
Submitted:
19 January 2026
Posted:
20 January 2026
You are already at the latest version
Abstract
Celiac disease (CD) is an autoimmune disorder triggered by immunogenic gluten peptides that resist gastrointestinal digestion. The only current treatment is a strict gluten-free diet, which is challenging to maintain. Lactic acid bacteria (LAB) with specific proteolytic systems offer a promising strategy for gluten detoxification. This study aims to isolate and characterize gluten-degrading LAB from traditional Spanish and Algerian dairy products. A total of 27 artisanal dairy samples were collected. LAB were isolated on MRS and Elliker agar. Gluten-degrading activity was screened using a well diffusion assay with cell-free supernatants and a spot assay with live cultures. Active isolates were identified by 16S rRNA gene sequencing. Out of 123 isolates, 40 (32.5%) were positive in the well assay, while 67 (54.5%) were positive in the spot assay, indicating the latter's higher sensitivity for detecting cell-associated proteases. Halo diameters ranged from 6 to 16 mm. Algerian isolates exhibited significantly stronger activity (mean halo: 12.6 ± 2.1 mm) compared to Spanish isolates (10.2 ± 2.0 mm; p < 0.001). Molecular identification of the 32 most active isolates revealed dominant species as Lactiplantibacillus plantarum, L. pentosus, Levilactobacillus brevis, and Enterococcus faecium. This study confirms that artisanal dairy fermentations are rich sources of LAB with robust gluten-degrading potential. The superior activity of Lactiplantibacillus spp. aligns with their complex peptidase systems. The geographical variation highlights the influence of local fermentation practices. Selected strains represent excellent candidates for developing adjunct cultures to produce gluten-reduced foods and novel probiotic therapeutics.
Keywords:
lactic acid bacteria
; gluten hydrolysis
; celiac disease
; fermented dairy
; probiotics
; Lactiplantibacillus
; proteolytic activity
; microbial diversity
1. Introduction
Celiac disease (CD) is a chronic immune-mediated enteropathy affecting approximately 0.7-1% of the global population [1]. It is triggered in genetically susceptible individuals by the ingestion of gluten proteins from wheat, barley, and rye [2]. Gluten's high proline and glutamine content renders it resistant to complete proteolysis by human digestive enzymes, leading to the persistence of immunogenic peptides in the gastrointestinal tract [3]. These peptides, such as the well-characterized 33-mer from α-gliadin, are deamidated by tissue transglutaminase, enhancing their affinity for human leukocyte antigen (HLA) molecules and triggering a T-helper 1-driven inflammatory response that results in villous atrophy, malabsorption, and systemic symptoms [4,5]. The cornerstone of CD management remains a strict, lifelong gluten-free diet (GFD). However, the GFD presents significant challenges, including limited food choices, higher costs, potential nutritional deficiencies, and the constant risk of cross-contamination [6,7]. Consequently, there is a pressing need for complementary or alternative therapeutic strategies [8]. Among these, the enzymatic degradation of gluten peptides before they reach the small intestine has emerged as a highly promising approach [9,10].
Lactic acid bacteria (LAB), widely used in food fermentations and recognized for their probiotic properties, possess sophisticated proteolytic systems [11]. These systems include cell-envelope proteinases that initiate the breakdown of proteins into oligopeptides, which are subsequently internalized and hydrolyzed by a broad array of intracellular peptidases, including proline-specific peptidases essential for cleaving gluten [12]. Several LAB strains, particularly from the genera Lactobacillus (recently reclassified into Lactiplantibacillus and Levilactobacillus), have demonstrated the ability to hydrolyze immunogenic gluten epitopes [13,14,15].
While sourdough ecosystems have been extensively explored for gluten-degrading LAB [16], traditional dairy fermentations represent an underexplored reservoir of microbial diversity with unique metabolic traits [17]. Artisanal cheeses, kefir, and other fermented milk products from regions like Spain and Algeria harbor complex, autochthonous LAB communities shaped by specific environmental conditions and traditional production methods [18]. These microbes have evolved to thrive in protein-rich matrices, suggesting a potentially high proteolytic capacity [19].
This study aimed to: (i) isolate LAB from a diverse collection of traditional Spanish and Algerian dairy products; (ii) screen their gluten-degrading potential using complementary agar-based assays; (iii) molecularly identify the most active isolates; and (iv) compare the functional and taxonomic profiles of isolates from the two geographical origins.
2. Materials and Methods
2.1. Sample Collection
A total of 27 artisanal dairy products were aseptically collected between May and August 2023 from small-scale producers in Spain (n=14) and Algeria (n=13). The Spanish samples included various cheeses from cow, goat, and sheep milk (Table 1). The Algerian samples encompassed traditional products such as Jben, Bouhaza cheese, Kefir, Klila, and Rayeb. Samples were transported to the laboratory under refrigerated conditions (4°C) and processed within 24 hours.
