
Submitted:
28 May 2025
Posted:
28 May 2025
You are already at the latest version
Abstract
Estimating human impact on parasites in wildlife is essential to create efficient strategies to control diseases and improve public health. This study assessed the human influence on the parasite burden of the two commonest canine mesocarnivores, the red fox (N=194) and the golden jackal (N=151), in Europe. We compared the infracommunity crowding index, parasite diversity, prevalence, and abundance of different parasite taxa, and the presence of Echinococcus multilocularis between groups of hosts, such as jackals and foxes hunted at different distances from human settlements. The analysis of hosts’ distribution ascertained that 61.2% of the investigated wild canids were hunted in a 500-1750 m ring around the human settlements. Ancylostomatidae proved the most prevalent taxon (>60%), while Echinococcus multilocularis and Dipylidium caninum occurred least frequently. The two taxa that mostly contributed to the crowding of infracommunities were Ancylostomatidae and Toxocaridae, with a negative correlation between their abundances (Spearmann's ρ=-0.445, p< 0.001). Echinococcus multilocularis infected parasite infracommunities were more crowded in both foxes (p=0.02) and jackals (p=0.001). Our study highlighted that human modification of landscapes worsened the parasite burden of wild mesocarnivores, which could increase the public health risk of zoonotic parasites.

Keywords:
helminth infracommunity
; Echinococcus multilocularis
; red fox
; golden jackal
; human impact
1. Introduction
Canine mesocarnivores of Europe, especially the red fox (Vulpes vulpes) and the golden jackal (Canis aureus) have high adaptability and can utilise the human-mediated environment, which resulted in population expansion of both species almost all over the continent [1]. The occurrence of both foxes [2] and jackals [3] in synanthropic environments can be observed in Europe frequently.
Urbanisation of wildlife changes the natural diet as a consequence of utilisation of anthropogenic food sources [3,4] which might cause alteration of gastrointestinal microbiome and thus the composition of parasitic infracommunity [5,6]. The adverse effect on intestinal bacteria and the parasitic status of the host population is confirmed by a study on coyotes in Calgary suburbs [7]. However, the anthropogenic environment has an additional influence on the parasites of urbanised wild carnivores. A higher parasite burden of the human environment, which is caused by dense domestic carnivore populations, can also affect the parasitic status of the carnivores, which regularly enter settlement areas) [2,8,9]. On the other hand, wild carnivores can harbour plenty of parasites, some with zoonotic potential, and carry these pathogens to the close vicinity of human communities [1,2,10]. Moreover, human presence decreases the home range size of urbanising mesocarnivores[11,12], therefore, a denser sympatric population might accumulate varied pathogens [2]. The mutual disadvantageous interdependence between the health of human communities and wildlife populations results in a special aspect of human-wildlife conflict, which needs a multidisciplinary approach to manage [13].
Our study aimed to investigate the diversity of the parasitic infracommunities, with a special focus on Echinococcus multilocularis, in the two most abundant European canine carnivore hosts in different human-influenced conditions. Our further goals were to estimate the potential human health risk of wild canine urbanisation and the detrimental effect of anthropogenic environment on wild carnivores' health.
2. Material and Methods
2.1. Parasitological Investigation
Between March 2022 and March 2024, carcasses of hunter-harvested red foxes and golden jackals were collected from the southwestern part of Hungary (Baranya County and Somogy County) to determine the small intestine helminth community diversity. All samples originated from animals that were killed by the legal hunt in the framework of authorised wildlife management plans, and none of them were shot for scientific purposes. Therefore, this study did not use live animals or tissue samples from live animals. The sample collection was carried out before the disposal of the carcasses of the hunted animals. Those specimens, which were carcasses of sexually mature animals (above 12 months old), in good body condition, without any visible signs of diseases or weak body condition score, entered the study. Hunters recorded the GPS coordinates and attached them to the carcasses submitted to the laboratory.
We opened the small intestines during their necropsy, and their content was placed into a plastic jar filled with lukewarm water. The gut mucosal layer was scraped two times between fingers. After 15 minutes of sedimentation, the thoroughly stirred content was decanted, and the procedure was repeated until the supernatant became transparent enough for parasite detection [14]. The remaining worms inside the sediment were collected and counted. The parasites were identified by comparing their morphological characteristics to previous descriptions [15,16,17] . The collected helminths were identified to the lowest admissible taxonomic level (species or genus). In the case of Taenia spp. and Mesocestoides spp., morphological identification of juvenile forms without mature proglottids is complicated. Therefore, these taxa were identified at the genus level. Because of similar pathological effects and trophic niche in the intestinal ecosystem, species belonging to the Ancylostomatidae and Toxocaridae genera were analysed at levels of both genus and species for a better appreciation of their role in the infracommunities.
At the infracommunity level, we determined the total parasite count; the number of parasite species, or genus (i.e. Taenia spp. and Mesocestoides spp.); and the number of parasites belonging to a species or a genus. All these data were collected into an Excel spreadsheet (Microsoft Excel 2010 version 16) for further data processing. The basic dataset was published at https://doi.org/10.5281/zenodo.15369276.
2.2. Estimation of Human Impact
To determine the potential influence of human settlements on the composition of the infracommunities, we used the shooting coordinates of the harvested animals. Based on several research data on home ranges of the two host species, various sizes are determined [18,19,20], which are also modified by human presence [11,12]. Therefore, we did not attempt to calculate a theoretical home range around the shooting point. Instead, we recorded the point's distance from the nearest human settlements. Then, we calculated the mean of the distance values. Based on the mean distance from human settlements, we sorted the specimens into a nearby group (closer than or at the average distance) and a farther group (farther than the average distance).
The direct presence of humans [2,21,22]and human activity manifested in land use [23,24,25] can affect host-parasite interactions in habitat and indirectly may change the parasite richness in host species. Based on these theories, we consider the closeness of human settlements as the strength of potential human impact on the composition of the helminth community.
The distribution of hosts around human settlements was illustrated by sorting the specimens in 250 m wide zones from settlements. We hypothesised that the wildlife managers endeavour to reduce mesocarnivore populations uniformly across the whole hunting territory; therefore, the hunting success depends on the frequency of occurrence of the potential hunting object.
2.3. Data Analysis
At the infracommunity level, we calculated the infracommunity crowding (ICr) [26]. For this purpose, we used the following equation:
where
xj = the abundance of the j-th parasite species,
S = the total number of parasite species (hereafter species richness) within the infracommunity,
N = the total parasite abundance.
