
Submitted:
16 January 2026
Posted:
19 January 2026
You are already at the latest version
Abstract
Reindeer in the southern Palearctic remain poorly documented, especially regarding helminths. Limited data exist for their small, isolated, and conservation-important populations. Because helminths affect survival, reproduction, and population stability, and act as biogeographic indicators, knowledge of their diversity in these southern regions is considered essential for research and management purposes. A total of 242 fecal samples were obtained from 2012 to 2025 from reindeer inhabiting China, Kazakhstan, Mongolia, and Russia. These samples were examined by coproscopy. Morphological diagnoses were supplemented with DNA analyses. Trematodes of Paramphistomoidea, cestodes of Moniezia, and nematodes identified as E. rangiferi, O. macrotis, the dimorphic O. gruehneri/O. arctica, as well as Nematodirus, Capillaria, and unidentified small strongylids were revealed. All taxa detected in this study have been reported previously for R. tarandus and for the Palearctic, exception for Nematodirus sp. for the southern area. However, examinations of reindeer from selected regions in Russia, as well as Mongolia and Inner Mongolia (China) were carried out for the first time. Southern range limits were established for E. rangiferi in China and O. macrotis in Russia. Species O. macrotis has been proposed as a biogeographical marker for wild reindeer in the Eastern Siberia, while Capillaria may indicate domestic herds.
Keywords:
Capillaria
; Orthostrongylus macrotis
; Elaphostrongylus rangiferi
; wild reindeer
; semiwild reindeer
; China
; Kazakhstan
; Mongolia
; Russia
1. Introduction
The reindeer (Rangifer tarandus) is regarded as an ecologically important animal. It is considered as a flagship species that attracts broad public attention [1]. It also functions as an umbrella species, because the conservation of reindeer, which often undertake extensive migrations, requires the protection of large landscapes [2]. Globally, the species’ conservation status in the IUCN Red List has been revised from Least Concern to Vulnerable nine years ago [3]. Even greater concern has been expressed at the regional level, where several reindeer populations have been placed in local Red Data Books as endangered, and strict protection has been required for these populations [4]. Many wild populations have been preserved only through the establishment of nature reserves. The reindeer is also an indicator species. Analyses of its tissues or excretes can reveal environmental contamination, including radionuclides [5].
The reindeer is represented by two forms — wild and semiwild (domestic) [6]. Unlike conventional livestock, most semiwild reindeer lead virtually the same migratory lifestyle as wild reindeer and are influenced by the same factors. The main difference is that domestic herds are accompanied by humans (Indigenous nomadic herders), who provide protection and care including, though not always, veterinary support. Wild reindeer may also lure individual semiwild reindeer into their herds, and the latter can sometimes be retrieved [7]. Thus, the boundary between wild and semiwild populations is permeable. For this reason, studies of the biology and ecology of wild reindeer should also take into account data from semiwild animals. Domestic reindeer are better studied and can, to some extent, serve as a model for the species as a whole.
The reindeer is commonly associated with high latitudes. This view is reflected even in its common names: the Russian term for reindeer, “severnyi olen”, points to the north (“sever”), and the North American word “caribou” refers to snow [8]. However, the species’ range is much broader. In the Pleistocene (12,000 years ago), it was likely continuous and extended to about 45° N [9,10]. Today, the range is fragmented and shifted northward. Only isolated southern populations persist, and these are little known outside specialist circles [11]. One of the most unexpected examples is the coexistence of reindeer with the Chara Desert in Transbaikalia (Russia) [12].
The global population of Rangifer species is estimated at roughly 4 million animals [6,11]. In Palearctic, reindeer are most numerous in the northern part: the wild herds on the Taymyr Peninsula or the semiwild herds in the Yamal-Nenets Autonomous Okrug (Russia) number in the hundreds of thousands. Individual herds may contain several thousand animals. In the southern part of the range, the situation is different: reindeer occur in small groups, usually consisting of a few dozen animals (sometimes even single individuals). Only rarely do such groups reach several hundred [13].
The helminths are limiting factors for their host populations and, the same time, potential indicators of these populations. Helminths can affect immunity, growth, and fecundity, compromise survival or cause the direct death of the reindeer hosts [6,14]. They are even able to influence the sex ratio of ruminants’ offsprings [15]. Meanwhile, stable host populations generally support a richer helminth fauna, whereas declining populations tend to harbor a depauperate parasite community [16]. Highly specialized helminths can also serve as biogeographic markers for stock discrimination of particular host populations [17].
The helminths of R. tarandus across the Holarctic have been studied quite thoroughly, but this applies primarily to northern regions and semiwild animals [6,14]. Data from the southern part of the Palearctic range are extremely limited, especially for wild reindeer, despite the high ecological and conservation value of these small, isolated populations [18,19,20,21,22,23,24]. Our research is the first attempt to examine the helminths of wild reindeer in the southern part of their continental Palearctic range. To provide a more complete picture, semiwild reindeer were also included in the analysis.
The aim of this work was to study the diversity and distribution of reindeer helminths in the southern continental Palearctic via coprological survey.
2. Materials and Methods
2.1. Fecal Sampling
Fecal samples were collected from both wild and semiwild reindeer inhabiting the southern part of the species’ continental range in the Palearctic. The dataset includes samples from China, Kazakhstan, Mongolia, and Russia, collected between 50° and 60° N latitude. The sampling period spanned from 2012 to 2025. Samples were collected at 12 sites (Figure 1). Four of them were inhabited by semiwild reindeer (sites 1, 4, 7, and 9), and eight were inhabited by wild reindeer (sites 2, 3, 5, and 12 within protected areas, and sites 6, 8, 10, and 11 in the wild). Site 5 was sampled on four separate occasions, resulting in a total of 15 fecal sample sets included in the study (Table 1).
Fecal samples were collected from the ground or snow without disturbing the reindeer. Most samples were obtained immediately after defecation. Sets 5, 7, 8, and 9 were exceptions; for these, sampling was conducted in reindeer habitat without direct observation of defecation. Thus, for these sets, the season of sample collection did not necessarily coincide with the season in which defecation occurred. Each fecal sample within each set was placed in an individual plastic container and labeled with the date and location of collection. In some cases, primarily involving semiwild reindeer, additional information was available, including the animal’s sex and age. These supplementary data are provided in the Supplementary Materials (S1).
Each fecal sample consisted of 12–15 fecal pellets. Most sets were delivered to the Center of Parasitology of the IEE RAS in Moscow, Russia in a moist state and stored at +4 °C. Sets 5, 6, and 12 dried naturally and were delivered in a dry state.
2.2. Fecal Analysis
A total of 242 fecal samples — 180 from wild and 62 from semiwild reindeer — were examined for helminths in accordance with the current state standard of the Russian Federation GOST R 54627-2011 [25]. Initially, all samples were inspected for adult helminths or their fragments, such as cestode proglottids. Each sample was then analyzed using three additional methods.
The first method was drip larvoscopy following Vajda’s technique [26]. Three drops of tap water were placed on a microscope slide at some distance from each other, and one fecal pellet was placed into each drop. The pellets were gently rinsed with several additional drops of water to ensure complete wetting of their surfaces. The preparation was left to stand for 40 minutes; for dry pellets, the exposure time was increased to 1.5 hours. The pellets were then removed with forceps, and the remaining liquid on the slide was examined under a microscope, without a coverslip, for larvae of zooparasitic nematodes.
The second method was combined ovoscopy according to Darling [27]. Three grams of feces were mixed with 60 ml of tap water, homogenized in a mortar with a pestle, filtered into a 20-ml centrifuge tube, and centrifuged at 1500 rpm for 5 minutes. The supernatant was decanted, and Darling’s flotation solution — a pre-prepared mixture of equal parts glycerol and saturated sodium chloride solution — was mixed with the sediment to fill the tube. The tube was centrifuged again under the same conditions. Using a fecal loop, several drops from the surface layer (depending on the loop diameter) were transferred to a microscope slide, covered with a coverslip, and examined for helminth eggs.
