Submitted:
15 January 2026
Posted:
19 January 2026
You are already at the latest version
Abstract
Sexual dimorphism in morphological traits is widespread across animals and can result from differing life-history strategies, sex-specific competitive pressures, and ecological interactions, which may be influenced by habitat structure and complexity. For epifaunal organisms, e.g., amphipods that inhabit structurally diverse benthic habitats, the structure of the habitat plays a key role in mediating access to food, mate encounters, and refuge provision. Here, we explored patterns of variation in body size and gnathopod 2 ratio (gnathopod 2 length/body length) among amphipod species in different marine habitats. We focus on two amphipod species, Ampithoe ramondi and Caprella acanthifera, across four benthic habitat types: rhodolith beds, macroalgae-
dominated reefs, seagrass meadows, and black coral forests. A. ramondi was present in all habitats except black coral forests, and males were significantly larger than females only in macroalgae-dominated reefs. Males also exhibited higher gnathopod ratios than females, increasing from macroalgae-dominated reefs to seagrass meadows and rhodolith beds. C. acanthifera was found in macroalgae-dominated reeds and black coral forests, where males were larger than females on average, but no significant habitat alterations were detected. Neither A. ramondi nor C. acanthifera was found in all four habitats. These results suggest that patterns of sexual dimorphism across coastal habitats are species-specific, with sexual selection operating more subtly in some taxa (e.g., C. acanthifera) than others, likely shaped by both habitat-specific ecological pressures and differences in life-history strategies. Expanding such analyses to more taxa and with balanced sampling across habitats and environmental gradients will offer deeper insight into how natural and sexual selection interact and inform how these dynamics may shift under changing climate regimes.
Keywords:
Amphipoda
; sexual dimorphism
; sexual selection
; marine ecology
; Canary Islands
1. Introduction
Sexual dimorphism refers to differences between males and females of a species, which can result from sexual or fecundity selection or from intersexual niche divergence [1,2,3]. These differences may be morphological, physiological, or behavioral, and their magnitude and direction are often shaped by abiotic (e.g., temperature, nutrient availability or habitat structure) and biotic factors (e.g., predation pressure or interactions with other organisms) [1,4]. For example, shared parental care, intraspecific competition, or mate availability can limit the development of certain traits, reducing dimorphism and favoring more similar (monomorphic) characteristics [5]. By contrast, when sexual selection strongly favors one sex, intense competition for mates can drive the evolution of exaggerated traits and behaviors, resulting in pronounced sexual dimorphism [2,6,7]. While sexual dimorphism across habitats has been widely studied in terrestrial organisms [8,9], there are very few studies in marine systems examining how sexual dimorphism varies within the same species across different habitats [10,11,12], which limits our understanding of how environmental context shapes the expression of dimorphic traits.
The Amphipoda is one of the most ecologically diverse crustacean orders, encompassing over 10,000 species [13,14] inhabiting all aquatic environments worldwide, from hadal depths to alpine freshwater streams, from lightless groundwaters to tropical forests, and from sea bottom sediments to the entrails of gelatinous plankton [15]. Amphipods display diverse mating strategies and pronounced sexual dimorphism, particularly in the enlarged male gnathopods, providing an opportunity to examine how ecological context shapes trait expression [16,17]. The first two pairs of pereopods are modified into gnathopods, with the second pair typically enlarged in males and used during copulation [18]. Such marked dimorphism in gnathopods suggests a role as sexually selected traits, analogous to weapons in other crustaceans and insects [19], as proposed or demonstrated in their use in intrasexual contests, territorial interactions, courtship displays, and assessment of female quality [20]. Although direct evidence for amphipod gnathopods as weapons is limited, both their morphology and associated behaviors indicate a role in mating competition, potentially shaped by selective pressures, such as mate guarding or signally in complex habitats [21]. Thus, understanding the functional morphology of sexually dimorphic amphipod structures necessitates the consideration of the ecological context, including habitat type, host associations, and feeding ecology, which influence the selective pressures acting on these traits [20,22,23]. Environmental conditions and habitat heterogeneity are expected to influence the intensity and expression of sexual dimorphism, for example, through intraspecific trait variation created by different selective pressures across gradients, and thus favouring distinct phenotypes in response to local conditions such as physical disturbance or resource availability [24]. At the habitat level, structural complexity plays a crucial role in shaping biodiversity by influencing species abundance, community composition, and key ecological processes. Complex habitats, such as dense macroalgae-dominated reefs or structurally diverse coral reefs, often provide increased shelter and refuge, which can reduce predation risk and enhance survival, particularly by allowing the coexistence of a wider range of phenotypes [25,26]. However, this relationship is not unidirectional. In habitats with lower structural complexity, limited refuge, and reduced food availability, such as black coral forests lacking epiphytic layers, smaller body sizes may be favored as a strategy to reduce predation risk and energetic demands under resource limitation [16]. From an evolutionary perspective, shifts in body size reflect trade-offs between ecological relaxation driven by empty niches and resource abundance versus constraints imposed by limited resources, a pattern well established in island biogeography [27]. The habitat complexity can also affect mate encounter rates and resource distribution, favoring the coexistence of multiple phenotypes within a population [1,4]. For example, in amphipods, habitats with intricate structures may allow males with different sizes or gnathopod morphologies to succeed under different microhabitat conditions, highlighting the role of habitat architecture in modulating sexual dimorphism [28].
The coastal habitats of the Canary Islands, which include black coral forests, rhodolith beds, seagrass meadows and macroalgae-dominated beds on reefs [29,30,31,32]. Such ecosystem engineers are species or communities that modify, create, or maintain habitat structures, thereby influencing resource availability, shelter, and interactions among other organisms [33,34]. These complex habitats can affect survival, predation risk, mate encounters, and the expression of morphological and behavioural traits. Although all four habitats share three-dimensional architecture, they differ in structural arrangement, a factor that may shape the selective pressures acting on amphipods and the development of sexually dimorphic traits [28]. Amphipods, in particular, maintain a close relationship with their habitats due to their direct development, limited mobility and, in some cases, specialized feeding strategies, which constrain their dispersal [16,35]. Taking advantage of this strong habitat association, we studied amphipods across habitats with varying structural complexity to assess how these differences influence trait expression and sexual dimorphism. We sampled amphipod species across the four aforementioned benthic habitats in Gran Canaria. Morphological traits (i.e., body length and gnathopod 2 size) were measured as functional traits to assess how habitat influences sexual dimorphism. We focused on two amphipod species, A.ramondi and C.acanthifera, exhibiting contrasting morphologies and mating strategies, allowing us to explore how sexual dimorphism varies across habitats. We hypothesize that sexual dimorphism is more pronounced in structurally intricate habitats, where fine-scale complexity intensifies selective pressures.
2. Materials and Methods
2.1. Study Region
Specimens were collected from two locations at each of four marine habitats around the eastern coastal areas of Gran Canaria (Figure 1), including seagrass meadows, rhodolith beds, macroalgae-dominated reefs, and black corals. All sampling was conducted between November and December from 2018 to 2022. Because the sites are geographically close and experience similar broad-scale environmental conditions, any differences observed in sexual dimorphism among habitats are likely attributable to habitat-specific structural features rather than overall environmental variation. The eastern coast of Gran Canaria is characterized by steep volcanic slopes and a mixture of hard and soft substrates that support diverse benthic habitats. Local hydrography is influenced by persistent northeast trade winds, which generate wind-driven waves and swells [36], while surrounding waters are largely oligotrophic, with occasional inputs of cooler, nutrient-rich water from coastal upwelling [37]. The eastern shoreline features a range of subtidal habitats including extensive rocky platforms, sandy plains, and biogenic assemblages. Based on existing knowledge of habitat distribution and ecological relevance to amphipod assemblages, representative sites were selected within the four most abundant and structurally distinct habitats in the region.
2.2. Sampling Design and Specimen Processing
Sampling was conducted by SCUBA diving at depths of 18–50 m for rhodolith beds, 8–12 m for seagrass meadows, 3–8 m for macroalgae-dominated reefs, and 25–32 m for black coral forests. Samples associated with each habitat were retrieved and processed in the laboratory to extract epifaunal amphipods. For rhodolith beds, five random 25 × 25 cm quadrats of rhodolith nodules were hand-collected by SCUBA divers, frozen at −20 ∘C, followed by epifaunal extraction and sorting. For seagrass meadows, five random 25 × 25 cm quadrats containing seagrass canopy were bagged, cut at sediment, and sieved for epifauna. For macroalgae-dominated reefs, holdfasts were cut within five random 25 × 25 cm quadrats, bagged, frozen, washed, sieved, and mobile fauna was identified. Following the method described by [38], for black coral forests, epifauna was collected by briefly vacuuming the top branches of Antipathella wollastoni underwater and preserved in ethanol. Collected epifaunal samples were transported to the laboratory in seawater-filled containers. Except for black coral samples, which were processed fresh to prevent epifaunal damage, all other samples were stored at −20 ∘C until sorting. In the laboratory, each samples was inspected, and the epifauna was separated from plant and and algal matter, sediment and coral mucus using a 250 m mesh. All organisms were preserved in 96% ethanol and identified to the lowest possible taxonomic level. No ethical approval was required for all but one species, the black corals [39], as none of the other sampled species are legally protected in Spain. All individuals were sorted under a stereomicroscope (OPTIKA SZP-10, Ponteranica, Italy) and a microscope (Leica DM1000 LED, Wetzlar, Germany), identified to the nearest taxonomic level [40,41,42], and preserved in absolute ethanol. Ampithoe ramondi and Caprella acanthifera were the primary focus of this study, as sufficient individuals of both sexes were collected across habitats to allow analysis of sexual dimorphism. However, neither species was observed across all four habitats. Ampithoe ramondi occurred in three habitats, rhodolith beds, seagrass meadows and macroalgae-dominated reefs, whereas Caprella acanthifera was restricted to only two habitats, macroalgae-dominated reefs and black coral forests.
