Submitted:
17 October 2023
Posted:
18 October 2023
You are already at the latest version
Abstract
The author attempts to reconstruct the climatic vicissitudes of the Mediterranean to explain why only broadly distributed Eurasian forest species have penetrated the Mediterranean peninsulas. He has carried out a bibliographic survey of the status of breeding forest birds in Italy over 15 decades (1872-2022) in order to establish an objective long-term trend (stable, increasing, decreasing, etc.). The number of breeding forest birds in Italy amounts to 61; their distribution, with a few exceptions, indicates that they are widespread in Eurasia, but only a small percentage of Eurasian forest species have colonized Italy and the other Mediterranean peninsulas, namely 49 in Iberian, 61 in Italian and 64 in Balkan peninsulas; a small percentage of them (between 10.9 and 14.3%) belongs to trans-Saharan migrants. The similarity between the forest species on the three peninsulas (Iberian, Italian and Balkan) results between 0.45 and 0.48, indicating a certain differ-ence in the overall fauna on the three territories. Not all species have penetrated southwards into the three peninsulas; for example, some that stopped in the Italian Alps have instead arrived to the forests of Greece, at a latitude corresponding to southern Italy, or species that in Italy stopped in the northern Apennines in the other two peninsulas have instead arrived far south. Iberian peninsula and the island of Corsica hold three endemic species among breeding forest birds, Italian and Balkan peninsulas have not endemic species. Overall, the Mediterranean presently hosts mainly neo-endemic taxa among forest bird species; the only paleo-endemics can be considered the three species of nuthatches living in Corsica, Algeria, and Turkey (other than Caucasus and the islet of Lesvos) and Le Vaillant’s woodpecker in the Maghreb (North Africa). Italian forests cover ca. 40% of land surface and since 1980’ are increasing, but 22% of them do not have a natural origin. However, it is difficult to know the true increase of forests, because some of them are fired every year. The presence of some ecologically demanding forest birds depends on the age of the trees, permanent open spaces and other characteristics at the edge of woodland.
Keywords:
list of breeding forest birds
; distribution over 150 years
; distribution limits
; peninsulas
; islands
; influence of man
; fires
1. Introduction
From the ornithological point of view, the Mediterranean area is one of the most explored regions, but for at least two millennia it has also been among the most densely populated areas by humans and most exploited by agriculture at the expense of forest areas. As for the forests that used to cover Italian territory, much of them have been taken away to make room for agriculture; this has undoubtedly caused a decline in forest bird species, however, less than species living in other environments. Italian forest bird populations, like those in the rest of the Mediterranean, are still substantial and undoubtedly deserve special attention from a biogeographical and conservationist point of view.
Altogether, over the last two centuries, the populations of some species of Italy's avifauna have gone into definitive extinction and others have undergone an unprecedented decline, due to direct and indirect anthropic pressure. The extinction of species in Italy is mainly due to man's increasingly intrusive activities over the millennia, but especially in the 20th century, when the vast majority of extinctions were recorded in the country. However, in the present century, as if to counterbalance the decline of some species, other birds are experiencing increases previously unreported in Italy, in some cases already recorded in other European countries. Recently, some forest species begun a process of colonization of environments that are covered by tree vegetation, but are generally planted by man, like urban habitats. For example, woodpigeon Columba palumbus, blackbird Turdus merula, magpie Pica pica, jay Garrulus glandarius and serin Serinus serinus since the 1990s have started to colonize urban areas and have become an integral part of the avifauna of built-up areas.
At the same time, it is difficult to understand the reasons for the decline of some species; it is not always clear whether it has been the excessive hunting, the destruction and fragmentation of some habitats, the climate change, or in the case of forest environments the fires, generally arson, that occur more frequently and violently each year. Increasing the extent of areas covered by large forest fires in Italy in the last fifteen years, in 2023 approaching 69,000 ha, including more than 10,000 ha of forest ecosystems; the forests involved are largely Mediterranean scrub and oakwood (61%) and areas covered by coniferous forests and afforestation (21%). Eighty-nine percent of the areas burned in 2023 are in Sicily (72%) and Calabria (17%) (data after the Istituto Superiore per la Protezione e la Ricerca Ambientale, https://www.isprambiente.gov.it/). Thus, undoubtedly the fires could be one of the main local causes of the decline of some forest and scrubland species, particularly those that are more demanding and have a rather specialized ecological niche (e.g., woodpeckers, treecreepers, etc.).
Italy has a total land area of 302,073 km2, while its present wooded surface is about 120,000 km2, amounting to ca. 40% of the total surface [1]. Forests historically occupied a large part of Italy's land area, but as early as Roman times they were reduced for various reasons (use of timber for shipbuilding, modification of the soil to make it arable, etc.). At the beginning of the last century, however, substantial afforestation activity took place that increased the wooded area. According to recent statistics, the surface of forests in Italy presently is increasing, but not all forests are equal, there is a big difference between natural forests and afforestation, of course. About 65,630 km2 (22% of total Italian land area) of the contemporary forests surfaces do not have a natural origin, but are the result of secondary successions occurred on abandoned farmed land since 1888; ca. 10,000 km2 (3.3%) are due to afforestation carried out between 1883 and 1939, and between 1952 and 1972. About 80% of them was done with conifers, mostly black pine, sometime replacing previous broadleaved woods [1].
During the course of our lives, it is rather difficult to have clear-cut answers as to the trend of species, but if we go through the past through the bibliography, we can gain a much better insight into the population dynamics of different species. The aim of this study was precisely to try to understand the old origin of Italian breeding forest birds and to investigate the evolution of their long-term trend over a fairly long period, from 1872 to 2022, i.e. 150 years, for which a conspicuous bibliography is available. Since, here we are talking about the entire Italian forest surface, it will not be possible to subdivide forests by essence and characteristics, but we will try to make some distinctions in the discussion.
2. Materials and Methods
Four periods were chosen between 1872 and 2022 in order to obtain an overall trend of breeding forest birds in Italy: a) before 1872 [2]; b) 1873-1929 [3,4,5]; c) 1930-1993 [6,7]; d) 1994-2022 [8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23]. The time periods were chosen on the basis of the available bibliography and therefore cover different periods of time, namely ca. 50, 56, 63, 28 years, respectively. By following the trend over 15 decades, and carefully studying species by species what the different authors have written, it was possible to understand the possible long-term status of each species (stable, increasing, decreasing, strongly decreasing, fluctuating). The choice to include some species within the fluctuating category may appear subjective, but it is the result of careful literature analysis and decades of personal experience; however, it is likely that another researcher had included some of them among different categories. The category of fluctuating species is generally overlooked by most ornithologists, but really there are species that have undergone positive and negative population number changes that are very difficult to understand (e.g., hazel grouse, hawfinch); other species, like the crossbill, due to the irregular cone seeds availability, fluctuate regularly in central-north Europe, however much less in the Mediterranean area.
The choice of forest species was established on the basis of their main habitat according to Brichetti and Fracasso [8,9,10,11,12,13,14,15,16,17,18] and Lardelli et al. [23]; e.g., some species, like blackbird, common redstart Phoenicurus phoenicurus, and European serin were excluded because although they live also in woody habitats, they occupy a very large part of arboreal habitats, including orchards and urban ones [23]. Additionally, African blue tit Cyanistes teneriffae and long-legged buzzard Buteo rufinus were excluded from the Italian list, being present outside their range only in the small island of Pantelleria (Sicily), just 80 km away from Tunisia (North Africa).
The habitat of each species was obtained from the bibliography and personal experience; the ecological niche fulfilled by each species was divided into three parts (trees, soil and undergrowth), in order to define the main activities (nesting, feeding) of the species in the three portions of the forest environment considered. Additionally, on the basis of species distribution and ability to colonize forest (or urban) habitats of little naturalistic interest the species was assigned one of the following three values: 1: of broad ecological value; 2: moderately ecologically demanding; 3: very ecologically demanding; a higher value of species corresponds to a greater number of ecologically demanding birds.
The conservation status of species in Europe has been obtained from BirdLife International (Red List) [24]. In order to carry out a comparison with forest birds of the Iberian and Balkan peninsulas, a list of forest birds breeding in those peninsulas was taken from the latest European Atlas [25] and other local references [26,27,28,29]. Sørensen's similarity index has been used, S = 2a/(2a+b+c), where a is the number of species in common in the two peninsulas considered, b and c are respectively the numbers of species found only in the first and only in the second peninsula compared. S can result between 0 (completely different) and 1 (identical) [30]. Concerning the trend of forest birds, a comparison has been tentatively carried out with the European trend simulated in the late 21th century in the climatic Atlas by Huntley et al. [31]. The systematics follow Baccetti, Fracasso and COI [32].
3. Results
3.1. Breeding Forest Species in Italy
The species regularly breeding in Italian forests, i.e. mainly present in these environments, number 61 (Table 1) and represent a modest percentage (20.8%) of the 293 species found breeding in Italy [23]. They belong to 20 families, namely: Phasianidae Tetraoninae (3 species), Columbidae (2 species), Caprimulgidae (1 species), Scolopacidae (1 species), Strigidae (6 species), Accipitridae (6 species), Picidae (8 species), Falconidae (1 species), Corvidae (2 species), Paridae (6 species), Phylloscopidae (3 species), Aegithalidae (1 species), Sylviidae (4 species), Certhiidae (2 species), Sittidae (1 species), Troglodytidae (1 species), Turdidae (4 species), Muscicapidae (2 species), Regulidae (2 species), and Fringillidae (5 species) (Table 2). Except for 12 species (19.7%), i. e. the three species of Phasianidae Tetraoninae (capercaillie Tetrao urogallus, black grouse Lyrurus tetrix, hazel grouse Bonasa bonasia), boreal owl Aegolius funereus, pygmy owl Glaucidium passerinum, Ural owl Strix uralensis, grey-faced woodpecker Picus canus, three-toed woodpecker Picoides tridactylus, northern nutcracker Nucifraga caryocatactes, willow tit Poecile palustris, and fieldfare Turdus pilaris, absent south of Alps, the other 49 species (80.3%) are present in different regions of the Italian peninsula. However, 2 species (3.3%) are present only in north Italy (barred warbler Sylvia nisoria, and crested tit Lophophanes cristatus), while the other 47 species cover much of the peninsula area. Only in the islands (Sicily and Sardinia) the number of breeding forest species is reduced, due to insularity species impoverishment. Out of 61 species, none became extinct through 150 years (but this occurred locally), 28 (45.9%) resulted stable, 2 (3.3%) strongly decreasing, 8 (13.1%) decreasing, 18 (29.5%) increasing, and 5 (8.2%) fluctuating (Figure 1). Most species are sedentary, but at least 16 (26.2%) are migrant breeder and wintering. This means that in some cases breeding populations are very small (e.g., Eurasian woodcock Scolopax rusticola, rare breeder in Italy), but conspicuous populations arrive during the winter; in other cases wintering populations are added to already conspicuous breeding populations (e.g., European robin Erithacus rubecula). Only 8 (13.1%) are trans-Saharan species, that is breeding in Europe and wintering in tropical Africa.
Though they are species related to forest cover, do not all have similar ecology, habits and trophic requirements. In fact, if we look closely at the different families, we can see that the forest is the habitat, but the ecological niche is sometimes fulfilled in the undergrowth, in clearings, at the edge of the forest, where the species find food, which may consist of invertebrates, berries, small mammals, etc. Table 2 summarizes the essential data highlighting the importance of trees, soil (litter) and undergrowth for the different species belonging to 20 bird families. It seems clear that while there are species groups closely related to trees (e.g., woodpeckers, treecreepers and nuthatch), many other species use forest trees only to place their nests and may forage both on the ground and in the undergrowth. Overall, 56 species (91.8%) use the trees for nesting. The most specialized species resulted to be those belonging to Picidae family, which are characterized by the zygodactily, that is the presence of two fingers pointing forward and two backward, other than the hardened rectrices that allow to lean on logs.
3.2. Biogeographical Peculiarities of the Breeding Forest Avifauna in Italy and the Other Peninsulas
Considering the Mediterranean broadly, three peninsulas project southwards into this great sea, the Iberian peninsula (which touches the Atlantic to the west), the Balkan peninsula (which touches the Black Sea to the east) and the Italian peninsula, the only one completely within the Mediterranean and proportionately narrow and long compared to the other two (Figure 2). In these regions the fauna generally has a north-south impoverishment, known as the 'peninsula effect', a form of impoverishment in 'quasi-island' territories, sensu Mac Arthur and Wilson [36; cf. also 37,38]. With regard to forest birds on the Italian peninsula, a north-south gradient of depletion has not been observed. The Apennines of peninsular Italy are home to large forests of various species with breeding bird populations in the southernmost part not substantially different from those in the north, with the exceptions above recorded.
Interestingly enough, are not the same northern species that have penetrated southwards into the peninsulas; for example, in the Iberian and Balkan peninsulas, crested tit has moved southwards, but in the Italian peninsula it has stopped in the northern Apennines. Conversely, Greece (latitude of S Italy) is the southernmost limit of western capercaillie, black grouse, hazel grouse, Eurasian pygmy-owl, boreal owl, grey-faced woodpecker, three-toed woodpecker, willow tit, and northern nutcracker [29], all species absent south of Italian Alps. Some species are present in the Balkan peninsula and northern Italy, but have not reached the Iberian peninsula: hazel grouse, grey-faced woodpecker, three-toed woodpecker, willow tit, northern nutcracker, wood warbler Phylloscopus sibilatrix, barred warbler, collared flycatcher Ficedula albicollis, and fieldfare. Others are present in the Balkan and Italian peninsulas, but reach only the north of Iberian peninsula: black woodpecker Dryocopus martius, middle spotted woodpecker Leiopicus medius, white-backed woodpecker Dendrocopos leucotos, ring ouzel Turdus torquatus, willow tit, and Eurasian treecreeper Certhia familiaris. Additionally, western Bonelli’s warbler Phylloscopus bonelli and western orphean warbler Sylvia hortensis are west Mediterranean species, present only in the Italian and Iberian peninsulas. Balkan peninsula has three forest birds not living in other Mediterranean areas: Syrian woodpecker Dendrocopos syriacus, Krüper’s nuthatch Sitta krueperi [living only on Lesvos Island (Greece), outside Turkey and Caucasus], and sombre tit Poecile lugubris [25]. Finally, Iberian peninsula has two endemic species, Iberian green woodpecker Picus sharpei (below discussed), and Iberian chiffchaff Phylloscopus ibericus.
The following numbers of breeding forest species were obtained from the Mediterranean peninsulas: Iberian peninsula 49 [25,26], Italian peninsula 61 [23], Balkan peninsula 64 [25,27,28,29]. The common component of the breeding forest avifauna of each peninsula is about 50%; altogether, 68 forest species breed in the three Mediterranean peninsulas, more or less equally distributed between them, but only part of species breeding in forests of each peninsula is in common with the other two (Sørensen's similarity index: 0.45-0.48; Table 3, Table 4, Figure 3, Figure 4). Seven species are trans-Saharan migrants in the Iberian and Balkan peninsulas, 14.3% and 10.9%, respectively, figures very similar to those obtained from Italian peninsula (13.1%).
Concerning the ecological value, in Table 3 are reported the sum of species values and their mean; while the sum of values may depend on the number of species, the mean is a more objective value. It resulted to be 2.0 for Iberian forest birds, and 2.1 for Italian and Balkan ones.
The conservation status of all Italian breeding forest birds resulted to be ‘Least Concern’ (LC), which means that no species is threatened.
