non-choice feeding assays, respectively. This fact suggest that the plant could be used, after a simple and accessible procedure as a biopesticide against this phytopathogenic insect.
The only study regarding the potential insecticide activity of M. pubescens is by Silva et al [35], who reported the insecticide irritability effect (avoidance after contact) similar to permethrin in its stem ethanolic extract against Sitophilus zeamais (maize weevil), a serious pest of stored maize. However, the present study is the first report regarding the antifeedant potential of M. pubescens roots. Furthermore, several chemical components have been identified in different parts of the plant, including flavonoids and volatile oils that are constituents of its flowers and leaves [32], and essential oils from inflorescences [33]. However, phytochemical analysis of the roots has not been reported. The findings reported herein highlight the effect of M. pubescens roots on phytopathogen insects and deserve future phytochemical investigation to identify the metabolites responsible for this effect with a view to developing biopesticides.
5. Conclusions
In summary, the ethanolic extract from Magonia pubescens roots was moderately efficient at suppressing the mycelial growth of the phytopathogenic fungi, Alternaria alternata, Brotrytis cinerea and Fusarium oxysporum, with a percentage of inhibition around 30 at 1 mg/mL. The liquid-liquid partition led to organic fractions exhibiting similar potency to the ethanolic extract, whereas the aqueous fraction showed from low to non-activity on the three phypathogenic fungi, indicating that the antifungal components possess range from low to medium polarity.
Furthermore, the ethanolic extract displayed a strong insect antifeedant activity against Chrysodeixis chalcites larvae, showing refusal rate values higher than 92% in both, the choice and non-choice feeding assays, comparable to the commercial insecticide used as a positive control, Steward 30WG. However, the organic fractions from the liquid-liquid fractionation behavior as appetite suppressors since larvae does not feed of treated disks in the non-choice assay, suggesting a synergism of constituents in the ethanolic extract.
Overall, M. pubescens roots could be applied in combination with other biological agents in an environmentally safe and economically acceptable integrated management of crop diseases, especially against C. chalcites insect. However, additional research is required to optimize extraction procedures and corroborate the results in field trials.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Antifungal effects (% growth inhibition) of plant extract, and fractions from Magonia pubescens roots against Alternaria alternata, Botrytis cinerea, and Fusarium oxysporum.
Author Contributions
Conceptualization, I.L.B. and I.A.J.; methodology, I.A.J., C.G. and L.P.D.; investigation, A.R.A.M., S.R.S., D.G.E., and G.E.; writing—original draft preparation, A.R.A.M., S.R.S. and G.F.S.; writing—review and editing, I.L.B. and R.C.; funding acquisition, I.L.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the project MACBIOPEST (MAC2/1.1a/289), the program Interreg MAC 2014-2020 co-funded by the European Regional Development Fund (FEDER), and the RTI2018-094356-B-C21 Spanish MINECO project, co-funded by the European Regional Development Fund (FEDER).
Acknowledgments
The authors thank Dr. Maria Olívia Mercadante-Simões (UNIMONTES - Brazil) for the collection and identification of the plant material. ARAM is grateful to the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES-Brazil) for the fellowship [CAPES-PrInt 88887.474270/2020-00), and SRS to the Program Interreg MAC2014–2020 (MACBIOPEST, MAC2/1.1a/289) for their financial support of this research.
Conflicts of Interest
The authors declare no conflict of interest.
References
- 2050: A third more mouths to feed. Available online: https://www.fao.org/news/story/en/item/35571/icode/ (accessed on 14 November 2022).