For further analysis, we sorted the sampled animals into four groups by host species and distance from human settlements (F1=nearby foxes; J1=nearby jackals, F2=farther foxes; J2=farther jackals). We rated an animal into a nearby group if the distance between its shooting coordinate and the nearest point of the nearest settlement was equal to or less than the average distance for the concerned host species. If the distance exceeded this value, the sample would be in the farther group.
We aimed to investigate the factors, which can contribute to E. multilocularis infection in wild carnivores. Therefore, we formed a further grouping with E. multilocularis infected and non-infected specimens among both foxes and jackals.
For each specimen group, we determined the following indices: species richness (number of helminth taxa in each host), overall prevalence (the percentage of hosts infected with at least one helminth taxon), the prevalence of a particular parasite taxon (percentage of infected hosts infected with a particular taxon), and mean abundance (the number of parasites found in any host, either infected or non-infected) [27,28].
For the calculation of prevalence, mean abundance and comparison of these indices between different host groups, we used the QPweb online application (https://www2.univet.hu/qpweb/qp10/index.php). In the case of prevalence, we applied Sterne’s method, while for mean abundance calculation, we used the BCa method with 2000 bootstrap replications. Prevalence values between groups were compared by Fisher’s exact test. Mean abundances were compared by a 2-sample t-test with 1000 bootstrap replications [28].
By QPweb, we compared prevalence and mean abundances between groups as follows: foxes vs. jackals; nearby carnivores vs. farther carnivores; nearby foxes vs. farther foxes; nearby jackals vs. farther jackals.
The number of identified parasite taxa was determined at both infracommunity and group levels. The richness of parasite fauna in groups was compared by t-test with 9999 bootstrap replication in PAST Paleontological Statistics Software version 5 [29]. The group pairs, which were compared in PAST, were the same analysed by QPweb.
We determined the correlation between the abundance of the two most abundant parasite taxa to find out whether any competition existed between them. In this analysis, those specimens were involved that carried at least one parasite of at least one of the concerned taxa. Spearman’s rank correlation was done by the PAST 5 software.
Based on the mean ICr, we compared the groups of specimens by t-test with 9999 bootstrap replications in PAST 5 software. The compared group pairs were as follows: foxes vs. jackals; nearby carnivores vs. farther carnivores; nearby foxes vs. farther foxes; nearby jackals vs. farther jackals; E. multilocularis infected vs. non-infected carnivores; E. multilocularis infected vs. non-infected jackals; E. multilocularis infected vs. non-infected foxes.
The distribution pattern of the carnivores around the human settlements was analysed based on the shooting point distribution. We compared the mean distances from settlements that belonged to the fox and jackal groups. This comparison was carried out by t-test with 9999 bootstrap replications in PAST 5 software. On the other hand, we compared the proportion of fox and jackal specimens that belonged to the nearby groups. For this, we applied a Chi-squared test with Yates correction and Monte Carlo permutation (N=999) in the PAST 5. Moreover, a finer distribution pattern was illustrated by sorting the specimens into 250 m wide zones around human settlements. These values were represented in a column diagram in MS Excel 2010 version 16 for better visualisation.
3. Results
During the study, 345 mesocarnivores' small intestines were investigated, of which 194 and 151 originated from carcasses of foxes and jackals, respectively. The specimens of the two hosts were collected between 0 and 4730 m far from human settlements (Figure 1).
In the cases of foxes and jackals, the mean distances were 1142.5 and 1256.8 m, respectively. The statistical comparison of means showed equality (p=0.196). Based on these two average distance values, we formed the following groups: the nearby fox group (F1), the farther fox group (F2), the nearby jackal group (J1) and the farther jackal group (J2), which contained 108, 86, 88 and 63 specimens, respectively. The probability of belonging to the nearby group was similar in the two host species (p=0.661 by Fisher’s exact). The finer distribution pattern of the investigated specimens around human settlements is represented in Figure 2.
The observed helminths belonged to seven taxa in both hosts, including Alaria alata, Dipylidium caninum, Mesocestoides spp., Echinococcus multilocularis, Taenia spp., Ancylostomatidae and Toxocaridae. The overall prevalence was calculated as 91.2% (CI95%=86.4-94.7%) and 89.4% (CI95%=83.5-93.5%) in red fox and golden jackal, respectively. There was no statistical difference between the infection levels of the hosts (p=0.59). In both carnivores, the most prevalent parasite taxon was the Ancylostomatidae, which exceeded the 60% infection rate in both host species. The Ancylostomatidae taxon was dominated by Uncinaria stenocephala, while Ancylostoma caninum was rare. The lowest detectable taxa were E. multilocularis and D. caninum in both fox and jackal samples. In comparing the taxa's prevalence and mean abundance, we did not detect differences between the hosts (Table 1), except the mean abundances of the Ancylostomatidae and the prevalence of the Toxocaridae. During a finer taxonomic identification, we could detect that within Ancylostomatidae and Toxocaridae, U. stenocephala and T. canis contributed mainly to the infracommunity crowding.
The parasite species richness proved equal in foxes and jackals (p=0.671). In all other comparisons, we found statistically confirmed dissimilarities. Between nearby and farther mixed host groups, nearby and farther jackal groups, and nearby and farther fox groups, the p-value was 0.0008, 0.0094, and 0.033, respectively.
Statistical analysis of the correlation between the intensities of two important taxa viz Ancylostomatidae and Toxocaridae showed a moderate negative relationship (Spearmann's ρ=-0.445, p<0.001). Infracommunities with zero intensity of both taxa were excluded from this calculation.
For comparison of ICr, we considered those specimens, which contained at least one parasite. Therefore, we could use 177 and 134 specimens of foxes and jackals, respectively. A moderate non-significant difference (p=0.36) was seen between the two canine species regarding individual infracommunity crowding; the mean ICr in jackals was 26.71 parasite/parasite (SD=45.51), while this index was proved to be 18.14 parasite/parasite (SD=23.84) in foxes. From the viewpoint of the E. multilocularis infected predators, the value of ICr was significantly higher in both species (Table 4). The distribution of E. multilocularis infected specimens were illustrated by Figure 2.
4. Discussion
In this study, we attempted to determine the human impact on parasite communities of two European mesocarnivore species and to evaluate the potential public health risk of these animals. For this reason, we sorted the sampled carcasses by location of hunting, i. e. nearby (close to a settlement) and farther (more than the species-specific average distance from any settlements). Since the success of hunting depended on the hunters’ skills and the visibility viz occurrence of the animals, we considered the hunting pattern as a result of chance events. Presenting the number of specimens by distant zones, we could ascertain that 23% of the foxes were hunted within the first 500 m from the borders of human settlements. We could see an accumulation of both species in a nearby zone between 500 and 1750 m around the settlements. Within this ring, 56% and 68% of foxes and jackals were hunted, respectively. Based on this finding, we assume that human presence exerted a strong attractive impact on both carnivores, though this phenomenon was more remarkable in the fox species.
