Submitted:
01 February 2026
Posted:
03 February 2026
You are already at the latest version
Abstract
The most widely held theory for the origin of bird migration states that temperate zone resident birds evolved over thousands of years the necessary adaptations to allow successful movement in winter to less seasonal regions (e.g., the tropics). A problem for this theory is the fact nearly half of species that migrate to the tropics have breeding populations there as well, indicative of tropical, rather than temperate, origin. Winger et al. (2014a,b) have proposed a mechanism explaining how temperate zone species might have invaded the tropics as migrants and remained as resident breeders ("Migration Drop-Off"). The methods used to reach these conclusions involve no ecological arguments; rather they use extant species' ranges along with phylogenetic data to construct a model for inferring the biogeographic history of migratory lineages. The fundamental error in Winger et al. lies in the assumption that a species’ range is a conservative genetic attribute evolved over millennia. Data from field studies and the crowd-source site, eBird, demonstrate that this assumption is incorrect. Ranges for humdreds of bird species have changed by hundreds of kilometers in a matter of years, evidently as a result of dispersal and changes in seasonal food availability (Rappole et al. 2011, Rappole 2013: Chapter 8). In addition, breeding, wintering, and residency ranges have appeared, disappeared, and/or fundamentally shifted for tens of thousands of years during the past 2.58 million years of the Quaternary for hundreds of species. These facts taken together with the ecological issues involved with a species entering a complex ecosystem and creating a niche for itself argue against a "migration drop-off" explanation. Dispersal provides a more likely hypothesis for the origins of migration.
Keywords:
migratory syndrome
; migration drop-off
; northern home theory
; migratory bird phylogeny
; migratory bird ranges
; climate change
; dispersal
; weather and migration
; Emberizoidea
; Thraupidae
Introduction
The most widely held theory for the origin of migration, at present and historically, is that the behavior evolves gradually over millennia as natural selection favors successful movement from a highly seasonal breeding area (e.g, temperate regions in summer) to a less seasonal wintering area (e.g., the tropics) in fall and return to the breeding area in spring (Newton 2024:474); a theory often titled the “Northern Home” or “Migratory Syndrome” hypothesis. The chief problem for this theory is that nearly half of the world’s long distance avian migrants that breed in temperate areas and winter in the tropics also have populations that breed in the tropics (Rappole et al. 1983:44, Rappole 1995, 2013, 2022). This fact would appear to pose a quandary for Northern Home and Migratory Syndrome theories. How could hundreds of species that evolved in temperate, boreal, and arctic zone habitats invade and establish resident breeding populations in tropical habitats?
Winger et al. (2014a,b) have proposed a mechanism for how this dilemma might have been resolved in an Old World species ancestral to the large New World songbird group, Emberizoidea (sparrows, blackbirds, orioles, wood-warblers, tanagers, and their close relatives). They argue that members of this species entered North America from Northeast Asian populations that spread across the Behring land bridge during the mid-Miocene into the Nearctic temperate zone. They further hypotheisize that migration in this species evolved through natural selection caused by gradual seasonal deterioration in weather occurring over thousands of years, pushing members of this temperate zone resident population southward during the winter period, eventually reaching the tropics where some wintering members remained to breed (migration drop-off) followed by subsequent diversification into the hundreds of tropical resident species of the group, including the large, mostly tropical family of tanagers (Thraupidae).
In this paper, I examine the assumptions and methods used to develop this hypothesis, and consider its conclusions in light of extensive biogeographic and ecological data.
Methods Summary for Winger et al.
The methods used by Winger et al. (2014a,b) to reach their conclusions involve no ecological arguments or analyses. Instead, the authors use omly the current range (breeding and wintering) of emberizoids (Ridgely (2003), Poole (1992-2010), del Hoyo et al. (1992-2014). They use these data to construct crude summaries called “dominos” (because of their similarity to the game piece) (Figure 1). These summaries show presence or absence as breeding (column 1) or wintering (column 2) populations for all New World emberizoids in three regions: temperate North America (row 1); tropical Middle America and the Caribbean (row 2); or tropical South America (row 3).
The range summaries are used along with phylogenetic data to construct a model for inferring the biogeographic history of migratory lineages. The emberizoid phylogeny used in the model was constructed by Barker et al. (2015) using DNA evidence to trace back each lineage to the point at which speciation occurred (cladogenesis), beginning with the current 823 species of emberizoids, and following each branch back from there roughly 14 million years to the assumed first New World colonist.
In essence, the model is a decision matrix in which the decisions are referred to as “transitions”. These “transitions” involve the change for the daughter species from the range of the ancestral species to the new range of the derivative species. The authors provide several “rules” controlling these transitions (Table 1). These “rules” allow them to conclude what the range of the ancestor was from the ranges of the two daughter species, which in turn provides the basis for their models and statistical treatments.
Using these “rules”, along with the current ranges of all New World emberizoids, i.e., all of the currently existent daughter species, the authors reconstruct the ranges of all species ancestral to these living daughter species all the way back to the first ancestor, about 14 million years before present (MyBP), at least at the level of their occupancy of the three defined regions. The authors use these reconstructions to determine whether or not ranges of daughter species tended to shrink (occupy fewer regions) or split (occupy different parts of the same region) when compared with ancestors. To provide a metric of this change, they assigned a “range separation” value to each emberizoid species throughout the entire phylogenetic tree, the current ranges being used along with the rules to derive the ancestral ranges all the way back to the first ancestor. This “range separation” metric was calculated as the number of different regions occupied in the breeding season but not in winter plus the number of regions occupied in the winter but not in the breeding season plus the number of regions separating each. Thus highest “range separation” would be a value of 4, which would occur for a species that breeds only in temperate North America (+1) and winters only in tropical South America (+1), with both breeding and wintering range being separated by a region (Middle America and the Caribbean) (+2). Lowest range separation value (0) would be for a species that breeds and winters in the same region. Each emberizoid species in the entire lineage dating back to the first colonist is thus assigned a range separation value. This value is compared with that of its putative ancestor by subtracting the range separation value for a daughter from the range separation value for an ancestor in order to calculate whether ranges in emberizoids tend to shrink or remain similar from ancestor to daughter. For instance, if an ancestral species breeds in temperate North America and winters in tropical South America (range separation value of 4) has a daughter species whose breeding and wintering range is tropical South America (range separation value 0) then the difference is 4. Thus, transitions with a positive value show decrease in range from ancestor to daughter while transitions with a negative value show increases.
As in the case of domino construction described above, determination of the range of the ancestor species is critical to this analysis. To address this issue the authors generated a “posterior probability distribution of ancestral states” using the following procedures: “For each posterior sample of µ and Q from the Markov Chain Monte Carlo (MCMC) runs, the conditional likelihoods of ancestral states given the observed dominos at descendant leaf nodes were computed for each internal node using the pruning algorithm of (17) [Felsenstein 1981]. These likelihoods were used to compute the maximum-likelihood set of ancestral states for µ and Q, first at the root node (i.e., the state with highest conditional likelihood), and then by a recursive preorder traversal of the phylogeny, selecting the state with highest likelihood given the domino at the start of the node’s ancestral (subtending) branch.” (Winger et al. 2014b:2.
In other words, in order to fdetermine the range of the ancestor, they looked at the range of the daughters and, based on a set of assumptions, calculated a probability as to which of the daughter ranges most resembled that of the ancestor’s, and assigned the range of that daughter to the ancestor. The assumptions used are not stated, but presumably they are the same “rules” as those used in the range transition analysis.
