
Submitted:
15 January 2026
Posted:
16 January 2026
You are already at the latest version
Abstract
The scientific understanding of cetacean pathology has advanced significantly in recent decades. However, data concerning the health status of members of the family Kogiidae remain scarce. This study presents a comprehensive pathological assessment and determination of causes of death in 45 stranded kogiids, comprising 35 Kogia breviceps and 10 K. sima, along the coasts of the Canary Islands between 1999 and 2018. Causes of death (CD) were classified as natural (30/45; 66.6%) or anthropogenic (11/45; 24.4%), while the cause remained undetermined in 4 cases (9%). Among natural causes, the most prevalent etiologies included trauma (13/30; 43.3%), infectious (7/30; 23.3%), parasitic (5/30; 16.6%), cardiomyopathy (3/30; 10%), malnutrition (1/30; 3.3%), and fetal distress (1/30; 3.3%). Anthropogenic causes were vessel collisions (7/11; 63.6%), interactions with fishing activities (2/11; 18.2%), and foreign body-associated pathology (2/11; 18.2%). Notably, intra-interspecific traumatic interactions were overrepresented in this cohort, and variable degrees of gross and histologic cardiomyopathic changes were observed in 68.9% of individuals (31/45) without evident sex bias. Septicemia caused by Clostridium perfringens and Cl. tertium was confirmed in three cases. Additionally, cervical gill slit adenitis by Crassicauda sp. (15/45; 33%) and parasitic gastritis by Anisakis sp. (27/45; 60%), were frequent findings. These findings provide novel insights into the pathologic spectrum affecting kogiids and enhance the current understanding of their health status, with implications for diagnostic protocols, stranding response, and conservation strategies.
Keywords:
pygmy sperm whale
; dwarf sperm whale
; cetacean
; pathology
; wildlife
1. Introduction
The kogiid whales (family Kogiidae) are medium-sized odontocete cetaceans including two species: the pygmy sperm whale (Kogia breviceps) and the dwarf sperm whale (K. sima) [1,2]. These species have been allocated in the Pacific, Atlantic, and Indian Ocean waters but there is insufficient information to define the population status [3,4]. Primary dietary sources are deepwater cephalopods (e.g., Histioteuthidae and Cranchiidae families) with feed depths down to 1,200 m for K. breviceps while K. sima are thought to feed in shallower water [3,5].
Concerning anatomic singularities, both species possess a distinctive colonic diverticulum, or “ink sac”, filled with brownish red pigment which is released under stressful or life-threatening situations [3,4]. The pygmy sperm whale has a cervical-lateral exocrine gland (“false gill split”) that is commonly parasitized by Crassicauda magna [6,7,8]. Specific anatomic investigations using advanced imaging techniques have described unique features e.g., supracranial basin, spermaceti chamber, hyoid apparatus [9]. Kogia spp. are often heavily parasitized by intestinal nematodes such as Anisakis spp. and Terranova cetecola, as well as larval cestodes of Clistobothrium delphini encysted in the blubber [3]. Heavy larval cestode infestations may correlate with shark predation prevalence [3]. Although overall level of predation on Kogia spp. remains unknown, there is evidence of attacks by lampreys, great white sharks (Carcharodon carcharias), and killer whales (Orcinus orca), with remains of pygmy sperm whales found in killer whale stomachs [3].
Long-term pathology-based health monitoring of uncommon cetacean species is rare. The Kogiidae are among the least studied odontocete taxa [10], largely due to their elusive and deep-diving behavior with short surface intervals, resulting in most biologic and pathologic knowledge being derived from stranded individuals [6,10,11,12,13,14,15,16,17,18]. Despite steadily increasing anatomic and diagnostic investigations, longitudinal health assessments remain limited, emphasizing the need for studies focused on their pathology and conservation efforts.
Cetaceans are exposed to environmental stressors either anthropogenic (e.g., chemical and acoustic pollution, fisheries, maritime traffic, tourism industry) and non-anthropogenic, hereafter ‘natural’ (e.g., biotoxins, pathogens such as bacteria, fungi, parasites and viruses) [19]. In general terms, physiologic and pathologic knowledge in kogiid whales is limited and fragmentary. Notably, the pygmy sperm whale is possibly the mammal with the highest prevalence of naturally occurring dilated cardiomyopathy. The cause(s) for such prevalence remain uncertain, although derangements in selenium and metallothionein have been argued [17,18,20]. Here we assessed retrospectively the epidemiology and pathologic findings and causes of death of K. breviceps and K. sima stranded in the Canary Islands between 1999 and 2018.
