
Submitted:
18 July 2025
Posted:
21 July 2025
You are already at the latest version
Abstract
Deep-water environments constitute the largest ecosystem on Earth and are insufficiently characterized. Lake Baikal, the only freshwater reservoir possessing rich deep-water fauna, provides unique opportunities to investigate the evolutionary processes that happened to such fauna independently and in parallel to adaptation to these environments in the ocean. Here, we focus on amphipods as one of the dominant elements of Baikal deep-water fauna and study the genetic diversity across a wide vertical (~1 km) and horizontal (~500 km) ranges, as well as depth-related traits in two deep-water scavengers Ommatogammarus flavus (Dybowsky, 1874) and Ommatogammarus albinus (Dybowsky, 1874). Our data revealed low intraspecific diversity of the cytochrome c oxidase subunit I gene marker fragment, at both vertical and horizontal scales, indicating no significant distribution barriers and a bottleneck event in the evolutionary history of both species. At the same time, we found depth-related stratification in carotenoid-based body coloration and eye shape in O. flavus and eye color over stable opsin expression in both species. These findings might indicate partial isolation between vertically stratified populations and characterize the ecological distinctions between the two studied species.
Keywords:
ancient lakes
; endemic speciation
; necrophagous amphipods
; genetic bottleneck
; carotenoid accumulation
; compound eyes
1. Introduction
Deep-water environments may seem almost empty and hostile, but yet they play many important roles for humanity [1]. Deep sea (commonly defined as depths below 200 m, i.e. beyond the continental shelf depth) is the largest ecosystem on Earth, covering around 65% of the planet’s surface and ~95% of the ocean volume [1,2,3]. Deep-sea habitats have many unique features connected to extreme values of ecological factors, such as temperatures below 4°C, which slow down metabolic processes; near total darkness, which severely restricts photosynthesis-based primary production; and high hydrostatic pressure, which also requires unique adaptations [4]. Despite some significant breakthroughs in studying these adaptations, mostly made with the help of modern sequencing techniques, many aspects of deep-water adaptation and related evolutionary changes remain to be explained [5].
Ancient Lake Baikal provides unique and, sometimes, more convenient opportunities to study the evolution of high depths-shaped organisms that happened in parallel to the ocean. Most deep freshwater lakes are anoxic below 250 m [6], while Baikal (maximum depth of 1642 m) is characterized by high oxygen content (up to 80% saturation) along the entire water column, which allowed for the independent emergence of the deep-water fauna [7,8]. Many of the deep-sea challenges apply to the inhabitants of deep-freshwater zone in this enormous lake: low temperatures (stable at 3.5–4.5°C below 300 m [9]); low-light conditions (even with exceptional transparency of Baikal water, the depth of the photic zone does not exceed 75 m; [10]); and relatively high hydrostatic pressure (up to 165 atmospheres). Thus, comparative studies of deep-water animals in the ocean and Lake Baikal pose a significant interest in terms of understanding the general evolutionary mechanisms of adaptation to deep-water environments.
While some taxa are exclusive to either saltwater or freshwater ecosystems and cannot thus be used for such comparative studies, some have followed remarkably similar evolutionary paths and underwent fast adaptive radiations. A primary example of such a group would be amphipods (Crustacea: Malacostraca: Amphipoda). This crustacean order lists over 10,000 species [11,12], which are known to form closely related groups of species, or so-called species flocks, in different seas and lakes [13,14,15,16,17]. Amphipods are the dominant animal taxon within the deepest ocean (hadal) fauna [18]. Similarly, amphipods represent a significant part of the deep-water fauna of Lake Baikal, with over 80 species and subspecies out of over 350 known encountered at depths below 500 m and over 55 at depths over 1000 m [19,20].
The unusual conditions for evolution in deep-water environments are the apparent lack of barriers for dispersal on the one hand [21] and the additional dimension for potential vertical stratification on the other. A general tendency has been proposed that the horizontal diversity is much less pronounced than the vertical one [2], and yet during the last 20 years not so much data have been accumulated on the genetic diversity of deep-sea amphipods across bathymetric (vertical) distances in comparison to geographic (horizontal) ones. The patterns of diversity have turned out to be varying for different species, frequently even within the same genus.
Some species indeed seem to lack dispersal barriers. For example, individuals of Abyssorchomene distinctus (Birstein & Vinogradov, 1960) sampled as far as the Pacific and Indian Ocean shared the same set of haplotypes [22]. Similarly, Paralicella tenuipes Chevreux, 1908 sampled from three oceans comprised only one genetic group [23]. Hirondellea dubia Dahl, 1959 sampled from three oceans at depths of 4,000 to 11,000 m turned out to be indistinguishable by mitochondrial markers [24].
At the same time, many morphological species turned out to be cryptic species complexes. Perhaps the most well-studied example is a giant bentho-pelagic scavenger Eurythenes gryllus (Lichtenstein, 1822), which is found in every world ocean and in a huge range of depths, from 550 to 7,800 m [25]. The species within the E. gryllus complex did not have strict geographic division, but almost all of them were distributed either above or below 3,000 m (the abyssal/hadal border), which might still be a sampling artifact [26]. Paralicella caperesca Shulenberger and Barnard, 1976 was also found to be a complex of potential cryptic species with overlapping ranges, the last common ancestor of which existed around 8 million years ago [23]. A cosmopolitan species Bathycallisoma schellenbergi (Birstein & M. Vinogradov, 1958) was revealed to comprise a number of genetic lineages correlated with their geography [27]. Finally, most hadal amphipods have been described as endemics of particular hadal features, and their genetic structure has not been revealed yet [18].
In this work, we aimed to shed the first light on the comparative roles of horizontal and vertical diversification of deep-water amphipod fauna in Lake Baikal. Over 20 species of Baikal amphipods have been previously studied in terms of genetic diversity, but most of the well-studied species unsurprisingly inhabit shallow depths [15]. Some widely distributed shallow-water morphological species comprise divergent genetic lineages, some of which have documented reproductive barriers [28,29,30,31]. At the same time, at least one species presents the same haplotypes for samples collected at several hundred kilometers [31]. Regarding deep-water species, there are only preliminary reports on the single pelagic Baikal amphipod species Macrohectopus branickii (Dybowsky, 1874), for which no geographically separated genetic lineages were revealed [32,33].
Here, we focused on two most frequently observed deep-water scavengers of the genus Ommatogammarus Stebbing, 1899, namely Ommatogammarus flavus (Dybowsky, 1874) = Abludogammarus flavus sensu Kamaltynov, 2009 and Ommatogammarus albinus (Dybowsky, 1874). They differ from other species of the genus Ommatogammarus sensu Takhteev, 2000, namely from three subspecies of Ommatogammarus carneolus [34], by high forehead [35] or by antenna I attached almost in the middle of the head in lateral view, as described by Karaman [36]. These two species have a number of morphological differences (File S1; Figure S1; Figure 1). The most obvious ones that can also be checked in live animals without dissection are (1) the characteristic shape of the eyes, which are irregularly triangular in O. flavus and with deeply serrated hind margin in O. albinus, and (2) the length of peduncles of antennae I, which are longer than the head segment in O. flavus but shorter or equal in length to the head segment in O. albinus (Figure 1). Together with O. carneolus melanophthalmus sensu Takhteev, 2000 (= Pretiositus melanophthalmus sensu Kamaltynov, 2009), these two species form stable phylogenetic group with genetic distances that are much smaller than within the genus Eulimnogammarus based on the fragments of COI, 16S rRNA and 18S rRNA genes [37,38]. For the purposes of this manuscript, we will name the species O. flavus and O. albinus, as they are named in NCBI taxonomy [39], txid75829 and txid315607, but not A. flavus and O. albinus, as in the World Amphipoda Database / WORMS [40], AphiaID 746011 and 746012) and BOLD [41], BOLD:AAX0702 and BOLD:AGK6709).
O. flavus and O. albinus are endemic to the lake and widely distributed both horizontally and vertically, being observed in all three basins of the lake at the depth ranges of 2.5-1300 m in the case of O. flavus and 50–1642 m in the case of O. albinus [19,20,34]. These species are frequently found together in baited traps. Even though the depth ranges of these species largely intersect, they tend to prefer different depths: O. flavus was most abundant at 100–300 m, while O. albinus was most frequent in the traps at 300–500 m [43]. Thus, relatively shallow-water borders between basins of Lake Baikal may be more important for geographic separation of O. albinus than O. flavus. Taking into account the wide depth ranges inhabited by both species, hydrostatic pressure and decrease in solar irradiation are the factors that potentially may drive their adaptive vertical separation into less interbreeding populations. However, since the laboratory exposures of these species showed ultraviolet to cause much more pronounced mortality than pressure decrease [43,44], here we hypothesized that the separation may be related to adaptations to different illumination conditions.
In this work, we explored the genetic diversity of O. flavus and O. albinus across the three basins of Lake Baikal, as well as their adaptations along the depth gradient at one sampling location. Apart from horizontal geographic diversity (>500-km distance between the furthest points), we sampled a range of depths at a small distance (1-km depth range at 2-km horizontal distance) to study vertical diversity, even though to a smaller scale than in the deep ocean. During the latter sampling we encountered phenotypic differences in body and eye color between individuals of O. flavus from different depths and explored this phenomenon further at the level of carotenoid concentration and opsin expression.
2. Materials and Methods
2.1. Sampling, Study Sites and Fixation
O. flavus and O. albinus were collected at 9 locations covering the southern, central and northern basins of Lake Baikal at different depths (ranging from 25 m to 1000 m) in 2020, 2023–2025 (Figure 2 and Table S1). Amphipods were captured from the lake ice in spring or from a boat in summer using bottom deep-water traps with rotten fish (for more dеtalis see [43]). Neither studied species is endangered or protected. All procedures were approved by the Animal Subjects Research Committee of the Institute of Biology at Irkutsk State University (protocol #9/2022).
Amphipods of the studied species were determined morphologically according to standard keys [34,35,45]. Amphipods from Bolshie Koty were fixed in liquid nitrogen immediately after capture. Amphipods from other locations were fixed in ethanol. From each location we sequenced from 2 to 27 animals of each species (Table S1). In total, COI sequences of 66 individuals of O. flavus and 57 of O. albinus were obtained.
Each animal from Bolshie Koty, which was frozen in liquid nitrogen, was unfrozen and quickly photographed on grey background with millimeter paper with an Olympus Tough TG-5 camera (Olympus, China). To assess eye morphology, photographs were taken under a SPM0880 stereomicroscope (Altami, Russia). Next, the head with eyes was dissected using scissors and scalpel, then immediately placed in Trizol for RNA isolation. Pereiopods were separated for genotyping. The rest of the body was used for carotenoid measurements. Animal body length and eye area were estimated in the ImageJ program [46], with body length measured from rostrum to telson. Quantitative analysis of pereon color was performed by normalizing white balance using grey background and identifying the RGB color of the random spot of the 6th or nearest visible segment of the pereon. The ratio of intensities of the red and green channels was used as the color index. The method was described in more detail earlier [47].
