1. Introduction
Cassava (
Manihot esculenta Crantz) is a drought-tolerant root crop that serves as a crucial staple food for nearly one billion people across 105 countries, particularly in sub-Saharan Africa [
1,
2]. Steaming, a widely adopted and simple preparation method, makes mealiness a key determinant of consumer acceptance and overall cassava quality [
3,
4]. Consumer preference studies consistently show a strong favor for a distinctly mealy texture, establishing it as a paramount target trait in breeding programs [
5]. Consequently, identifying the components that govern mealiness and determining their predictive thresholds is essential for developing rapid identification methods.
To address this, numerous studies have primarily focused on the relationships between mealiness and components. A well-studied factor like dry matter content illustrates this limitation: a sub-25% level is necessary but not sufficient for a non-mealy phenotype [
6]. Furthermore, studies on gene-environment interactions indicate that both biological and abiotic factors during crop growth can influence related gene expression, leading to phenotypic deviations [
7]. These findings collectively underscore that cassava mealiness is a complex, multifactorial trait arising from synergistic physicochemical interactions.
For further research, the factors determining mealiness are often broadly attributed to pectin, starch, and cell wall properties [
4,
6]. However, specific findings point decisively to cell wall pectin, revealing no consistent link with starch properties or granule size [
8,
9,
10,
11,
12]. A similar mechanism has been observed in lotus roots, where the binding of pectin carboxyl groups to Ca
2+ ions inhibit cell wall degradation during heating, thereby playing a critical role in the development of a mealy texture [
13]. Beyond these compositional factors, the cooked quality of potatoes is also influenced by physical-mechanical factors such as cell turgor pressure, cell wall strength, and pectin adhesion [
14,
15,
16]. These insights further highlight the multifactorial nature of mealiness and underscore the particular importance of cell wall components.
As mealiness lacks a direct quantitative metric and is often assessed categorically, correlating it with continuous variables presents an effective strategy for its rapid identification. Decision tree analysis is particularly suitable for this purpose due to its capacity to model complex, non-parametric relationships, a strength confirmed in similar agricultural applications [
17,
18].
This study analyzes variations in cell wall composition and microstructure among cassava roots with differing mealiness levels at two growth stages to identify the key underlying factors. Using decision tree analysis, we establish a predictive threshold model for this trait, providing a theoretical foundation for targeted breeding and rapid postharvest grading of varieties with superior steaming quality.
2. Materials and Methods
2.1. Materials
Five cassava cultivars (SC9, GR11, GR10, GR891, and SC6068) were sampled at two distinct physiological maturity stages: 8 and 12 months after planting. All plant materials were grown under controlled field conditions at the Tropical Crops Genetic Resources Institute, Chinese Academy of Tropical Agricultural Sciences (CATAS), in Danzhou, Hainan Province, China. The experimental field was established during the regular growing sea-son (planted in April), following standardized agronomic practices. For each cultivar, biological triplicates were maintained under uniform soil and environmental conditions.
2.2. Flour Preparation
The method for preparing steamed cassava flour (SCF) was adapted from Jia et al, with minor modifications [
19]. Peeled cassava roots were sliced and steamed for 45 minutes, then oven-dried until the moisture content dropped below 14%. The dried samples were subsequently milled and sieved through a 178 μm mesh for further analysis.
2.3. Sensory Evaluation
To evaluate the degree of mealiness in steamed cassava tubers, a modified version of the method described by Franck et al [
4]. The five cassava varieties were washed, peeled, cut into uniform segments, and steamed for 45 minutes. A sensory panel comprising 10 individuals, including consumers and cassava producers, was assembled to assess tuber mealiness. Each panelist evaluated the samples by tasting and assigning a binary score: 0 for non-mealy and 1 for mealy.
2.4. Microstructure
Tissue samples from both fresh and boiled tubers of five cassava varieties, harvested at 8 and 12 months after planting, were sectioned using a sharp razor blade into approximately 3 × 3 mm (length × width) pieces. The sections were immediately transferred into tubes containing 20 mL of FAA fixative solution [90% ethanol (v/v), 5% formalin (v/v), and 5% acetic acid (v/v)] and fixed for a minimum of 24 hours. All procedures were conducted according to the established protocol, with each staining experiment performed in triplicate [
20].
2.5. Determination the Content of Dry Matter and Starch
Starch content (SC) was determined following the method described by Wang et al. [
21]. Similarly, dry matter (DM) content was measured according to previously established protocol [
22].
2.6. Determination the Content of Soluble Pectin, Water-Soluble Pectin, Covalently Bound Pectin in Steamed Cassava Flour
The contents of covalently-soluble pectin (CSP), ion-soluble pectin (ISP), and water-soluble pectin (WSP) were measured using commercially available detection kits (Su-zhou Michy Biomedical Technology Co., Ltd., Jiangsu, China) according to the manufacturer’s instructions.
For CSP and ISP determination, 0.1 g of sample was homogenized in 80% ethanol, heated at 95°C for 20 minutes, then washed with an ethanol/acetone mixture. After treatment with a starch-removal reagent for 15 hours, the samples were extracted using CSP/ISP-specific solutions and centrifuged to collect the supernatants.
WSP extraction involved homogenization in ethanol followed by two heating steps at 95°C for 30 minutes, with ethanol replenishment between steps. This was followed by water extraction at 50°C for 30 minutes and centrifugation.
All assays utilized galacturonic acid standards at 0.5 mg/mL. Absorbance was measured at 530 nm after color development induced by sulfuric acid. Specifically, CSP and ISP samples (50 μL) were reacted with carbazole reagent at 95°C for 5 minutes, while soluble pectin supernatants (30 μL) underwent hydroxy biphenyl derivatization, first at 90°C for 10 minutes, then at 25°C for 30 minutes.
2.7. Determination of Pectin Methylation in Steamed Cassava Flour
The degree of pectin methylation was determined using a micro-method kit (Suzhou Comin Biotechnology). Tissue samples (0.1 g) were homogenized in 95% ethanol and centrifuged at 10,000 × g, 4°C. The pellet was washed with distilled water and then extracted with extraction buffer at 90°C for 2 hours.
Methanol released from methyl ester bonds was quantified by measuring absorbance at 550 nm following derivatization with methanol oxidase and 4-amino-3-hydrazino-5-mercapto-1,2,4-triazole. The derivatization involved incubation at 37°C for 10 minutes, followed by 30°C for 30 minutes.
Total pectin content, reflected by galacturonic acid, was measured at 450/400 nm after sulfuric acid treatment at 70°C for 10 minutes and subsequent reaction with 3,5-dimethylphenol.