The prevalence of small intestinal helminthosis proved almost equal in the two host species. Moreover, most of the parasite taxa reached very similar prevalence and intensity values in both hosts. It should be noted that the mean intensity of small intestinal parasites was very low in both hosts, moreover the highest crowding were 166 and 360 in the fox and the jackal, respectively. However, the prevalence of helminthosis was high, about 90%, in both hosts. This phenomenon might be in accordance with our previous observation that clinical diseases or weak body condition scores were rare in the carnivore populations of the study site. In the background, we presume the outstanding carrying capacity of the area. Countryside abandonment provides non-disturbed habitats for these animals [30]. The change in land use and the climate contributed to the population expansion of field voles (Microtus arvalis) [31], which supported the population increase of opportunistic mesocarnivores of the area [13,32]. Another feed source for canid carnivores is hunting offal, which is available in huge amounts all year round, since our study area is the densest big game hunting area in Hungary [33]. Based on these data, the low intensity of helminthosis in adult wild canids is not an inexplicable phenomenon. In healthy animals with good body condition, most parasite taxa cannot cause conspicuous health damage [7,34]. Therefore, in a population with an advantageous parasitological status, dissimilarities between groups of host are complicated to detect.
Our study revealed a more remarkable difference in diversity, prevalence, and mean abundance of infracommunities by distance from settlements than by host species. The distance-dependent dissimilarity was caused mostly by the taxa Ancylostomatidae and Toxocaridae. In the case of foxes, both these taxa contributed equally to the variation of diversity between the F1 and F2 groups. While in jackals, Ancylostomatidae produced 40% of the dissimilarity between the nearby (J1) and the distant (J2) groups.
The importance of Ancylostomatidae was characterised by its high prevalence, whereas more than 60% of the animals were infected with these parasites. Comparing the nearby and the distant groups of hosts, we could ascertain that Ancylostomatidae proved to be more important in synanthropic environment in both fox and jackal hosts; however, jackals could harbour more parasites of this taxon. The accumulation of hookworm-positive carnivores around human settlements was observed previously in a study on foxes [35]. The dominance of Acylostomatidae above Toxocaridae in the golden jackal host was confirmed in a work in Serbia [36], while studies in Greece ascertained similar importance of the two taxa [37]. Another investigation in Iran found that Toxocaridae outnumbers Ancylostomatidae in dog, red fox, and golden jackal host species, as well [38].
Toxocaridae proved to be a more abundant parasite taxon in foxes in both types of habitats, though the distant subpopulation (F2) seemed more infected. This finding agrees with previous studies, which concluded that the red fox is the most important wildlife reservoir of Toxocaridae thus the main source of infection for pets and humans [8]. Our finding that Toxocaridae characterised rather the natural environment than the human settlements suggests that foxes pose a higher risk of toxocariasis for domesticated carnivores than vice versa.
These two taxa of intestinal helminths with direct life cycles behave partly differently in the host's body. Though both can modulate the immune response of the host, they follow dissimilar ways. Hookworms can down-regulate their own genes, which are responsible for immune suppression if they enter an immunodeficient environment. This phenotypic plasticity is a result of a long-term co-evolution with their hosts, which helps the parasite to create an optimal balance with the host's body. Thus, hookworms can be found in hosts of any age, and the worm abundance depends on the host's carrying capacity, which is determined mainly by the host's immune response ability [39]. On the other hand, roundworms mostly infect young animals. Adult hosts can rarely acquire patent infection; therefore, they cannot contribute efficiently to the egg burden of the environment [40]. Mostly, the scientific evidence supports the definite immune suppressive capability of Toxocaridae [40,41,42].
The Ancylostomatidae parasite family applies highly efficient evolutionary answers for successful transmission and survival in the hosts. These parasites can enter the host's body by both oral intake and skin penetration of infective larvae. Then, these larvae and the adult stages can effectively escape the immune defence mechanisms of the body [43]. This behaviour of hookworms gives the impression of being opportunistic throughout the host's life. However, Toxocaridae destroy the intestinal integrity, inhibit the immune response and decrease bile-acid secretion, thus reducing peristalsis by excreted metabolites [42], though these abilities can be expressed mostly in the younger ages of the host [40].
Despite the apparently dissimilar invasion strategies, Ancylostomatidae and Toxocaridae seemed to compete with each other within the infracommunities, whereas a significant negative correlation could be confirmed in our investigation between their abundance. The competitive exclusion might be influenced parallelly by both the host species and the human impact on the host. The high density of hosts, regardless of their age, supports the transmission of hookworms throughout the year. Inside a human settlement, dense populations of domestic carnivores shed huge amounts of parasites into the environment [2,9]. Jackals yet live in highly social small families of mixed age in very close physical contact [44]. In these circumstances, the transmission of parasites has a high risk.
On the other hand, foxes are solitary animals that meet their counterparts exclusively during the breeding season. However, they can acquire parasite eggs in their dens, which can harbour huge amounts of infective material. Foxes' den use differs from that of the golden jackals. While jackals occupy dens only for the pup-rearing two months [44], foxes use their dens all year round [45,46]. Those dens, which are used for pup-rearing during spring, accumulate roundworm eggs shed by the fox pups [40]. Regarding the viability of infective larvae inside the roundworm eggs, fox dens can be infinite sources of infection. This hypothesis was supported by our finding that Toxocaridae were more important taxon in foxes than in golden jackals in both types of habitats. However, in those foxes, which were harvested in the close vicinity of human settlements, Ancylostomatidae outcompeted Toxocaridae. This finding suggested that in an environment of high hookworm burden, roundworms cannot succeed in efficient transmission.
The opposite effect of Toxocaridae was not confirmed by our findings. In golden jackals, both hookworms and roundworms were more abundant around the human settlements, thus in high host density. However, the abundance of Ancylostomatidae proved to be three and two times higher than Toxocaridae under human impact and in natural circumstances, respectively. Our results suggested that the golden jackal population of the study site showed increased susceptibility to Ancylostomatidae. However, this susceptibility does not mean more severe clinical signs of the infection but higher risk of getting infected. Comparing our results to other studies, jackals of Serbia showed a similar characteristic [36], while the importance of Toxocaridae seems to increase eastward [37,38]. Although these studies were not carried out synchronously with the same sampling pattern, this phenomenon might be explained by the expansion direction of the golden jackal. The westernmost resident population of jackals, which occupied its habitat in the last two decades, lives in our study site. The direction of their migration is from east and southeast to west and northwest [4]. For this reason, we presumed that the study site possessed the most mobile resident jackal population in Europe.
