
Submitted:
22 January 2026
Posted:
23 January 2026
You are already at the latest version
Abstract
Garlic (Allium sativum L.) cultivars in Korea, particularly the widely adaptable ‘Hongsan’, are challenging to identify in processed forms or seedlings due to the plasticity of phenotypic traits such as clove tip greening, which risks mislabeling and infringement of UPOV breeders' rights. This study aimed to develop a stable SCAR marker for ‘Hongsan’-specific identification using RAPD-bulked segregant analysis (BSA). Sixty Operon primers (>60% GC) were screened against ‘Hongsan’ gDNA versus a non-’Hongsan’ BSA pool (‘Daeseo’, ‘Uiseong’, ‘Danyang’, and ‘Namdo’); OPE-01 consistently amplified a unique 1.3 kb band, cloned and sequenced to reveal a 1,272 bp sequence with translocation junction (878+394 bp), 18 bp insertion, and EcoRI site on chromosome 2 (NCBI GCA_030737875.1). SCAR primers SaH191R/SaH513F produced a specific 545 bp amplicon in ‘Hongsan’, clearly distinguishing it from other cultivars and parental lines, confirming paternal origin (9209). This RAPD-to-SCAR marker overcomes reproducibility limitations, enabling authentication in processing (powders, black garlic) irrespective of environmental factors. The cost-effective and rapid assay ensures industry transparency, quality control, and IP protection for Korean garlic production.
Keywords:
garlic (Allium sativum L.)
; ‘Hongsan’
; RAPD
; SCAR
; cultivar identification
; molecular marker
1. Introduction
Garlic (Allium sativum L.) is a perennial crop belonging to the family Liliaceae and is widely used as a seasoning vegetable globally due to its unique flavor and various bioactive compounds. In Korea, garlic is mainly divided into the introduced southern ecotype and the indigenous northern ecotype based on ecological types [1]. Southern ecotype garlic matures early and includes cultivars such as ‘Namdo’ and ‘Daeseo’. Northern ecotype garlic, mainly cultivated in central regions, includes cultivars like ‘Danyang’ and ‘Uiseong’. Northern ecotype garlic is characterized by a long dormancy period, late maturity compared to southern ecotype, 6-8 cloves, excellent storage quality, strong pungency, and rich flavor [2]. Additionally, there is the ‘Hongsan’ cultivar, a cross-bred variety suitable for both regions. Since most garlic cultivars are sterile, breeding has largely relied on clonal selection [3]. However, the discovery of fertile garlic clones in the late 20th century [4,5], and the confirmation of true seed production via crossing [6,7,8] allowed breeding research to commence in Korea in 1986 [9]. Consequently, three cultivars, ‘Dasan’, ‘Hwasan’, and ‘Cheonwoon’ were bred using fertile lines (18 fertile lines, including 8661) and 12 male sterile lines, including 8614 [10].
Following the registration of garlic as a protected crop under the UPOV convention in 2009, the need to develop new cultivars with superior environmental adaptability and consumer appeal arose [11,12]. ‘Hongsan’ was developed from true seeds derived from a cross between the fertile maternal line '8902' and the paternal line '9209' in 2003. It was registered as a new cultivar in 2015 after passing artificial seed production, acclimatization, productivity tests, and regional adaptability trials [2,13,14]. ‘Hongsan’ garlic can be cultivated in both northern and southern regions, exhibits higher yields than existing cultivars, and is suitable for labor-saving cultivation due to its easy harvesting [2,15].
Although ‘Hongsan’ can be visually distinguished by the light green chlorophyll expression at the clove tip, this trait may not appear depending on cultivation or drying methods, making identification difficult in processed or seedling states [16]. Furthermore, when ‘Hongsan’ garlic is used as a food ingredient after grinding or processing, or when products are developed using its bulbils, determining whether the product contains the ‘Hongsan’ variety becomes challenging. This raises the risk of ‘Hongsan’ being sold as a different variety or other varieties being misrepresented as ‘Hongsan’, highlighting the need for research on DNA molecular markers for specific variety identification.
DNA molecular markers are not influenced by cultivation methods, weather conditions, or environmental factors, making them highly useful for cultivar identification [17,18,19,20]. Furthermore, they can be effectively applied to assess genetic diversity within species [21,22]. Among these, the RAPD method is often integrated with other techniques; for instance, RAPD and RFLP have been used to analyze genetic relationships among fig cultivars [23], and an analysis of differences in the evaluation of genetic relationships among Brassica cultivars using RFLP and RAPD markers [24]. RAPD analysis has also been employed to identify polymorphic bands, followed by sequencing to develop species-specific SCAR markers, Allium cepa as demonstrated in the identification of Commiphora species [25] and native Malaysian orchid species [26].
Previous molecular marker studies in garlic have utilized morphological and chemical traits for classification [27], RAPD markers for genetic variation and cultivar identification [28], EST-derived SSR markers to analyze genetic diversity and population structure in 127 garlic germplasm accessions [29], AFLP markers to evaluate genetic diversity among 20 Brazilian garlic cultivars [30], and SLAF-seq specific length amplified fragment sequencing-derived SNP markers for genetic diversity and population structure analysis of 102 Chinese garlic cultivars [31]. However, studies on domestic Korean garlic cultivars remain limited.
Therefore, this study aimed to provide baseline data on domestically cultivated garlic cultivars and develop reliable molecular markers for varietal identification to prevent cultivar admixture. To achieve this, polymorphic bands were obtained using 60 random primers, followed by sequence analysis and SCAR marker development to create varietal-specific markers. The developed markers successfully distinguish the ‘Hongsan’ cultivar from representative domestic garlic varieties, including ‘Daese’, ‘Uiseong’, ‘Danyang’, and ‘Namdo’. These markers hold significant potential for ensuring transparency and quality control in the garlic processing industry, while also supporting intellectual property protection for breeders of new varieties.
2. Materials and Methods
2.1. Garlic Cultivars and gDNA Extraction
This study utilized garlic cultivars ‘Hongsan’, ‘Daeseo’, ‘Uiseong’, ‘Danyang’, and ‘Namdo’ grown at the Allium Research Center, National Institute of Horticultural and Herbal Science, Rural Development Administration (RDA), as test materials. Fresh shoots from each cultivar were collected for gDNA extraction.
Genomic DNA (gDNA) was extracted using the DNeasy Plant Mini Kit (QIAGEN, Hilden, Germany) following the manufacturer's protocol. The concentration and purity of extracted gDNA were measured with a Multiskan SkyHigh Microplate Spectrophotometer (Thermo Fisher Scientific, Waltham, USA), and all samples were diluted to 10 ng/µL for subsequent experiments.
2.2. Random Amplified Polymorphic DNA (RAPD) PCR
RAPD experiments were conducted using a bulked segregant analysis (BSA) approach with a DNA pool from four cultivars excluding ‘Hongsan’. The 60 Operon random primers selected had a high GC content of over 60% for stability and were chosen to maximize inclusion of Operon A- and C-type primers effective for detecting polymorphic bands in garlic, as reported by Alimoradi et al. [32]. The PCR reaction mixture (total 10 µL) consisted of 3.4 µL nuclease-free water, 1 µL gDNA (10 ng/µL), 0.6 µL random primer (5 pmol), and 5 µL rTaq (Takara, Seoul, Republic of Korea). PCR conditions included initial denaturation at 95°C for 4 min, followed by 40 cycles of 94°C denaturation for 40 s, 40°C annealing for 1 min, and 72°C extension for 2 min, with a final extension at 72°C for 5 min.

Table 1.
Information on 48 random primers.