The third method was sedimentation ovoscopy via sequential washings. Three grams of feces were mixed with 60 ml of tap water, homogenized, and filtered into a beaker. After allowing the suspension to settle for several minutes, the supernatant was carefully poured off and replaced with fresh tap water. The procedure was repeated until the supernatant became clear. The remaining water was then decanted by inverting the beaker, and the residual drops containing the sediment were transferred to a slide, covered with a coverslip, and examined microscopically for helminth eggs.
The use of two ovoscopic techniques was justified by their differing sensitivity to light versus heavy eggs. However, in practice, each method occasionally allowed the detection of other developmental stages, such as eggs with the Vajda’ method or larvae with the ovoscopic methods (see Table S1).
Quantitative analysis was carried out using a VIGIS counting chamber from the Diapar kit (VIGIS, Moscow, USSR), which is an analogue of the McMaster chamber.
2.3. Helminth Identification
Preparations were initially examined, and helminths were identified based on their morphological and morphometric characteristics. Morphological criteria for eggs included their shape, shell structure, presence of an operculum or polar plugs, color, and the type of internal contents. For larvae, diagnostic features included the structure of the intestine (homogeneous or visible intestinal cells), the morphology of the tail (including the presence of spines or spikes), and the presence of a sheath. Position of larva at rest was also taken into consideration.
Morphometric characteristics included, for eggs, their length and width. For larvae, the following measurements were considered: total body length, maximum width at the level of the esophagus, distance from the anterior end to the nerve ring, excretory pore, junction of the esophagus and intestine, genital primordium, and anus, as well as tail length, spine or spike length (when present), length of the caudal portion of the sheath, and length of the sheath filament.
Morphological and morphometric analyses were performed using a Mikmed-6 light microscope (LOMO-MA, Russia) equipped with a digital Canon 5D Mark II camera (Canon, Japan) connected via an optical–mechanical C-mount adapter (LOMO-MA, Russia). Photomicrographs were obtained at 400× magnification (200× for third-stage larvae). Measurements were taken from the resulting images using Fiji/ImageJ Version 1.2.4, RRID:SCR_003070 (National Institutes of Health, USA). Prior to analysis, the software was calibrated using an officially certified OMP micrometer calibration slide (LOMO-MA, Russia). Linear dimensions of eggs were measured using the Straight Line tool, while larval measurements additionally employed the Segmented Line tool when specimens were curved. To immobilize live larvae, they were briefly heated over a lighter flame for 3–5 seconds (heat shocked).
2.4. DNA Analysis
The first- or third-stage larvae (L1s or L3s) of zooparasitic nematodes were placed into 0.5-ml Eppendorf tubes. DNA extraction was performed using the QIAamp DNA Accessory Set, Micro Kit (Qiagen, Netherlands) according to the manufacturer’s protocol. From each nematode sample, 1.0 µl of extracted DNA was used for polymerase chain reaction (PCR). Taxonomic identification of nematodes was based on sequencing of the internal transcribed spacer regions of ribosomal DNA (ITS rDNA). The study employed two conserved 20-base oligonucleotide primers, NC1 (5’-ACG-TCT-GGT-TCA-GGG-TTG-TT-3’) and NC2 (5’-TTA-GTT-TCT-TTT-CCT-CCG-CT-3’), originally described by Gasser [34]. For this purpose, PCR was carried out on a T100 Thermal Cycler (Bio-Rad, USA). The PCR followed a standard thermocycling scheme comprising an initial denaturation phase (3 min at 95 °C), 35 cycles of denaturation (30 s at 95 °C), primer annealing (45 s at 95 °C), and elongation (1 min at 72 °C), and concluded with a final elongation period (5 min at 72 °C).
Amplification success was assessed by electrophoretic separation of PCR products in a 1% agarose gel for 30 minutes at 100 V using the Mini-Sub Cell GT + PowerPac Basic system (Bio-Rad, USA), followed by visualization on an EXC-F15.M transilluminator (Vilber Lourmat, Germany). When a product of the expected size was obtained, as determined from published data or prior experience, its yield was evaluated. In cases where only faint bands were observed, an additional round of amplification was performed. Adequate quantities of PCR product were pooled and purified in a 0.8% agarose gel through prolonged electrophoresis (70–90 minutes). The DNA band was excised, and PCR product was extracted using the SV Gel and PCR Clean-Up kit (Promega, USA).
The eluate was further purified by ethanol precipitation in the presence of ammonium acetate. The resulting pellet was resuspended in high-purity autoclaved molecular grade water (G-Biosciences, USA). The volume of added water (10–40 µl) was adjusted depending on band intensity in the gel. DNA concentration was measured using a Nanodrop 2000 spectrophotometer (Thermo Fisher Scientific, USA). Samples with high DNA concentrations were diluted to 15–30 ng/µl. Direct sequencing was subsequently performed at the Genotech® (Moscow, Russia).
2.5. Statistical Analysis
Statistical analyses were performed using the Microsoft Excel 2007 Analysis ToolPak, RRID SCR_016137 (Microsoft Corporation, USA). The numbers of eggs and larvae of each detected helminth kind were first determined for every 3-g fecal sample from reindeer. These values were obtained by summing the results generated by the three diagnostic methods (see Supplementary Materials, S1). The counts were then recalculated per 1 g of feces, yielding invasion-intensity values for each helminth kind in every positive fecal sample [37]. Arithmetic means and standard errors of the mean (SEM) were subsequently calculated for invasion intensity within each positive Sample Set for each identified kind of helminth. These values represent the mean number of eggs or larvae of a given helminth kind per 1 gram of reindeer feces for samples collected at a single site. Data from Sample Sets 5, 6, 7 and 8 were pooled to obtain a single estimate of mean invasion intensity for site 5, because these Sample Sets had been collected at that site in different years.
Sample prevalence was calculated as the proportion of positive fecal samples relative to the total number of examined samples for every Sample Set.
Attempts to identify any influence of sex or age of reindeer on helminth infection were abandoned, as such information was largely unavailable. The effect of season on infection with helminths also could not be assessed reliably, because, as noted above, the season of sample collection did not always correspond to the season in which the feces were originally deposited by the reindeer. Therefore, the dataset was insufficient for the use of a General Linear Model or comparable analytical tools.
However, a trend of different infection with helminths between wild and semiwild reindeer was highlighted. All positive sample sets (including combined positive sample sets for the site 5) were divided into two groups, consisting of fecal samples from wild and semiwild animals, respectively. For each group, the proportion of positive sites for each helminth kind relative to the total number of positive sites was calculated (see Supplementary Materials, S2). These data were then visualized as a bar chart.
3. Results
In the fecal samples from reindeer, no intact helminths or their fragments were found during the external examination. Eggs of trematodes of the superfamily Paramphistomoidea were found via sedimentation. Eggs of cestodes of the genus Moniezia were also detected via flotation and sedimentation. Eggs and L3 larvae of small strongylid nematodes, eggs of Nematodirus, eggs of Capillaria, and L1 larvae of Protostrongylidae were detected as well (see S1). These Protostrongylidae L1 larvae included both dorsal-spined larvae (DSL) and Protostrongylus-type morphotypes. The DSL were identified as Elaphostrongylus rangiferi, and the Protostrongylus morphotype was identified as Orthostrongylus macrotis. For E. rangiferi larvae from sample set 12 (site 9), DNA analysis confirmed the primarily identification. DNA analysis failed for all other DSLs, so their identification was based on morphology and expected host specificity. For O. macrotis, DNA analysis was successful. In sample set 15 (site 12), the L3 larvae were identified as Ostertagia gruehneri based on morphology and DNA analysis. The nucleotide sequences obtained were deposited in GenBank (Table 2).