4. Discussion
4.1. Comparison with the Trend Simulated in the late 21th Century
An attempt was made to compare the data collected from the Italian bibliography over 150 years and the trend simulated in the late 21st century by Huntley et al. [31]; it is not possible to make this comparison as the two methodologies are profoundly different. However, it is considered useful to make a few practical considerations. According to Huntley et al. [31] in the late 21st century of the 61 forest species considered here 35 (57.4%) will be decreasing in southern Europe, 14 (22.9%) decreasing in central Europe, only 8 (13.1%) remain stable, 2 (3.3%) increasing in southern and central Europe, respectively (Figure 5). For 12 species (19.7%), the trend matches both with the literature search through 150 years and with the simulation in the late 21st century; namely seven species (black grouse, capercaillie, eagle owl, stock dove, garden warbler, fieldfare, and ring ouzel) resulted to be decreasing in both the 150-year considered period and the late 21st century simulations, two species (short-toed snake-eagle, and booted eagle) were found to be increasing, and three species (blue tit, great tit, and wren) resulted stable in the two time periods. It seems obvious that the two methodologies give different results; in particular the simulation for forest species based on climatic trend is very pessimistic, as altogether 80.3% are expected to decline in the late 21st century, while the trend calculated through the bibliographic analysis gives a much lower figure (16.4%). Even if a predictive analysis that takes into account future climate projections should not disregard a bibliographic reference of long-term species trends, which implicitly includes information on each species' potential for possible adaptation to changes in habitat or climate, predictions for the late 21st century may be sadly true.
4.2. Forest Birds on Italian Peninsula and Islands
Considering that the Iberian peninsula is the extreme western apex of Eurasia, a minimal depletion of species seems normal, which in contrast is not evident in the Balkan peninsula, an appendage of Asia. The higher number of forest birds in the Balkan peninsula can be explained by the fact that it is an area in continuity with Asia and suffers less from the peninsula effect, more evident in the other two Mediterranean peninsulas. In addition, the extreme south-western land of Eurasia (Iberian peninsula), with its isolation, allowed the evolution of two endemic species of forest birds, Iberian woodpecker (Pyrenees), and Iberian chiffchaff (westernmost French and Spanish Pyrenees and western parts of the peninsula). Concerning endemic taxa of Mediterranean Europe, another species lives in Corsica, the Corsican nuthatch Sitta whiteheadi, linked to black pine Pinus nigra forests, taxonomically related to other two forest endemic species, one living on Kabyle, the Algerian nuthatch Sitta ledanti linked to forest remains dominated by Abies numidica, and the other, Krüper nuthatch Sitta krueperi, linked to Turkish pine Pinus brutia, living in Turkey, Lesvos Is. (Greece), and Caucasus.
The forests of peninsular Italy do not host any endemic representatives among the birds; all the species can be found in forest environments elsewhere in Europe. However, the Apennines are home to forest bird species that are most likely relicts of the last ice age, i.e. around 18,000-20,000 years old. Among these there is the trans-Adriatic subspecies of the white-backed woodpecker Dendrocopos leucotos lilfordi. This taxon, distributed in the Iberian and French Pyrenees to Asia Minor, Caucasus and Transcaucasia, may have arrived from the Balkans to Apennines and Pyrenees during the last ice age; the typical subspecies (leucotos), widespread across Eurasia from north, central and eastern Europe to northeast Asia, may instead have arrived in central Europe from the north. A similar case is that of the Eurasian green woodpecker; the species-complex consists of Picus viridis, distributed from western Europe to the Caucasus and Iran, the Iberian green woodpecker, distributed in the Iberian and French Pyrenees, and the related Le Vaillant’s woodpecker P. vaillantii, distributed in north-western Africa from central Morocco to Tunisia (Maghreb). The divergence event between the Iberian and the European lineages took place during the mid-Pleistocene (0.7–1.2 Ma), and a post-glacial range expansion of two lineages from distinct refugia located in the Iberian peninsula and in eastern Europe occurred. During the Pleistocene much of the forests suitable for green woodpeckers in central and northern Europe were covered by ice, tundra, steppe or other unsuitable habitat; thus, the woodpeckers might have occupied the current range during the past 20,000 years [39,40]. A similar case is reported also for a mammal, the chamois, present with Rupicapra rupicapra in the Alps and with another species, Rupicapra pyrenaica (subdivided into two subspecies, pyrenaica and ornata) in Pyrenees and Apennines, respectively [41].
Other possible relicts of the last ice age in the Apennines are black woodpecker, middle spotted woodpecker (Figure 6), goldcrest, Eurasian treecreeper, collared flycatcher (Figure 7), the western European subspecies of the bullfinch Pyrrhula pyrrhula europoea, howfinch Coccothraustes coccothraustes (with sedentary populations also in Sardinia), Eurasian siskin Spinus spinus (with a small breeding population also in Sicily, on Etna) (Figure 8) and red crossbill Loxia curvirostra ‘sensu lato’. The latter, unlike the northern populations (linked to cone seeds of larch and spruce pines), is associated with the black pine (with bigger cones than spruces and larches) and characterized by a more massive beak, smaller size, evident discoloration in both males and females, and marked sedentariness (Figure 9). Similar populations of sedentary crossbills evolved not only in the Italian peninsula, but also in the Iberian and Balkan peninsulas, Corsica, Sicily (Etna), Cyprus, and North Africa. Some of them live at the expense of the cones of the Aleppo pine Pinus halepensis, also characterized by big size. Polytypical species status has recently been proposed for the three main sedentary populations on Mediterranean peninsulas, islands and North Africa (L. balearica in the Balearics and Iberian peninsula, L. poliogyna in North Africa, Italian peninsula and Corsica, L. guillemardi in Cyprus and Balkan peninsula), as was previously the case for Loxia scotica [42].
The situation is different on the large Italian islands, which are poorer in species than peninsular Italy. Sicily has been definitively an island for only 20 thousand years [43], so it is assumed that numerous species reached it earlier and then remained isolated for around twenty thousand generations. However, this was enough to give rise to local endemic forms, as below reported. Far more isolated is Sardinia, which, in accordance with the reconstructions of Alvarez et al. [44], together with Corsica and other microplates (among which those that formed the Kabyle in Algeria) would have broken away from the Iberian peninsula and migrated towards the center of the Tyrrhenian Sea. This would have happened over 20 million years ago; this ancient isolation undoubtedly allowed numerous invertebrate species to differentiate, but as far as forest birds are concerned, there must have been some gene flow with other Mediterranean populations in fairly recent times. However, the isolation allowed the subspecific differentiation of some forest birds: jay Garrulus glandarius ichnusae, coal tit Periparus ater sardus, blue tit Cyanistes caeruleus ogliastrae, great tit Parus major corsus, wren Troglodytes troglodytes koenigi, sparrowhawk Accipiter nisus wolterstorffi, goshawk Accipiter gentilis arrigonii, and great spotted woodpecker Dendrocopos major harterti. Three of them are non-Passeriformes, which generally require longer isolation times than Passeriformes to differentiate themselves from continental populations. Most of them are subspecies in the Sardinian-Corsican system, recognized by taxonomists [45,46].
In Sicily, by far more recent island than Sardinia, live some isolated populations not yet taxonomically recognized, but certainly differentiated from continental ones. For example, Sicilian great and blue tits are smaller in size compared to continental individuals [pers. obs.]. The smaller size of these birds can be explained by Bergmann's rule according to which there is an inverse correlation between body size and average temperature in the geographical area of origin. The relict populations of the above-mentioned sedentary species may have remained isolated for several millennia, although for some of them probably there may have been mixing with more recent northern immigrant populations. It would be interesting to establish whether the isolation caused adaptations and differentiations or whether the gene flow was maintained, a fact that would deserve to be studied in depth with genomic investigations. A particular case is represented by the Sicilian population of long-tailed tit Aegithalos caudatus siculus (Figure 10), a taxon found mainly in broadleaved woods; its isolation in Sicily for at least 20,000 years has led to a clear differentiation, not only in the grey back, but also in the coloring of the head, which generally lacks the characteristic dark lateral bands, in the grey (not reddish) scapulars and in the size of the wing and tail, which are on average shorter than those of continental individuals [47,48]. It seems that interbreeding with peninsular populations does not occur, thanks to the 3 km long sea arm of the Strait of Messina between Sicily and south Italy. A similar case of isolation, but not occurred in a forest insular species, is that of the Sicilian rock partridge Alectoris graeca whitakeri, clearly differentiated from the peninsular taxon.
Probably due to the loss of mature woodland areas in Sicily, the eagle owl and three woodpecker species became extinct in the 20th century, namely black woodpecker, Eurasian green woodpecker, and lesser spotted woodpecker Dryobates minor [49]; the latter seems that disappeared also from Sardinia [22,33].
4.3. Palaeogeographical Vicissitudes of the Mediterranean
The Mediterranean sea is subject to very complex tectonic activities, due primarily to the strong collision between the large Eurasian Plate to the north and the African Plate to the south. Between them there are other microplates; the Hellenic Arc is an arc-shaped tectonic structure about 1,000 km long that stretches from the Ionian Islands, located north-west of Greece, to the island of Rhodes from which the Cyprus Arc originates.
Generating this complex structure was the subduction (still ongoing) of the African plate, moving S-N at a rate of about 10 mm/year, under the Aegean microplate, which in turn moves in a NE-SW direction by about 45 mm/year [50].
In order to understand the origin of the breeding forest avifauna in Italy, it should useful to trace the various vicissitudes that Mediterranean vegetation has gone through since the Tertiary. During the Late Cretaceous and Paleocene periods, tropical and subtropical forests dominated by oaks, laurels and palms covered the lower-middle latitudes of Eurasia and North America; at the beginning of the Tertiary period, North Africa was characterized by tropical forests and savannah. With the expansion of drier climates in Europe and Asia, xerophyllous species appeared in the Middle Eocene, which include ancestor species of today's woody plants with leathery leaves. In the early Oligocene, xerophyllous floras were widespread; environmental conditions from Spain to Central Asia favored Mediterranean-type plants [48,51].
In the Miocene (around seven million years ago), the climate of much of the earth's surface was torrid and the waters were inhabited by tropical species. In the second half of the Messinian (between about five and six million years ago), an exceptional event occurred: the Messinian salinity crisis, i.e. the drying up of the Mediterranean Sea. The relationship between the Atlantic Ocean and the Mediterranean Sea was interrupted; only a few large brackish lagoons remained, water was scarce for at least half a million years, and lions, buffalos, monkeys and elephants moved to the only areas where water still existed [51]. The pollen research carried out by Fauquette et al. [52] established that the Mediterranean area was then characterized by two types of vegetation: in the southernmost part conditions were dry and warm and the vegetation was relatively open, while in the northernmost part the warm-humid conditions allowed the growth of forest-type vegetation. The researchers were also able to establish that the salinity crisis was not directly caused by the climate and that the climate did not change substantially during this period, particularly in the southernmost part of the area, where it remained very dry and warm before, during and after the salinity crisis [52].
The Quaternary was a period of exceptional climatic instability [53]. During the cold-dry episodes, forests were replaced by steppes and tundra in the north and by savannah-like formations from the tropics to southern Europe; the continental vertebrate faunas of peninsular Italy and Sicily included elephants, lions and hippos, indicating a savannah environment even at our latitudes. In the Pliocene (between four and two and half million years ago), the climate must still have been relatively warm; the Mediterranean climate stabilized more or less three million years ago, but considerable climatic changes also occurred in the Pleistocene (between two and half million and eleven thousand seven hundred years ago), particularly during glaciations and interglacial periods.
The effects of each glaciation on the fauna were cancelled out by the next one, so that the last glaciation explains the distribution and diversification patterns of today's avifauna. Although the deciduous forests of Europe were reduced and compressed in the Mediterranean peninsulas during the Pleistocene, areas covered by forests remained in North Africa and the Middle East. During glacial periods there was a drop in sea level of the order of 60 meters in the first glaciation and as much as 130 meters in the last (Würm, around eighteen thousand years ago); the maximum temperatures would have allowed the coexistence of European vegetation and associated fauna [54]. It is from the period before the first glaciation (around 50,000 years ago) that pollen shows the presence of a Mediterranean-type vegetation, i.e. scrubland characterized by Quercus ilex (or Q. coccifera/calliprinos), Pistacia and Ericaceae (mainly represented by Arbutus unedo) [55]. Pollens dating back to the Würmian glaciation (30,000-25,000 years ago) show that much of southern Europe was composed of Artemisia steppes with scattered patches of forest (the average temperature was ca. 8 °C lower than today) and only later, after the retreat of the glaciers, 15,000 years ago, there was an expansion of Juniperus and Pinus, while 13,500 years ago, a re-growth of Quercus forests was observed [56]. This kind of vegetation favored the spread of bird species associated to coniferous (e.g., crossbills, siskin) and to evergreen broadleaved (many species of passerines, woodpeckers, etc.) (see Table 1).
In Mediterranean habitats today evergreen sclerophylls are found, which have adapted to the abundant winter rains that alternate with very dry summers; they have small, thick, evergreen leaves, usually with a villous covering and frequently with a robust cuticle, which allow them to retain moisture during long dry periods [57]. Their deep root systems allow them to capture water at depth; in addition, many species are well adapted to withstand fire disturbance, promoting rapid re-growth of vegetation after fire. This type of structure seems to have originated in relatively warm and moderately mesic conditions and has pre-adapted different species to climates with shorter periods of rainfall and greater temperature variability. Such modifications are ancient, as can be judged from the presence of essentially modern scrub and woodland evergreen sclerophyll taxa in the Miocene and Oligocene floras, which experienced very wet summers. These characters are therefore not an evolutionary response to an ancient and persistent Mediterranean-type climate; from a functional point of view, adaptation to the Mediterranean climate is a fine-tuning when considering the ability to reproduce in a short period of wet weather during the coldest part of the year [57]. Aridity in summer is the major limiting factor for plant activity, not the cold of winter; flowering and growth take place mainly in winter and spring and only a few species and rare individuals vegetate in summer [51,53]. The Mediterranean climate is therefore much younger than the sclerophylls, as it only appeared after the first glacial events.
4.4. The Climatic Phases
The climatic phases have been studied both by sampling and identifying fossil pollen, and by analyzing the ratio of oxygen 18O to 16O, which changes with changing temperatures. When temperatures are very low due to ice caps, i.e. during glaciations, water and marine organisms are enriched in 18O and thus the ratio between 18O and 16O is in favor of the former; the opposite occurs when temperatures rise, i.e. during interglacial stages [52]. Thanks to the study of the ratio between 18O and 16O, it was discovered that glacial and interglacial stages amount to about sixty and glaciations to about thirty. These periodic climatic changes make us understand how shifts in the range of fauna and flora could have taken place southwards and then northwards, and also how so many endemic invertebrate species could have formed in mountainous areas of the Mediterranean peninsulas, where populations could remain imprisoned and isolated for long periods of time and thus undergo differentiation, thanks to small adaptive changes. Examples are the Iberian mountains, the Apennines in Italy and the Balkans in the east, areas rich in taxa of endemic invertebrates but with only single cases of endemic forest bird species [25,58,59]. It is difficult to explain why certain species did not reach the central and southern Apennines (e.g., crested tit), especially when one considers that they were able to colonize the other two Mediterranean peninsulas; however, the recent expansion in north Apennines of crested tit [23] shows its present ability to live at lower Apennine latitudes.
It is very interesting to highlight that when comparing the entire breeding avifauna with the forest birds of the three peninsulas the same Sørensen's similarity index values were found [59]; in a sense, the forest species represent a significant sample of the entire avifauna of the three peninsulas and show that a similar biogeographical story occurred.
As stated above, corroborated by paleobotanical and paleontological data, at least during the last two million years the expansion and contraction phases of glacial and Arctic conditions and the consequent expansion of ecological zones forced the entire European avifauna to take refuge in the Mediterranean [60]. Thirteen thousand years ago deciduous oak forests were located at the southern tip of the Balkan, Italian and Iberian peninsulas; they reached Scandinavia only six thousand years later. Mediterranean vegetation never completely disappeared during the Pleistocene, even under the most severe climatic conditions. During the interglacial period, Mediterranean forests at low to medium altitudes were widespread, mainly dominated by deciduous and non-evergreen broadleaf trees. The expansion of vegetation took place between twelve thousand and nine thousand five hundred years ago [53]. This has allowed the majority of species to continue living in these Mediterranean-type habitats.