- Pandian, S.; Ramesh, M. Development of pesticide resistance in pests: A key challenge to the crop protection and environmental safety. Pesticides in Crop Production: Physiological and Biochemical Action, 1st ed.; Srivastava, P.K., Singh, V.P., Singh, A., Tripathi, D.K., Singh, S., Prasad, S.M., Chauhan, D.K., Eds.; Willey: New Jersey, NJ, USA, 2020; pp. 1–13. [Google Scholar]
- De Simone, N.; Pace, B.; Grieco, F.; Chimienti, M.; Tyibilika, V.; Santoro, V.; Capozzi, V.; Colelli, G.; Spano, G.; Russo, P. Botrytis cinerea and table grapes: a review of the main physical, chemical, and bio-based control treatments in post-harvest. Foods 2020, 9, 1138. [Google Scholar] [CrossRef]
- Rocha, A. J.; Soares, J. M. S.; Nascimento, F. S.; Santos, A. S.; Amorim, V. B. O.; Ferreira, C. F.; Haddad, F.; Santos-Serejo, J. A.; Amorim, E. P. Improvements in the resistance of the banana species to Fusarium Wilt: A systematic review of methods and perspectives. J. Fungi 2021, 7, 249. [Google Scholar] [CrossRef]
- Wenderoth, M.; Garganese, F.; Schmidt-Heydt, M.; Soukup, S. T.; Ippolito, A.; Sanzani, S. M.; Fischer, R. Alternariol as virulence and colonization factor of Alternaria alternata during plant infection. Mol. Microbiol. 2019, 112, 131–146. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, D. M. Q.; Martins, S. D. S.; Guterres, D. C.; Martins, M. D.; Araújo, L.; Guimaraes, L. M. S.; Alfenas, A. C.; Furtado, G. Q. Diversity, prevalence and phylogenetic positioning of Botrytis species in Brazil. Fungal Biol. 2020, 124, 940–957. [Google Scholar] [CrossRef] [PubMed]
- Würz, D. A.; Rufato, L.; Bogo, A.; Allebrandt, R.; Bem, B. P.; Marcon-Filho, J. L.; Brighenti, A. F.; Bonin, B. F. Effects of leaf removal on grape cluster architecture and control of Botrytis bunch rot in Sauvignon Blanc grapevines in Southern Brazil. Crop Prot. 2020, 131, 105079. [Google Scholar] [CrossRef]
- Maia, J. N.; Beger, G.; Pereira, W. V.; De Mio, L. L. M.; Duarte, H. S. S. Gray mold in strawberries in the Paraná state of Brazil is caused by Botrytis cinerea and its isolates exhibit multiple-fungicide resistance. Crop Prot. 2021, 140, 105415. [Google Scholar] [CrossRef]
- Chitolina, G. M.; Silva-Junior, G. J.; Feichtenberger, E.; Pereira, R. G.; Amorim, L. First report on quinone outside inhibitor resistance of Alternaria alternata causing Alternaria brown spot in tangerines in São Paulo, Brazil. Plant Health Prog. 2019, 20, 94. [Google Scholar] [CrossRef]
- FAOSTAT. Available online: https://www.fao.org/faostat/en/#rankings/countries_by_commodity (accessed on 14 November 2022).
- Deltour, P.; França, S. C.; Heyman, L.; Pereira, O. L.; Höfte, M. Comparative analysis of pathogenic and nonpathogenic Fusarium oxysporum populations associated with banana on a farm in Minas Gerais, Brazil. Plant Pathol. 2018, 67, 707–718. [Google Scholar] [CrossRef]
- Chitolina, G. M.; Silva-Junior, G. J.; Feichtenberger, E.; Pereira, R. G.; Amorin, L. Distribution of Alternaria alternata isolates with resistance to quinone outside inhibitor (QoI) fungicides in Brazilian orchards of tangerines and their hybrids. Crop Prot. 2021, 141, 105493. [Google Scholar] [CrossRef]
- CABI. Chrysodeixis chalcites datasheet; 2022 [cited 2022 August 22]. Database: Invasive Species Compendium [Internet]. Available online: https://www.cabi.org/isc/datasheet/13243.