Summary of Results and Conclusions in Winger et al.
After performing the treatments described above on the range and ancestry data, Winger et al. then calculate the number of transitions in which the range separation value increased from ancestor to daughter or decreased. They find that in the vast majority the number decreased, i.e., daughter species tended to occupy fewer regions for breeding and wintering than did ancestors. Based on these and related analyses, the authors reach the following conclusions:
1. The first emberizoid colonist to the New World arrived via dispersal from east Asia across Beringia into the northern regions of temperate North America.
2. This first colonist was a temperate zone resident.
3. Over a long period (thousands of years), seasonal deterioration in weather pushed this first temperate zone resident southward during temperate winter, eventually resulting in the evolution of the suite of characters required for successful long-distance migration (the Migratory Syndrome hypothesis).
4. Some members of this long-distance migrant population wintered as far south as Middle America, the Caribbean, and South America.
5. Some of the wintering members of this population remained to breed in their wintering grounds, establishing resident populations in the tropics. This behavior is referred to as “migration drop-off” by the authors.
6. Subsequent diversification of the group in the tropics of Middle and South America occurred in three ways: fragmentation of widespread ranges; expansion of resident ranges of nonmigratory populations from temperate to tropical latitudes; and expansion of a migratory species’ breeding range into a tropical area already occupied by their winter range (migration drop-off).
7. They emphasize that the most important of these sources involved early evolution of migration in the first colonist for temperate North America followed by migration drop-off in the tropics, resulting in tropical diversification. The authors point to the tanager family, Thraupidae, as the premier example of this process in action. According to Barker et al. (2015), this family arose roughly 13 MyBP and is restricted in distribution entirely to the New World tropics of Middle and South America, at least as it is currently recognized by the authors. This family is the largest group of New World emberizoids. It consists of 374 species. They are pan-tropical in the New World, but their greatest diversity occurs in the northern Andes. Most of the other emberizoid families also have their centers of diversity located in the tropics. As hypothesized for Thraupidae, the authors propose that these families also arose through migration drop-off of temperate zone emberizoids followed by in situ diversification in the tropics.
8. Long distance migration between North America and the Neotropics has evolved many times in this group, principally if not entirely due to shifts of winter ranges from North America into the tropics as opposed to breeding-range shifts from tropical latitudes into temperate regions.
9. Shifts of the winter range imposed by seasonal climate deterioration are the main drivers of the evolution of long-distance migration throughout Emberizoidea and, probably, most other migratory species as well.
10. Long-distance migration is the primary method through which temperate zone emberizoids established resident breeding populations in the tropics. This occurred through “migration drop-off”, i.e., when wintering birds of temperate breeding populations remained on the wintering ground to breed. Subsequent diversification within the tropics occurred via the normal processes of fragmentation of widespread ranges.
11. Expansion of resident ranges of nonmigratory populations from temperate to tropical latitudes also occurred in some instances, but at a much lower frequency than migration drop-off.
Discussion
The validity of the Winger et al. hypothesis for the origin and evolution of migration rests on a set of assumptions that allow the authors to reconstruct the range of each ancestor in the entire emberizoid lineage (Ree et al. 2005). The authors refer to these assumptions as “rules”, specifically, “the rules governing transitions between dominos”, which means the transition from the range of the ancestor to the ranges of the daughters. The reason these assumptions are referred to as “rules” is presumably because the logic of the analysis requires that they always be correct. A few deviations from the “rules” derails the line of reasoning that leads back to the range of the first ancestor.
All of these “rules” (Table 1) involve assumptions concerning the breeding and wintering ranges of migrants that derive from a single fundamental assumption, namely that the total range of a species is an evolved, genetic attribute.
Data on actual range change in migrants raise questions as to the validity of this fundamental assumption as well as those derived from it. Some of the most problematic ones are discussed below:
“Rule” 4. Breeding range expansion for a daughter species into a region south of the ancestral breeding range must be accompanied by comparable expansion of the winter range.
Hundreds of exceptions to this “rule” are known to occur. Many species of birds that are resident in the South American, Asian, and African tropics appear to have given rise to closely-related migratory populations that breed in temperate regions south of the tropical breeding and wintering areas of their presumptive ancestors, but winter north of their breeding areas in the tropics (McClure 1974, Jones 1996, Chesser and Levey 1998, Chesser 2005) including at least one species of the emberizoid family, Thraupidae, the Black-backed Tanager (Tangara peruviana) (Hilty 2011:229). By stating categorically that an apparently common “transition” cannot occur, the authors limit consideration of the evolution of migration to a specific set of assumed circumstances (i.e., a north temperate zone ancestor for every tropical daughter species), and in so doing, they limit understanding of how migration originates or evolves.
“Rule” 8. Neither breeding nor wintering range for a daughter species can jump a region as compared with the ancestral species, e.g., an ancestral breeding range in South America cannot give rise to a daughter species’ breeding range in temperate North America.
There is a significant body of evidence documenting the invalidity of this assumption. Closest relatives for many species of migrants from tropical South America to the temperate region of North America are tropical South or Middle American residents (Mayr 1946, Mayr and Short 1970, Rappole et al. 1983, Rappole 1995, 2013). For instance, the Red-eyed Vireo’s (Vireo olivaceus olivaceus) closest relative (considered by some authors to be a separate species) is the Chivi Vireo (Vireo olivaceus chivi), a resident or partial migrant of the Amazon basin. There are many similar examples from tropical Africa and Asia as well (Rappole 1995, Rappole and Jones 2002, Helbig 2003, Rappole 2005). One could argue, as the authors have, that all of the tropical relatives of long-distance migrants to the temperate zone are the result of migration drop-off. This argument seems highly unlikely given the abundant ecological and taxonomic evidence to the contrary (Rappole 2013:385-444).
“Rule” 9. There is, however, an exception to rule #8: an ancestral migratory species with a breeding range in temperate North America can have a daughter species that breeds in tropical South America.
This assumption is the “migration drop-off” rule from which, the authors believe, most South American emberizoids are derived, either directly or indirectly. Taken in combination with “Rule” 8, it allows the authors to reconstruct ancestral ranges entirely consistent with the migration drop-off hypothesis. However, if there are exceptions to “Rule” 8 in which tropical residents give rise to long-distance migrants to the temperate zone, “Rule” 9 is circular and invalid, and there are indeed many known exceptions (Rappole 2013:385-444). To take one well-documented example, “The Cattle Egret [Ardea ibis], which invaded South America from the Old World Tropics in the late 1800s (Meyer de Schauensee 1966), is a tropical resident throughout most of its range. First records for the Western Hemisphere are from Suriname in 1877 (Gladstone 1983). The first dispersing wanderers arrived in the United States around 1952 (Florida). Since then, members of the species have been recorded in most states of the United States as well as in many Canadian provinces. Northern populations of the species have established breeding colonies as far north as New York (Bull 1974). The populations of these northern-breeding egrets are Nearctic migrants, arriving in early summer to set up breeding colonies and leaving in autumn for their tropic winter quarters (Peterson 1980).” (Rappole 1995:114).
“Rule” 10. During speciation, the ancestral range can be inherited by daughter species or fragmented, but the entire ancestral range must be cumulatively present in the daughter species.
This assumption allows the authors to assume that the most complex range (i.e., combination of the two daughter ranges) is always the ancestral range. This assumption is absolutely critical to construction of ancestral ranges, especially the farther back you go in time along the lineage. Following this rule allows the authors to go all the way back to the first colonist. However, it can only be valid if “Rule” 11 is correct. If there are exceptions to “Rule” 11, then “Rule” 10 is circular and invalid, and, as documented below, there are many exceptions to “Rule” 11.