2. Materials and Methods
2.1. Stranding Epidemiologic Data and Autopsy Examination
The required permission for the management of stranded cetaceans anywhere within the Canarian archipelago was issued by the General Subdirectorate for Terrestrial and Marine Biodiversity, Ministry for the Ecological Transition and the Demographic Challenge, Government of Spain. No experiments were performed on live animals because our work was based on dead stranded cetaceans.
Autopsies were conducted on kogiids stranded dead or alive along the coasts of the Canary Islands from 1999 to 2018. Life history data (i.e., age category, sex, body condition [BC], and decomposition status [DS], morphometrics, and stranding conditions of the individuals) were recorded systematically following established criteria [11,12] (Appendix A Table ). Three age categories based on total body length (TBL) and gonadal macroscopic and histologic appearance were considered: neonate/calf, juvenile/subadult, and adult [21,22]. The BC of each animal was established morphologically based on anatomical parameters such as the osseous prominence of the spinous and transverse vertebral processes and ribs, the mass of the epaxial musculature (longissimus dorsi, multifidus, spinalis), and the amount of subcutaneous and cavitary fat deposits, taking account of the species and the age of the animal. These parameters allowed us to classify their BC as good, moderate, poor, and emaciated. The DS was classified as very fresh (individuals known to live-strand and autopsied immediately), fresh, moderate autolysis, advanced autolysis, or very advanced autolysis [21,22].
A standardized autopsy protocol was used [21,22]. Special attention was paid to the cervical gland to establish the presence/absence of Crassicauda sp. adult or larval forms. Grading of Crassicauda parasitic burden in the cervical gill gland was based on adult and larval nematode counts wherein <10 individuals represented grade +, 11–20 grade ++, and >21 grade +++.
2.2. Statistical Analysis
The association between sex and age category among stranded individuals was assessed using the Fisher–Freeman–Halton exact test in Microsoft Excel (Microsoft Corporation, Redmond, WA, USA) with the Real Statistics add-in. Sex and age category were treated as categorical variables. Statistical significance was set at p < 0.05. Generative artificial intelligence (ChatGPT, OpenAI) was used to assist with the statistical analysis and interpretation of data
2.3. Histopathology and Pathologic Classification
Selected samples collected during necropsy were fixed in 10% neutral buffered formalin, routinely processed, paraffin-embedded, sectioned at 5 μm, and stained with Hematoxylin and Eosin (H&E) for histopathological examination. Additional histochemical techniques, including periodic Acid-Schiff (PAS), Masson’s trichrome, and Gram, were performed as needed.
Morphologic diagnoses were established following international and standardized protocols. The prevalence of gross and histologic pathologic findings was assessed per organ/system (Appendix B1 Table and Appendix B2 Table). When possible, and in order of relevance if numerous, etiologic diagnoses were also determined. Finally, macroscopic and microscopic postmortem examination information, together with the stranding circumstances, were scrutinized to determine the “cause of death” (CD) and associated etiologic diagnoses (Appendix C Table). The CD was classified as “natural”, “anthropogenic”, and “not determined”. Major etiologic diagnoses within the natural CDs included: trauma, infectious, parasitic, cardiomyopathy (CMP), and fetal distress. Three major etiologic diagnoses were considered within the anthropic-related CDs: vessel collision (VC), interaction with fishing activities (IFA), and foreign body-associated pathology (FBAP). Furthermore, gross and microscopic lesions typically associated with the “stress response syndrome” or “alarm reaction” [23] and “capture myopathy" [24,25,26] were also considered.
2.4. Microbiology
Fresh tissue samples (skin, skeletal muscle, lung, prescapular, pulmonary, mediastinal and mesenteric lymph nodes, liver, intestine, kidney, spleen, brain) collected routinely during autopsy, were frozen (-80°C) and selectively submitted for bacteriologic analysis. These included routine culture and surface plating on routine media, e.g., Columbia blood agar and preliminary identification of isolates via API® system (API® 20E, API® Rapid 20E, API® Staph, API® 20 Strep, API® Coryne, API® 20A). The tissues cultured and the respective results are recorded in Appendix D Table.
