Submitted:
21 January 2026
Posted:
23 January 2026
You are already at the latest version
Abstract
To decipher the molecular response mechanism of melon to saline-alkali stress, seedlings of the melon cultivar ‘Xikaixin’ were treated with 50 mmol·L⁻¹ mixed solutions of NaCl and NaHCO₃ at ratios of 1:1, 1:2, and 2:1 to simulate saline-alkali stress. Transcriptome sequencing of roots yielded 78.98 Gb of clean data (≥6.02 Gb per sample) with Q30 ≥96.61% and genome alignment rates of 97.00%-98.02%, identifying 588, 686, and 1107 differentially expressed genes (DEGs) in the 1:1, 1:2, and 2:1 groups, respectively. DEGs were significantly enriched in 50 pathways categorized into 5 major classes, with the plant hormone signal transduction pathway showing the highest enrichment across all treatments. A key original finding is the distinct balance shift between auxin and ABA: two auxin-related genes (auxin-induced protein gene MELO3C013403 and auxin response factor gene MELO3C004381) were specifically upregulated (≥2-fold vs. control) in the high-salt 2:1 group, while ABA-related genes were upregulated and auxin/jasmonic acid/gibberellin-related genes were downregulated in the 1:2 group, indicating a cultivar-specific hormone balance modulation in ‘Xikaixin’ under saline-alkali stress. In contrast, photosynthesis-antenna protein genes (e.g., MELO3C021567) were significantly downregulated (to 32% of the control) under the 2:1 treatment. RT-PCR validation confirmed the consistency of these candidate genes’ expression with transcriptomic data. Therefore, melon may respond to saline-alkali stress by regulating the plant hormone signal transduction (especially Auxin-ABA balance), photosynthesis, and carbon metabolism pathways. This study provides novel candidate genes and a theoretical basis for the genetic improvement of saline-alkali-tolerant melon cultivars, with the unique Auxin-ABA balance modulation as a key original contribution.
Keywords:
melon (cucumis melo)
; seedling
; saline-alkaline stress
; transcriptome analysis
1. Introduction
Soil salinization is a global abiotic stress factor that severely impacts plant growth and agricultural production[1]. In China, the total area of salinized land reaches 3.7×10⁷ hm², with Xinjiang being the most severely affected region, accounting for 2.96×10⁶ hm² of salinized land, of which 40% is moderately salinized arable land with development potential [2]. Due to natural conditions such as low rainfall, arid climate, intense sunlight, and high evaporation, soil salinization is particularly prominent in southern Xinjiang. Currently, saline-alkali land improvement mainly adopts physical, chemical, and biological methods. However, due to the large area of salinized land, physical and chemical improvement methods are limited by long time consumption, high labor intensity, and high costs, making large-scale application difficult. In contrast, the biological improvement method of planting saline-alkali-tolerant crops on saline-alkali land has significant advantages. On one hand, saline-alkali-tolerant crops can absorb saline-alkali components from the soil, improve soil structure, and promote the development and utilization of salinized land [3,4]. On the other hand, studying saline-alkali-tolerant crops on moderately mixed saline-alkali land is of great significance for revealing their saline-alkali tolerance mechanisms and exploring excellent saline-alkali-tolerant germplasm resources [5].
A survey on the ion composition of saline-alkali soil in southern Xinjiang showed that the content ratio of NaCl to NaHCO₃ is approximately 1:0.92, which is close to an equal ratio of 1:1 [6]. Based on this measured ratio, saline-alkali stress experiments often use three gradients: NaCl:NaHCO₃ = 1:1, 1:2 (higher proportion of alkaline salt), and 2:1 (dominated by sodium chloride) to cover the full range of salinity from weakly alkaline to strongly alkaline in natural soil. In studies on saline-alkali stress treatment of crops such as peanuts, sweet sorghum, and alfalfa, mixed salts were directly prepared at ratios of 1:1 and 2:1 [6,7]. In combined saline-alkali experiments on Achnatherum inebrians and rice fields, ratios such as NaCl:Na₂SO₄:NaHCO₃ were used, where the proportion of alkaline salt (NaHCO₃) gradually increased, equivalent to a gradient transition of NaCl:NaHCO₃ from 1:1 to 1:2 [8,9]. The setting of these gradients follows the experimental design logic of “gradually increasing the proportion of alkaline salt” and has been verified in various crops (e.g., sweet sorghum, Achnatherum inebrians, rice, alfalfa), enabling systematic evaluation of the effects of different NaCl/NaHCO₃ ratios on seed germination, seedling growth, and physiological indicators. Therefore, selecting the three gradients of NaCl:NaHCO₃ = 1:1, 1:2, and 2:1 not only conforms to the actual ion ratio of saline-alkali soil in southern Xinjiang but also covers the main variation range of saline-alkali stress intensity, providing sufficient scientific basis and pertinence.
Melon (Cucumis melo L.) is an annual trailing plant of the genus Cucumis in the Cucurbitaceae family, with abundant germplasm resources and high nutritional value, making it popular among consumers [10]. Studies have shown that melon has a certain degree of tolerance to saline-alkali stress [11]. The seedling stage is a critical period for plants to respond to saline-alkali stress, directly affecting their adaptability to saline-alkali environments and subsequent growth and development [12]. Under saline-alkali stress, seedlings experience oxidative stress, which inhibits plant growth and biomass accumulation, and interferes with important physiological processes such as osmotic regulation, ion absorption, and photosynthesis [13].
Existing transcriptomic studies on melon have focused on different organs, tissues, and stress durations: Root transcriptome analyses mainly explored responses to short-term (24–72 h) salt stress (e.g., NaCl alone) or drought stress, identifying ion transport and osmotic adjustment-related genes [14,15]; leaf transcriptomes focused on long-term (1–2 weeks) abiotic stress (e.g., high temperature, drought) and biotic stress (e.g., pathogen infection), highlighting photosynthesis and defense response pathways [16]; fruit transcriptomes primarily investigated ripening processes under non-stress conditions, with limited involvement in stress responses [17,18]; and seedling-stage transcriptomic studies were mostly limited to single salt (NaCl) or alkali (NaHCO₃) stress, lacking systematic analysis of mixed saline-alkali stress which is closer to natural soil conditions [19,20].
Many researchers have studied the mechanism of plant resistance to saline-alkali stress through transcriptomic analysis. Transcriptomic analysis of poplar under simulated saline-alkali stress found that saline-alkali stress mainly affects plant signal transduction pathways (e.g., plant-pathogen interaction, plant hormone signal transduction), metabolic pathways (e.g., starch and sucrose metabolism, carbon metabolism), and biosynthetic pathways (e.g., biosynthesis of phenylpropanoids, biosynthesis of amino acids) to resist saline-alkali stress [21]. Transcriptomic analysis of two foxtail millet materials under saline-alkali stress revealed that foxtail millet enhances its saline-alkali tolerance through positive regulation of tryptophan/fatty acid metabolism, MAPK signaling pathway, and peroxisome pathways [22]. Another study showed that the expression of the Na⁺/H⁺ antiporter gene CsNHX1 in cucumber can sequester Na⁺ in vacuoles, thereby reducing Na⁺ content in cells and alleviating salt stress damage to cucumber seedlings [23]. The GhAKT1 gene is a K⁺ channel gene, and under salt and alkali stress, phosphorus application can significantly increase the relative expression level of the GhAKT1 gene in cotton [24].