4.5. Man's Influence On Present-Day Forest Avifauna
The great naturalist Gaius Plinius Secundus (Pliny the Elder) wrote ‘Summum munus homini datum arbores silvaeque intelligebantur’ [translated from Latin: Trees and forests must be understood as the supreme gift given to mankind]. It seems clear that already at the beginning of the first millennium, men of science were aware of the importance of forests and individual trees. The Mediterranean, and in particular the three peninsulas, were then covered with important forests, which, due to overexploitation and the need to obtain cultivable land, were considerably reduced over the following hundreds of years. It is obvious that the animal populations that were linked to those forests also declined; however, to the best of our knowledge, no forest birds have become extinct in historical times, except for in single insular regions.
The present extension of the maquis is associated with deforestation by man, which has not stopped since the beginning of the Neolithic period, nine thousand years ago. The most favorable time for the natural expansion of the maquis was probably in the Xerothermal period, between eight and four thousand years ago, when the climate was drier and warmer than today [51]. At the beginning of the Holocene, 10,000 years ago, the warming of the climate became more pronounced, the incident solar radiation at Mediterranean latitudes was about 7% higher than today, and precipitation more numerous. It should be noted that in the Fezzan (Matkandush, Sahara, Libya) there are rock carvings of species such as giraffes, elephants, dating back some seven thousand years, indicating the existence of a savannah where today there is a rocky desert (Figure 11). This is evidence of the natural relatively rapid climate change a few thousand years ago, and the youth of the Sahara desert. This was the most important period for the pedogenesis of the Mediterranean where there was a climatic optimum; it was during this period that the first Neolithic cultures and the very first agricultural activities were born. The pastoralism and agricultural practices marked the first changes to the environment that would later become increasingly accentuated. According to Le Houérou [60], between twelve and four thousand years ago, during the Neolithic and Protohistoric revolutions, man exerted a very important transformative action on vegetation, initially in the eastern and then in the western Mediterranean. The definition of ‘revolution’ is due to the fact that in the 2,500 years of that period there was a large increase in the world’s human population, which grew from 10 million to 100 million [60].
Between five and four thousand years ago there was also a climatic deterioration towards aridity throughout the Mediterranean, and for at least 2,500-4,000 years shepherds and farmers have been burning down forests to obtain better pastures and new land for cultivation. This has had a particular significance on Mediterranean vegetation, which can undoubtedly be considered fragile and vulnerable, due to the climatic conditions in which it lives (relatively mild winter, long aridity in summer), especially susceptible to fire and overgrazing. In much of the Mediterranean region historically logging and fire have degraded the original oak forests to the maquis stage and altered the relative abundance of other smaller tree species. Overgrazing and inappropriate cultivation practices subsequently caused the disappearance of shrub species, changing maquis into garrigue, a type of woody Mediterranean vegetation consisting of very low evergreen shrubs (e.g., rosemary, thyme, broom, dwarf palm, etc.) and allowing soil erosion. Thus, little by little the ancient forest landscape was converted into heaths covered with grasses and other wind-resistant and undemanding herbaceous species. Human activity has produced vast expanses of garrigue; bare spaces and small rocky areas are often interspersed among the plants. Due to regular fires or overgrazing, garrigue can cover large expanses and is perhaps one of the most widespread environments in the Mediterranean because of these very negative factors.
The present extent of forests in Italy amounts to 120,000 km2, accounting for ca. 40% of the country's area. Most forests are found in the Alps and Apennines, but a fair amount of forest cover also exists in coastal (e.g., pine forests), lowland areas, and islands. Over nearly a century, humans have also reforested, mostly with conifers, and in many cases terraced hillsides and mountainsides in order to plant trees. Thus a fair percentage of Italy's forest cover is made by tree planting.
Throughout the Mediterranean area, and particularly in islands characterized by rugged hills and mountains, man has sought to obtain arable land by terracing with great difficulty and powerful manpower the slopes, even steep hills and mountains, in order to obtain soils where to cultivate. Cyprus, Crete, Sicily have mountain terraces that have served both for the planting of coniferous forests and for the cultivation of olive groves (Figure 12). Eventually, although tree cover of limited ecological value, these have served for the establishment of some of the most widespread species of forest birds. These events have contributed in no small measure to the recent distribution of bird species, especially those associated with tree vegetation.
According to available statistics, tree cover in Italy is increasing, mainly since 1980’ [1; Figure 13]; recently the extent of the forest has increased remarkably, rising from 91,414 km2 in 1990, to 94.704 km2 in 2000, and reaching 96,532 km2 in 2008 (for all three dates, the error standard of the estimate is very small, amounting to 0.1%) [61]. Thus, during the period 1990-2008, there was an increase in forest area of 5,1 km2. It was certainly the same phenomenon of progressive abandonment of rural land that sees the triggering of processes natural recolonization by shrub and tree species in marginal areas [61]. However, it should be noted that while it is true that due to the abandonment of agriculture, mainly in central and northern Italy, forests are expanding naturally, much of the recent forest increase (22%, according to Agnoletti et al. [1]) is due to afforestation, mainly with conifers.
There is an obvious difference between natural forests and afforestation; many species of invertebrates or even herbaceous or shrubby plants established within natural forests are lacking in afforestation, and thus some bird species select only certain forest environments and not others. Data collected in the Iberian peninsula [62] allowed to find out the relationships between a series of environmental variables and the abundance of individual species of forest birds. The variable most highly correlated with the abundance of species was floristic composition and the diameter of trees; bird density was also correlated with mean annual rainfall (positive correlation: e. g., chiffchaff, goldcrest, robin, bullfinch; negative correlation: e. g., Bonelli’s warbler) [62]. This shows that obviously each forest has its own characteristics, and these in turn allow the presence of certain species and not others.
Although, in general, from 1980 to 2003 populations of many common forest birds in some parts of Europe appeared to be declining by 13-18% [63], in particular in Great Britain [64], Gil-Tena et al. [65] found a certain degree of variation across the Iberian peninsula. Interestingly, the percentage of diminution is very similar both for species and populations. Overall, in Italy a similar percentage (16.4%) of forest bird species resulted to be decreasing, both over a long time (150 years) and in present times [23], also similar to that found (20%) by Gil-Tena et al. [65] in Spanish forest bird populations. Latter authors highlighted that forest maturation is most influencing dynamic of forest birds, which are more related with advanced forest development stages than with initial succession stages as those of young forests. Overall, species that inhabit a forest do not remain in that forest forever; the greatest species diversity can be found in forests where different stages of succession are represented at all times [66]. In conclusion, presently this may depend on the forest management by man, as well as on the age of the trees, the presence of permanent open spaces (important to birds), and other characteristics at the edge of woodland.
The forest consists of numerous microhabitats that depend on forest species (conifers, broadleaf trees of different species, etc.), age, exposure, altitude, amount of litter, and the presence of undergrowth and clearings. These ecological factors enable the establishment of different species of birds. An afforestation of allochthonous Eucalyptus is certainly unattractive to most forest species, not only birds; however, it can be easily understood that the most ecologically demanding species are found only in certain types of forest. Generally if a species is uncommon then it will occur only in wide woods with singular ecological characteristics.
Additionally, it is very difficult to know the true increase in forest area in Italy, as this should be offset by that lost to fires; statistics on the area of forest destroyed by fire are very poor and in some cases absent. The Italian regions more affected by fire generally are Sicily, Calabria, and Sardinia. In terms of percentage, the habitats more affected by fires from 2000 to 2023 are Mediterranean pine forests, and Italy resulted to be the 3rd country more affected by forest fires in Europe (after Spain and Portugal) [67]; ca. 1,784,000 hectares of Italian land have been fired, mainly arson, between 2000 and 2023, that is 5.9% of total Italian land area (Figure 14). Of course, not all of the fire passed through forests, some of it affected pastures, shrubs, garrigue, and even cultivated land; the majority of the forests traversed by the fire, however, were coniferous afforestation, which once burned are obliterated forever.
Following Blondel [68] Mediterranean forest birds originated in the large forest blocks of Eurasia; every species living in the Italian forests, as well as in the other Mediterranean peninsulas, are almost certainly also found in central European forest environments. For the reasons stated above, species of true Mediterranean origin are hardly found in mature evergreen oak forests today. The nearly absence of endemic taxa among Mediterranean Europe forest birds is surprising, especially when one considers how many endemic plants there are and how many opportunities species have had to isolate themselves [68,69]. Overall, the Mediterranean avifauna is no different from that of Europe today, but barely impoverished, probably due to the fact that the Mediterranean region falls on the south-western edge of Eurasia [68]. Only in the Iberian peninsula two endemic forest birds live; however, the fact that all forest species present in Italy are Least Concern [24] also means that generally researchers who dealt with the red lists did not consider them to be at risk because these birds are widespread in Europe and do not face any kind of present threat. However, according to the Italian red list [70] six forest bird species are considered at local risk: barred warbler (Critically Endangered), black grouse (Endangered), western capercaillie (Vulnerable), middle spotted woodpecker (Vulnerable), white-backed woodpecker (Vulnerable), and garden warbler (Vulnerable); in addition the Sardinian subspecies of goshawk Accipiter gentilis arrigonii is considered Endangered, while the other forest species are listed as Least Concern. The long-term study calculated over 150 years, however, shows a less risky trend for the two woodpecker species above cited (‘stable’ in Table 1), while for the other four species in Italy the status is not at all positive. Also, the mean ecological value (2.0-2.1; Table 2) shows a good presence of ecologically demanding species in all the three peninsulas.
More recently, in the last few decades, perhaps due to climate change or the increased trophic availability within cities, some forest species have begun to colonize the tree cover of towns and have become heavily urbanized. Some species (e.g., woodpigeon, blackbird, serin) had already carried out this urban colonization process many years earlier in central Europe, others (e.g., jay) have begun to do so in Italian towns. The reasons for this recent immigration may be many: better winter temperatures inside cities compared to forests, less likelihood of predation for some species, greater availability of trophic resources, which allow a greater number of reproductions per year (e.g., woodpigeon, three times a year and sometimes from January to November) [71], and finally lower mortality of fledglings. In addition, other species, like siskin, benefited from recent conifer afforestation, mainly in south Italy [G. Congi, pers. comm.].
Among the causes of the expansion of bird species there are creation of new habitats, increase in trophic availability, and climatic fluctuations. It is not always easy to understand the cause of the expansion of certain species, but certainly new trophic factors and probably, at least as far as urban environments are concerned, significantly higher average temperatures in association with the anthropic activities from which the birds sometimes benefit, contribute. This takes place because in nature, animal populations undergo colonization events, declines, increases or simple fluctuations over time [72,73,74].
5. Conclusions
Overall, today the Mediterranean forest avifauna is no different from that of Europe, but hardly impoverished, very likely due to the fact that the Mediterranean region falls on the south-western edge of Eurasia. The nearly absence of endemic species among Mediterranean forest birds is unexpected, especially when one considers how many endemic plants there are and how many opportunities species have had to isolate themselves. However, from the reconstruction made in the preceding pages, it can be understood that the avifauna of the forests of Mediterranean Europe is of fairly recent origin and, above all, the result of the isolation of Eurasian populations during the ice ages. The effects of each glaciation on the fauna were cancelled out by the next one, so that the last glaciation explains the distribution and diversification patterns of presently forest avifauna. Most species became definitively established during the last ice age and can be considered in this sense as glacial relicts. A portion of species that colonized the Balkan peninsula in turn colonized the Italian peninsula via the trans-Adriatic route and are therefore absent from the Alps. However, those which have differentiated from north-eastern populations should be considered neo-endemic taxa, dating back some 18-20,000 years. Thus, this time frame was sufficient for differentiation of isolated populations of some species, at the subspecific or specific level. This took place in the Iberian peninsula, Sicily and probably the Apennines. Only the Iberian peninsula has two endemic species, which have become isolated due to the Pyrenean barrier and the fact that the Iberian peninsula is the extreme south-western apex of Eurasia, separated from North Africa by the Strait of Gibraltar. It remains quite unknown what has happened in the Sardinian-Corsican system, which, although very old, presently hosts only a few differentiated populations of some forest species of birds, except for Corsican nuthatch, a species endemic to Corsica, and absent elsewhere, related to Algerian nuthatch (Kabyle) and Krüper nuthatch (Turkey, Caucasus and Lesvos islet). Here briefly another Mediterranean forest species deserves to be mentioned, Le Vaillant woodpecker in North-West Africa, belonging to green woodpecker group and isolated in the Maghreb, probably when the Strait of Gibraltar reopened around 5 million years ago. This woodpecker, and the three species of nuthatches mentioned above are undoubtedly among the oldest forest species in the Mediterranean, and may be considered paleo-endemic taxa.
Italian forest species, like those in the rest of the Mediterranean, will face some challenges in the coming decades because of the problems faced by their elective habitat, the forest. The main threats to forests are climate change, global warming, reduction of old-growth forests, increased fires and CO2 emissions (Figure 15). As we have seen above Huntley et al. [31] have attempted to simulate the future of European avifauna in the late 21st century; the very pessimistic vision they present, much more than the trend currently calculated on the basis of the bibliography consulted, may be grimly realistic, but this will depend only on humans making important decisions for environmental protection.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
First of all, due credit should be given to Jacques Blondel for first outlining the possible origin of Mediterranean forest birds; without his work this article would not have seen the light of day. I also would like to thank the editors of the recent Atlas of Italian breeding birds, who invited me to write a chapter on the biogeography of Italian birds; this gave me the inspiration to continue exploring the topic in depth. Many thanks are due to many friends, specialists on Mediterranean birds, for their advices, suggestions and fruitful discussions on biogeographical, taxonomic and habitat issues. In addition, I would like to thank Fulvio Fraticelli for the stimulating discussions on forest ornithological topics, Tommaso La Mantia for some bibliographical references and suggestions on forestry issues, Rafael de Silveira Bueno for his support in the processing of Italian forest area, Gianluca Congi for his interesting advices and nice photos of forest bird species from south Italy kindly provided, Mathia Coco, who has kindly made available the photo of the Sicilian long-tailed tit, and Violetta Massa, who depicted the Figure 14.
Dedication
This article is dedicated to the author Italo Calvino (1923-1985) on the centenary of his birth. Calvino, the only humanist in a family of scientists, father agronomist, mother botanist, two uncles chemist, and a brother geologist, undoubtedly felt the need to recompose the rift with his family, writing in an ecological vein ‘Il Barone rampante’ (1957) an indictment against the building speculation of the 1950s, when no one was yet talking about ecology. One sentence within this short story is interesting: «I do not know if it is true what one reads in books, that in ancient times a monkey that had set out from Rome jumping from tree to tree could reach Spain without ever touching the ground. In my time of places so dense with trees there was only the gulf of Ombrosa from end to end and its valley to the crests of the mountains». Calvino testified to the ancient continuity between Italian forests and those in France and Spain, which is discussed in this article regarding forest birds, some of which due to habitat fragmentation are declining. Furthermore, in the short story 'La nuvola di smog' (1958) he talks about a cloud of smog that gravitates over the city in which the protagonist lives causing dust and dirt to pervade everything, soiling rooms, clothes, walls and streets; in the same book he included another short story 'La formica argentina', already published in 1952, where he treats among the first in Italy of an alien species: the unpleasant invasion in the 1920s of the Argentine ant in San Remo and the Riviera di Ponente (Liguria, Italy).
| 1 | |
| 2 | The presence in Sicily of middle spotted woodpecker was erroneously reported by Minà Palumbo [34; see also 35]. |
| 3 | According to Lardelli et al. [23] the willow tit is absent from Apennines, where evidences of its presence are lacking. |
References
- Agnoletti, M.; Piras, F.; Venturi, M.; Santoro, A. Cultural values and forest dynamics: The Italian forests in the last 150 years. Forest Ecol. Manage, 2022; 503. [Google Scholar] [CrossRef]
- Salvadori, T. Fauna d’Italia. Uccelli. Vallardi, Milano, 1872.