2.2. Nucleic Acid Extraction, PCR and Sequencing
DNA was isolated from limbs with a commercial kit “S-sorb” (Syntol, Russia) with modifications described earlier [48]. Total RNA was isolated from the heads with eyes of O. flavus or O. albinus individuals by homogenizing frozen tissues in TriReagent (MRC, Germany) according to the manufacturer's protocol using two 3-mm stainless steel beads (Qiagen, Germany) in a TissueLyser LT instrument (Qiagen, Germany). The methodology for isolation of total RNA is described in detail in [49]. The homogenate was centrifuged to precipitate steel beads and tissue debris, transferred to a new tube and mixed with chloroform according to the manufacturer's protocol, followed by centrifugation and phase separation. The upper phase was carefully removed without touching the interphase and then followed the protocol of the RNeasy Mini kit (Qiagen, Germany). RNA integrity was checked by agarose gel electrophoresis, and the concentration was measured on a Nano-300 spectrophotometer (ALLSHENG, China). The resulting RNA was purified from residual genomic DNA using RapidOut DNA Removal Kit (Thermo Scientific, Lithuania) following the manufacturer's instructions. To make sure that the sequences that were obtained in different ways (with cDNA or genomic DNA) match, we additionally isolated DNA from the limbs of several animals of both species (from Bolshie Koty). The sequences matched.
PCR of the cytochrome c oxidase subunit I (COI) gene fragment for O. flavus DNA samples was performed with primers 398_Eve_F4 (TAAACTATAAGCCTTCCAAGC) and 399_Eve_R4 (TGTGAAGTAAGCTCGGGTAT) [31]. If we took O. flavus cDNA (only samples from Bolshie Koty), the primers COI_Ofla_MD_F7 (GTGACTATTTTCTACTAACCA) and COI_Ofla_MD_R7 (AGCCTAGAAAACCAATAGCCAGT) (this study) were used. Primers were designed in the SnapGene Viewer (Insightful Science; available at snapgene.com), using mitochondrial sequences of the corresponding species from transcriptomes assemblies. COI sequences were retrieved from the transcriptomes [50] reassembled earlier [51]. The transcripts used were TRINITY_DN16_c2_g1_i2 for O. flavus and TRINITY_DN2055_c0_g1_i3 for O. albinus. These sequences were found in the assemblies with exonerate version 2.4.0 [52] using partial sequences published earlier [38] as queries. The sequences with matching primers are available in the Supplementary (Files S2 and S3).
PCR of COI fragment for DNA and cDNA (only samples from Bolshie Koty) of O. albinus were performed with primers 0303_LCO_Eve_F1 (TCTCTACTAATCATAAAGATATCGG) and 399_Eve_R4 (TGTGAAGTAAGCTCGGGTAT) [31]. The size of the COI product for both species was about 950 bp. In some cases when concentration of the obtained PCR product was not sufficient for sequencing, re-amplification with different primers was used. See Table S2 for the list of primers and Table S3 for more information on primers used for each sample. PCR of the long wavelength-sensitive (LWS) opsins gene region for both species were performed with primers LWS_4F (GCGGAACTGGAACTGACTACCTCA) and LWS_7R (CACGATGGGGGGTTGTAGAC), which amplify the part of the open reading frame corresponding to amino acids 204-332 [51]. The product length for both species was slightly more than 350 bp.
Amplification of COI and LWS fragments was performed with 2.5x qPCR mix (Syntol, Russia) in the volume of 30–40 μl. Cycling conditions for COI fragments of both species and all primer pairs were as follows: at 95 °C for 5 min; 30 cycles of (95 °C for 30 s; 56 °C for 1 min; 72 °C for 1 min); and 72 °C for 5 min. The conditions for LWS amplification are described in detail by [51]. PCR products were visualized in a 1% agarose gel in TAE buffer. PCR products were purified with Cleanup Standard or Cleanup Mini kits (Evrogen, Russia). The quality and concentration of purified PCR products were assessed using a Nano-300 spectrophotometer (ALLSHENG, China). PCR products were sequenced according to the Sanger method with a Nanophor 05 sequencer (Institute for Analytical Instrumentation RAS, Russia) in both directions (for combination of primers used for each sample, see Table S3). The BigDye Terminator v3.1 Cycle Sequencing kit (Life Technologies, USA) was used for the amplification reaction; see [48] for details. For the reverse sequencing reaction, primer COI_HCO2198 (TAAACTTCAGGGTGACCAAAAAATCA) [53] was used for samples from Bolshie Koty.
Quantitative PCR (qPCR) was performed with samples collected in Bolshie Koty. Complementary DNA (cDNA) synthesis was performed using Thermo Scientific RevertAid RT Kit RevertAid FirstStrand cDNA synthesis kit (Thermo Scientific, Lithuania) with random primers according to the manufacturers' recommendations. Up to 5% of the cDNA volume was used for qPCR. Amplification was performed on a StepOnePlus Applied Biosystems (USA) using 5X qPCRmix-HS SYBR (Evrogen, Russia). The same amount of RNA sample, which was treated similarly but without the addition of reverse transcriptase (-RT), was used to test for genomic DNA contamination. Expression levels of the lws genes were estimated using the housekeeping glyceraldehyde-3-phosphate dehydrogenase (gapdh) gene as a reference. The following primer sequences were used for the qPCR. Primers LWS_4F (GCGGAACTGACTGACTACCTCA) and LWS_4R (GACTCCCATCTTCTTTCTTGGC) flank a 200-bp region of the gene [51]. Expression of the reference gene gapdh was quantified with the primers F-ACTCTACTCACGGCGTCTTCAAG and R-CGCTGGACTCTACGATGTACTCAG (KF293381) (Protopopova et al, 2014). Experimental samples (cDNA) with quantification cycle values over 30 cycles or -RT samples with less than 30 cycles for gapdh were considered below detection or contaminated, respectively, and were therefore discarded from further analysis. Only one sample failed to isolate a high-quality RNA (O. flavus from 25 m #5).
2.3. Extraction and Estimation of Carotenoid Concentration
Total carotenoid content was measured according to the method described in detail earlier [47]. Carotenoid concentration was estimated spectrophotometrically at wavelengths from 200 to 800 nm on a Cary 50 UV/VIS spectrophotometer (Varian Inc., Australia). The purity of the extract was monitored by absorbance at 600 nm, and the concentration of carotenoids was determined in parts per million (ppm) by absorbance at 450 nm (A450) as 4 × A450 × V / M, where V is the volume of petroleum ether used for re-extraction (mL) and M is the crude weight of the sample (g).
2.4. Data Analysis
The raw Sanger sequencing data were processed and basecalled using Mutation Surveyor V 5.1 [54]. Consensus sequences were obtained in the UGENE V 41.0 program [55] by aligning to the reference sequence of the corresponding species retrieved from transcriptome assemblies (see above). Reference LWS opsin sequences were also retrieved from transcriptomes and published earlier [51]. All sequences obtained in this work were added to the NCBI GenBank (https://www.ncbi.nlm.nih.gov/nucleotide/) database with the following accession numbers: PV474290–PV474412 for COI and PV491384-PV491410 for LWS opsins and the Barcode of Life Datasystems (BOLD) [41] database as the OMM project. The sequences were aligned and trimmed to the shortest sequence in the UGENE program [55] using the MUSCLE algorithm [56]. The haplotype network was constructed in R [57] (R Core Team, 2024) with the pegas package (Population and Evolutionary Genetics Analysis System) [58], using the maximum parsimony method. The presence of a gap between intraspecific and interspecific variability (barcode gap) was estimated using the ABGD (Automatic Barcode Gap Discovery) and ASAP (Assemble Species by Automatic Partitioning) algorithms in the SpartExplorer web interface (https://spartexplorer.mnhn.fr/; [59,60,61].
To construct the bathymetric map of Lake Baikal we used the GEBCO project data [62], as well as the marmap v1.0.10 [63] rnaturalearth v1.0.1 [64] packages for R. The qPCR data were presented as relative quantitative cycles (ΔCt) obtained by subtracting the quantitative cycle value for the reference gene from the corresponding value of the target gene. Four to seven biological repeats were measured at each point (depth of capture). Each biological repeat was measured in three technical repeats. The values obtained for lws gene expression and carotenoid from each depth were compared using the Wilcoxon-Mann-Whitney criterion [65], with Holm's correction for multiple comparisons [66] implemented in the R environment. All data were analyzed and visualized with the R software network v4.1.2 using suitable packages: ape v5.8.1 [67]; ggplot2 v3.4.2 [68] for boxplots; pegas package v1.3 [58] for haplotype network build; openxlsx v4.2.5 (Schauberger & Walker, 2023); dplyr v1.1.4 [69]; ggrepel v0.9.6 [70]. The graphs were finalized with Inkscape software V0.92.3 (https://inkscape.org/). All scripts are available on GitHub (https://github.com/Telnes/Population-structure-of-Ommatogammarus-flavus-and-O.-albinus).