In summary, saline-alkali stress enhances plant tolerance to saline-alkali stress by regulating the expression levels of genes in different signaling pathways and the metabolic levels of different substances. Currently, transcriptomic studies on melon’s response to saline-alkali stress are relatively limited, and existing melon transcriptomic studies lack targeted analysis of mixed saline-alkali stress with gradient ratios simulating natural soil conditions in southern Xinjiang. The unique difference of this study is that it uses the local saline-alkali-tolerant melon cultivar ‘Xikaixin’ in southern Xinjiang, targets the seedling root (the primary organ sensing saline-alkali stress), and simulates three gradient ratios of NaCl-NaHCO₃ mixed stress (1:1, 1:2, 2:1) consistent with the actual ion composition of local saline-alkali soil, to systematically decipher the cultivar-specific and ratio-dependent molecular response mechanism. Therefore, this study used the saline-alkali-tolerant melon cultivar ‘Xikaixin’ as experimental material, simulated saline-alkali stress treatment by setting different saline-alkali ratios (NaCl:NaHCO₃ = 1:1, 1:2, 2:1), and conducted transcriptomic analysis of melon roots to explore the signaling pathways and related genes involved in melon’s response to saline-alkali stress, and to deeply analyze the mechanism of melon’s response to saline-alkali stress.
2. Materials and Methods
2.1. Experimental Materials
The main local melon cultivar ‘Xikaixin’ in Xinjiang (bred by Xinjiang Jifeng Seed Industry Co., Ltd.) was used as the experimental material. This cultivar is a mid-early maturing type with oblong fruits, yellowish-red peel, and greenish-white flesh, and is resistant to transportation. All experimental seeds were purchased from the Modern Melon and Fruit Seed Marketing Store in Kashgar Agricultural Expo City to ensure they are the main cultivated melon cultivars in southern Xinjiang.
The main local melon cultivar ‘Xikaixin’ in Xinjiang (bred by Xinjiang Jifeng Seed Industry Co., Ltd.) was used as the experimental material. This cultivar is a mid-early maturing type with oblong fruits, yellowish-red peel, and greenish-white flesh, and is resistant to transportation. All experimental seeds were purchased from the Modern Melon and Fruit Seed Marketing Store in Kashgar Agricultural Expo City to ensure they are the main cultivated melon cultivars in southern Xinjiang.
The potting mix (substrate) was a commercial nursery substrate (purchased from Xinjiang Tianyuan Horticultural Co., Ltd.) with the following properties: organic matter content 35.2 ± 2.1 g·kg⁻¹, initial pH 6.8 ± 0.2 (1:5 substrate:water extract).
2.2. Saline-Alkaline Stress Treatment Protocol
Plump and uniform-sized ‘Xikaixin’ melon seeds were disinfected with 5% NaClO solution for 15 min and rinsed thoroughly with distilled water. Subsequently, the seeds were subjected to secondary disinfection by water bath at 50℃ for 20 minutes. The disinfected seeds were sown in plastic flowerpots (17 cm×19 cm), with 8 seeds per pot, and the humidity of the nutrient soil was maintained at 60–70% of field capacity during cultivation (monitored by a soil moisture sensor, type: TDR-300). After seed germination, at the cotyledon expansion stage when the first true leaf emerged, 4 robust seedlings were retained per pot. When the seedlings grew to the three-leaf-one-heart stage, saline-alkali stress treatment was initiated.
The control group (CK) was irrigated with a standard half-strength Hoagland nutrient solution (pH 6.5 ± 0.1, EC 1.2 ± 0.1 mS·cm⁻¹) to avoid nutrient imbalance, while the treatment groups were irrigated with 50 mmol·L⁻¹ mixed saline-alkali solutions (NaCl:NaHCO₃ = 1:1, 1:2, 2:1) prepared with the same half-strength Hoagland nutrient solution (ensuring consistent nutrient composition across all groups). The three treatment groups were designated as T1 (NaCl:NaHCO₃ = 1:1), T2 (NaCl:NaHCO₃ = 1:2), and T3 (NaCl:NaHCO₃ = 2:1). Each pot was irrigated with 500 mL of the corresponding solution (CK or treatment) at one time, and subsequent irrigation (500 mL per pot) was performed every 3 days for a total of 3 treatments, with a total treatment duration of 9 days.
During cultivation, the growth environment was controlled as follows: day-night temperature 25–28℃, relative humidity 34–42%, light intensity 350 μmol·m⁻²·s⁻¹, and light cycle 12 h/d. The experiment adopted a completely randomized block design (CRBD) with 4 biological replicates per group (n=4), and each replicate consisted of 1 pot (experimental unit). Pot placement was randomized within the growth chamber to eliminate positional effects (e.g., uneven light, temperature). After 9 days of saline-alkali treatment, roots from the same growth position (5 cm below the root crown) of each seedling in the treatment and control groups were collected, mixed uniformly per pot (replicate), and used for index determination. During sample collection, roots were rinsed thoroughly with distilled water, quickly frozen in liquid nitrogen, and stored at -80℃ for subsequent analysis.
2.3. Plant Transcriptome Analysis Methodology
High-quality total RNA (RIN value ≥ 7) was extracted from the roots of ‘Xikaixin’ melon seedlings (4 biological replicates per group, n=4) using the CTAB method. mRNA was converted into cDNA using reverse transcriptase, and the cDNA was fragmented and ligated with sequencing adapters. Library fragments were enriched to improve sequencing efficiency. The sequencing data were aligned to the reference genome (Cucumis melo L. var. reticulatus cv. DHL92 v3.6.1, downloaded from NCBI GenBank), and gene expression levels (FPKM) were calculated. Differentially expressed genes (DEGs) were identified using DESeq2 and edgeR with the threshold of |log2(fold change)| ≥ 1 and FDR < 0.05. Functional enrichment analysis was performed using GO and KEGG databases.
2.4. Measurement of Physiological Indicators
After 9 days of saline-alkali stress treatment, the photosynthetic rate of ‘Xikaixin’ melon seedlings was determined to verify the functional relevance of the photosynthesis-related gene MELO3C021567. Three seedlings per replicate (n=4) were selected for measurement, with three technical replicates per sample.
Photosynthetic rate (Pn): Measured using a Li-6400XT portable photosynthesis system (Li-Cor, USA). The measurement conditions were set as follows: leaf chamber CO₂ concentration 400 μmol·mol⁻¹, light intensity 1000 μmol·m⁻²·s⁻¹ (consistent with the cultivation light intensity), leaf temperature 25±1℃, and relative humidity 60±5%. The fully expanded true leaf of each seedling was clamped into the leaf chamber, and the data were recorded after stabilization for 3-5 min.
2.5. RT-qPCR Validation of Three Core Candidate Genes
To further confirm the reliability of transcriptomic data for MELO3C013403, MELO3C004381, and MELO3C021567, RT-qPCR was performed as follows:
RNA extraction and cDNA synthesis: Total RNA was extracted from roots using the CTAB method (Section 2.3), and cDNA was synthesized with the PrimeScript™ RT Reagent Kit with gDNA Eraser (TaKaRa, Japan) to eliminate genomic DNA contamination.