2.8. Determination the Content of Cellulose in Steamed Cassava Flour
Cellulose content (CLL) was determined using commercially available detection kits (Suzhou Michy Biomedical Technology Co., Ltd., Jiangsu, China) following the manufacturer’s instructions.
Briefly, 0.1 g of sample was homogenized with Reagent 1 and Reagent 2 (prepared with H2SO4) and incubated overnight at 60°C. The mixture was then washed with a Rea-gent 3/ethanol solution, followed by reaction with Reagent 2 at 95°C for 10 minutes. The absorbance of the resulting supernatants was measured at 620 nm to quantify cellulose content.
2.9. Determination of Enzyme Activity
Cellulase activity was measured using a DNS-based assay kit (Suzhou Michy Bio-medical Technology). Briefly, 0.5 g of sample was homogenized and sonicated in 1 mL extraction buffer (200 W, 3 s on/10 s off cycles, 30 cycles, with an ice bath). The homogenate was then centrifuged at 8,000 × g for 10 minutes at 4°C, and the supernatant collected. For the assay, 50 μL of supernatant was incubated with 150 μL of Reagent I (test) or 300 μL of Reagent II (control) at 37°C for 3 hours. The mixture was heated at 95°C for 15 minutes, followed by centrifugation at 8,000 × g for 10 minutes at 25°C. Then, 150 μL of the resulting saccharified solution was reacted with 150 μL of Reagent III at 95°C for 5 minutes. After cooling, absorbance was measured at 540 nm.
Pectin methylesterase (PME) activity was determined using a sodium hydroxide titration kit (Suzhou Michy Biomedical Technology). Samples (0.5 g) were homogenized in ice-cold extraction buffer using a mortar pre-cooled to –20°C, then centrifuged at 12,000 × g for 15 minutes at 4°C. The supernatant was collected, and 0.2 mL was mixed with 50 μL of Reagent II and 8 mL of pre-warmed Reagent I (37°C). The mixture’s pH was adjusted to 7.8 using Reagent IV, then incubated at 37°C for 60 minutes. During incubation, the pH was maintained at 7.8 by continuous titration with Reagent IV, and the volume of titrant consumed was recorded for activity calculation.
2.10. Model Validation
Ten edible cassava germplasms—SC9, SC6068, SC101, Guimushu 9, Guimushu 8, GR891, GR10, GR11, GR13, and Reyin—were selected to determine cellulose content, dry matter content, and mealiness after steaming. These measurements were used to validate the accuracy of the decision tree model. Agreement between experimental results and model predictions was indicated by “Y,” while disagreement was marked as “N”.
2.11. Statistical Analysis
Data were processed using ORIGIN 2024 software and presented as mean ± standard deviation from three replicates. Statistical analyses were performed using ANOVA followed by Duncan’s multiple range test in SPSS for Windows. Additionally, the CART decision tree algorithm was applied to classify the data and develop the predictive model.
3. Results
3.1. The Mealiness of Steamed Cassava Sensory Evaluation
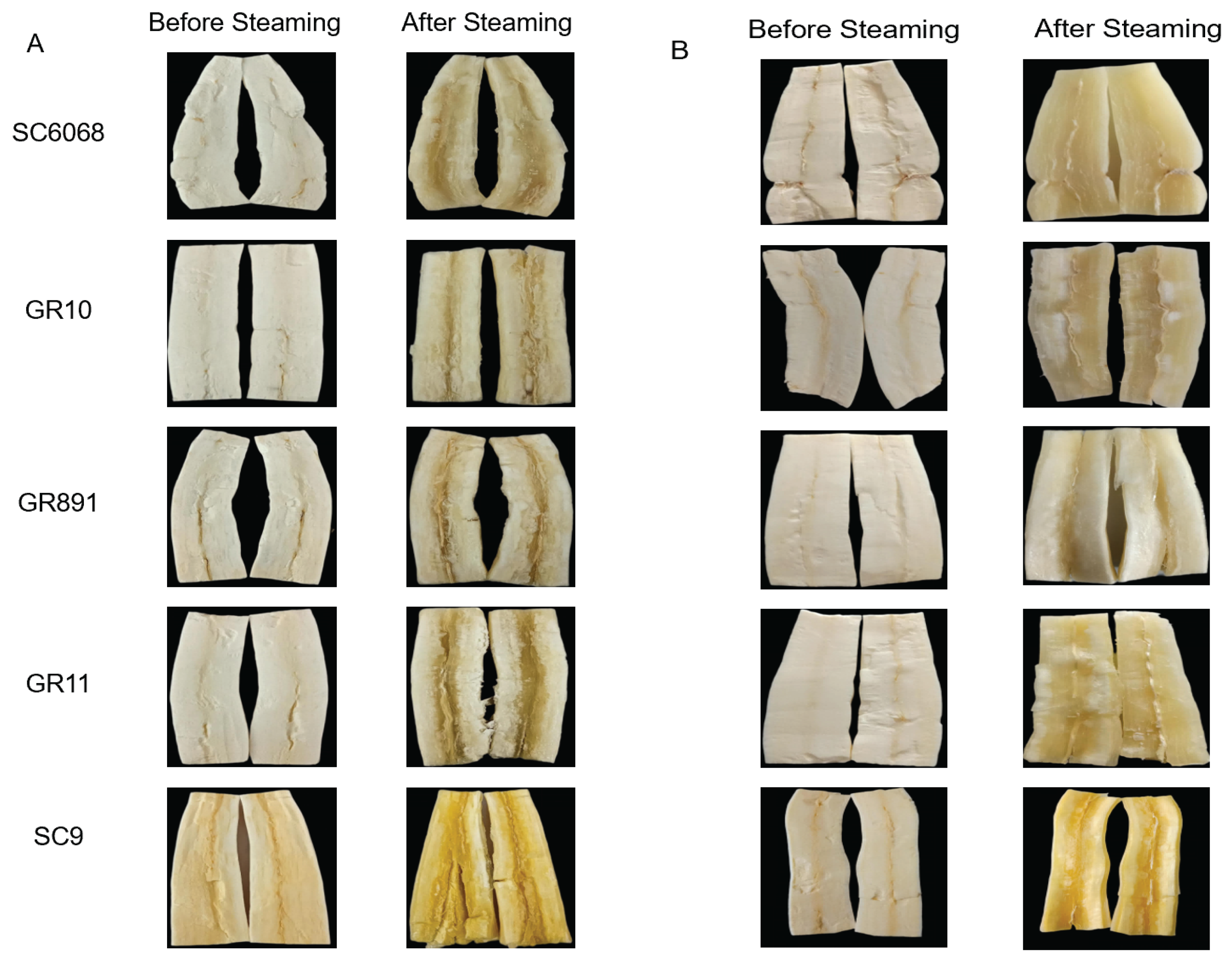
As shown in
Table 1, sensory evaluation revealed that three varieties—SC9, GR11, and GR891—exhibited mealiness at both 8 and 12 months after planting. In contrast, GR10 and SC6068 were mealy at 8 months but not at 12 months.