- Martorelli, G. Gli Uccelli d’Italia. Rizzoli and C., Milano, 1906.
- Giglioli, H.E. Secondo resoconto dei risultati dell'Inchiesta Ornitologica in Italia. Avifauna Italica. Tip. S. Giuseppe, Firenze, 1907.
- Arrigoni Degli Oddi, E. Ornitologia italiana. Hoepli, Milano, 1929.
- Schenk, H. Analisi della situazione faunistica in Sardegna. Uccelli e Mammiferi. In: SOS Fauna. Animali in pericolo in Italia. WWF Ed., Camerino, 1976, pp. 465‒556.
- Meschini, E.; Frugis, S. (Eds.) ; Atlante degli Uccelli nidificanti in Italia. Suppl. Ric. Biol. Selvaggina, 1993, 20, 1‒344.
- Brichetti, P.; Fracasso, G. Ornitologia italiana. Vol. 1. Gaviidae-Falconidae. A. Perdisa Ed., Bologna, 2003.
- Brichetti, P.; Fracasso, G. Ornitologia italiana. Vol. 2. Tetraonidae-Scolopacidae. A. Perdisa Ed., Bologna, 2004.
- Brichetti, P.; Fracasso, G. Ornitologia italiana. Vol. 3. Stercorariidae-Caprimulgidae. A. Perdisa Ed., Bologna, 2006.
- Brichetti, P.; Fracasso, G. Ornitologia italiana. Vol. 4. Apodidae-Prunellidae. A. Perdisa Ed., Bologna, 2007.
- Brichetti, P.; Fracasso, G. Ornitologia italiana. Vol. 5. Turdidae-Cisticolidae. A. Perdisa Ed., Bologna, 2008.
- Brichetti, P.; Fracasso, G. Ornitologia italiana. Vol. 6. Sylviidae-Paradoxornithidae. A. Perdisa Ed., Bologna, 2010.
- Brichetti, P.; Fracasso, G. Ornitologia italiana. Vol. 7. Paridae- Corvidae. A. Perdisa Ed., Bologna, 2011.
- Brichetti, P.; Fracasso, G. Ornitologia italiana. Vol. 8. Sturnidae-Fringillidae. A. Perdisa Ed., Bologna, 2013.
- Brichetti, P.; Fracasso, G. Ornitologia italiana. Vol. 9. Emberizidae-Icteridae. A. Perdisa Ed., Bologna, 2015.
- Brichetti, P.; Fracasso, G. The birds of Italy. Volume 1. Anatidae-Alcidae. Belvedere Ed., Latina, Italy, 2018.
- Brichetti, P.; Fracasso, G. The birds of Italy. Volume 2. Pteroclidae-Locustellidae. Belvedere Ed., Latina, Italy, 2020.
- Corso, A. Avifauna di Sicilia. L’Epos Ed., Palermo, 2005.
- Gustin, M.; Brambilla, M.; Celada, C. Stato di conservazione e valore di riferimento favorevole per le popolazioni di uccelli nidificanti in Italia. Riv. ital. Orn, 2016, 86 (2), 3‒58. [https://sisn.pagepress.org/index.php/rio/article/view/332]. [CrossRef]
- Massa, B.; Ientile, R.; Aradis, A.; Surdo, S. One hundred and fifty years of ornithology in Sicily, with an unknown manuscript by Joseph Whitaker. Biodiversity J. 2021, 12, 27‒89. [CrossRef]
- Grussu, M. New checklist of the birds of Sardinia (Italy). Edition 2022. Aves Ichnusae, 2022, 12, 3‒62. [https://www.researchgate.net/publication/366733621_New_checklist_of_the_birds_of_Sardinia_Italy_Edition_2022].
- Lardelli, R.; Bogliani, G.; Caprio, E.; Celada, C.; Fraticelli, F.; Gustin, M.; Janni, O.; Pedrini, P.; Puglisi, L.; Rubolini, D.; Ruggieri, L.; Spina, F.; Tinarelli, R.; Calvi, G.; Brambilla, M. (Eds.) ; Atlante degli Uccelli nidificanti in Italia. Ed. Belvedere, Latina, Italy, 2022.
- BirdLife International, European Red List of Birds. Office for Official Publications of the European Communities, Luxembourg, 2015. [https://www.iucnredlist.org/species/pdf/attachment].
- Keller, V.; Herrando, S.; Voříšek, P.; Franch, M.; Kipson, M.; Milanesi, P.; Martí, D.; Anton, M.; Klvaňová, A.; Kalyakin, M.V.; Bauer, H.-G.; Foppen, R.P.B. European Breeding Bird Atlas 2: Distribution, Abundance and Change. European Bird Census Council and Lynx ed., Barcelona, 2020.
- De Juana, E.; Varela, J.M. Birds of Spain. Lynx and SEO BirdLife, Barcelona, 2016.
- Matveiev, S.D.; Vasić, V.F. Catalogus Faunae Jugoslaviae. IV/3. Aves. Academia Scientiarum et Artium Slovenica, Ljubljana, 1973.
- Matveiev, S.D.; Vasić, V.F. Catalogus Faunae Jugoslaviae. Aves. Addenda et corrigenda. Larus, 1977, 29‒30, 123‒136.
- Handrinos, G.; Akriotis, T. The Birds of Greece. C. Helm, London, 1997.
- Pielou, E.C. Mathematical Ecology. John Wiley and Sons, New York, 1977.
- Huntley, B.; Green, R.E.; Collingham, Y.C.; Willis, S.G. A climatic Atlas of European breeding birds. Lynx ed., Barcelona, 2007.
- Baccetti, N.; Fracasso, G. ; Commissione Ornitologica Italiana (COI). CISO-COI Checklist of the Italian Birds – 2020. Avocetta, 2021, 44, 21‒82. [https://ciso-coi.it/coi/checklist-ciso-coi-degli-uccelli-italiani/].
- Massa, B.; Schenk, H. Similarità tra le avifaune della Sicilia, Sardegna e Corsica. Lav. Soc. it. Biogeogr, 1983, 8, 757‒799. [https://escholarship.org/uc/item/4tm114bn].
- Minà Palumbo, F. Aggiunte al catalogo degli Uccelli delle Madonie. La Favilla, 1857, 1 (10-11), 84‒86.
- Massa, B.; Sarà, M. Uccelli/Birds. Iconografia della Storia Naturale delle Madonie. Vol. IV. Sellerio ed., Palermo, 2011.
- Mac Arthur, R.H.; Wilson, E.O. The theory of island Biogeography. Princeton University Press, Princeton, 1967.
- Massa, B. Il gradiente faunistico nella penisola italiana e nelle Isole. Atti Soc. ital. Sc. nat. Mus. civ. St. nat. Milano.
- Battisti, C.; Testi, A. Peninsular patterns of breeding landbird richness in Italy: on the role of climatic, orographic and vegetational factors. Avocetta, 2001, 25, 289–297. [https://www.avocetta.org/articles/vol-25-2-vdd-peninsular-patterns-of-breeding-landbird-richness-in-italy-on-the-roleem-emof-climatic-orographic-and-vegetational-factors/].
- Pons, J.-M.; Olioso, G.; Cruaud, C.; Fuchs, J. Phylogeography of the Eurasian green woodpecker (Picus viridis). J. Biogeogr, 2011, 38, 311–325. [CrossRef]
- Perktas, U.; Barrowclough, E.F.; Groth, J.G. Phylogeography and species limits in the green woodpecker complex (Aves: Picidae): multiple Pleistocene refugia and range expansion across Europe and the Near East. Biol. J. Linnean Soc. 2011, 104, 710–723. [https://www.researchgate.net/publication/231814818_Phylogeography_and_species_limits_in_the_Green_Woodpecker_complex_Aves_Picidae_multiple_Pleistocene_refugia_and_range_expansion_across_Europe_and_the_Near_East].
- Nascetti, G.; Lovari, S.; Lanfranchi, P.; Berducou, C.; Mattiucci, S.; Rossi, L.; Bullini, L. Revision of Rupicapra Genus. III. Electrophoretic studies demonstrating species distinction of chamois populations of the Alps from those of the Apennines and Pyrenees. In: Lovari, S.; Ed. The biology and management of mountain ungulates. Croom Helm, London, 1985, pp. 56-62. [https://www.researchgate.net/publication/310842731_Revision_of_Rupicapra_genus_III_electrophoretic_studies_demonstrating_species_distinction_of_chamois_populations_of_the_Alps_from_those_of_the_Apennines_and_Pyrenees].
- Massa, B.; Canale, E.D.; Lo Verde, G.; Congi, G.; La Mantia, T.; Ientile, R. Mediterranean Crossbills Loxia curvirostra sensu lato (Aves, Passeriformes): new data and directions for future research. Riv. ital. Orn. 2022, 92, 41‒60. [https://sisn.pagepress.org/index.php/rio/article/view/574]. [CrossRef]
- Antonioli, F.; Lo Presti, V.; Morticelli, M.G.; Mannino, M.A.; Lambeck, K.; Ferranti, L.; Bonfiglioli, C.; Mangano, V.; Sannino, G.M.; Furlani, S.; Canese, S.P. The land bridge between Europe and Sicily over the past 40 kyrs: timing of emersion and implications for the migration of Homo sapiens. Rend. Online Soc. geol. ital, 2012, 21, 1167‒1169. [https://www.rendicontisocietageologicaitaliana.it/297/article-690/the-land-bridge-between-europe-and-sicily-over-the-past-40-kyrs-timing-of-emersion-and-implications-for-the-migration-of-em-homo-sapiens-em.html].
- Alvarez, W.; Cocozza, T.; Wezel Forese, C. Fragmentation of the Alpine orogenetic belt by microplate dispersal. Nature, 1974, 248, 309‒314.
- Vaurie, C. The birds of the Palearctic fauna. Passeriformes. H.F. and G. Witherby, London, 1959. [CrossRef]
- Vaurie, C. The birds of the Palearctic fauna. Non Passeriformes. H.F. and G. Witherby, London, 1965.
- Ientile, R. Il Codibugnolo siciliano, Aegithalos caudatus siculus (Whitaker 1901): eco-etologia, morfologia e caratterizzazione genetica. Doctoral Thesis, on Evolutionary Biology, Università degli Studi di Catania, 2010. [Google Scholar]
- Lo Valvo, F. The Sicilian Long-tailed Tit, Aegithalos caudatus siculus Whitaker 1901, a subspecies not to be underestimated. In: Life on Islands. 1. Biodiversity in Sicily and surrounding islands. Studies dedicated to Bruno Massa; La Mantia, T.; Badalamenti, E.; Carapezza, A.; Lo Cascio, P.; Troia, A., Eds.; Danaus Ed., Palermo, 2020, pp. 349‒354.
- La Mantia, T.; Buscemi, I.; Mingozzi, T.; Massa, B. Data analysis of extinct and living Woodpeckers (Aves Picidae) in Sicily and Calabria (Southern Italy). Naturalista sicil, 2015, 39, 29‒49. [http://www.sssn.it/indice4.htm].
- Chamot-Rooke, N.; Rabaute, A.; Kreemer, C. Western Mediterranean Ridge mud belt correlates with active shear strain at the prism-backstop geological contact. Geology, 2005, 33, 861–864. [CrossRef]
- Axelrod, D.J. , Evolution and biogeography of madrean-tethyan sclerophyll vegetation. Ann. Missouri bot. Garden, 1975, 62, 280-334.
- Fauquette, S.; Suc, J.-P.; Bertini, A.; Popescu, S.-M.; Warny, S.; Bachiri Taoifiq, N.; Perez Villa, M.-J.; Chikhi, H.; Feddi, N.; Subally, D.; Clauzon, G.; Ferrier, J. How much did climate force the Messinian salinity crisis? Quantified climatic conditions from pollen records in the Mediterranean region. Palaeogeogr., Paleoclim., Palaeoecol, 2006, 238, 281‒301. [https://www.sciencedirect.com/science/article/abs/pii/S0031018206001970]. [CrossRef]
- Blondel, J. Synthesis: the history of forest bird avifaunas of the world. In: Biogeography and ecology of forest bird communities, Keast, A., Ed.; SPB Academic Publ., The Hague, Netherlands, 1990, pp. 371‒377.
- Keast, A. Distribution and origins of forest birds. In: Biogeography and ecology of forest bird communities, Keast, A., Ed.; SPB Academic Publ., The Hague, Netherlands, 1990, pp. 45‒59.
- Pons, A. The History of the Mediterranean Shrublands. In: Ecosystems of the World-Mediterranean Type Shrublands, Vol. 11, Di Castri, F.; Goodall, W.; Specht, R.L., Eds.; Elsevier, Amsterdam, 1981, pp. 131‒138.
- Grove, A.T. The Historical context: before 1850. In: Mediterranean Desertification and Land Use, Brandt, C.J.; Thornes, J.B., Eds.; J. Wiley and Sons, Chichester, England, 1996, pp. 13‒28.
- Shugart, H.H. Patterns and ecology of forests. In: Biogeography and ecology of forest bird communities, Keast, A., Ed.; SPB Academic Publ., The Hague, Netherlands, 1990, pp. 7‒26.
- Massa, B. Gli Uccelli della fauna italiana. XIX Sem. Accad. Naz. Lincei, 1993, 79‒86.
- Massa, B. Biogeografia degli uccelli nidificanti in Italia. In: Atlante degli Uccelli nidificanti in Italia, Lardelli, R.; Bogliani, G.; Caprio, E.; Celada, C.; Fraticelli, F.; Gustin, M.; Janni, O.; Pedrini, P.; Puglisi, L.; Rubolini, D.; Ruggieri, L.; Spina, F.; Tinarelli, R.; Calvi, G.; Brambilla, M., Eds.; Ed. Belvedere, Latina, 2022, pp. 37‒46.
- Le Houérou, H.-N. L'impact de l'homme et de ses animaux sur la forêt méditerranéenne. 1980, 2, 31‒44, 155‒174. [https://www.foret-mediterraneenne.org].
- Marchetti, M.; Bertani, R.; Corona, P.; Valentini, R. Cambiamenti di copertura forestale e dell’uso del suolo nell’inventario dell’uso delle terre in Italia. Forest, 2012, 9, 170–184. [http://www.sisef.it/forest@/contents/?id=efor0696-009].
- Telleria, J.J.; Santos, T. Factors involved in the distribution of forest birds in the Iberian Peninsula. Birds Study, 1994, 41, 161‒169. [https://www.researchgate.net/publication/233249779_Factors_involved_in_the_distribution_of_forest_birds_in_the_Ibenan_Pennsula].
- Gregory, R.; Vorisek, P.; Van Strien, A.; Gmelig Mayling, A.W.; Jiguet, F.; Fornasari, L.; Reif, J.; Chylarecki, P.; Burfield, I.J. Population trends of widespread woodland birds in Europe. Ibis, 2007, 149 (suppl. 2), 78–97. [CrossRef]
- Fuller, R.J.; Noble, D.G.; Smith, K.W.; Vanhinsbergh, D. Recent declines in populations of woodland birds in Britain: a review of possible causes. British Birds, 2005, 98, 116–143.
- Gil-Tena, A.; Brotons, L.; Saura, S. Mediterranean forest dynamics and forest bird distribution changes in the late 20th century. Global Change Biol, 2009, 15, 474‒485. [CrossRef]
- Avery, M.; Leslie, R. Birds and Forestry. T. & A.D. Poyser, London, 1990.