In the case of wolves, Lesniak et al. observed that immigration increased the parasite diversity in the host population. That study revealed that growing population density also contributed to the per capita endoparasite species richness [34]. The same phenomenon could be confirmed in Florida, where the range expansion of the coyote population resulted in the emergence of novel parasites [47].
However, our findings confirmed that rather the vicinity of human settlements than the host species affected the composition of parasite infracommunities. Though, apart from Ancylostomatidae and Toxocaridae, all other taxa reached extremely low prevalence and intensity, a higher overall diversity could be confirmed in nearby infrapopulations. Previous studies have confirmed the disadvantageous effect of human settlements. In a carnivore community of wolves and foxes in central Italy, the researchers revealed that the predominance of hookworms characterised the synanthropic foxes, while wolves that lived in a human-influenced environment accumulated coccidia [35]. In coyotes of North America, an increased prevalence of E. multilocularis was detected due to the synanthropic environment. The authors suggested that carbohydrate-rich anthropogenic food sources altered the composition of the microbiota, thus the intestinal microenvironment, which provided beneficial conditions for parasitic helminths [7].
The excessive utilisation of anthropogenic food is a well-known phenomenon in golden jackals [3]. The switch in feeding habits changes the intestinal microbiota in this mesocarnivore, as well. These changes can alter the accessibility of monosaccharides in the chyme, the integrity of the intestinal barrier, and the efficiency of local immunity [5]. Though direct evidence has not been detected, yet a population-level correlation between human-impacted microbiota alteration and the prevalence of E. multilocularis is confirmed [7].
In our study, the low prevalence of E. multilocularis could not allow us to confirm any difference in E. multilocularis infection between either host species or habitat types. However, the pattern of the infected hosts' distribution showed that E. multilocularis infected hosts accumulated in the nearby (0-1750 m) zones of settlements, which are mostly populated with carnivores (Figure 1). In the cases of foxes and jackals, 72% and 87% of E. multilocularis infected individuals were hunted in this very close vicinity of human settlements. On the other hand, we revealed that infracommunity crowding had a significant positive effect on E. multilocularis occurrence. Although cestodes need completely degraded nutrients for survival in the intestinal ecosystem, the coexistence of other parasites seemed beneficial for them. In the background of this contradictory phenomenon, the immune modulatory effect of nematodes was assumed. Both Ancylostomatidae [39,46] and Toxocaridae [38,40] possess a level of immune suppression ability. Moreover, adult Toxocaridae helminths reduce the bile-acid production of the host [42]. Bile acids exert a strong lytic effect on the tegument of protoscoleces; thus, can impede the occupation of intestinal segments by E. multilocularis [48]. Consequently, any decrease in bile acid level might support the establishment of cestodes in an intestinal ecosystem.
The disadvantageous impact of crowded nematode infracommunities on human health is observed explicitly in low-income areas where the human larva migrans cases show remarkably high prevalence [41]. The vulnerability of low-income human communities reveals itself also in human echinococcosis. The high public health risk in less developed regions originates mainly from a lower educational level of the population [49,50], which might result in inefficient disease prevention in both humans and domestic carnivores. Considering that unmanaged nematode infection in domestic carnivores might increase the risk of E. multilocularis occurrence in the synanthropic environment thus it might indirectly multiply the public health risk of human alveolar echinococcosis.
Besides anthropogenic food sources and the high density of domestic carnivores, human-mediated landscapes have further adverse influences on ecosystem health. Direct or indirect human presence can alter the host community structure, cause disturbance that reduces host health and immunity, and disrupt ecosystem services capable of regulating pathogen transmission [9].
Since our study was based on the investigation of hunter-harvested carcasses, it could provide solely a snapshot of the parasite community. Therefore, our observations could provide indirect evidence of interactions between different parasite populations. The statistical analysis of data showed differences between the parasite infracommunities of hosts at different distances from human settlements.
In this study, we did not investigate the diet and the microbiome composition of the hosts. Therefore, we could not claim that our findings are the explicit results of different life-style of synanthropic and sylvatic mesocarnivores. However, the unambiguous dissimilarities in parasite infracommunities at different distances from settlements could be confirmed. In the lack of investigating background causes, we could solely report that those carcasses, which were harvested close to human communities, carried more diverse, more abundant infracommunities with higher prevalence than those hunted farther from humans.
The opposition of Ancylostomatidae and Toxocaridae could not be confirmed directly by the current study design. We could reveal a negative correlation between the abundances of the two parasites' populations. A limitation of this study is that we did not examine the localisation of the parasites in the small intestine. Therefore, we could not investigate whether the two species compete with each other for space and resources within the intestinal lumen. Moreover, we did not collect data on the hosts' health status in details that could indicate the function of their immune systems. For mitigation of considerable dissimilarities of hosts, we did not involve carcasses of premature, underweight, or diseased animals. However, our deduction on competition between the two taxa relies entirely on the statistical association between the summarised numbers of parasites in the infracommunities.
In the case of E. multilocularis, comprehensive population biological research regularly faces obstacles because of the low prevalence of the parasite. The low number of achievable infracommunities containing the members of E. multilocularis species very frequently impedes the statistical comparison of these infracommunities. In our study, we could compare the crowding indices of the infracommunities that contained or did not contain E. multilocularis. We could find statistically confirmed dissimilarity between them, which led us to report that crowding might influence the probability of infection by E. multilocularis. The direction of the correlation could not be determined by the applied statistical method. Nevertheless, we concluded that rather crowding should have been a risk for E. multilocularis infection and not vice versa. This hypothesis was based on the facts that the total biomass of E. multilocularis could not cause a recognisable resource limitation for the whole infracommunity; moreover, the relationship between crowding and E. multilocularis infection was independent of the number of E. multilocularis parasites in the concerned infracommunity. However, our conclusion was a result of deduction.
In this study, we aimed to determine the human impact on parasite infection of the two most abundant European mesocarnivore. Our investigations revealed that land use seemed to exert a significant influence on the diversity of parasite infracommunities in both foxes and golden jackals. The disadvantageous epidemiological effect of human presence was confirmed, though solely indirectly. The most affected parasite taxa were Ancylostomatidae and Toxocaridae, which contributed to the dissimilarities between infracommunities of the human-mediated and the rather natural habitat types. In the case of E. multilocularis, we obtained indirect evidence that human presence could contribute to the occurrence of this parasite as a consequence of nematode burden in synanthropic hosts. Our study highlighted the importance of population interactions in parasite communities, which were modified by environmental factors. However, these factors proved to be able to adjust the host-parasite interaction, as well. The complex interdependence between the host, the members of the parasite infracommunity, and the environment that affects all needs a holistic approach, which considers parasitic infracommunities rather than separate parasite species. The most relevant limitation of our study was that our findings provided indirect evidence of human impact on parasite communities; therefore, further research is needed to confirm our results directly.