| No. | Primer | Sequence | GC(%) | No. | Primer | Sequence | GC(%) |
| 1 | OPA-07 | 5'-CAGGCCCTTC-3 | 70 | 25 | OPB-01 | 5'-GTTTCGCTCC-3’ | 60 |
| 2 | OPA-14 | 5'-TCTGTGCTGG-3' | 60 | 26 | OPA-04 | 5'-AATCGGGCTG-3' | 60 |
| 3 | OPE-16 | 5'-CCTGATCACC-3' | 60 | 27 | OPA-08 | 5'-GTGACGTAGG-3' | 60 |
| 4 | OPB-18 | 5'-CCACAGCAGT-3’ | 60 | 28 | OPA-09 | 5'-GGGTAACGCC-3’ | 70 |
| 5 | OPA-19 | 5'-CAAACGTCGG-3’ | 60 | 29 | OPA-10 | 5'-GTGATCGCAG-3' | 60 |
| 6 | OPC-07 | 5'-GTCCCGACGA-3’ | 70 | 30 | OPC-06 | 5'-GAACGGACTC-3' | 60 |
| 7 | OPB-13 | 5'-TTCCCCCGCT-3' | 70 | 31 | OPA-12 | 5'-TCGGCGATAG-3' | 60 |
| 8 | OPE-01 | 5'-CCCAAGGTCC-3' | 70 | 32 | OPE-14 | 5'-TGCGGCTGAG-3’ | 70 |
| 9 | OPA-17 | 5'-GACCGCTTGT-3’ | 60 | 33 | OPA-15 | 5'-TTCCGAACCC-3' | 60 |
| 10 | OPE-08 | 5'-GGTCGGAGAA-3' | 60 | 34 | OPA-16 | 5'-AGCCAGCGAA-3' | 60 |
| 11 | OPE-09 | 5'-TCGGACGTGA-3' | 60 | 35 | OPA-18 | 5'-AGGTGACCGT-3' | 60 |
| 12 | OPB-02 | 5'-TGATCCCTGG-3’ | 60 | 36 | OPA-01 | 5'-CAGGCCCTTC-3’ | 70 |
| 13 | OPE-11 | 5'-GGAAGTCGCC-3' | 70 | 37 | OPA-11 | 5'-CAATCGCCGT-3' | 60 |
| 14 | OPC-05 | 5'-GATGACCGCC-3' | 70 | 38 | OPA-20 | 5'-GTTGCGATCC-3' | 60 |
| 15 | OPE-17 | 5'-CTACTGCCGT-3' | 60 | 39 | OPC-09 | 5'-CTCACCGTCC-3' | 70 |
| 16 | OPE-18 | 5'-GGACTGCAGA-3’ | 60 | 40 | OPC-13 | 5'-AAGCCTCGTC-3' | 60 |
| 17 | OPE-20 | 5'-AACGGTGACC-3' | 60 | 41 | OPD-01 | 5'-ACCGCGAAGG-3' | 70 |
| 18 | OPH-01 | 5'-GGTCGGAGAA-3' | 60 | 42 | OPD-03 | 5'-GTCGCCGTCA-3' | 70 |
| 19 | OPC-04 | 5'-CCGCATCTAC-3’ | 60 | 43 | OPH-02 | 5'-TCGGACGTGA-3' | 60 |
| 20 | OPH-04 | 5'-GGAAGTCGCC-3' | 70 | 44 | OPE-03 | 5'-CCAGATGCAC-3' | 60 |
| 21 | OPF-03 | 5'-CCTGATCACC-3' | 60 | 45 | OPE-12 | 5'-TTATCGCCCC-3’ | 60 |
| 22 | OPG-18 | 5'-GGCTCATGTG-3' | 60 | 46 | OPB-06 | 5'-TGCTCTGCCC-3' | 70 |
| 23 | OPG-19 | 5'-GTCAGGGCAA-3' | 60 | 47 | OPG-02 | 5'-GGCACTGAGG-3' | 70 |
| 24 | OPA-02 | 5'-TGCCGAGCTG-3’ | 70 | 48 | OPG-03 | 5'-GAGCCCTCCA-3' | 70 |
2.3. Cloning
Cloning was performed using the Mighty TA-Cloning Kit (Takara, Seoul, Republic of Korea) according to the manufacturer's protocol, with DH5α competent cells (Enzynomics, Daejeon, Republic of Korea). The cloning process involved purification of PCR products using the PCR Prep Kit (Takara, Seoul, Republic of Korea) before ligation. The T-Vector pMD20 provided in the TA-Cloning Kit was used as the cloning vector.
2.4. Target Insert Confirmation by PCR and Sequencing
To confirm proper insertion of target bands into plasmids extracted from cloned E. coli, PCR was performed using M13 primers, followed by restriction enzyme digestion targeting sites within the vector. The PCR reaction mixture (total 10 µL) consisted of 3.4 µL nuclease-free water, 1 µL gDNA (10 ng/µL), 0.3 µL M13 F primer, 0.3 µL M13 R primer, and 5 µL ExTaq (Takara, Seoul, Republic of Korea). PCR conditions included initial denaturation at 95°C for 4 min, followed by 35 cycles of 94°C denaturation for 30 s, 55°C annealing for 30 s, and 72°C extension for 40 s, with a final extension at 72°C for 5 min. Takara Q-Cut XbaI and KpnI (Takara, Seoul, Republic of Korea) were used for digestion. Both enzymes were incubated at 37°C for 16 h due to their compatible reaction temperatures. Selected samples were diluted to a minimum concentration of 150 ng/µL for sequencing and submitted to Macrogen (Sejong, Republic of Korea) for four reactions using M13-pug F and M13 R primers.
2.5. Analysis of PCR Product and DNA Sequence Data
PCR products were analyzed by electrophoresis on 1.8% agarose gels at 100 V for 45 min, visualized under UV light using a FireReader V10 (UVITEC, England, United Kingdom). And sequence data were reassembled using the Staden Package [33,34,35,36]. Assembled sequences were compared to garlic reference sequences (GCA_030737875.1) in the NCBI (National Center for Biotechnology Information, Bethesda, USA) database.
2.6. SCAR-PCR Validation
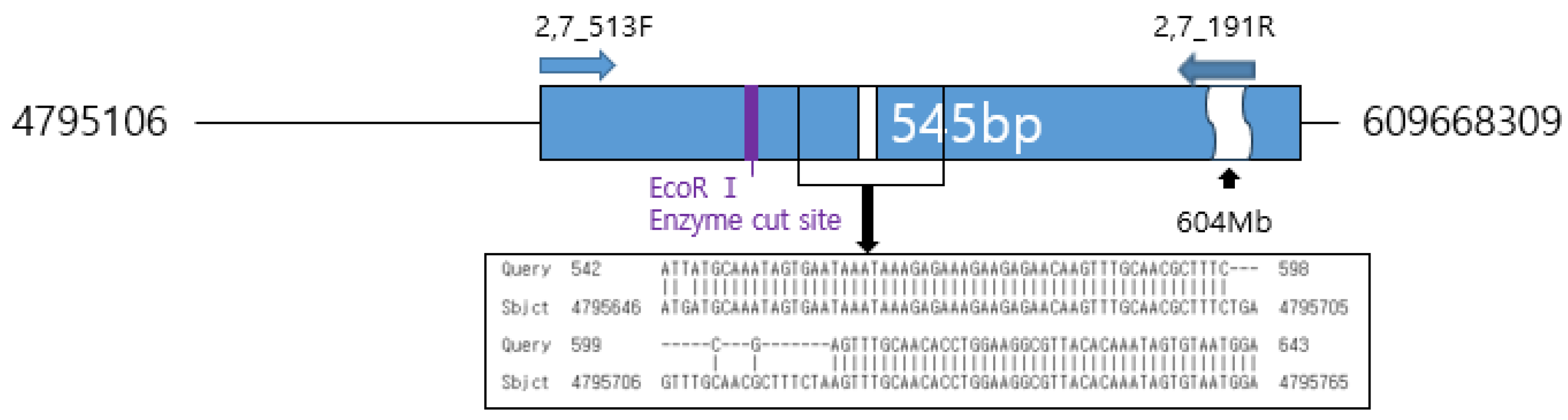
SCAR primers SaH191R (junction-specific) and SaH513F (gap/EcoRI site-targeting) were designed from the 1,272 bp sequence (Figure 8; Table 2) using Primer3 software [37] with parameters: amplicon size 500-600 bp, primer length 18-22 mer, GC content 50-60%, Tm 58-65°C. PCR reactions (10 μL) followed RAPD conditions: 1 μL gDNA (10 ng μL⁻¹), 0.5 μL each primer (10 pmol μL⁻¹), 5 μL rTaq polymerase (Takara, Seoul, Republic of Korea), 3 μL nuclease-free water. Thermal cycling: initial denaturation at 95°C for 4 min; 35 cycles of 94°C for 40 s, 60°C for 1 min, 72°C for 1 min 30 s; final extension at 72°C for 7 min.
Validation tested five garlic cultivars (‘Hongsan’, ‘Daeseo’, ‘Uiseong’, ‘Danyang’, and ‘Namdo’) and parental lines (maternal 8902, paternal 9209). PCR products were separated on 1.8% agarose gels (100 V, 45 min) and visualized under UV light (FireReader V10, UVITEC, UK). Expected amplicon size was 545 bp for ‘Hongsan’. To confirm specificity against repetitive sequences, PCR was repeated three times independently. Annealing temperature gradient (55-65°C) optimized specificity, with 60°C selected for reproducibility.
3. Results
3.1. RAPD PCR
For RAPD PCR templates, genomic DNA (gDNA) from the ‘Hongsan’ cultivar and a bulked gDNA mixture (BSA; bulked segregant analysis) of ‘Danyang’, ‘Uiseong’, ‘Namdo’, and ‘Daeseo’ cultivars were utilized. RAPD analysis using 60 random primers revealed that 12 primers produced no bands, while 48 generated valid bands. Consequently, the reproducibility of these 48 primers was assessed (Figure 1, Figure 2 and Figure 3). The results indicated that 8 primers, including OPE-09, lacked reproducibility. However, 13 primers, such as OPA-07 (Figure 1: {1}, {2}, {3}, {7}, {8}, {9}; Figure 2: {25}, {29}, {32}; Figure 3: {36}, {37}, {38}), yielded bands that clearly distinguished ‘Hongsan’ from the other cultivars. The remaining 27 primers were excluded from selection due to insufficient specificity.