- Del Pino, M.; Cabello, T.; Hernández-Suárez, E. Age-stage, two-sex life table of Chrysodeixis chalcites (Lepidoptera: Noctuidae) at constant temperatures on semi-synthetic diet. Environ. Entomol. 2020, 49, 777–788. [Google Scholar] [CrossRef]
- Çakmak, T.; Simón, O.; Kaydan, M. B.; Tange, D. A.; Rodríguez, A. M. G.; Díaz, A. P.-B.; Murillo, P. C.; Suárez, E. H. Effects of several UV-protective substances on the persistence of the insecticidal activity of the Alphabaculovirus of Chrysodeixis chalcites (ChchNPV-TF1) on banana (Musa acuminata, Musaceae, Colla) under laboratory and open field conditions. PLoS One 2021, 16, e0250217. [Google Scholar] [CrossRef] [PubMed]
- López-Cepero, J.J. Análisis de residuos y gestión integrada de plagas en platanera. Phytoma España 2015, 271, 48–52. [Google Scholar]
- Soares, J. R. S.; Da Silva, R. S.; Ramos, R. S.; Picanço, M. C. Distribution and invasion risk assessments of Chrysodeixis includens (Walker, [1858]) (Lepidoptera: Noctuidae) using CLIMEX. Int. J. Biometeorol. 2021, 65, 1137–1149. [Google Scholar] [CrossRef] [PubMed]
- Dionisio, M.A.; Calvo, F.J. Integrated Management of Chrysodeixis chalcites Esper (Lepidoptera: Noctuidae) based on Trichogramma achaeae releases in commercial banana crops in the Canary Islands. Agronomy 2022, 12, 2982. [Google Scholar] [CrossRef]
- Gikas, G. D.; Parlakidis, P.; Mavropoulos, T.; Vryzas, Z. Particularities of fungicides and factors affecting their fate and removal efficacy: A review. Sustainability 2022, 14, 4056. [Google Scholar] [CrossRef]
- Isman, M. B. Botanical insecticides in the twenty-first century–fulfilling their promise? Annu. Rev. Entomol. 2020, 65, 233–249. [Google Scholar] [CrossRef]
- Acheuk, F.; Basiouni, S.; Shehata, A. A.; Dick, K.; Hajri, H.; Lasram, S.; Yilmaz, M.; Emekci, M.; Tsiamis, G.; Spona-Friedl, M.; May-Simera, H.; Eisenreich, W.; Ntougias, S. Status and prospects of botanical biopesticides in Europe and Mediterranean countries. Biomolecules 2022, 12, 311. [Google Scholar] [CrossRef]
- Souto, A. L.; Sylvestre, M.; Tölke, E. D.; Tavares, J. F.; Barbosa-Filho, J. M.; Cebrián-Torrejón, G. Plant-derived pesticides as an alternative to pest management and sustainable agricultural production: prospects, applications and challenges. Molecules 2021, 26, 4835. [Google Scholar] [CrossRef]
- Tavares, W. R.; Barreto, M. C.; Seca, A. M. L. Aqueous and ethanolic plant extracts as bio-insecticides— Establishing a bridge between raw scientific data and practical reality. Plants 2021, 10, 920. [Google Scholar] [CrossRef]
- Filho, J. G. S.; Almeida, A. S.; Pinto-Zevallos, D.; Barreto, I. C.; Cavalcanti, S. C. H.; Nunes, R.; Teodoro, A. V.; Xavier, H. S.; Filho, J. M. B.; Guan, L.; Neves, A. L. A.; Duringer, J. M. From plant scent defense to biopesticide discovery: Evaluation of toxicity and acetylcholinesterase docking properties for Lamiaceae monoterpenes. Crop Prot. 2023, 164, 106126. [Google Scholar] [CrossRef]
- Bellonzi, T. K.; Dutra, F. V.; Souza, C. N.; Rezende, A. A.; Gasparino, E. C. ; Pollen types of Sapindaceae from Brazilian forest fragments: apertural variation. Acta Bot. Bras. 2020, 34, 327–341. [Google Scholar] [CrossRef]
- Sofidiya, M. O.; Jimoh, F. O.; Aliero, A. A.; Afolayan, A. J.; Odukoya, O. A.; Familoni, O. B. ; Evaluation of antioxidant and antibacterial properties of six Sapindaceae members. J. Med. Plan. Res. 2012, 6, 154–160. [Google Scholar]