3. Results
3.1. Intraspecific Genetic Diversity Is Low and Similar Across Locations and Depths
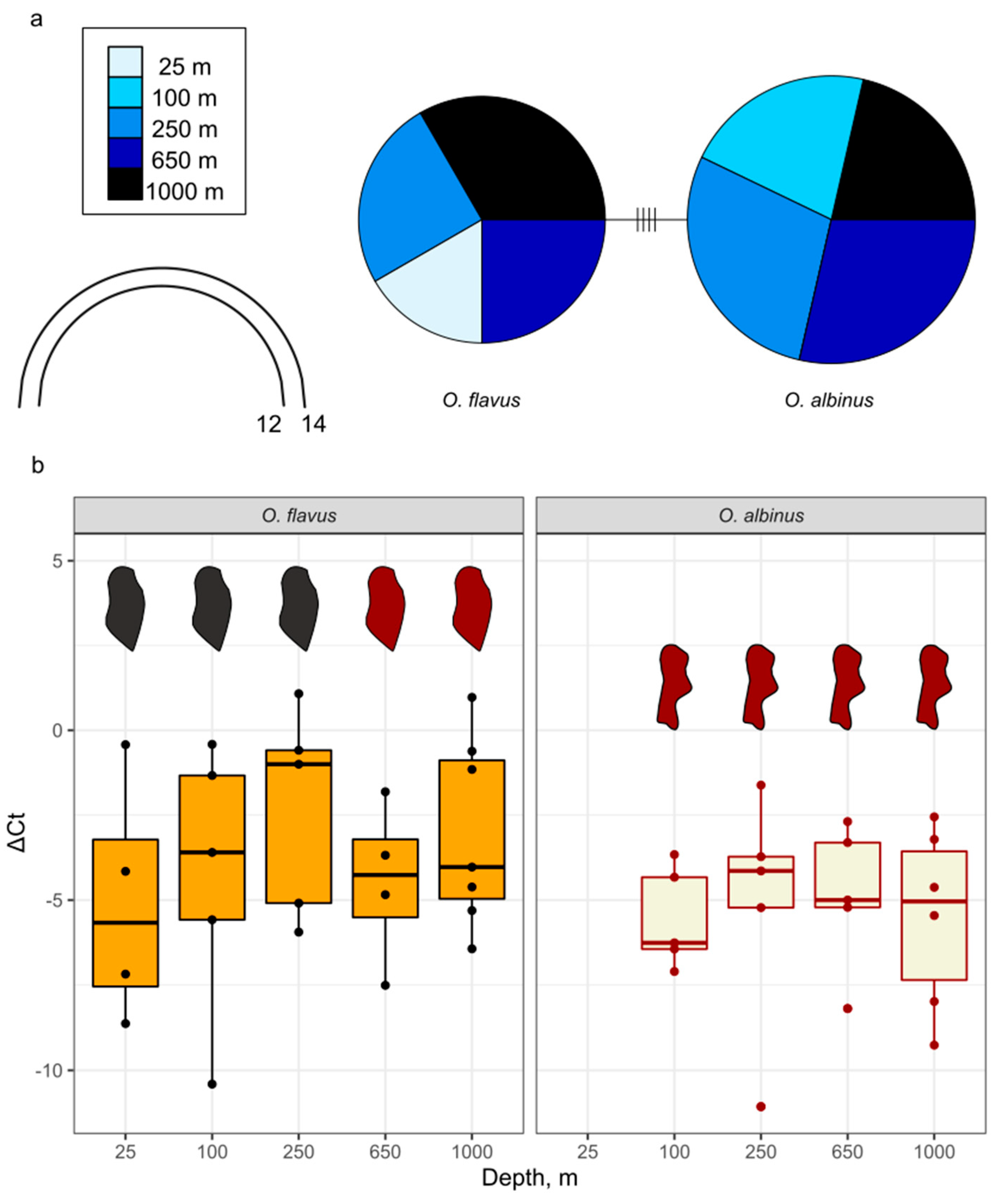
We started by exploring the genetic diversity of O. flavus and O. albinus from different depths in Bolshie Koty, near the Biological Station of the ISU. This station is located in the southwestern part of the lake, which is characterized by steep declining slopes ([71]; http://bic.iwlearn.org/en/atlas/atlas/127-angles-of-inclination-of-map)), thus providing us with a rare opportunity to sample at different depths (25 to 1000 m) at relatively short horizontal distances (around 2-km distance from the shore to 1000-m point). We used the Folmer COI fragment as the marker sequence, as this approach has been successfully applied to study genetic diversity in other Baikal amphipods before and other deep-water amphipods (see the Introduction section). Both studied species were characterized by low COI diversity with two (O. flavus) or one (O. albinus) prevailing haplotypes and several singleton haplotypes differing from the major ones by one substitution (Figure 3). The low diversity means that the morphologically united species at different depths did not form isolated populations, at least for evolutionary meaningful times. Even more importantly, this homogeneity allowed us to compare the horizontal diversity across the lake because it was not physically possible to sample the same depth everywhere.
Then, we went on to sample O. flavus and O. albinus from nine points in all three basins of the lake to analyze their horizontal diversity (Figure 2). Even though a few unique haplotypes were found, the overall COI genetic diversity was still low (two major and 16 singleton haplotypes for O. flavus, one major and five singleton haplotypes for O. albinus) and did not correlate with sampling geography (Figure 4 and File S4). Thus, either these species do not form isolated groups of populations in the basins or formed such groups too recently to be distinguishable by mitochondrial sequences. The delimitation of the two species was confirmed with both ABGD and ASAP methods as the most optimal number of subsets.
3.2. Analysis of Coloration and Eye Shape Reveals Depth-Related Phenotypes
However, notwithstanding the genetic homogeneity within each species across the depth range, we found that O. flavus differed in their visual appearance, most importantly and most clearly in coloration. While the animals sampled from 250 m and above had their characteristic orange coloration and black eyes, the ones sampled from 650 m and below were pale and had red eyes, resembling the description of O. albinus (Figure 5a–d; see also Figures S2–S4). However, the main distinguishing morphological character, the shape of the eye, was consistent with molecular species identification. The intensity of orange coloration in amphipods and other crustaceans is mainly determined by the concentration of carotenoids [72,73]. So, in order to quantitatively access the visible differences in color, we measured carotenoid contents in four adult animals from each sampling depth. Even though we could not detect statistically significant differences in pairwise comparisons corrected for multiple comparisons, the values obtained from O. flavus samples definitely fell into two groups, up to 250 m vs. 600 m and below (Figure 5e). Moreover, linear regression analysis confirmed the correlation between sampling depth and carotenoid content for O. flavus (Figure 5f). The specimens of O. albinus from different depths differed neither in their visible body color nor in measured carotenoid content.
In order to check the trend to a paler body coloration at larger depths in adult O. flavus, we quantified the color using R/B color index in a larger sample from each depth (12–21 animals). Indeed, there was a clear tendency towards a lower R/B index at lower depths (Figure 6a). Importantly, the trend of paler O. flavus coloration with greater depths was not absolute, as smaller animals at 1000 m had a brighter orange color than the large ones. This observation, combined with the fact that O. flavus from this lowest depth looked unusual, led us to the decision to revisit the characterize the color of individuals of different sizes specifically from this depth. We found that some small-bodied O. flavus from 1000 m, in contrast to larger animals from the same depth, had a much brighter orange coloration (Figure 6b).
Moreover, the shape and coloration of eyes also differed within the adult representatives of O. flavus across sampling depths by becoming more ragged and paler with higher depths (Figure 7 and Figure S4). Reddish eye color in O. flavus from greater depths had been noted earlier [74] but never explored in detail. Color change was also noted for O. albinus eyes but the difference in shape was not so apparent.
Since the observed difference in eye coloration and shape may be attributed to an adaptation to extremely low-light conditions, we analyzed their opsins (visual proteins). According to our previous study based on transcriptome analysis, both species are characterized by a single expressed opsin gene, which belongs to the long wavelength-sensitive class [51]. To verify this result and also check if there was any difference between opsins sequences between depths, at which eyes looked different, we compared the sequences of opsins in animals from different depths. The same animals that were used for the eye photographs (Figure 7) and carotenoid extraction (Figure 5) were used. The sequences of the opsin fragment were identical within species and had four substitutions between the species; three of them led to amino acid changes (Figure 8a and File S5). The relative expression levels were also not different between depth or species (Figure 8b). However, it is worth noting that as the expression was normalized per the expression of a housekeeping gene, it should be proportionally higher in animals with larger eyes.
4. Discussion
In this work, we explored the genetic diversity and illumination-related traits of two deep-water scavenger amphipod species from Lake Baikal, namely O. flavus and O. albinus. The first important result is that we fully confirm the delimitation between these two species with molecular methods on a large sample. Previous works have also shown molecular distance between samples of these species [37,38,50], but none of them analyzed more than four samples per species or specimens from different locations, which could have largely affected the results. Thus, there is still a question of how two closely related and sympatric species had diverged. A plausible hypothesis could be that these species originally formed to occupy deep-water niches in two different basins of the lake and then contacted when the lake progressively deepened. According to paleogeographic scenarios, depths of hundreds of meters existed in the future northern and southern Baikal basins already at the early substage of the Proto-Baikal stage 30–10 million years ago, while the central part of the lake was much shallower [75]. However, before approximately 6 million years ago the water temperature was much higher, probably rendering the deep-water zone anoxic and uninhabitable for multicellular life [71,76]. Alternatively, adaptation-driven speciation with O. flavus more resistant to solar radiation and O. albinus better suited to darkness [44] also cannot be excluded at this stage. A further analysis of diversity in Ommatogammarus with more variable markers could be used to trace the origin of each species and test this hypothesis.
While the between-species genetic distance is obvious, we found that the genetic diversity of the COI fragment was homogeneous and quite low across 500-km horizontal distance and almost 1000-m vertical distance. This low intraspecific diversity of both species attracts considerable attention. The usage of the COI fragment may be seen as a pitfall to this work, as it is not variable enough to capture the variation happening within a smaller evolutionary scale. However, we need to underline that littoral Eulimnogammarus species, which are closely related to Ommatogammarus sp. [50], formed divergent COI lineages [31]. Thus, we believe that there is no particular reason for Ommatogammarus species to not accumulate COI differences. The low intraspecific diversity should mean that each of these species have recently come through a bottleneck. The plausibility of this hypothesis may depend on the age of the habitable (cold and thus oxygenated) deep-water zone in Lake Baikal. High depths in Baikal existed for a long time (see above), but the lake has undergone several episodes of significant warming [71], which could be the reason for shrinking of the habitable spaces and thus for a catastrophic population decrease. Specialization of O. flavus and O. albinus on scavenging [77] could also make them susceptible to variations in the ecosystem productivity and cause periodic depopulations for both species. Similar low intraspecific variability and evidence of a bottleneck event have been described for other Baikal species, particularly amphipods Gmelinoides fasciatus (Stebbing, 1899) and a gastropod mollusc Maackia herderiana (Lindholm, 1909) [78]. Some other Baikal animal groups, such as Lumbriculidae (Oligochaeta), underwent explosive speciation around 4–3 million years ago, probably also inhabiting the deeper parts of the lake at that time [79].
The absence of geographically partially or completely isolated genetic lineages suggests each species is highly connected throughout the lake. However, it does not mean that the individuals within each species have the same probability of mating to each other, i.e. that the whole lake is the habitat of one panmictic population. Even though these animals are highly mobile and can presumably cover large distances while swimming to access food sources based on the chemical cues, it sounds unrealistic that they could cover 100-km distances during a lifetime of several years. This is especially the case for sub-water elevations bringing the bottom to shallower depths than preferred by O. albinus, such as the Academician Ridge separating the northern and central basins of the lake, the central part of which is mostly above 350-m depth [80]. Still, we found no clues that the elevations can disrupt interbreeding of O. albinus on evolutionary meaningful times.
Yet, the most intriguing finding of this study is the vertical variability of O. flavus. In spite of genetic homogeneity, this species demonstrates less pigmented eyes with a more ragged edge and carotenoid-related shift in body coloration with greater depths, which are mostly not the case for O. albinus except for a slight change in eye pigmentation (Figure 5 and Figure 7). Importantly, both species have similar and stable opsin expression along the depth gradient (Figure 8), which indicates the leading role of pigments in adaptation of eyes to different illumination levels. These findings make us question how inclined the scavengers are to migrate to different depths and also highlight the potential adaptation-driven speciation of O. flavus and O. albinus as a plausible hypothesis.
It is hard to imagine how an eye shape could change in an adult animal. At the same time, it is conceivable to suggest that coloration may change throughout life, but this process is slow. Carotenoids are not synthesized by the vast majority of animal species and are thus obtained from food [73]. In our previous work with a shallow-water Baikal amphipod species Eulimnogammarus cyaneus (Dybowsky, 1874), we noticed differences in coloration upon a two-month experimental feeding with feeds differing in carotenoid abundance and did not even detect changes in the carotenoid contents at all [81]. Thus, it would be logical to suggest that the change that was seen in O. flavus would take at least months. For deep-ocean amphipods Eurythenes gryllus and Anonyx sp. Krøyer, 1838, body color intensity and carotenoid levels have been shown to positively correlate with body length, i.e. animal age [82]. However, in the case of Ommatogammarus, we did not find any clear evidence of ontogenetic depth stratification. Here we focused on adult individuals due to the complexity of species identification for small body-sized animals, but closer examination of the samples from 1000 m revealed a number of smaller individuals with typical O. flavus appearance (Figure 6).