2.2. Isolation and Preservation of Lactic Acid Bacteria
From each sample, 5 g were homogenized with 45 mL of sterile saline solution (0.90% NaCl). Serial decimal dilutions were prepared and spread-plated on de Man, Rogosa, and Sharpe (MRS) agar and Elliker agar. Plates were incubated at 30°C for 24-48 hours. Colonies with distinct morphologies were selected, purified and stored at -20°C in MRS broth with 20% (v/v) glycerol.
2.3. Screening for Gluten-Degrading Activity
2.3.1. Preparation of Gluten Agar
The basal medium for gluten degradation assays contained (per liter): 10 g peptone, 5 g NaCl, and 10 g bacteriological agar. After autoclaving, the medium was cooled to 50°C and supplemented with 0.8 g/L of vital wheat gluten (Sigma-Aldrich, G5004) [20].
2.3.2. Well Diffusion Assay
LAB cultures were centrifuged to obtain cell-free supernatants (CFS). Sterile stainless-steel cylinders were placed on gluten agar, covered with a tempered gluten agar overlay, and removed after solidification. Wells were filled with 75 µL of CFS and plates were incubated at 30°C for 24 hours.
2.3.3. Spot Assay
Five microliters of active overnight MRS cultures were spotted onto gluten agar plates, air-dried, and incubated at 30°C for 24 h.
2.3.4. Visualization and Measurement
After incubation, the plates were flooded with Coomassie Brilliant Blue G-250 staining solution (1 g/L in a mixture of methanol: acetic acid: water, 4:1:5) for 30 min. The plates were then destained overnight with a solution of methanol: acetic acid: water (5:1:4). Clear, transparent zones (halos) around the wells or spots indicated proteolysis of gluten. The diameters of the halos were measured using a caliper.
2.4. Molecular Identification of Active Isolates
The thirty-two most promising isolates, selected based on halo diameter of ≥10 mm in a well-diffusion assay, were subjected to molecular identification. Bacterial genomic DNA was extracted using the GenElute Bacterial Genomic DNA Kit (Sigma-Aldrich, USA) according to the manufacturer's instructions. The 16S rRNA gene was amplified via PCR with the universal bacterial primers 27F (5'-AGAGTTTGATCCTGGCTCAG-3') and 1492R (5'-GGTTACCTTGTTACGACTT-3'). Successful amplification was verified by electrophoresis on a 1% agarose gel. Amplicons were purified and sent for Sanger sequencing (Stab Vida, Portugal). The resulting sequences were queried against the NCBI GenBank database using the BLASTN algorithm, and species-level identification was assigned to isolates with ≥98.5% sequence similarity to a reference strain.
2.5. Statistical Analysis
Data are presented as mean ± standard deviation (SD). Differences in halo diameters between geographical origins were analyzed using an unpaired two-sample t-test. Differences across bacterial species were analyzed using a one-way ANOVA followed by Tukey's post-hoc test. A p-value of < 0.05 was considered significant. Analyses were performed using R software (v4.3.0).
3. Results
3.1. Isolation and Screening for Gluten-Degrading Activity
A total of 123 presumptive LAB isolates were obtained, 84 from Spain and 39 from Algeria (Table 2 and Table 3, respectively). Screening revealed that 40 isolates (32.5%) were positive in the well diffusion assay which detects extracellular proteases. In contrast, the spot assay, which detects both extracellular and cell-associated activity, identified a higher number of positive isolates 67 isolates (54.5%) were positive in the spot assay (Table 4), indicating a significant contribution of cell-bound enzymes.
This comprehensive analysis of 123 lactic acid bacteria (LAB) isolates from traditional Spanish (n=84) and Algerian (n=39) dairy products revealed significant gluten-degrading capabilities, with distinct geographical patterns emerging. The spot assay proved more sensitive than the well diffusion assay, detecting 54.5% positive isolates overall compared to 32.5% in the well assay, indicating substantial contribution of cell-associated proteases alongside extracellular enzymes. As shown in Figure 1, Algerian samples demonstrated superior proteolytic activity, particularly in extracellular enzyme production (38.5% positivity in well assays versus 29.8% for Spanish samples), while both regions showed comparable effectiveness in spot assays (56.4% Algeria vs 53.6% Spain). Notably, several Algerian products including Kefir (KIF), Bouhaza Picante (BP), and Klila (KL) showed exceptional performance with 100% positivity in well diffusion assays, while strong activity (≥13 mm halo diameter) was observed in isolates from both Spanish (QF2, OBII series, EM3) and Algerian (KIF5, OB1, BP2) samples. These findings highlight traditional dairy fermentations, particularly Algerian products, as valuable reservoirs of LAB with robust gluten-degrading potential, offering promising candidates for developing adjunct cultures and novel probiotic therapeutics for celiac disease management.