Figure 1 further demonstrates that the mealy varieties developed cracked roots and dry centers after steaming. By comparison, SC6068 and GR10, at 12 months, retained their structural integrity, showing no signs of cracking and maintaining moist centers [
4].
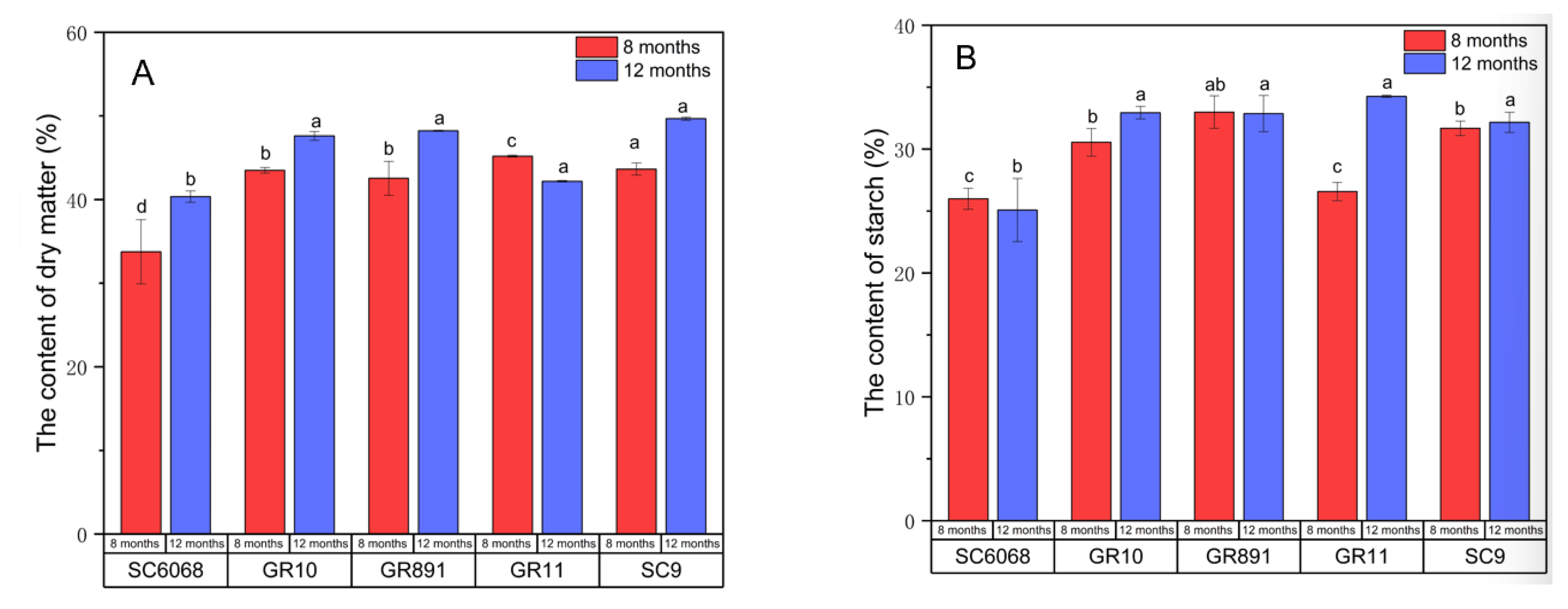
3.2. Dry Matter and Starch Content
As shown in
Figure 2, with the exception of GR11, the dry matter content of each variety generally increased with longer cultivation duration, whereas starch content exhibited no consistent trend. Previous studies have indicated that dry matter content has a decisive effect on mealiness only when it falls below 25% [
4,
5].
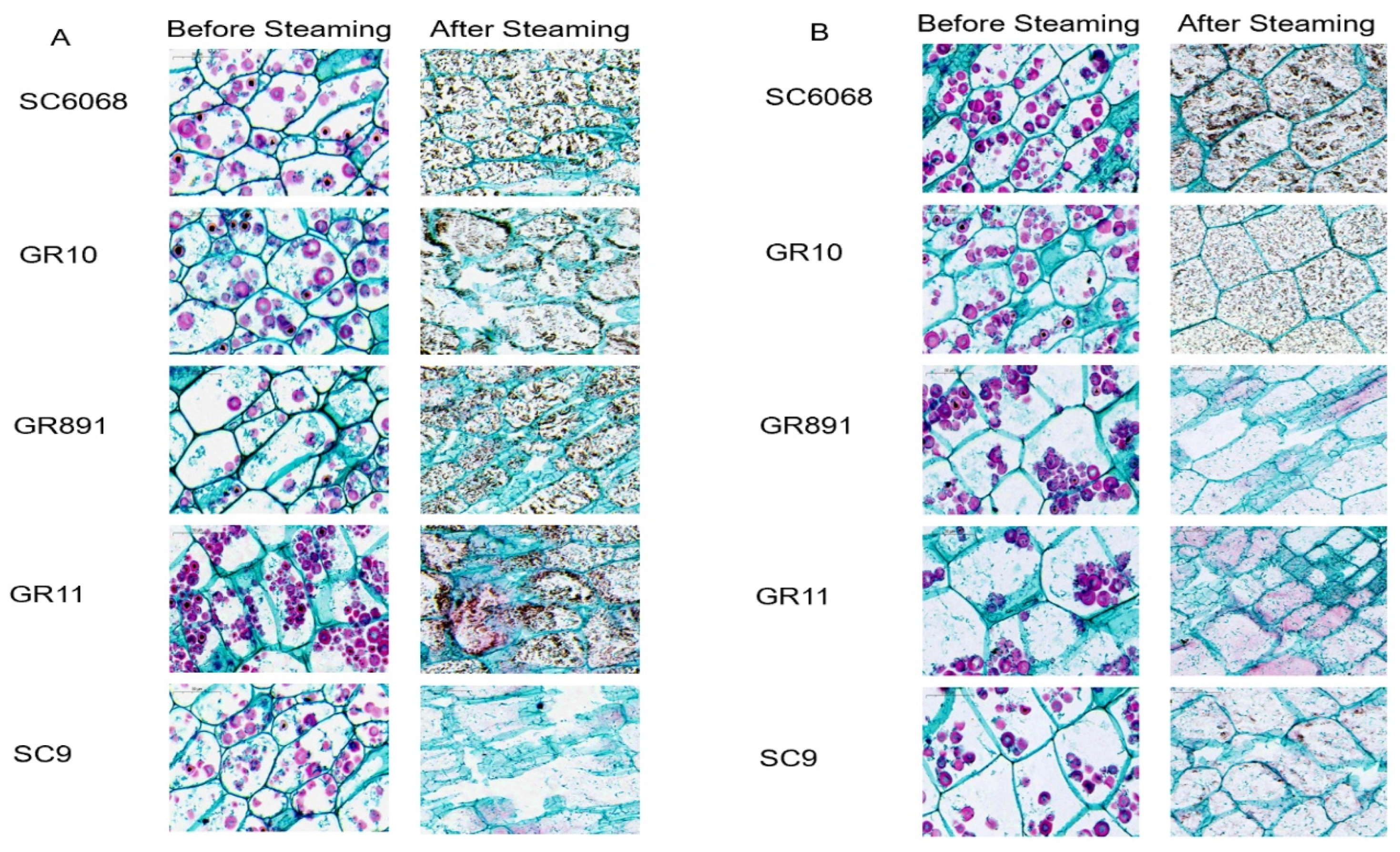
3.3. Microstructure Observation
As shown in
Figure 3, starch granules within the cells were stained red by Congo red. Light microscopy revealed that, although cell size and shape varied among the cassava varieties, the cells were densely packed across all samples. Prior to steaming, the cellular structures of all varieties remained intact. Following steaming, varying degrees of cell wall damage were observed in all varieties harvested at 8 months, as well as in SC9, GR11, and GR891 at 12 months. In contrast, SC6068 and GR10 harvested at 12 months retained a more intact cellular morphology.