Additional research exploring more variable genetic markers or tracking the distance, which an Ommatogammarus individual can travel, could shed more light at the stratification within these species. However, it is already safe to say that O. flavus is a convenient and promising model species to study the molecular mechanisms behind phenotypic plasticity of deep-water amphipods in the context of illumination-related traits.
5. Conclusions
In this work, we show that two species of endemic Baikal deep-water scavenger amphipods, O. flavus and O. albinus, have low intraspecific diversity of the marker fragment of the standard mitochondrial marker, cytochrome c oxidase subunit I gene, at both vertical (bathymetric) and horizontal (geographical) scales, suggesting absence of significant barriers for their distribution in the lake and also a bottleneck event in their history. At the same time, we did find even some visible differences (coloration, eye shape and color, and carotenoid contents) between O. flavus from different depths, indicating that the adult animals might not extensively migrate between depths within the lifespan of an individual. The differences in phenotypic plasticity point at probable evolutionary reduction of molecular mechanisms of pigment accumulation in O. albinus and potentially characterize the distinction in ecological niches between these two closely related species.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: representative photographs of O. flavus and O. albinus coxae I; Figure S2: photographs of 48 Ommatogammarus sp. individuals from different depths in Bolshie Koty showing differences in body color; Figure S3: photographs of larger samples of O. flavus from different depths in Bolshie Koty showing differences in body coloration; Figure S4: photographs of heads of 48 Ommatogammarus sp. individuals from different depths in Bolshie Koty showing differences in eye color and shape; Table S1: Coordinates of sampling points; Table S2: Primers used in this work; Table S3: Primer combinations used for PCR and sequencing for each sample; Table S: Measurements for the sample of O. flavus and O. albinus from different depths in Bolshie Koty; Table S5: Color and body length measurements for O. flavus from different depth; File S1: Morphological descriptions of Ommatogammarus flavus and Ommatogammarus albinus; File S2: map of the partial mitchondrial genome assembly for O. flavus containing the COI gene with annealed primers; File S3: map of the partial mitchondrial genome assembly for O. albinus containing the COI gene with annealed primers; File S4: multiple alignment of COI sequences obtained in this work; File S5: multiple alignment of long wavelength-sensitive opsin gene fragments obtained in this work.
Author Contributions
Conceptualization, E.T., A.G., M.T. and P.D.; Data curation, E.T., Y.S. and P.D.; Formal analysis, E.T., A.G. and P.D.; Funding acquisition, M.T.; Investigation, Y.S., T.P., A.M., Y.R, A.G. and P.D.; Methodology, E.T., A.G. and P.D.; Project administration, A.G. and P.D.; Resources, E.T., Y.S., T.P., A.M., Y.R., A.F. and A.G.; Software, E.T. and P.D.; Supervision, M.T.; Validation, P.D.; Visualization, E.T. and P.D.; Writing – original draft, E.T. and P.D.; Writing – review & editing, Y.S., T.P., A.M., Y.R., A.F., A.G. and M.T.
Funding
This research was funded by the Russian Science Foundation, grant number 23-14-00165 (https://rscf.ru/en/project/23-14-00165/).
Institutional Review Board Statement
The animal study protocol was approved by the Animal Subjects Research Committee of the Institute of Biology at Irkutsk State University (protocol #9/2022).
Data Availability Statement
The original data presented in the study are openly available in GitHub at https://github.com/Telnes/Population-structure-of-Ommatogammarus-flavus-and-O.-albinus/ and in the supplementary material.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Thurber, A.R.; Sweetman, A.K.; Narayanaswamy, B.E.; Jones, D.O.B.; Ingels, J.; Hansman, R.L. Ecosystem Function and Services Provided by the Deep Sea. Biogeosciences 2014, 11, 3941–3963. [Google Scholar] [CrossRef]
- Taylor, M.L.; Roterman, C.N. Invertebrate Population Genetics across Earth’s Largest Habitat: The Deep-Sea Floor. Mol. Ecol. 2017, 26, 4872–4896. [Google Scholar] [CrossRef]
- Webb, T.J.; Vanden Berghe, E.; O’Dor, R. Biodiversity’s Big Wet Secret: The Global Distribution of Marine Biological Records Reveals Chronic Under-Exploration of the Deep Pelagic Ocean. PLOS ONE 2010, 5, e10223. [Google Scholar] [CrossRef]
- Danovaro, R.; Snelgrove, P.V.R.; Tyler, P. Challenging the Paradigms of Deep-Sea Ecology. Trends Ecol. Evol. 2014, 29, 465–475. [Google Scholar] [CrossRef]
- Feng, J.-C.; Liang, J.; Cai, Y.; Zhang, S.; Xue, J.; Yang, Z. Deep-Sea Organisms Research Oriented by Deep-Sea Technologies Development. Sci. Bull. 2022, 67, 1802–1816. [Google Scholar] [CrossRef]
- Schön, I.; Martens, K. Adaptive, Pre-Adaptive and Non-Adaptive Components of Radiations in Ancient Lakes: A Review. Org. Divers. Evol. 2004, 4, 137–156. [Google Scholar] [CrossRef]
- Piccolroaz, S.; Toffolon, M. The Fate of Lake Baikal: How Climate Change May Alter Deep Ventilation in the Largest Lake on Earth. Clim. Change 2018, 150, 181–194. [Google Scholar] [CrossRef]
- Chapelle, G.; Peck, L.S. Polar Gigantism Dictated by Oxygen Availability. Nature 1999, 399, 114–115. [Google Scholar] [CrossRef]
- Shimaraev, M.N.; Domysheva, V.M. Trends in Hydrological and Hydrochemical Processes in Lake Baikal under Conditions of Modern Climate Change. Clim. Change Glob. Warm. Inland Waters Impacts Mitig. Ecosyst. Soc. 2013, 43–66. [Google Scholar] [CrossRef]
- Hampton, S.E.; Gray, D.K.; Izmest’eva, L.R.; Moore, M.V.; Ozersky, T. The Rise and Fall of Plankton: Long-Term Changes in the Vertical Distribution of Algae and Grazers in Lake Baikal, Siberia. PLoS One 2014, 9, e88920. [Google Scholar] [CrossRef]
- Arfianti, T.; Wilson, S.; Costello, M.J. Progress in the Discovery of Amphipod Crustaceans. PeerJ 2018, 6, e5187. [Google Scholar] [CrossRef]
- Horton, T.; De Broyer, C.; Bellan-Santini, D.; Coleman, C.O.; Copilaș-Ciocianu, D.; Corbari, L.; Daneliya, M.E.; Dauvin, J.-C.; Decock, W.; Fanini, L.; et al. The World Amphipoda Database: History and Progress. Rec. Aust. Mus. 2023, 75, 329–342. [Google Scholar] [CrossRef]
- Bowen, B.W.; Forsman, Z.H.; Whitney, J.L.; Faucci, A.; Hoban, M.; Canfield, S.J.; Johnston, E.C.; Coleman, R.R.; Copus, J.M.; Vicente, J.; et al. Species Radiations in the Sea: What the Flock? J. Hered. 2020, 111, 70–83. [Google Scholar] [CrossRef]
- Chenuil, A.; Saucède, T.; Hemery, L.G.; Eléaume, M.; Féral, J.-P.; Améziane, N.; David, B.; Lecointre, G.; Havermans, C. Understanding Processes at the Origin of Species Flocks with a Focus on the Marine Antarctic Fauna. Biol. Rev. 2018, 93, 481–504. [Google Scholar] [CrossRef]
- Drozdova, P.B.; Madyarova, E.V.; Gurkov, A.N.; Saranchina, A.E.; Romanova, E.V.; Petunina, J.V.; Peretolchina, T.E.; Sherbakov, D.Y.; Timofeyev, M.A. Lake Baikal Amphipods and Their Genomes, Great and Small. Vavilov J. Genet. Breed. 2024, 28, 317–325. [Google Scholar] [CrossRef]
- Copilaș-Ciocianu, D.; Sidorov, D. Taxonomic, Ecological and Morphological Diversity of Ponto-Caspian Gammaroidean Amphipods: A Review. Org. Divers. Evol. 2022. [Google Scholar] [CrossRef]
- Cristescu, M.E.; Adamowicz, S.J.; Vaillant, J.J.; Haffner, D.G. Ancient Lakes Revisited: From the Ecology to the Genetics of Speciation. Mol. Ecol. 2010, 19, 4837–4851. [Google Scholar] [CrossRef]
- Jamieson, A.J.; Weston, J.N.J. Amphipoda from Depths Exceeding 6,000 Meters Revisited 60 Years On. J. Crustac. Biol. 2023, 43, ruad020. [Google Scholar] [CrossRef]
- Kamaltynov, R.M. Amphipods (Amphipoda: Gammaroidea). In Index of animal species inhabiting Lake Baikal and its catchment area; Novosibirsk Branch of “Nauka” Publishing House: Novosibirsk, 2001; ISBN 978-0-02-018978-2. [Google Scholar]
- Kozhova, O.M.; Izmest’eva, L.R. Lake Baikal: Evolution and Biodiversity; Backhuys Publishers: Leiden, 1998; p. 225. [Google Scholar]
- Palumbi, S.R. Genetic Divergence, Reproductive Isolation, and Marine Speciation. Annu. Rev. Ecol. Syst. 1994, 547–572. [Google Scholar] [CrossRef]
- Dupont, D.W.E.; Patel, T.; Kochzius, M.; Schön, I. Evidence for a Single Population Expansion Event across 24,000 Km: The Case of the Deep-Sea Scavenging Amphipod Abyssorchomene distinctus. Hydrobiologia 2024, 851, 2309–2327. [Google Scholar] [CrossRef]