- Macedo, M. C.; Scalon, S. P.; Sari, A. P. ; Biometria de frutos e sementes e germinação de Magonia pubescens St. Hil (Sapindaceae); Rev. Bras. Sementes 2009, 31, 202–211. [Google Scholar] [CrossRef]
- Rocha, J.D.; Carneiro, F.M.; Fernandes, A.S.; Morais, J.M.; Borges, L.L.; Chen-Chen, L.; de Almeida, L.M.; Bailão, E.F.L.C. Toxic potential of Cerrado plants on different organisms. Int. J. Mol. Sci. 2022, 23, 3413. [Google Scholar] [CrossRef] [PubMed]
- Mesquita, M. L.; Paula, J. E.; Pessoa, C.; Moraes, M. O.; Costa-Lotufo, L. V.; Gourgnet, R.; Michel, S.; Tillequin, F.; Espindola, L. ; Cytoxic activity of Brazilian Cerrado plants used in traditional medicine against cancer cell lines. J. Ethnopharmacol. 2009, 123, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Arruda, W.; Oliveira, G. M.; Silva, I. G. Toxicidade do extrato etanólico de Magonia pubescens sobre larvas de Aedes aegypti. Rev. Soc. Bras. Med. Trop. 2003, 36, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Mendes, J. M. Ação leishmanicida de extratos de plantas no desenvolvimento de promastigotas de Leishmania amazonensis e estudo do perfil metabólico utilizando a técnica de cromatografia líquida de alta eficiência (CLAE). Master’s thesis, Universidade Federal de Goiás, Goiania, 2006. [Google Scholar]
- Aguiar, M. M.; Santos, K. T.; Royo, V. A.; Fonseca, F. S.; Menezes, E. V.; Mercadante-Simões, M. O.; Oliveira, D. A.; Junior, A. F. J. Antioxidant activity, total flavonoids and volatile constituents of Magonia pubescens A. St.-Hil. Med. Plants Res. 2015, 9, 1089–1097. [Google Scholar]
- Moraes, A. R. A.; Camargo, K. C.; Simões, M. O. M.; Ferraz, V. P.; Pereira, M. T.; Evangelista, F. C. G.; Sabino, A. P.; Duarte, L. P.; Alcântara, A. F. C.; Sousa, G. F. Chemical composition of Magonia pubescens essential oils and gamma-radiation effects on its constituents and cytotoxic activity in leukemia and breast cancer model. Chem. Biodiversity 2021, 18, e2100094. [Google Scholar] [CrossRef]
- Filho, J. C.; Coelho, M. F.; Albuquerque, M. C.; Azevedo, R. A. Seedling emergence of Magonia pubescens St. Hil. Sapindaceae a function of temperature. Amaz. J. Agric. Environ. Sci. 2011, 54, 137–143. [Google Scholar]
- Silva, L. B.; Silva, J. C.; Pavan, B. E.; Pereira, F. F.; Maggioni, K.; Andrade, L. H.; Candido, A. C. S.; Peres, M. T. L. P. Insecticide irritability of plant extracts against Sitophilus zeamais. Afr. J. Agric. Res. 2013, 8, 978–983. [Google Scholar]
- Souza, M. J.; Mercadante-Simões, M. O.; Ribeiro, L. M. Secondary-cell-wall release: a particular pattern of secretion in the mucilaginous seed coat of Magonia pubescens. Am. J. Bot. 2020, 107, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Kupchan, S.M. Recent advances in the chemistry of terpenoid tumor inhibitors. Pure Appl. Chem. 1970, 21, 227–246. [Google Scholar] [CrossRef] [PubMed]
- Cosoveanu, A.; Hernandez, M.; Iacomi-Vasilescu, B.; Zhang, X.; Shu, S.; Wang, M.; Cabrera, R. Fungi as endophytes in Chinese Artemisia spp.: Juxtaposed elements of phylogeny, diversity and bioactivity. Mycosphere 2016, 7, 102–117. [Google Scholar] [CrossRef]
- Fosbel-Plus. Available online: https://www.infoagro.com/agrovademecum/fito_m.asp?nreg=22421 (accessed on 14 November 2022).