The appearance of the helminths found in reindeer feces is shown in Figure 2.
In the fecal samples collected at sites 1, 3, 6, and 11, no helminths were detected. The data on sample prevalence and intensity are presented in the Table 3.
The intensity of infection by all detected helminth kinds in all samples is interpreted as low according to GOST R 54627-2011 terminology [25]. The ranges from minimum to maximum values for each helminth kind are presented in Table 4.
Comparative distribution of different helminths among sites corresponding to the wild and semiwild reindeer habitats is presented in Figure 3. This analysis is purely descriptive, and the authors acknowledge several limitations. Sample sizes are small, with fewer than ten positive sites per each group. The number of samples per positive site is uneven, ranging from 8 to 90. The data represent frequencies in the samples rather than in individual animals, as a single sample cannot always be linked to a single reindeer when defecation was not directly observed. The data also do not represent population-level prevalence.
4. Discussion
The limited available sources indicate that during the 20th century helminths of the reindeer in the Palearctic between 50° and 60° N were studied only in the USSR. Published records exist for reindeer from the Khabarovsk Krai, present-day Buryatia, and Tuva (Russia) [18,19,20,21,22,38]. In most of these studies, the Scriabin’s helminthological dissection method [39] was used, and domestic reindeer were taken as the study animals. In the 21st century, only two relevant studies have been produced. One concerned Kazakhstan and the other concerned the Magadan Region of Russia [23,24]. Both were conducted by Loginova, who is also one of the authors of the present article. Thus, no site (included in our study) has been examined before for the same reindeer type and by the same methodological approach, resulting in no possible adequate comparison. However, within the combined area that was surveyed, the helminth fauna detected in our study has already been documented, with the exception of the representative of the family Molineidae. Machulsky [18], Sulimov [19], and Kadenatsi [21] reported Nematodirella longissimespiculata, whereas Nematodirus spp. were found in our material. Mizkewitsch noted that these two genera often occur together [14]. Given the low infection intensity (<1 egg / 1 g of feces) and the fact that eggs were detected in only 4 of 242 samples, it can be assumed that cases of such mixed infection simply did not appear in our dataset.
Eggs of the rumen flukes in the superfamily Paramphistomoidea that were detected in our material may belong to species of the genus Paramphistomum (Paramphistomidae), such as P. cervi, widely reported for reindeer [6,14,21]. They may also belong to Calicophoron daubneyi (Paramphistomidae), reported from cervids in the Palearctic [40], or to Fischoederius elongatus (Gastrothylacidae), which was recorded by Zhaltsanova in reindeer from Buryatia [22]. The eggs of these trematodes are very similar, and their size ranges overlap [28]. This group is still in need of profound taxonomic revision [41]. For these reasons, identification was restricted to the superfamily level.
Species of Moniezia reported for reindeer in the south was identified either as M. expansa in semiwild reindeer in Buryatia [18], M. baeri in semiwild reindeer in Tuva [19] or M. benedeni in wild reindeer in Tuva [20] and semiwild reindeer in Khabarovsk Krai [21]. The Moniezia eggs found in our material tended toward a square shape in two-dimensional projection. For this reason, the species M. expansa, which is characterized by triangular eggs, has been excluded from the diagnosis. The diagnosis remains at the genus level, because a recent revision has shown that its taxonomy requires further development [42].
The small strongylid eggs that were detected varied in both size and shape. This variation provides a basis for assuming the presence of multiple species from different genera, including, for instance, Cooperia, Haemonchus, Ostertagia, Spiculopteragia, Teladorsagia, Trichostrongylus, and others. A species-level diagnosis cannot be determined without molecular-genetic analyses, because these nematodes are characterized by egg polymorphism and by a dependence of egg size on the host species and its body size. The possibility of interspecific hybridization, along with other influencing factors, cannot be excluded [30]. The material from wild reindeer at site 12 allowed the species identity of the small strongylid to be determined through molecular-genetic analysis. The species was identified as Ostertagia gruehneri. This species is currently regarded as dimorphic, with the major morph corresponding to O. gruehneri and the minor morph to O. arctica [43]. It is noteworthy that O. gruehneri was originally described by Skryabin in 1929 from domestic reindeer precisely in the southern part of the species’ range (Khabarovsk Krai) [38]. In the same year (1929), the minor morph was described by Mizkewitsch from material obtained in Arkhangelsk [14,38]. Findings of O. gruehneri and O. arctica from both domestic and wild reindeer in Tuva were reported by Sulimov [19,20], while Kadenatsi documented O. gruehneri in domestic reindeer from Khabarovsk Krai [21].
The Palearctic reindeer can host two known protostrongylid species with the DSL larval morphotype, E. rangiferi and Varestrongylus eleguneniensis [6,14]. Although an exact identification of larvae can be achieved only through DNA analysis, there are some morphological differences between the larvae of these two species. For domestic reindeer from China (site 9), the morphological diagnosis was supported by molecular-genetic confirmation. In all other cases, the species was identified on the basis of morphological characters and the well-documented broad distribution of E. rangiferi. In contrast, V. eleguneniensis has been recorded from wild reindeer in the Palearctic only once [44]. Unidentified species of Elaphostrongylus was also reported for semiwild reindeer in Tuva in 20th centiry [19].
Another protostrongylid found in reindeer feces was O. macrotis. This species was described for Nearctic [31]. For the first time in Palearctic, it was found (and identified genetically) in 2022 in wild reindeer feces collected at the site with coordinates 71.18833 N, 98.07083 E [45]. Later, it was found (and identified morphologically) in wild reindeer feces collected in Magadan Oblast (63.72640 N, 158.07980 E) [24]. In our research, it was found (and identified genetically) in the feces of wild reindeer collected at the site 12. These data are not enough for a solid conclusion, but they allow suggestion that there is a region in the Eastern Siberia harboring O. macrotis. Another suggestion is that O. macrotis can be considered as a biogeographic marker for wild reindeer of this region. Testing these two hypotheses require further investigation and broader sampling.
In contrast, Capillaria sp. eggs were found only in semiwild reindeer (in Mongolia and China, sites 4 and 9, consequently). For the southern range of reindeer, it was also reported for semiwild reindeer in Tuva [19] and Khabarovsk Krai [21]. In all known previous studies for Palearctic, Capillaria was found only in semiwild reindeer [6,14,33,44]. This trend supports the suggestion that Capillaria can be considered as a biological marker for semiwild reindeer in general [44].
It should be noted that no natural populations of reindeer are present in Kazakhstan. In both documented cases ([23] and this study), the animals were kept in two private menageries, where several individuals had been brought from Russia in the 21st century. However, these private introduction attempts still require monitoring. For this reason, their inclusion in the present study is considered important.
In all cases with positive samples, the intensity was low. This may be regarded as indirect evidence that the animals producing these feces were in satisfactory health. The authors acknowledge the following biases. The number of eggs detected does not always correlate directly with the number of mature helminths. Helminth reproductive output shows daily and seasonal fluctuations. Tissue-dwelling and body-cavity helminths are not detected by coproscopy, including larval Taeniidae and adult Setaria, Onchocerca, and other such parasites. Coproscopic results may be falsely negative when the helminths are still immature and not yet producing eggs or larvae. Animals with extremely heavy infections may be missing from the sample because they die in remote areas and thus remain unsampled. However, the format of coproscopy was chosen because it is non-invasive, unlike lethal sampling required for the Scriabin’s helminthological dissection method [39]. This approach allows work with live, protected wild reindeer, and it permits reliable comparison with data obtained from semiwild animals.