- Jażdżewska, A.M.; Horton, T.; Hendrycks, E.; Mamos, T.; Driskell, A.C.; Brix, S.; Arbizu, P.M. Pandora’s Box in the Deep Sea–Intraspecific Diversity Patterns and Distribution of Two Congeneric Scavenging Amphipods. Front. Mar. Sci. 2021. [Google Scholar] [CrossRef]
- Weston, J.N.J.; Jamieson, A.J. The Multi-Ocean Distribution of the Hadal Amphipod, Hirondellea dubia Dahl, 1959 (Crustacea, Amphipoda). Front. Mar. Sci. 2022. [Google Scholar] [CrossRef]
- Havermans, C.; Sonet, G.; d’Udekem d’Acoz, C.; Nagy, Z.T.; Martin, P.; Brix, S.; Riehl, T.; Agrawal, S.; Held, C. Genetic and Morphological Divergences in the Cosmopolitan Deep-Sea Amphipod Eurythenes gryllus Reveal a Diverse Abyss and a Bipolar Species. PloS One 2013, 8, 1–15. [Google Scholar] [CrossRef]
- Havermans, C. Have We so Far Only Seen the Tip of the Iceberg? Exploring Species Diversity and Distribution of the Giant Amphipod Eurythenes. Biodiversity 2016, 17, 12–25. [Google Scholar] [CrossRef]
- Weston, J.N.J.; Jensen, E.L.; Hasoon, M.S.R.; Kitson, J.J.N.; Stewart, H.A.; Jamieson, A.J. Barriers to Gene Flow in the Deepest Ocean Ecosystems: Evidence from Global Population Genomics of a Cosmopolitan Amphipod. Sci. Adv. 2022, 8, eabo6672. [Google Scholar] [CrossRef]
- Daneliya, M.E.; Kamaltynov, R.M.; Väinölä, R. Phylogeography and Systematics of Acanthogammarus s. Str., Giant Amphipod Crustaceans from Lake Baikal. Zool. Scr. 2011, 40, 623–637. [Google Scholar] [CrossRef]
- Drozdova, P.; Saranchina, A.; Madyarova, E.; Gurkov, A.; Timofeyev, M. Experimental Crossing Confirms Reproductive Isolation between Cryptic Species within Eulimnogammarus verrucosus (Crustacea: Amphipoda) from Lake Baikal. Int. J. Mol. Sci. 2022, 23, 10858. [Google Scholar] [CrossRef]
- Gomanenko, G.V.; Kamaltynov, R.M.; Kuzmenkova, Zh.V.; Berenos, K.; Sherbakov, D.Yu. Population Structure of the Baikalian Amphipod Gmelinoides Fasciatus (Stebbing). Russ. J. Genet. 2005, 41, 907–912. [Google Scholar] [CrossRef]
- Gurkov, A.; Rivarola-Duarte, L.; Bedulina, D.; Fernández Casas, I.; Michael, H.; Drozdova, P.; Nazarova, A.; Govorukhina, E.; Timofeyev, M.; Stadler, P.F.; et al. Indication of Ongoing Amphipod Speciation in Lake Baikal by Genetic Structures within Endemic Species. BMC Evol. Biol. 2019, 19, 138. [Google Scholar] [CrossRef]
- Petunina, J.V.; Vavrishchuk, N.V.; Romanova, E.V. Variabel’nost’ Morfologicheskikh i Geneticheskikh Priznakov Macrohectopus Branickii [Variability of Morphological and Genetic Traits of Macrohectopus Branickii]. In Proceedings of the Development of physicochemical biology, bioengineering and bioinformatics at the present stage; Publishing house of ISU: Irkutsk, October 25, 2023; pp. 111–113. [Google Scholar]
- Zaidykov, I.Y.; Naumova, E.Y.; Sukhanova, L.V. MtDNA Polymorphism of Macrohectopus Branickii Dybowsky, 1974 (Amphipoda) – An Endemic Pelagic Key Species of Lake Baikal. In Proceedings of the Complex Investigation of the World Ocean (CIWO-2023); Chaplina, T., Ed.; Springer Nature Switzerland: Cham, 2023; pp. 223–229. [Google Scholar]
- Bazikalova, A.Y. Amfipody Ozera Baikala [Amphipods of Lake Baikal]. Proc. Baikal Limnol. Stn. 1945, 11, 1–440. [Google Scholar]
- Takhteev, V.V. Ocherki o Bokoplavakh Ozera Baikal (sistematika, sravnitel’naya ekologiya, evolyutsiya) [Essays on the amphipods of Lake Baikal (systematics, comparative ecology, evolution)]; Irkutsk University Publishing House: Irkutsk, 2000; ISBN 5-7430-0123-5. [Google Scholar]
- Karaman, G.S. New Genus of Family Gammaridae from Baikal Lake, Abludogammarus, n. Gen. with References to Genus Ommatogammarus Stebb. (Contribution to the Knowledge of the Amphipoda 108). Glas. Od. Prir. Nauka Crnog. Akad. Nauka Umjet. Titogr. 1980, 3, 149–169. [Google Scholar]
- Macdonald III, K.S.; Yampolsky, L.; Duffy, J.E. Molecular and Morphological Evolution of the Amphipod Radiation of Lake Baikal. Mol. Phylogenet. Evol. 2005, 35, 323–343. [Google Scholar] [CrossRef]
- Shirokova, Y.A.; Saranchina, A.E.; Shatilina, Zh.M.; Kashchuk, N.D.; Timofeyev, M.A. Comparison of Olfactory Sensilla Structure in Littoral and Deep-Water Amphipods from the Baikal Region. Inland Water Biol. 2023, 16, 873–883. [Google Scholar] [CrossRef]
- Schoch, C.L.; Ciufo, S.; Domrachev, M.; Hotton, C.L.; Kannan, S.; Khovanskaya, R.; Leipe, D.; Mcveigh, R.; O’Neill, K.; Robbertse, B.; et al. NCBI Taxonomy: A Comprehensive Update on Curation, Resources and Tools. Database 2020, 2020, baaa062. [Google Scholar] [CrossRef]
- Horton, T.; Lowry, J.; Broyer, C.D.; Bellan-Santini, D.; Copilaș-Ciocianu, D.; Corbari, L.; Costello, M.J.; Daneliya, M.; Dauvin, J.-C.; Gasca, R.; et al. World Amphipoda Database 2025. Available online: https://www.marinespecies.org/amphipoda/ (accessed on 10th May, 2025).
- Ratnasingham, S.; Wei, C.; Chan, D.; Agda, J.; Agda, J.; Ballesteros-Mejia, L.; Boutou, H.A.; El Bastami, Z.M.; Ma, E.; Manjunath, R.; et al. BOLD v4: A Centralized Bioinformatics Platform for DNA-Based Biodiversity Data. In DNA Barcoding: Methods and Protocols; DeSalle, R., Ed.; Springer US: New York, NY, 2024; ISBN 978-1-0716-3581-0. [Google Scholar]
- Dybowsky, B.N. Beiträge Zur Näheren Kenntniss Der in Dem Baikal-See Vorkommenden Niederen Krebse Aus Der Gruppe Der Gammariden [Contributions to a more detailed knowledge of the lower crustaceans of the gammarid group found in Lake Baikal]; Buchdr. von W. Besobrasoff: St. Petersburg, 1874. [Google Scholar]
- Madyarova, E.; Shirokova, Y.; Gurkov, A.; Drozdova, P.; Baduev, B.; Lubyaga, Y.; Shatilina, Z.; Vishnevskaya, M.; Timofeyev, M. Metabolic Tolerance to Atmospheric Pressure of Two Freshwater Endemic Amphipods Mostly Inhabiting the Deep-Water Zone of the Ancient Lake Baikal. Insects 2022, 13. [Google Scholar] [CrossRef]
- Kondrateva, E.; Gurkov, A.; Rzhechitskiy, Y.; Saranchina, A.; Diagileva, A.; Drozdova, P.; Vereshchagina, K.; Shatilina, Z.; Sokolova, I.; Timofeyev, M. UV Sensitivities of Two Littoral and Two Deep-Freshwater Amphipods (Amphipoda, Crustacea) Reflect Their Preferred Depths in the Ancient Lake Baikal. Biology 2024, 13. [Google Scholar] [CrossRef]
- Takhteev, V.; Didorenko, S. Fauna i ekologiya bokoplavov ozera Baikal [Fauna and ecology of amphipods of Lake Baikal]; Publ. House Inst Geog VB Sochavy SB RAS: Irkutsk, Russia, 2015; ISBN 978-5-94797-244-3. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Drozdova, P.; Saranchina, A.; Morgunova, M.; Kizenko, A.; Lubyaga, Y.; Baduev, B.; Timofeyev, M. The Level of Putative Carotenoid-Binding Proteins Determines the Body Color in Two Species of Endemic Lake Baikal Amphipods. PeerJ 2020, 8, e9387. [Google Scholar] [CrossRef]
- Saranchina, A.; Mutin, A.; Govorukhina, E.; Rzhechitskiy, Y.; Gurkov, A.; Timofeyev, M.; Drozdova, P. Genetic Diversity in a Baikal Species Complex Eulimnogammarus verrucosus (Amphipoda: Gammaroidea) in the Angara River, the Only Outflow of Lake Baikal. Zool. Scr. 2024, 53, 867–879. [Google Scholar] [CrossRef]
- Drozdova, P.; Rivarola-Duarte, L.; Bedulina, D.; Axenov-Gribanov, D.; Schreiber, S.; Gurkov, A.; Shatilina, Z.; Vereshchagina, K.; Lubyaga, Y.; Madyarova, E.; et al. Comparison between Transcriptomic Responses to Short-Term Stress Exposures of a Common Holarctic and Endemic Lake Baikal Amphipods. BMC Genomics 2019, 20, 712. [Google Scholar] [CrossRef]
- Naumenko, S.A.; Logacheva, M.D.; Popova, N.V.; Klepikova, A.V.; Penin, A.A.; Bazykin, G.A.; Etingova, A.E.; Mugue, N.S.; Kondrashov, A.S.; Yampolsky, L.Y. Transcriptome-Based Phylogeny of Endemic Lake Baikal Amphipod Species Flock: Fast Speciation Accompanied by Frequent Episodes of Positive Selection. Mol. Ecol. 2017, 26, 536–553. [Google Scholar] [CrossRef]
- Drozdova, P.; Kizenko, A.; Saranchina, A.; Gurkov, A.; Firulyova, M.; Govorukhina, E.; Timofeyev, M. The Diversity of Opsins in Lake Baikal Amphipods (Amphipoda: Gammaridae). BMC Ecol. Evol. 2021, 21, 81. [Google Scholar] [CrossRef]
- Slater, G.S.C.; Birney, E. Automated Generation of Heuristics for Biological Sequence Comparison. BMC Bioinformatics 2005, 6, 31. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA Primers for Amplification of Mitochondrial Cytochrome c Oxidase Subunit I from Diverse Metazoan Invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Minton, J.A.L.; Flanagan, S.E.; Ellard, S. Mutation Surveyor: Software for DNA Sequence Analysis. In PCR Mutation Detection Protocols; Theophilus, B.D.M., Rapley, R., Eds.; Humana Press: Totowa, NJ, 2011; ISBN 978-1-60761-946-8. [Google Scholar]
- Okonechnikov, K.; Golosova, O.; Fursov, M. Unipro UGENE: A Unified Bioinformatics Toolkit. Bioinforma. Oxf. Engl. 2012, 28, 1166–1167. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing 2024.