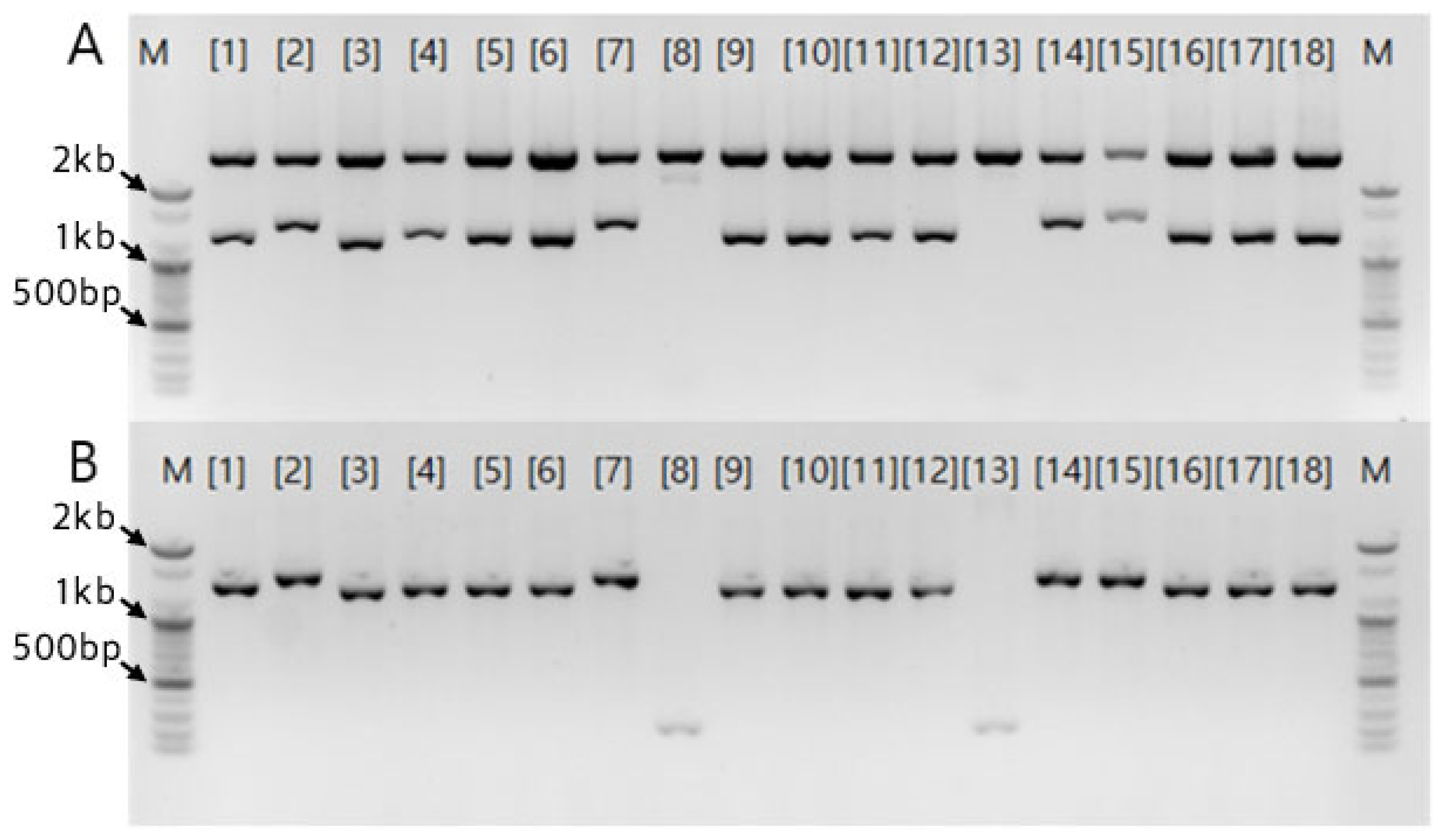
Using the 13 selected primers, independent band patterns were analyzed for each of the five garlic cultivars using individual genomic DNA samples rather than the BSA method (Figure 4). The ‘Uiseong’, ‘Namdo’, and ‘Danyang’ cultivars generally exhibited similar band patterns, whereas ‘Hongsan’ and ‘Daeseo’ displayed patterns similar to one another. Notably, a polymorphic band specific to the ‘Hongsan’ cultivar was identified at approximately 1.3 kb using the OPE-01 primer {8}. This specific band was subsequently selected for cloning.
The observation of a unique band in the ‘Hongsan’ cultivar suggests that specific primers are capable of detecting polymorphisms in the ‘Hongsan’ cultivar. These results demonstrate, through comparison with BSA, that the RAPD technique is a useful tool for detecting genetic differences among cultivars. Meanwhile, the similar band patterns among the ‘Uiseong’, ‘Namdo’, and ‘Danyang’ cultivars appear to be involved with the genetic similarity of highly repetitive sequences in garlic. This aligns with results stating that although there are cultivation differences between northern and southern ecotypes, they are not clearly distinguished by RAPD analysis [38]. Furthermore, the fact that almost no genetic variation appears among cultivars in the highly repetitive sequences of garlic, as reported by Lee [39], also supports these findings.
The similar band patterns observed between the ‘Hongsan’ and ‘Daeseo’ cultivars suggest the possibility that the two cultivars originate from similar ecotypes or share a similar genetic background. However, the specific band detected exclusively in ‘Hongsan’ represents a unique polymorphism, confirming the potential for developing RAPD-based molecular markers. In particular, the polymorphic band cloned based on these RAPD results is expected to serve as foundational data for the molecular identification of the ‘Hongsan’ cultivar.
3.2. Cloning and Sequence Analysis
Colony PCR was performed on 48 white colonies using M13 primers (Figure 5). Six distinct bands of varying sizes were observed, suggesting that all amplicons generated by the OPE-01 primer were individually cloned. Among these, the band at approximately 1.4 kb was identified as a potential specific marker for the ‘Hongsan’ cultivar. Additionally, a band at approximately 1.3 kb was detected; thus, it was decided to extract plasmids from the colonies containing these fragments. Consequently, a total of 18 colonies, including OPE-01-H colony 2 and OPE-01-H colony 5, were cultured in a shaking incubator at 37°C for 16 hours.
Following colony propagation, plasmids were extracted, and the results of PCR and restriction enzyme digestion are presented in Figure 5. A total of 18 plasmids were isolated, and the two band sizes (1.3 kb and 1.4 kb) observed in the colony PCR were consistently detected.
The results of restriction enzyme digestion (Figure 5A) appeared approximately 100 bp larger than the original bands. This occurred because the internal restriction sites of the target bands were unknown, requiring the use of restriction sites within the vector that were closest to the insert. Additionally, as shown in Figure 5B, the amplified bands were larger than the original bands, which is attributed to the influence of the M13 primer binding sites located at both ends of the pMD20 T-vector. Although previous studies suggest using the same primers from the initial PCR for colony PCR [40], this study used 10-mer random primers, necessitating the use of universal primers flanking the cloning site.
Consistent with the previous colony PCR results, 11 colonies, including {1} and {3}, exhibited bands at approximately 1.3 kb, while 5 colonies, including {2} and {4}, showed bands at approximately 1.4 kb. However, bands were not detected in some plasmids. According to Popov et al. [41], plasmids can be irregularly distributed between mother and daughter cells during E. coli replication, which may lead to plasmid loss. Plasmid loss likely occurred during the incubation, a phenomenon occasionally observed in unstable clones. Consequently, excluding the two plasmids where no bands were detected, the remaining 16 plasmids were submitted to Macrogen for sequencing.
Figure 6.
PCR results of the 18 selected plasmids using the restriction enzymes XbaI and BamHI and the M13 primers. (A) Restriction enzyme digestion results of 18 plasmids using XbaI and KpnI. (B) PC results of 18 plasmids using the M13 primer. {M} 100bp Size Marker, {1} E01H_P1, {2} E01H_P2, {3} E01H_P3, {4} E01H_P4, {5} E01H_P5, {6} E01H_P6, {7} E01H_P7, {8} E01H_P8, {9} E01H_P9, {10} E01H_P10, {11} E01H_P11, {12} E01H_P12, {13} E01H_P13, {14} E01H_P14, {15} E01H_P15, {16} E01H_P16, {17} E01H_P17, {18} E01H_P18.
Figure 6.
PCR results of the 18 selected plasmids using the restriction enzymes XbaI and BamHI and the M13 primers. (A) Restriction enzyme digestion results of 18 plasmids using XbaI and KpnI. (B) PC results of 18 plasmids using the M13 primer. {M} 100bp Size Marker, {1} E01H_P1, {2} E01H_P2, {3} E01H_P3, {4} E01H_P4, {5} E01H_P5, {6} E01H_P6, {7} E01H_P7, {8} E01H_P8, {9} E01H_P9, {10} E01H_P10, {11} E01H_P11, {12} E01H_P12, {13} E01H_P13, {14} E01H_P14, {15} E01H_P15, {16} E01H_P16, {17} E01H_P17, {18} E01H_P18.
Assembly of the sequences using the Staden Package yielded two fragments of 1,123 bp and 1,272 bp. Comparison with the NCBI database confirmed the highest homology with chromosome 2 of the garlic genome. Further analysis indicated that over 30 regions with >98% similarity to the 1,123 bp sequence were present on chromosome 1, with similar frequencies on other chromosomes (Figure 7A). Consequently, this sequence was identified as repetitive and excluded. Detailed analysis focused on the 1,272 bp sequence. BLAST analysis split this sequence into 878 bp and 394 bp fragments. The 878 bp fragment matched chromosome 2 (4,795,106–4,796,000 bp). The 394 bp fragment aligned to two locations: one at a distance of 11 Mb and another at 604 Mb. However, as the 11 Mb site exhibited reversed primer orientation, the location separated by 604 Mb was determined as the correct match (Figure 7B). The full 878 bp sequence was unique to the specified chromosome 2 locus, whereas a 578 bp partial sequence (lacking the 5' 300 bp) appeared in three or more copies on each chromosome (Figure 7C).