- Liu, T.-X.; Zhang, Y. Side effects of two reduced-risk insecticides, indoxacarb and spinosad, on two species of Trichogramma (Hymenoptera: Trichogrammatidae) on cabbage. Ecotoxicol. 2012, 21, 2254–2263. [Google Scholar] [CrossRef]
- Bajonero, J. G.; Parra, J. R. P. Selection and suitability of an artificial diet for Tuta absoluta (Lepidoptera: Gelechiidae) based on physical and chemical characteristics. J. Insect Sci. 2017, 17, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Simón, O.; Bernal, A.; Williams, T.; Carnero, A.; Hernández-Suárez, E.; Muñoz, D.; Caballero, P. Efficacy of an alphabaculovirus-based biological insecticide for control of Chrysodeixis chalcites (Lepidoptera: Noctuidae) on tomato and banana crops. Pest Manag. Sci. 2015, 71, 1623–1630. [Google Scholar] [CrossRef] [PubMed]
- Escoubas, P.; Lajide, L.; Mitzutani, J. An improved leaf-disk bioassay and its application for the screening of Hokkaido plants. Entomol. Exp. Appl. 1993, 66, 99–107. [Google Scholar] [CrossRef]
- González-Coloma, A.; Terrero, D.; Perales, A.; Escoubas, P.; Fraga, B. M. Insect antifeedant ryanodane diterpenes from Persea indica. J. Agric. Food Chem. 1996, 44, 296–300. [Google Scholar] [CrossRef]
- Song, Z.; Li, X.; Xu, K.; Sun, G.; Yang, L.; Huang, L.; Liu, J.; Yin, P.; Huang, S.; Gao, F.; Zhou, X.; Chen, L. Design, synthesis and insecticidal activity and mechanism research of chasmanthinine derivatives. Sci. Rep. 2022, 12, 15290. [Google Scholar] [CrossRef]
- Schneider, C. A.; Rasband, W. S.; Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
Figure 1.