2.3. Morphometric Measurements
In the laboratory, individuals were examined under a stereomicroscope (OPTIKA, SZP- 10, C-P8, Ponteranica, Italy) mounted with its associated camera. Images were analysed using ImageJ (Schneider et al., 2012). Morphometric traits measured included body length (determined by measuring the distance from the dorsal margin at the base of the first antennae to the posterior end of the telson following the methodology described by Wilhelm et al., 1998) and Gnathopod 2 length which was calculated following the methodology of Navarro-Mayoral et al., 2025 (refer Supplementary Table 1 and Figure 2). Measurement precision was estimated at 0.01–0.05 mm, depending on image scale calibration (either 0.5 mm or 1 mm per image). To avoid underestimating mean body size, only the largest individuals were included in analyses.
2.4. Data Analysis
The Sexual Dimorphism Index (SDI) was calculated using the standard formula:
SDI = (mean male trait – mean female trait) / mean female trait,
which was applied to each of the traits analyzed, refer Supplementary Figure 2. A positive SDI value indicates that males have larger average trait values, reflecting male-biased sexual selection, whereas a negative SDI suggests female-biased sexual dimorphism, reflecting fecundity-based selection. This approach follows the widely used method proposed by [44] for quantifying sexual dimorphism in morphological traits.
Data wrangling and statistical analysis were carried out in RStudio version 2024.09.1+394 “Cranberry Hibiscus” using R version 4.4.2 [45]. For interspecific comparisons, we included a minimum of five individuals per sex of the two targeted species and fitted linear models to test the effects of species, sex, and their interaction on body length and gnathopod ratio. Gnathopod ratio was calculated by dividing the gnathopod length by the individual’s body length to account for differences in body size. For species-specific analyses, univariate linear models were applied separately to each species to test for the effects of sex and habitat on each morphometric trait. For gnathopod ratio data, which are constrained between 0 and 1, beta regression models were implemented using the glmmTMB package [46]. with a logit link. Model diagnostics- including residual distribution, heteroscedasticity, and normality, were assessed visually and using simulation-based methods via the DHARMa package [47]. We computed estimated marginal means (EMMs) using the emmeans package [48] to examine interaction effects (Sex × Habitat), followed by visualization of these contrasts. Visualizations were generated using ggplot2 [49].
3. Results
3.1. Sex-Specific Allometry Comparisons
In A. ramondi, gnathopod 2 length scaled positively with body size (, ; Figure 3), with males exhibiting a marginally steeper slope than females (, ), although this difference was not statistically significant. The sex effect alone was also non-significant (, ), indicating that while males tend to have longer gnathopods, the scaling relationship between gnathopod length and body length is broadly similar across sexes. Conversely, in C. acanthifera, neither body length (, ), sex (, ), nor their interaction (, ) significantly predicted gnathopod 2 length. These results suggest that sexual dimorphism in gnathopod 2 exaggeration is less pronounced or absent in C. acanthifera, with both sexes exhibiting comparable scaling patterns.
3.2. Species-Specific Models
3.2.1. Ampithoe ramondi
Body Length
Ampithoe ramondi occurred in rhodolith beds, macroalgae-dominated reefs, and seagrass meadows and was absent from black coral forests. Body length in Ampithoe ramondi was significantly influenced by habitat, sex, and their interaction (adjusted , ; Figure 4). Individuals from macroalgae-dominated reefs were significantly smaller than those from rhodolith beds (, ), while body length in seagrass meadows did not differ significantly from rhodolith beds (, ). Although males were on average larger than females, this difference was not statistically significant overall (, ). A significant Sex × Habitat interaction () indicated that the effect of sex on body length varied among habitats, with the greatest difference observed in macroalgae-dominated reefs, where males were larger than females. In contrast, sex-related differences were negligible in seagrass meadows and rhodolith beds.
Gnathopod ratio
Beta regression analysis revealed significant effects of sex (p < 0.001), habitat (macroalgae-dominated reefs: p = 0.035; seagrass meadows: p = 0.016), and their interaction (male × macroalgae-dominated reefs: p < 0.001 ) on gnathopod ratio (Figure 5). The difference between males and females was negligible in macroalgal-dominated reefs, but males had higher gnathopod ratios than females in seagrass meadows and rhodolith beds.
3.2.2. Caprella acanthifera
Body Length
Caprella acanthifera was recorded only in macroalgae-dominated reefs and black coral forests and was not observed in the other sampled habitats. The linear model indicated a significant effect of sex on body length (, Figure 6), with males larger than females (, ). Habitat had a significant effect, with individuals in black coral forests being smaller than those in macroalgae-dominated reefs (, ), but the interaction between sex and habitat was not significant (, ), indicating that the male–female size difference was consistent across habitats.
Gnathopod Ratio
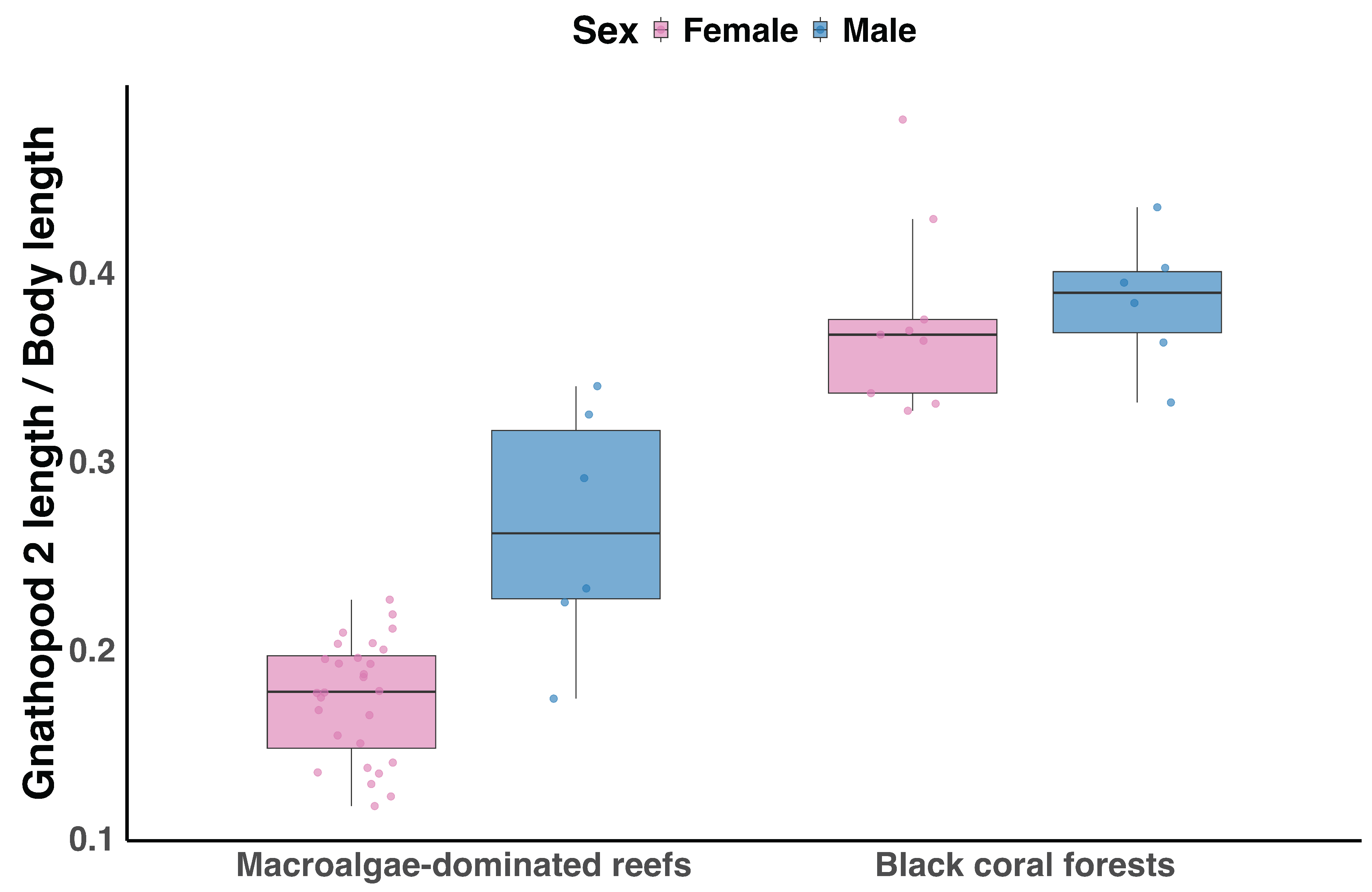
Beta regression analysis revealed significant effects of sex (p < 0.001), habitat (p < 0.001), and their interaction (p < 0.001) on gnathopod ratio in C. acanthifera (Figure 7). Males had higher gnathopod ratios than females in macroalgae-dominated reefs, whereas the male-female difference was reduced in black coral forests.
4. Discussion
Sexual dimorphism in amphipod morphology varied significantly by species and habitat. Ampithoe ramondi exhibited pronounced male-biased dimorphism in body length and gnathopod ratio, particularly in macroalgae-dominated reefs. Similarly, Caprella acanthifera showed male-biased dimorphism in body length in macroalgae-dominated reefs, but gnathopod size was similar between sexes across habitats. Differences in reproductive strategies, mating competition [50,51], and habitat-specific factors such as resource availability or predation [52] likely underlie the observed patterns. However, direct comparisons between Ampithoe ramondi and Caprella acanthifera are constrained by the fact that macroalgae-dominated reefs were the only habitat common to both species.