Halo diameters ranged from 6 to 16 mm. Isolates from Algerian products produced significantly larger halos than those from Spanish products (p < 0.001). The most potent activity (+++) was associated with isolates from Algerian Kefir (KIF1, KIF2, KIF5) and Klila (KL3, KL4).
3.2. Molecular Identification of Gluten-Degrading Isolates
Molecular identification of the 32 most active isolates revealed a diverse taxonomy, with Lactiplantibacillus being the most prominent among highly active strains (Table 5). 40% were identified as Lactiplantibacillus, 25% as Enterococcus, 15% as Levilactobacillus, 10% as Leuconostoc, and the remaining 10% as Lactococcus (Figure 2). The phylogenetic relationships of these isolates are depicted in Figure 3, highlighting their clustering with closely related species.
The analysis of the proteolytic strains reveals distinct microbial profiles between the two regions. The Spanish samples exhibit exceptional biodiversity with remarkable performances: strains of Lactococcus lactis (QF2, OBII1, OBII2, OBII4) from fresh and sheep's milk cheeses show the highest proteolytic activity (14 mm), while Leuconostoc mesenteroides (EM3, AZ3) from blue cheese and Emmental, and Levilactobacillus brevis (EMI1, EMI2) from mixed goat-sheep milk cheeses, demonstrate activities between 12-13 mm, complemented by various Enterococcus strains (OBII3, CH5, QF3) with halo diameters of 12-13 mm. The Algerian samples show marked specialization around the Lactiplantibacillus genus, with strains KIF5 (13 mm), BP2 (12 mm), BP3 (11 mm) and KIF1-KIF2 (12 mm) demonstrating consistent activity, particularly in traditional fermented products like Kefir and Bouhaza Picante, accompanied by Enterococcus faecalis (OB1, OB4, OB5) and Leuconostoc mesenteroides (KL4) mainly isolated from sheep's milk cheeses and Klila, with halo diameters ranging between 11-13 mm. This distribution reflects both the influence of terroirs and traditional manufacturing methods on the selection of microbial communities with differentiated proteolytic capabilities. Figure 4 shows some of the inhibition halos resulting from the proteolytic activity of some of the selected strains.
4. Discussion
This study provides evidence that traditional dairy fermentations, particularly from Algeria, are a rich source of LAB with significant gluten-degrading potential. The higher sensitivity of the spot assay (56.4% positive) compared to the well diffusion assay (38.5%) strongly suggests that cell-wall-associated proteinases play a crucial role in the initial cleavage of gluten proteins, consistent with the known proteolytic system of LAB [11,21].
The most potent activity was unequivocally associated with Lactiplantibacillus plantarum and L. pentosus. This agrees with previous studies highlighting their robust proteolytic systems, including proline-specific peptidases essential for hydrolyzing proline-rich gluten peptides [13,15,22]. Our results corroborate recent studies demonstrating the efficacy of L. plantarum in degrading gliadin and reducing immunogenicity [19,23].
The significantly stronger activity in Algerian isolates may be attributed to several factors. Traditional Algerian products like Kefir and Klila are complex ecosystems involving symbiotic communities of LAB and yeasts, which may drive evolution towards enhanced proteolytic capabilities [24]. Differences in raw milk microbiota, animal diet, and spontaneous fermentation processes unique to Algerian artisanship likely contribute to selecting a more proteolytically active microbiome [17,25].
Beyond Lactiplantibacillus, we identified moderate activity in Levilactobacillus brevis and Leuconostoc mesenteroides, which have been reported to contribute to gluten hydrolysis [26]. The activity of some Enterococcus strains, while notable, necessitates careful safety evaluation due to potential virulence factors in this genus [27,28]. The variable activity observed among Lactococcus lactis strains suggests strain-specific proteolytic capabilities, though this species is generally recognized for its primary focus on lactose fermentation [11].
From an application perspective, the selected Lactiplantibacillus strains are prime candidates for development as adjunct cultures for "gluten-reduced" fermented foods [29]. Furthermore, their probiotic potential could offer dual benefits by detoxifying gluten and modulating the gut immune response [30,31]. Future work must validate efficacy under physiologically relevant conditions, characterizing hydrolysis products and assessing residual immunogenicity [32,33].
5. Conclusions
This study successfully identified a diverse collection of LAB from Spanish and Algerian artisanal dairy products with the ability to degrade gluten. The spot assay proved more sensitive for detecting this activity, highlighting the importance of cell-associated enzymes. Strains of Lactiplantibacillus plantarum and L. pentosus, particularly from Algerian Kefir and Klila, exhibited the highest degradation potential. These findings highlight the value of traditional food ecosystems as a source of microbes with unique functional properties. The selected strains represent promising biological tools for developing novel dietary strategies for individuals with celiac disease and other gluten-related disorders, though further studies are needed to validate their efficacy in vivo and assess the safety of potential probiotic applications.