Antifungal effects (% growth inhibition, % GI) of the ethanolic extract (EE) and fractions (F-H/D: hexane/dichloromethane, and F-B: n-butanol) from M. pubescens roots against A. alternata. Fosbel-Plus was used as a positive control. Results are expressed as percentage relative to the negative control. Data are presented as meas ± SD (Standard Deviation, n = 8). F-E (ethyl acetate) and F-W (water) fractions were inactive (GI < 10%). Extract/fractions with a GI > than 20% at 1 mg/mL were assayed at lower concentrations (0.5 and 0.1 mg/mL).
Figure 1.
Antifungal effects (% growth inhibition, % GI) of the ethanolic extract (EE) and fractions (F-H/D: hexane/dichloromethane, and F-B: n-butanol) from M. pubescens roots against A. alternata. Fosbel-Plus was used as a positive control. Results are expressed as percentage relative to the negative control. Data are presented as meas ± SD (Standard Deviation, n = 8). F-E (ethyl acetate) and F-W (water) fractions were inactive (GI < 10%). Extract/fractions with a GI > than 20% at 1 mg/mL were assayed at lower concentrations (0.5 and 0.1 mg/mL).
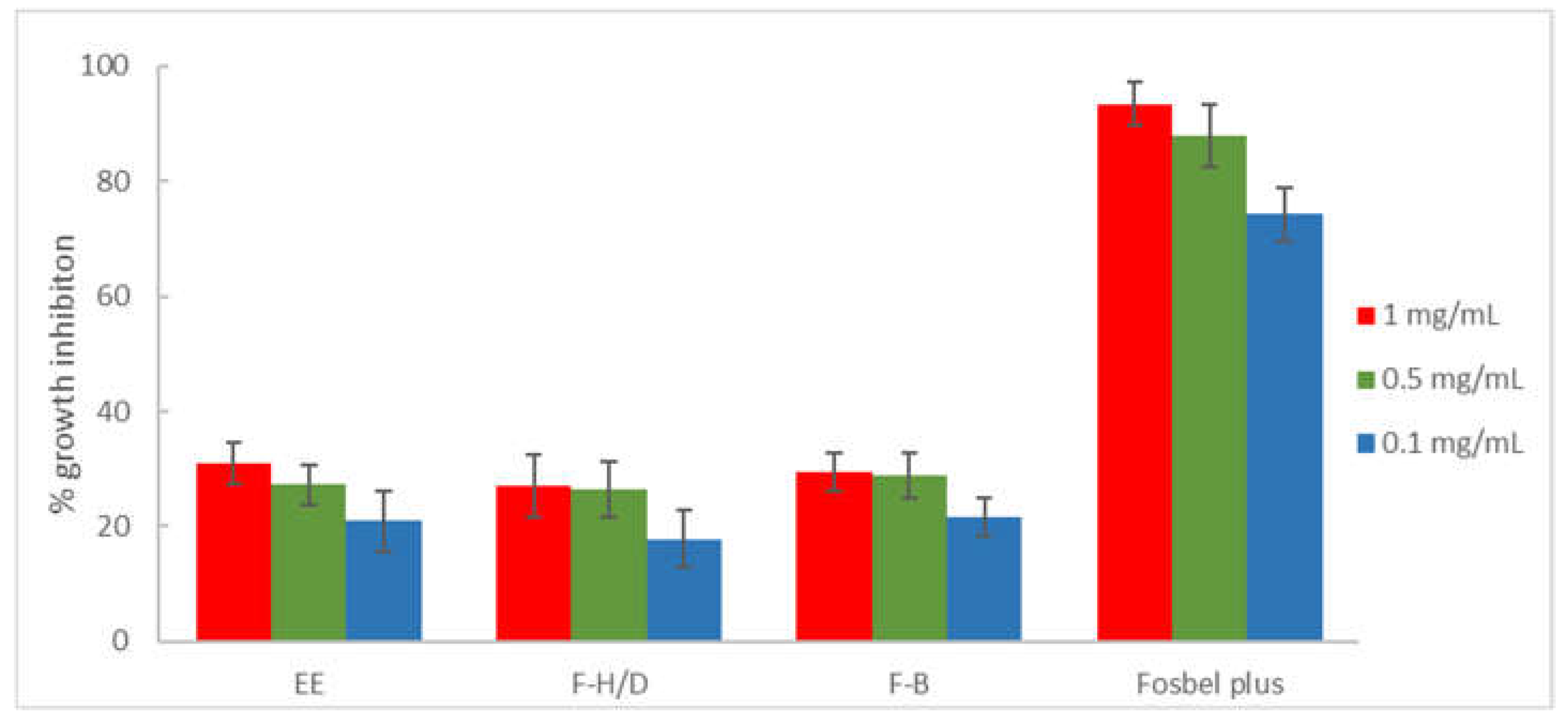
Figure 2.
Antifungal effects (% growth inhibition, % GI) of the ethanolic extract (EE) and fractions (F-H/D: hexane/dichloromethane, F-E: ethyl acetate, and F-B: n-butanol) from M. pubescens roots against B. cinerea. Fosbel-Plus was used as a positive control. Results are expressed as percentage relative to the negative control. Data are presented as means ± SD (Standard Deviation, n = 8). F-W (water) fraction was inactive (GI < 10%). Extract/fractions with a GI > than 20% at 1 mg/mL were assayed at lower concentrations (0.5 and 0.1 mg/mL).
Figure 2.