Habitat plays a central role in modulating trait expression. In A. ramondi, individuals from rhodolith beds were consistently larger than those from macroalgae-dominated reefs or seagrass meadows. This pattern may reflect differences in available colonizable space and niche diversity among benthic habitats, where structurally complex environments can reduce competition for space and resources and support larger body sizes [16]. Bathymetric range may further mediate these patterns through differences in environmental stability and resource availability, including light penetration and epiphytic development, which increase habitat complexity and vary temporally. Shallow habitats are often subject to strong intra-annual variability, such as winter storms and swell, favoring species adapted to fluctuating habitat structure and resource inputs, whereas deeper habitats tend to experience lower but more stable resource availability. Compared to the other habitats considered in this study, rhodolith beds occur at greater depths and are characterized by extensive interstitial spaces [53] that may buffer hydrodynamic stress, potentially allowing greater energetic investment in body growth despite reduced oxygen levels.
The largest degree of sexual dimorphism in body size in A.ramondi, occurred in macroalgae-dominated reefs, where males were significantly larger than females. Macroalgae provide both shelter [54] and trophic resources supporting diverse and stable amphipod communities [55]. They are well-established facilitators of amphipod assemblages, with more diverse macroalgae-dominated reefs supporting higher biodiversity richness due to enhanced niche partitioning and habitat complexity [56]. The three-dimensional structure of macroalgae-dominated reefs promotes sediment accumulation and increases prey availability, benefiting both detritivorous and carnivorous species [57]. Amphipods often function as mesoherbivores, consuming macroalgae directly [58], and forming close associations with these habitats [59]. This pattern may reflect higher mating competition or more favorable energetic and ecological conditions for trait exaggeration in macroalgae-dominated reefs, which offer increased food resources, structural complexity, and refuge [56,57].
4.1. Sexual Selection, Trait Scaling, and Ecological Modulation
Allometric scaling of gnathopod 2 length in A. ramondi indicated a consistent positive relationship with body size, and a trend toward steeper scaling for males. Though not statistically significant, this pattern is consistent with male-specific investment in sexually selected structures, as seen in other amphipods and crustaceans [50]. Alternatively, lower relative investment in gnathopod 2 length could reflect selection for larger overall body size, which may also influence reproductive success [21] or vice versa [20]. No such pattern was detected in C. acanthifera, where scaling relationships did not differ by sex, and trait values were more conserved across habitats. C. acanthifera exhibits limited sexual dimorphism, likely due to its mobile lifestyle and generalist diet that includes macroalgae, detritus and epiphytes, minimizing resource monopolization [60,61]. Mobility possibly reduces the need for males to defend fixed territories, lowering male–male competition. Together, these factors decrease sexual selection pressures for exaggerated male traits [62].
Unlike carrier species that grasp females by hooking unenlarged gnathopods under the coxae or anterior pereon segments, caprellid males typically guard mates by attending, i.e., maintaining close proximity, often by positioning themselves over the female or sharing her tube [50]. In these mate-guarding taxa, the enlarged gnathopods are held free and not used for grasping; instead, they are employed in mate takeovers, agonistic interactions, defense, and possibly to assess the female’s reproductive state through contact pheromones [50]. Due to their sedentary and often gregarious habits, attending males are not energetically burdened by physically carrying a mate, but since they remain close to the female and must actively defend her, there is selective pressure for mechanisms that signal dominance or reproductive quality [50]. As a result, some attending species exhibit strong sexual selection on exaggerated traits wherein some males monopolize females [51], and others defend mating clusters [63]. Several species evolve conspicuous weaponry on the gnathopods for use in fights, such as poison spines in Caprella gorgonia [64] or thumb-like projections in Dyopedos porrectus [65]. That both C. acanthifera and A. ramondi belong to mate-attending superfamilies (Caprellidea and Corophioidea, respectively) yet show contrasting levels of dimorphism suggests that even within a shared guarding strategy, habitat use, and competitive context can shape the strength and nature of sexual selection.
These findings suggest that sexual dimorphism in Caprelloidea may be less labile or subject to different selective pressures compared to Corophioidea.
4.2. Ecological Complexity and Habitat Structure
A. ramondi and C. acanthifera share broadly similar feeding habits: both species rely heavily on detritus (over 65% of their diets), with A. ramondi supplementing its detritivorous diet with herbivory [66,67] while C. acanthifera also consuming algae as an herbivore and exhibiting omnivory [68]. Despite this overlap in dietary preferences, differences in their mobility, habitat use, competitive strategies, or mating behaviours may better explain variations in sexual dimorphism than diet alone.
The four habitats studied, viz. rhodolith beds, macroalgae-dominated reefs, seagrass meadows, and black coral forests, differ markedly in structural complexity, resource distribution, and potential for predator avoidance. Macroalgae-dominated reefs are relatively open but structurally intricate at a small scale, offering numerous attachment sites and food sources [54], possibly enhancing male–female encounter rates and facilitating the evolution of conspicuous or exaggerated male traits. Rhodolith beds, composed of free-living coralline algae, form three-dimensional matrices that vary in complexity depending on rhodolith size and aggregation [25]; they may support high amphipod densities and moderate encounter rates but impose spatial constraints on movement. Seagrass meadows, while highly productive, are often more homogenous in structure [32] and may support lower refuge from predation compared to macroalgae-dominated reefs or rhodolith beds, possibly dampening selective pressures for extreme trait expression. However, more structurally complex seagrass meadows have been shown to support higher densities and species richness of amphipods [26], suggesting that ranking habitats by complexity and heterogeneity is not always straightforward.
In contrast, black coral forests are among the most complex and vertically stratified habitats. Despite this, black coral forests can support high predator diversity and, particularly in deeper habitats, experience low light levels, which may limit active mate searching and favor cryptic behaviors or morphologies [38]. Their three-dimensional structure atop rocky substrates also creates numerous microhabitats that support diverse associated species [69]. When sufficiently dense, they form dense aggregations known as ‘coral forests’ (a type of marine animal forest, see [70]) and provide protection against currents and predators [71]. Thus, habitat characteristics likely shape amphipod abundance and behavior, but their influence on trait exaggeration and sexual dimorphism varies between species, reflecting species-specific responses to ecological complexity.
4.3. Future Directions
While this study demonstrates the influence of habitat and sex on amphipod morphological traits, incorporating adult sex ratio (ASR) would provide deeper insight into the demographic mechanisms driving sexual dimorphism. ASR is often used as a proxy for the strength and direction of sexual selection [72], and its variation across habitats may correlate with shifts in dimorphism intensity. For example, female-biased ASRs, potentially arising in energetically constrained or high-predation habitats, could lead to reduced male-male competition and diminished exaggeration of sexually selected traits [73]. Conversely, male-biased ASRs may amplify selection for competitive traits and accentuate dimorphism [73]. ASR could not be reliably calculated due to sampling limitations, including selective counts (e.g., stopping after obtaining enough males in A. ramondi) and damaged specimens that prevented sex identification and use in analysis.
As habitat structure shifts under ongoing environmental change, these alterations will reshape the ecological context in which sexual traits evolve. Seagrass meadows which are key ecosystem engineers, support diverse epifaunal communities as well as epiphytic macroalgae [74], are experiencing widespread decline at local, regional, and global scales [75]. Under changing environmental conditions, sexual selection may increase rates of adaptation and aid evolutionary rescue [76]. In this context, tracking future trends in sexual dimorphism, and how such changes influence both population dynamics and life-history traits like mating behavior and reproductive strategies, will be crucial. Temperature-dependent increases in amphipod metabolic rates can accelerate life history traits such as growth and reproduction [77], potentially leading to higher population growth and faster age at maturity. These shifts may intensify competition for mates or alter mate choice dynamics, ultimately influencing sexual selection intensity and contributing to variation in sexual dimorphism. Future studies should explore how these temperature-driven changes impact sexual selection processes and sexual dimorphism patterns.
A key limitation of this study is the absence of direct measurements of habitat complexity, which constrains our ability to explicitly link morphological variation to environmental structure. Future research should therefore include quantitative assessments of habitat complexity to better connect habitat heterogeneity with sexually dimorphic trait divergence.
5. Conclusions
Our findings show that sexual dimorphism in amphipod morphology is influenced by both sex-specific selective pressures and ecological context, with outcomes that vary across species and habitats. Ampithoe ramondi demonstrated clear sexual dimorphism in body size and gnathopod lengths, with habitat-specific expression suggesting plasticity or condition-dependent trait development. In contrast, Caprella acanthifera showed weaker, more conserved patterns. Due to limited within-species comparisons across all four habitats and mixed patterns across traits, no uniform relationship could be established. The interaction between natural and sexual selection, modulated by habitat structure and possibly adult sex ratios, governs the expression of sexually dimorphic traits. Species with greater plasticity in trait development may adjust their morphological strategies depending on local competition, resource availability, or predation risk.