“Rule” 11. Migratory daughter species cannot be derived from a non-migratory ancestor.
This assumption is the key on which all of the other “rules”, and indeed the entire argument of the paper, hinges. If it is correct, then the key parts of the argument hold. If there are exceptions, then the entire methodology is circular and invalid. This assumption has to be true in order for the authors to perform the analyses that prove it is true.
As the authors note, there are several migration theorists who have argued that most temperate-zone migrants to the tropics derive from tropical zone residents. Perhaps arguments of these “southern home” theorists do not achieve the level of a “paradigm” as the authors claim, but certainly they have published a considerable amount of field data in support of their thesis (Dixon 1897, Mayr and Meise 1930, Williams 1958, Rappole et al. 1983, Cox 1985, Rappole and Tipton 1992, Levey and Stiles 1992, Rappole 1995, Safriel 1995, Rappole and Jones 2002, Helbig 2003, Rappole e al 2003, Rappole 2013:385-444). Convention and logical process require that Winger et al. answer these arguments before they dismiss them. Simply mentioning them in disparaging terms is not sufficient, especially when the procedures, models, and statistical treatments on which the conclusions in Winger et al. are based are circular without proof that no migratory daughter species has been derived from a non-migratory ancestor. In fact, the reality, as documented by the phylogenetic work of Helbig 2003, which documents the likely tropical ancestry for most migratory Phylloscopus species, and in numerous other studies cited in Rappole 2013, is quite the contrary.
The strength of the approach taken in Winger et al. is that, if the methods and reasoning are sound, then the conclusions are irrefutable, thereby providing an explanation for how the Northern Home and Migratory Syndrome theories for evolution of migration from a more seasonal environment to a less seasonal one might be made to fit the many observed violations of this situation for hundreds of species. Thus, they do not couch their conclusions in the usual cautious manner used in hypothetico-deductive reasoning, as in “our results indicate....” etc., etc. Instead, they state their conclusions more in the form of a mathematical proof ending with an implied QED, as in “...we found that long-distance migration primarily evolved through evolutionary shifts of geographic range south for the winter out of North America, as opposed to north for the summer.” (Winger et al. 2014a:1). However, the flaws in methodology and reasoning detailed above raise questions regarding the validity of these conclusions. Portions of their hypothesis could be correct, but the element of certainty is lost. Other explanations are possible for the colonization by emberizoids of the New World, their subsequent invasion and diversification in the tropics, and evolution of migration in the various members of the group.
Accordingly, I address the three major conclusions of Winger et al. below under the headings “Entry into the New World”; “Survival and Diversification in the New World Tropics”, and“Evolution of Migration.”
Entry into the New World. The authors assume that the first emberizoid colonists to the New World arrived via dispersal across Beringia about 14 MyBP. Thus this progenitor of New World Emberizoidea is assumed to be a north temperate zone resident. This assumption derives from the phylogenetic work of Barker et al. (2015), which presents the entire phylogeny of the New World Emberizoidea, and provides the assumed region of origin for each major branch of the phylogeneitc tree. At the base of the tree, the first New World member of Emberizoidea, from which all other emberizoids are assumed to have been derived, is shown as a resident of the north temperate zone of North America. However, Barker et al. (2015) do not know that this story of Old World temperate origin of New World emberizoids is true. They have no fossils documenting the presence of this proto emberizoid in Beringia or northern North America, and even if they did, one cannot tell from a fossil whether or not it is a resident of the temperate zone or a migrant that breeds in the temperate zone, winters in the tropics, and perhaps evolved in the tropics. Nevertheless, the assumption is logical. Beringia served as a connection between the Eurasian and North American land masses of varying width north to south for most of the past 100 million years (Cooper et al. 1992, Galdenkov et al. 2002, Sewall et al. 2007, Fiorillo 2008) (Figure 2). As such, it served as the main pathway for New World colonization by many different groups of animals and plants for nearly 60 million years (Pigg et al. 2004, Vila et al. 2011, Guo et al. 2012).
However, it is not the only possible pathway for invasion, at least for birds. In fact, the first emberizoid to colonize the New World could have flown there. And, if that was the mode of entry, then the most likely place of entry was tropical South America, since the very first branching of the phylogenetic tree constructed by Barker et al. (2015) produces South American daughter species. Indeed, over half of all emberizoids are South American residents.
But is such a founding flight possible? As Barker et al. (2015) note, there are other examples of long distance colonizations by emberizoids. Emberizoids colonized the Galapagos, 900 km from the nearest land, and emberizoids colonized Tristan da Cunha Island in the South Atlantic, 2,900 km from south Africa. Also, as if these historical examples were not enough, there are current examples. For instance, Brosset (1968) reported that a small population of two New World migrants, the Red-eyed Vireo (Vireo olicaceus) and the Summer Tanager (Piranga rubra) (an emberizoid), winter regularly in Gabon in equatorial Africa, 6,000 km outside their normal winter range (Figure 3). Clearly, these examples demonstrate that the first New World emberizoid could have entered tropical South America directly from tropical Africa rather than indirectly via Beringia.
The model for the likely method of mid-Miocene colonization of the New World by an emberizoid ancestor is provided by a similar avian colonization, namely that of the Galapagos Islands. Of the current 13 islands, seven are estimated to have existed for at least five million years (Geist et al. 2014). These islands are now inhabited by 18 indigenous species of emberizoid tanagers whose originating ancestor evidently was a relative of the modern genus Tiaris that arrived about three million years ago (Lamichhaney et al. 2015). Other indigenous species include four species of mockingbird (Mimus), a hawk (Buteo), a dove (Zenaida), a rail (Laterallus), a martin (Progne), and a flycatcher (Myiarchus) (Reaney et al. 2020). There are also four species presumably more recently arrived and as yet undifferentiated from their South American parent species, including Dark-billed Cuckoo (Coccyzus melacoryphus), Common Gallinule (Gallinula galeata), Paint-billed Crake (Mustelirallus erythrops), and Vermilion Flycatcher (Pyrocephalus rubinus).
Survival and Diversification in the New World Tropics Whether it was migration drop-off or dispersal from the New World temperate zone or Old World tropics, the new arrival in tropical South America would confront the same problem presented to any bird attempting to enter into a new environment, namely, “Moreau’s paradox” (Moreau 1952, 1972). How can an individual whose niche supposedly evolved in the temperate zone survive, and, in the case of a migration drop-off scenario, breed and diversify in a complex, highly evolved avian community like that which likely existed in South America 13-14 MyBP? Previous migration theorists recognized that this ecological question had to be explained, and so far, only three rationales have been provided: 1) superabundant resources (Moreau 1952, 1972, Morel and Bourlière 1962, Willis 1966, Leck 1972, MacArthur 1972, Karr 1976), 2) available empty niches (Moreau 1952, 1972) , or 3) seasonally returning migrants of tropical origin (Williams 1958, Rappole et al. 1983:44, Safriel 1995, Helbig 2003, Rappole 2013). Since the first emberizoid colonists cannot be returning to their origin, the third possibility is denied it, and since it stays to breed, excess resources also are not a likely explanation. Therefore the second explanation seems most likely, i.e., the first colonist was able to fill an empty niche, as in the case described above for the Galapagos. There had to be a new environment available to this new colonist, otherwise highly evolved resident avifauna would have outcompeted them. It may be that Andean orogeny presented just such an environment (Orme 2007, Gutierrez et al. 2013), one to which lowland forest residents were not well-adapted to exploit, could explain how a newly arrived emberizoid ancesto might have been able to colonize South America 14 MyBP, whether they came from the Nearctic or the African Tropics. However, this hypothesis cannot explain how emberizoids from the Nearctic might have invaded Neotropical environments more recently, as both Winger et al. and Barker et al. suggest (through migration drop-off). So far as we know, there have been no vast areas of empty niches to invade for several million years.