Antifungal effects (% growth inhibition, % GI) of the ethanolic extract (EE) and fractions (F-H/D: hexane/dichloromethane, F-E: ethyl acetate, and F-B: n-butanol) from M. pubescens roots against B. cinerea. Fosbel-Plus was used as a positive control. Results are expressed as percentage relative to the negative control. Data are presented as means ± SD (Standard Deviation, n = 8). F-W (water) fraction was inactive (GI < 10%). Extract/fractions with a GI > than 20% at 1 mg/mL were assayed at lower concentrations (0.5 and 0.1 mg/mL).
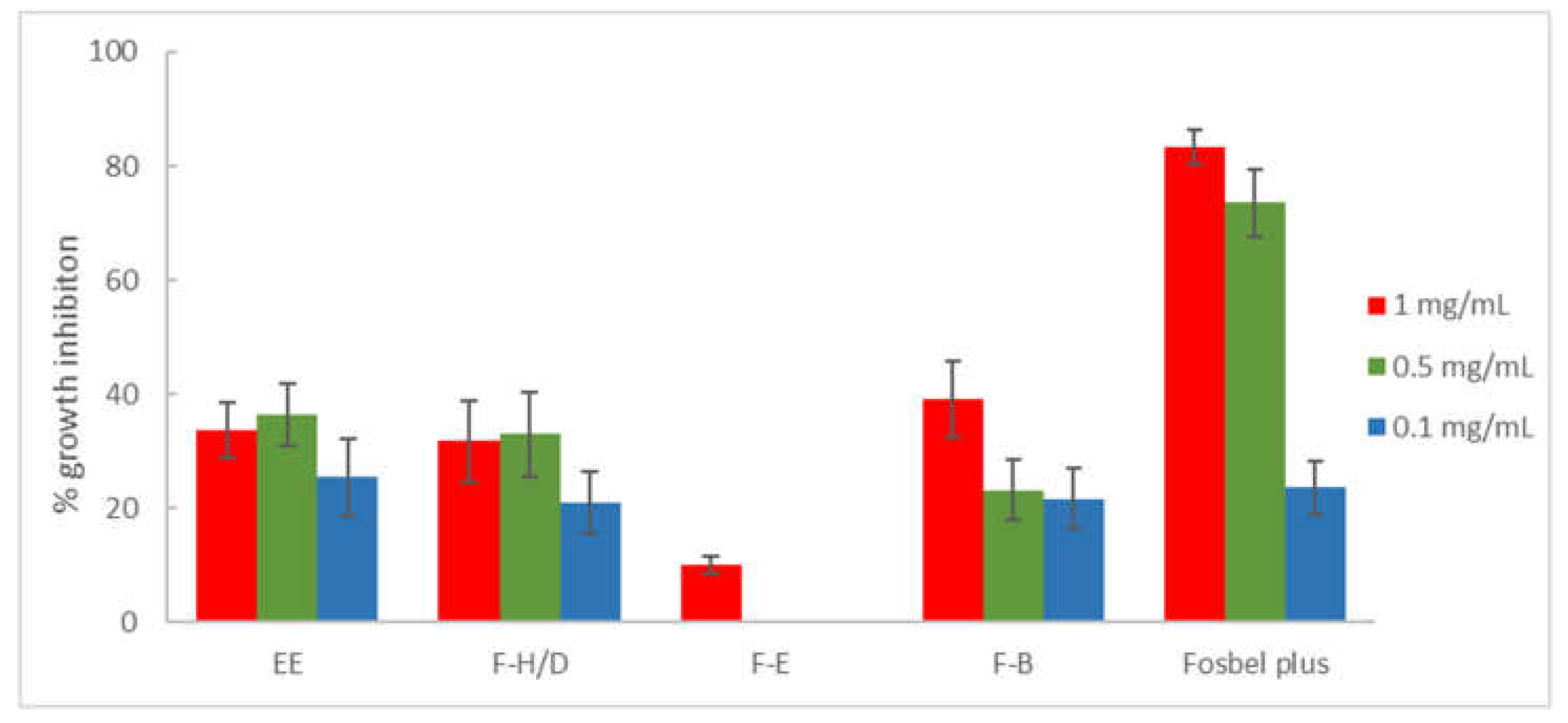
Figure 3.
Antifungal effects (% growth inhibition) of the ethanolic extract (EE) and fractions (F-H/D: hexane/dichloromethane, F-E: ethyl acetate fraction, and F-B: n-butanol) from M. pubescens roots against F. oxysporum. Results are expressed as percentage relative to the negative control. Fosbel-Plus was used as a positive control. Data are presented as means ± SD (Standard Deviation, n = 8). F-W (water) fraction was inactive (GI < 10%). Extract/fractions with a GI > than 20% at 1 mg/mL were assayed at lower concentrations (0.5 and 0.1 mg/mL).
Figure 3.
Antifungal effects (% growth inhibition) of the ethanolic extract (EE) and fractions (F-H/D: hexane/dichloromethane, F-E: ethyl acetate fraction, and F-B: n-butanol) from M. pubescens roots against F. oxysporum. Results are expressed as percentage relative to the negative control. Fosbel-Plus was used as a positive control. Data are presented as means ± SD (Standard Deviation, n = 8). F-W (water) fraction was inactive (GI < 10%). Extract/fractions with a GI > than 20% at 1 mg/mL were assayed at lower concentrations (0.5 and 0.1 mg/mL).