Previous studies have reported that pectin forms a matrix within the cell wall network, playing a critical role in regulating cell wall permeability and thickness [
23]. Similarly, sweet potato varieties with higher pectin content in their cell walls exhibit greater structural stability and integrity after the same cooking duration [
24]. These findings highlight the importance of further investigating cell wall composition and structure.
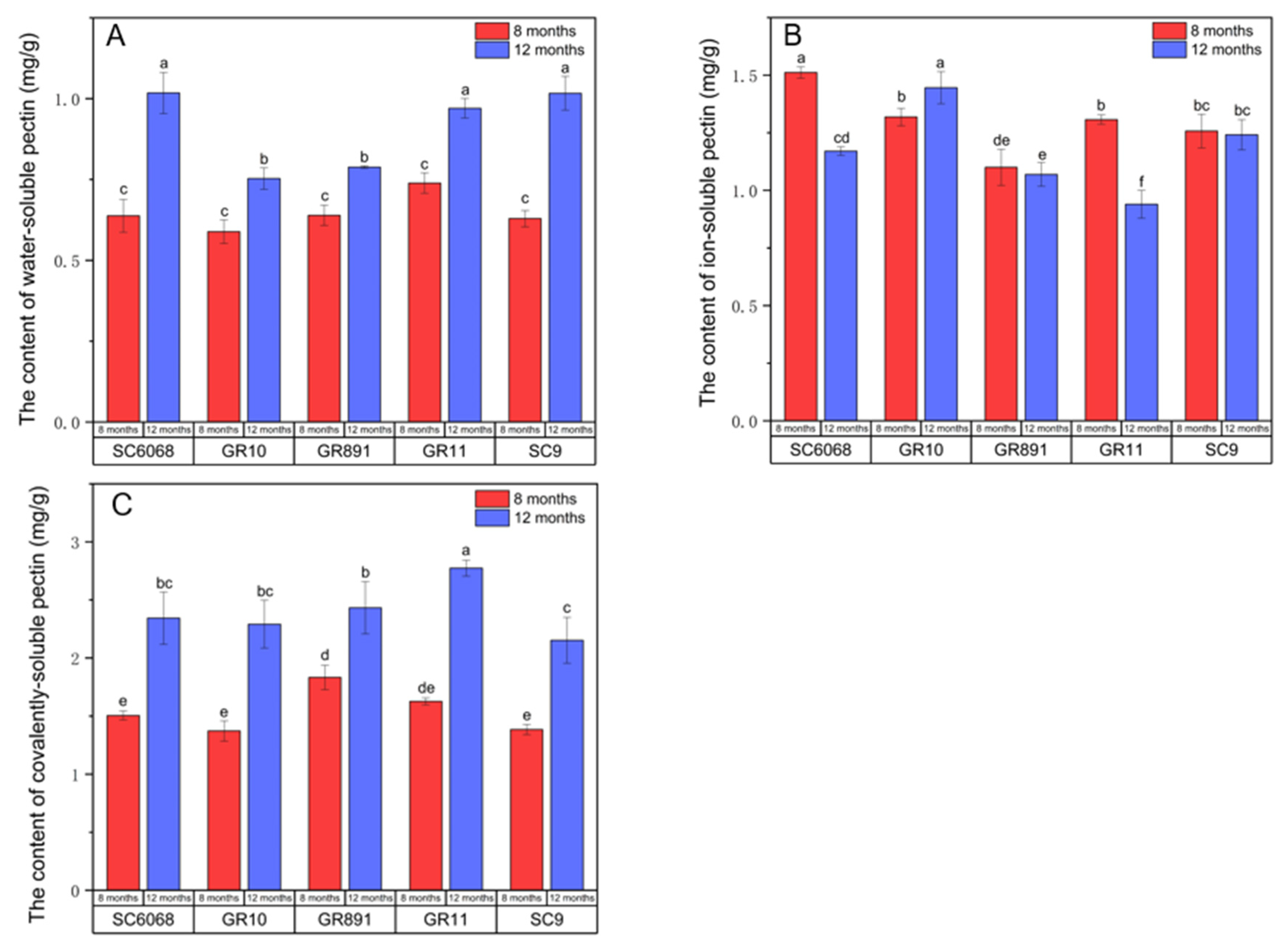
3.4. Analysis of the Pectin Components
CSP, ISP, and WSP represent three distinct pectin fractions, and the dynamic balance among them influences the structural integrity of the cell wall [
25]. As shown in
Figure 4, with increasing growth duration, the contents of WSP and CSP increased across all varieties, whereas ISP content decreased in all varieties except GR10.