So how did they do it? The temperate zone origin hypothesis of migration drop-off does not address this question. Winger et al. simply state that these temperate emberizoid colonists just did it, as proven by the transition maps.
Evolution of Migration. Winger et al. hypothesize that the first emberizoid species to enter the New World was a temperate zone resident derived from the Old World temperate region via Beringia roughly 14 MyBP. These temperate zone resident populations were subjected to increasingly severe weather seasonally and over millenia evolved a suite of characters that allowed some members of the population to temporarily escape the weather by migrating south for the winter, returning each summer to the New World temperate zone to breed. To summarize, they state, “long-distance migration [in emberizoids] primarily evolved though evoltionay shifts of geographic range south for the winter out of North America.”
As noted above, proof of the validity of the Winger et al. theory is derived entirely from extant ranges of emberizoid migrants and their phylogenetic history. However, Helbig (2003:8), whose work was also based on avian phylogenies in the Old World warbler genus, Phylloscopus, stated, “phylogenetic relationships should be poor predictors of whether a species is highly migratory or not.”
Winger et al. contrast their work to a theory of “southern home origin” in which birds that breed in the Nearctic and migrate to the Neotropics for the winter originated in the Neotropics. This approach tends to obscure the true differences between their theory and others. The real differences are the way in which the influences of resources and time are understood.
Winger et al. follow the Migratory Syndrome theory (a more recent form of the Northern Home theory) according to which movement is a necessity, i.e., lowered survival due to lack of resources drives evolution of a suite of characters (i.e., the “syndrome”) that allow migration. Responding to this necessity takes evolutionary time to develop. The birds can’t just leave when they need to as the resources decline. They have to wait generations for the proper adaptations to evolve.
This concept is quite different from the Dispersal Theory proposed by Rappole (2013, 2022, 2024) in which movement is an opportunity, and, because it is an opportunity that is always a possibility for any dispersing individual, all dispersing individuals of all species already possess the basic equipment to capitalize on the opportunity, e.g., the ability to fly, to fatten, and to find their way. According to this theory, migration is a form of dispersal driven by food availability. Andreas Helbig put the matter succinctly “Preadaptations related to habitat choice are the main prerequisites enabling a species to expand its range northward during an interglacial period.” (Helbig 2003:16).
Contrary to the claims of Winger et al., the Dispersal Theory does not necessarily envision a southern home origin. Rather, what the Dispersal theory proposes is that dispersal, often by young of the year, is random in terms of direction. The dispersers move away from competition with conspecifics in search of new areas with suitable habitat and available resources to colonize. If they find it, they settle. If the habitat is aseasonal, they stay. If it is seasonal, they stay until resources deteriorate, and then return to where they came from (Rappole et al. 2003). Thus, migration is predicted to develop from the less seasonal environment (e.g, the tropics), where the colonizing birds originated from a resident population, to a more seasonal environment (e.g., the temperate zone), where the colonists go to breed. In the Northern Hemisphere, this seasonal movement will more often be northward in spring toward areas where resources are seasonally superabundant and southward in fall. In the Southern Hemisphere, the pattern will be reversed (Chesser and Levey 1998, Chesser 2005). In places where the seasonal environment is a mountain, the movement will be upslope in spring and downslope in fall (Rappole and Schuchmann 2003); and in areas where rainfall rather than temperature is the driver of resource availability, the movement will follow seasonal rainfall patterns, which could be north, south, east or west of the less seasonal environment - as is the case in sub Saharan Africa and much of Australia (Jones 1996, Rappole and Jones 2002, Rappole 2005).
A further weakness of the Winger et al. theory of how migration evolves is its failure to explain the pattern of Pleistocene obliteration of temperate/boreal breeding ranges for hundreds of long-distance migrant species (Finlayson 2011) followed by reoccupation of those ranges at intervals of roughly 41 thousand years (during the first 1.6 million years of the Quaternary) to 100,000 years (during the most recent million years) (based on climate data from Köhler and van de Wal 2020), a phenomenon that has occurred at least 20 times. In addition, Pleistocene tropical ranges for most species likely underwent immense shifts as habitats responded to radical changes in rainfall distribution (Bradley and Diaz 2021).
These facts of paleogeography demonstrate the extraordinaty flexibility of the migrant range. Migration drop-off does not provide an explanation for how birds could re-occupy temperate zone ranges abandoned for tens of thousands of years or track abrupt changes in habitat distribution in their tropical ranges. The Dispersal Theory does, by proposing that dispersing individuals are always searching for suitable new environments, regardless of time intervals of their absence. Helbig (2003:8) summarized this theory quite well based on his study of the phylogenitic work with phylloscopid warblers, “Highly migratory taxa probably originated repeatedly and independently from less migratory or sedentary populations living in climatically stable areas where no long-range movements were required.”
The Dispersal Theory for the origin of migration is hypothetical. Nevertheless, the argument is well-supported by ecological data on migrants from throughout the world (Rappole 2013) as well as phylogenetic data on Old World warblers (Helbig 2003). The same cannot be said for the Winger et al. hypothesis, which rests solely on a specious statistical treatment of phylogenetic data and false assumptions concerning the nature of range change.
Conclusions
The abstract for the article on origin and evolution of avian migration by Winger et al. (2014a) states, “The complicated geography of seasonal migration has long posed a challenge for inferring the geographic origins of migratory species as well as evolutionary sequences of change in migratory behavior. To address this challenge, we developed a phylogenetic model of the joint evolution of breeding and nonbreeding (winter) ranges and applied it to the inference of biogeographic history in the emberizoid passerine birds.”
This statement presents the fundamental error in the Winger et al. migration evolution hypothesis, namely the assumption that a species’ range is a conservative genetic attribute evolved over millennia. Data from field studies in Texas (Rappole, Blacklock et al. 2007, Rappole, Pine et al. 2007, Rappole et al. 2011, Faridani et al. 2009, Rappole 2013, 2022, 2024), the crowd-source site, ebird (eBird - Discover a new world of birding...), and numerous other sources published over the past quarter century demonstrate that this is not the case; ranges for hundreds of bird species have changed by hundreds of kilometers in a matter of years, evidently as a result of dispersal and changes in seasonal food availability presumably related to climate change (Rappole, Pine et al. 2007, Rappole 2013:329, Rappole 2022:86, 141-143, 2024:311).
Certainly, aspects of the range are somehow a part of the species’ genetic make-up. Otherwise how would young born in the temperate breeding range be capable of navigating correctly to a tropical wintering range? Nevertheless, the ability for individuals to exploit changes in food distribution, dramatically changing their known breeding and wintering distribution; expanding, contracting, or ceasing migration altogether (Rappole, Pine et al. 2007, Becker et al. 2019) demonstrate that we have a great deal to learn with regard to how genes and environment interact to produce observed migration phenomena.
The strength of any hypothesis lies in its ability to explain observations. By this measure, as explained herein, the Northern Home theory and its relative, the Migratory Syndrome, for evolution of migration from more seasonal environments (e.g., temperate regions) to less seasonal ones (e.g., tropical regions) as influenced by gradual deterioration in winter weather are lacking, despite the ideas offered by Winger et al.