PCR analyses for the detection of cetacean morbillivirus (CeMV) and cetacean herpesvirus (CeHV) followed published protocols [27,28,29,30]. DNA and RNA were simultaneously extracted from 300 µl homogenized tissue samples using pressure filtration with the QuickGene Mini80 system and the DNA Tissue Kit S, with RNA carrier added during the lysis step. When suspected, CeMV was assessed using one-step RT-PCR targeting a 426 bp fragment of the P gene, nested RT-PCR also targeting the phosphoprotein (P) gene, and semiquantitative (sq) RT-PCR targeting a 192 bp region of the Fusion (F) gene. Similarly, CeHV DNA was investigated, when relevant, using nested PCR targeting the DNA polymerase gene. For Brucella spp. detection, a duplex sq PCR targeting the IS711 insertion sequence and conventional PCR amplifying a 223 bp fragment of the bcsp31 gene were used [27,31].
2.5. Osmium Tetroxide (OsO4) and Chromic Acid (H2CrO4)
Detection of lipid emboli in lung tissue was undertaken using two established histochemical techniques following published methodologies [32]. Available and selected lung samples were fixed in 2% osmium, or alternatively, fixed in chromic acid before paraffin embedding, as both approaches enable preservation and visualization of intravascular neutral lipid droplets and are routinely used in cetaceans to support the diagnosis of ante-mortem blunt trauma associated with vessel collision.
3. Results
3.1. Stranding epidemiology
A total of 67 kogiids (K. breviceps, n=51; K. sima, n=13; unknown kogiid species, n=3) stranded between January 1st, 1999, and December 31st, 2018. Postmortem examinations were conducted on 45/67 (67%) animals encompassing 35 K. breviceps and 10 K. sima. Sex distribution of animals studied was 24 (53.3%) males, 18 (40%) females, and 3 (7%) undetermined. Age categories were: 27/45 (60%) adult, 11/45 (24.4%) juvenile/subadult, 5/45 (11.1%) neonate/calf. Age category could not be determined in 2/45 (4.4%) animals. The Fisher–Freeman–Halton exact test indicated no significant association between sex and age class (exact p > 0.05), suggesting these variables were independent in this cohort. Annual strandings through this 20-year period averaged 4 kogiids without a clear yearly stranding bias. Autopsies were performed on 2/45 (4%) very fresh, 7/45 (15%) fresh, 18/45 (40%) moderate autolysis, 5/45 (11%) advanced autolysis and 13/45 (29%) very advanced autolysis carcasses. Five of 45 (11%) animals stranded alive; 40/45 (88.9%) stranded dead or were retrieved adrift. Distribution based on NS included: 9 (25%) good, 12 (27%) moderate, 9 (29%) poor and 3 (7%) cachectic animals. The NS could not be determined in 12 (12%) animals due to advanced decomposition.
The most probable CD was identified in 41/45 (91%) examined individuals. The CD was not determined in 4/45 (9%) individuals. Etiologic diagnoses within the natural CDs included trauma (13/30; 43.3%), infectious (7/30; 23.3), parasitic (5/30; 16.6%), CMP (3/30; 10%), malnutrition (1/30; 3.3%), and fetal distress (1/30; 3.3%) while trauma, encompassing vessel collision (7/11; 18.2%) and IFA (2/11; 18.2%), and FBAP (2/11) were the most prevalent within the anthropic CDs. Non-anthropogenic CDs accounted for 30/45 (66.6%) of deaths, whereas anthropogenic activities included 11/45 (24.4%) cases.
3.2. Natural causes of death
Natural CD was determined in a total of 30 animals, including 27 K. breviceps and 3 K. sima (animals no. 1, 10, 14 ,16, 24, 28, 33, 34, 37, 38, 39, 42, 44, 45).