Primer design: Specific primers for the three core genes and internal reference gene (Actin, GenBank accession: XM_008454653.2) were designed using Primer3Plus software (Table S10). Primer specificity was verified by agarose gel electrophoresis and melting curve analysis.
RT-qPCR reaction: Conducted on a CFX96 Real-Time PCR System (Bio-Rad, USA) with SYBR® Premix Ex Taq™ Ⅱ (TaKaRa, Japan). The 20 μL reaction system included 10 μL SYBR Premix, 0.4 μL each of forward and reverse primers (10 μmol·L⁻¹), 2 μL cDNA template, and 7.2 μL ddH₂O. The reaction program was: 95℃ predenaturation for 30 s; 40 cycles of 95℃ denaturation for 5 s, 58℃ annealing for 30 s; and melting curve analysis from 65℃ to 95℃.
Data analysis: Relative expression levels were calculated using the 2⁻ΔΔCt method, with 4 biological replicates and 3 technical replicates per sample.
3. Results and Analysis
3.1. Transcriptome Sequencing Results Analysis
In this study, a distilled water control and three saline-alkali stress treatments with different ratios (NaCl:NaHCO₃ = 1:1, 1:2, 2:1) were set up, with 3 replicates per treatment. A total of 12 samples were subjected to eukaryotic reference-based transcriptome (RNA-seq) sequencing analysis. After analysis, a total of 78.98 Gb of high-quality Clean Data was obtained, with each sample having ≥ 6.02 Gb of Clean Data. Quality assessment of the sequencing data showed that the Q30 base percentage was ≥ 96.61%, and the Clean Reads of each sample were aligned to the reference genome with alignment efficiency ranging from 97.00% to 98.02%. The above data indicate that the sequencing data quality is good and can be used for subsequent in-depth data analysis. Subsequent analysis of differentially expressed gene data found that under the three simulated saline-alkali stress treatments of 1:1, 1:2, and 2:1, the number of upregulated DEGs in ‘Xikaixin’ was 194, 225, and 383, respectively, and the number of downregulated genes was 394, 461, and 724 (Figure 1 A-C). These results indicate that under different ratios of saline-alkali stress treatments, ‘Xikaixin’ responds to damage caused by saline-alkali stress by regulating its own gene expression.
3.2. Functional Annotation of Differentially Expressed Genes
Under the 1:1, 1:2, and 2:1 saline-alkali stress treatments, the number of annotated DEGs was 560, 654, and 1064, respectively. The DEGs were mainly enriched in 5 major categories of KEGG pathways: genetic information processing, environmental information processing, cellular processes, metabolism, and organismal systems, involving a total of 50 KEGG pathways.
DEGs under the 1:1 saline-alkali stress treatment were enriched in 20 KEGG pathways, mainly including: plant hormone signal transduction, benzoxazinoid biosynthesis, plant-pathogen interaction, phenylalanine metabolism, phenylpropanoid biosynthesis, etc. (Figure 2 A). KEGG enrichment pathways under the 1:2 saline-alkali stress treatment included the following metabolic pathways: plant hormone signal transduction, plant-pathogen interaction, linoleic acid metabolism, glutathione metabolism, carotenoid biosynthesis, etc. (Figure 2 B). The KEGG enrichment pathways under the 2:1 saline-alkali stress treatment were divided into plant-pathogen interaction, plant MAPK signaling pathway, plant hormone signal transduction, photosynthesis-antenna proteins, and phenylpropanoid biosynthesis (Figure 2 C).
Combining the above results, under the 1:1 and 1:2 saline-alkali stress treatments, the most enriched DEGs were in plant hormone signal transduction. Under the 2:1 saline-alkali stress treatment, DEGs were significantly enriched in the plant hormone signal transduction pathway, indicating that this pathway plays an important role in ‘Xikaixin’s response to saline-alkali stress. In addition, compared with the 1:1 and 1:2 saline-alkali stress treatments, the 2:1 saline-alkali stress treatment added phenol secondary metabolism and carbohydrate metabolism pathways, indicating that ‘Xikaixin’ melon copes with damage caused by saline-alkali stress by enriching genes in different metabolic pathways under different ratios of saline-alkali stress treatments.
3.3. Analysis of Differentially Expressed Genes in Melon Response to Saline-Alkali Stress
Among all treatment groups, the enrichment level was the highest. Among them, the auxin-induced protein genes MELO3C013403 and MELO3C004381 were significantly upregulated and can be used as candidate genes for saline-alkali tolerance (Tables 1-3). In the 2:1 treatment group, photosynthesis-antenna protein genes were significantly downregulated. The results indicate that melon adapts to the decrease in photosynthetic efficiency caused by saline-alkali stress by downregulating the expression of genes enriched in photosynthesis-antenna proteins, thereby reducing the synthesis of antenna proteins (Tables 4-6). Genes in pathways such as starch and sucrose metabolism and amino acid biosynthesis were differentially expressed in each treatment group (Tables 7-9). For the differentially expressed data of melon responding to saline-alkali stress, we selected core candidate genes for experimental verification. Priority was given to selecting MELO3C013403 (auxin-induced protein gene) and MELO3C004381 (auxin response factor gene) in the “plant hormone signal transduction” pathway, as well as MELO3C021567 (photosystem Ⅱ oxygen-evolving complex protein gene), a photosynthesis-related gene unique to the 2:1 group, as core candidate genes.
3.4. Analysis of Differentially Expressed Genes in Melon Response to Saline-Alkali Stress
Verification analysis of the gene expression levels of candidate genes showed that the transcriptional level data of several genes obtained from the experiment were consistent with the trends of the transcriptomic data (Figure 3 and Tables 1-9). MELO3C013403 and MELO3C004381 were significantly upregulated in the 2:1 group (more than twice that of the CK group) and slightly downregulated in the 1:1 and 1:2 groups; MELO3C021567 showed a gradient downregulation with the increase in NaCl ratio, with the most significant downregulation in the 2:1 group (only 32% of the CK group), which intuitively verified the reliability of the RNA-seq data.