3.3. Design of SCAR Molecular Markers
Within the 1,272 bp sequence of the ‘Hongsan’ cultivar, a putative translocation site was identified that differs from the garlic reference sequence in the NCBI database. Consequently, the SaH_191R primer was designed to target the junction of the 878 bp and 389 bp regions to ensure specific amplification in ‘Hongsan’. Additionally, the SaH_513F primer was designed to encompass an approximately 18 bp gap at position 595 bp and an EcoR I restriction site at position 486 bp. This design was adopted to ensure that, even if the suspected translocation region involves highly repetitive sequences, differentiation would be possible either through the size difference resulting from the 18 bp gap (producing a longer amplicon in ‘Hongsan’) or via digestion at the EcoR I restriction site (Figure 8). The resulting primer pair, SaH_191R and SaH_513F (Table 2), was designed to specifically amplify a 545 bp fragment in the ‘Hongsan’ cultivar.
Figure 8.
The amplification positions of the SaH_191R, SaH_513F primers on the garlic nucleotide sequence in NCBI.
Figure 8.
The amplification positions of the SaH_191R, SaH_513F primers on the garlic nucleotide sequence in NCBI.
PCR analysis using the designed primers (Figure 9) successfully resulted in the amplification of a 545 bp band in the ‘Hongsan’ cultivar. This indicates that both primers target specific nucleotide sequences unique to the ‘Hongsan’ cultivar. Consequently, the SaH_191R and SaH_513F primers were confirmed as effective molecular markers for the identification of the ‘Hongsan’ cultivar.
As mentioned in the Introduction, ‘Hongsan’ was developed through a cross between the maternal line '8902' and the paternal line '9209'. Since the amplified region targeted by the developed primers was expected to be present in either the maternal or paternal parent, PCR was performed on both parental lines using the SaH_191R and SaH_513F primers (Figure 10). The results revealed a band in the paternal line that appeared slightly shorter than the band in ‘Hongsan’, confirming that this region was inherited from the paternal parent.
4. Discussion
RAPD-PCR analysis showed that 48 out of 60 Operon primers generated valid bands, with 13 primers (e.g., OPA-07, OPE-01) producing clear polymorphic bands specific to ‘Hongsan’ compared to the BSA pool (‘Daeseo’, ‘Uiseong’, ‘Danyang’, and ‘Namdo’). Notably, the OPE-01 primer consistently amplified a ~1.3 kb ‘Hongsan’-specific band, making it the prime candidate for cloning; this outcome underscores the stability of high GC content (70%) primers and the efficiency of the BSA strategy. Individual analysis of the five cultivars (Figure 4) showed similar band patterns among ‘Uiseong’, ‘Namdo’, and ‘Danyang’, reflecting high repetitive sequence homology despite ecological differences between northern-type (central region cultivation) and southern-type (‘Namdo’) cultivars [39], consistent with Kwon et al. [38] who reported challenges in distinguishing ecotypes via RAPD. In contrast, partial similarity between ‘Hongsan’ and ‘Daeseo’ suggests a shared southern-type background (early maturity; [13]), yet the OPE-01 band confirmed ‘Hongsan’-unique polymorphism suitable for cultivar identification.
In the cloning process, colony PCR of 48 white colonies (Figure 5) identified 1.3-1.4 kb bands, leading to plasmid extraction and sequencing from 18 colonies; the 1,123 bp sequence was excluded due to its repetitive nature (>30 sites on chromosome 1 with 98% similarity), while the 1,272 bp sequence (878 bp + 394 bp) emerged as the key marker. Assembly with the Staden Package [36] and NCBI BLAST analysis (GCA_030737875.1, chromosome 2: 4,795,106–4,796,000 bp) identified a putative translocation junction, 18 bp insertion (position 595 bp), and EcoRI site (position 486 bp) as ‘Hongsan’-specific variants; PCR of the paternal line 9209 (Figure 10) yielded a distinct amplicon smaller than that of ‘Hongsan’, confirming paternal inheritance while maintaining marker specificity through size polymorphism (2003 cross: 8902 × 9209). Plasmid loss in 2/18 cases [41] was likely due to prolonged incubation (16 h), but sufficient data were obtained from the 16 successful sequences.
Previous garlic marker studies employed RAPD for genetic diversity [28,42] or rust resistance screening, yet reproducibility and specificity limitations necessitated SCAR conversion. This study's RAPD-to-SCAR approach mirrors Sharma et al. [25] for Commiphora and Yap et al. [26] for Coelogyne SCAR markers, providing a novel cultivar-specific SCAR marker for the Korean garlic cultivar ‘Hongsan’ following Bae et al.'s [28] RAPD identification. Compared to SSR [29], AFLP [30], and SLAF-seq [31], RAPD-SCAR offers low cost (10 μL PCR), rapidity (40 cycles), and compatibility with simple equipment, optimizing it for the processing industry and overcoming phenotypic limitations (clove tip greenness lost upon drying; [16]).
SCAR primer design (SaH191R: junction-specific; SaH513F: gap/EcoRI-utilizing; Table 2) enhanced discrimination of repetitive sequences (GC 50-60%, Tm 59.8-64.1°C), achieving a 545 bp ‘Hongsan’-only amplicon (Figure 9). This demonstrates superior cultivar specificity over AFLP [18] or SSR [19] approaches and fulfills the RDA project (PJ01512803) goals for cultivar protection. Limitations include validation limited to five cultivars, necessitating multi-regional (national Allium Centers), multi-year testing, and international germplasm comparisons. Practically, it enables authentication of processed products (black garlic, bulbil items), UPOV rights enforcement, and export traceability.
5. Conclusions
This study successfully developed a reliable molecular marker (SaH191R/SaH513F, 545 bp amplicon) for specific identification of the Korean garlic cultivar ‘Hongsan’ from major cultivars (‘Daeseo’, ‘Uiseong’, ‘Danyang’, and ‘Namdo’) using BSA-aided RAPD analysis and SCAR conversion. Cloning and sequencing of the 1.3 kb polymorphic band from OPE-01 revealed ‘Hongsan’-specific variants in the 1,272 bp sequence—a putative translocation junction (878 bp + 394 bp), 18 bp insertion gap (position 595 bp), and EcoRI site (position 486 bp)—originating from the paternal parent line 9209, as confirmed by PCR (Figure 10). These genetic features reflect structural variations unique to ‘Hongsan’ relative to the NCBI garlic reference genome (GCA_030737875.1, chromosome 2), with exclusion of repetitive sequences (1,123 bp fragment) and accurate Staden Package assembly being pivotal to success.
The SCAR marker overcomes RAPD reproducibility limitations [28,38], enabling 100% specific detection of ‘Hongsan’ irrespective of environmental factors (cultivation/drying conditions), processing states (grinding, black garlic production), or seedling stages, fully addressing visual identification constraints (clove tip greenness; [16]). It is directly applicable to preventing cultivar mislabeling in the garlic processing industry (powders, fermented products, bulbil foods) and protecting breeder rights under UPOV (2015 registration).
Furthermore, this marker serves as foundational data for Korean garlic genetic diversity studies, complementing SSR/AFLP/SNP approaches [29,31] with its low-cost, rapid PCR (10 μL reaction) suited for field use. Although limited to five cultivars, future work should include expanded validation across >20 national/international germplasms (China, Central Asia), multi-year/multi-site reproducibility tests, and NGS-linked high-resolution markers. Commercialization efforts, such as real-time PCR kits, LAMP-based on-site diagnostics, and standardized protocols, are recommended to bolster global competitiveness and sustainable cultivar management in the Korean garlic industry.
Author Contributions
Conceptualization, K.J., and C.M.; Data curation, R.D. and A.S.; Formal analysis, K.J. and C.M.; Funding acquisition, K.J., and C.M.; Investigation, R.D. and A.S.; Methodology, K.J. and R.D.; Project administration, K.J. and C.M.; Resources, K.J. and C.M.; Software, R.D. and A.S.; Supervision, K.J.; Validation, C.M.; Visualization, R.D. and A.S.; Writing—original draft, R.D.; writing—review and editing K.J. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Rural Development Administration (Project No. PJ01512803) and by 2024 collaborative research program between university and Rural Development Administration, Republic of Korea. the Cooperative Research Program for Agriculture Science & Technology Development, Rural Development Administration, Republic of Korea.
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Choi, H.-S.; Kim, K.-T.; Ahn, Y.-G.; Kim, D.-S.; Woo, J.-G.; Lim, Y.-P. Analysis of Genetic Relationships in Garlic Germplasm and Fertile Garlic by RAPD. Horticulture, Environment, and Biotechnology 2003, 44(5), 595–600. Available online: https://zrr.kr/zkjVRG.
- Rural Development Administration (RDA). Agricultural Technology Guide_117_Garlic. Available online: https://zrr.kr/aSZMVo (accessed on 02 Dec 2025).
- Ahn, Y. K.; Yoon, M. K. Effect of Short Daylength and Temperature Control on Garlic Florogenesis. Horticultural Science and Technology 2010, 28(2), 180–185. Available online: https://www.dbpia.co.kr/journal/articleDetail?nodeId=NODE11877595.
- Etoh, T. Germination of Seeds Obtained from a Clone of Garlic, Allium sativum L. Proceedings of the Japan Academy 1983, 59 (B), 83–87. [Google Scholar] [CrossRef]
- Katarzhin, M. S.; Katarzhin, I. M. Experiments on the Sexual Reproduction of Garlic. Byulleten' Vsesoyuznogo Ordena Lenina I Ordena Druzhby Narodov Instituta Rastenievodstva Imeni N.I. Vavilov 1978, 80, 74–76. Available online: https://www.cabidigitallibrary.org/doi/full/10.5555/19801687714.