At the population level, it can be concluded that our data support the view that relatively greater helminth diversity and higher helminth loads are associated with a stable host population. Thus, the known groups of wild northern Siberian forest reindeer (Rangifer tarandus valentinae Flerov, 1932) preserved in the Altai (site 3) and Baikal Nature Reserves (site 5) are characterized by very small numbers, often only a few dozen individuals. This subspecies is listed in the Red Data Book of the Russian Federation as an endangered population [4]. In 128 samples collected from this population, only three helminth kinds were detected, and these occurred in just 10 samples. In contrast, in Tokinsko-Stanovoy National Park (site 15), where wild reindeer are also present, five helminth kinds were detected in 10 samples, with sample prevalence reaching 50 percent, while the local reindeer population there is estimated at 700–800 individuals.
5. Conclusions
In our coprological survey conducted to detect helminths of reindeer in the southern part of their Palearctic range, trematodes of the superfamily Paramphistomoidea were found, along with cestodes of the genus Moniezia; nematodes identified as E. rangiferi, O. macrotis, and the dimorphic species O. gruehneri/O. arctica; as well as members of the genera Nematodirus and Capillaria, and small strongylids that were not identified to genus. All helminths detected in this study have been documented previously for R. tarandus and for the Palearctic region itself, with the exception of Nematodirus sp. for southern part of the area. However, examinations of helminths in wild reindeer from Vasyugansky State Nature Reserve (Novosibirsk Oblast, Russia), Altai State Nature Reserve (Altai Republic, Russia), Baikal State Nature Reserve (Buryatia, Russia), Irkutsk Oblast (Russia), Amur Oblast (Russia), as well as in semiwild reindeer from Zabaykalsky Krai (Russia), Mongolia, and Inner Mongolia (China), were carried out for the first time.
The southern limits of the known ranges were established for E. rangiferi (51.35080 N, 121.50146 E; China) and O. macrotis (55.44280 N, 130.48006 E; Russia).
It has been suggested that O. macrotis could be used as a biogeographical marker for wild reindeer in Eastern Siberia, while the genus Capillaria could serve as a biological marker for domestic reindeer in the Palearctic.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org; Table S1: Fecal Tests; Table S2: Statistics.
Author Contributions
Conceptualization, O.A.L. and L.M.B.; methodology, O.A.L. and S.E.S.; software, O.A.L.; validation, L.M.B. and S.E.S.; formal analysis, O.A.L.; investigation, O.A.L. and S.E.S.; resources, O.A.L., A.V.S., A.V.K., V.V.K., S.B.R., Y.N.K., M.A.Z., E.V.L., V.I.A., S.M.Z., and S.A.P.; data curation, O.A.L.; writing—original draft preparation, O.A.L.; writing—review and editing, O.A.L., L.M.B., A.V.S., A.V.K., V.V.K., S.B.R., Y.N.K., M.A.Z., E.V.L., V.I.A., S.M.Z., S.A.P., and S.E.S.; visualization, O.A.L.; supervision, L.M.B.; project administration, S.E.S.; funding acquisition, S.E.S. and S.A.P. All authors have read and agreed to the published version of the manuscript.
Funding
This study was mainly performed under state order (project FFER-2024-0027; 1023032000082-1-1.6.12) for IEE RAS. Partly it was performed under state order (project FMWZ-2025-0002; 125030703344-8) for WPI RAS, as well as during the preparation of the Chronicle of Nature of the Zeisky Nature Reserve and the Tokinsko-Stanovoy National Park.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original analyzed and generated data presented in the study are openly available as Supplementary Materials.
Acknowledgments
Authors are deeply grateful to the administrations of the Altai, Baikal, Vasyugansky, and Zeisky State Nature Reserves, and the Tokinsko-Stanovoy National Park for their administrative and technical support. We would like to thank Veterinary service of the Yamalo-Nenets Autonomous Okrug (Salekhard, Russia) and Evgeniy Popov personally for their assistance. We also extend our gratitude to reindeer herders of all participated countries for their cooperation. During the preparation of this manuscript, the author used the GPT-5.1 family of large language models (LLM) for the purposes of superficial text editing (e.g., grammar, spelling, punctuation, and formatting). The authors have reviewed and edited the output and take full responsibility for the content of this publication.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| DNA | Deoxyribonucleic Acid |
| DSL | Dorsal-Spined Larva |
| E | East |
| ID | Identifier |
| IEE RAS | A. N. Severtsov Institute of Ecology and Evolution of the Russian Academy of Sciences |
| ITS | Internal Transcribed Spacer |
| GPT | Generative Pre-trained Transformer |
| L1 | First stage nematode larva |
| L3 | Third stage nematode larva |
| LLM | Large Language Model |
| N | North |
| NCBI | National Center for Biotechnology Information |
| PCR | Polymerase Chain Reaction |
| USA | United States of America |
| USSR | Union of Soviet Socialist Republics |
| WPI RAS | Water Problems Institute of the Russian Academy of Sciences |
References
- Kaltenborn, B.P.; Andersen, O.; Gundersen, V. The role of wild reindeer as a flagship species in new management models in Norway. Nor. J. Geogr. 2014, 68, 168–177. [Google Scholar] [CrossRef]
- Mysterud, A.; Tveraa, T.; Hansen, B.B.; Gundersen, V.; Tømmervik, H.; Erlandsson, R.; Røed, K.H; Våge, J.; Andersen, R.; Brænd, E.; Bøthun, S.W.; Elgaaen, M.; Holand, Ø.; Kjørstad, M.; Kvie, K.; Mossing, A.; Myren, I.S.; Panzacchi, M.; Peeters, B.; Punsvik, T.; Romtveit, L.; Skarin, A.; Moorter, B.V.; Veiberg, V.; Jaren, V.; Strand, O.; Rolandsen, C.M. A quality standard for conservation of wild reindeer. Wildl. Monogr. 2025, 219, e70005. [Google Scholar] [CrossRef]