CRediT authorship contribution statement
Sibusiso Moloi: conceptualisation, investigation, data curation; Ágnes Csivincsik: conceptualisation, funding acquisition, writing - original draft; Eszter Nagy: investigation, data curation, formal analysis; Tamás Tari: data curation, writing - review and editing; Tibor Halász: investigation; Klaudia Polgár: data management; Gábor Nagy: conceptualisation; data curation; investigation; writing - original draft, review and editing
Declaration of competing interests
The authors declare no competing interests.
Acknowledgements
This study was supported by the Flagship Research Groups Programme of the Hungarian University of Agriculture and Life Sciences.
Ethics declaration
The authors declare the collected animals were killed by the legal hunt in the framework of authorised wildlife management plans, and none of them was shot for scientific purposes.
Data availability
The original data presented in the study are openly available in the Zenodo repository https://doi.org/10.5281/zenodo.15369276.
References
- Veronesi, F.; Deak, G.; Diakou, A. Wild mesocarnivores as reservoirs of endoparasites causing important zoonoses and emerging bridging infections across Europe. Pathogens. 2023, 12(2), 178. [CrossRef]
- Mackenstedt, U.; Jenkins, D.; Romig, T. The role of wildlife in the transmission of parasitic zoonoses in peri-urban and urban areas. Int. J. Parasitol. Parasites Wildl. 2015, 4(1), 71-79. [CrossRef]
- Ćirović, D.; Penezić, A.; Krofel, M. Jackals as cleaners: Ecosystem services provided by a mesocarnivore in human-dominated landscapes. Biol. Conserv. 2016, 199, 51-55. [CrossRef]
- Tsunoda, H.; Saito, M.U. Variations in the trophic niches of the golden jackal Canis aureus across the Eurasian continent associated with biogeographic and anthropogenic factors. J. Vertebr. Biol. 2020, 69(4), 20056-1. [CrossRef]
- Lapid, R.; Motro, Y.; Craddock, H.; Khalfin, B.; King, R.; Bar-Gal, G.K.; Moran-Gilad, J. Fecal microbiota of the synanthropic golden jackal (Canis aureus). Anim. Microbiome. 2023, 5(1), 37. [CrossRef]
- Speer, K. A. Microbiomes mediate host−parasite interactions. Mol. Ecol. 2022, 31(7), 1925–1927. [CrossRef]
- Sugden, S.; Sanderson, D.; Ford, K.; Stein, L.Y.; St. Clair, C.C. An altered microbiome in urban coyotes mediates relationships between anthropogenic diet and poor health. Sci. Rep. 2020, 10(1), 22207. [CrossRef]
- Holland, C.V. A walk on the wild side: A review of the epidemiology of Toxocara canis and Toxocara cati in wild hosts. Int. J. Parasitol. Parasites Wildl. 2023, 22, 216-228. [CrossRef]
- Wilson, A.G.; Wilson, S.; Alavi, N.; Lapen, D.R. Human density is associated with the increased prevalence of a generalist zoonotic parasite in mammalian wildlife. Proc. Royal Soc. B. 2021, 288(1961), 20211724. https://doi.org/. [CrossRef]
- Otranto, D.; Cantacessi, C.> Dantas-Torres, F.; Brianti, E.; Pfeffer, M.; Genchi, C.; Guberti, V.; Capelli, G.; Deplazes, P. The role of wild canids and felids in spreading parasites to dogs and cats in Europe. Part II: Helminths and arthropods. Vet. Parasitol. 2015, 213(1-2), 24-37. [CrossRef]
- Šálek, M.; Drahníková, L.; Tkadlec, E. Changes in home range sizes and population densities of carnivore species along the natural to urban habitat gradient. Mamm. Rev. 2015, 45, 1–14. [CrossRef]
- O’Donnell, K.; delBarco-Trillo, J. Changes in the home range sizes of terrestrial vertebrates in response to urban disturbance: a meta-analysis, J. Urban Ecol. 2020, 6(1), juaa014. [CrossRef]
- Thompson, R.A. Parasite zoonoses and wildlife: one health, spillover and human activity. Int. J. Parasitol. 2013, 43(12-13), 1079-1088. [CrossRef]
- Hofer, S.; Gloor, S.; Müller, U.; Mathis, A.; Hegglin, D.; Deplazes, P. 2000. High prevalence of Echinococcus multilocularis in urban red foxes (Vulpes vulpes) and voles (Arvicola terrestris) in the city of Zürich, Switzerland. Parasitology. 2000, 120, 135–142. [CrossRef]
- Anderson, C.R.; Chabaud, A.G.; Willmott, S. Keys to the nematode parasites of vertebrates: Archival Volume. CABI, Publishing: Wallingford, Oxfordshire, UK, 2009; 44-69 pp, 309-324 pp.
- Gibson, D.I.; Jones, A.; Bray, R.A.; Keys to the trematoda: Volume 1. CABI Publishing: Wallingford, Oxfordshire, UK, 2002; 178-185 pp.
- Khalil, L.F.; Jones, A.; Bray, R.A. Keys to the cestode parasites of vertebrates. CABI Publishing: Wallingford, Oxfordshire, UK, 1994; 309-315 pp, 355-359 pp, 665-673 pp.