Mechanistically, the labile chemical bonds in WSP promote progressive hydration of the cell wall matrix. Elevated levels of WSP reduce structural rigidity and increase cell wall porosity, thereby enhancing cellulase accessibility [
26,
27]. In contrast, CSP contributes to cell wall stability through ester bond-mediated crosslinking, playing a critical role in maintaining structural rigidity [
28]. Heat treatment typically softens plant tissues by increasing WSP levels and reducing CSP content [
29,
30].
However, the data from this study suggest that these three pectin fractions do not have a significant impact on cell wall strength. The underlying mechanisms responsible for this discrepancy remain to be elucidated.
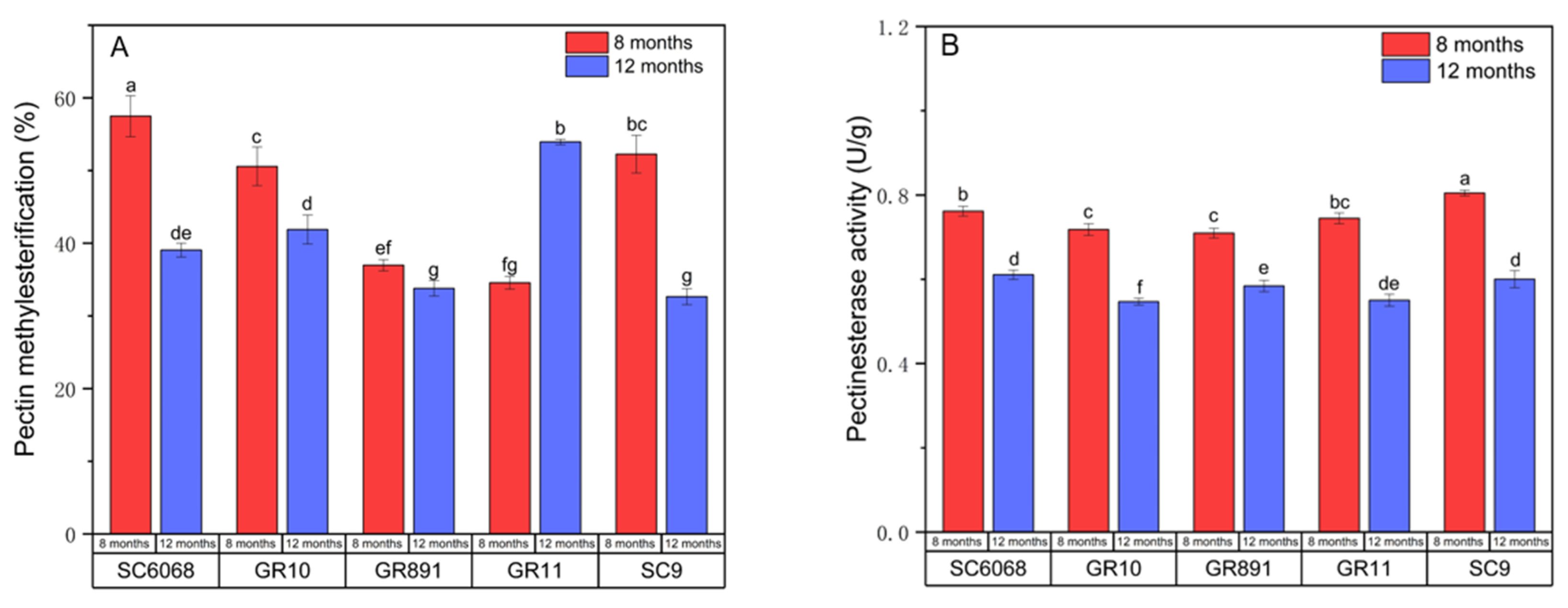
3.5. Analysis of Pectin Methylesterification and Enzyme Activity
Pectin degradation involves the action of pectin methylesterase (PME), which increases cell wall rigidity through demethylation [
31]. As shown in
Figure 5, except for GR11, all other varieties exhibited a higher degree of pectin esterification at 8 months compared to 12 months. Correspondingly, PME activity was higher at 8 months across all varieties.
Notably, SC6068 and GR10 displayed mealy and non-mealy characteristics at different growth stages, which may be linked to their demethylation patterns. Previous studies have reported that block wise demethylation enhances cell wall rigidity, whereas random demethylation results in loosening of the cell wall structure [
32].
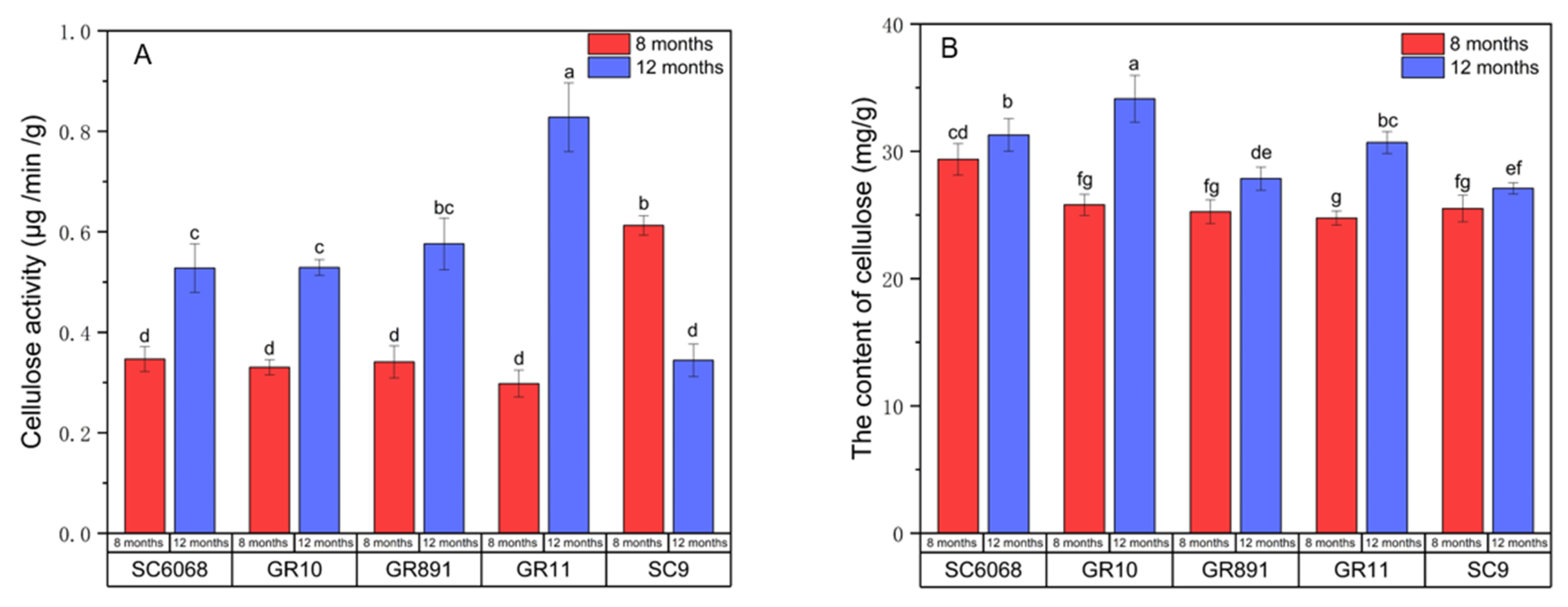
3.6. Analysis of Cellulose Content and Enzyme Activity
The cell wall is considered an internal barrier to starch hydrolysis, and studies confirm that cellulose in the cell wall is degraded by cellulase treatment, thus affecting cell wall integrity [
33,
34,
35]. In
Figure 6, cellulose content increased in all five varieties with longer growth time. The cellulose content of the non-mealy varieties SC6068 and GR10 at 12 months was significantly higher than that of the mealy varieties SC9, GR11, and GR891, which may also contribute to maintaining a more intact cell wall after cooking. Regarding enzyme activity, cellulase activity showed no significant differences among the varieties at 8 months. However, at 12 months, cellulase activity in SC6068 and GR10 was lower than in the other three varieties. This indicates that the degree of cell wall degradation is one important factor affecting mealiness, but the underlying mechanism remains to be elucidated.