- Etoh, T.; Noma, Y.; Nisfutarumizu, Y.; Wakomoto, T. Seed Productivity and Germinability of Various Clones Collected in Soviet Central Asia. Memoirs of the Faculty of Agriculture, Kagoshima University 1988, 24, 29–139. Available online: https://www.jstage.jst.go.jp/article/jjshs1925/55/3/55_3_312/_pdf.
- Dhall, R. K.; Cavagnaro, P. F.; Singh, H.; Mandal, S. History, Evolution and Domestication of Garlic: A Review. Plant Syst Evol 2023, 309, 33. [Google Scholar] [CrossRef]
- Pooler, M. R.; Simon, P. W. Characterization and Classification of Isozyme and Morphological Variation in a Diverse Collection of Garlic Clones. Euphytica 1993, 68, 121–130. [Google Scholar] [CrossRef]
- Choi, H. S.; Chae, W. B.; Kwack, Y. B.; Jeong, M. I. A New Early Harvest Garlic Cultivar 'Allkae'. Korean Journal of Horticultural Science and Technology 2008, 26 (II), 9–12. Available online: https://www.dbpia.co.kr/journal/articleDetail?nodeId=NODE11877429.
- Yoon, M.; Ahn, Y.; Chae, S. Development of Garlic Cultivar (Allium sativum L.) Using Fertile Garlic. Horticultural Science and Technology 2007, 25 (I), 79–79. Available online: https://www.dbpia.co.kr/journal/articleDetail?nodeId=NODE11871658.
- Kim, J.-S.; Ra, J. H. Comparison of Phytochemical Composition and Physiological Activity of ‘Hongsan’ and 'Hansan', a New Variety of Garlic. Korean Journal of Food Science and Technology 2019, 51(2), 147–151. [Google Scholar] [CrossRef]
- Park, J.-M.; Kim, A.-J. Evaluation of the Biological Activities of ‘Hongsan’ Garlic Bulbil. Journal of the Korean Society of Food Science and Nutrition 2023, 52(6), 773–779. [Google Scholar] [CrossRef]
- Kwak, J.-H.; Ahn, Y. K.; Kim, C.-W.; Kwon, Y.-S.; Choi, K. J.; Yoon, M. K. New Garlic Variety ‘Hongsan’ for High Yield and Nationwide Cultivation. Korean Journal of Horticultural Science and Technology 2016, 34 (I), 115–115. Available online: https://www.dbpia.co.kr/journal/articleDetail?nodeId=NODE06685275.
- Lee, J.; Yoon, S.; Lee, M.; Kwon, J.; Hong, K. Effect of Bulblet Harvest Timings on Bulb and Bulblet Yield of Garlic cv. ‘Hongsan’. Korean Journal of Horticultural Science and Technology 2018, 36 (II), 79–79. Available online: https://www.dbpia.co.kr/journal/articleDetail?nodeId=NODE07555674.
- Han, J. W.; Kwon, Y. S.; Kim, C. W.; Kwak, J. H. Changes on the Yield and Components in New Garlic Variety ‘Hongsan’ According to Preharvesting Treatment and Harvesting Season. Korean Journal of Horticultural Science and Technology 2017, 35 (I), 66–66. Available online: https://www.dbpia.co.kr/journal/articleDetail?nodeId=NODE07181725.
- Luitel, B. P.; Lee, H. J.; Choi, M. S.; Lim, T. J. Effects of Planting Depth and Sunshine Drying on Expression of Greenness at the Tip of Peeled ‘Hongsan’ Garlic Cloves. Korean Journal of Environmental Agriculture 2024, 43, 128–136. [Google Scholar] [CrossRef]
- Jenderek, M. M.; Schierenbeck, K.; Hannan, R. M. Random Amplified Polymorphic DNA Analysis of Garlic (Allium sativum L.) Germplasm Collection. HortScience 1997, 32(3), 452F. [Google Scholar] [CrossRef]
- Wang, H.; Li, X.; Liu, X.; Qiu, Y.; Song, J.; Zhang, X. Genetic Diversity of Garlic (Allium sativum L.) Germplasm from China by Fluorescent-Based AFLP, SSR and InDel Markers. Plant Breed 2016, 135, 743–750. [Google Scholar] [CrossRef]
- Ipek, M.; Sahin, N.; Ipek, A.; Cansev, A.; Simon, P. W. Development and Validation of New SSR Markers from Expressed Regions in the Garlic Genome. In Scientia Agricola; FapUNIFESP (SciELO), 2015. [Google Scholar] [CrossRef]
- Mane-Deshmukh, V.; Ghodake, B.; Kharpude, P.; Patil, A. Morphological and Molecular Characterization of Onion (Allium cepa L.) Genotypes Using RAPD Markers. The Pharma Innovation 2023, 12(2), 1925–1930. [Google Scholar] [CrossRef]
- Devos, K. M.; Gale, M. The Use of Random Amplified Polymorphic DNA Markers in Wheat. Theoretical and Applied Genetics 1992, 84, 567–572. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, J.; Chen, Q.; Chang, Y.; Du, J.; Meng, H. Analysis of the Genetic Diversity of Garlic (Allium sativum L.) Germplasm by SRAP. Biochemical Systematics and Ecology 2013, 50, 139–146. [Google Scholar] [CrossRef]
- Cabrita, L. F.; Aksoy, U.; Hepaksoy, S.; Leitão, J. M. Suitability of Isozyme, RAPD and AFLP Markers to Assess Genetic Differences and Relatedness Among Fig (Ficus carica L.) Clones. Scientia Horticulturae 2001, 87(4), 261–273. [Google Scholar] [CrossRef]
- Thormann, C. E.; Ferreira, M. E.; Camargo, L. E.; Tivang, J. G.; Osborn, T. C. Comparison of RFLP and RAPD Markers to Estimating Genetic Relationships Within and Among Cruciferous Species. Theoretical and Applied Genetics 1994, 88(8), 973–980. [Google Scholar] [CrossRef]
- Sairkar, P. K.; Sharma, A.; Shukla, N. P. SCAR Marker for Identification and Discrimination of Commiphora wightii and C. myrrha. Molecular Biology International 2016, 1482796. [Google Scholar] [CrossRef] [PubMed]
- Kok Hon, Y.; Yong, C. S.-Y.; Abdullah, J. O.; Go, R. Development of Species-Specific SCAR Markers for Identification and Authentication of Three Rare Peninsular Malaysian Endemic Coelogyne (Orchidaceae) Orchids. F1000Research 2021, 9, 1161. [Google Scholar] [CrossRef]
- Al-Mathidy, A. M.; Al-Talab, N. N.; Kharabe, K. B. E. Identification of Garlic (Allium sativum L.) Cultivars by Using Morphological and Chemical Characters. NVEO-NATURAL VOLATILES and ESSENTIAL OILS Journal 2021, 8893–8907. Available online: https://zrr.kr/sZTGW7.
- Bae, S.-K.; Jung, E.-A.; Kwon, S.-T. Genetic Variation and Identification of RAPD Markers from Some Garlic Cultivars in Korea and Mongolia. Korean Journal of Plant Resources 2010, 23(5), 458–464. Available online: https://koreascience.or.kr/article/JAKO201006159731691.page.
- Li, X.; Qiao, L.; Chen, B.; Zheng, Y.; Zhi, C.; Zhang, S.; Pan, Y.; Cheng, Z. SSR Markers Development and Their Application in Genetic Diversity Evaluation of Garlic (Allium sativum L.) Germplasm. Plant Diversity 2022, 44(5), 481–491. [Google Scholar] [CrossRef] [PubMed]
- Morales, R. G.; Resende, J. T.; Resende, F. V.; Delatorre, C. A.; Figueiredo, A. S.; Da-Silva, P. R. Genetic Divergence Among Brazilian Garlic Cultivars Based on Morphological Characters and AFLP Markers. Genetics and Molecular Research 2013, 12(1), 270–281. Available online: https://pubmed.ncbi.nlm.nih.gov/23408414/. [CrossRef]
- Yang, J.; Sun, M.; Ren, X.; Li, P.; Hui, J.; Zhang, J.; Lin, G. Revealing the Genetic Diversity and Population Structure of Garlic Resource Cultivars and Screening of Core Cultivars Based on Specific Length Amplified Fragment Sequencing (SLAF-Seq). Genes 2024, 15(9), 1135. [Google Scholar] [CrossRef]
- Alimoradi Askar, S.; Nasr Esfahani, M.; Shirazi, K.; Nasr Esfahani, A.; Zeinalzadeh-Tabrizi, H.; Mohammadi, M. Unveiling Genetic Variation in Garlic Genotypes in Response to Rust Disease Using RAPD Markers. OBM Genetics 2024, 8(2), 231. [Google Scholar] [CrossRef]
- Staden, R. The Staden Sequence Analysis Package. Molecular Biotechnology 1996, 5, 233–241. [Google Scholar] [CrossRef]
- Staden, R.; Beal, K. F.; Bonfield, J. K. The Staden Package, 1998. Bioinformatics Methods and Protocols 1998, 115–130. [Google Scholar] [CrossRef]
- Bonfield, J.; Beal, K.; Jordan, M.; Cheng, Y. The Staden Package Manual; Medical Research Council, Laboratory of Molecular Biology: Cambridge, UK, 1999. [Google Scholar]
- Lee, Y.-J.; Lee, J.-B.; Lim, G.-S.; Kim, B.-K.; Kim, J.-H.; Park, H.-B. Analysis of Sanger Sequencing Data Using the Open-Source STADEN Package for Single Nucleotide Polymorphism Detection. Journal of Animal Breeding and Genomics 2023, 7(1), 1–8. Available online: https://www.jabg.org/0701-01/.