- Pepe, A.; Caverni, L.; Maluccio, S. Rete Rurale Nazionale RRN 2014-2020. The state of italian forests, executive summary. Ministero delle politiche agricole alimentari e forestali. 2020. [https://www. reterurale.it].
- Blondel, J. Biogeography and history of forest bird faunas in the Mediterranean zone. Biogeography and ecology of forest bird communities, Keast, A., Ed.; SPB Academic Publ., The Hague, Netherlands, 1990, pp. 95‒107.
- Covas, R.; Blondel, J. Biogeography and history of the Mediterranean bird fauna. Ibis, 2008, 140, 395‒407. [https://www.researchgate.net/publication/229859515_Biogeography_and_history_of_the_Mediterranean_bird_fauna].
- Gustin, M.; Nardelli, R.; Brichetti, P.; Battistoni, A.; Rondinini, C.; Teofili, C. (Eds.) ; Lista Rossa IUCN degli uccelli nidificanti in Italia 2019. Comitato Italiano IUCN e Ministero dell’Ambiente e della Tutela del Territorio e del Mare, Roma, 2019.
- La Mantia, T. Regolare nidificazione di Colombaccio, Columba palumbus, in gennaio in Sicilia. Riv. ital. Orn, 1994, 64, 77. [http://www.ornitologiasiciliana.it/bibliografia.php].
- Isenmann, P. Some recent bird invasions in Europe and the Mediterranean basin. In Biological Invasions in Europe and the Mediterranean Basin. Di Castri, F.; Hansen, A.J.; Debussche, M., Eds.; Kluwer Acad. Publ., Dordrecht, 1990, pp. 245‒261.
- Massa, B.; La Mantia, T. Forestry, pasture, agriculture and fauna correlated to recent changes in Sicily. Forest, 2007, 4 (4), 418‒438. [http://foresta.sisef.org/contents/?id=ifor0495-002Cached]. [CrossRef]
- La Mantia, T.; Bonaviri, L.; Massa, B. Ornithological communities as indicators of recent transformations on a regional scale: the case of the Sicily island. 2014, 38, 67‒81. [https://www.avocetta.org/articles/vol-38-2-xo-ornithological-communities-as-indicators-of-recent-transformations-on-a-regional-scale-sicilys-case/].
Figure 1.
Percentage of different categories of abundance (stable, increasing, decreasing, strongly decreasing, fluctuating) of 61 breeding forest birds in Italy, during the 150 years considered.
Figure 1.
Percentage of different categories of abundance (stable, increasing, decreasing, strongly decreasing, fluctuating) of 61 breeding forest birds in Italy, during the 150 years considered.
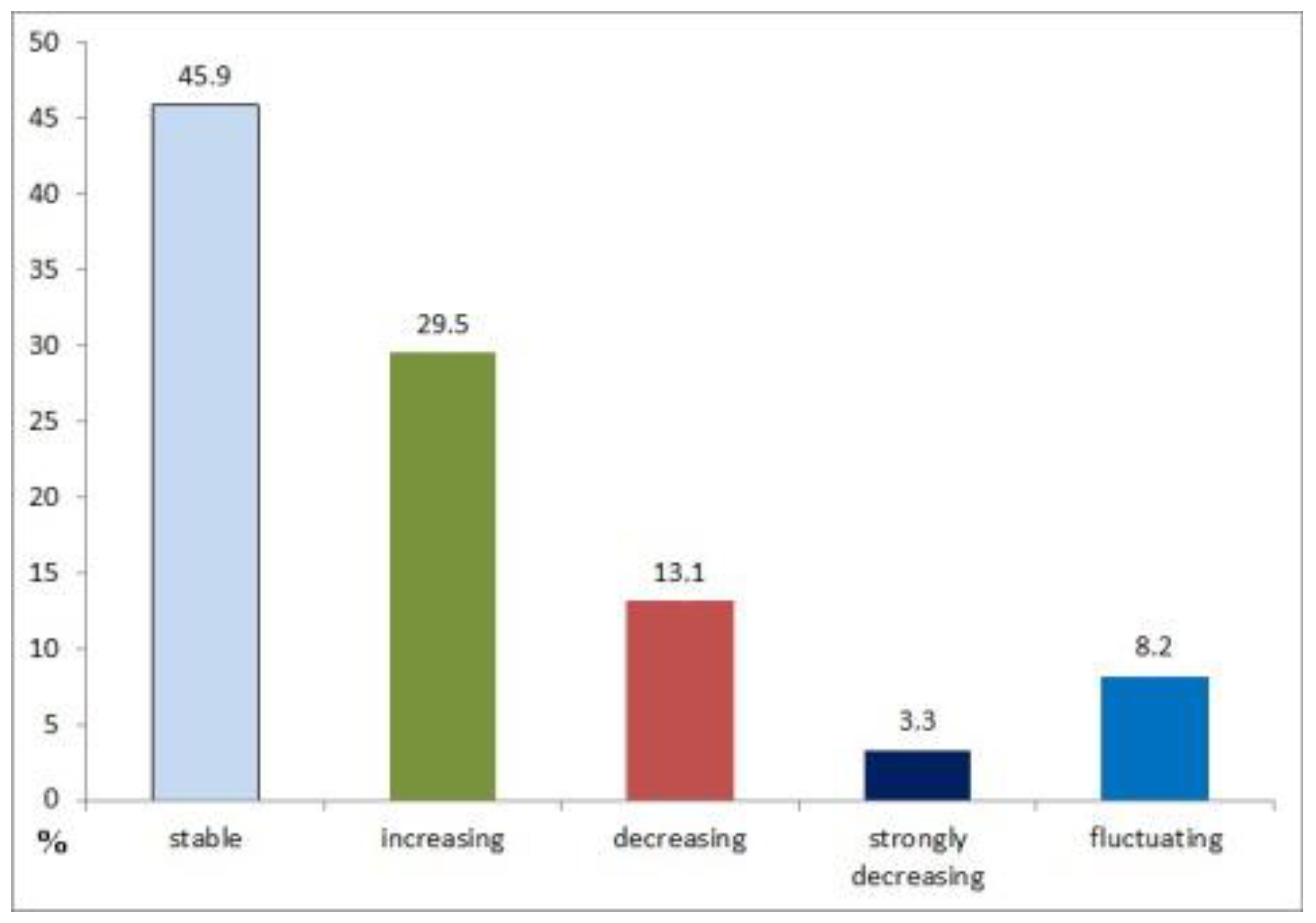
Figure 2.
Satellitar map of the Mediterranean area with the three peninsulas that characterize it; dark green areas correspond to those covered with forest (after Google Earth, modified).
Figure 2.
Satellitar map of the Mediterranean area with the three peninsulas that characterize it; dark green areas correspond to those covered with forest (after Google Earth, modified).
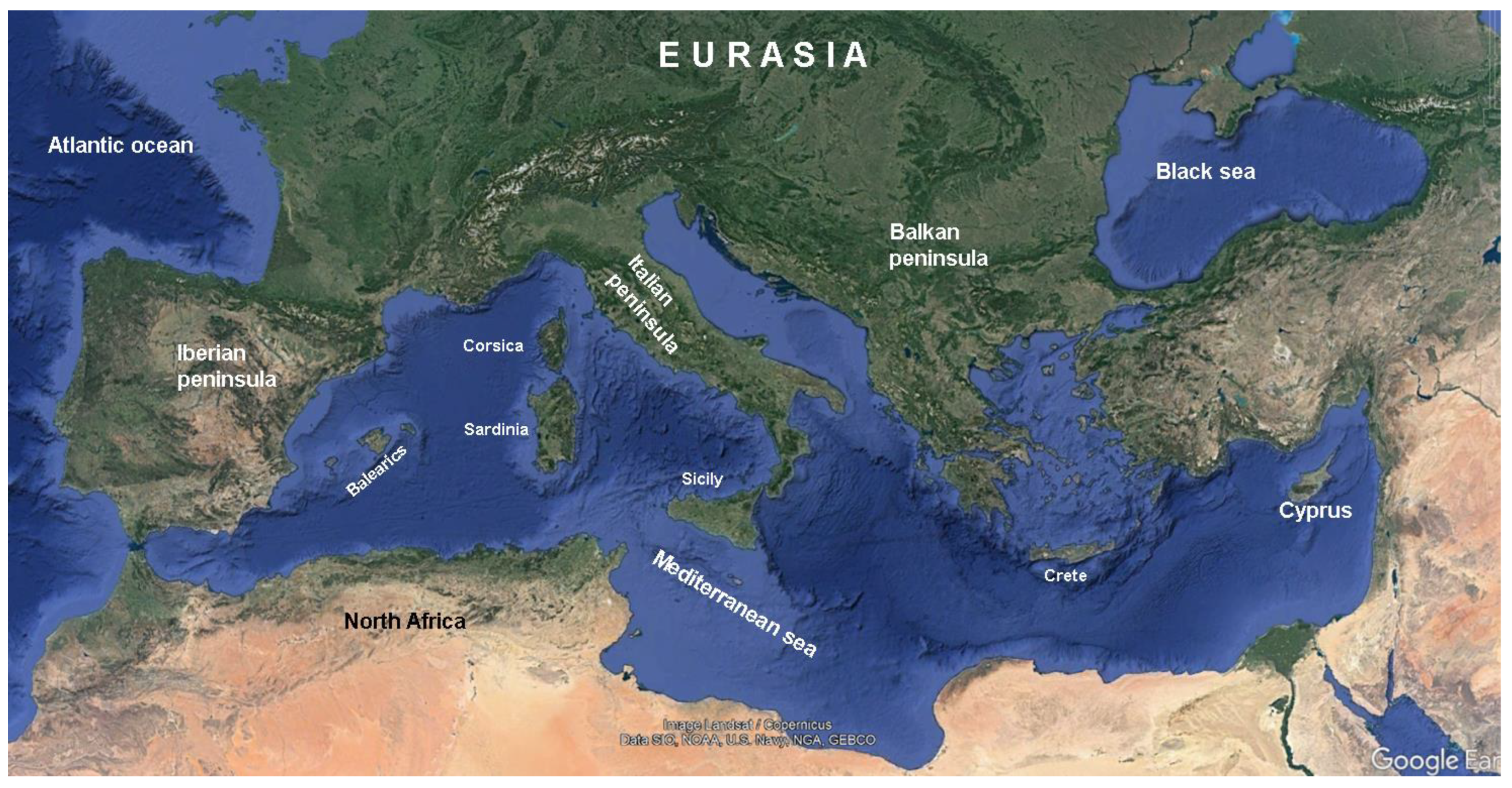
Figure 3.
Number of breeding forest birds in the three Mediterranean peninsulas.
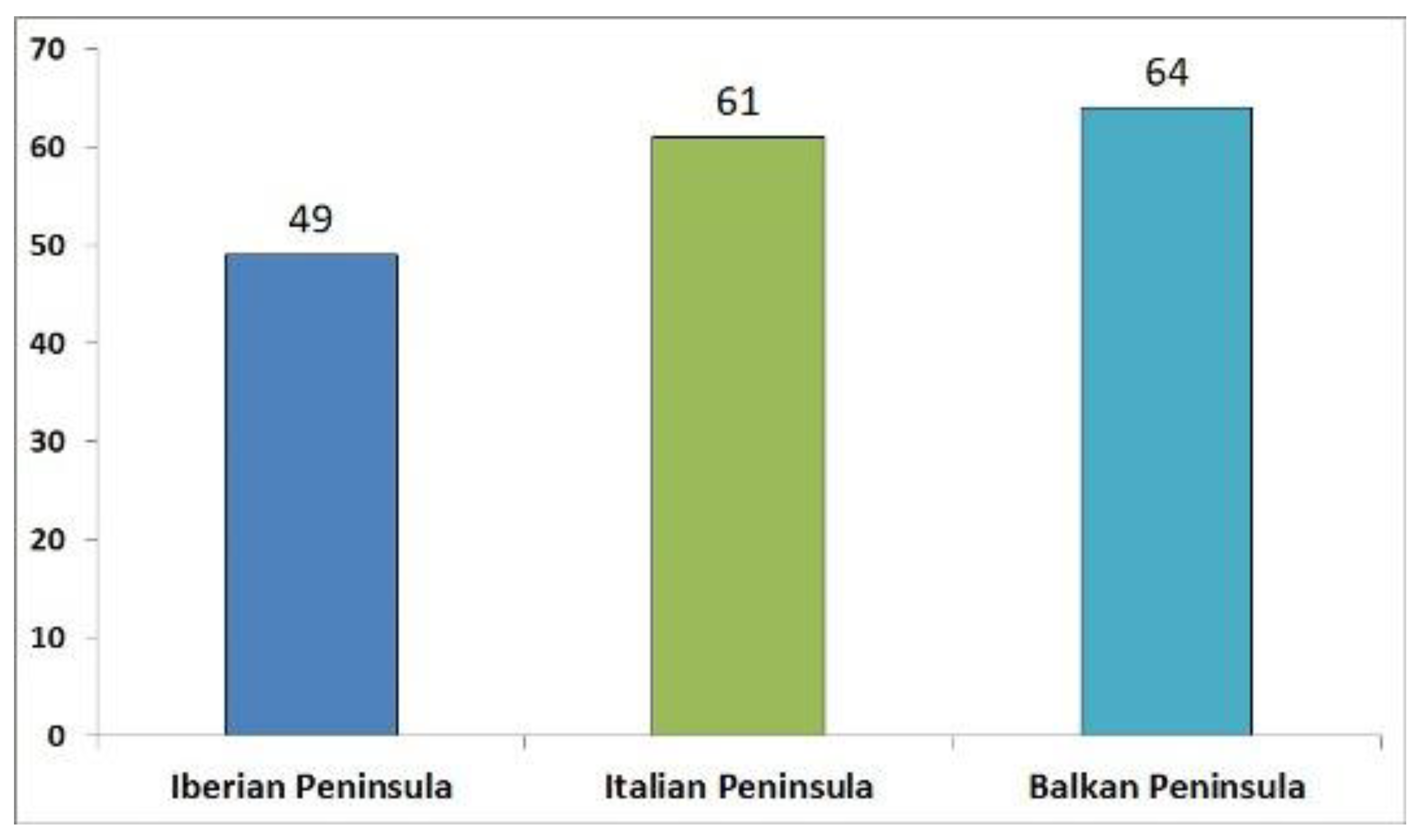
Figure 4.
Sørensen's similarity index between pairs of breeding forest birds of three Mediterranean peninsulas.
Figure 4.
Sørensen's similarity index between pairs of breeding forest birds of three Mediterranean peninsulas.