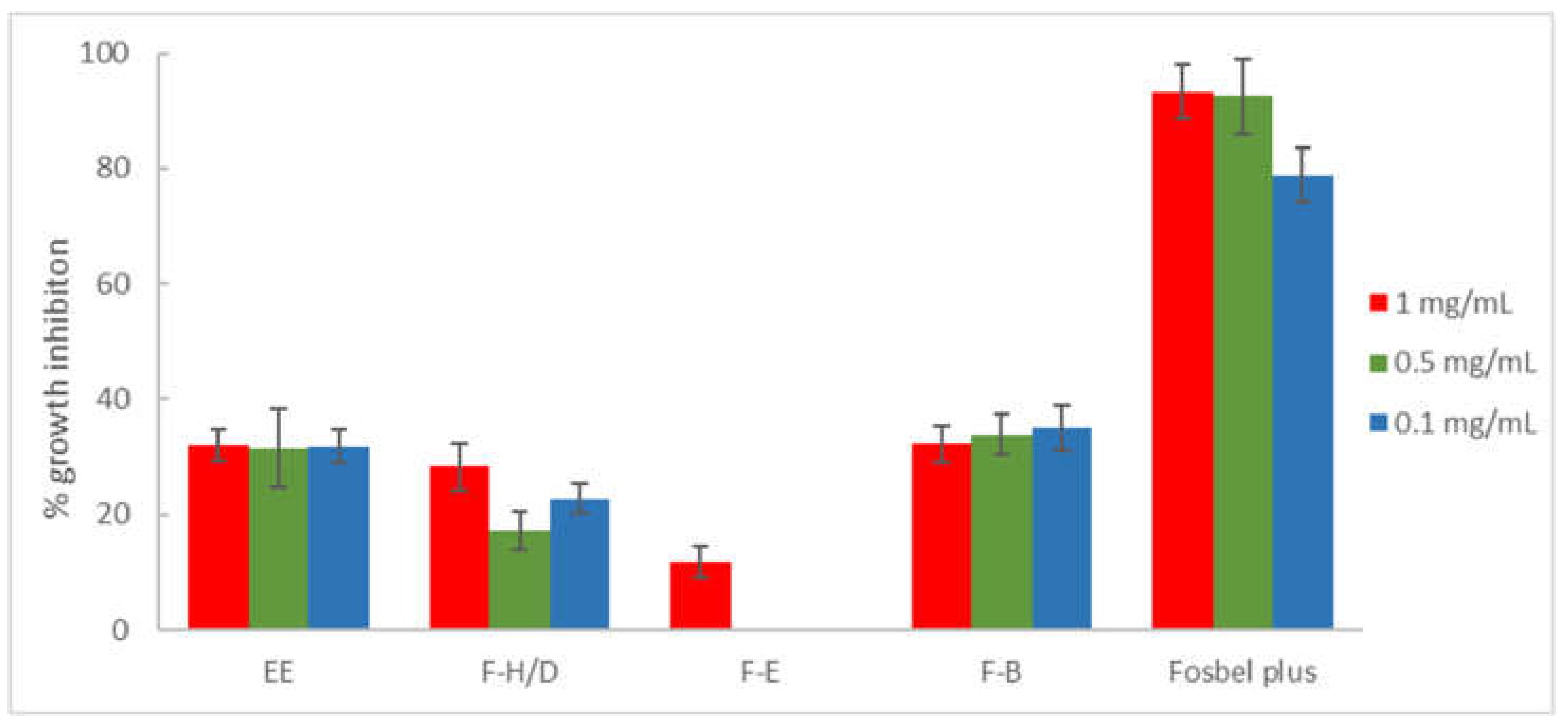

Table 1.
Insect antifeeding activity (% FR)1 of the ethanolic extract and fractions from Magonia pubescens roots against Chrysodeixis chalcites. .
Table 1.
Insect antifeeding activity (% FR)1 of the ethanolic extract and fractions from Magonia pubescens roots against Chrysodeixis chalcites. .
| Sample2 | Choice Feeding assay | Non-choice Feeding assay | Antifeedant category |
|---|---|---|---|
| EE3 | 92.14 | 95.16 | strong insect antifeedant |
| F-H/D3 | 60.52 | -27.67 | weak insect antifeedant |
| F-E3 | -108.09 | 61.10 | appetite suppressor |
| F-B | -6.69 | 60.71 | appetite suppressor |
| F-W3 | -74.57 | 44.73 | inactive |
| C4 | 97.95 | 99.04 | strong insect antifeedant |
1 Values are expressed as refusal rate (FR) = [1 - (% Treated disks consumed / % Control disks consumed)] x 100. 2 Samples were dissolved in ethanol: 5 µL per disk–0.2 mg/cm2. 3 EE: ethanolic extract, F-H/D: hexane-dichloromethane fraction, F-E ethyl acetate fraction, FB: n-butanol fraction, F-W: water fraction. 4 C: Indoxacarb 30% (STEWARD® 30WG) was used as a positive control.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