- Fenton, S.; Moorcroft, P.R.; Ćirović, D.; Lanszki, J.; Heltai, M.; Cagnacci, F.; Breck,S.; Bogdanović, N.; Pantelić, I.; Ács, K.; Ranc, N. Movement, space-use and resource preferences of European golden jackals in human-dominated landscapes: Insights from a telemetry study. Mamm. Biol. 2021, 101(5), 619–630. [CrossRef]
- Main, M.T.; Davis, R.A.; Blake, D.; Mills, H.; Doherty, T.S.; Human impact overrides bioclimatic drivers of red fox home range size globally. Divers. Distrib. 2020, 26, 1083–1092. [CrossRef]
- Torretta, E.; Dondina, O.; Delfoco, C.; Riboldi, L.; Orioli, V.; Lapini, L.; Meriggi, A. 2020. First assessment of habitat suitability and connectivity for the golden jackal in north-eastern Italy. Mammal. Biol. 2020, 100, 631–643. [CrossRef]
- Bradley, C.A.; Altizer, S. 2006. Urbanization and the ecology of wildlife diseases. Trends Ecol. Evol. 2006, 22(2), 95-102. [CrossRef]
- Gecchele, L.V.; Pedersen, A.B.; Bell, M. Fine-scale variation within urban landscapes affects marking patterns and gastrointestinal parasite diversity in red foxes. Ecol. Evol. 2020, 10(24), 13796-13809. [CrossRef]
- Chakraborty, D.; Reddy, M.; Tiwari, S.; Umapathy, G. Land use change increases wildlife parasite diversity in Anamalai Hills, Western Ghats, India. Sci. Rep. 2019, 9(1), 11975. [CrossRef]
- Gibb, R.; Redding, D.W.; Chin, K.Q.; Donnelly, C.A.; Blackburn, T.M.; Newbold, T.; Jones, K.E. Zoonotic host diversity increases in human-dominated ecosystems. Nature. 2020, 584(7821), 398-402. [CrossRef]
- Guo, F.; Bonebrake, T.C.; Gibson, L. Land-use change alters host and vector communities and may elevate disease risk. Ecohealth. 2019, 16(4), 647-658. [CrossRef]
- Ferrari, N.; Citterio, C.V.; Lanfranchi, P. 2016. Infracommunity crowding as an individual measure of interactive-isolationist degree of parasite communities: disclosing the effects of extrinsic and host factors. Parasite Vectors. 2016, 17(9), 88. [CrossRef]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak A.W. Parasitology meets ecology on its own terms: Margolis et al. revisited. J. Parasitol. 1997, 83, 575-583.
- Reiczigel, J.; Marozzi, M.; Fabian, I.; Rozsa, L. 2019. Biostatistics for parasitologists – a primer to Quantitative Parasitology. Trends Parasitol. 2019, 35(4), 277-281. [CrossRef]
- Hammer, Ø.; Harper, D.A. Paleontological data analysis, 2nd ed.; John Wiley & Sons Ltd., Chichester, West Sussex, UK, 2024; 216-262 pp.
- Martínez-Abraín, A.; Jiménez, J.; Jiménez, I.; Ferrer, X.; Llaneza, L.; Ferrer, M.; Palomero, G.; Ballesteros, F.; Galán, P.; Oro, D. (2020). Ecological consequences of human depopulation of rural areas on wildlife: A unifying perspective. Biol. Conserv. 2020, 252, 108860. [CrossRef]
- Balčiauskas, L.; Balčiauskienė, L. Habitat and body condition of small mammals in a country at mid-latitude. Land. 2024, 13(8), 1214. [CrossRef]
- Lanszki, Z.; Horváth, G.F.; Bende, Z.; Lanszki, J. Differences in the diet and trophic niche of three sympatric carnivores in a marshland. Mamm. Res. 2020, 65, 93-104. [CrossRef]
- Kemenszky, P.; Jánoska, F.; Nagy, G.; Csivincsik, Á. The golden jackal (Canis aureus) and the African swine fever pandemic: Its role is controversial but not negligible (a diet analysis study). Vet. Med. Sci. 2022, 8(1), 97-103. [CrossRef]
- Lesniak, I.; Heckmann, I.; Heitlinger, E.; Szentiks, C.A.; Nowak, C.; Harms, V.; Jarausch, A.; Reinhardt, I.; Kluth, G.; Hofer, H.; Krone, O. Population expansion and individual age affect endoparasite richness and diversity in a recolonising large carnivore population. Sci. Rep. 2017, 7(1), 41730. [CrossRef]
- Perrucci, S.; Maestrini, M.; Coppola, F.; Di Marco, M.; Rosso, A.D.; Pacini, M.I.; Zintu, P.; Felicioli, A. Gray wolf (Canis lupus italicus) and red fox (Vulpes vulpes) parasite survey in anthropized and natural areas of central Italy. Vet. Sci. 2023,10(2), 108. [CrossRef]
- Miljević, M.; Lalošević, D.; Simin, V.; Blagojević, J.; Čabrilo, B.; Čabrilo, O.B. Intestinal helminth infections in the golden jackal (Canis aureus L.) from Vojvodina: Hotspot area of multilocular echinococcosis in Serbia. Acta Vet. Hung. 2021, 69(3), 274-281. [CrossRef]
- Tsokana, C.N.; Sioutas, G.; Symeonidou, I.; Papadopoulos, E. Wildlife and parasitic infections: A One Health perspective in Greece. Curr. Res. Parasitol. Vector Borne Dis. 2024, 6, 100184. [CrossRef]
- Dalimi, A.; Sattari, A.; Motamedi, G.H.; A study on intestinal helminthes of dogs, foxes and jackals in the western part of Iran. Vet. Parasitol. 2006, 142(1-2), 129-133. [CrossRef]
- Ferguson, A.A.; Inclan-Rico, J.M.; Lu, D., Bobardt, S.D.; Hung, L.; Gouil, Q.; Baker, L.; Ritchie, M.E.; Jex, A.R.; Schwarz, E.M.; Rossi, H.L.; Nair, M.G.; Dillman, A.R.; Herbert, D.R. 2023. Hookworms dynamically respond to loss of Type 2 immune pressure. PLoS Pathog. 2023, 19(12), e1011797. [CrossRef]
- Abou-El-Naga, I.F.; Mogahed, N.M. 2023. Potential roles of Toxocara canis larval excretory secretory molecules in immunomodulation and immune evasion. Acta Trop. 2023, 238, 106784. [CrossRef]
- Amor, D.A.L.M.; Santos, L.N.; Silva, E.S.; de Santana, M.B.R.; Belitardo, E.M.M.A.; Sena, F.A.; Pontes-de-Carvalho, L.; Figueiredo, C.A.; Alcântara-Neves, N.M. 2022. Toxocara canis extract fractions promote mainly the production of Th1 and regulatory cytokines by human leukocytes in vitro. Acta Trop. 2022, 234, 106579. [CrossRef]
- Wang, N.; Sieng, S.; Liang, T.; Xu, J.; Han, Q. Intestine proteomic and metabolomic alterations in dogs infected with Toxocara canis. Acta Trop. 2024, 252, 107140. [CrossRef]
- Cassar, M.; Dagenais, D. An overview of hookworm virulence mechanisms. Curr. Trop. Med. Rep. 2023, 10(4), 295-299. [CrossRef]
- Mukherjee, A.; Kumara, H.N.; Bhupathy, S. 2018. Golden jackal’s underground shelters: natal site selection, seasonal burrowing activity and pup rearing by a cathemeral canid. Mammal Res. 2018, 63, 325-339. [CrossRef]
- Reshamwala, H.S.; Mahar, N.; Dirzo, R.; Habib, B. Successful neighbour: interactions of the generalist carnivore red fox with dogs, wolves and humans for continued survival in dynamic anthropogenic landscapes. Glob. Ecol. Conserv. 2021, 25, e01446. [CrossRef]
- Zaman, M.; Tolhurst, B.A.; Zhu, M.; Jiang, G. Den-site selection at multiple scales by the red fox (Vulpes vulpes subsp. montana) in a patchy human-dominated landscape. Glob. Ecol. Conserv. 2020, 23, e01136. [CrossRef]
- Grigione, M.M.; Burman, P.; Clavio, S.; Harper, S.J.; Manning, D.L.; Sarno, R.J. A comparative study between enteric parasites of coyotes in a protected and suburban habitat. Urban Ecosyst. 2014, 17, 1-10. [CrossRef]
- Smyth, J.D. Lysis of Echinococcus granulosus by surface-active agents in bile and the role of this phenomenon in determining host specificity in helminths. Proc. Royal Soc. B. 1962, 156(965), 553-572. [CrossRef]
- Conraths, F.J.; Probst, C.; Possenti, A.; Boufana, B.; Saulle, R.; La Torre, G.; Busani, L.; Casulli, A. Potential risk factors associated with human alveolar echinococcosis: Systematic review and meta-analysis. PLoS Negl. Trop. Dis. 2017, 11(7), e0005801. [CrossRef]
- Casulli, A. 2020. Recognising the substantial burden of neglected pandemics cystic and alveolar echinococcosis. Lancet Glob. Health. 2020, 8(4), e470-e471. [CrossRef]
Figure 1.