Author Contributions
Conceptualization, A.M., R.P.P., and M.J.G.B.; methodology, A.M., R.P.P.; formal analysis, A.M., R.P.P., and M.J.G.B.; investigation, A.M.; resources, R.L. and A.G.; writing—original draft preparation, A.M., R.P.P., and M.J.G.B.; writing—review and editing, J.R.L. and A.G.; supervision, R.L.; funding acquisition, A.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by University of Jaén, grant number AGR230. The APC was funded by University of Jaén.
Acknowledgments
We acknowledge the university of Jaén “Plan de apoyo a la investigación”.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Iversen, R.; Sollid, L.M. The immunobiology and pathogenesis of celiac disease. Annu. Rev. Pathol. Mech. Dis. 2023, 18, 47–70. [Google Scholar] [CrossRef] [PubMed]
- Balakireva, A.V.; Zamyatnin, A.A. Properties of gluten intolerance: Gluten structure, evolution, pathogenicity and detoxification capabilities. Nutrients 2016, 8, 644. [Google Scholar] [CrossRef] [PubMed]
- Shan, L.; Molberg, Ø.; Parrot, I.; Hausch, F.; Filiz, F.; Gray, G.M.; Khosla, C. Structural basis for gluten intolerance in celiac sprue. Science 2002, 297, 2275–2279. [Google Scholar] [CrossRef] [PubMed]
- Sollid, L.M.; Jabri, B. Triggers and drivers of autoimmunity: lessons from coeliac disease. Nat. Rev. Immunol. 2013, 13, 294–302. [Google Scholar] [CrossRef]
- Lebwohl, B.; Sanders, D.S.; Green, P.H. Coeliac disease. Lancet 2018, 391, 70–81. [Google Scholar] [CrossRef]
- Lamacchia, C.; Camarca, A.; Picascia, S.; Di Luccia, A.; Gianfrani, C. Cereal-based gluten-free food: how to reconcile nutritional and technological properties of wheat proteins with safety for celiac disease patients. Nutrients 2014, 6, 575–590. [Google Scholar] [CrossRef]
- Vargas, F.M.; Cardoso, L.T.; Didoné, A.; Lima, J.P.M.; Venzke, J.G.; de Oliveira, V.R. Celiac disease: Risks of cross-contamination and strategies for gluten removal in food environments. Int. J. Environ. Res. Public Health 2024, 21, 124. [Google Scholar] [CrossRef]
- Yoosuf, S.; Makharia, G.K. Evolving therapy for celiac disease. Front. Pediatr. 2019, 7, 193. [Google Scholar] [CrossRef]
- Gänzle, M.G.; Loponen, J.; Gobbetti, M. Proteolysis in sourdough fermentations: mechanisms and potential for improved bread quality. Trends Food Sci. Technol. 2008, 19, 513–521. [Google Scholar] [CrossRef]
- Kõiv, V.; Tenson, T. Gluten-degrading bacteria: Availability and applications. Appl. Microbiol. Biotechnol. 2021, 105, 3045–3059. [Google Scholar] [CrossRef]
- Savijoki, K.; Ingmer, H.; Varmanen, P. Proteolytic systems of lactic acid bacteria. Appl. Microbiol. Biotechnol. 2006, 71, 394–406. [Google Scholar] [CrossRef] [PubMed]
- Brzozowski, B.; Stasiewicz, K.; Ostolski, M.; Adamczak, M. Reducing Immunoreactivity of Gliadins and Coeliac-Toxic Peptides Using Peptidases from L. acidophilus 5e2 and A. niger Catalysts 2020, 10(8), 923. [Google Scholar] [CrossRef]
- De Angelis, M.; Rizzello, C.G.; Fasano, A.; Clemente, M.G.; De Simone, C.; Silano, M.; Gobbetti, M. VSL#3 probiotic preparation has the capacity to hydrolyze gliadin polypeptides responsible for celiac disease. Appl. Environ. Microbiol. 2006, 72, 4490–4495. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Curiel, J.A.; Nionelli, L.; Vincentini, O.; Di Cagno, R.; Silano, M.; Coda, R. Use of fungal proteases and selected sourdough lactic acid bacteria for making wheat bread with an intermediate content of gluten. Food Microbiol. 2014, 37, 59–68. [Google Scholar] [CrossRef]
- Álvarez-Sieiro, P.; Redruello, B.; Ladero, V.; Martín, M.C.; Fernández, M.; Alvarez, M.A. Screening sourdough samples for gliadin-degrading activity revealed Lactobacillus casei strains able to individually metabolize the coeliac-disease-related 33-mer peptide. Can. J. Microbiol. 2016, 62, 422–430. [Google Scholar] [CrossRef]
- Gänzle, M.G. Enzymatic and bacterial conversions during sourdough fermentation. Food Microbiol. 2014, 37, 2–10. [Google Scholar] [CrossRef]
- Yu, J.; Wang, W.H.; Menghe, B.L.; Jiri, M.T.; Wang, H.M.; Liu, W.J.; Bao, Q.H.; Lu, Q.; Zhang, J.C.; Wang, F.; Xu, H.Y.; Sun, T.S.; Zhang, H.P. Diversity of lactic acid bacteria associated with traditional fermented dairy products in Mongolia. J. Dairy Sci. 2011, 94, 3229–3241. [Google Scholar] [CrossRef]