- Paradis, E. Pegas: An R Package for Population Genetics with an Integrated–Modular Approach. Bioinformatics 2010, 26, 419–420. [Google Scholar] [CrossRef]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for Primary Species Delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef]
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: Assemble Species by Automatic Partitioning. Mol. Ecol. Resour. 2021, 21, 609–620. [Google Scholar] [CrossRef]
- Miralles, A.; Ducasse, J.; Brouillet, S.; Flouri, T.; Fujisawa, T.; Kapli, P.; Knowles, L.L.; Kumari, S.; Stamatakis, A.; Sukumaran, J.; et al. SPART: A Versatile and Standardized Data Exchange Format for Species Partition Information. Mol. Ecol. Resour. 2022, 22, 430–438. [Google Scholar] [CrossRef]
- GEBCO Compilation Group. GEBCO 2024 Grid. [CrossRef]
- Pante, E.; Simon-Bouhet, B.; Irisson, J. Marmap: Import, Plot and Analyze Bathymetric and Topographic Data. R Package Version 1.0. Available online at https://CRAN.R-project.org/package=marmap. (accessed on 14 March 2025).
- Massicotte, P.; South, A. Rnaturalearth: World Map Data from Natural Earth. R Package Version 03. Available online at https://CRAN.R-project.org/package=rnaturalearth. (accessed on 14 March 2025).
- Hothorn, T.; Hornik, K.; Wiel, M.A. van de; Zeileis, A. Implementing a Class of Permutation Tests: The Coin Package. J. Stat. Softw. 2008, 28. [Google Scholar] [CrossRef]
- Holm, S. A Simple Sequentially Rejective Multiple Test Procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar]
- Paradis, E.; Schliep, K. Ape 5.0: An Environment for Modern Phylogenetics and Evolutionary Analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer Verlag, 2016.
- Wickham, H. Forcats: Tools for Working with Categorical Variables (Factors) Available online:. Available online: https://cran.r-project.org/web/packages/forcats/index.html (accessed on 14 March 2025).
- Slowikowski, K.; Schep, A.; Hughes, S.; Lukauskas, S.; Irisson, J.-O.; Kamvar, Z.N.; Ryan, T.; Christophe, D.; Hiroaki, Y.; Gramme, P. ggrepel: Automatically Position Non-Overlapping Text Labels with 'ggplot2' https://CRAN.R-project.org/package=ggrepel. (accessed on 14 March 2025).
- Sherstyankin, P.P.; Alekseev, S.P.; Abramov, A.M.; Stavrov, K.G.; De Batist, M.; Hus, R.; Canals, M.; Casamor, J.L. Computer-Based Bathymetric Map of Lake Baikal. Dokl. Earth Sci. 2006, 408, 564–569. [Google Scholar] [CrossRef]
- Wade, N.M.; Gabaudan, J.; Glencross, B.D. A Review of Carotenoid Utilisation and Function in Crustacean Aquaculture. Rev. Aquac. 2017, 9, 141–156. [Google Scholar] [CrossRef]
- Maoka, T. Carotenoids as Natural Functional Pigments. J. Nat. Med. 2020, 74, 1–16. [Google Scholar] [CrossRef]
- Stebbing, T.R.R. Amphipoda; Friedlaender (R.), Berlin, 1906.
- Mats, V.D.; Shcherbakov, D.Y.; Efimova, I.M. Late Cretaceous-Cenozoic History of the Lake Baikal Depression and Formation of Its Unique Biodiversity. Stratigr. Geol. Correl. 2011, 19, 404–423. [Google Scholar] [CrossRef]
- Martin, P.; Martens, K.; Goddeeris, B. Oligochaeta from the Abyssal Zone of Lake Baikal (Siberia, Russia). Hydrobiologia 1999, 406, 165–174. [Google Scholar] [CrossRef]
- Takhteev, V.V. Trends in the Evolution of Baikal Amphipods and Evolutionary Parallels with Some Marine Malacostracan Faunas. In Advances in Ecological Research; Ancient Lakes: Biodiversity, Ecology and Evolution; Academic Press, 2000. [Google Scholar]
- Fazalova, V.; Nevado, B.; Peretolchina, T.; Petunina, J.; Sherbakov, D. When Environmental Changes Do Not Cause Geographic Separation of Fauna: Differential Responses of Baikalian Invertebrates. BMC Evol. Biol. 2010, 10, 320. [Google Scholar] [CrossRef]
- Kaygorodova, I.A.; Sherbakov, D.Y.; Martin, P. Molecular Phylogeny of Baikalian Lumbriculidae (Oligochaeta): Evidence for Recent Explosive Speciation. Comp. Cytogenet. 2007, 1, 71–84. [Google Scholar]
- Khlystov, O.M.; Kononov, E.E. Mesorelief of the Submarine Academician Ridge in Lake Baikal (Based on Newly Gathered Instrument Readings). Geogr. Nat. Resour. 2024, 45, 390–396. [Google Scholar] [CrossRef]
- Saranchina, A.; Drozdova, P.; Mutin, A.; Timofeyev, M. Diet Affects Body Color and Energy Metabolism in the Baikal Endemic Amphipod Eulimnogammarus Cyaneus Maintained in Laboratory Conditions. Biol. Commun. 2021, 66. [Google Scholar] [CrossRef]
- Thoen, H.H.; Johnsen, G.; Berge, J. Pigmentation and Spectral Absorbance in the Deep-Sea Arctic Amphipods Eurythenes Gryllus and Anonyx Sp. Polar Biol. Polar Biol. 2011, 34, 83–93. [Google Scholar] [CrossRef]
Figure 1.
Typical representatives of two species of the genus Ommatogammarus Stebbing, 1899. The left part of the image shows O. flavus (Dybowsky, 1874), and the right part depicts O. albinus (Dybowsky, 1874). a, photographs of live amphipods in a Petri dish on millimeter paper. The scale bar on photos is equal to 5 mm. b, morphology of the head segment of the body and shape of the eyes (redrawn from [42]). Representative photos of coxae I are shown in Figure S1.
Figure 1.
Typical representatives of two species of the genus Ommatogammarus Stebbing, 1899. The left part of the image shows O. flavus (Dybowsky, 1874), and the right part depicts O. albinus (Dybowsky, 1874). a, photographs of live amphipods in a Petri dish on millimeter paper. The scale bar on photos is equal to 5 mm. b, morphology of the head segment of the body and shape of the eyes (redrawn from [42]). Representative photos of coxae I are shown in Figure S1.
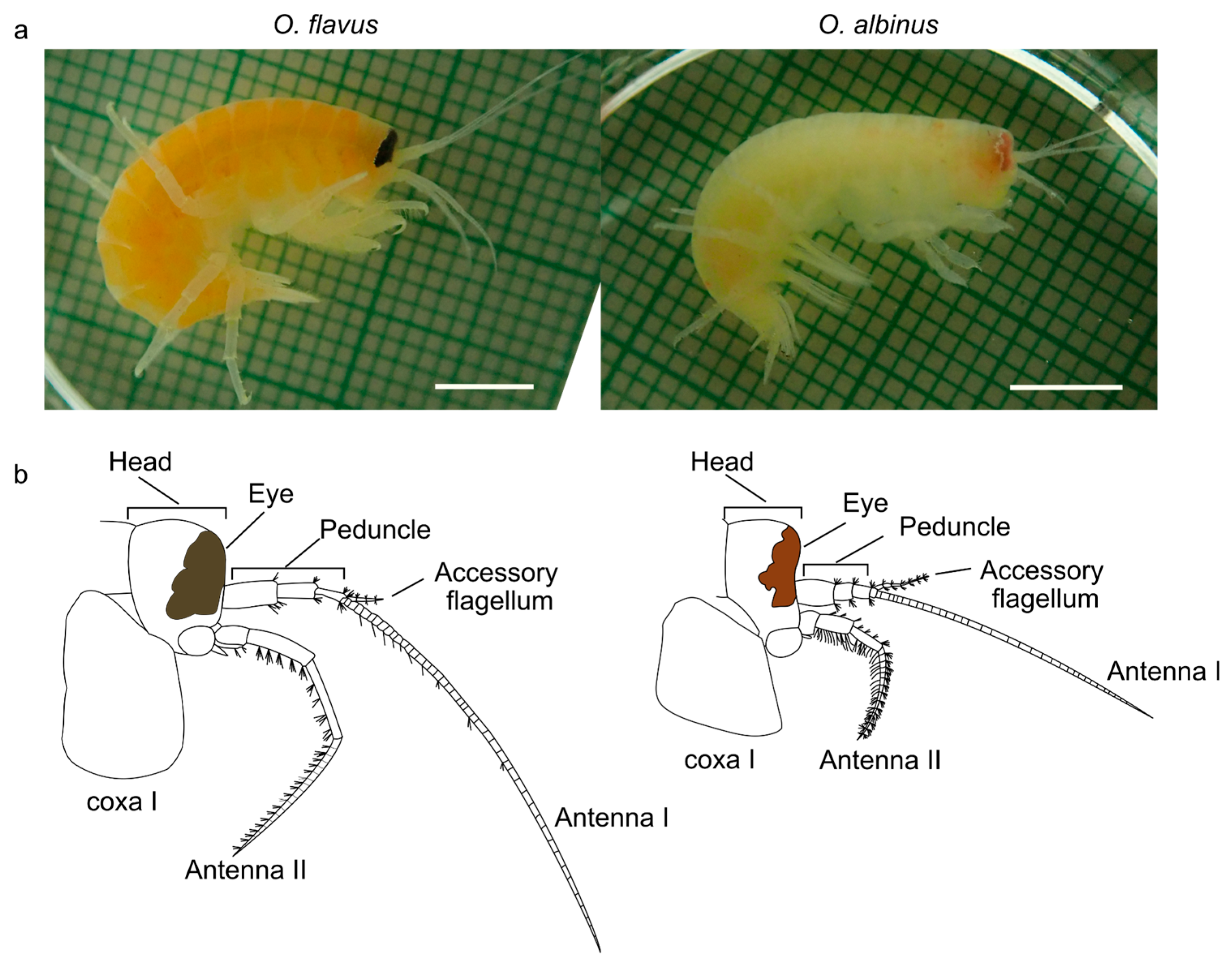
Figure 2.
Sampling points for all analyzes and sample sizes for COI sequencing. In the southern basin, five sampling points were analyzed, namely Bolshoe Goloustnoe (March 2020); Bolshie Koty (March 2023); Listvyanka (March 2025), Baikalsk (July 2023); Posolskoe (April 2024); Boyarsky (February 2025). In the central basin, one point was analyzed, namely Buguldeika (June 2023). In the northern basin, there were two points, specifically Ayaya Bay (August 2023) and Slyudyanka Bay (August 2023). Exact coordinates and sample sizes are listed in Table S1.
Figure 2.
Sampling points for all analyzes and sample sizes for COI sequencing. In the southern basin, five sampling points were analyzed, namely Bolshoe Goloustnoe (March 2020); Bolshie Koty (March 2023); Listvyanka (March 2025), Baikalsk (July 2023); Posolskoe (April 2024); Boyarsky (February 2025). In the central basin, one point was analyzed, namely Buguldeika (June 2023). In the northern basin, there were two points, specifically Ayaya Bay (August 2023) and Slyudyanka Bay (August 2023). Exact coordinates and sample sizes are listed in Table S1.