- Gunn, A. Rangifer tarandus. The IUCN Red List of Threatened Species 2016 2016, e.T29742A22167140. [Google Scholar] [CrossRef]
- Panchenko, D.V.; Mizin, I.A.; Rozhnov, V.V. Severnyy olen’ Rangifer tarandus Linnaeus, 1758 (Reindeer Rangifer tarandus Linnaeus, 1758). In Krasnaya kniga Rossiyskoy Federatsii. Zhivotnyye (Red Data Book of the Russian Federation. Animals), (in Russian), 2nd ed.; Pavlov, D.S., Ed.; FGBU “VNII Ekologiya”: Moscow, Russia, 2021; pp. 1020–1025. ISBN 978-5-6047425-0-1. [Google Scholar]
- Stavridis, M.A.; Røed, S.B.; Hansen, B.B.; Mikkelsen, Ø.; Ciesielski, T.M.; Jenssen, B.M. Tracing the footprints of Arctic pollution: Spatial variations in toxic and essential elements in Svalbard reindeer (Rangifer tarandus platyrhynchus) faeces. Sci. Total Environ. 2024, 906, 167562. [Google Scholar] [CrossRef] [PubMed]
- Reindeer and Caribou. Health and Disease; Tryland, M., Kutz, S.J., Eds.; CRC Press (Taylor and Francis Group): Boca Raton, FL, USA; London, UK; New York, NY, USA, 2019; p. 533. ISBN 978-1-4822-5068-8. [Google Scholar]
- Klein, D.R. Conflicts between domestic reindeer and their wild counterparts: a review of Eurasian and North American experience. Arctic 1980, 33, 739–756. Available online: https://www.jstor.org/stable/40509079. [CrossRef]
- Guillou, S. Le Renne: Habitant des régions froides (Reindeer: Inhabitant of cold regions); (In French). Éd. Atlas jeunesse: Paris, France, 2005; p. 29. ISBN 978-2-7312-3308-7. [Google Scholar]
- Kurtén, B. Pleistocene mammals in Europe; Weidenfeld and Nicolson: London, UK, 1968; p. 317. ISSN 0345-0074. [Google Scholar]
- Kurtén, B.; Anderson, E. Pleistocene mammals in North America; Columbia University Press: New York, USA, 1980; p. 442. ISBN 0-231-03733-3. [Google Scholar]
- Baskin, L.; Danell, K. Ecology of Ungulates: A Handbook of Species in Eastern Europe and Northern and Central Asia; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2003; pp. 378–385. ISBN 3-540-43804-1. [Google Scholar]
- Zaplatin, M.A. Chara: With a movie camera across Transbaikalia (Chara: S kinoapparatom po Zabaykal’yu); (in Russian). Mysl’: Moscow, USSR, 1964; p. 142. [Google Scholar]
- Smirnov, M.N. Severnyy olen’ na yuge Sibiri (Reindeer in the South of Siberia); (in Russian). Siberian Federal University: Krasnoyarsk, Russia, 2016; p. 230. ISBN 978-5-7638-3461-1. [Google Scholar]
- Mizkewitsch, V.Y. Gel’minty Severnogo Olenya i Vyzyvayemyye Imi Zabolevaniya (Reindeer Helminths and the Diseases They Cause); 308p. (In Russian). Kolos: Leningrad, USSR, 1967. [Google Scholar]
- Aleuy, O.A.; Serrano, E.; Ruckstuhl, K.E.; Hoberg, E.P.; Kutz, S. Parasite intensity drives fetal development and sex allocation in a wild ungulate. Sci. Rep. 2020, 10, 15626. [Google Scholar] [CrossRef] [PubMed]
- Hudson, P.J.; Dobson, A.P.; Lafferty, K.D. Is a healthy ecosystem one that is rich in parasites? Trends Ecol. Evol. 2006, 21, 381–385. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, K.; Abaunza, P. Parasites as Biological Tags for Stock Discrimination of Marine Fish: a Guide to Procedures and Methods. Fish. Res. 1998, 38, 45–56. [Google Scholar] [CrossRef]
- Machulsky, S.N. Dikiye parnokopytnyye kak rezerventy gel’mintoznykh zabolevaniy dlya sel’skokhozyaystvennykh zhivotnykh Buryat-Mongol’skoy ASSR (Wild ungulates as reservoirs of Helminthic Diseases for Farm Animals of the Buryat-Mongolian ASSR). In Proceedings of the Buryat-Mongolian Zoological Veterinary Institute; (In Russian). BurGiz: Ulan-Ude, USSR, 1955; Volume 9, pp. 163–172. [Google Scholar]
- Sulimov, A.D. Gel’minty severnykh oleney Tuvy (Helminths of Reindeer in Tuva). In Proceedings of the Omsk Veterinary Institute; (In Russian). Zapadno-Sibirskoye knizhnoye Izdatel’stvo: Omsk, USSR, 1966; Volume 24, pp. 135–141. [Google Scholar]
- Sulimov, A.D. K gel’mintofaune dikikh zhvachnykh Tuvy (To the Helminth Fauna of Wild Ruminants of Tuva). In Proceedings of the Omsk Veterinary Institute; (In Russian). Zapadno-Sibirskoye knizhnoye izdatel’stvo: Omsk, USSR, 1968; Volume 25, pp. 238–242. [Google Scholar]
- Kadenatsi, A.N. Gel’minty severnogo olenya v Khabarovskom kray (Helminths of Reindeer in Khabarovsk Krai). In Proceedings of the All-Union Scientific Conference dedicated to the 90th anniversary of the Kazan Veterinary Institute; (In Russian). KVI: Kazan’, USSR, 1963; pp. 146–147. [Google Scholar]
- Zhaltsanova, D.S.D. Obnaruzheniye Fischoederius elongatus (Poirier, 1883) v Buryatskoy ASSR (Detection of Fischoederius elongatus (Poirier, 1883) in the Buryat ASSR). Epizootologiya, epidemiologiya i profilaktika gel’mintozov (In Russian). 1972, 24, 49–50. [Google Scholar]
- Loginova, O.A. Gel’minty severnykh oleney Kazakhstana (Helminths of Reindeer in Kazakhstan). In Proceedings of the ХVII All-Russian Scientific and Practical Conference “Modern Problems of General and Applied Parasitology”, (In Russian). Voronezh, Russia, 17–18 October 2024. [Google Scholar] [CrossRef]
- Loginova, O.A.; Sipko, T.P.; Hernandez-Blanco, J.A.; Plotnikova, Iu.K. Gel’mintokoproskopiya dikogo severnogo olenya i ovtsebyka v Magadanskoy oblasti (Helminthocoproscopy of Wild Reindeer and Muskoxen in the Magadan Region). In Proceedings of the International Scientific Conference “Biological problems of the North”, (In Russian). Magadan, Russia, 07–11 October 2024. [Google Scholar] [CrossRef]