The inclusion of demographic, behavioral, and genetic variables in future studies will be essential to disentangle these complex evolutionary pressures. While sexual selection may not strongly drive adaptation under changing environmental conditions, it remains relevant. Environmental shifts can alter the costs of sexually selected traits, potentially leading to evolutionary hedging of those traits.Habitats such as seagrass meadows are often structurally simple, but can attain fine-scale complexity through epiphytic communities, including macroalgae, which increase surface area, enhance nutrient-rich food availability, provide habitat for epifauna, and modulate predation pressure [78,79]. Rhodolith beds too gain a secondary structural layer from epiphytic algae, creating a cascade effect that enhances habitat complexity. These epiphytes provide numerous microhabitats, refuge from predators, and a wider range of body sizes, reducing intra- and interspecific competition. Macroalgae-dominated reefs similarly form dense, structurally intricate habitats that trap sediment and detritus, supplying both physical structure and trophic resources. In contrast, black coral forests lack epiphytic layers, resulting in a more uniform and competitive environment. Accounting for these layered habitat components in future studies will be crucial for better understanding how habitat-specific factors influence the intensity and expression of sexual dimorphism.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org
Author Contributions
Conceptualization, A.D., F.T. and S.N.-M.; Methodology, S.N.-M., F.O.-F., F.T. and S.D.-V.; Validation, F.T. and S.N.-M.; Formal analysis, A.D.; Investigation, A.D.; Data curation, A.D.; Writing—original draft preparation, A.D.; Writing—review and editing, A.D., F.T., S.N.-M. and F.O.-F.; Visualization, A.D.; Supervision, F.T. and S.N.-M.; Project administration, F.T. and S.N.-M. All authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
All procedures involving animals in this study were conducted in accordance with the ethical guidelines and regulations of Universidad de Las Palmas de Gran Canaria.
Data Availability Statement
All data and analysis scripts used in this study, including the final processed dataset and R scripts for analyses, are publicly available in the following GitHub repository: https://github.com/amey-danole/Habitat-driven-variation-in-sexual-dimorphism-of-amphipods.
Acknowledgments
I would like to sincerely thank Lucia Castillo for her daily guidance, valuable insights, and support throughout the project, which greatly contributed to the development of this work. During the preparation of this manuscript, I used ChatGPT (GPT-5 mini) to improve clarity, conciseness, and overall flow of the writing, as well as to assist with analyzing results, including generating R scripts and interpreting outcomes. I have reviewed and edited all outputs and take full responsibility for the content of this publication.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results’.
References
- Shine, R. Ecological causes for the evolution of sexual dimorphism: A review of the evidence. The Quarterly Review of Biology 1989, 64, 419–461. [Google Scholar] [CrossRef]
- Andersson, M. Sexual selection; Princeton University Press, 1994. [Google Scholar]
- Stillwell, R.C.; Blanckenhorn, W.U.; Teder, T.; Davidowitz, G.; Fox, C.W. Sex differences in phenotypic plasticity affect variation in sexual size dimorphism in insects: From physiology to evolution. Annual Review of Entomology 2010, 55, 227–245. [Google Scholar] [CrossRef]
- Endler, J.A. Signals, signal conditions, and the direction of evolution. American Naturalist 1992, 139, S125–S153. [Google Scholar] [CrossRef]
- Clutton-Brock, T. Sexual selection in males and females. Science 2007, 318, 1882–1885. [Google Scholar] [CrossRef]
- Darwin, C. The descent of man, and selection in relation to sex; John Murray, 1871; Vol. 1. [Google Scholar]
- Lande, R. Sexual dimorphism, sexual selection, and adaptation in polygenic characters. Evolution 1980, 34, 292–305. [Google Scholar] [CrossRef]
- Ford, S.M. Evolution of sexual dimorphism in body weight in platyrrhines. American Journal of Primatology 1994, 34, 221–244. [Google Scholar] [CrossRef]
- Geist, V.; Bayer, M. Sexual dimorphism in the Cervidae and its relation to habitat. Journal of Zoology 1988, 214, 45–53. [Google Scholar] [CrossRef]
- Cleasby, I.R.; Wakefield, E.D.; Bodey, T.W.; Davies, R.D.; Patrick, S.C.; Newton, J.; Votier, S.C.; Bearhop, S.; et al. Sexual segregation in a wide-ranging marine predator is a consequence of habitat selection. Marine Ecology Progress Series 2015, 518, 1–12. [Google Scholar] [CrossRef]
- Losos, J.B.; Butler, M. Sexual Dimorphism in Body Size and Shape in Relation to Habitat Use among. Lizard social behavior 2003, 356. [Google Scholar]
- Wikelski, M.; Trillmich, F. Body size and sexual size dimorphism in marine iguanas fluctuate as a result of opposing natural and sexual selection: An island comparison. Evolution 1997, 51, 922–936. [Google Scholar] [CrossRef]
- Arfianti, T.; Wilson, S.; Costello, M.J. Progress in the discovery of amphipod crustaceans. PeerJ 2018, 6, e5187. [Google Scholar] [CrossRef]
- Horton, T.; De Broyer, C.; Bellan-Santini, D.; Coleman, C.O.; Copilaș-Ciocianu, D.; Corbari, L.; Daneliya, M.E.; Dauvin, J.C.; Decock, W.; Fanini, L.; et al. The World Amphipoda Database: History and progress. Australian Museum 2023. [Google Scholar]
- Lowry, J.K.; Myers, A.A. A phylogeny and classification of the Amphipoda with the establishment of the new order Ingolfiellida (Crustacea: Peracarida). Zootaxa 2017, 4265, 1–89. [Google Scholar] [CrossRef]
- Navarro-Mayoral, S.; Díaz-Vergara, S.; Bosch, N.E.; Tuya, F.; Bramanti, L.; Fernandez-Gonzalez, V.; Terrana, L.; Espino, F.; Haroun, R.; Otero-Ferrer, F. Inside the mesophotic zone: Taxonomic and trait diversity of epifauna associated with black coral forests across an oceanic archipelago. Coral Reefs 2025, 1–19. [CrossRef]
- Copilaș-Ciocianu, D.; Boros, B.V.; Šidagytė-Copilas, E. Morphology mirrors trophic niche in a freshwater amphipod community. Freshwater Biology 2021, 66, 1968–1979. [Google Scholar] [CrossRef]
- Hume, K.D.; Elwood, R.W.; Dick, J.T.; Morrison, J. Sexual dimorphism in amphipods: The role of male posterior gnathopods revealed in Gammarus pulex. Behavioral Ecology and Sociobiology 2005, 58, 264–269. [Google Scholar] [CrossRef]
- Emlen, D.J. The evolution of animal weapons. Annual review of ecology, evolution, and systematics 2008, 39, 387–413. [Google Scholar] [CrossRef]
- Wellborn, G.A. Selection on a sexually dimorphic trait in ecotypes within the Hyalella azteca species complex (Amphipoda: Hyalellidae). The American Midland Naturalist 2000, 143, 212–225. [Google Scholar] [CrossRef]
- Clutton-Brock, T. The functions of antlers. Behaviour 1982, 79, 108–124. [Google Scholar] [CrossRef]
- Wellborn, G.A. Trade-off between competitive ability and antipredator adaptation in a freshwater amphipod species complex. Ecology 2002, 83, 129–136. [Google Scholar] [CrossRef]
- Wellborn, G.A.; Bartholf, S.E. Ecological context and the importance of body and gnathopod size for pairing success in two amphipod ecomorphs. Oecologia 2005, 143, 308–316. [Google Scholar] [CrossRef]
- Bolnick, D.I.; Svanbäck, R.; Fordyce, J.A.; Yang, L.H.; Davis, J.M.; Hulsey, C.D.; Forister, M.L. The ecology of individuals: Incidence and implications of individual specialization. The American Naturalist 2003, 161, 1–28. [Google Scholar] [CrossRef]
- Navarro-Mayoral, S.; Fernandez-Gonzalez, V.; Otero-Ferrer, F.; Tuya, F. Spatio-temporal variability of amphipod assemblages associated with rhodolith seabeds. Marine and Freshwater Research 2020, 72, 76–83. [Google Scholar] [CrossRef]
- Navarro-Mayoral, S.; Tuya, F.; Prado, P.; Marco-Méndez, C.; Fernandez-Gonzalez, V.; Fernández-Torquemada, Y.; Espino, F.; de la Ossa, J.A.; Vilella, D.M.; Machado, M.; et al. Drivers of variation in seagrass-associated amphipods across biogeographical areas. Marine Environmental Research 2023, 186, 105918. [Google Scholar] [CrossRef]
- Fernández-Palacios, J.M.; et al. Introducción a las islas. Ecología Insular/Island Ecology 2004, 21–55. [Google Scholar]
- Premate, E.; Fišer, Ž.; Biró, A.; Copilaş-Ciocianu, D.; Fromhage, L.; Jennions, M.; Borko, Š.; Herczeg, G.; Balázs, G.; Kralj-Fišer, S.; et al. Sexual dimorphism in subterranean amphipod crustaceans covaries with subterranean habitat type. Journal of Evolutionary Biology 2024, 37, 487–500. [Google Scholar] [CrossRef] [PubMed]
- Bosch, N.E.; Espino, F.; Tuya, F.; Haroun, R.; Bramanti, L.; Otero-Ferrer, F. Black coral forests enhance taxonomic and functional distinctiveness of mesophotic fishes in an oceanic island: Implications for biodiversity conservation. Scientific Reports 2023, 13, 4963. [Google Scholar] [CrossRef]
- Pérez-Peris, I.; Navarro-Mayoral, S.; de Esteban, M.C.; Tuya, F.; Peña, V.; Barbara, I.; Neves, P.; Ribeiro, C.; Abreu, A.; Grall, J.; et al. Effect of depth across a latitudinal gradient in the structure of rhodolith seabeds and associated biota across the eastern Atlantic Ocean. Diversity 2023, 15, 103. [Google Scholar] [CrossRef]
- Alfonso, B.; Hernández, J.C.; Sangil, C.; Martín, L.; Expósito, F.J.; Díaz, J.P.; Sansón, M. Fast climatic changes place an endemic Canary Island macroalga at extinction risk. Regional Environmental Change 2021, 21, 113. [Google Scholar] [CrossRef]
- Barbera, C.; Tuya, F.; Boyra, A.; Sanchez-Jerez, P.; Blanch, I.; Haroun, R.J. Spatial variation in the structural parameters of Cymodocea nodosa seagrass meadows in the Canary Islands: A multiscaled approach. In Botanica marina (Print); 2005. [Google Scholar]
- Jones, C.G.; Lawton, J.H.; Shachak, M. Organisms as ecosystem engineers. In Ecosystem management: Selected readings; Springer, 1994; pp. 130–147. [Google Scholar]