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B. C.; Remm, M.; Rozen, S. G. Primer3—New Capabilities and Interfaces. Nucleic Acids Research 2012, 40(15), e115. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.-T.; Oh, S.-M. Genetic Relationship Among Garlic Cultivars Based on RAPD Analysis. Journal of Life Science 1999, 9(6), 671–676. Available online: https://koreascience.kr/article/JAKO199911919955413.page.
- Eom, E. M.; Lee, D. H. Characterization of Chromosomal DNA and DNA Polymorphism in Korean Cultivars of Allium sativum L. Journal of Plant Biology 1999, 42, 159–167. [Google Scholar] [CrossRef]
- Bergkessel, M.; Guthrie, C. Colony PCR. Methods in Enzymology 2013, 529, 299–309. [Google Scholar] [CrossRef]
- Popov, M.; Petrov, S.; Nacheva, G.; Ivanov, I.; Reichl, U. Effects of Recombinant Gene Expression on ColE1-like Plasmid Segregation in Escherichia coli. BMC Biotechnology 2011, 11, 1–12. [Google Scholar] [CrossRef]
- Nabulsi, I.; Al-Safadi, B.; Ali, N. M.; Arabi, M. I. Evaluation of Some Garlic (Allium sativum L.) Mutants Resistant to White Rot Disease by RAPD Analysis. Annals of Applied Biology 2001, 138, 197–202. [Google Scholar] [CrossRef]
Figure 1.
PCR results of primers 1 to 11 for ‘Hongsan’ and BSA samples. M 1kp Size Marker, m 100bp Size Marker, {1} OPA-07, {2} OPA-14, {3} OPE-16, {4} OPB-18, {5} OPE-15, {6} OPB-06, {7} OPB-13, {8} OPE-01, {9} OPA-17, {10} OPE-08, {11} OPE-09.
Figure 1.
PCR results of primers 1 to 11 for ‘Hongsan’ and BSA samples. M 1kp Size Marker, m 100bp Size Marker, {1} OPA-07, {2} OPA-14, {3} OPE-16, {4} OPB-18, {5} OPE-15, {6} OPB-06, {7} OPB-13, {8} OPE-01, {9} OPA-17, {10} OPE-08, {11} OPE-09.

Figure 2.
PCR results of primers 12 to 33 for ‘Hongsan’ and BSA samples. {M} 1kp Size Marker, m 100bp Size Marker, {12} OPE-09, {13} OPE-11, {14} OPC-05, {15} OPE-17, {16} OPE-18, {17} OPE-20, {18} OPH-01, {19} OPC-04, {20} OPH-04, {21} OPF-03, {22} OPG-18, {23} OPG-19, {24} OPA-02, {25} OPB-01, {26} OPA-04, {27} OPA-08, {28} OPA-09, {29} OPA-10, {30} OPC-06, {31} OPA-12, {32} OPE-14, {33} OPA-15.
Figure 2.
PCR results of primers 12 to 33 for ‘Hongsan’ and BSA samples. {M} 1kp Size Marker, m 100bp Size Marker, {12} OPE-09, {13} OPE-11, {14} OPC-05, {15} OPE-17, {16} OPE-18, {17} OPE-20, {18} OPH-01, {19} OPC-04, {20} OPH-04, {21} OPF-03, {22} OPG-18, {23} OPG-19, {24} OPA-02, {25} OPB-01, {26} OPA-04, {27} OPA-08, {28} OPA-09, {29} OPA-10, {30} OPC-06, {31} OPA-12, {32} OPE-14, {33} OPA-15.
Figure 3.
PCR results of primers 34 to 48 for ‘Hongsan’ and BSA samples. {M} 1kp Size Marker, m 100bp Size Marker, {34} OPA-16, {35} OPA-18, {36} OPA-01, {37} OPA-11, {38} OPA-20, {39} OPC-09, {40} OPC-13, {41} OPD-01, {42} OPD-03, {43} OPH-02, {44} OPE-03, {45} OPE-12, {46} OPB-06, {47} OPG-02, {48} OPG-03.
Figure 3.
PCR results of primers 34 to 48 for ‘Hongsan’ and BSA samples. {M} 1kp Size Marker, m 100bp Size Marker, {34} OPA-16, {35} OPA-18, {36} OPA-01, {37} OPA-11, {38} OPA-20, {39} OPC-09, {40} OPC-13, {41} OPD-01, {42} OPD-03, {43} OPH-02, {44} OPE-03, {45} OPE-12, {46} OPB-06, {47} OPG-02, {48} OPG-03.
Figure 4.
Thirteen primers, selected from a set of 48 random primers, were used for PCR analysis of five garlic cultivars: ‘Hongsan’', ‘Daeseo’, ‘Uiseong’, ‘Namdo’, and ‘Danyang’. M 100bp Size Marker, {1} OPB-01, {2} OPA-10, {3} OPE-14, {4} OPA-07, {5} OPE-16, {6} OPA-14, {7} OPB-13, {8} OPE-01, {9} OPA-17, {10} OPA-01, {11} OPA-11, {12} OPA-20, {13} OPH-02, {A} ‘Hongsan’, {B} ‘Daeseo’, {C} ‘Uiseong’. {D} ‘Danyang’. {E} ‘Namdo’, {b} Blank.
Figure 4.
Thirteen primers, selected from a set of 48 random primers, were used for PCR analysis of five garlic cultivars: ‘Hongsan’', ‘Daeseo’, ‘Uiseong’, ‘Namdo’, and ‘Danyang’. M 100bp Size Marker, {1} OPB-01, {2} OPA-10, {3} OPE-14, {4} OPA-07, {5} OPE-16, {6} OPA-14, {7} OPB-13, {8} OPE-01, {9} OPA-17, {10} OPA-01, {11} OPA-11, {12} OPA-20, {13} OPH-02, {A} ‘Hongsan’, {B} ‘Daeseo’, {C} ‘Uiseong’. {D} ‘Danyang’. {E} ‘Namdo’, {b} Blank.
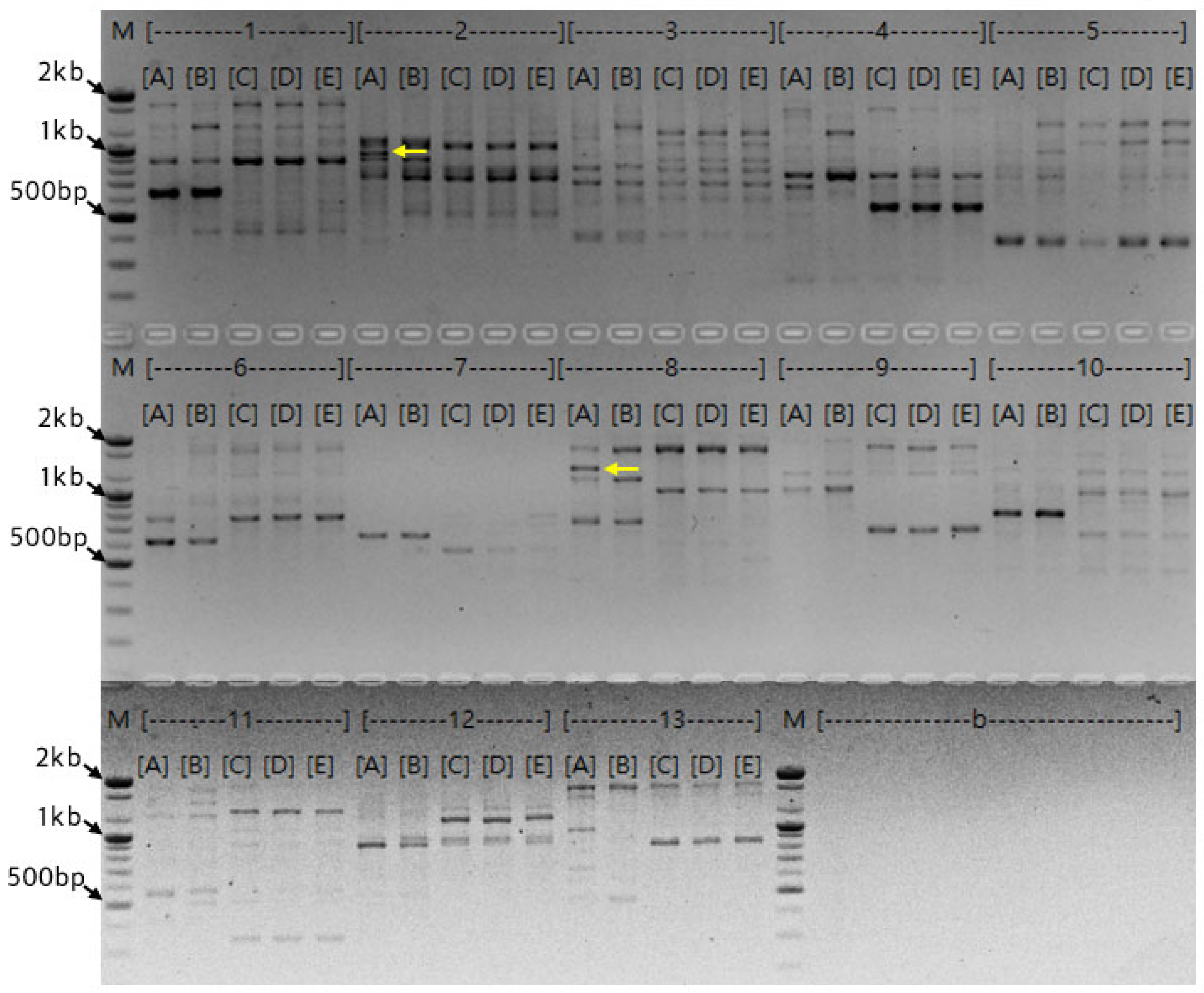
Figure 5.