- National Standard of the Russian Federation GOST R 54627-2011 Zhivotnyye sel’skokhozyaystvennyye zhvachnyye. Metody laboratornoy diagnostiki gel’mintozov (Agricultural Ruminant Animals. Methods of Laboratory Helminthological Diagnostics); Standartinform. 20p. (in Russian). Moscow, Russia, 2013.
- Vajda, T. Gyors eljárás a juhok tüdőférgességének megállapítására (Rapid method for diagnosing lung helminths in sheep). Állatorvosi Lapok (In Hungarian). 1931, 54, 187–190. [Google Scholar]
- Darling, S.T.; Barber, M.A.; Hacker, H.P. Hookworm and Malaria. Research in Malaya, Java, and the Fiji Islands (Report of Uncinariasis Commission to the Orient 1915–1917); The Rockefeller Foundation International Health Board: New York, USA, 1920; p. 25. [Google Scholar]
- Skrjabin, K.I. Trematody Zhivotnykh i Cheloveka. Osnovy Trematodologii (Trematodes of Animals and Man. Principles of Trematodology); (In Russian). Izdatelstvo Akademii Nauk SSSR: Moscow, USSR, 1949; Volume 3, p. 623. [Google Scholar]
- Spassky, A.A. Osnovy Tsestodologii. Anoplotsefalyaty – lentochnyye gel’minty domashnikh i dikikh zhivotnykh (Fundamentals of Cestodology. Anoplocephalates – tapeworms of domestic and wild animals); (In Russian). Izdatelstvo Akademii Nauk SSSR: Moscow, USSR, 1951; Volume 1, p. 735. [Google Scholar]
- Skrjabin, K.I.; Shikhobalova, N.P.; Schulz, R.S. Trikhostrongilidy Zhivotnykh i Cheloveka. Osnovy Nematodologii (Trichostrongylides of Animals and Man. Principles of Nematodology); (In Russian). Izdatelstvo Akademii Nauk SSSR: Moscow, USSR, 1954; Volume 3, p. 683. [Google Scholar]
- Boev, S.N. Prostrongilidy. Osnovy Nematodologii (Protostrongylides. Principles of Nematodology); (In Russian). Nauka SSSR: Moscow, USSR, 1975; Volume 25, p. 267. [Google Scholar]
- Kontrimavichus, V.L.; Delyamure, S.L.; Boev, S.N. Metastrongiloidei Zhivotnykh i Cheloveka. Osnovy Nematodologii (Metastrongyliodeas of Animals and Man. Principles of Nematodology); (In Russian). Nauka SSSR: Moscow, USSR, 1976; Volume 26, p. 239. [Google Scholar]
- Skrjabin, K.I.; Shikhobalova, N.P.; Orloff, I.W. Trikhotsefalidy i Kapillyariidy Zhivotnykh i Cheloveka i Vyzyvaemye Imi Zabolevaniya. Osnovy Nematodologii (Trichocephalides and Capillariides of Animals and Man and the Diseases They Cause. Principles of Nematodology); (In Russian). Izdatelstvo Akademii Nauk SSSR: Moscow, USSR, 1957; Volume 6, p. 587. [Google Scholar]
- Gasser, R.B.; Chilton, N.B.; Hoste, H.; Beveridge, I. Rapid sequencing of rDNA from single worms and eggs of parasitic helminths. Nucleic Acids Res. 1993, 21, 2525–2526. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A greedy algorithm for aligning DNA sequences. J. Comput. Biol. 2004, 7, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Morgulis, A.; Coulouris, G.; Raytselis, Y.; Madden, T.L.; Agarwala, R.; Schäffer, A.A. Database indexing for production MegaBLAST searches. Bioinformatics 2008, 24, 1757–1764. [Google Scholar] [CrossRef] [PubMed]
- Blaker, H. Confidence curves and improved exact confidence intervals for discrete distributions. Can. J. Stat. 2000, 28, 783–798. [Google Scholar] [CrossRef]
- Skrjabin, K.I. Glistnyye invazii severnogo olenya (Worm infestations in reindeer); (In Russian). Sel’khozgiz: Moscow and Leningrad, USSR, 1930; p. 88. [Google Scholar]
- Skrjabin, K.I. Metod polnykh gel’mintologicheskikh vskrytiy pozvonochnykh, vklyuchaya cheloveka. (Method of complete helminthological dissections of vertebrates, including humans.); (In Russian). Izdaniye 1-go Moskovskogo gosudarstvennogo Universiteta: Moscow, USSR, 1928; p. 45. [Google Scholar]
- Rehbein, S.; Vymyslická, P.J.; Peterka, T.; Strube, C.; Visser, M.; Mayr, S.; Lackerschmid, J. Calicophoron daubneyi (Paramphistomidae) in deer of the Šumava National Park, Czech Republic – Consequence of Prevalent Rumen Fluke Infection in Cattle. Vet. Parasitol.: Reg. St. Rep. 2024, 50, art. 101012. [Google Scholar] [CrossRef] [PubMed]
- Eduardo, S.L. The taxonomy of the family Paramphistomidae Fischoeder, 1901 with special reference to the morphology of species occurring in ruminants. I. General considerations. Syst. Parasitol. 1982, 4, 7–57. [Google Scholar] [CrossRef]
- Haukisalmi, V.; Laaksonen, S.; Oksanen, A.; Beckmen, K.; Halajian, A.; Yanagida, T.; Nakao, M. Molecular taxonomy and subgeneric classification of tapeworms of the genus Moniezia Blanchard, 1891 (Cestoda, Anoplocephalidae) in northern cervids (Alces and Rangifer). Parasitol. Int. 2018, 67, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Dróżdż, J. Polymorphism in the Ostertagiinae Lopez-Neyra, 1947 and Comments on the Systematics of These Nematodes. Syst. Parasitol. 1995, 32, 91–99. [Google Scholar] [CrossRef]
- Loginova, O.A.; Rozenfeld, S.B.; Sipko, T.P.; Mizin, I.A.; Panchenko, D.V.; Laishev, K.A.; Bondar, M.G.; Kolpashchikov, L.A.; Gruzdev, A.R.; Kulemeev, P.S.; et al. Diversity and Distribution of Helminths in Wild Ruminants of the Russian Arctic: Reindeer (Rangifer tarandus), Muskoxen (Ovibos moschatus), and Snow Sheep (Ovis nivicola). Diversity 2023, 15, 672. [Google Scholar] [CrossRef]
- Loginova, O.A.; Kolpashchikov, L.A.; Spiridonov, S.E. First report of Orthostrongylus sp. (Nematoda: Protostrongylidae) in wild reindeer (Rangifer tarandus) from the Taimyr, Russia: Nearctic parasites in a Palearctic host. Parasitol. Res. 2023, 122, 685–689. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Map indicating the sampling sites. Numbers correspond to identification numbers in Table 1. Sampling area is marked with a red rectangle on the world map in the insert.
Figure 1.
Map indicating the sampling sites. Numbers correspond to identification numbers in Table 1. Sampling area is marked with a red rectangle on the world map in the insert.
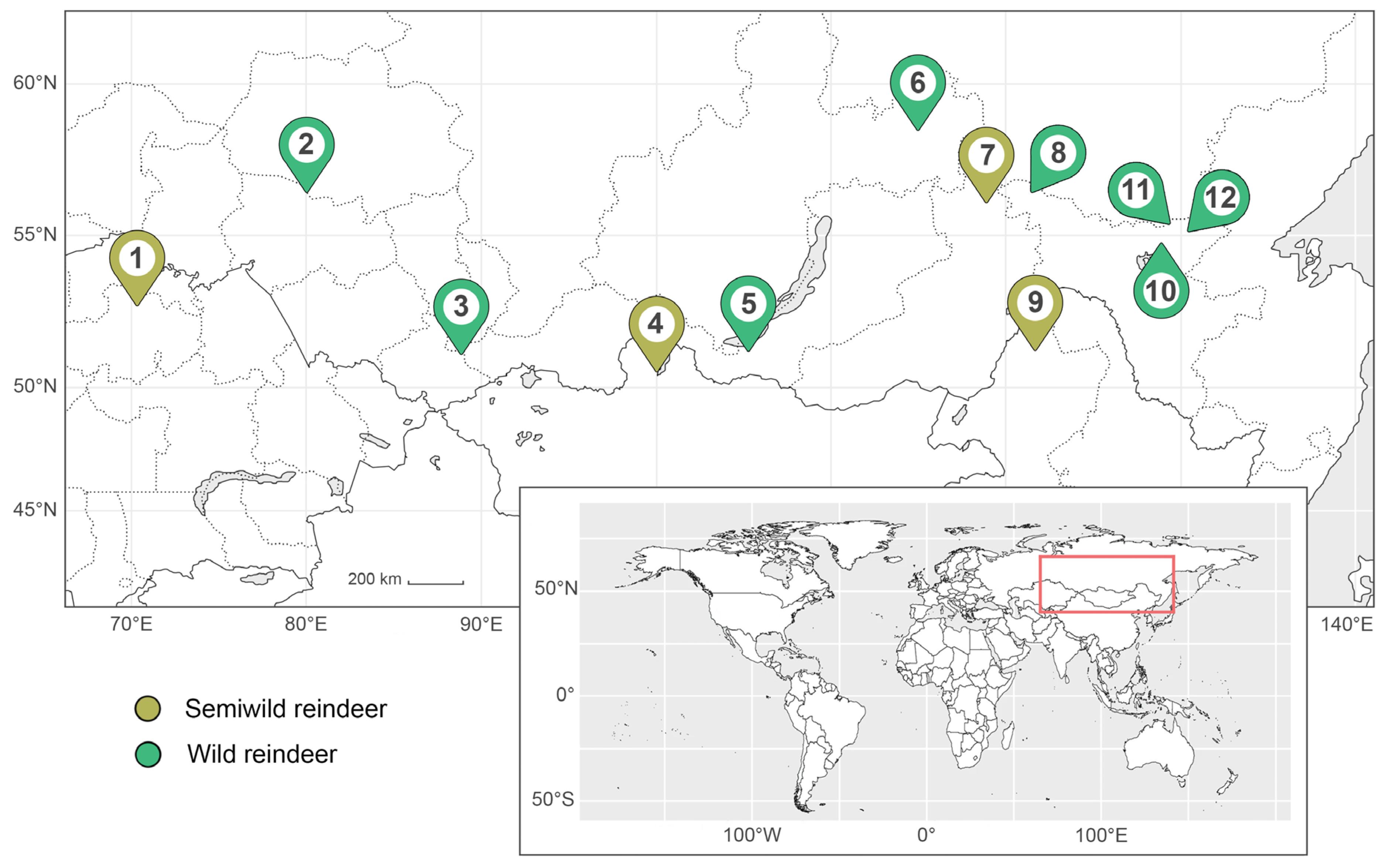
Figure 2.