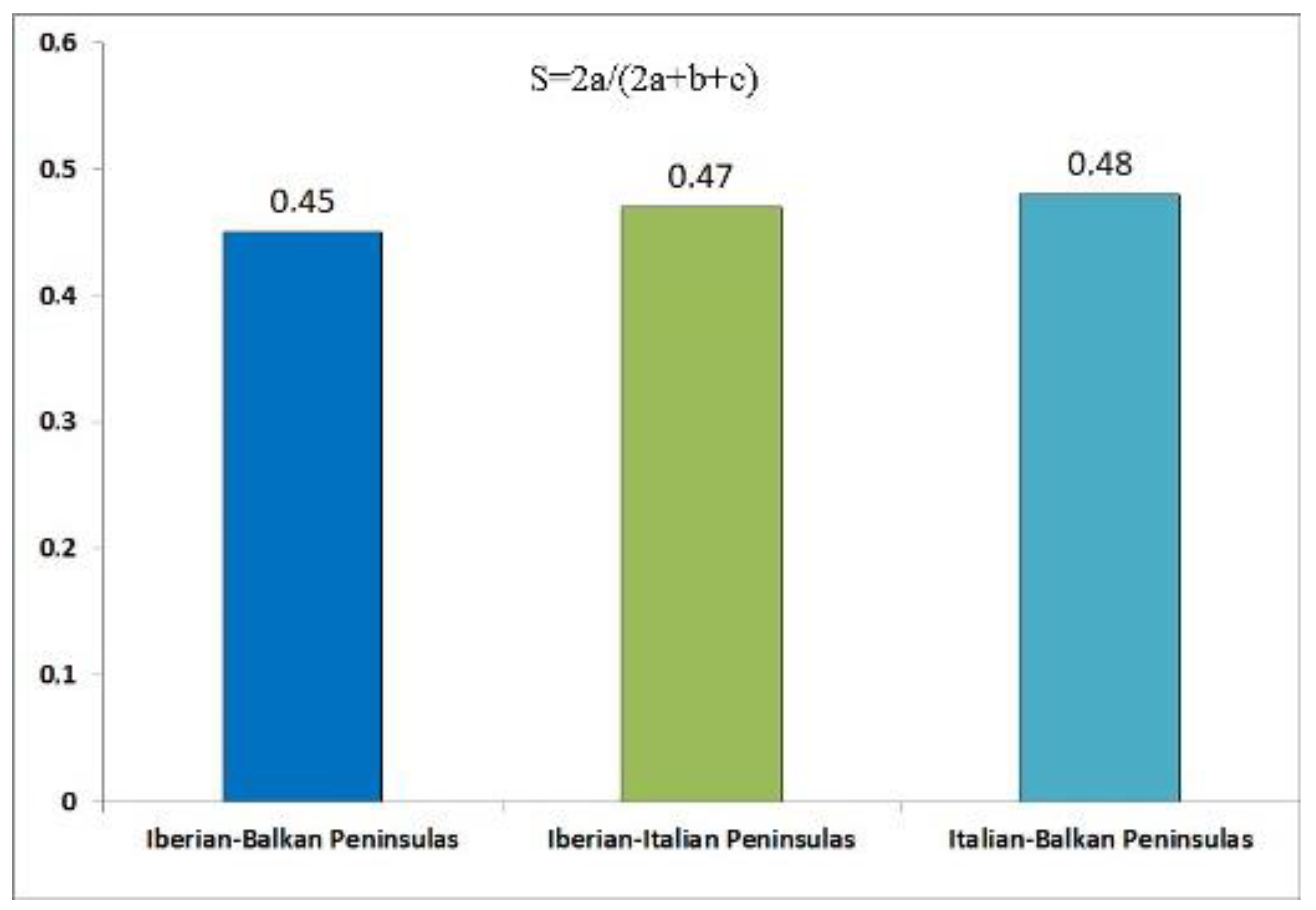
Figure 5.
Simulated trend of percentage of Italian breeding forest birds in the late XXI century according to Huntley et al. [31].
Figure 5.
Simulated trend of percentage of Italian breeding forest birds in the late XXI century according to Huntley et al. [31].
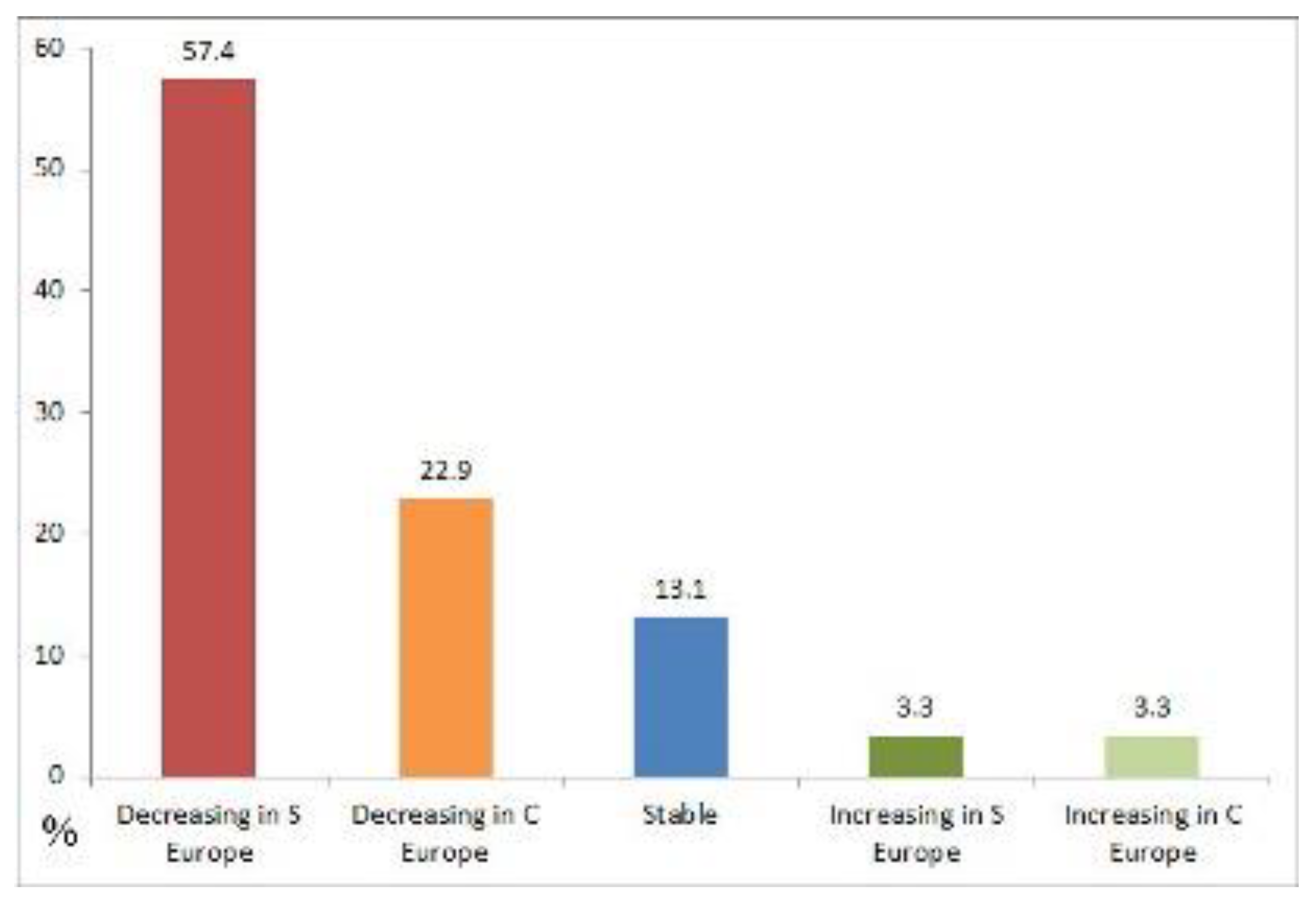
Figure 6.
Two species of woodpeckers found as far south as southern Italy (Calabria): black woodpecker (increasing) and middle spotted woodpecker (uncommon, but stable over a long period) (Photos by G. Congi, Calabria, Park of Sila).
Figure 6.
Two species of woodpeckers found as far south as southern Italy (Calabria): black woodpecker (increasing) and middle spotted woodpecker (uncommon, but stable over a long period) (Photos by G. Congi, Calabria, Park of Sila).

Figure 7.
The collared flycatcher is a trans-Saharan species breeding in the Apennines to Calabria, where Europe's southernmost nesting sites are found; it is an ecologically demanding bird of secondary cavities (produced by branch breakage or excavated by woodpeckers) found in mature trees, which then play a really important role in sustaining populations of these birds (Photo by G. Congi, Calabria, Park of Sila).
Figure 7.
The collared flycatcher is a trans-Saharan species breeding in the Apennines to Calabria, where Europe's southernmost nesting sites are found; it is an ecologically demanding bird of secondary cavities (produced by branch breakage or excavated by woodpeckers) found in mature trees, which then play a really important role in sustaining populations of these birds (Photo by G. Congi, Calabria, Park of Sila).

Figure 8.
The Eurasian siskin is generally linked to coniferous trees and recently it benefited of afforestation with exotic species; in the photo (by G. Congi) one siskin extracting seeds from cones of the native black pine in south Italy (Calabria, Park of Sila).
Figure 8.
The Eurasian siskin is generally linked to coniferous trees and recently it benefited of afforestation with exotic species; in the photo (by G. Congi) one siskin extracting seeds from cones of the native black pine in south Italy (Calabria, Park of Sila).

Figure 9.
Crossbill male photographed on Park of Sila (Calabria, South Italy); it is evident the massive bill and the discolored plumage compared to north European individuals (photo by G. Congi).
Figure 9.
Crossbill male photographed on Park of Sila (Calabria, South Italy); it is evident the massive bill and the discolored plumage compared to north European individuals (photo by G. Congi).

Figure 10.
The Sicilian subspecies of long-tailed tit Aegithalos caudatus siculus, which lives in oakwood, is clearly differentiated from the Italian peninsular populations; it has grey back, the head lacks the characteristic dark lateral bands, the scapulars are grey (not reddish), and its size on average is smaller than those of continental individuals (Photo by M. Coco).
Figure 10.
The Sicilian subspecies of long-tailed tit Aegithalos caudatus siculus, which lives in oakwood, is clearly differentiated from the Italian peninsular populations; it has grey back, the head lacks the characteristic dark lateral bands, the scapulars are grey (not reddish), and its size on average is smaller than those of continental individuals (Photo by M. Coco).

Figure 11.
Rock carvings of giraffes and elephants, dating back some seven thousand years, at Matkandush, in the Libyan Sahara (Fezzan), showing that what today is desert was a savannah only a few thousands of years ago.
Figure 11.
Rock carvings of giraffes and elephants, dating back some seven thousand years, at Matkandush, in the Libyan Sahara (Fezzan), showing that what today is desert was a savannah only a few thousands of years ago.

Figure 12.
Terracing in the islands of Cyprus (above) and Crete (below) carried out in the steep slopes of mountains to obtain arable soil and plant olive trees.
Figure 12.
Terracing in the islands of Cyprus (above) and Crete (below) carried out in the steep slopes of mountains to obtain arable soil and plant olive trees.

Figure 13.
Increase of wooded area in Italy from 1861 to 2015; 22% of the recent increase (since 1980’) is due to afforestation (after Agnoletti et al. [1], reprocessed).
Figure 13.
Increase of wooded area in Italy from 1861 to 2015; 22% of the recent increase (since 1980’) is due to afforestation (after Agnoletti et al. [1], reprocessed).
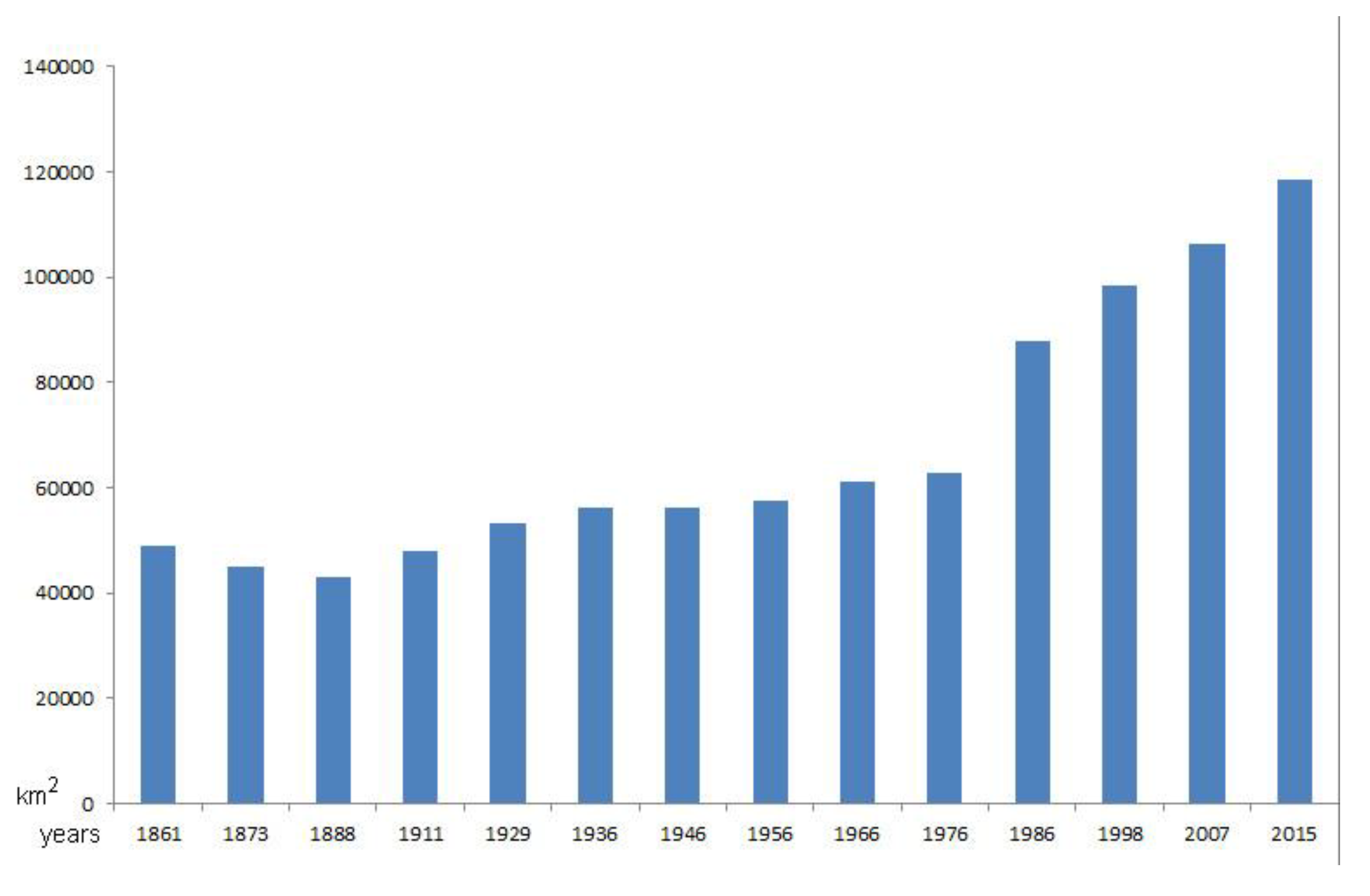
Figure 14.
Hectares of land interested by Italian fires between 2000 and 2023; the total surface of fires, mainly arson, which affected Italian land amounts to ca. 1,784,000 hectares, much covered by forests (after Pepe et al. [67], updated).
Figure 14.
Hectares of land interested by Italian fires between 2000 and 2023; the total surface of fires, mainly arson, which affected Italian land amounts to ca. 1,784,000 hectares, much covered by forests (after Pepe et al. [67], updated).