PCR results of 48 colonies using the M13 primer for colony PCR. M 100bp Size Marker, {1}~ {48} OPE-01-H colony 1~ 48.
Figure 5.
PCR results of 48 colonies using the M13 primer for colony PCR. M 100bp Size Marker, {1}~ {48} OPE-01-H colony 1~ 48.
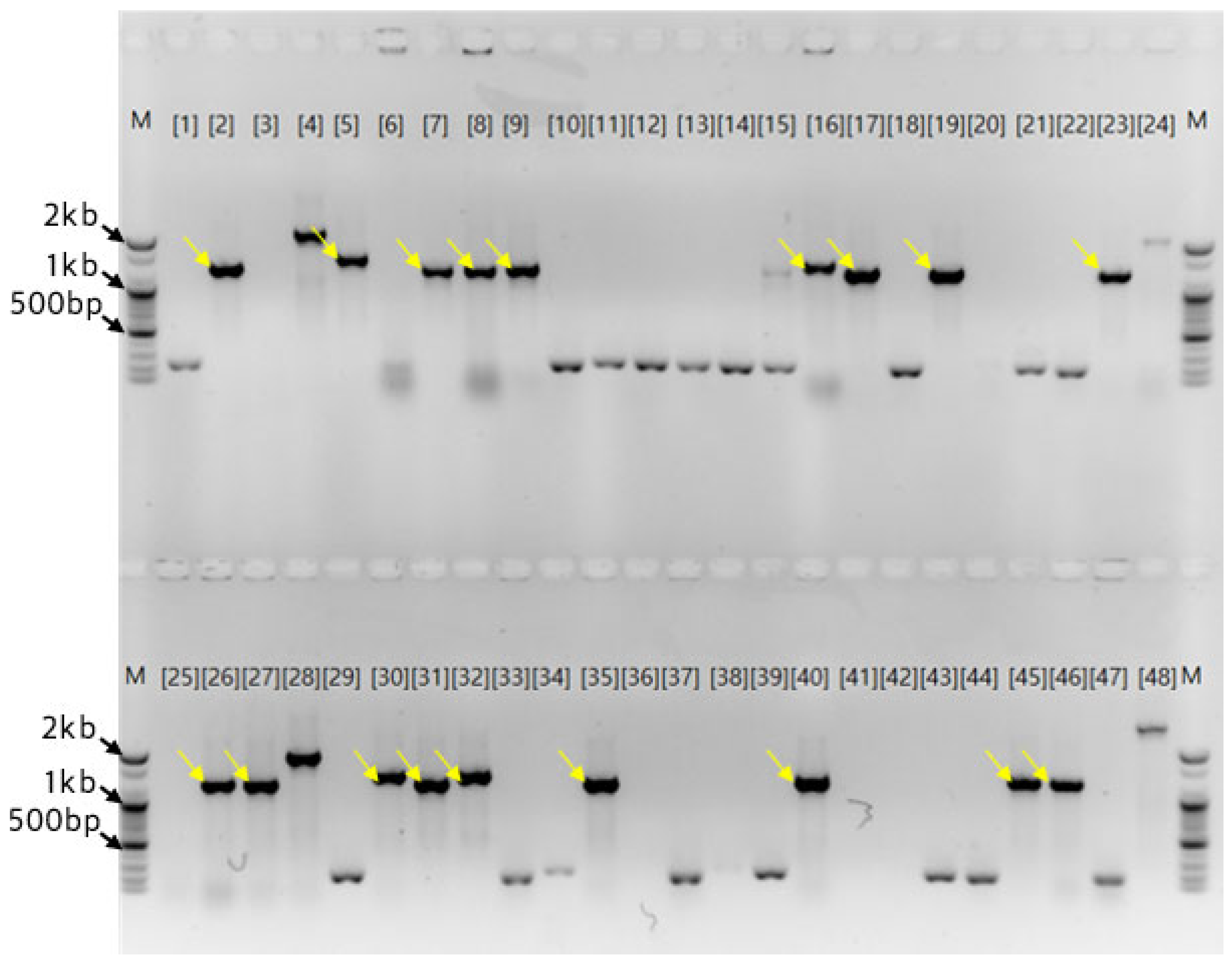
Figure 7.
Schematic diagram of the two nucleotide sequences detected in the plasmid from ‘Hongsan’. (A) Schematic of the binding site on chromosome 1 when a 1172 bp nucleotide sequence is compared to the garlic nucleotide sequence in NCBI. (B) Schematic of the comparison of a 1272 bp nucleotide sequence with the garlic nucleotide sequence in NCBI. (C) Schematic of the alignment of a 1272 bp nucleotide sequence with the matching position in the garlic nucleotide sequence in NCBI.
Figure 7.
Schematic diagram of the two nucleotide sequences detected in the plasmid from ‘Hongsan’. (A) Schematic of the binding site on chromosome 1 when a 1172 bp nucleotide sequence is compared to the garlic nucleotide sequence in NCBI. (B) Schematic of the comparison of a 1272 bp nucleotide sequence with the garlic nucleotide sequence in NCBI. (C) Schematic of the alignment of a 1272 bp nucleotide sequence with the matching position in the garlic nucleotide sequence in NCBI.
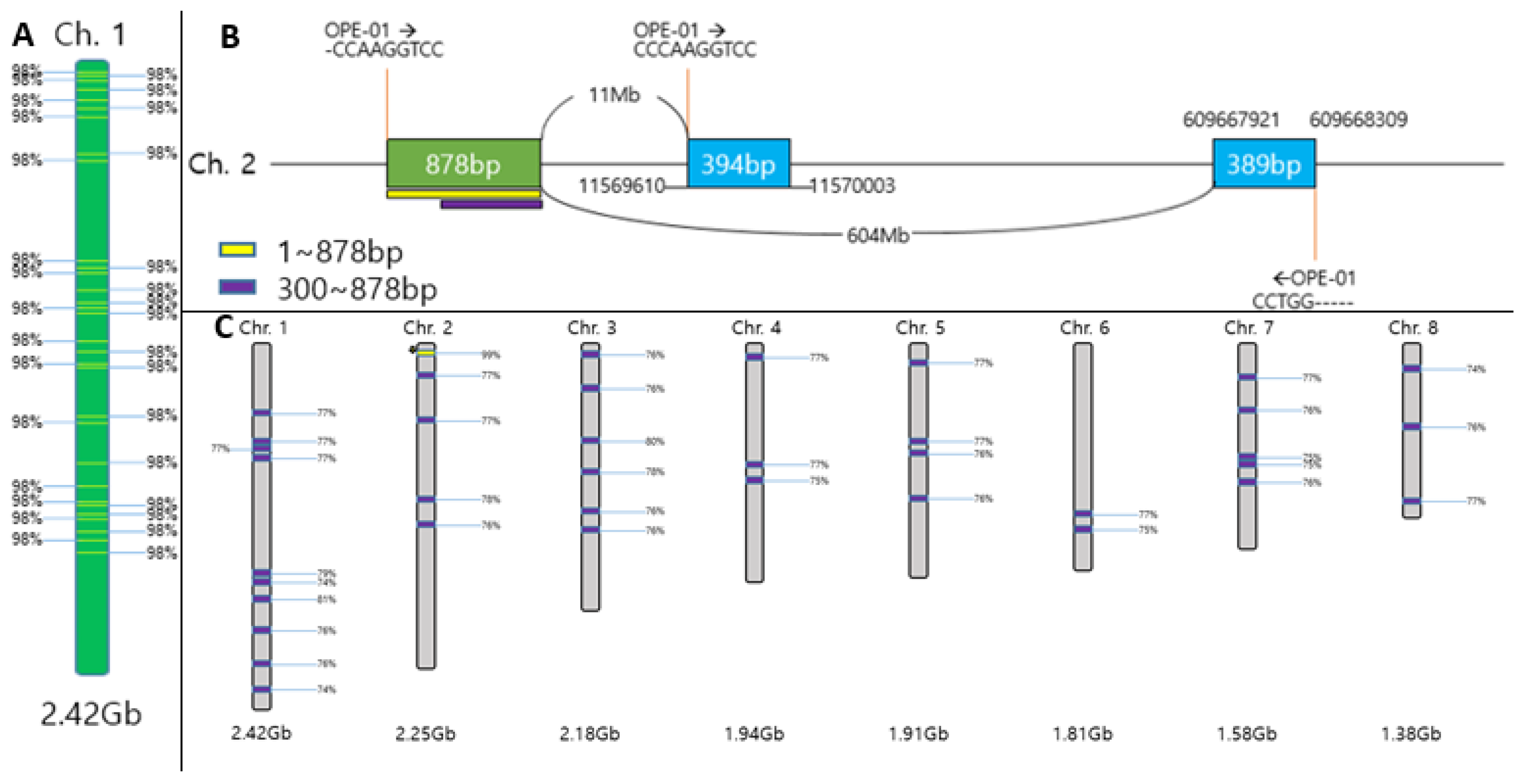
Figure 9.