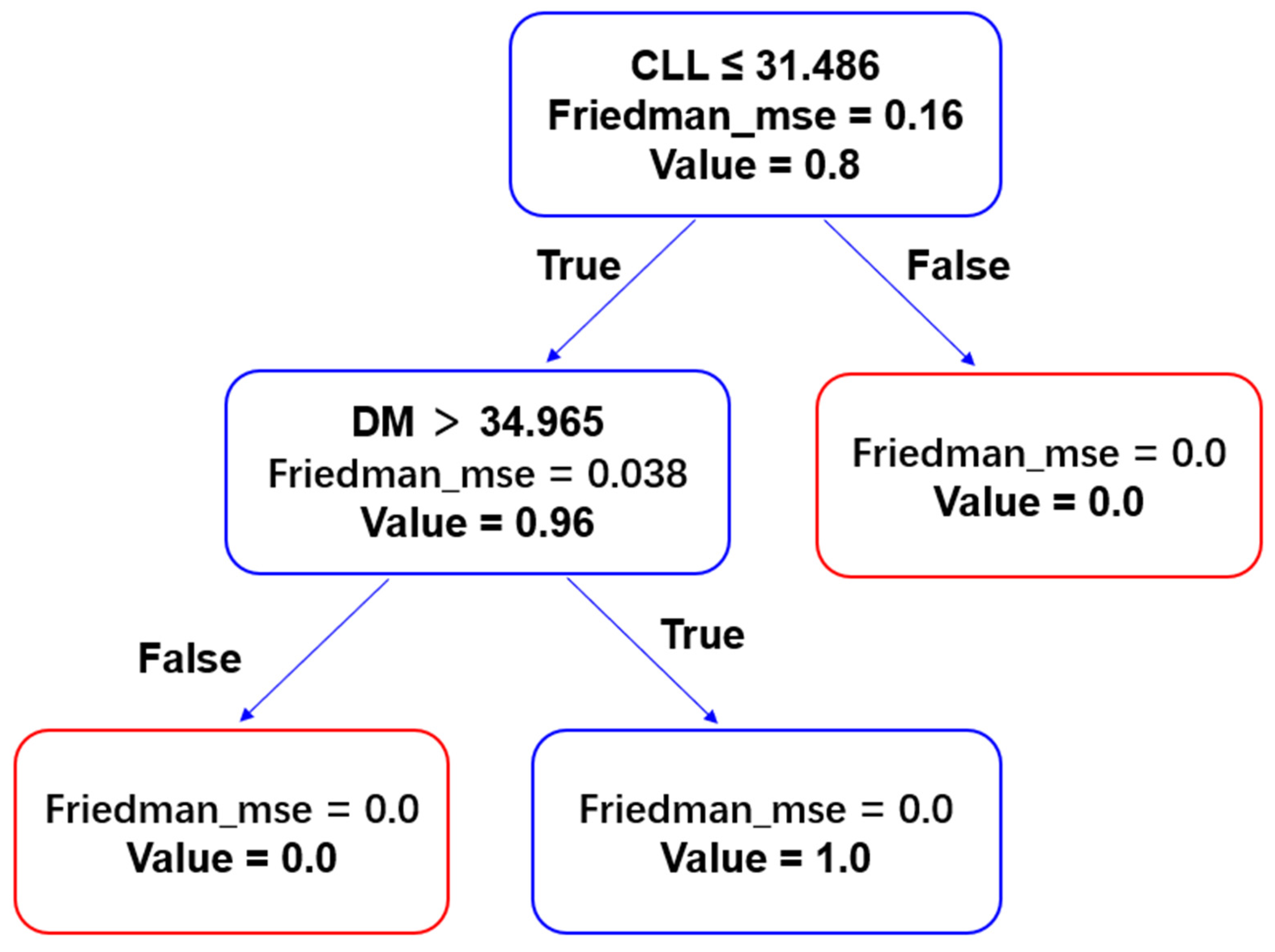
3.7. Decision Tree Analysis
As shown in
Figure 7, decision tree analysis identified cellulose content as the primary factor and dry matter content as the secondary factor influencing mealiness. Specifically, when the cellulose content of the tuber is less than or equal to 31.486 mg/g, there is an 80% probability that the tuber will exhibit a mealy texture. Moreover, when cellulose content is ≤31.486 mg/g and dry matter content exceeds 34.965%, the prediction accuracy increases to 96%.
Friedman’s mean squared error (MSE) was used to evaluate the quality of node splits in the decision tree, with higher MSE values indicating greater splitting error. As shown in
Table 2, cellulose content (CLL) accounts for 80% of the feature importance, while dry matter content (DM) contributes the remaining 20%.
3.8. Verify the Accuracy of the Decision Tree Analysis Model
As shown in
Table 3, the cellulose content, dry matter content, and mealiness of 10 cassava germplasm resources were evaluated. Among them, the mealiness of 7 germplasms aligned with the predictions of the decision tree model, while 3 did not, resulting in an overall prediction accuracy of 70%. This discrepancy may be attributed to the limited size of the dataset used to develop the model, underscoring the need for larger and more diverse datasets in future studies to enhance its predictive performance.