- Feldens, P.; Held, P.; Otero-Ferrer, F.; Bramanti, L.; Espino, F.; Schneider von Deimling, J. Can black coral forests be detected using multibeam echosounder “multi-detect” data? Frontiers in Remote Sensing 2023, 4, 988366. [Google Scholar] [CrossRef]
- Fernandez-Gonzalez, V.; Navarro-Mayoral, S.; Sanchez-Jerez, P. Connectivity patterns for direct developing invertebrates in fragmented marine habitats: Fish farms fouling as source population in the establishment and maintenance of local metapopulations. Frontiers in Marine Science 2021, 8, 785260. [Google Scholar] [CrossRef]
- Smith, A.; Jackson, D.W.; Cooper, J.A.G.; Beyers, M.; Breen, C. Whole-island wind bifurcation and localized topographic steering: Impacts on aeolian dune dynamics. Science of the Total Environment 2021, 763, 144444. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, J.; Arístegui, J.; Hernández-Hernández, N.; Fernández-Méndez, M.; Riebesell, U. Oligotrophic phytoplankton community effectively adjusts to artificial upwelling regardless of intensity, but differently among upwelling modes. Frontiers in Marine Science 2022, 9, 880550. [Google Scholar]
- Navarro-Mayoral, S.; Gouillieux, B.; Fernandez-Gonzalez, V.; Tuya, F.; Lecoquierre, N.; Bramanti, L.; Terrana, L.; Espino, F.; Flot, J.F.; Haroun, R.; et al. “Hidden” biodiversity: A new amphipod genus dominates epifauna in association with a mesophotic black coral forest. Coral Reefs 2024, 43, 655–672. [Google Scholar] [CrossRef]
- Wagner, D.; Luck, D.G.; Toonen, R.J. The biology and ecology of black corals (Cnidaria: Anthozoa: Hexacorallia: Antipatharia). Advances in Marine Biology 2012, 63, 67–132. [Google Scholar] [PubMed]
- Bellan-Santini, D.; Karaman, G.; Krapp-Schickel, G.; Ledoyer, M.; Myers, A.; Ruffo, S.; Schiecke, U. The Amphipoda of the Mediterranean. Part 1: Gammaridae (Acanthonotozomatidae to Gammaridae) 1982.
- Bellan-Santini, D.; Diviacco, G.; Krapp-Schickel, G.; Ruffo, S. The Amphipoda of the Mediterranean. Part 2. Gammaridea (Haustoriidae to Lysianassidae) 1989.
- Bellan-Santini, D.; Karaman, G.; Krapp-Schickel, G.; Ledoyer, M.; Ruffo, S. The amphipoda of the mediterranean. Part 3: Gammaridea (melphidippidae to talitridae), ingolfiellidea, caprellidea; 1993. [Google Scholar]
- Wilhelm, F.M.; Lasenby, D.C. Seasonal trends in the head capsule length and body length/weight relationships of two amphipod species. Crustaceana 1998, 399–410. [Google Scholar] [CrossRef]
- Lovich, J.E.; Gibbons, J.W. A review of techniques for quantifying sexual size dimorphism. Growth, Development, and Aging: GDA 1992, 56, 269–281. [Google Scholar]
- R Core Team. R: A language and environment for statistical computing; R Foundation for Statistical Computing; 2024.
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Maechler, M.; Bolker, B.M. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. The R Journal 2017, 9, 378–400. [Google Scholar] [CrossRef]
- Hartig, F. DHARMa: Residual diagnostics for hierarchical (multi-level / mixed) regression models, 2024. R package version 0.4.7.
- Lenth, R. emmeans: Estimated marginal means, aka least-squares means, 2025. R package version 1.11.1.
- Wickham, H. ggplot2: Elegant graphics for data analysis; Springer-Verlag, 2016. [Google Scholar]
- Conlan, K.E. Precopulatory mating behavior and sexual dimorphism in the amphipod Crustacea. Hydrobiologia 1991, 223, 255–282. [Google Scholar] [CrossRef]
- Borowsky, B.; Borowsky, R. The reproductive behaviors of the amphipod crustacean Gammarus palustris (Bousfield) and some insights into the nature of their stimuli. Journal of Experimental Marine Biology and Ecology 1987, 107, 131–144. [Google Scholar] [CrossRef]
- Bazikalova, A.Y. Taxonomy, ecology, and distribution of genera Micruropus Stebbing and Pseudomicruropus nov. gen. (Amphipoda; Gammaridea). Tr. Limnol. Inst., Akad. Nauk SSSR, Sib. Otd 1962, 2, 3–140. [Google Scholar]
- Cabrito, A.; de Juan, S.; Hinz, H.; Maynou, F. Morphological insights into the three-dimensional complexity of rhodolith beds. Marine Biology 2024, 171, 127. [Google Scholar] [CrossRef]
- Lippert, H.; Iken, K.; Rachor, E.; Wiencke, C. Macrofauna associated with macroalgae in the Kongsfjord (Spitsbergen). Polar Biology 2001, 24, 512–522. [Google Scholar] [CrossRef]
- Hughes, A.R.; Stachowicz, J.J. Genetic diversity enhances the resistance of a seagrass ecosystem to disturbance. Proceedings of the National Academy of Sciences 2004, 101, 8998–9002. [Google Scholar] [CrossRef]
- Ferreira, A.P.; Jacobucci, G.B.; Leite, F.P.P. Title TBD. In press. 2025.
- Edgar, G.J.; Aoki, M. Resource limitation and fish predation: Their importance to mobile epifauna associated with Japanese Sargassum. Oecologia 1993, 95, 122–133. [Google Scholar] [CrossRef]
- Poore, A.G.; Campbell, A.H.; Coleman, R.A.; Edgar, G.J.; Jormalainen, V.; Reynolds, P.L.; Sotka, E.E.; Stachowicz, J.J.; Taylor, R.B.; Vanderklift, M.A.; et al. Global patterns in the impact of marine herbivores on benthic primary producers. Ecology letters 2012, 15, 912–922. [Google Scholar] [CrossRef]
- Leite, F.P.P.; Tanaka, M.O.; Gebara, R.S. Structural variation in the brown alga Sargassum cymosum and its effects on associated amphipod assemblages. Brazilian Journal of Biology 2007, 67, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Rivera, E.; Hay, M.E. The effects of diet mixing on consumer fitness: Macroalgae, epiphytes, and animal matter as food for marine amphipods. Oecologia 2000, 123, 252–264. [Google Scholar] [CrossRef] [PubMed]
- Guerra-García, J.M.; Sánchez, J.A.; Ros, M. Distributional and ecological patterns of caprellids (Crustacea: Amphipoda) associated with the seaweed Stypocaulon scoparium in the Iberian Peninsula. Marine Biodiversity Records 2009, 2, e151. [Google Scholar] [CrossRef]
- Caine, E.A. Reproductive behavior and sexual dimorphism of a caprellid amphipod. Journal of Crustacean Biology 1991, 11, 56–63. [Google Scholar] [CrossRef]
- Emlen, S.T.; Oring, L.W. Ecology, sexual selection, and the evolution of mating systems. Science 1977, 197, 215–223. [Google Scholar] [CrossRef]
- Lewbel, G.S. Sexual dimorphism and intraspecific aggression, and their relationship to sex ratios in Caprella gorgonia Laubitz & Lewbel (Crustacea: Amphipoda: Caprellidae). Journal of experimental marine Biology and Ecology 1978, 33, 133–151. [Google Scholar]
- Mattson, S.; Cedhagen, T. Aspects of the behaviour and ecology of Dyopedos monacanthus (Metzger) and D. porrectus Bate, with comparative notes on Dulichia tuberculata Boeck (Crustacea: Amphipoda: Podoceridae). Journal of Experimental Marine Biology and Ecology 1989, 127, 253–272. [Google Scholar] [CrossRef]
- Castejón-Silvo, I.; Jaume, D.; Terrados, J. Feeding preferences of amphipod crustaceans Ampithoe ramondi and Gammarella fucicola for Posidonia oceanica seeds and leaves. In CSIC-Instituto de Ciencias del Mar (ICM); 2019. [Google Scholar]
- Guerra-García, J.M.; Martínez-Pita, I.; Pita, M.L. Fatty acid composition of the Caprellidea (Crustacea: Amphipoda) from the Strait of Gibraltar. Scientia Marina 2004, 68, 501–510. [Google Scholar] [CrossRef]
- Michel, L.N.; Dauby, P.; Gobert, S.; Graeve, M.; Nyssen, F.; Thelen, N.; Lepoint, G. Dominant amphipods of Posidonia oceanica seagrass meadows display considerable trophic diversity. Marine Ecology 2015, 36, 969–981. [Google Scholar] [CrossRef]
- Czechowska, K.; Feldens, P.; Tuya, F.; Cosme de Esteban, M.; Espino, F.; Haroun, R.; Otero-Ferrer, F. Testing side-scan sonar and multibeam echosounder to study black coral gardens: A case study from Macaronesia. Remote Sensing 2020, 12, 3244. [Google Scholar] [CrossRef]
- Rossi, S.; Bramanti, L.; Gori, A.; Orejas, C. An overview of the animal forests of the world. In Marine animal forests: The ecology of benthic biodiversity hotspots; Rossi, S., Bramanti, L., Gori, A., Orejas, C., Eds.; Springer, 2017; pp. 1–28. [Google Scholar]
- Buhl-Mortensen, L.; Buhl-Mortensen, P.; Rungruangsak-Torrissen, K.; Schwach, V.; Hjort, J.; Jakobsen, T.; Toresen, R. Cold temperate coral habitats. Corals in a changing world 2018, 9. [Google Scholar]
- Janicke, T.; Morrow, E.H. Operational sex ratio predicts the opportunity and direction of sexual selection across animals. Ecology letters 2018, 21, 384–391. [Google Scholar] [CrossRef]
- Kokko, H.; Jennions, M.D. Parental investment, sexual selection and sex ratios. Journal of Evolutionary Biology 2008, 21, 919–948. [Google Scholar] [CrossRef]
- Tuya, F.; Png Gonzalez, L.; Riera, R.; Haroun, R.; Espino, F. Ecological structure and function differs between habitats dominated by seagrasses and green seaweeds. Marine Environmental Research 2014, 98, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Tuya, F.; Ribeiro-Leite, L.; Arto-Cuesta, N.; Coca, J.; Haroun, R.; Espino, F. Decadal changes in the structure of Cymodocea nodosa seagrass meadows: Natural vs. human influences. Estuarine, Coastal and Shelf Science 2014, 137, 41–49. [Google Scholar] [CrossRef]
- Martinossi-Allibert, I.; Rueffler, C.; Arnqvist, G.; Berger, D. The efficacy of good genes sexual selection under environmental change. Proceedings of the Royal Society B 2019, 286, 20182313. [Google Scholar] [CrossRef] [PubMed]
- Sainte-Marie, B. A review of the reproductive bionomics of aquatic gammaridean amphipods: Variation of life history traits with latitude, depth, salinity, and superfamily. Hydrobiologia 1991, 223, 189–227. [Google Scholar] [CrossRef]
- Leber, K.M. The influence of predatory decapods, refuge, and microhabitat selection on seagrass communities. Ecology 1985, 66, 1951–1964. [Google Scholar] [CrossRef]
- Moksnes, P.O.; Gullström, M.; Tryman, K.; Baden, S. Trophic cascades in a temperate seagrass community. Oikos 2008, 117, 763–777. [Google Scholar] [CrossRef]
Figure 1.