Distribution of the sampling points in Hungary.
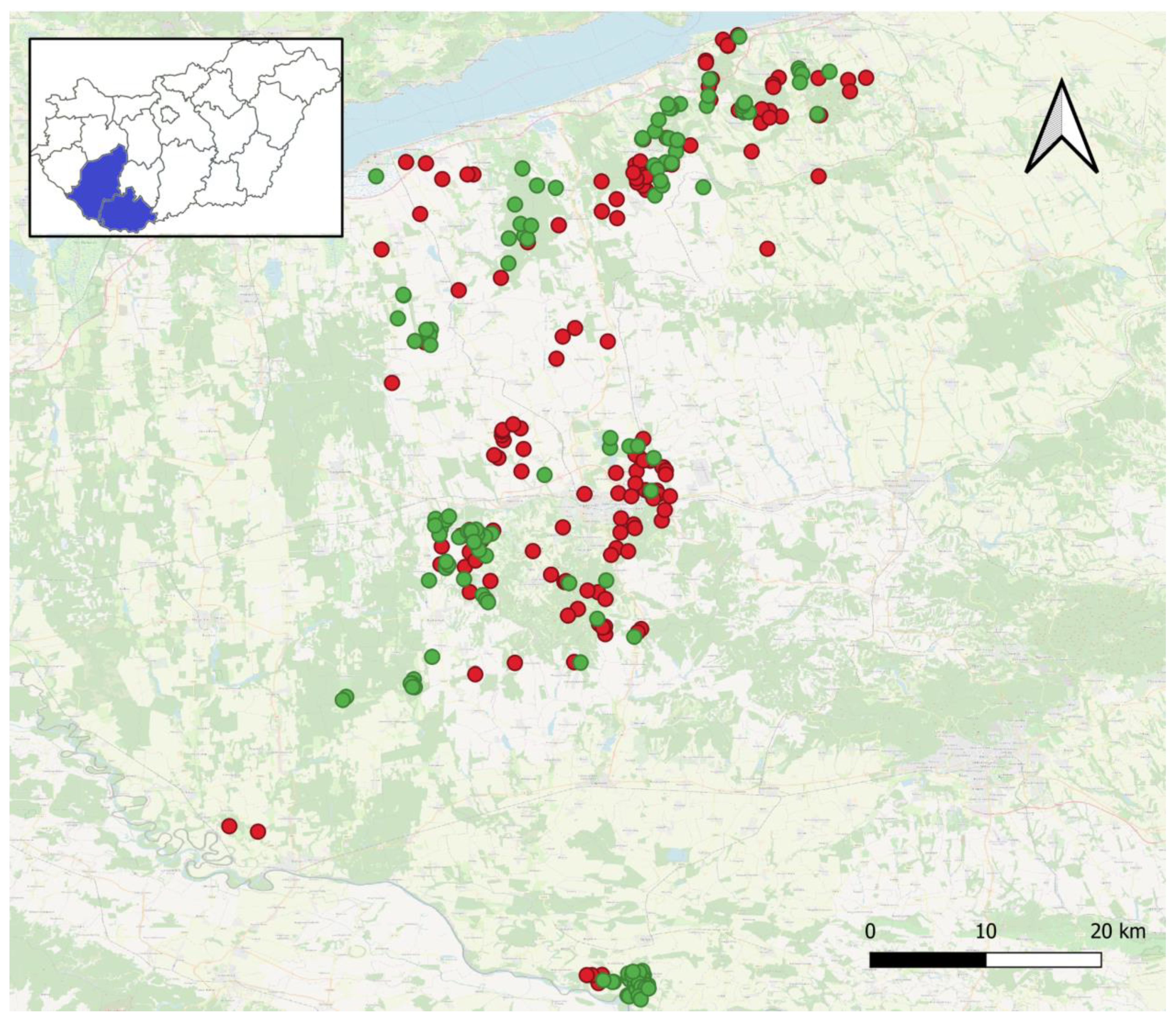
Figure 2.
Distribution of specimens and Echinococcus multilocularis infected specimens around human settlements (x-axis= the distance in m from the settlement border, y-axis= number of specimens).
Figure 2.
Distribution of specimens and Echinococcus multilocularis infected specimens around human settlements (x-axis= the distance in m from the settlement border, y-axis= number of specimens).