- Mefleh, M.; Darwish, A.M.G.; Mudgil, P.; Maqsood, S.; Boukid, F. Traditional Fermented Dairy Products in Southern Mediterranean Countries: From Tradition to Innovation. Fermentation 2022, 8, 743. [Google Scholar] [CrossRef]
- Leszczyńska, J.; Szczepankowska, A.K.; Majak, I.; Mańkowska, D.; Smolińska, B.; Ścieszka, S.; Diowksz, A.; Cukrowska, B.; Aleksandrzak-Piekarczyk, T. Reducing Immunoreactivity of Gluten Peptides by Probiotic Lactic Acid Bacteria for Dietary Management of Gluten-Related Diseases. Nutrients 2024, 16, 976. [Google Scholar] [CrossRef]
- Rashmi, B.S.; Gayathri, D.; Vasudha, M.; Prashantkumar, C.S.; Kumbhar, Y.B. Gluten hydrolyzing activity of Bacillus spp. isolated from sourdough. Microb. Cell Fact. 2020, 19, 130. [Google Scholar] [CrossRef]
- Liang, J.; Chai, K.; Wang, X.; Li, J.; Zhu, L.; Zhang, G. From gluten structure to immunogenicity: Investigating the effects of lactic acid bacteria and yeast co-fermentation on wheat allergenicity in steamed buns. Food Chem. 2025, 492, 145356. [Google Scholar] [CrossRef] [PubMed]
- Rizzello, C.G.; De Angelis, M.; Di Cagno, R.; Camarca, A.; Silano, M.; Losito, I.; Gobbetti, M. Highly efficient gluten degradation by lactobacilli and fungal proteases during food processing: New perspectives for celiac disease. Appl. Environ. Microbiol. 2007, 73, 4499–4507. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Jia, X.; Liu, C.; Meng, X.; Zhang, K.; Tao, S.; Xue, W. Sourdough yeast-bacteria interactions results in reduced immunogenicity by increasing depolymerization and hydrolysis of gluten. Innov. Food Sci. Emerg. Technol. 2023, 84, 103281. [Google Scholar] [CrossRef]
- Usal, M.; Özgölet, M.; Arici, M.; Törnük, F. Enzymatic and antimicrobial activities of lactic acid bacteria and yeasts isolated from boza, a traditional fermented grain based beverage. Food Biosci. 2024, 61, 104681. [Google Scholar] [CrossRef]
- Zolnikova, O.; Dzhakhaya, N.; Bueverova, E.; Sedova, A.; Kurbatova, A.; Kryuchkova, K.; Butkova, T.; Izotov, A.; Kulikova, L.; Yurku, K.; et al. The Contribution of the Intestinal Microbiota to the Celiac Disease Pathogenesis along with the Effectiveness of Probiotic Therapy. Microorganisms 2023, 11, 2848. [Google Scholar] [CrossRef]
- Mickowska, B.; Romanova, K.; Urminská, D. Reduction of immunoreactivity of rye and wheat prolamins by lactobacilli and flavourzyme proteolysis during sourdough fermentation—A way to obtain low-gluten bread. J. Food Nutr. Res. 2019, 58, 153–166. Available online: http://www.vup.sk/en/download.php?bulID=2015.
- Hanchi, H.; Mottawea, W.; Sebei, K.; Hammami, R. The genus Enterococcus: Between probiotic potential and safety concerns—An update. Front. Microbiol. 2018, 9, 1791. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.; Harris, H.M.; Mattarelli, P.; Lebeer, S. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Francavilla, R.; Cristofori, F.; Vacca, M.; Barone, M.; De Angelis, M. Advances in understanding the potential therapeutic applications of gut microbiota and probiotic mediated therapies in celiac disease. Expert Rev. Gastroenterol. Hepatol. 2020, 14, 323–333. [Google Scholar] [CrossRef]
- Cristofori, F.; Dargenio, V.N.; Dargenio, C.; Miniello, V.L.; Barone, M.; Francavilla, R. Anti-inflammatory and immunomodulatory effects of probiotics in gut inflammation: A door to the body. Front. Immunol. 2021, 12, 578386. [Google Scholar] [CrossRef]
- Salvi, P.S.; Cowles, R.A. Butyrate and the Intestinal Epithelium: Modulation of Proliferation and Inflammation in Homeostasis and Disease. Cells 2021, 10, 1775. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Curiel, J.A.; Nionelli, L.; Vincentini, O.; Di Cagno, R.; Silano, M.; Gobbetti, M.; Coda, R. Use of fungal proteases and selected sourdough lactic acid bacteria for making wheat bread with an intermediate content of gluten. Food Microbiol. 2014, 37, 59–68. [Google Scholar] [CrossRef]
- Kõiv, V.; Tenson, T. Gluten-degrading bacteria: availability and applications. Appl. Microbiol. Biotechnol. 2021, 105, 3045–3059. [Google Scholar] [CrossRef]
Figure 1.