(A) The Canary Islands Archipelago (28.292° N, 16.629° W) lies in the Atlantic Ocean off northwestern Africa. (B) Gran Canaria (27.9202° N, 15.5474° W) is located approximately 150 km from the Moroccan coast; sampling was focused on the eastern shore (grey inset). (C) Map of sampling sites across four benthic habitats - black coral, macroalgae-dominated reefs, rhodolith beds, and seagrass meadows, surveyed at five distinct locations along the eastern coast of Gran Canaria, viz. Jinámar, Tufia, Gando, Risco Verde, and Arinaga (from north to south). (D) Representative habitat images: (i) black coral forests, (ii) macroalgae-dominated reefs, (iii) seagrass meadows, and (iv) rhodolith beds, photographed at study sites around Gran Canaria. (@photo credit: Franck Gazzola and Fernando Espino
Figure 1.
(A) The Canary Islands Archipelago (28.292° N, 16.629° W) lies in the Atlantic Ocean off northwestern Africa. (B) Gran Canaria (27.9202° N, 15.5474° W) is located approximately 150 km from the Moroccan coast; sampling was focused on the eastern shore (grey inset). (C) Map of sampling sites across four benthic habitats - black coral, macroalgae-dominated reefs, rhodolith beds, and seagrass meadows, surveyed at five distinct locations along the eastern coast of Gran Canaria, viz. Jinámar, Tufia, Gando, Risco Verde, and Arinaga (from north to south). (D) Representative habitat images: (i) black coral forests, (ii) macroalgae-dominated reefs, (iii) seagrass meadows, and (iv) rhodolith beds, photographed at study sites around Gran Canaria. (@photo credit: Franck Gazzola and Fernando Espino
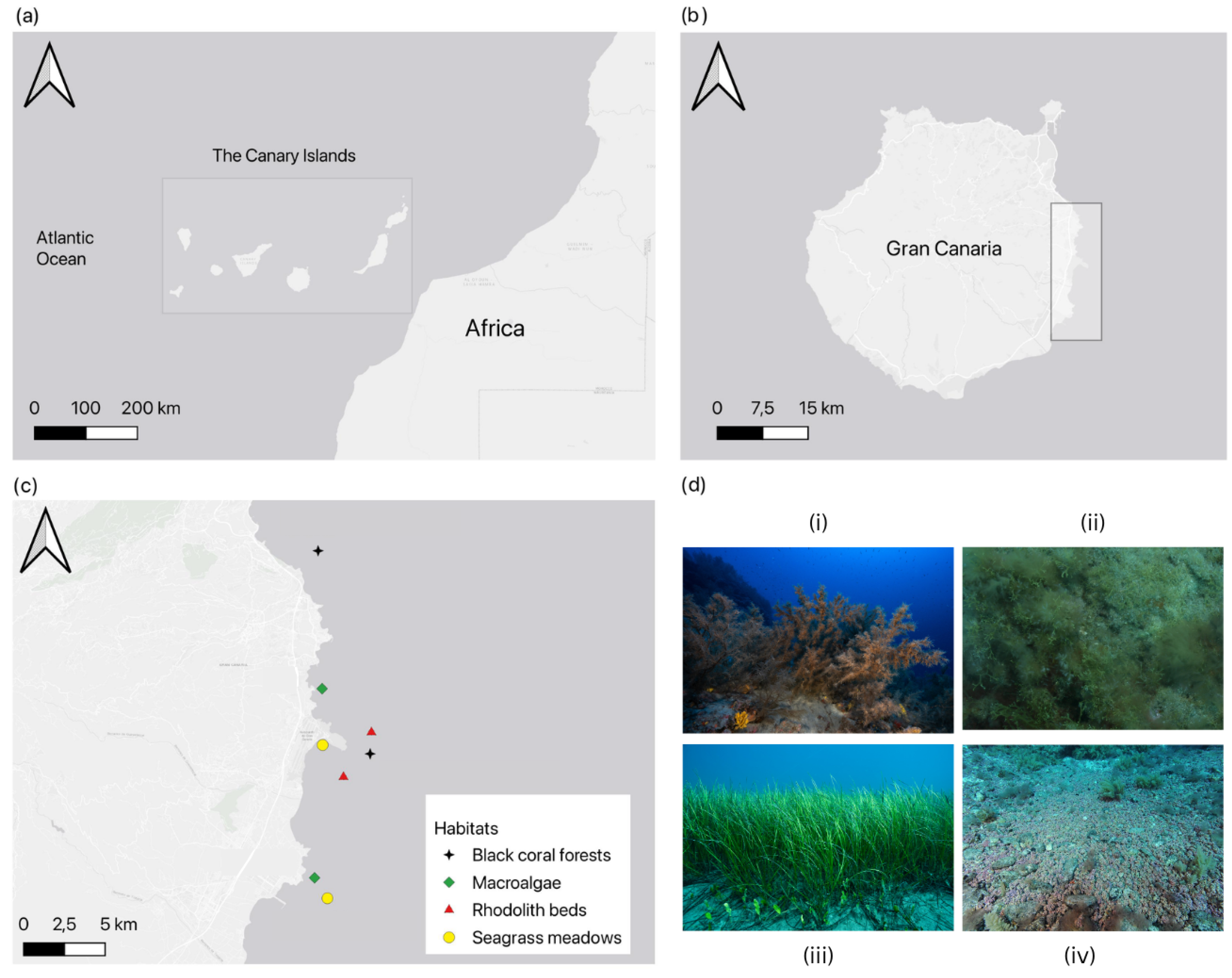
Figure 2.
Images for body length (panel a) and gnathopod 2 length (panel b) measurements in a male Ampithoe ramondi individual. Representative images for other species are provided in Figure 1 of Supplementary Information.
Figure 2.
Images for body length (panel a) and gnathopod 2 length (panel b) measurements in a male Ampithoe ramondi individual. Representative images for other species are provided in Figure 1 of Supplementary Information.