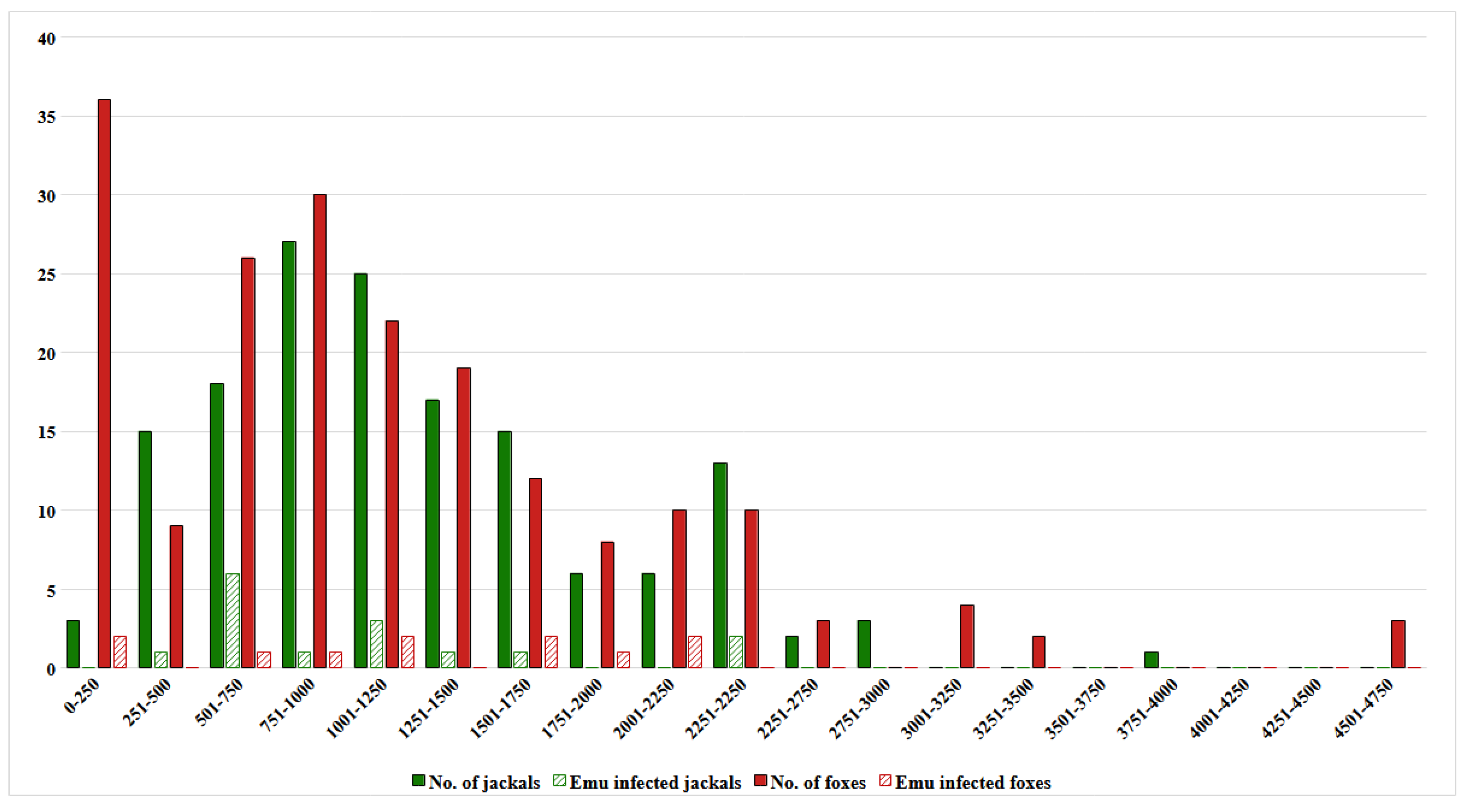

Table 1.
True prevalence and mean abundance of the detected helminth taxa in red fox and golden jackal. .
Table 1.
True prevalence and mean abundance of the detected helminth taxa in red fox and golden jackal. .
| parasite taxa | host | prevalence (%) (CI95%)* |
mean abundance (CI95%) | |
|---|---|---|---|---|
| Ancylostomatidae | fox | 61.3 (54.1-68.1) | 5.11 (3.92-6.99)** | |
| jackal | 63.6 (55.6-70.9) | 10.7 (7.8-16.7) | ||
| Uncinaria stenocephala | fox | 57.7 (50.5-64.7) | 4.88 (3.63-6.78) | |
| jackal | 51.7 (43.7-59.6) | 8.64 (5.97-15.4) | ||
| Ancylostoma caninum | fox | 4.1 (1.9-7.9) | 0.23 (0.08-0.6) | |
| jackal | 15.9 (10.8-22.8) | 2.09 (1.15-3.89) | ||
| Toxocaridae | fox | 49 (42-56.2) ** | 4.53 (3.39-6.45) | |
| jackal | 28.5 (21.8-36.4) | 3.4 (1.97-6.18) | ||
| Toxocara canis | fox | 29.4 (23.2-36.3) | 2.81 (1.9-5.24) | |
| jackal | 21.9 (15.8-29.1) | 2.64 (1.51-5.05) | ||
| Toxascaris leonina | fox | 20.1 (14.9-26.5) | 1.72 (1.07-2.72) | |
| jackal | 6.6 (3.5-11.8) | 0.76 (0.14-3.07) | ||
| Alaria alata | fox | 22.3 (16.8-28.7) | 1.63 (1.04-2.74) | |
| jackal | 20.5 (14.7-27.8) | 2.31 (1.35-4.21) | ||
| Dipylidium caninum | fox | 5.7 (3-10) | 0.13 (0.06-0.27) | |
| jackal | 7.3 (3.8-12.5) | 0.37 (0.17-0.76) | ||
| Mesocestoides spp. | fox | 29.9 (23.7-36.8) | 2.06 (1.2-3.77) | |
| jackal | 29.8 (22.8-37.7) | 1.38 (0.84-3.07) | ||
| Echinococcus multilocularis | fox | 5.7 (3-10) | 0.68 (0.31-1.47) | |
| jackal | 9.9 (5.8-15.8) | 4.44 (1.62-12.9) | ||
| Taenia spp. | fox | 26.3 (20.5-33) | 2.42 (1.27-5.17) | |
| jackal | 33.1 (25.8-41) | 1.07 (0.74-1.96) | ||
*95% confidence interval, **bold letters indicate the significant difference in prevalence or mean abundance between fox and jackal (p<0.05).
Table 4.
The infracommunity crowding index comparison between E. multilocularis infected animals in red foxes and golden jackals.
Table 4.
The infracommunity crowding index comparison between E. multilocularis infected animals in red foxes and golden jackals.
| red fox | ||
| non-infected (n=166) | 17 parasite/parasite (SD=23.41) | p=0.02 |
| infected (n=11) | 35.27 parasite/parasite (SD=24.41) | |
| golden jackal | ||
| non-infected (n=119) | 21.39 parasite/parasite (SD=33.23) | p=0.001 |
| infected (n=15) | 68.93 parasite/parasite (SD=90.66) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