Percentage of Positive Isolates by Test and Origin.
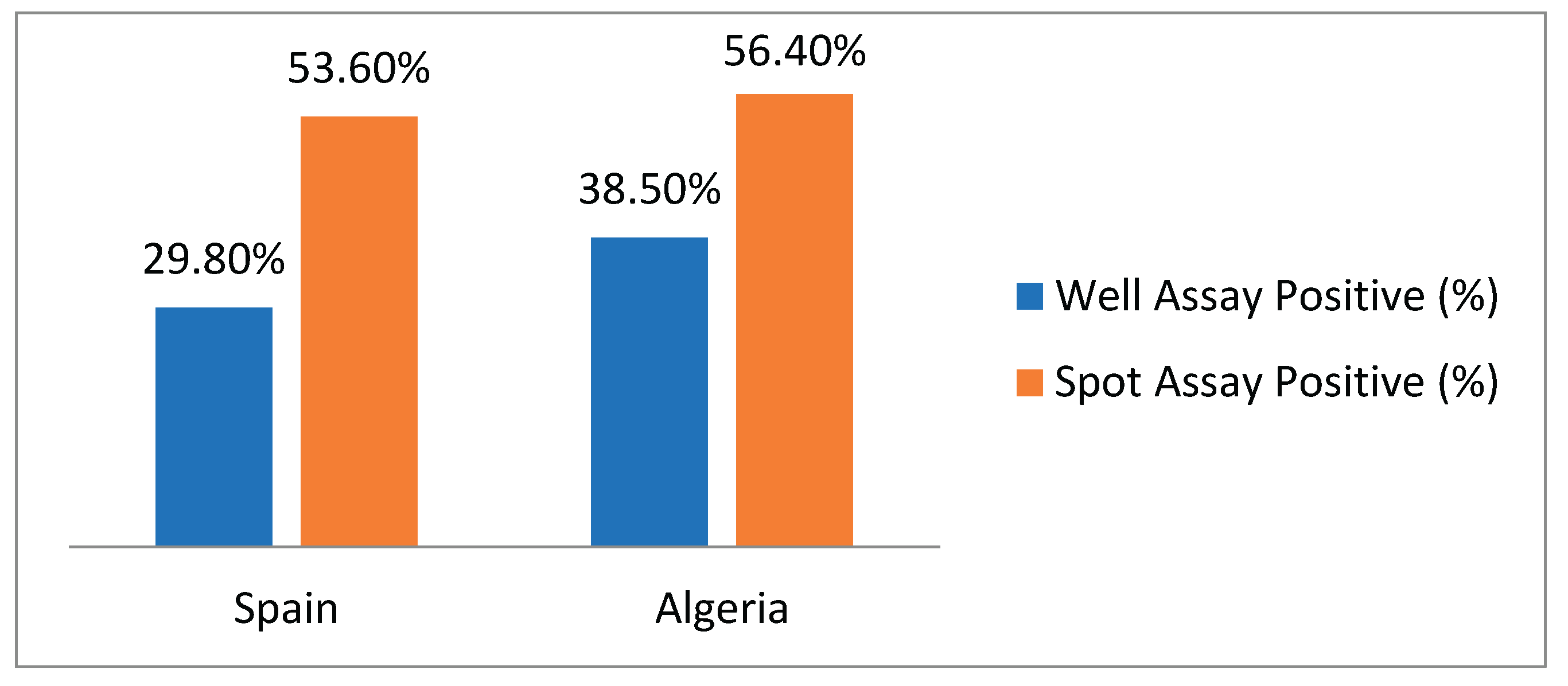
Figure 2.
Taxonomic Distribution of the Most Active Isolates.