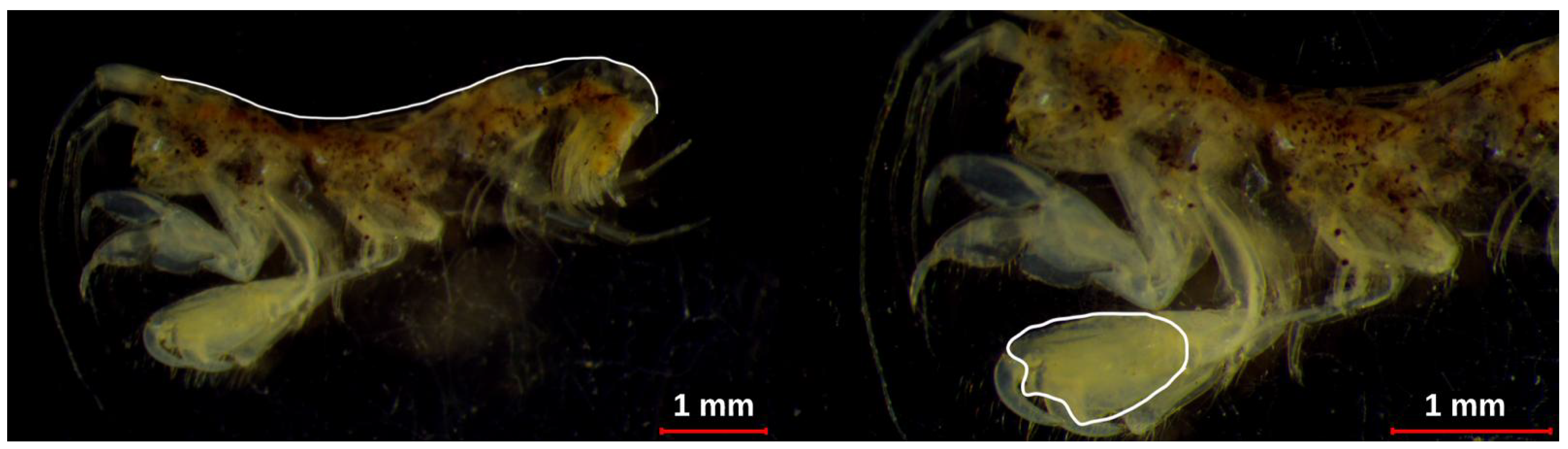
Figure 3.
Sex-specific allometry of gnathopod 2 length in Ampithoe ramondi and Caprella acanthifera. Points represent individual measurements, and shaded areas around regression lines indicate 95% confidence intervals.
Figure 3.
Sex-specific allometry of gnathopod 2 length in Ampithoe ramondi and Caprella acanthifera. Points represent individual measurements, and shaded areas around regression lines indicate 95% confidence intervals.
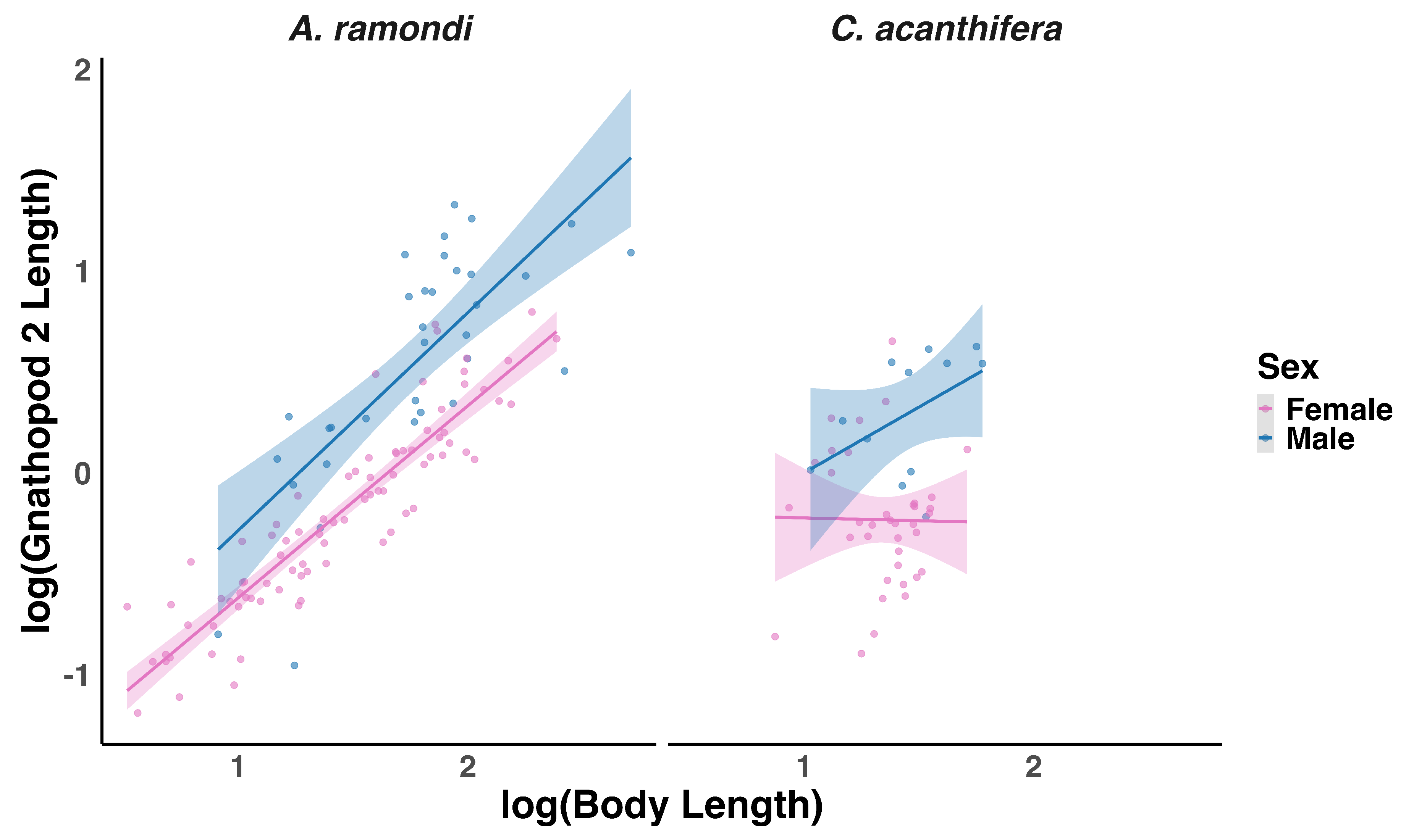
Figure 4.
Box plot showing body length differences between females (pink) and males (blue) of Ampithoe ramondi across rhodolith beds, macroalgae-dominated reefs, and seagrass meadows. Sample sizes (F = female, M = male) for each habitat are as follows: rhodolith beds — F = 10, M = 12; macroalgae-dominated reefs — F = 45, M = 9; seagrass meadows — F = 29, M = 12.
Figure 4.
Box plot showing body length differences between females (pink) and males (blue) of Ampithoe ramondi across rhodolith beds, macroalgae-dominated reefs, and seagrass meadows. Sample sizes (F = female, M = male) for each habitat are as follows: rhodolith beds — F = 10, M = 12; macroalgae-dominated reefs — F = 45, M = 9; seagrass meadows — F = 29, M = 12.
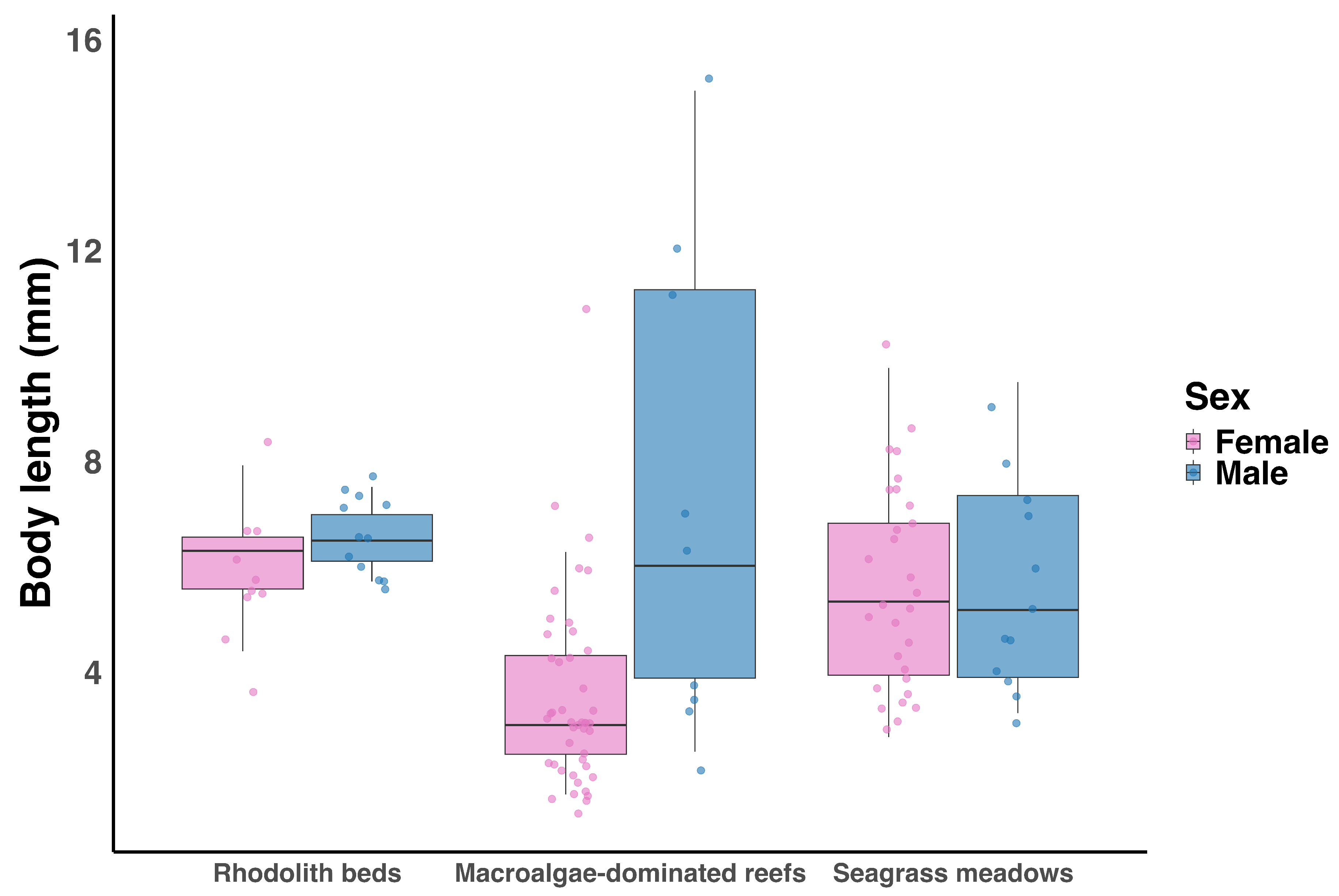
Figure 5.