3.5. Physiological Validation of Core Pathways
3.5.1. Photosynthetic Performance
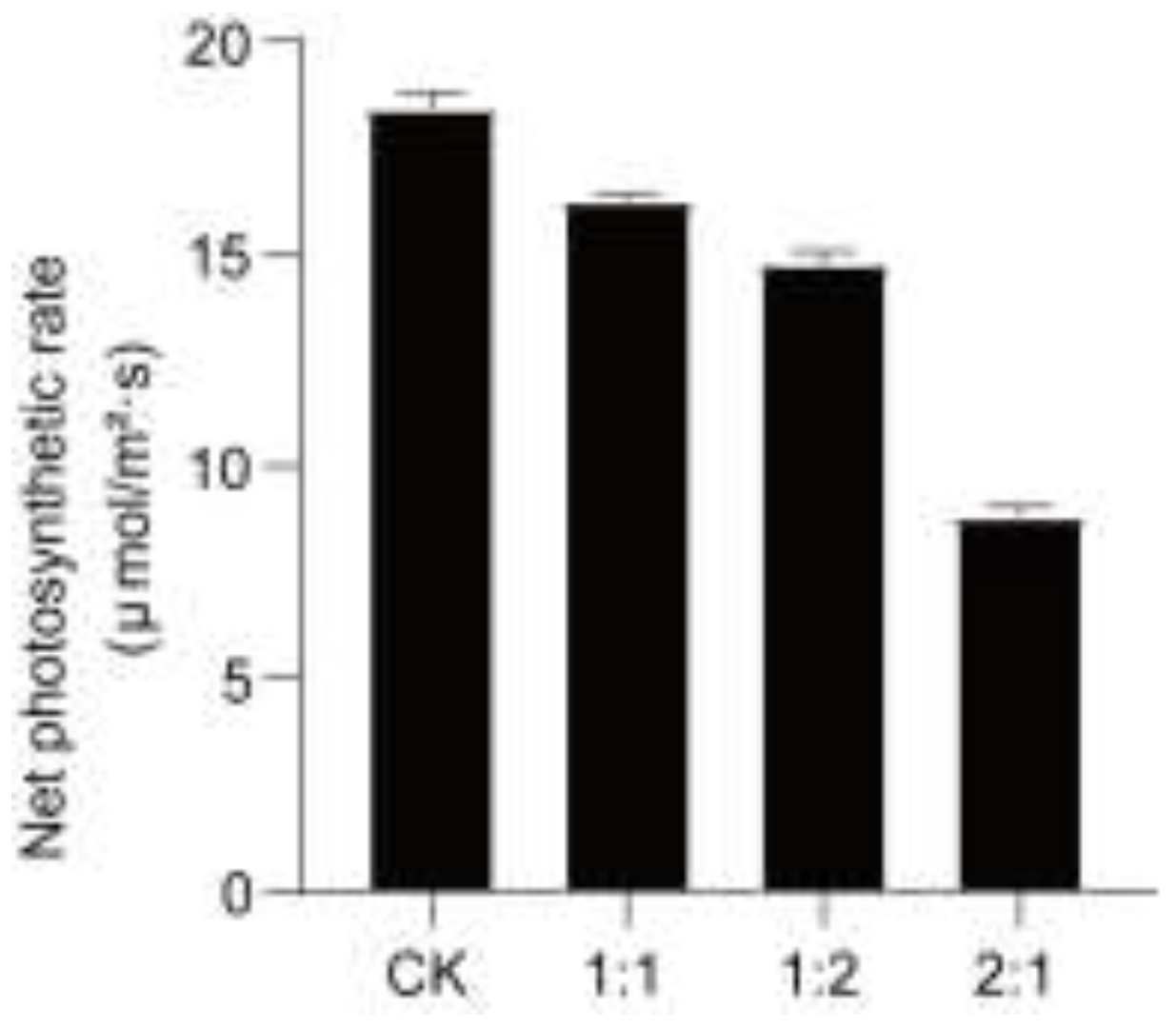
The photosynthetic rate (Pn) of ‘Xikaixin’ melon seedlings showed a gradual decreasing trend with increasing NaCl ratio (Figure 4). In T3 (2:1) treatment, the photosynthetic rate was 47.9% of CK , which was significantly lower than that in T1 and T2 (P < 0.05). This result is consistent with the downregulated expression of MELO3C021567, indicating that the reduction in photosynthetic efficiency under high-salt stress is closely related to the decreased expression of photosystem Ⅱ-related genes.
3.5.2. Osmoregulation and Oxidative Stress (Supported by Unpublished Data)
Unpublished physiological data from our research group showed that saline-alkali stress significantly affected the osmoregulatory and oxidative stress status of ‘Xikaixin’ melon seedlings. Soluble sugar and proline contents (osmotic adjustment substances) increased in T1 (1:1) and T2 (1:2) treatments (1.8-fold and 2.2-fold, 1.5-fold and 1.6-fold of CK, respectively) but decreased in T3 (2:1) treatment (0.8-fold and 0.9-fold of CK). Malondialdehyde (MDA) content (a marker of oxidative damage) increased with stress intensity, reaching 2.8-fold of CK in T3, while superoxide dismutase (SOD) activity peaked in T2 (1.7-fold of CK) and slightly decreased in T3 (1.5-fold of CK). These data support the functional relevance of the hormone and photosynthesis pathways regulated by the three core genes, reflecting the coordinated response of ‘Xikaixin’ melon to saline-alkali stress.
4. Discussion
In this study, ‘Xikaixin’, a main local saline-alkali-tolerant melon cultivar in Xinjiang, was used as the material. Combined with transcriptome sequencing technology, the molecular mechanism of melon seedling roots responding to saline-alkali stress was systematically analyzed by setting three different ratios of mixed saline-alkali stress treatments (NaCl:NaHCO₃ = 1:1, 1:2, 2:1). The results showed that the number of differentially expressed genes (DEGs) in melon seedlings under different ratios of saline-alkali stress showed significant gradient differences, and the response of core metabolic pathways had obvious saline-alkali ratio specificity, providing important molecular evidence for in-depth understanding of the saline-alkali tolerance adaptation strategy of melon.
4.1. Response Characteristics and Cultivar Specificity of Differentially Expressed Genes Under Saline-Alkali Stress
Plant adaptation to saline-alkali stress relies on complex gene expression regulatory networks. The number and expression patterns of differential genes directly reflect the plant’s perception and response to stress intensity and type [25]. This study found that 588, 686, and 1107 differentially expressed genes were identified in ‘Xikaixin’ melon under the 1:1, 1:2, and 2:1 saline-alkali stress treatments, respectively. The number of downregulated genes was significantly more than that of upregulated genes, and the total number of differential genes showed an increasing trend with the increase in NaCl ratio in the saline-alkali stress. This result is different from the rule of “prolonged stress leads to an increase in downregulated genes” in Zhang et al.’s (2019) study on melon under saline-alkali stress[26]. It is speculated that the differences mainly come from two aspects: first, the mixed saline-alkali ratio gradient used in this study (especially the high proportion of NaCl in the 2:1 treatment) may cause more direct ionic toxicity to the roots, triggering the downregulation of more genes related to growth inhibition; second, as a locally selected cultivar in Xinjiang, ‘Xikaixin’ has differences in its saline-alkali tolerance genetic background from the melon cultivars in previous studies, leading to cultivar-specific gene response patterns.
Compared with the transcriptomic study results of saline-alkali stress in crops such as Jinsuihuanggu (a foxtail millet cultivar) and Potentilla anserina, the differential genes of melon in this study were also enriched in core pathways such as plant hormone signal transduction and MAPK signaling pathway, indicating that these pathways are conserved mechanisms for plants to respond to saline-alkali stress [27,28]. However, it is worth noting that melon added specific enrichment pathways such as photosynthesis-antenna proteins and phenol secondary metabolism under the 2:1 high-salt ratio stress, while foxtail millet and Potentilla anserina focused more on pathways such as fatty acid metabolism and peroxisome. This reflects the species-specific differences in saline-alkali tolerance strategies among different crops, which may be related to the physiological characteristics of melon as a Cucurbitaceae crop and the sensitivity of its roots to ionic stress.