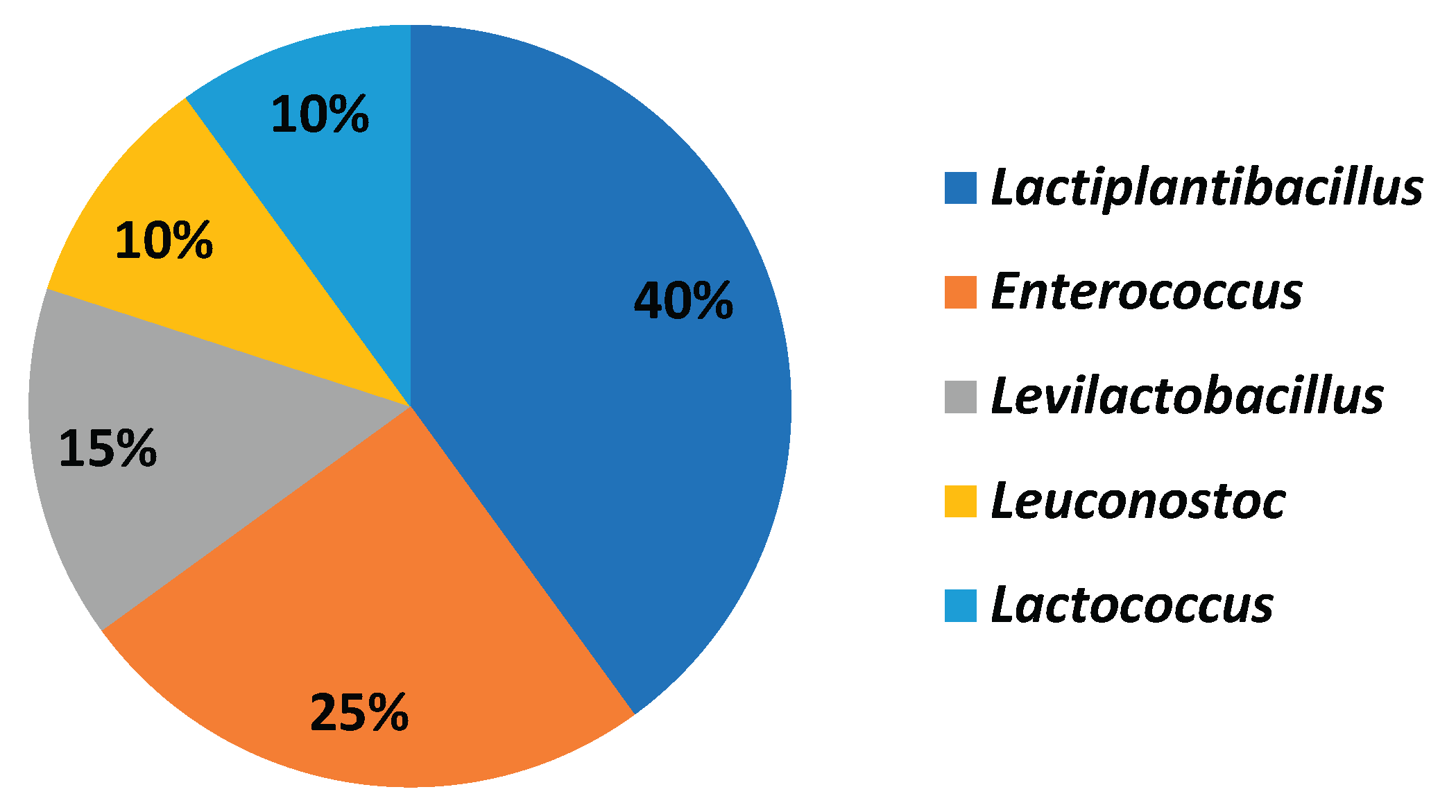
Figure 3.
Phylogenetic Tree of Selected Strains.
Figure 4.
Proteolytic Activity of Selected Strains.


Table 1.
Nomenclature of collected samples.
| Spanish Samples (ES) | Algerian Samples (DZ) |
|---|---|
| QB: Cow Cheese | BZ: Traditional Bouhaza Cheese |
| LF: Fermented Milk | BP: Spicy Bouhaza |
| QS: Solid Cheese | CAB: Goat Cheese |
| CH: Goat Cheese | KIF: Kefir |
| QF: Fresh Cheese | CAM: Camembert Cheese |
| BO: Cow and Sheep Cheese | QS: Solid Cheese |
| CUI: Cooked Cheese | QG: Gouda Cheese |
| B: Buffalo Cheese | CUI: Cooked Cheese |
| OB: Sheep Cheese | QFS: Fresh Solid Cheese |
| COV: Goat and Sheep Cheese | OBN: Sheep Cheese |
| CAB: Goat Cheese | QF: Fresh Cheese |
| AZ: Blue Cheese | OB: Sheep Cheese |
| EM: Emmental Cheese | KL: Klila (Traditional Solid Fermented Milk) |
| LF: Fermented Milk |
Table 2.
Results of Spanish Samples (ES).
| Sample | Total Isolates | Well Assay Positive (n) | Well Assay Positive (%) | Spot Assay Positive (n) | Spot Assay Positive (%) |
|---|---|---|---|---|---|
| QB | 3 | 0 | 0.0% | 3 | 100.0% |
| LF | 6 | 0 | 0.0% | 3 | 50.0% |
| QS | 1 | 0 | 0.0% | 1 | 100.0% |
| CH | 4 | 1 | 25.0% | 1 | 25.0% |
| QF | 5 | 4 | 80.0% | 3 | 60.0% |
| BO | 5 | 0 | 0.0% | 4 | 80.0% |
| CUI | 3 | 0 | 0.0% | 3 | 100.0% |
| B | 13 | 1 | 7.7% | 6 | 46.2% |
| OB | 9 | 5 | 55.6% | 7 | 77.8% |
| COV | 10 | 4 | 40.0% | 5 | 50.0% |
| CAB | 5 | 3 | 60.0% | 5 | 100.0% |
| AZ | 10 | 3 | 30.0% | 2 | 20.0% |
| EM | 10 | 4 | 40.0% | 2 | 20.0% |
| Total | 84 | 25 | 29.8% | 45 | 53.6% |
Table 3.