Box plot showing gnathopod ratio differences between females (pink) and males (blue) of Ampithoe ramondi in rhodolith beds, macroalgae-dominated reefs and seagrass meadows. Sample sizes for each species and sex are provided in Figure 4).
Figure 5.
Box plot showing gnathopod ratio differences between females (pink) and males (blue) of Ampithoe ramondi in rhodolith beds, macroalgae-dominated reefs and seagrass meadows. Sample sizes for each species and sex are provided in Figure 4).
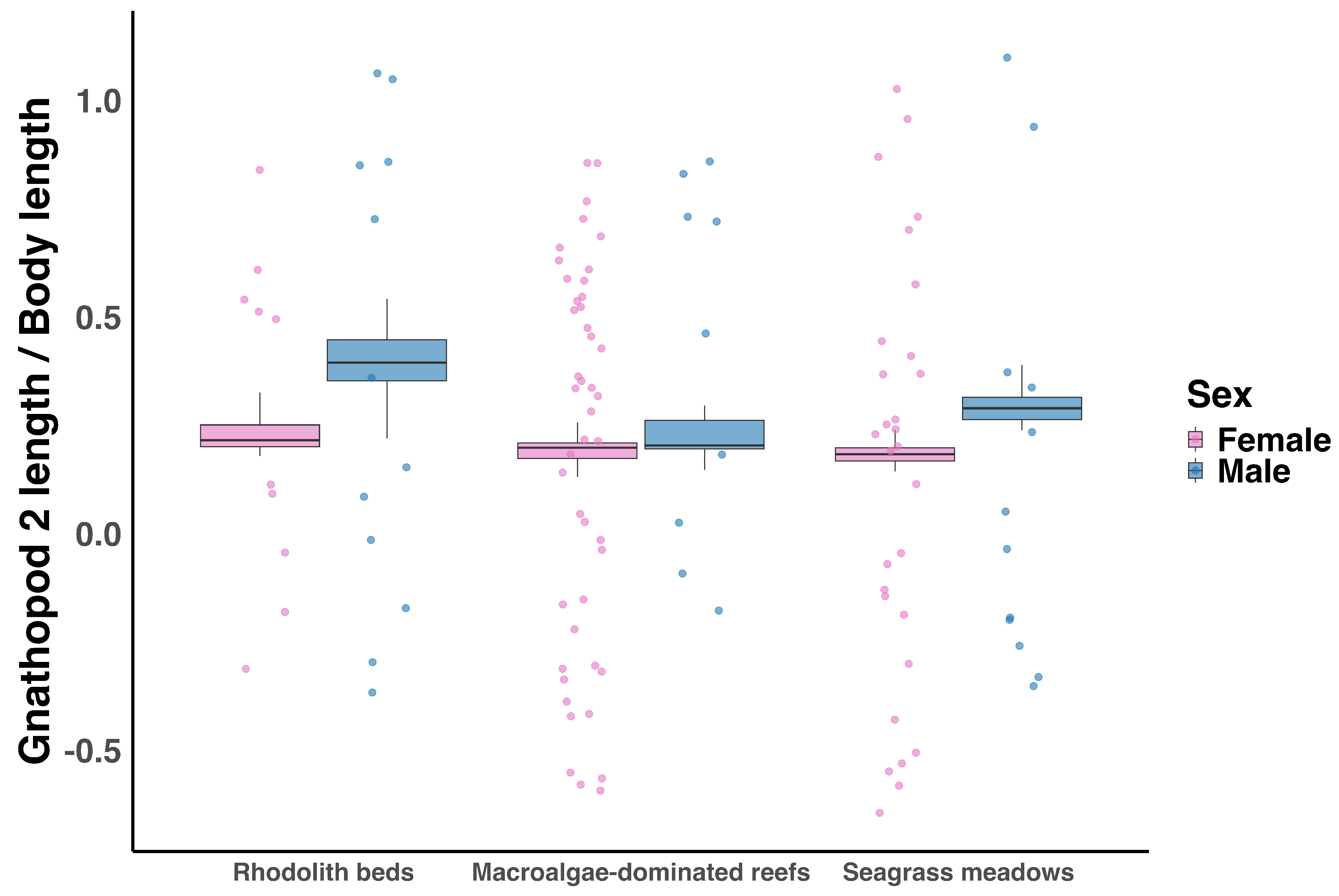
Figure 6.
Box plot showing body length differences between females (pink) and males (blue) of Caprella acanthifera in macroalgae-dominated reefs and black coral forests. Sample sizes for each habitat are as follows (F = female, M = male): Macroalgae-dominated reefs — F = 28, M = 6; Black coral forest — F = 9, M = 6.
Figure 6.
Box plot showing body length differences between females (pink) and males (blue) of Caprella acanthifera in macroalgae-dominated reefs and black coral forests. Sample sizes for each habitat are as follows (F = female, M = male): Macroalgae-dominated reefs — F = 28, M = 6; Black coral forest — F = 9, M = 6.
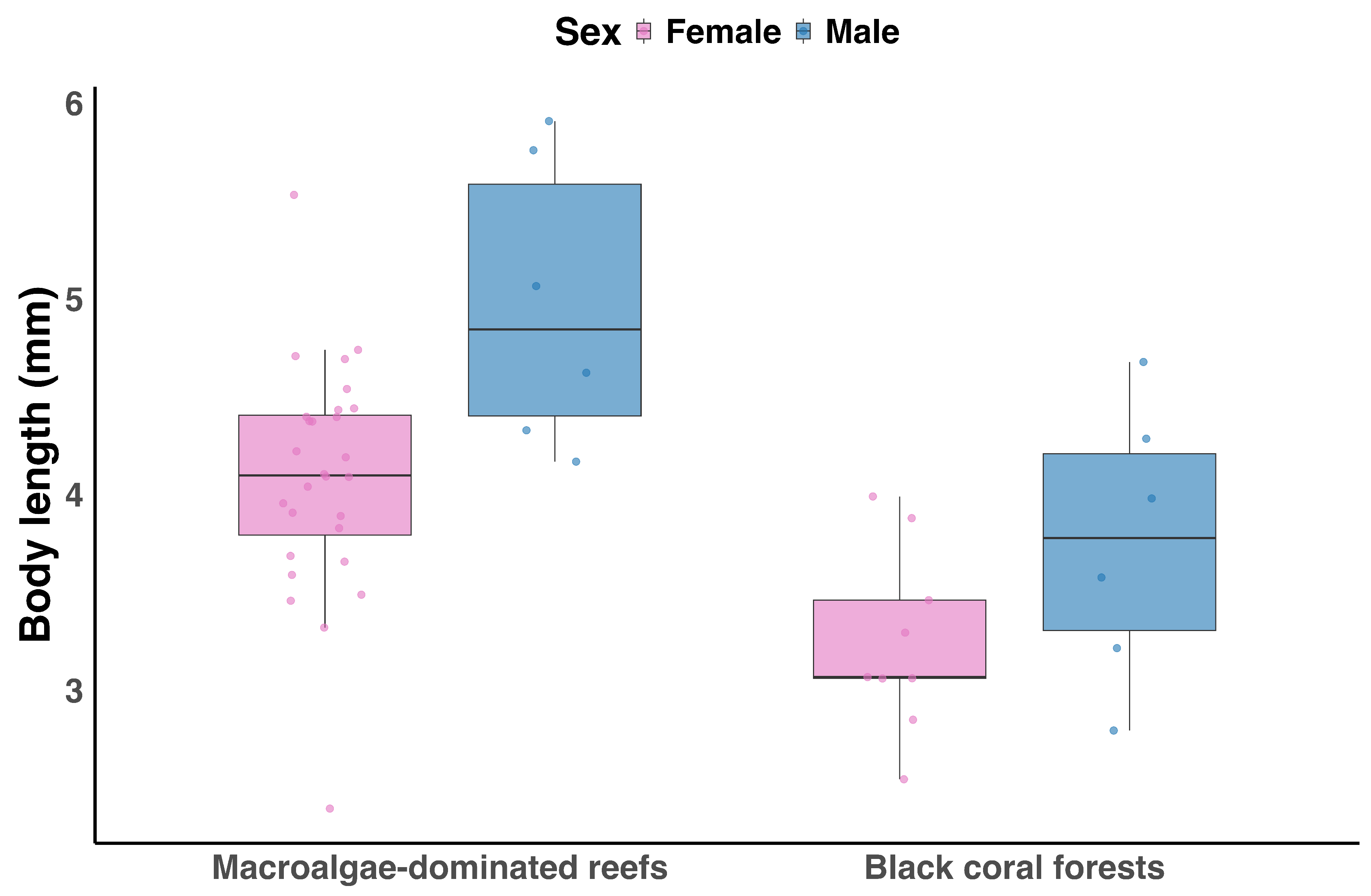
Figure 7.
Box plot showing gnathopod ratio differences between females (pink) and males (blue) of Caprella acanthifera in macroalgae-dominated reefs and black coral forests. Sample sizes for each species and sex are provided in Figure 6).
Figure 7.
Box plot showing gnathopod ratio differences between females (pink) and males (blue) of Caprella acanthifera in macroalgae-dominated reefs and black coral forests. Sample sizes for each species and sex are provided in Figure 6).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