In contrast, the Dispersal Theory for the origin of migration deriving from the dispersal of individuals in search of suitable habitats in which they can survive and breed has significant power. Extensive data from a variety of sources show the extraordinary flexibility for the development and modification of migration in space and time, a flexibility that only dispersal can allow.
Acknowledgments
I thank Kevin Winker, Barbara Helm, and David King for review of the manuscript. I thank F. K. Barker for sending me a pre-publication version of his article in The Auk, and J. Klicka for copies of several relevant articles on related phylogenetic analyses.
References
- Barker, F. K.; Burns, K. J.; Klicka, J.; Lanyon, S. M.; Lovette, I. J. New insights into New World biogeography: an integrated view from the phylogeny of blackbirds, cardinals, sparrow, tanagers, warblers, and allies. Auk 2015, 132, 333–348. [Google Scholar] [CrossRef]
- Becker, D. J.; Schultz, E. M.; Atwell, J. W.; Ketterson, E. D. Urban residency and leucocyte profiles in a traditional migratory songbird. Animal Migration 2019, 6, 49–59. [Google Scholar] [CrossRef]
- Bradley, R. S.; Diaz, H. F. Late Quaternary abrupt climate change in the tropics and sub-tropics: The continental signal of tropical hydroclimatic events (THEs). Review of Geophysics 2021, 59, e2020RG000732. [Google Scholar] [CrossRef]
- Brosset, A. Ecological localization of migratory birds in the equatorial forest of Gabon [in French]. Alauda 1968, 52, 81–101. [Google Scholar]
- Bull, J. L. Birds of New York State; Doubleday: Garden City, New Jersey, 1974. [Google Scholar]
- Chesser, R. T. Seasonal distribution and ecology of South American austral migrant flycatchers. In Birds of two worlds; Greenberg, R., Marra, P., Eds.; Johns Hopkins University Press: Baltimore, 2005; pp. 168–181. [Google Scholar]
- Chesser, R. T.; Levey, D. J. Austral migrants and the evolution of migration in New World birds: Diet, habitat,, and migration revisited. American Naturalist 1998, 152, 311–319. [Google Scholar] [CrossRef]
- Cooper, A. K.; Scholl, D. W.; Marlow, M. S. Plate tectonic model for the evolution of the eastern Bering Sea basin. Geological Society of American Bulletin 1992, 87, 1119–1126. [Google Scholar] [CrossRef]
- Handbook of the Birds of the World; del Hoyo, J., Elliott, A., Christie, D., Eds.; Lynx Edicions: Barcelona, Spain, 1992-2014. [Google Scholar]
- Dixon, C. The migration of birds; Horace Cox, Windsor House, London, 1897. [Google Scholar]
- Faridani, S.; Lee, B.; Glasscock, S.; Rappole, J.; Song, D.; Goldberg, K. A networked telerobotic observatory for collaborative remote observation of avian activity and range change. In Proceedings of the International Robotics Conference, 2009. [Google Scholar]
- Felsenstein, J. Evolutionary trees from gene frequencies and quantitative characters: finding maximum likelihood estimates. Evolution 1981, 35, 1229–1242. [Google Scholar] [CrossRef] [PubMed]
- Finlayson, C. Avian Survivors: The History and Biogeography of Palearctic Birds; T & A. D. Poyser, London, 2011. [Google Scholar]
- Fiorillo, A. R.; Blodgett, R. B.; Stanley, G. D. Dinosaurs of Alaska: implications for the Cretaceous origin of Beringia. Pp. 313-326 In The terrane puzzle: new perspectives on paleontology and stratigraphy from the North American Cordillera; Geological Society of America, Special Paper # 442, 2008. [Google Scholar]
- Geist, D. J.; Snell, Howard; Snell, Heidi; Goddard, C.; Kurz, M. D. A Paleogeographic Model of the Galápagos Islands and Biogeographical and Evolutionary Implications. Chapter 8, Pp. 145-166 In The Galápagos: A Natural Laboratory for the Earth Sciences; Harpp, K. S., Mittelstaedt, E., d’Ozouville, N., Graham, D. W., Eds.; American Geophysical Union, Geophysical Monograph 204, John Wiley & Sons: New York, 2014. [Google Scholar]
- Gladenkov, A. Y.; Oleinik, A. E.; Marincovich, L., Jr.; Barinov, K. B. A refined age for the earliest opening of the Bering Strait. Palaeogeography, Palaeoclimatology, Palaeoecology 2002, 183, 321–328. [Google Scholar] [CrossRef]
- Gladstone, D. E. Bubulcus ibis (Garcilla Bueyera, Cattle Egret). Pp. 550-551 In Costa Rican Natural History; Janzen, D., Ed.; University of Chicago Press: Chicago, 1983. [Google Scholar]
- Guo, P.; Liu, Q.; Xu, Y.; Jiang, K.; Hou, M.; Ding, L.; Pyron, R. A.; Burbrink, F. T. Out of Asia: Natricine snakes support the Cenozoic Beringian dispersal hypothesis. Molecular Phylogenetics and Evolution 2012, 63, 825–833. [Google Scholar] [CrossRef]
- Gutierrez, N. M.; Hinjosa, L. F.; LeRoux, J. P.; Pedroza, V. Evidence for an early-middle Miocene age of the Navidad Formation (central Chile): Paleontological, paleoclimatic and tectonic implications. Andean Geology 2013, 40, 66–78. [Google Scholar]
- Helbig, A. J. Evolution of bird migration: A phylogenetic and biogeographic perspective. Pp. 3–21 in Avian migration; Berthold, P., Gwinner, E., Sonnenschein, E., Eds.; Springer: Heidelberg, 2003. [Google Scholar]
- Hilty, S. L. Family Thraupidae (tanagers). In Handbook of the birds of the world; del Hoyo, J., Elliott, A., Christie, D. A., Eds.; Lynx Edicions: Barcelona, 2011; Volume 16, pp. 46–329. [Google Scholar]
- Jones, P. Community dynamics of arboreal insectivorous birds in African savannas in relation to seasonal rainfall patterns and habitat change. In Dynamics of tropical communities; Newberry, D. M., Prins, H. H. T., Brown, N. D., Eds.; Blackwell, Oxford, 1996; pp. 421–447. [Google Scholar]
- Karr, J. R. On the relative abundance of migrants from the North Temperate Zone in tropical habitats. Wilson Bulletin 1976, 88, 433–458. [Google Scholar]
- Köhler, P.; van de Wal, R. S. W. Interglacials of the Quaternary defined by northern hemispheric land ice distribution outside of Greenland. Nature Communications 2020. [Google Scholar] [CrossRef] [PubMed]
- Lamichhaney, S.; Berglund, J.; Almén, M. Sällman; Maqbool, K.; Grabherr, M.; Martinez- Barrio, A.; Promerová, M.; Rubin, C.-J.; Wang, C.; Zamani, N.; Grant, B. R.; Grant, P. R.; Webster, M. T.; Andersson, L. Evolution of Darwin’s finches and their beaks revealed by genome sequencing. Nature 2015, 518, 371–375. [Google Scholar] [CrossRef] [PubMed]
- Leck, C. F. The impact of some North American migrants at fruiting trees in Panama. Auk 1972, 89, 842–850. [Google Scholar] [CrossRef]