Results of Algerian Samples (DZ).
| Sample | Total Isolates | Well Assay Positive (n) | Well Assay Positive (%) | Spot Assay Positive (n) | Spot Assay Positive (%) |
|---|---|---|---|---|---|
| BZ | 1 | 0 | 0.0% | 0 | 0.0% |
| BP | 3 | 3 | 100.0% | 3 | 100.0% |
| CAB II | 5 | 0 | 0.0% | 4 | 80.0% |
| KIF | 3 | 3 | 100.0% | 3 | 100.0% |
| CAM | 5 | 0 | 0.0% | 4 | 80.0% |
| QS | 1 | 0 | 0.0% | 0 | 0.0% |
| QG | 1 | 0 | 0.0% | 0 | 0.0% |
| CUI | 1 | 0 | 0.0% | 0 | 0.0% |
| QFS | 5 | 0 | 0.0% | 1 | 20.0% |
| OBN | 2 | 0 | 0.0% | 2 | 100.0% |
| QF | 1 | 0 | 0.0% | 1 | 100.0% |
| CAB1 | 1 | 0 | 0.0% | 1 | 100.0% |
| OB | 5 | 5 | 100.0% | 0 | 0.0% |
| KL | 4 | 4 | 100.0% | 3 | 75.0% |
| Total | 39 | 15 | 38.5% | 22 | 56.4% |
Table 4.
Summary of gluten-degrading activity among LAB isolates.
| Origin | Total Isolates | Well Assay Positive (%) | Spot Assay Positive (%) | Mean Halo Diameter ± SD (mm) | Halo Range (mm) |
|---|---|---|---|---|---|
| Spain | 84 | 25 (29.8%) | 45 (53.6%) | 10.2 ± 2.0 | 6 – 14 |
| Algeria | 39 | 15 (38.5%) | 22 (56.4%) | 12.6 ± 2.1 | 8 – 16 |
| Total | 123 | 40 (32.5%) | 67 (54.5%) | 11.1 ± 2.3 | 6 – 16 |
Table 5.
Taxonomic identification and activity of selected gluten-degrading LAB isolates.
| Isolate Code | Closest Relative | Origin | Halo (mm) |
|---|---|---|---|
| BP1 | Lactiplantibacillus pentosus | DZ | 10 |
| BP2 | Lactiplantibacillus pentosus | DZ | 12 |
| BP3 | Lactiplantibacillus plantarum | DZ | 11 |
| KIF1 | Lactiplantibacillus plantarum | DZ | 12 |
| KIF2 | Lactiplantibacillus plantarum | DZ | 12 |
| KIF5 | Lactiplantibacillus plantarum | DZ | 13 |
| KL3 | Enterococcus faecalis | DZ | 11 |
| KL4 | Leuconostoc mesenteroides | DZ | 11 |
| OB1 | Enterococcus faecalis | DZ | 13 |
| OB3 | Enterococcus faecalis | DZ | 10 |
| OB4 | Enterococcus faecalis | DZ | 12 |
| OB5 | Enterococcus faecalis | DZ | 12 |
| CH5 | Enterococcus faecium | ES | 12 |
| QF1 | Enterococcus faecium | ES | 11 |
| QF2 | Lactococcus lactis | ES | 14 |
| QF3 | Enterococcus faecium | ES | 12 |
| QF5 | Enterococcus faecium | ES | 10 |
| CAB1 | Levilactobacillus brevis | ES | 11 |
| CAB4 | Enterococcus faecium | ES | 10 |
| CAB5 | Enterococcus hirae | ES | 10 |
| EM3 | Leuconostoc mesenteroides | ES | 13 |
| BI2 | Leuconostoc mesenteroides | ES | 10 |
| COVI1 | Levilactobacillus brevis | ES | 10 |
| AZ3 | Leuconostoc mesenteroides | ES | 12 |
| EMI1 | Levilactobacillus brevis | ES | 13 |
| EMI2 | Levilactobacillus brevis | ES | 12 |
| EMI3 | Levilactobacillus brevis | ES | 10 |
| OBII1 | Lactococcus lactis | ES | 14 |
| OBII2 | Lactococcus lactis | ES | 13 |
| OBII3 | Enterococcus faecalis | ES | 13 |
| OBII4 | Lactococcus lactis | ES | 12 |
| OBII5 | Lactiplantibacillus pentosus | ES | 13 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