Appearance of the helminths found in reindeer feces. (A) Paramphistomoidea sp. egg; (B) Moniezia sp. egg; (C) Capillaria sp. egg; (D) Strongyle-type egg; (E) Nematodirus sp. egg; (F) Elaphostrongylus rangiferi L1; (G) Orthostrongylus macrotis L1; (H) Ostertagia gruehneri L3 (dead). Bright field microscopy. A to G magnification: 400×, H: 200×. A, E, F, G, H: helminths in water; B, C, D: helminths in Darling’ solution. Cover slips used. No staining added. Scale bars equal 50 μm.
Figure 2.
Appearance of the helminths found in reindeer feces. (A) Paramphistomoidea sp. egg; (B) Moniezia sp. egg; (C) Capillaria sp. egg; (D) Strongyle-type egg; (E) Nematodirus sp. egg; (F) Elaphostrongylus rangiferi L1; (G) Orthostrongylus macrotis L1; (H) Ostertagia gruehneri L3 (dead). Bright field microscopy. A to G magnification: 400×, H: 200×. A, E, F, G, H: helminths in water; B, C, D: helminths in Darling’ solution. Cover slips used. No staining added. Scale bars equal 50 μm.
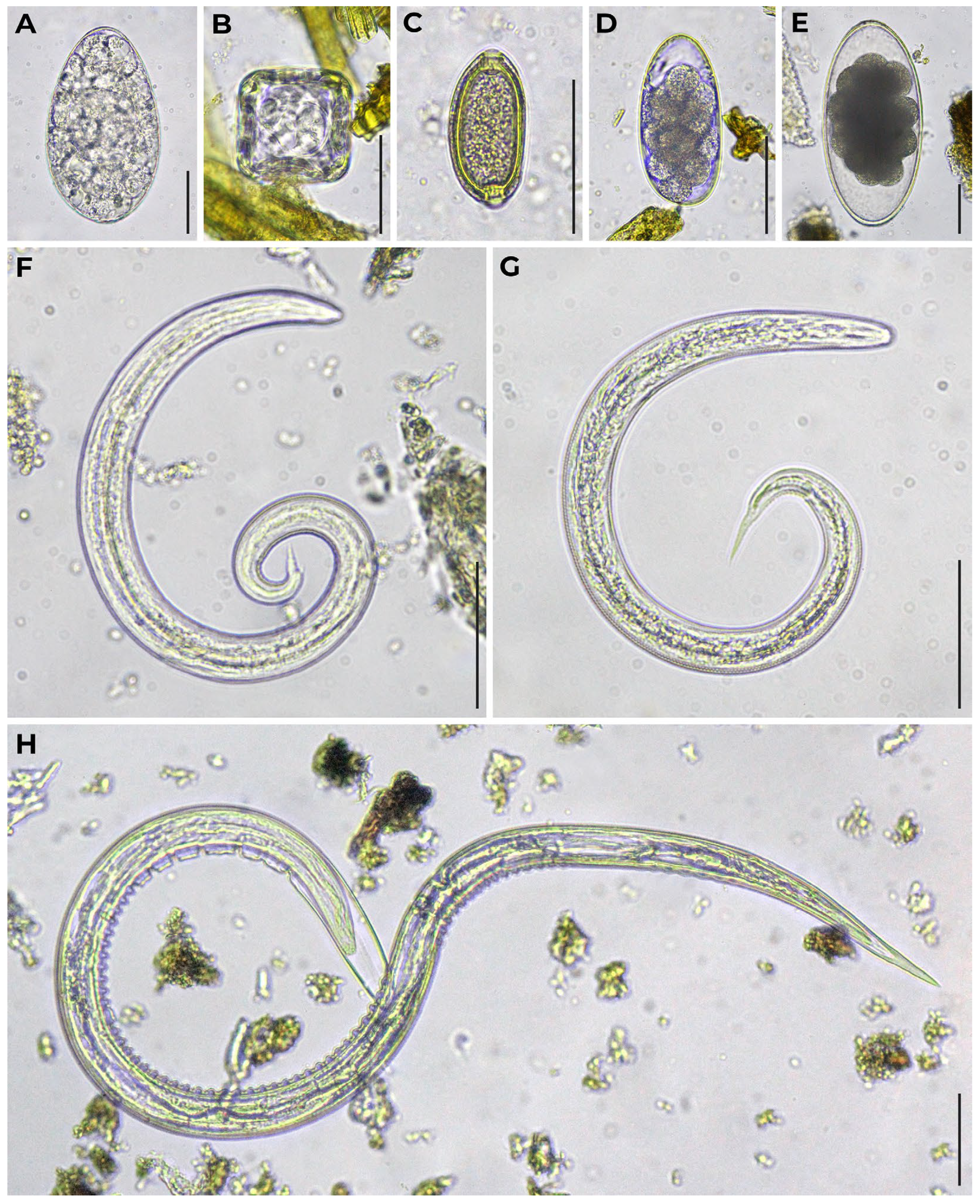
Figure 3.
Comparative distribution of different helminths among sites corresponding to the wild and semiwild reindeer habitat.
Figure 3.
Comparative distribution of different helminths among sites corresponding to the wild and semiwild reindeer habitat.
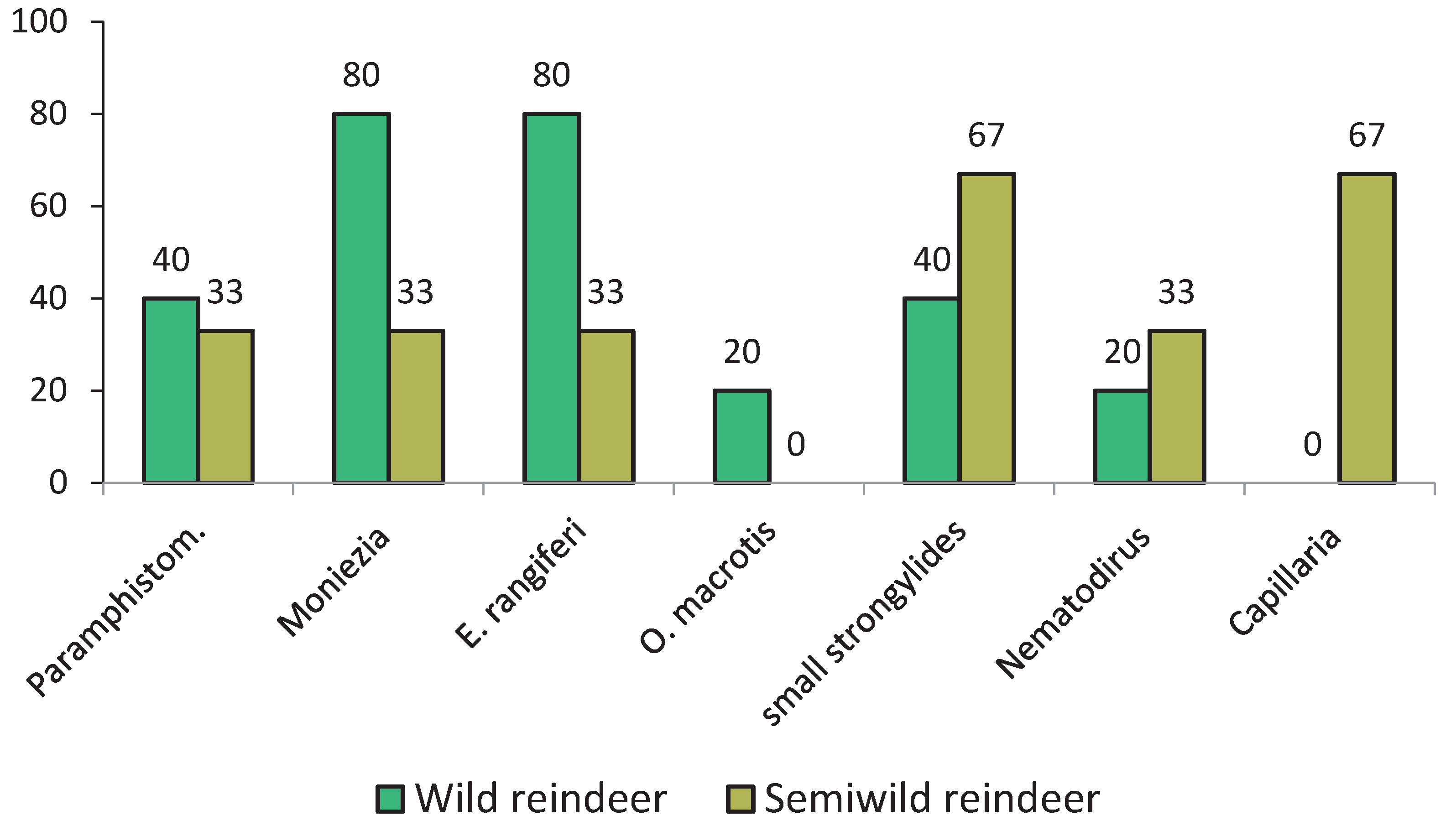

Table 1.