- Levey, D. J.; Stiles, F. G. Evolutionary precursors to long-distance migration: Resource availability and movement patterns in neotropical landbirds. American Naturalist 1992, 140, 447–476. [Google Scholar]
- MacArthur, R. H. Geographical Ecology: Patterns in the Distribution of Species; Harper & Row: New York, 1972. [Google Scholar]
- Mayr, E. History of the North American bird fauna. Wilson Bulletin 1946, 58, 2–41. [Google Scholar]
- Mayr, E.; Meise, W. [Theories on the history of migration]. Vogelzug (in German). 1930, 1, 149–172. [Google Scholar]
- McClure, E. Migration and survival of the birds of Asia. U. S. Army Component, SEATO Medical Research Laboratory, Bangkok. The Place of Africa in the Palaearctic; 1974. [Google Scholar]
- Meyer de Schauensee, R. The Species of Birds of South America and their Distribution; Livingston: Wynnewood, Pennsylvania, 1966. [Google Scholar]
- Moreau, R. E. The place of Africa in the Palaearctic migration system. Animal Ecology 1952, 21, 250–271. [Google Scholar] [CrossRef]
- Moreau, R. E. The Palaearctic-African Bird Migration System; Academic Press: New York, 1972. [Google Scholar]
- Morel, G.; Bourlière, F. Ecological relations of the sedentary and migratory avifauna in a Sahel savannah of lower Senegal [in French]. Terre et Vie 1962, 4, 371–393. [Google Scholar]
- Newton, I. The Migration Ecology of Bird; Academic Press: New York, 2024. [Google Scholar]
- Orme, A. R. The tectonic framework of South America. In Physical Geography of South America; Veblen, T. T., Young, K. R., Orme, A. R., Eds.; Oxford University Press: Oxford, 2007; pp. 12–17. [Google Scholar]
- Peterson, R. T. A Field Guide to the Birds; Houghton Mifflin: Boston, 1980. [Google Scholar]
- Pigg, K. B.; Ickert-Bond, S. M.; Wen, amd J. Anatomically preserved Liquidambar (Altingiaceae) from the middle Miocene of Yakima Canyon, Washington state, USA, and its biogeographic implications. American Journal of Botany 2004, 91, 499–509. [Google Scholar] [CrossRef]
- The Birds of North America; Poole, A., Ed.; Cornell Laboratory of Ornithology: Ithaca, New York, 1992-2010. [Google Scholar]
- Rappole, J. H. The ecology of migrant birds: A Neotropical perspective; Smithsonian Institution Press: Washington, D.C, 1995. [Google Scholar]
- Rappole, J.H. Evolution of old and new world migration systems: a response to Bell. Ardea 2005, 93(1), 125–131. [Google Scholar]
- Rappole, J. H. The Avian Migrant; Columbia University Press: New York, 2013. [Google Scholar]
- Rappole, J. H. Bird Migration: A New Understanding; Johns Hopkins University Press: Baltimore, Maryland, 2022. [Google Scholar]
- Rappole, J. H. Migration Mysteries; Texas A&M University Press: College Station, Texas, 2024. [Google Scholar]
- Rappole, J. H.; Blacklock, G. W.; Norwine, J. Apparent rapid range change in South Texas birds: Response to climate change? In Texas climate 2100; Norwine, J., John, K., Eds.; Texas A&M University: Kingsville, Texas, 2007. [Google Scholar]
- Rappole, J. H.; Glasscock, S.; Goldberg, K.; Song, D.; Faridani, S. Range change among New World tropical and subtropical birds. Pp. In International Symposium on Tropical Ecology; Schuchmann, K., Ed.; Alexander Koenig Museum Monographs: Bonn, Germany, 2011; Volume 57, pp. 151–167. [Google Scholar]
- Rappole, J. H.; Helm, B.; Ramos, M. A. An integrative framework for understanding the origin and evolution of avian migration. Journal of Avian Biology 2003, 34, 124–128. [Google Scholar] [CrossRef]
- Rappole, J. H.; Jones, P. Evolution of Old and New World migration systems. Ardea 2002, 90, 525–537. [Google Scholar]
- Rappole, J. H.; Morton, E. S.; Lovejoy, T. E., III; Ruos, J. S. Nearctic avian migrants in the Neotropics; U.S. Fish and Wildlife Service: Washington, D.C, 1983. [Google Scholar]
- Rappole, J. H.; Pine, A.; Swanson, D.; Waggerman, G. Conservation and management For migratory birds: Insights from population data and theory for the White-winged Dove. In Proc. 25th Anniversary Symposium for the Caesar Kleberg Wildlife Research Institute; Fulbright, T., Hewitt, D., Eds.; CRC Press: Gainesville, Florida, 2007. [Google Scholar]
- Rappole, J. H.; Schuchmann, K.-L. The ecology and evolution of hummingbird population movements: A review. In Avian migration; Berthold, P., Gwinner, E., Sonnenschein, E., Eds.; Springer: Heidelberg, 2003; pp. 39–51. [Google Scholar]
- Rappole, J. H.; Tipton, A. R. The evolution of avian migration in the Neotropics. Ornitología Neotropical 1992, 3, 45–55. [Google Scholar]
- Reaney, A. M.; Bouchenak-Khelladi, Y.; Tobias, J. A.; Abzhanov, A. Ecological and morphological determinants of evolutionary diversification in Darwin’s finches and their relatives. In Ecology and Evolution; 2020. [Google Scholar] [CrossRef]
- Ree, R. H.; Moore, B. R.; Webb, C. O.; Donoghue, M. J. A likelihood framework for inferring the evolution of geographic range on phylogenetic trees. Evolution 2005, 59, 2299–2311. [Google Scholar] [CrossRef]
- Ridgely, R. S.; Allnutt, T. F.; Brooks, T.; McNicol, D. K.; Mehlman, D. W.; Young, B. E.; Zook, J. R. Digital Distribution Maps of the Birds of the Western Hemisphere. Version 1.0; NatureServe: Arlington, Virginia, USA, 2003; Available online: www.natureserve.org.
- Safriel, U. N. The evolution of Palearctic migration - the case for southern ancestry. Israel Journal of Zoology 1995, 41, 417–431. [Google Scholar]
- Sewall, J. O.; Van De Wal, R. S. W.; Van Der Zwan, K.; Van Oosterhout, C.; Dijkstra, H. A.; Scotese, C. R. Climate model boundary conditions for four Cretaceous time slices. Climate of the past, European Geosciences Union 2007, 3(4), 647–657, hal-00298095. [Google Scholar] [CrossRef]
- Vila, R.; Bell, C. D.; Macniven, R.; Goldman-Huertas, B.; Ree, R. H.; Marshall, C. R.; Bálint, Z.; Johnson, K.; Benyamini, D.; Pierce, N. Phylogeny and palaeoecology of Polyommatus blue butterflies show Beringia was a climate-regulated gateway to the New World. Proceedings of the Royal Society B: Biological Sciences 2011, 278, 2737–2744. [Google Scholar] [CrossRef] [PubMed]
- Williams, G. G. Evolutionary aspects of bird migration. In Lida Scott Brown Lectures in Ornithology; University of California, Los Angeles, 1958; pp. 53–85. [Google Scholar]
- Willis, E. O. The role of migrant birds at swarms of army ants. Living Bird 1966, 5, 187–231. [Google Scholar]
- 2014a. Temperate origins of long-distance seasonal migration in New World songbirds. Proceedings of the National Academy of Sciences www.pnas.org/cgo/doi/10.1073/pnas.1405000111:1-6.