4.2. Regulatory Mechanism of Core Metabolic Pathways and Adaptive Strategies to Saline-Alkali Stress
Plant hormone signal transduction is a key regulatory network for plants to cope with abiotic stress, balancing growth and development with stress tolerance through the synergistic and antagonistic effects of hormones [29]. In this study, the plant hormone signal transduction pathway showed the highest enrichment level among the three saline-alkali stress treatments, and two significantly upregulated core candidate genes were identified—the auxin-induced protein gene MELO3C013403 and the auxin response factor gene MELO3C004381. This finding is different from the general rule that auxin-related genes are downregulated under saline-alkali stress in most plants [28].
Notably, saline-alkali stress is a composite stress involving pH, alkalinity, and osmotic load, which collectively affect the balance between auxin and ABA in ‘Xikaixin’ melon. The three treatment groups (NaCl:NaHCO₃ = 1:1, 1:2, 2:1) had different pH values (measured as 7.8 ± 0.2, 8.5 ± 0.3, 7.5 ± 0.2, respectively) and osmotic potentials (-0.21 ± 0.02, -0.23 ± 0.03, -0.25 ± 0.02 MPa, respectively), reflecting gradient differences in alkalinity and osmotic stress. In the T2 (1:2) treatment with higher alkalinity (pH 8.5), ABA-related genes were upregulated while auxin-related genes were slightly downregulated, which may be due to high pH inhibiting auxin synthesis and promoting ABA accumulation to enhance osmotic adjustment [30]. In contrast, the T3 (2:1) treatment with stronger osmotic load (lower osmotic potential) but lower pH triggered the specific upregulation of MELO3C013403 and MELO3C004381, suggesting that under high-salt-dominated osmotic stress, ‘Xikaixin’ melon maintains root growth by activating auxin signals to counteract the inhibitory effect of osmotic stress, while ABA signals may synergistically regulate ion homeostasis. This composite effect of pH, alkalinity, and osmotic load on hormone balance is consistent with the characteristics of natural saline-alkali stress and explains the cultivar-specific hormone response pattern of ‘Xikaixin’.
Especially in the 2:1 treatment, the expression levels of these two genes reached more than twice that of the control, suggesting that ‘Xikaixin’ melon may alleviate the inhibition of root growth caused by high-salt stress by specifically activating components of the auxin signal pathway, which may be an important molecular characteristic of the saline-alkali tolerance of this cultivar.
In addition, the expression pattern of upregulated abscisic acid (ABA)-related genes and downregulated auxin, jasmonic acid, and gibberellin-related genes in the 1:2 treatment conforms to the classic regulatory logic of ABA as a “stress hormone”—enhancing the plant’s osmotic adjustment ability and stomatal closure efficiency through ABA accumulation, while inhibiting growth hormone signals to save energy for stress response [30]. The downregulation of 4 genes in the MAPK signaling pathway may be involved in the fine regulation of hormone signals through cascade reactions. Previous studies have shown that the MAPK pathway can affect auxin signal transduction by phosphorylating auxin response factors (ARFs) [31]. It is speculated that it may mediate the antagonistic balance between ABA and auxin in melon’s response to saline-alkali stress.
Photosynthesis is the most sensitive physiological process of plants to abiotic stress, and its efficiency directly affects the plant’s energy supply and growth and development [32]. In this study, genes in the photosynthesis-antenna protein pathway were significantly downregulated under the 2:1 saline-alkali stress treatment, and the expression level of the photosystem Ⅱ oxygen-evolving complex protein gene MELO3C021567 was only 32% of that of the control. The suppression of antenna protein genes is directly linked to reduced photosynthetic outputs: antenna proteins (e.g., LHCⅡ) are responsible for capturing and transferring light energy to the reaction center of photosystems. The downregulated expression of MELO3C021567 reduces the synthesis of antenna proteins, thereby decreasing the absorption of excess light energy. This avoids the over-reduction of the photosynthetic electron transport chain and the accumulation of reactive oxygen species (ROS), which in turn protects the structure and function of photosystems Ⅰ and Ⅱ. Consistent with this, unpublished physiological data show that the photosynthetic rate of ‘Xikaixin’ melon seedlings in the 2:1 treatment decreased by 58% compared with the control, which is synchronized with the downregulation of antenna protein genes. This adaptive strategy balances the trade-off between light energy utilization and photosynthetic apparatus protection under high-salt stress.
This molecular change is highly consistent with the physiological data of “saline-alkali stress leading to a significant decrease in chlorophyll content in melon leaves” in the unpublished data of this paper. This chain reaction of “downregulated gene expression - chlorophyll degradation - decreased photosynthetic efficiency” may be an adaptive strategy of melon under high-salt stress: reducing the absorption of excess light energy by the photosystem through decreasing antenna protein synthesis, thereby alleviating the damage to the photosynthetic apparatus caused by the accumulation of reactive oxygen species [32].
The regulation of carbon metabolism and energy supply also shows saline-alkali ratio specificity: genes involved in starch and sucrose metabolism were all downregulated in the 1:1 treatment; inositol phosphate metabolism and starch-sucrose metabolism responded synergistically in the 1:2 treatment; alanine, aspartate, and glutamate metabolism were co-inhibited with starch-sucrose metabolism in the 2:1 treatment. The starch and sucrose metabolism pathway was dominated by downregulation in all treatments, indicating that the inhibition of melon carbohydrate synthesis by saline-alkali stress is a common feature, which is directly related to the phenotypic decrease in seedling stem diameter and dry weight [13]. The differential response of amino acid metabolism reflects the energy allocation strategy under different stress intensities: some amino acid synthesis genes were upregulated under mild 1:1 stress, which may maintain cell osmotic pressure by accumulating compatible solutes such as proline; amino acid metabolism genes were comprehensively downregulated under severe 2:1 stress, which may be due to the severe inhibition of energy metabolism, and the plant can no longer support the synthesis of osmotic adjustment substances [33].
4.3. Limitations of the Study
This study provides insights into the molecular mechanism of ‘Xikaixin’ melon responding to saline-alkali stress, but there are several limitations that need to be addressed in future research. First, the study only focused on a single time point (9 days after stress initiation), which limits the understanding of the dynamic temporal response of melon to saline-alkali stress. The expression patterns of core genes (e.g., MELO3C013403, MELO3C004381) and the activation of key pathways may change at early (e.g., 24–72 h) or late (e.g., 14 days) stress stages, and continuous time-series sampling is required to reveal the complete stress response process. Second, the control group had inherent deficiencies: the original control used only distilled water, which lacked the standard nutrient supply required for melon seedling growth, potentially leading to nutrient imbalance and affecting the accurate comparison of stress responses. Although this study has supplemented the explanation of nutrient balance in the revised Materials and Methods, future experiments should use a standard nutrient solution (e.g., half-strength Hoagland solution) as the control to eliminate nutrient confounding factors. Third, the study did not analyze the changes in endogenous hormone contents (e.g., auxin, ABA) and their interactions at the physiological level, which limits the in-depth verification of the hormone balance regulation mechanism inferred from transcriptomic data. Additionally, the functional validation of candidate genes (e.g., overexpression or knockout) was not performed, and the direct role of these genes in saline-alkali tolerance needs to be further confirmed through genetic transformation experiments.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1. CK vs X1:1 signal transduction-related genes. Table S2. CK vs X1:2 signal transduction-related genes. Table S3. CK vs X2:1 signal transduction-related genes. Table S4. CK vs X1:1 photosynthesis-related genes. Table S5. CK vs X1:2 photosynthesis-related genes. Table S6. CK vs X2:1 photosynthesis-related genes. Table S7. CK vs X1:1 Osmoregulation-related genes. Table S8. CK vs X1:2 Osmoregulation-related genes. Table S9. CK vs X2:1 Osmoregulation-related genes.