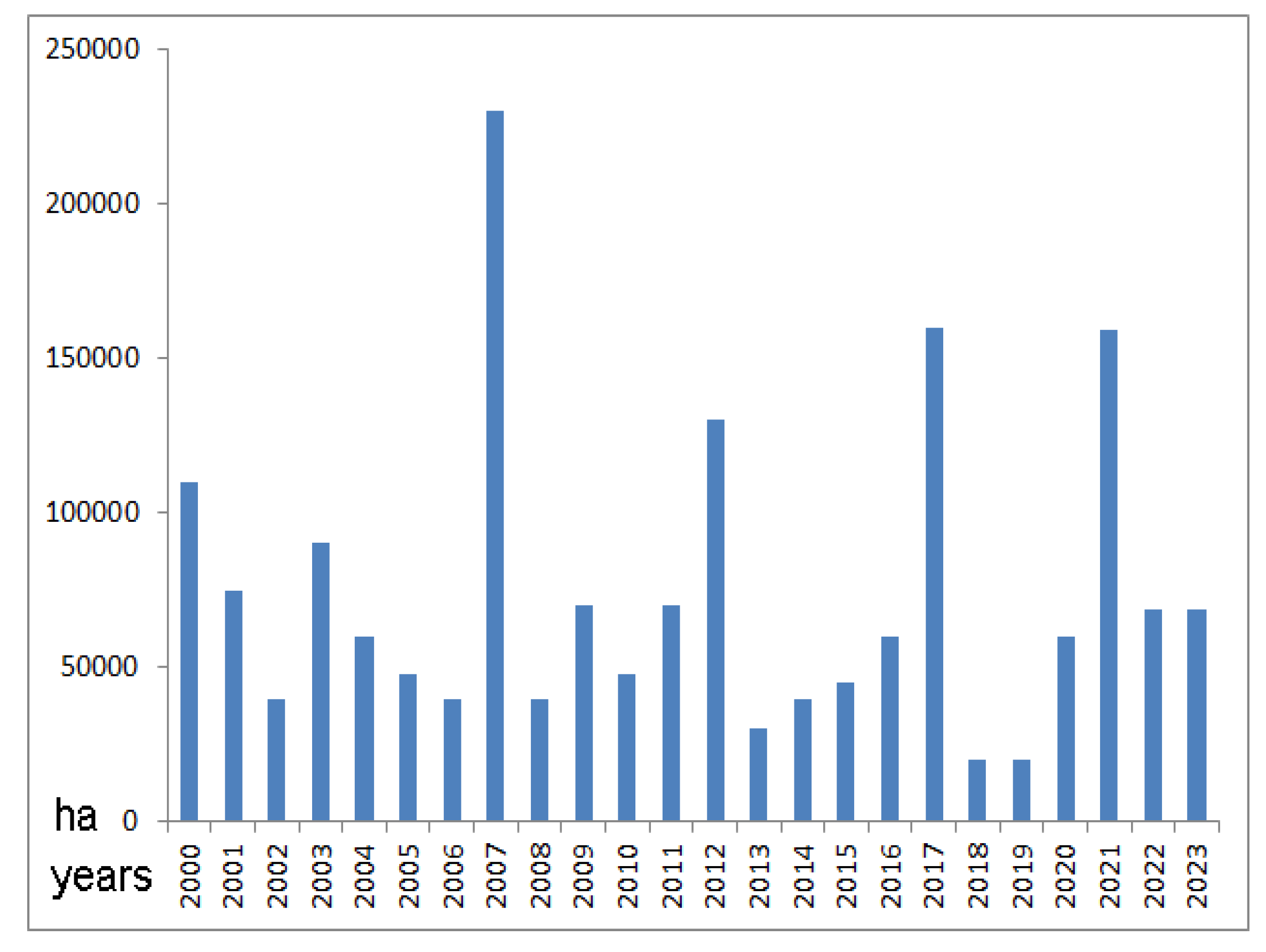
Figure 15.
Main threats forest birds will face in the coming decades: climate change, global warming, reduction of old-growth forests, increased fire, and CO2 emissions (drawing by V. Massa).
Figure 15.
Main threats forest birds will face in the coming decades: climate change, global warming, reduction of old-growth forests, increased fire, and CO2 emissions (drawing by V. Massa).


Table 1.
List of 61 Italian breeding forest birds since 1872 to 2022. The Category in the European Red List in the first column after BirdLife International [24]. At the beginning of the columns 2, 3, 4, 5 are reported the bibliographical references, from which the status was obtained. In the column 6 is reported the reliable trend through 150 years (stable, decreasing, strongly decreasing, increasing, fluctuating as breeding). In the column 7 the simulated trend in the late 21th century after the Climate Atlas, and in the column 8 the main habitat of each species are recorded. Trans-Saharan species are indicated with an asterisk (*).
Table 1.
List of 61 Italian breeding forest birds since 1872 to 2022. The Category in the European Red List in the first column after BirdLife International [24]. At the beginning of the columns 2, 3, 4, 5 are reported the bibliographical references, from which the status was obtained. In the column 6 is reported the reliable trend through 150 years (stable, decreasing, strongly decreasing, increasing, fluctuating as breeding). In the column 7 the simulated trend in the late 21th century after the Climate Atlas, and in the column 8 the main habitat of each species are recorded. Trans-Saharan species are indicated with an asterisk (*).
| Species (threat) | Salvadori [2] | Martorelli [3]; Giglioli [4]; Arrigoni degli Oddi [5] | Schenk [6]; Meschini and Frugis [7] | Brichetti and Fracasso [8,9,10,11,12,13,14,15,16,17,18]; Corso [19]; Gustin et al. [20]; Massa et al. [21]; Grussu [22]; Lardelli et al. [23] | Trend 1872-2022 | Huntley et al. [31], simulated trend in late XXI century | Habitat |
|---|---|---|---|---|---|---|---|
| Western capercaillie Tetrao urogallus (LC) | Sedentary in Alps, decreasing | Sedentary in Alps, very decreasing | Sedentary in CE Alps, decreasing | Sedentary in CE Alps, strongly decreasing | Strongly decreasing | Decreasing in C Europe | Coniferous mixed with broadleaved woods |
| Black grouse Lyrurus tetrix (LC) | Sedentary in Alps, fairly common | Sedentary in Alps, decreasing | Sedentary in Alps, decreasing | Sedentary in Alps, decreasing | Decreasing | Decreasing in C Europe | Coniferous woods with pastures |
| Hazel grouse Bonasa bonasia (LC) | Sedentary in Alps, decreasing | Sedentary in Alps, very decreasing | Sedentary in Alps, recently increasing | Sedentary in Alps, in recent westward expansion | Fluctuating | Decreasing in S Europe | Coniferous mixed with broadleaved woods and common hazel |
| Common woodpigeon Columba palumbus (LC) | Common sedentary breeder in all Italy | Common sedentary breeder in all Italy, also in parks of some towns | Common sedentary breeder in all Italy, also in urban parks, increasing | Sedentary and migrant breeder, increasing in all Italian regions | Increasing | Stable | Coniferous and broadleaved woods |
| Stock dove Columba oenas (LC) | Common passage migrant, rare as breeding | Scarce in all Italy as breeding, more common as migrant and wintering | Very scarce migrant and rare breeder in scattered sites of Italian peninsula and Sicily | Migrant breeder fluctuating, disappearing in C-S Italy, breeding only in NW Italy | Strongly decreasing | Decreasing in S Europe | Coniferous and broadleaved woods |
| European nightjar Caprimulgus europaeus (LC) | Migrant breeder in all Italy | Migrant breeder in all Italy | Migrant breeder in all Italy | Migrant breeder in all Italy, increasing | Increasing | Decreasing in S Europe | Wooded areas with clearings |
| Eurasian woodcock Scolopax rusticola (LC) | Common wintering, some pairs breed in N Italy | Common migrant wintering, rare breeder in Tuscany, Piedmont, Veneto, Lombardy, Trentino | Rare migrant breeder in N Italy, not confirmed in Tuscany, common wintering | Rare migrant breeder in Alps and N Apennines, common wintering | Stable | Decreasing in S Europe | Broadleaved and coniferous woods |
| Eurasian eagle-owl Bubo bubo (LC) | All Italian peninsula and Sicily | Sedentary in all Italian peninsula and Sicily, decreasing | Sedentary in Italian peninsula, decreasing, extinct in Sicily | Sedentary in Italian peninsula, probably fluctuating | Decreasing | Decreasing in S Europe | Cliffs next to woods |
| Boreal owl Aegolius funereus (LC) | Accidental | Very rare | Sedentary breeder in Alps | Sedentary breeder in Alps | Increasing | Decreasing in C Europe | Pure and mixed coniferous woods |
| Eurasian pygmy-owl Glaucidium passerinum (LC) | Accidental | Rare breeding in E Alps | Rare sedentary breeder in Alps | Scarce sedentary breeder in Alps | Stable | Decreasing in C Europe | Uneven-aged coniferous woods |
| Northern long-heared owl Asio otus (LC) | Common in Italian peninsula | Rare breeder in N-C Italy and Sicily | Scarce migrant breeder in N Italy, rarer in C-S Italy and Sicily, wintering | Scarce migrant and sedentary breeder in N Italy, C-S Italy, Sardinia and Sicily, wintering | Increasing | Decreasing in S Europe | Coniferous and broadleaved woods |
| Tawny owl Strix aluco (LC) | Common in Italian peninsula and Sicily | Common sedentary in Italian peninsula and Sicily | Common sedentary breeder in all Italian peninsula and Sicily | Common sedentary breeder in all Italian peninsula and Sicily | Stable | Decreasing in S Europe | Broadleaved and coniferous woods |
| Ural owl Strix uralensis (LC) | Accidental | Very rare | Unrecorded as breeding | First breeding in 1994 (Udine), later increasing westwards | Increasing | Decreasing in C Europe | Beech and alpine coniferous woods |
| European honey-buzzard Pernis apivorus (LC) | Uncommon migrant | Uncommon migrant breeder in mounts over Po Valley, Tuscany, etc., decreasing | Uncommon migrant breeder in Italian peninsula, mainly in N Italy | Uncommon migrant breeder in Italian peninsula, mainly in N Italy | Stable | Decreasing in S Europe | Broadleaved and coniferous woods |
| Short-toed snake-eagle Circaetus gallicus (LC)* | Common migrant breeder in Latium and Tuscany, rarely wintering | Common sedentary breeder from Alps to islands | Migrant breeder in Italian peninsula, mainly NW Italy | Migrant breeder in Italian peninsula, absent in islands | Increasing | Increasing in C Europe | Broadleaved and coniferous woods with clearings |
| Booted eagle Hieraetus pennatus (LC) | Migrant, very rare | Migrant, very rare | Unrecorded as breeding | Migrant and wintering, common in some regions, breeding in scattered sites | Increasing | Increasing in S Europe | Broadleaved and coniferous woods |
| Eurasian sparrowhawk Accipiter nisus (LC) | Breeding in N Italy | Common breeder in all Italy | Common sedentary breeder in all Italy | Sedentary breeder, increasing in all Italy since mid ‘1980 | Increasing | Decreasing in S Europe | Broadleaved and coniferous woods |
| Northern goshawk Accipiter gentilis (LC) | Uncommon in Sardinia and Italian peninsula | Uncommon breeder in Tuscany, Abruzzo, Molise, Alps, common in Sardinia | Sedentary breeder in Italian peninsula, mainly N Italy, and Sardinia | Sedentary breeder in all Italian peninsula and Sardinia | Stable | Decreasing in S Europe | Broadleaved and coniferous woods |
| Eurasian buzzard Buteo buteo (LC) | Very common in all Italy | Very common breeder in all Italy | Common sedentary breeder in all Italy | Common sedentary breeder in all Italy | Stable | Decreasing in S Europe | Broadleaved and coniferous woods |
| Black woodpecker Dryocopus martius (LC) | Alps, central Apennines and Sicily (rare) | Sedentary in Alps, to be confirmed in Apennines and Sicily | Sedentary in Alps and scattered zones of C-S Apennines, extinct in Sicily (1900) | Sedentary in Alps and scattered zones of C-S Apennines, accidental in Sicily | Increasing | Decreasing in S Europe | Mature broadleaved and coniferous woods |
| Grey-faced woodpecker Picus canus (LC) | Sedentary in Alps | Sedentary in Alps | Sedentary breeder in C-E Alps | Sedentary breeder in C-E Alps | Stable | Decreasing in C Europe | Broadleaved and coniferous woods |
| Eurasian green woodpecker Picus viridis (LC) | Very common, rare in Sicily, never found in Sardinia | Common sedentary breeder, rare in Sicily, absent in Sardinia | Sedentary breeder in all Italy, extinct in Sicily (1930) | Sedentary breeder in the peninsula, increasing | Increasing | Decreasing in S Europe | Broadleaved and coniferous woods |
| Three-toed woodpecker Picoides tridactylus (LC) | Alps (rare) | Accidental | Very local sedentary breeder in E Alps | Very local sedentary breeder in C-E Alps | Stable | Decreasing in C Europe | Coniferous woods |
| Lesser spotted woodpecker Dryobates minor (LC) | All Italy, but no abundant | Local in all Italy, including Sicily and Sardinia | Uncommon sedentary breeder in W Alps and Apennines, extinct in Sicily (1930), no data in Sardinia from 1980 1 | Sedentary breeder in all Italian peninsula, increasing | Increasing | Decreasing in S Europe | Broadleaved and coniferous woods |
| Middle spotted woodpecker Leiopicus medius (LC) | Central Italy (rare) | Sedentary in mounts over Po Valley, Umbria, Tuscany, Abruzzo, Latium and Sicily 2 | Sedentary breeder in scattered sites of C-S Apennines | Sedentary breeder in scattered sites of C-S Apennines, colonized Friuli | Stable | Decreasing in S Europe | Broadleaved woods |
| Great spotted woodpecker Dendrocopos major (LC) | Common in all Italy | Sedentary breeder in all Italy, decreasing for deforestation | Sedentary breeder in all Italy, decreasing for deforestation | Sedentary breeder in all Italy, increasing | Increasing | Decreasing in S Europe | Broadleaved and coniferous woods |
| White-backed woodpecker Dendrocopos leucotos (LC) | East Alps | Accidental, breeder in Abruzzo | Sedentary in C Apennines and Gargano (Apulia) | Sedentary in C Apennines, unconfirmed in Apulia | Stable | Decreasing in S Europe | Broadleaved woods |
| Eurasian hobby Falco subbuteo (LC)* | Migrant, breeding not confirmed | Migrant, breeding not confirmed | Migrant breeder in all Italy | Migrant breeder in all Italy, increasing | Increasing | Decreasing in S Europe | Edge of broadleaved and coniferous woods |
| Eurasian jay Garrulus glandarius (LC) | Common breeder in all Italy | Common breeder in all Italy | Sedentary breeder in all Italy | Sedentary breeder in all Italy, increasing in urban habitats | Increasing | Stable | Broadleaved and coniferous woods |
| Northern nutcracker Nucifraga caryocatactes (LC) | Uncommon breeder in Alps | Uncommon breeder in Alps, mainly Trentino, Veneto, Lombardy | Sedentary breeder in Alps | Sedentary breeder in Alps | Stable | Decreasing in C Europe | Alpine woods of Pinus cembra |
| Coal tit Periparus ater (LC) | Uncommon breeder in all Italy | Uncommon breeder in montane areas of Italy | Sedentary breeder in all Italy | Sedentary breeder in all Italy, increasing | Increasing | Decreasing in S Europe | Coniferous and broadleaved woods |
| Crested tit Lophophanes cristatus (LC) | Breeding in Alps | Breeding in Alps | Sedentary breeder in Alps | Sedentary breeder in Alps and N Apennines, increasing | Increasing | Decreasing in S Europe | Coniferous woods, also mixed with broadleaved trees |
| Marsh tit Poecile palustris (LC) | Uncommon breeder in all Italy, unrecorded in Sardinia | Uncommon breeder in all Italy, vagrant in Sardinia | Sedentary breeder in Italian peninsula and local in Sicily | Sedentary breeder in Italian peninsula and local in Sicily | Stable | Decreasing in S Europe | Mature broadleaved and coniferous woods |
| Willow tit Poecile montanus (LC) | Breeding in Alps | Breeding in Alps | Sedentary breeder in Alps and C Apennines 3 | Sedentary breeder in Alps | Stable | Decreasing in C Europe | Montane coniferous woods |
| Great tit Parus major (LC) | Very common breeder in all Italy | Very common sedentary breeder in all Italy | Sedentary breeder in all Italy | Sedentary breeder in all Italy | Stable | Stable | Woods and wooded areas |
| Eurasian blue tit Cyanistes caeruleus (LC) | Common breeder in all Italy | Common sedentary breeder in all Italy | Sedentary breeder in all Italy | Sedentary breeder in all Italy | Stable | Stable | Broadleaved and coniferous woods |
| Western Bonelli’s warbler Phylloscopus bonelli (LC)* | Breeding? | Migrant breeder, more in N Italy | Migrant breeder in Alps and Apennines, rare in S Italy | Migrant breeder in Alps, Apennines and Sardinia, rare in S Italy | Fluctuating | Increasing in C Europe | Coniferous and broadleaved woods |
| Common chiffchaff Phylloscopus collybita (LC) | Breeding in all Italy | Common migrant and sedentary breeding in all Italy, wintering south of Tuscany | Migrant and sedentary breeder in Italian peninsula and Sicily | Migrant and sedentary breeder in Italian peninsula and Sicily, colonized Sardinia | Increasing | Decreasing in S Europe | Broadleaved and coniferous woods |