4. Discussion
Mealiness is a textural attribute perceived during the chewing of plant tissues. It is widely perceived as a quality defect in many fruits but is a desired characteristic in steamed cassava roots. This property is closely linked to cell wall integrity [
36], in which cellulose serves as the primary structural framework. The rigidity imparted by cellulose contributes to cell wall strength and resistance to rupture [
37].
Our results show that non-mealy varieties possess higher cellulose content and greater cellular integrity than mealy ones, implying that cell wall properties are a primary determinant of textural variation. This aligns with findings in lotus root, where textural differences between crisp and mealy have similarly been linked to cellulose degradation [
36] and higher moisture content [
38]. These physiological relationships are reinforced by our decision tree model, which identifies high dry matter and low cellulose as key markers for mealiness. While existing research has largely focused on identifying individual components associated with mealiness, the interactive mechanisms through which these components collectively drive texture remain poorly understood.
Current methods for identifying mealiness are hampered by their reliance on post-steaming analysis, which is inherently time-consuming and low-throughput, limiting their practical utility. The evidence presented in this study collectively indicates that the composition and structure of the cell wall are fundamental to mealiness development. This insight lays the groundwork for novel rapid detection technologies, which could significantly enhance the efficiency of cassava breeding and postharvest grading.
5. Conclusions
This study investigated the cell wall composition and structure of five cassava varieties across two planting seasons. The decision tree model identified cellulose content and dry matter content as the primary and secondary predictors of mealiness in steamed cassava roots, respectively. These findings provide valuable insights to inform future breeding programs and processing strategies.
Author Contributions
Conceptualization, L.L. and Z.Z.; methodology, Q.W.; software, L.L.; validation, M.L.; formal analysis, L.L. and P.D.; investigation, L.L. and Y.H.; data curation, L.L. and M.L.; writing—original draft, L.L.; writing—review and editing, L.L. and Z.Z.; supervision, Z.Z.; funding acquisition, Z.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the earmarked fund for CARS, grant number CARS-11 and The APC was funded by CARS-11.
Data Availability Statement
All relevant data have been supplied in this manuscript.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Okogbenin, E.; Setter, T.L.; Ferguson, M.; Mutegi, R.; Ceballos, H.; Olasanmi, B.; Fregene, M. Phenotypic approaches to drought in cassava: Review. Front. Physiol, 2013, 4, 93. [CrossRef]
- Zainuddin, I.M.; Fathoni, A.; Sudarmonowati, E.; Beeching, J.R.; Gruissem, W.; Vanderschuren, H. Cassava postharvest physiological deterioration: From triggers to symptoms. Postharvest Biol. Technol, 2018, 142,115-123. [CrossRef]
- Bechoff, A.; Tomlins, K.; Fliedel, G.; Lopez-Lavalle, L.A.B.; Westby, A.; Hershey, C.; Dufour, D. Cassava traits and end-user preference: Relating traits to consumer liking, sensory perception, and genetics. Crit. Rev. Food Sci. Nutr, 2017, 58, 547–567. [CrossRef]
- Franck, H.; Christian, M.; Noël, A.; Brigitte, P.; D. Joseph, H.; Cornet, D.; C. Mathurin, N. Effects of cultivar and harvesting conditions (age, season) on the texture and taste of boiled cassava roots. Food Chem, 2011, 126, 127-133. [CrossRef]
- Kelechi, U.; Damian, N.N.; Ugochukwu, N.I.; Daniel, D.; Pangirayi, T.; Samuel, O.; Chiedozie, E. Genotype-by-environment interaction and stability of root mealiness and other organoleptic properties of boiled cassava roots. Sci Rep, 2022, 12, 20909. [CrossRef]
- Padonou, W.; Mestres, C.; Nago, C.M. The quality of boiled cassava roots: instrumental characterization and relationship with physicochemical properties and sensorial properties. Food Chem, 2005, 89, 261-270. [CrossRef]
- Roberto, T. Quantitative Genetics, Genomics, and Plant Breeding. AOB, 2004, 93,2,225. [CrossRef]
- Menoli, A.V.; Beleia, A. Starch and pectin solubilization and texture modification during pre-cooking and cooking of cassava root (Manihot esculenta Crantz). LWT – Food Sci. Technol, 2007, 40, 744–747. [CrossRef]
- Favaro, S.P.; Beléia, A.; da Silva, F.J. N.; Waldron, K.W. The roles of cell wall polymers and intracellular components in the thermal softening of cassava roots. Food Chem, 2008, 108, 220–227. [CrossRef]
- Kelechi, U.; Damian, N.N.; Agre, P.; Ismail, Y.R.; Daniel, D.; Pangirayi T.; Samuel, O.; Chiedozie, E. Genome-Wide Association Study of Root Mealiness and Other Texture-Associated Traits in Cassava. Front. Plant. Sci, 2021, 12,770434. [CrossRef]
- Paniagua, C.; Posé, S.; Morris,V.J.; Kirby, A.R.; Quesada, M.A.; Mercado, J.A. Fruit softening and pectin disassembly: an overview of nanostructural pectin modifications assessed by atomic force microscopy, Annals of Botany, 2014, 114,1375-1383.