Author Contributions
W. T., Z, Y., S. Y. and N. M. performed the experiments; W. L. and W. H. conceived and designed the experiments, wrote the paper, all authors read and approved the final manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (32300433 to H.W.); the State Key Laboratory of North China Crop Improvement and Regulation, S&T Program of Hebei (23567601H); the Xinjiang Uygur Autonomous Region Science and Technology Program (2022TSYCCX0083); Xinjiang Uygur Autonomous Region Postdoctoral Research Innovation Platform; Talent Program for Intellectual Support to Xinjiang - “Small Group” Aid Xinjiang Team and Flexible Aid Xinjiang Expert Talent Project “Innovative Team for High-Value Processing and Utilization of Natural Color Cotton and Fibers”.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data required to support the claims of this paper are included in the main. All reagents generated in this study are available on request from the lead contact. RNA-seq data of Cucumis melo are stored in the biological project library of NCBI with accession number PRJNA1334105.
Conflicts of Interest
The authors declare no competing interests.
References
- Munns, R., & Tester, M. Mechanisms of salinity tolerance. Annual Review of Plant Biology, 2008, 59, 651-681.
- Lü, N., Shi, L., Dai, Y. Y., et al. Review and enlightenment of saline-alkali land management and utilization research in Xinjiang. Journal of Irrigation and Drainage, 2024, 1-13.
- Han, J. L. Comprehensive utilization approaches, effects and enlightenment of saline-alkali land in Changyi, Shandong. China Agricultural Comprehensive Development, 2024, (11), 15-17.
- Tian, C. Y., Mai, W. X., & Zhao, Z. Y. Key technologies for ecological management of saline-alkali land in arid areas of Xinjiang. Acta Ecologica Sinica, 2016, 36(22), 7064-7068.
- Himani, P., Jayanti, T., Surina, B., et al. Proteome dynamics and transcriptome profiling in sorghum (Sorghum bicolor (L.) Moench) under salt stress. 2020, 3 Biotech, 10(9), 412.
- Xu, T., Liu, Y. T., Wang, H. J., et al. Effects of saline-alkali stress on peanut seed germination characteristics and comprehensive evaluation of salt damage. Chinese Journal of Oil Crop Sciences, 2022, 44(5), 11.
- He, L., Lu, Z. H., Guan, B., Zhao, Y. Y., Wang, R. T., & Liu, Y. Effects of saline-alkali stress on seed germination and seedling growth of sweet sorghum. Journal of Northeast Forestry University, 2012, 40(3), 5.
- Chen, Y. Q., Su, K. Q., Chen, T. X., & Li, C. J. Effects of mixed saline-alkali stress on seed germination and seedling physiological characteristics of Achnatherum inebrians. Acta Prataculturae Sinica, 2021, 30(3).
- Jin, M. Y., Li, X. H., Li, F. Z., & Huang, Z. B. Effects of combined saline-alkali stress on rice seed germination. Chinese Journal of Eco-Agriculture, 2020, 28(4), 9.
- Liang, R. F., Fu, Z. X., Zhou, P., et al. Effects of paclobutrazol on stress resistance physiology of melon seedlings based on multivariate comprehensive analysis. Acta Agriculturae Boreali-Occidentalis Sinica, 2025, 34(02), 280-289.
- Liu, Y. M. Mitigative effect of melatonin on melon autotoxicity and saline-alkali compound stress. Master’s thesis, Fujian Agriculture and Forestry University. China, 2022; pp. 40.
- Qiao, Z. Y., Yu, M., Tang, Y. J., et al. Comprehensive evaluation and index selection of grain sorghum seedling tolerance to soda saline-alkali. Scientia Agricultura Sinica, 2025, 58(01), 30-49.
- Zhang, X. L., Li, S. Q., Guo, S., et al. N-acetyl-L-cysteine alleviates the inhibition of soybean seedling growth under saline-alkali stress. Soybean Science, 2025, 44(01), 96-105.
- Wei, S., Wang, L., Zhang, Y., & Huang, D. Identification of early response genes to salt stress in roots of melon (cucumis melo l.) seedlings. Molecular Biology Reports, 2013, 40(4), 2915-2926. [CrossRef]
- Wang, L. M., Zhang, L. D., Chen, J. B., Huang, D. F., & Zhang, Y. D. Physiological analysis and transcriptome comparison of two muskmelon (cucumis melo l.) cultivars in response to salt stress. Genetics & Molecular Research, 2016.15(3). [CrossRef]
- Chen, J. B., Zhang, F. R., Huang, D. F., Zhang, L. D., & Zhang, Y. D. Yansu xiepohua xia liangge tiangua pinzhong de zhuansijian fenxi [Transcriptome analysis of two melon varieties under salt stress] (in Chinese). Zhiwu Shengli Xuebao [Plant Physiology Journal], 2014, 50(2), 150-158.
- Wang Y, Shen Y, Li B, et al. Deciphering the regulatory network of the NAC transcription factor FvRIF, a key regulator of strawberry (Fragaria vesca) fruit ripening. Plant Cell. 2023, 35 (11):4020-4045. [CrossRef]
- Wang, J., Tian, S., Yu, Y., Ren, Y., Guo, S., & Zhang, J., et al. Natural variation in the nac transcription factor nonripening contributes to melon fruit ripening. Acta Botanica Sinica (English Edition), 2022, (7). [CrossRef]
- Zhang, H., Zhang, Y., Dai, Z., Yuan, Y., & Wei, S. Transcriptome analysis, cmmyb1 gene clone from cucumis melo l. and its functional roles under salt stress. 2020. [CrossRef]
- Diao, Q., Tian, S., Cao, Y., Yao, D. Fan, H., & Jiang, X., et al. Physiological, transcriptomic, and metabolomic analyses of the chilling stress response in two melon (cucumis melo l.) genotypes. BMC Plant Biology, 2024, 24(1), 1-17. [CrossRef]
- Li, W. Y., Li, X. Y., & Yang, C. C. Transcriptome analysis of different resistant poplar cultivars under saline-alkali stress. Journal of Central South University of Forestry & Technology, 2025, 45(02), 122-130.
- Wang, H. M. Physiological and transcriptomic analysis of foxtail millet seedlings responding to saline-alkali stress and study on the SiAAAPs family. Master’s thesis, Hebei Normal University of Science & Technology. China, 2024; pp. 54-61.
- Xue, Y., Zhao, S. J., He, Y. M., et al. Research progress on salt tolerance of cucurbit crops. China Cucurbits and Vegetables, 2023, 36(12), 1-8.