| Wood warbler Phylloscopus sibilatrix (LC)* | Migrant breeder in montane areas | Migrant breeder, rare in Sardinia and Apulia | Migrant breeder in Alps and Apennines | Migrant breeder in Alps, Apennines and possible breeder in Apulia | Stable | Decreasing in C Europe | Broadleaved woods |
| Long-tailed tit Aegithalos caudatus (LC) | Very common breeder in all Italy, unrecorded in Sardinia | Common breeder in all Italy, absent in Sardinia | Sedentary breeder in Italian peninsula and Sicily | Sedentary breeder in Italian peninsula and Sicily | Stable | Decreasing in S Europe | Broadleaved and coniferous woods |
| Garden warbler Sylvia borin (LC)* | Rare migrant breeder in C-N Italy | Migrant breeder in montane areas, Po Valley, C Italy | Migrant breeder in N Italy, a single population in C Apennines | Migrant breeder in Alps and N Apennines, decreasing | Decreasing | Decreasing in S Europe | Woods with undergrowth |
| Eurasian blackcap Sylvia atricapilla (LC) | Very common sedentary breeder in all Italy | Very common sedentary breeder in all Italy | Sedentary breeder in all Italy | Sedentary breeder in all Italy | Stable | Decreasing in S Europe | Broadleaved and coniferous woods, gardens |
| Barred warbler Sylvia nisoria (LC)* | Rare migrant breeder in N-E Italy | Local migrant breeder in the Po Valley | Local migrant breeder in N Italy | Local migrant breeder in Lombardy and Friuli, decreasing | Decreasing | Stable | Thermophilic bushy woods |
| Western orphean warbler Sylvia hortensis (LC)* | Migrant breeder in montane areas of N Italy | Migrant breeder in hillsides of N Italy | Migrant breeder in scattered sites of all Italian peninsula | Migrant breeder in scattered sites of all Italian peninsula, decreasing | Decreasing | Stable | Edges of woods |
| Short-toed treecreeper Certhia brachydactyla (LC) | Common sedentary breeder in all Italy, unrecorded in Sardinia | Common sedentary breeder in all Italy, absent in Sardinia | Sedentary breeder in Italian peninsula and Sicily | Sedentary breeder in Italian peninsula and Sicily | Stable | Decreasing in S Europe | Mature broadleaved and coniferous woods |
| Eurasian treecreeper Certhia familiaris (LC) | Breeding in Alps | Breeding in Alps and probably in Apennines | Sedentary breeder in Alps and C Apennines | Sedentary breeder in Alps and C Apennines | Stable | Decreasing in C Europe | Mature coniferous woods |
| Eurasian nuthatch Sitta europaea (LC) | Common in all Italy, unrecorded in Sardinia | Common in all Italy, absent in Sardinia | Sedentary breeder in Italian peninsula and Sicily | Sedentary breeder in Italian peninsula and Sicily | Stable | Decreasing in S Europe | Mature broadleaved and coniferous woods |
| Northern wren Troglodytes troglodytes (LC) | Common breeder in all Italy | Common breeder in all Italy, mainly in montane and hilly areas | Sedentary breeder in all Italy | Sedentary breeder in all Italy | Stable | Stable | Woods with undergrowth |
| Fieldfare Turdus pilaris (LC) | Breeding in Alps | Breeding in Alps? | Partially sedentary and migrant breeder in Alps | Partially sedentary and migrant breeder in Alps, declining | Decreasing | Decreasing in C Europe | Coniferous woods |
| Mistle thrush Turdus viscivorus (LC) | Common breeder in all Italy | Common breeder in all Italy | Sedentary breeder in all Italy | Sedentary breeder in all Italy | Stable | Decreasing in S Europe | Broadleaved and coniferous woods |
| Song thrush Turdus philomelos (LC) | Uncommon breeder in N Italy | Uncommon breeder from Alps to Apennines | Migrant breeder in Alps and Apennines | Migrant breeder in Alps and Apennines | Stable | Decreasing in S Europe | Broadleaved and coniferous woods |
| Ring ouzel Turdus torquatus (LC) | Rare breeder in C-N Italy | Rare breeder in Alps and Apennines | Migrant breeder in Alps and scattered sites of C Apennines | Migrant breeder in Alps and N Apennines, not confirmed in C Apennines | Decreasing | Decreasing in S Europe | Coniferous woods with bushy environments |
| European robin Erithacus rubecula (LC) | Very common breeder in all Italy | Very common breeder in montane areas of all Italy | Migrant breeder in all Italy | Abundant in all Italy, locally increasing | Increasing | Decreasing in S Europe | Woods with abundant undergrowth |
| Collared flycatcher Ficedula albicollis (LC)* | Rare migrant breeder | Local migrant breeding in Veneto, Lombardy, Piedmont, Liguria, Tuscany, Latium, Molise and probably Calabria | Local migrant breeder in Alps, N Italy, C-S Apennines, mainly in Abruzzo, Molise and Calabria | Local migrant breeder in Alps, N Italy, C-S Apennines, fluctuating | Fluctuating | Decreasing in S Europe | Mature broadleaved woods |
| Goldcrest Regulus regulus (LC) | Scarce breeder in all Italy | Scarce breeder in Italian peninsula and Sardinia | Sedentary breeder in Alps and Apennines, wintering in S Italy, unrecorded in Sardinia | Sedentary breeder in Alps and Apennines, wintering in S Italy, absent in Sardinia and Sicily | Stable | Decreasing in C Europe | Coniferous woods with undergrowth |
| Common firecrest Regulus ignicapilla (LC) | Scarce in all Italy | Scarce in all Italy | Sedentary breeder in all Italy | Sedentary breeder in all Italy | Stable | Decreasing in S Europe | Coniferous and broadleaved woods with undergrowth |
| Common chaffinch Fringilla coelebs (LC) | Very common wintering and breeder in all Italy | Very common wintering and breeder in all Italy | Migrant and sedentary breeder in all Italy | Migrant and sedentary breeder in all Italy, fluctuating, locally decreasing | Decreasing | Stable | Coniferous and broadleaved woods |
| Hawfinch Coccothraustes coccothraustes (LC) | Uncommon breeder in C-N Italy | Uncommon breeder in montane areas, common in Sardinia | Sedentary and migrant breeder in scattered sites of Italian peninsula and Sardinia | Sedentary and migrant breeder in scattered sites of Italian peninsula and Sardinia | Fluctuating | Decreasing in S Europe | Broadleaved woods |
| Red crossbill Loxia curvirostra (LC) | Irregularly breeder in C-N Italy | Sedentary breeder in C-N Italy | Sedentary and migrant breeder in Alps, scattered sites of Apennines and Sicily | Sedentary and migrant breeder in Alps, Apennines, Sardinia and Sicily | Fluctuating | Decreasing in S Europe | Coniferous woods |
| Eurasian bullfinch Pyrrhula pyrrhula (LC) | Uncommon breeder in beech woods of C-N Italy | Uncommon breeder in beech woods of C-N Italy | Sedentary breeder in Alps and Apennines | Sedentary breeder in Alps and Apennines | Stable | Decreasing in S Europe | Broadleaved and coniferous woods |
| Eurasian siskin Spinus spinus (LC) | Breeding in N Italy? | Migrant, some pairs breed in Alps | Sedentary and migrant breeder in Alps, S Apennines and Sardinia | Sedentary and migrant breeder in Alps, S Apennines, Sicily, Sardinia (?) | Stable | Decreasing in S Europe | Coniferous woods, also afforestation |
Table 2.
Main habitat and ecological niche of Italian forest bird species belonging to 20 families.
| Ecological niche | ||||
|---|---|---|---|---|
| Family | Habitat | Trees | Soil (litter) | Undergrowth |
| Phasianidae (3 species) | Coniferous mixed with broadleaved woods, coniferous woods with pastures and common hazel | Feeding | Nesting | Feeding |
| Columbidae (2 species) | Coniferous and broadleaved woods, with clearings | Feeding and nesting | Feeding | Feeding |
| Caprimulgidae (1 species) | Wooded areas with wide clearings and shrubs (aeroplanktophagous species) | Perching | Nesting | Feeding |
| Scolopacidae (1 species) | Broadleaved and coniferous woods with pastures at the edge | Nesting and feeding | ||
| Strigidae (6 species) | Cliffs next to woods, broadleaved and coniferous woods, pure and mixed coniferous woods, uneven-aged coniferous woods, beech and alpine coniferous woods, generally with big trees and holes for nesting | Nesting | Feeding | |
| Accipitridae (6 species) | Broadleaved and coniferous woods with clearings | Nesting | Feeding | |
| Picidae (8 species) | Mature broadleaved and coniferous woods with clearings | Nesting and feeding | Feeding | |
| Falconidae (1 species) | Edge of broadleaved and coniferous woods | Nesting | Feeding | |
| Corvidae (2 species) | Broadleaved and coniferous woods, alpine woods of Pinus cembra (nutcracker) | Nesting | Feeding | Feeding |
| Paridae (6 species) | Mature broadleaved and coniferous woods, also mixed with clearings | Nesting and feeding | Feeding | Feeding |
| Phylloscopidae (3 species) | Broadleaved and coniferous woods with clearings | Nesting and feeding | Feeding | |
| Aegithalidae (1 species) | Broadleaved and coniferous woods | Nesting and feeding | Feeding | |
| Sylviidae (4 species) | Woods with undergrowth, edges of woods, thermophilic bushy woods (barred warbler) | Nesting and feeding | Feeding | Nesting and feeding |
| Certhiidae (2 species) | Mature broadleaved and coniferous woods | Nesting and feeding | ||
| Sittidae (1 species) | Mature broadleaved and coniferous woods | Nesting and feeding | ||
| Troglodytidae (1 species) | Woods with undergrowth | Nesting | Feeding | Nesting and feeding |
| Turdidae (4 species) | Broadleaved and coniferous woods, coniferous woods with bushy environments | Nesting and feeding | Feeding | Feeding |
| Muscicapidae (2 species) | Woods with abundant undergrowth, mature broadleaved woods | Nesting and feeding | Feeding | Nesting and feeding |
| Regulidae (2 species) | Coniferous and broadleaved woods with undergrowth | Nesting and feeding | ||
| Fringillidae (5 species) | Coniferous and broadleaved woods | Nesting and feeding | Feeding | Feeding |
Table 3.
List of breeding forest bird species in the three Mediterranean peninsulas. * = trans-Saharan species; X = present. Ecological values of species: 1: of broad ecological value; 2: moderately ecologically demanding; 3: very ecologically demanding.
Table 3.
List of breeding forest bird species in the three Mediterranean peninsulas. * = trans-Saharan species; X = present. Ecological values of species: 1: of broad ecological value; 2: moderately ecologically demanding; 3: very ecologically demanding.
| Iberian peninsula | Italian peninsula | Balkan peninsula | |
|---|---|---|---|
| Tetrao urogallus (3) | X | X | X |
| Lyrurus tetrix (3) | X | X | |
| Bonasa bonasia (3) | X | X | |
| Columba oenas (3) | X | X | X |
| Columba palumbus (1) | X | X | X |
| Caprimulgus europaeus (2) | X | X | X |
| Scolopax rusticola (3) | X | X | X |
| Bubo bubo (2) | X | X | X |
| Strix aluco (1) | X | X | X |
| Strix uralensis (3) | X | X | |
| Glaucidium passerinum (3) | X | X | |
| Aegolius funereus (3) | X | X | |
| Asio otus (1) | X | X | X |
| Buteo rufinus (2) | X | ||
| Buteo buteo (1) | X | X | X |
| Circaetus gallicus* (2) | X | X | X |
| Hieraetus pennatus (3) | X | X | X |
| Pernis apivorus* (2) | X | X | X |
| Accipiter nisus (2) | X | X | X |
| Accipiter gentilis (2) | X | X | X |
| Dryobates minor (2) | X | X | X |
| Leiopicus medius (3) | X | X | X |
| Dendrocopos leucotos (3) | X | X | X |
| Dendrocopos major (1) | X | X | X |
| Dendrocopos syriacus (1) | X | ||
| Picoides tridactylus (3) | X | X | |
| Dryocopus martius (2) | X | X | X |
| Picus sharpei (3) | X | ||
| Picus viridis (2) | X | X | |
| Picus canus (3) | X | X | |
| Falco subbuteo* (2) | X | X | X |
| Garrulus glandarius (1) | X | X | X |
| Nucifraga caryocatactes (3) | X | X | |
| Poecile lugubris (3) | X | ||
| Poecile palustris (2) | X | X | X |
| Poecile montanus (3) | X | X | |
| Periparus ater (2) | X | X | X |
| Lophophanes cristatus (2) | X | X | X |
| Parus major (1) | X | X | X |
| Cyanistes caeruleus (1) | X | X | X |
| Phylloscopus trochilus (2) | X | ||
| Phylloscopus collybita (2) | X | X | X |
| Phylloscopus ibericus* (3) | X | ||
| Phylloscopus bonelli* (3) | X | X | |
| Phylloscopus sibilatrix* (2) | X | X | |
| Aegithalos caudatus (1) | X | X | X |
| Sylvia atricapilla (1) | X | X | X |
| Sylvia borin* (2) | X | X | X |
| Sylvia nisoria* (3) | X | X | |
| Sylvia hortensis* (3) | X | X | |
| Certhia familiaris (2) | X | X | X |
| Certhia brachydactyla (1) | X | X | X |
| Sitta europaea (2) | X | X | X |
| Sitta krueperi (2) | X | ||
| Troglodytes troglodytes (1) | X | X | X |
| Turdus torquatus (3) | X | X | X |
| Turdus pilaris (2) | X | X | |
| Turdus philomelos (2) | X | X | X |
| Turdus viscivorus (2) | X | X | X |
| Ficedula albicollis* (3) | X | X | |
| Erithacus rubecula (1) | X | X | X |
| Regulus regulus (2) | X | X | X |
| Regulus ignicapilla (1) | X | X | X |
| Fringilla coelebs (2) | X | X | X |
| Loxia curvirostra (2) | X | X | X |
| Spinus spinus (2) | X | X | X |
| Pyrrhula pyrrhula (2) | X | X | X |
| Coccothraustes coccothraustes (3) | X | X | X |
| Total No. of species | 49 | 61 | 64 |
| Sum and mean of ecological value of species | 96, 2.0 | 130, 2.1 | 134, 2.1 |
Table 4.
Number of breeding forest birds in the three Mediterranean peninsulas and Sørensen's similarity index [S = 2a/(2a+b+c)] between pairs of peninsulas.
Table 4.
Number of breeding forest birds in the three Mediterranean peninsulas and Sørensen's similarity index [S = 2a/(2a+b+c)] between pairs of peninsulas.
| Iberian peninsula | Italian peninsula | Balkan peninsula | |
|---|---|---|---|
| Number of breeding forest birds | 49 | 61 | 64 |
| Iberian/Balkan peninsulas | Iberian/Italian peninsulas | Italian/Balkan peninsulas | |
| Sørensen's similarity index | 0.45 | 0.47 | 0.48 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