The result of PCR using the primers SaH_191R and SaH_513F on the garlic cultivars ‘Hongsan’, ‘Daeseo’, ‘Uiseong’, ‘Namdo’, and ‘Danyang’. {M} 100bp Size Marker, {1} ‘Hongsan’, {2} ‘Daeseo’, {3} ‘Uiseong’, {4} ‘Namdo’, {5} ‘Danyang’.
Figure 9.
The result of PCR using the primers SaH_191R and SaH_513F on the garlic cultivars ‘Hongsan’, ‘Daeseo’, ‘Uiseong’, ‘Namdo’, and ‘Danyang’. {M} 100bp Size Marker, {1} ‘Hongsan’, {2} ‘Daeseo’, {3} ‘Uiseong’, {4} ‘Namdo’, {5} ‘Danyang’.
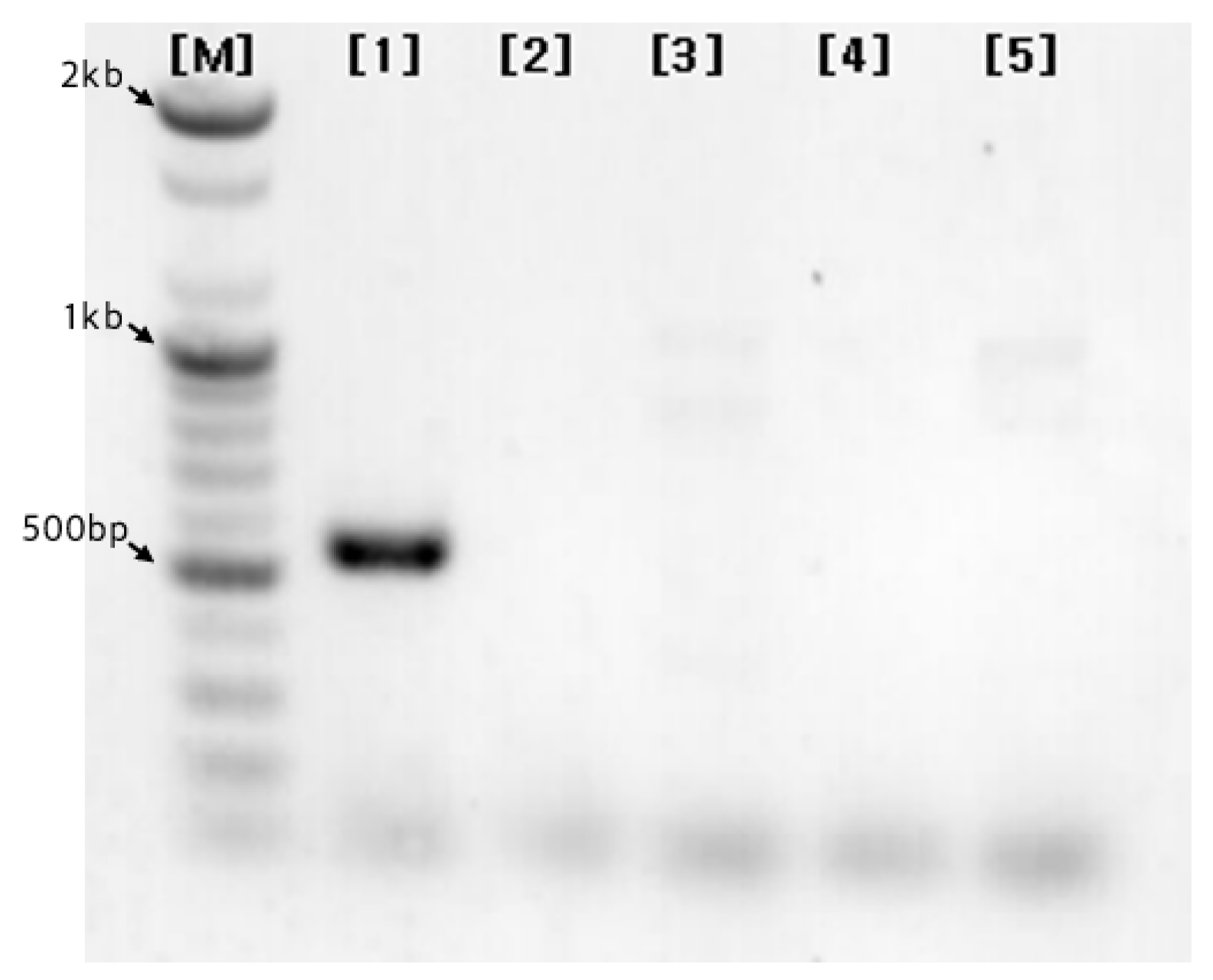
Figure 10.
The result of PCR using the primers SaH_191R and SaH_513F on the maternal line '8902' and the paternal line '9209' of the garlic cultivar ‘Hongsan’ was as follows. {M} 100bp Size Marker, {1} Seed parent 8902, {2} Pollen parent 9209.
Figure 10.
The result of PCR using the primers SaH_191R and SaH_513F on the maternal line '8902' and the paternal line '9209' of the garlic cultivar ‘Hongsan’ was as follows. {M} 100bp Size Marker, {1} Seed parent 8902, {2} Pollen parent 9209.
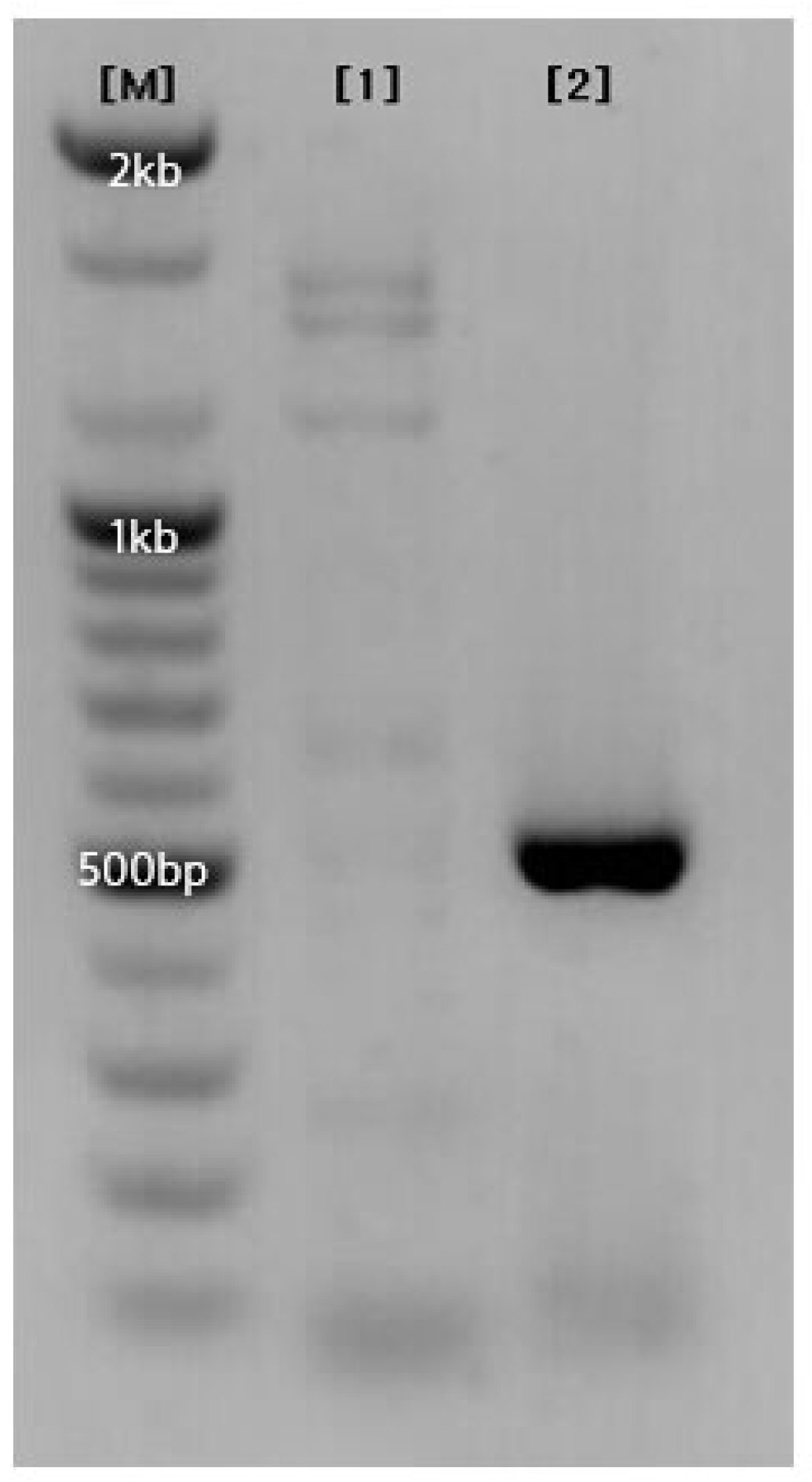
Table 2.
Information on the developed molecular marker.
| Name | Sequence(5’-3’) | GC(%) | Tm(℃) | mer |
| SaH_191R | GGTGTTTCATTTGCAAGGCC | 50 | 59.8 | 20 |
| SaH_513F | CTCGGAGGAAGGTAATGCCC | 60 | 64.1 | 20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