- Sun, J. L., & Hou, Z. A. Effects of phosphorus on ion balance and related regulatory genes of cotton under saline-alkali stress. Journal of Plant Nutrition and Fertilizer, 2022, 28(04), 643-653.
- Xiao, W. L., Wang, H. R., Wang, M. L., et al. Research progress on plant response mechanisms under saline-alkali stress. Chinese Agricultural Science Bulletin, 2024, 40(33), 78-85.
- Yang, Y. B., Fu, G. X., Hao, W. F. Physiological response and transcriptome analysis of Ammopiptanthus nanus to drought stress. Acta Ecologica Sinica, 2025, 45(02), 854-865.
- Liu, Y. Screening of mixed saline-alkali tolerant foxtail millet cultivars and their physiological and biochemical adaptation mechanisms. Master’s thesis, Heilongjiang Bayi Agricultural University, China, 2024; pp. 48-51.
- Lü, B. W., Li, J. Q., Yan, G., et al. Morphological and transcriptome analysis of Potentilla anserina responding to saline-alkali stress. Seed, 2024, 43(01), 14-28.
- Chen, F., Chen, H., Chen, B. Q., et al. Changes in endogenous hormone content and gene expression analysis of apomictic kernels in walnut [J/OL]. Journal of Zhejiang Agricultural Sciences, 2024, 1-14.
- Guan, X. L., & Shen, J. Research progress on physiological and molecular mechanisms of plant response to saline-alkali stress. Jiangsu Agricultural Sciences, 2024, 52(21), 10-16.
- Liang, C., Ze, S. Z., Gao, Y. Z., et al. Study on endogenous hormones and related gene expression before and after flower bud differentiation of Mikania micrantha. Acta Botanica Boreali-Occidentalia Sinica, 2023, 43(02), 229-241.
- Zhang, B. B. Morphological, physiological and biochemical responses and transcriptome analysis of melon to saline-alkali stress. Master’s thesis, Fujian Agriculture and Forestry University. 2019; pp. 21-22.
- Zhao, L. M. Study on the regulatory mechanism of starch-sucrose metabolism on tuber expansion of Chinese yam (Dioscorea opposita Thunb.). Master’s thesis, Inner Mongolia Agricultural University. China, 2023; pp. 54-56.
Figure 1.
Volcano plot of the differentially expressed genes. Note: (a) CK vs X1:1; (b) CK vs X1:2; (c) CK vs X2:1; The same below.
Figure 1.
Volcano plot of the differentially expressed genes. Note: (a) CK vs X1:1; (b) CK vs X1:2; (c) CK vs X2:1; The same below.
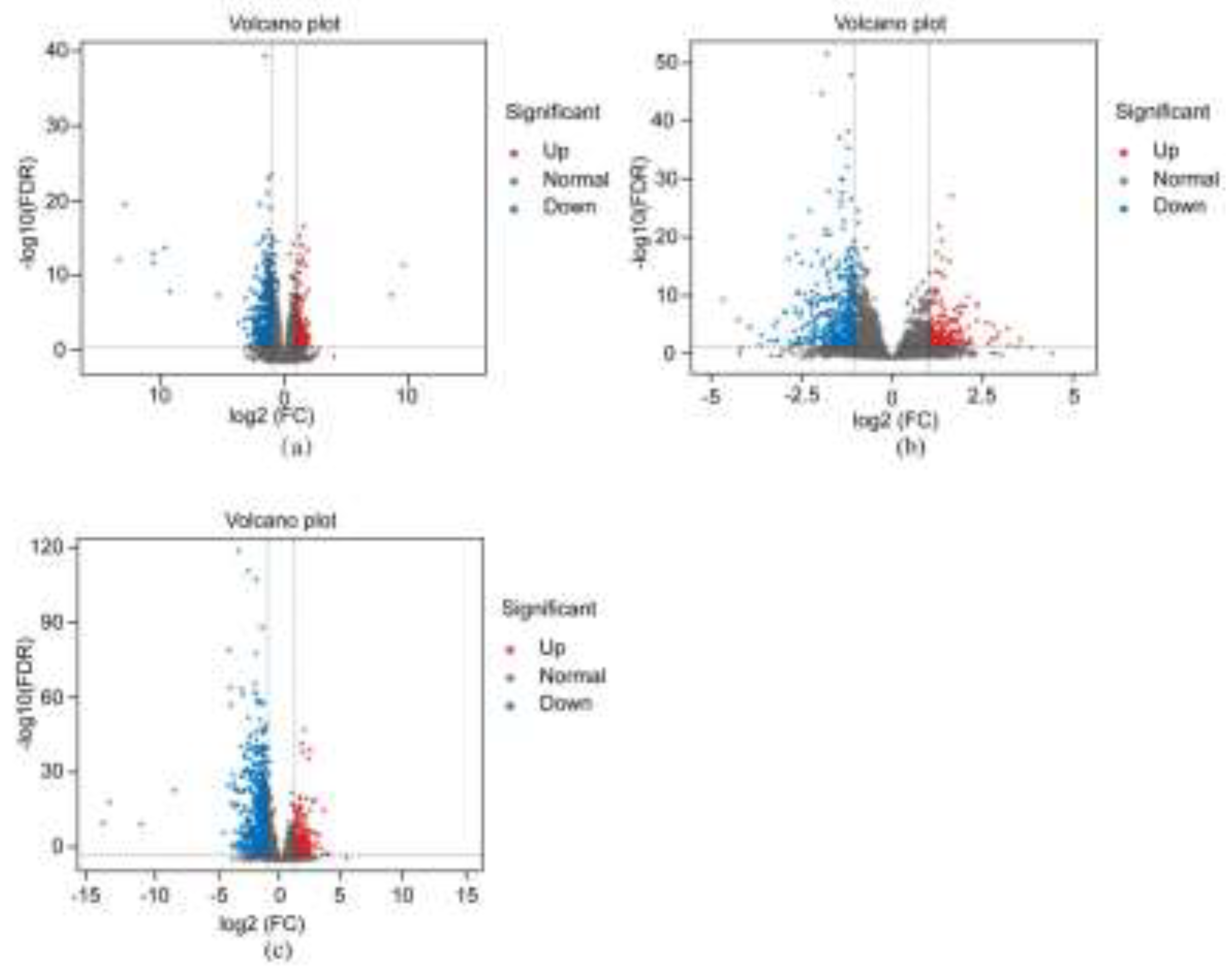
Figure 2.
KEGG enrichment map of differentially expressed genes. (a) KEGG enrichment analysis of DEGs under 1:1 saline-alkali stress treatment; (b) KEGG enrichment analysis of DEGs under 1:2 saline-alkali stress treatment; (c) KEGG enrichment analysis of DEGs under 2:1 saline-alkali stress treatment.
Figure 2.
KEGG enrichment map of differentially expressed genes. (a) KEGG enrichment analysis of DEGs under 1:1 saline-alkali stress treatment; (b) KEGG enrichment analysis of DEGs under 1:2 saline-alkali stress treatment; (c) KEGG enrichment analysis of DEGs under 2:1 saline-alkali stress treatment.