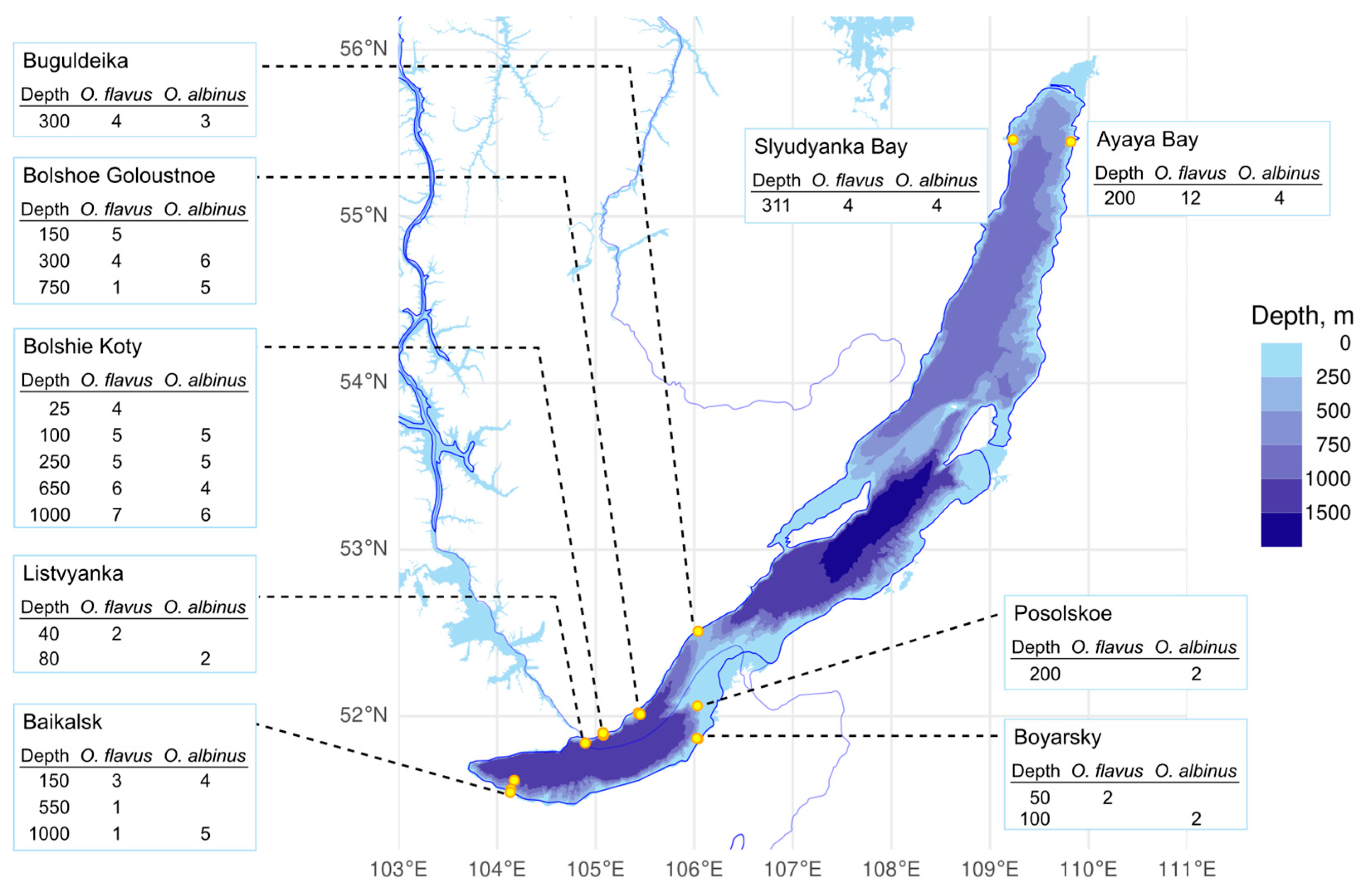
Figure 3.
Haplotype network for O. flavus and O. albinus from different depths near the Bolshie Koty settlement based on the 753-bp COI fragment.
Figure 3.
Haplotype network for O. flavus and O. albinus from different depths near the Bolshie Koty settlement based on the 753-bp COI fragment.
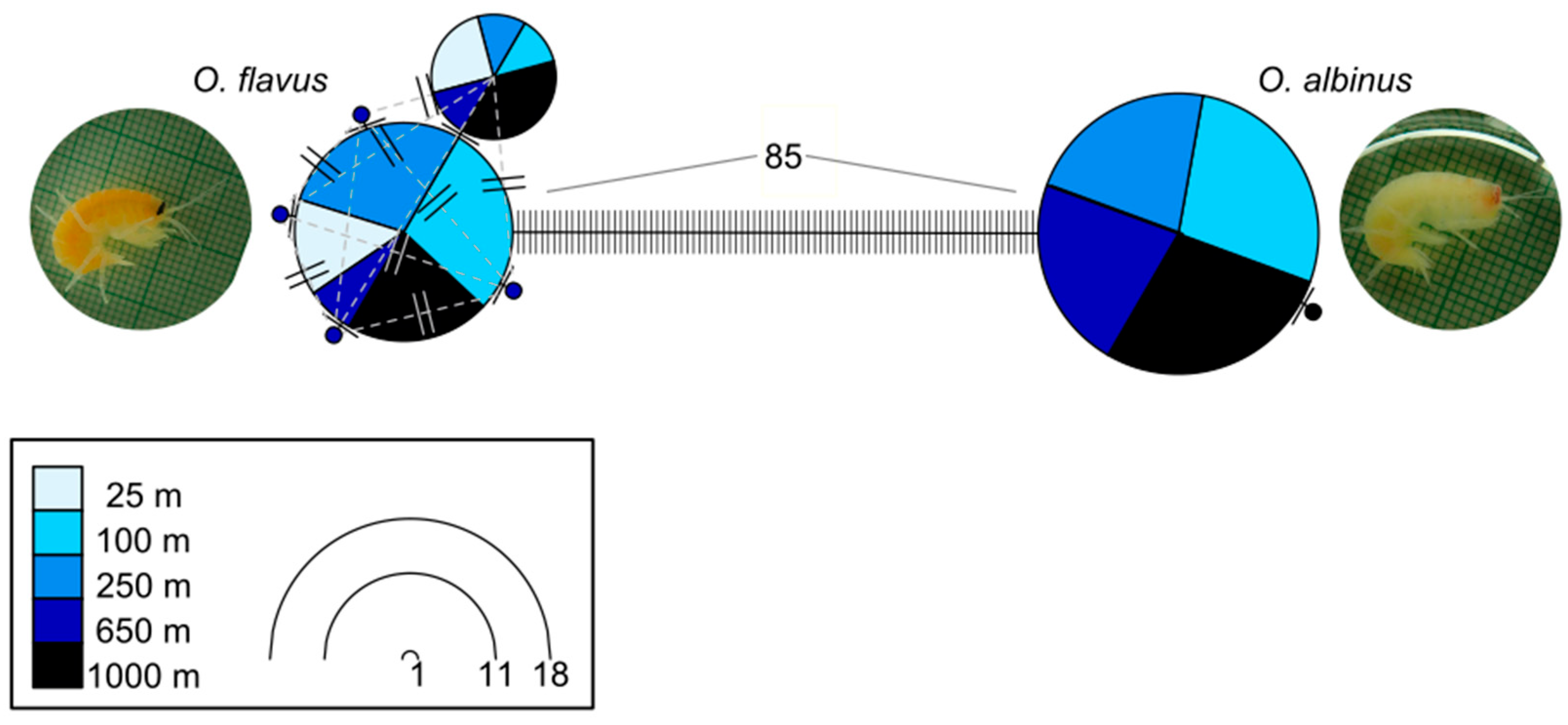
Figure 4.
Haplotype network for O. flavus and O. albinus from different sampling points across Lake Baikal based on a 608-bp COI fragment.
Figure 4.
Haplotype network for O. flavus and O. albinus from different sampling points across Lake Baikal based on a 608-bp COI fragment.
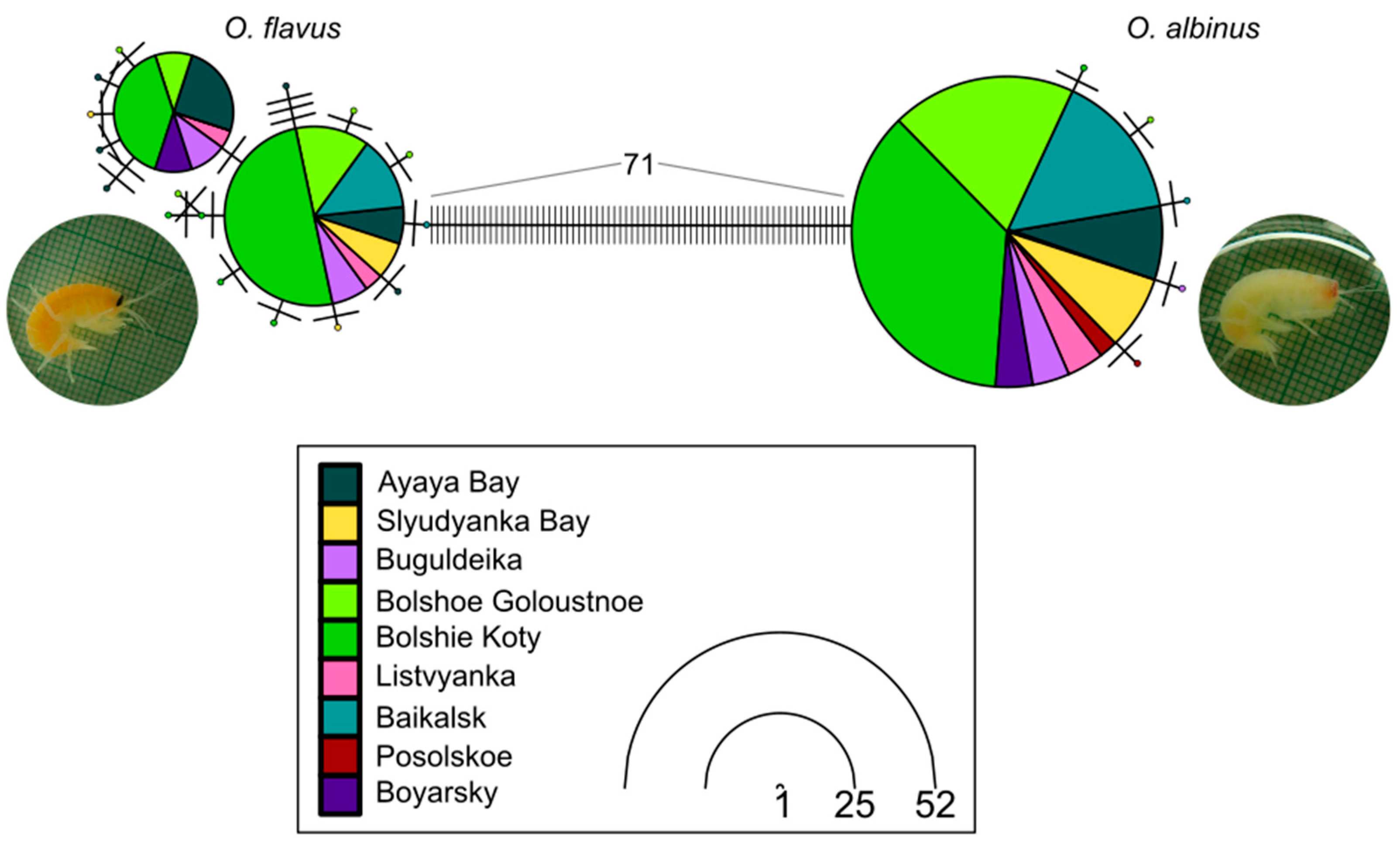
Figure 5.