Collection data for fecal samples from reindeer (Rangifer tarandus) across the Southern continental Palearctic (sampling sites arranged from west to east).
Table 1.
Collection data for fecal samples from reindeer (Rangifer tarandus) across the Southern continental Palearctic (sampling sites arranged from west to east).
| Sample Set ID | Map Reference |
Type of Reindeer | Number of Fecal Samples |
Location (Country, Administrative Unit) |
Protected Area |
Coordinates (Decimal Degrees) | Altitude (Meters) |
Date Collected |
|---|---|---|---|---|---|---|---|---|
| 1 | 1 | semiwild | 2 | Kazakhstan, Akmola Region |
— | 52.93877 70.24527 |
450 | April 2025 |
| 2 | 2 | wild | 8 | Russia, Novosibirsk Oblast |
Vasyugansky State Nature Reserve | 56.636553 79.981242 |
129 | February 2025 |
| 3 | 3 | wild | 3 | Russia, Altai Republic |
Altai State Nature Reserve |
51.38385 88.98008 |
1671 | February 2024 |
| 4 | 4 | semiwild | 20 | Mongolia, Khövsgöl Province |
— | 50.566855 100.127792 |
1915 | August 2023 |
| 5 | 5 | wild | 20 | Russia, Buryatia |
Baikal State Nature Reserve |
51.30777 105.24722 |
1370 | February 2012 |
| 6 | 5 | wild | 35 | Russia, Buryatia |
Baikal State Nature Reserve |
51.30777 105.24722 |
1370 | July 2023 |
| 7 | 5 | wild | 35 | Russia, Buryatia |
Baikal State Nature Reserve |
51.30777 105.24722 |
1370 | July 2024 |
| 8 | 5 | wild | 35 | Russia, Buryatia |
Baikal State Nature Reserve |
51.30777 105.24722 |
1370 | July 2025 |
| 9 | 6 | wild | 1 | Russia, Irkutsk Oblast |
— | 58.609750 114.980806 |
884 | September 2021 |
| 10 | 7 | semiwild | 30 | Russia, Zabaykalsky Krai |
— | 56.579214 118.886167 |
882 | February 2024 |
| 11 | 8 | wild | 7 | Russia, Amur Oblast |
— | 56.73530 121.50033 |
1396 | February 2023 |
| 12 | 9 | semiwild | 10 | China, Inner Mongolia |
— | 51.35080 121.50146 |
788 | August 2023 |
| 13 | 10 | wild | 17 | Russia, Amur Oblast |
— | 55.142893 128.850543 |
653 | February 2023 |
| 14 | 11 | wild | 9 | Russia, Amur Oblast |
— | 55.679389 129.329878 | 1064 | February 2023 |
| 15 | 12 | wild | 10 | Russia, Amur Oblast |
Tokinsko-Stanovoy National Park | 55.44280 130.48006 |
1518 | July 2021 |
Table 2.
Parasitic nematode species detected in reindeer feces and identified by their ITS region.
| Sample Set ID | Species | GenBank 1 | Voucher 2 |
|---|---|---|---|
| 12 | Elaphostrongylus rangiferi | PX720411 | IPEE_Parasites 14460 |
| 15 | Orthostrongylus macrotis | PX720450 | IPEE_Parasites 14459 |
| 15 | Ostertagia gruehneri | PX716654 | IPEE_Parasites 14458 |
1 GenBank accession numbers are provided for sequences obtained from individual larvae at the first (L1) or third (L3) developmental stage, each representing its respective species. 2 Voucher specimens with definitive identifications and accession numbers are archived in the Museum of Helminthological Collections, Parasitology Center, IEE RAS (Moscow, Russia).
Table 3.
Helminths found in reindeer feces. Sample prevalence: absolute number (ratio, %); sample intensity: mean ± standard error of the mean.
Table 3.
Helminths found in reindeer feces. Sample prevalence: absolute number (ratio, %); sample intensity: mean ± standard error of the mean.
| Map Site | Trematodes | Cestodes | Nematodes | ||||
|---|---|---|---|---|---|---|---|
| Paramphistomoidea | Moniezia | Strongyle-type | Nematodirus | Elaphostrongylus rangiferi | Orthostrongylus macrotis | Capillaria | |
| 1 | – | – | – | – | – | – | – |
| 2 | – | 5 (63%); 1.2±0.3 | 1(13%); 0.3±0.0 | – | 6 (75%); 0.6±0.1 | – | – |
| 3 | – | – | – | – | – | – | – |
| 4 | 2 (10%); 0.5±0.2 | 1 (5%); 0.3±0.0 | 9 (45%); 1.0±0.4 | – | – | – | 2 (10%); 0.3±0.0 |
| 5 | 2 (2%); 0.3±0.0 | 7 (6%); 0.6±0.1 | – | – | 1 (1%); 2.0±0.0 | – | – |
| 6 | – | – | – | – | – | – | – |
| 7 | – | – | – | 3 (10%); 0.6±0.3 | – | – | – |
| 8 | – | 1 (14%); 0.3±0.0 | – | – | 3 (43%); 1.0±0.3 | – | – |
| 9 | – | – | 3 (30%); 1.1±0.5 | – | 2 (20%); 36.2±20.3 | – | 1(10%); 0.7±0.0 |
| 10 | – | 5 (29%); 0.7±1.2 | – | – | – | – | – |
| 11 | – | – | – | – | – | – | – |
| 12 | 3 (30%); 0.6±0.1 | – | 5 (50%)*; 2.6±1.2 | 1 (10%); 0.3±0.0 | 1 (10%); 1.3±0.0 | 1 (10%); 4.0±0.0 | – |
* Including Ostertagia gruehneri.
Table 4.
Sample intensity of invasion according to GOST R 54627-2011.
| Helminths | Intensity of Invasion Depending on the Number of Found Helminth Eggs and Larvae, Specimens per 1 g of Feces | |||
|---|---|---|---|---|
| Low | Medium | High | Very High | |
| Nematodes, Cestodes | 1–100 | 101–500 1 | 501–1000 | >1000 |
| GINs (eggs + L3s) 2 | 0.3–6.7 (sites 2, 4 ,9, 12) | – | – | – |
| Nematodirus (eggs) | 0.3–1.3 (sites 7, 12) | – | – | – |
| E. rangiferi (L1s) | 0.3–75.3 (sites 2, 5, 8, 9, 12) | – | – | – |
| O. macrotis (L1s) | 4.0 (site 12) | – | – | – |
| Capillaria (eggs) | 0.3–0.7 (sites 4, 9) | – | – | – |
| Moniezia (eggs) | 0.3–2.0 (sites) | – | – | – |
| Trematodes | 1–10 | 11–100 | >100 | – |
| Paramphistomoidea (eggs) | 0.3–0.7 (sites 4, 5, 12) | – | – | – |
1 Low intensity in case of adult animals. 2 Including Ostertagia gruehneri.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