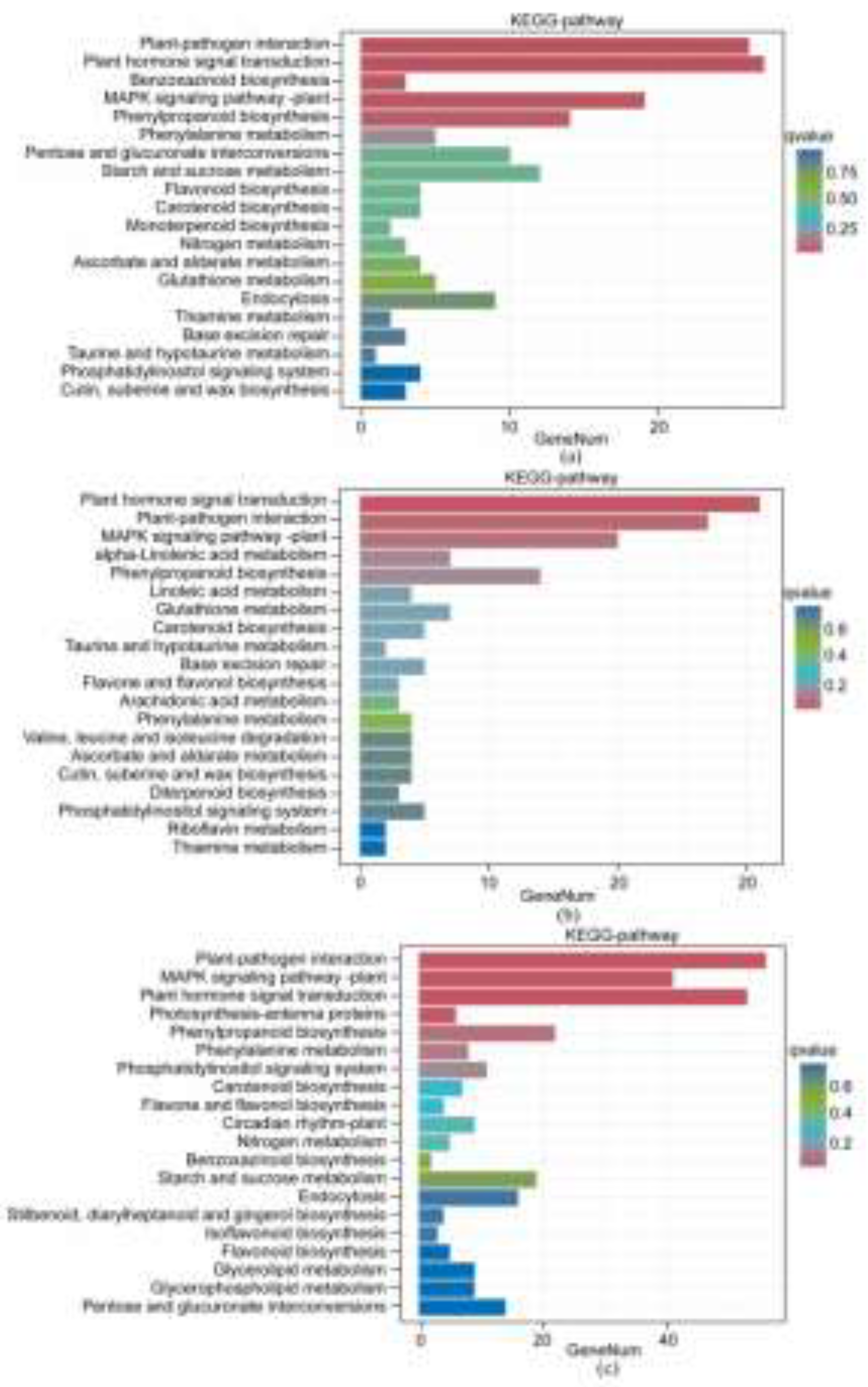
Figure 3.
Analysis of gene expression levels of core candidate genes. (a) Detection of the expression level of MELO3C013403 under different saline-alkali treatments; (b) Detection of the expression level of MELO3C004381 under different saline-alkali treatments; (c) Detection of the expression level of MELO3C021567 under different saline-alkali treatments.
Figure 3.
Analysis of gene expression levels of core candidate genes. (a) Detection of the expression level of MELO3C013403 under different saline-alkali treatments; (b) Detection of the expression level of MELO3C004381 under different saline-alkali treatments; (c) Detection of the expression level of MELO3C021567 under different saline-alkali treatments.
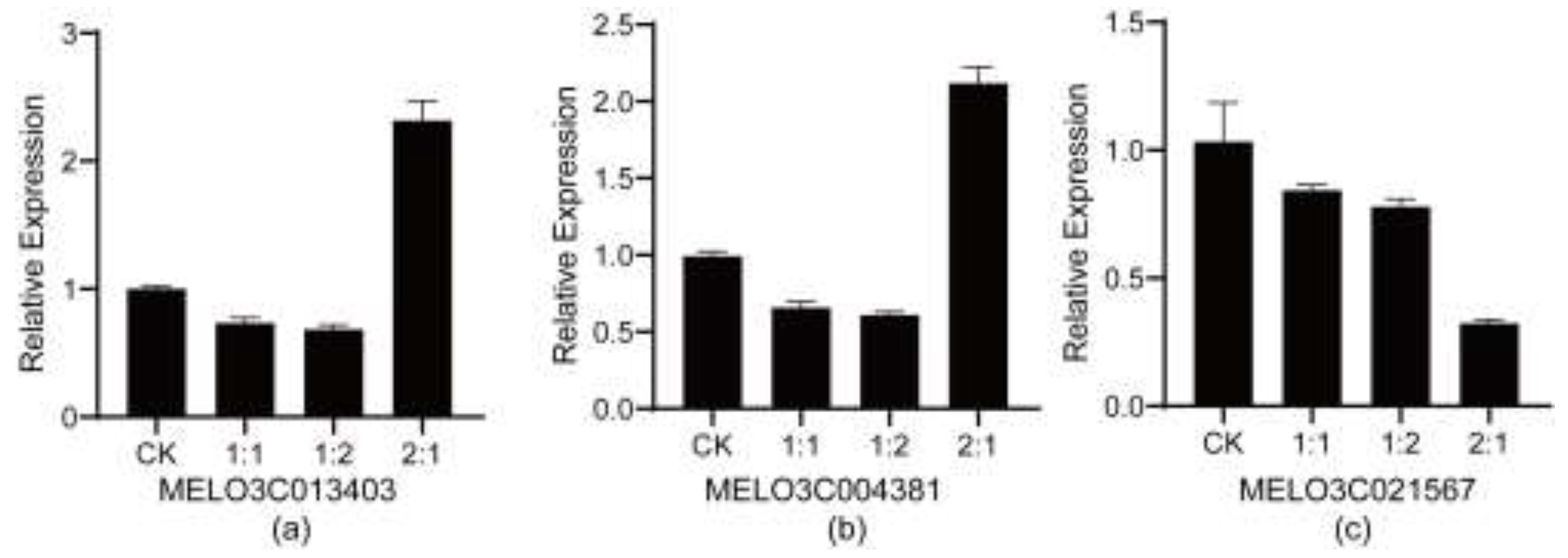
Figure 4.
Analysis of the photosynthetic rate of the gene MELO3C021567.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