- Winger, B. M.; Barker, F. K.; Ree, R. H. 2014b. Supporting information for “Temperate origins of long-distance seasonal migration in New World songbirds”. Proceedings of the National Academy of Sciences www.pnas.org/cgo/content/short/1405000111:1-9.
Figure 1.
Range summary “domino” for a neotropical migrant, e.g., the Blackpoll Warbler (Setophaga striata), that breeds (gray circle) in temperate (or boreal) North America and winters (black circle) in tropical South America.
Figure 1.
Range summary “domino” for a neotropical migrant, e.g., the Blackpoll Warbler (Setophaga striata), that breeds (gray circle) in temperate (or boreal) North America and winters (black circle) in tropical South America.
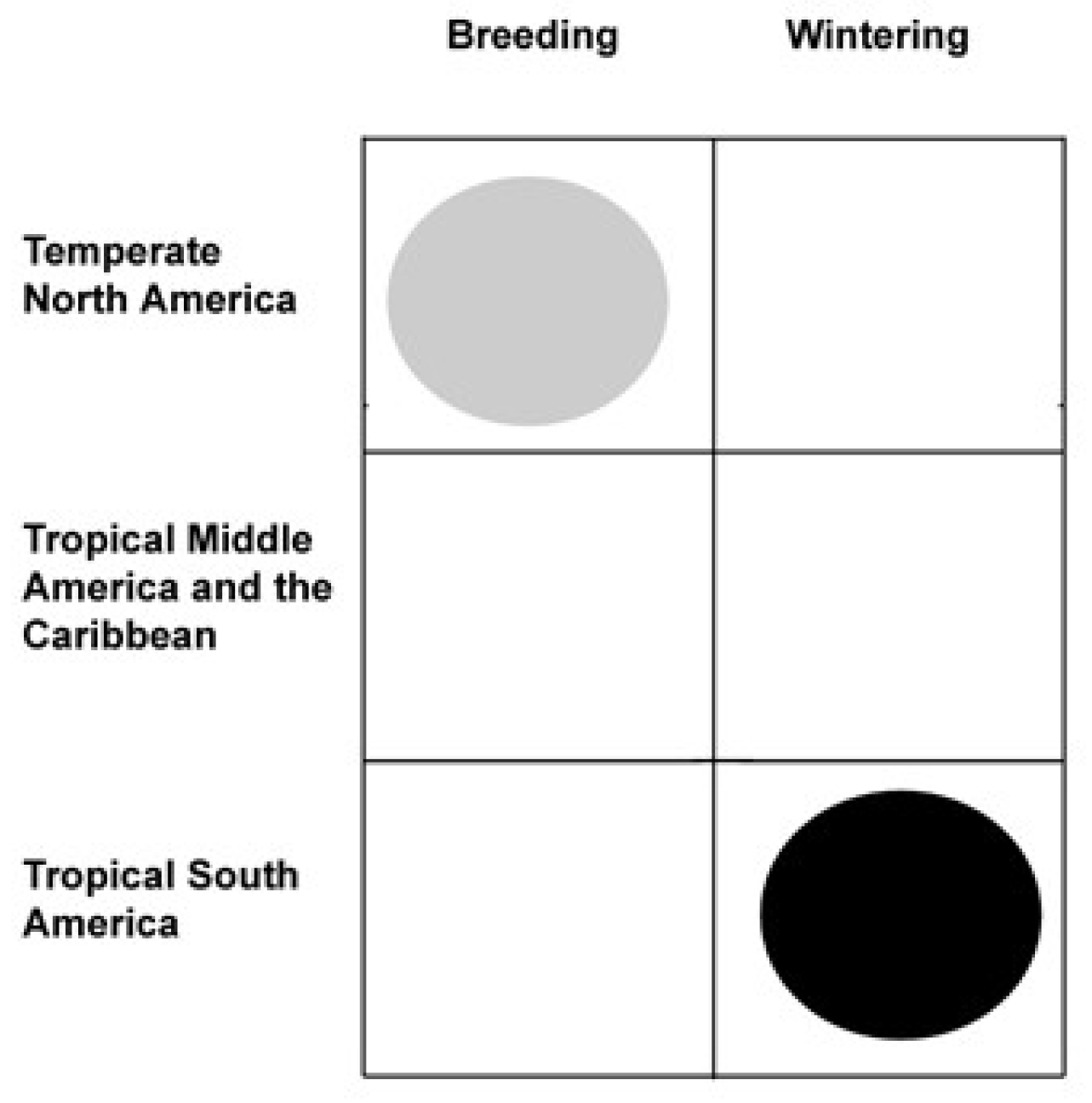
Figure 2.
Beringia (based on an online map from the National Park Service).
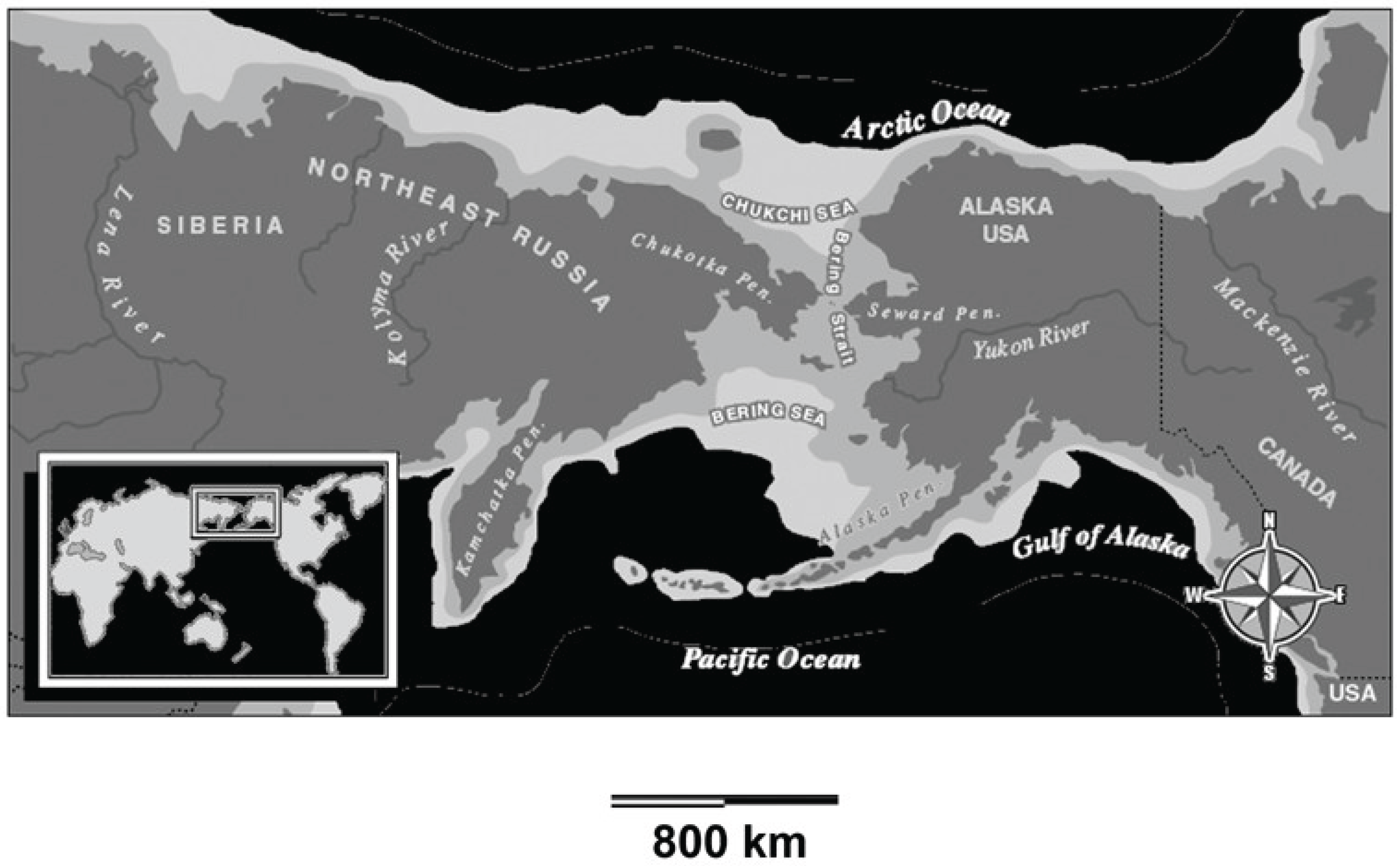
Figure 3.