- . [CrossRef]
- Charoenkul, N.; Uttapap, D.; Pathipanawat, W.; Takeda, Y. Physicochemical characteristics of starches and flours from cassava varieties having different cooked root textures, LWT - Food Sci. Technol, 2011, 391 44 ,1774-1781. [CrossRef]
- Liu, G.; Liu, Y.; Yan, S.; Li, J. Acetic acid reducing the softening of lotus rhizome during heating by regulating the chelate-soluble polysaccharides. Carb pol, 2020, 240,116209. [CrossRef]
- Bordoloi, A.; Kaur, L.; Singh, J. Parenchyma cell microstructure and textural characteristics of raw and cooked potatoes. Food Chem, 2012, 133, 1092–1100. [CrossRef]
- Botero-Uribe, M.; Fitzgerald, M.; Gilbert, R. G.; Midgley, J. Effect of pulsed electrical fields on the structural properties that affect French fry texture during processing. Trends Food Sci Tech, 2017, 67, 1–11. [CrossRef]
- Dite Hunjek, D., Pranji´c, T., Repaji´c, M., & Levaj, B. Fresh-cut potato quality and sensory: Effect of cultivar, age, processing, and cooking during storage. J Food Sci, 2020, 85, 2296–2309. [CrossRef]
- Ramos, M. F.; Ribeiro, D. E.; Cirillo, M. Â.; Borém, F. M. Discrimination of the sensory quality of the Coffea arabica L. (cv. Yellow Bourbon) produced in different altitudes using decision trees obtained by the CHAID method. J Sci Food Agric, 2015, 96(10): 3543-3551. [CrossRef]
- Perri, P.F.; van der Heijden, P.G.M. A property of the CHAID partitioning method for dichotomous randomized response data and cate gorical predictors. J Classif, 2012, 29, 76–90. [CrossRef]
- Jia, M.; Ma, R.R.; Liu, C.; Yang, T.Y.; Zhan, J.L.; Shen, W.Y.; Tian, Y.Q. Isolated cassava cells: Comparison of structure and physicochemical properties with starch and whole flour. Carb pol, 2024, 343,122467. [CrossRef]
- Li, H.; Wang, Y.; Liu, H.; Lin, S.J.; Han, M.H.; Zhuang, J. Genomic analyses of the crosstalk between gibberellins and brassinosteroids metabolisms in tea plant (Camellia sinensis (L.) O. kuntze). Sci Hortic, 2020, 268:109368. [CrossRef]
- Wang, C.; Wang, Y.; Liu, X.; Cao, P.; Hu, X.; Chen, Y.; Li, S.; Xiao, Y.; Min, Y. Commercial potato starch standards cannot be used in place of cassava starch standards when measuring the starch content of cassava samples. Starch, 2019, 71, 1-10. [CrossRef]
- Karima, M.; Thierry, T.; Luis, F.D.; Maria, A.O.; Jhon, L.M.; Jorge, L.; Luis, L.; Dominique, D.; Fabrice, D. Hyperspectral imaging for the determination of relevant cooking quality traits of boiled cassava. J Sci Food Agric, 2023, 104(8), 4782-4792. [CrossRef]
- Broxterman, S.; Schols, H. Interactions between pectin and cellulose in primary plant cell walls. Carb pol, 2018, 192, 263-272. [CrossRef]
- Chen, X.; Zhu, L.; Zhang, H.; Zhang, Y.; Cheng, L.; Wu, G. Influence of cell-wall permeability on starch digestion in sweet potato cells, Food hyd, 2025, 159, 110718. [CrossRef]
- Xu, H.; Chen, Y.; Ding, S.; Qin, Y.; Jiang, L.; Zhou, H.; Deng, F.; Wang, R. Changes in texture qualities and pectin characteristics of fermented minced pepper during natural and inoculated fermentation process. Int. J. Food Sci. Technol, 2021, 56,6073-6085. [CrossRef]
- Willats, W.; McCartney, L.; Mackie, W.; Knox, J. Pectin: cell biology and prospects for functional analysis. Plant mol biol, 2001, 47(1-2), 9-27. [CrossRef]
- Posé, S.; Paniagua, C.; Matas, A.; Gunning, A.; Morris, V.; Quesada, M.; Mercado, J. A nanostructural view of the cell wall disassembly process during fruit ripening and postharvest storage by atomic force microscopy, Trends Food Sci Tech, 2019, 87,47-58. [CrossRef]
- Sila, D.; Van Buggenhout, S.; Duvetter, T.; Fraeye, I.; De Roeck, A.; Van Loey, A.; Hendrickx, M. Pectins in Processed Fruits and Vegetables: Part II—Structure–Function Relationships. Compr Rev Food Sci F, 2009, 8,86-104. [CrossRef]
- Deng, L.; Mujumdar, A.; Yang, X.; Wang, J.; Zhang, Q.; Zheng, Z.; Gao, Z.; Xiao, H. High humidity hot air impingement blanching (HHAIB) enhances drying rate and softens texture of apricot via cell wall pectin polysaccharides degradation and ultrastructure modification. Food Chem, 2018, 261(30),292-300. [CrossRef]
- Wang, J.; Mujumdar, A.S.; Deng, L.Z.; Gao, Z.J.; Xiao, H.W.; Raghavan, G.S.V. High-humidity hot air impingement blanching alters texture, cell-wall polysaccharides, water status and distribution of seedless grape. Carb pol, 2018, 194,15,9-17. [CrossRef]
- Xu, Z.; Yang, M.; Li, Z.; Xiao, J.; Yang, X.; Wang, H.; Wang, X. Tissue-specific pectin methylesterification and pectin methylesterase activities play a role in lettuce seed germination. Sci Hortic-amsterdam, 2022, 301,111134. [CrossRef]
- Hocq, L.; Pelloux, J.; Lefebvre, V. Connecting Homogalacturonan-Type Pectin Remodeling to Acid Growth. Trends Plant Sci, 2017, 22, 20-29. [CrossRef]
- Chaffey, N.; Alberts, B.; Johnson, A.; Lewis, J.; Raff, M.; Roberts, K.; Walter, P. Molecular Biology of the Cell. 4th edn. Ann. Bot, 2003, 91(3), 401. [CrossRef]
- Abramson, M., Shoseyov, O., Shani, Z. Plant Cell Wall Reconstruction toward Improved Lignocellulosic Production and Processability. Plant Sci, 2010, 178, 61–72. [CrossRef]
- Sun, Y.M.; Cheng, S.; Cheng, J.Y.; Langrish, T.A.G. Mass Transfer Resistance and Reaction Rate Kinetics for Carbohydrate Digestion with Cell Wall Degradation by Cellulase. Foods, 2024, 13(18), 2881. [CrossRef]
- Li, X.N.; Liang, Q.Y.; Gao, M.P.; Ou, Y.X.; Hu, Y.F.; Jiang, W.; Jiang, H.P.; Yang, S.D. Comparative Analysis of Soil Microbial Community Structures in Rhizosphere of Two Texture-Differentiated Lotus Root Varieties. Microorganisms, 2025, 13(7), 1637. [CrossRef]
- Wang, H.; Wang, H.; Yi, Y.; Hou, W.; Wang, L.; Ai, Y.; Min, T. Varying concentrations of ethephon induce antioxidant defences and cell wall degradation to regulate storage quality of fresh-cut lotus root. Food Biosci. 2024, 61, 104975. [CrossRef]
- Wei, Y.F.; Zhou, X.Y.; Gao, M.P.; Ou, Y.X.; Hu, Y.F.; Jiang, W.; Jiang, H.P.; Yang, S.D. Different Endophytes Colonized in Various Lotus Root Varieties and Their Associated Mealy and Crunchy Properties, Int J Mol Sci, 2025, 26(10), 4529. [CrossRef]
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).