Carotenoid levels in O. flavus change with sampling depth and correlate with body color. Photos (a–d) show typical appearance of animals sampled from low (a, c) or high (b, d) depths. Boxplots (e) and regression lines (f) show the correlation between sampling depth and carotenoid content (present in O. flavus and absent in O. albinus). Each dot corresponds to one animal.
Figure 5.
Carotenoid levels in O. flavus change with sampling depth and correlate with body color. Photos (a–d) show typical appearance of animals sampled from low (a, c) or high (b, d) depths. Boxplots (e) and regression lines (f) show the correlation between sampling depth and carotenoid content (present in O. flavus and absent in O. albinus). Each dot corresponds to one animal.
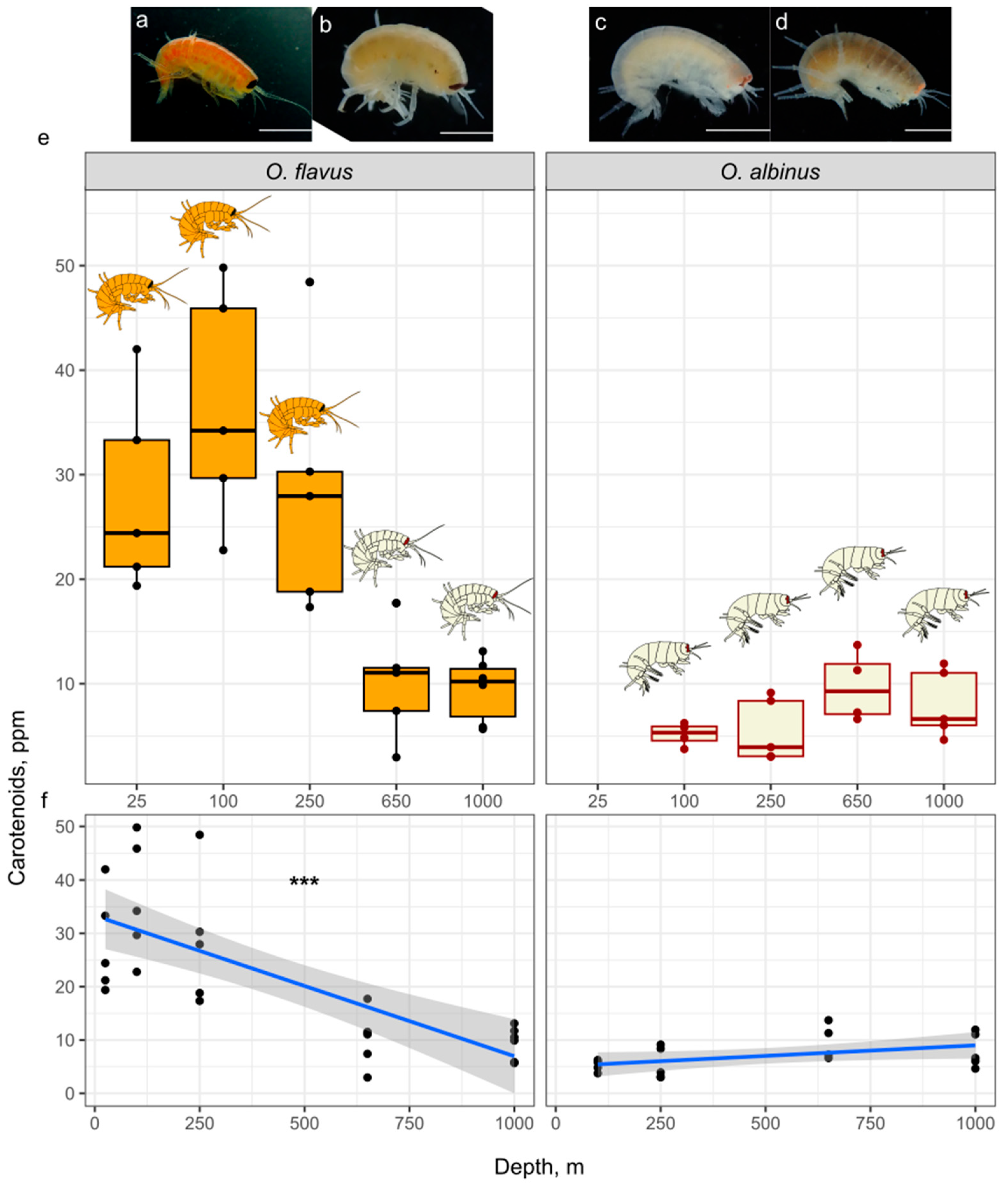
Figure 6.
Body coloration of adult O. flavus becomes paler with larger depth (a), but some small animals from the greatest depth (1000 m) do not follow this tendency.
Figure 6.
Body coloration of adult O. flavus becomes paler with larger depth (a), but some small animals from the greatest depth (1000 m) do not follow this tendency.
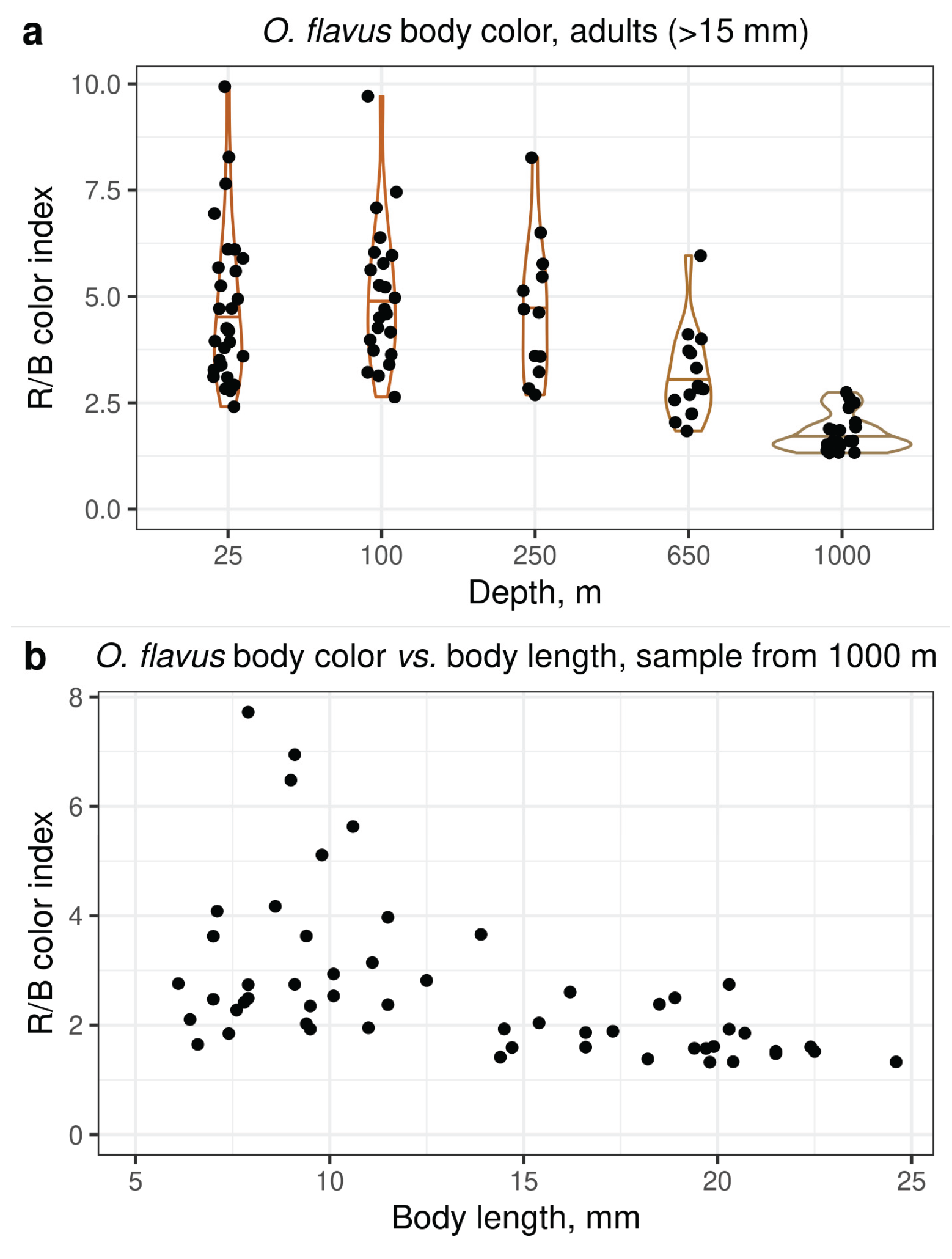
Figure 7.
Eye color and shape changes with sampling depth. Example photographs of eyes of O. flavus (upper row) and O. albinus (lower row) from different depths. In both species, the pigment becomes paler with depth, and the edge of the eye becomes more ragged (see also Figure S4 for all photographs).
Figure 7.
Eye color and shape changes with sampling depth. Example photographs of eyes of O. flavus (upper row) and O. albinus (lower row) from different depths. In both species, the pigment becomes paler with depth, and the edge of the eye becomes more ragged (see also Figure S4 for all photographs).

Figure 8.
The sequences of opsins do not differ across depths in sequence or expression. The haplotype network in (a) shows the haplotype diversity of 340-bp the opsin gene fragment in O. flavus and O. albinus (corresponding to amino acids 212–324 in Ommatogammarus sp. LWS opsins and amino acids 226–298 in Bos taurus rhodopsin). In this analysis, three to four animals per depth were analyzed. The boxplots in (b) depict expression levels of opsin genes relative to the housekeeping gene gapdh; each point corresponds to one animal. The schematic eye shapes and colors above the plots are drawn to illustrate the difference in pigmentation.
Figure 8.
The sequences of opsins do not differ across depths in sequence or expression. The haplotype network in (a) shows the haplotype diversity of 340-bp the opsin gene fragment in O. flavus and O. albinus (corresponding to amino acids 212–324 in Ommatogammarus sp. LWS opsins and amino acids 226–298 in Bos taurus rhodopsin). In this analysis, three to four animals per depth were analyzed. The boxplots in (b) depict expression levels of opsin genes relative to the housekeeping gene gapdh; each point corresponds to one animal. The schematic eye shapes and colors above the plots are drawn to illustrate the difference in pigmentation.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