World map showing the breeding (dark gray) and wintering (black) distribution of the Summer Tanager (Piranga rubra) (an emberizoid). Gabon, where a small population of this species winters, and Tristan da Cunha, an island colonized by emberizoids, are also shown. See text for discussion.
Figure 3.
World map showing the breeding (dark gray) and wintering (black) distribution of the Summer Tanager (Piranga rubra) (an emberizoid). Gabon, where a small population of this species winters, and Tristan da Cunha, an island colonized by emberizoids, are also shown. See text for discussion.
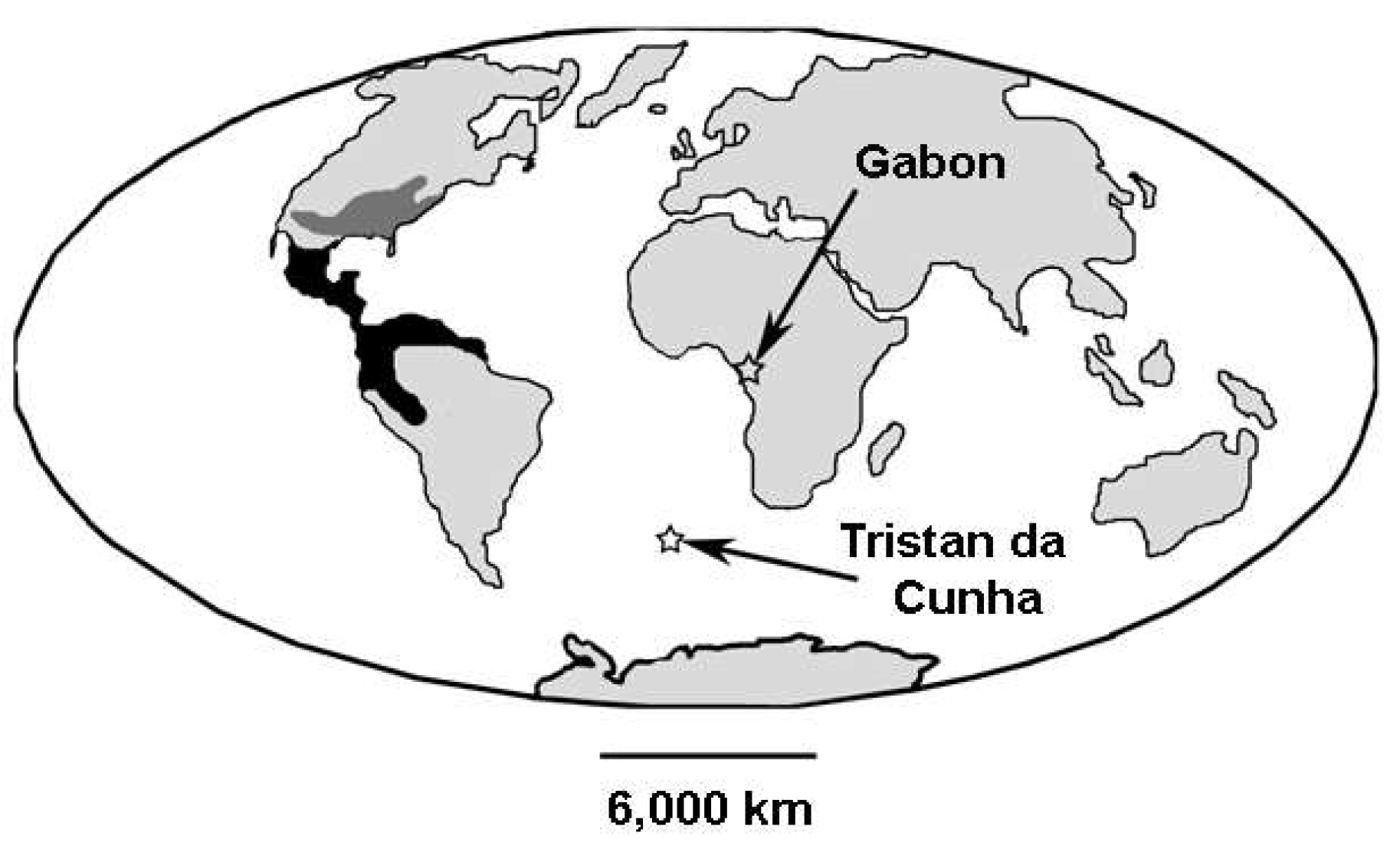

Table 1.
“Rules”* governing transitions from ancestral range to daughter ranges in the emberizoid lineage (Winger et al. 2014b:1).
Table 1.
“Rules”* governing transitions from ancestral range to daughter ranges in the emberizoid lineage (Winger et al. 2014b:1).
| 1 | Both breeding and winter range [of the two daughter species] can show expansion or contraction separately or in unison into neighboring regions [i.e., the three regions defined in Figure 1 - temperate North America; Middle America and the Caribbean; and tropical South America] relative to the range of their immediate ancestor. |
| 2 | The new breeding range [of the two daughter species] cannot expand or contract to result in breeding-range occupancy into regions south of the winter range [of the immediate ancestor]. |
| 3 | The new winter range [of a daughter species] cannot occupy a region north of the breeding range [of the ancestor]. |
| 4 | Breeding range expansion [for a daughter species ] into a region south of the [ancestral] breeding range must be accompanied by comparable expansion of the winter range. |
| 5 | Winter range expansion [for a daughter species] into a region north of the [ancestral] winter range must be accompanied by a comparable expansion of the breeding range. |
| 6 | Breeding range [for a daughter species] can expand into, or contract from, a region north of the [ancestral] breeding range without change in the winter range. |
| 7 | Winter range [for a daughter species] can expand into, or contract from, a region south of the [ancestral] breeding range without change in the breeding range [of the daughter species]. |
| 8 | Neither breeding nor wintering range [for a daughter species] can jump a region [as compared with the ancestral species], e.g., an ancestral breeding range in South America cannot give rise to a daughter species’ breeding range in temperate North America. |
| 9 | There is, however, an exception to rule #8: an ancestral migratory species with a breeding range in temperate North America can have a daughter species that breeds in tropical South America. |
| 10 | During speciation, the ancestral range can be inherited by daughter species or fragmented, but the entire ancestral range must be cumulatively present in the daughter species. |
| 11 | Migratory daughter species cannot be derived from a non-migratory ancestor. |
* The list of “rules” is derived from the “Supporting Information” portion of the paper (Winger et al. 2014b:1). They are not cited here exactly as written, and include some explanatory notes not provided in the original text.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