Traumatic intra-interespecific interaction was identified as the CD in 11 K. breviceps and 2 K. sima (animals no. 11, 14, 16, 24, 28, 33, 34, 37, 38, 39, 42, 44, 45). From the adults, a total of 5 animals were female and 5 were male. Among females, 3 were pregnant (animal no. 10 and 11) or exhibited uterine post-partum regression (animal no. 44). Additionally, most of the traumatic events were recorded in animals stranded along the coasts of the western islands (7/13; Tenerife=4, La Gomera=2, La Palma=1). Main macroscopic lesions in these animals were: bone fractures (i.e., skull, mandibular, teeth, vertebral, hyoid apparatus), multiorgan (e.g., pulmonary, central nervous system, lymph nodes) hemorrhage, hemoabdomen, hemothorax, hemopericardium, cutaneous erosions/lacerations, and interspecific rake marks (by killer whales and/or sharks) with occasional extensive loss of muscle bundles (flank, head, thoracic, and central region), hemarthrosis, pulmonary perforation, and pulmonary oedema (Fig. 1A, B, C). Parasitic gastritis (e.g., Anisakis spp., P. gasterophilus) was identified in 10/13 animals, being moderate to severe in 7/13. Most relevant microscopic findings encompassed: multiorgan hemorrhage, more prominently affecting the lungs and central nervous system (CNS), myodegeneration and necrosis of skeletal muscle, and pulmonary edema. Myocardial degeneration and necrosis were common findings in most individuals. Animal no. 14 exhibited pulmonary, cardiac and lymphatic intravascular gas or fat emboli. Moderate to abundant partially digested ingesta was observed in 7/13 individuals.
Singular cases in this category included an ulcerative dermatitis with vasculitis, epithelial necrosis, with occasional amphophilic intranuclear inclusions identified as herpesvirus through PCR (animal no. 33) and lymphoplasmacytic balanitis with renal urolithiasis (animal no. 34).
Gross and histologic findings aligning with an infectious disease process were identified in 5 K. breviceps and 2 K. sima (animals no. 3, 12, 23, 26, 31, 32, 3). Animal no. 32 exhibited multifocal intestinal serosal emphysema (pneumatosis intestinalis), lymphohistiocytic hepatitis with necrosis, splenic necrosis, and multinodal (hepatic, tracheobronchial, retromandibular) necrosuppurative lymphadenitis with intralesional sporulated Gram-positive bacilli (Fig. 1D). Animal no. 36 had fibrinosuppurative peritonitis with intralesional Gram-positive, sporulated bacilli. Bacteriologic analysis identified Cl. perfringens in both cases. Animal no. 31 displayed a lymphoplasmacytic meningoencephalitis with myelitis (Fig. 1E). Other relevant cases included a fibrinosuppurative omphalophlebitis and peritonitis in animal no. 26, lymphoplasmacytic periportal hepatitis, endometritis, bronchointerstitial pneumonia, and nephritis in animal no. 12, and suppurative bronchopneumonia with type II pneumocyte hyperplasia in animal no. 23. Intravascular pulmonary fat emboli were identified in animal no. 12 by using osmium tetroxide (OsO4) staining.
Fatal parasitism was identified in 3 K. breviceps and 2 K. sima (animals no. 13, 10, 17, 22, 25). Suppurative cervical adenitis of the gill slit gland by Crassicauda spp. nematodes, with or without multisystemic nematode egg emboli with regional deep and superficial myositis and fasciitis was observed in animals no. 1, 13, 14, 16, 17, 19, 22, 25, 8, 28, 31, 32, 34, 36, 42 (Fig. 1F). From these, animal no. 13 had a 15 cm in diameter abscess of the cervical gill slit. Severe ulcerative gastritis with numerous Anisakis spp. was identified in animals no. 17 and 22. A pyogranulomatous cholangiohepatitis with intralesional Brachycladiidae trematodes was also identified in animal no. 17. Additionally, neuroparenchymal vacuolation including presumptive intramyelinic edema with axonal degeneration, astrocytosis, and neuronal satellitosis were seen within the thalamus and cerebral cortex of this animal. Animal no. 13 exhibited multiple sarcoplasmic masses within skeletal myofibers, associated with changes in sarcolemmal nuclear number and localization, variation in myofiber size and shape, and altered fiber type proportions. Animal no. 25 exhibited moderate catarrhal bronchitis with intraluminal nematodes (undetermined species). A hepatocellular carcinoma was noted in this animal.
Severe CMP with subsequent multiorgan failure was identified as the main cause of death in 3 K. breviceps (animals no. 2, 5, 35). Two were adults (male and female) and one was a juvenile male. Prominent gross changes included dilation of the right ventricle (Fig. 1G) and pulmonary congestion, edema, and hemorrhage, along with chronic passive hepatic congestion. Frequent histologic features comprised: acute cardiomyocyte degeneration, contraction band necrosis, myocardial fiber disarray, juxtanuclear vacuolization, cardiomyocyte hypertrophy, and subendocardial and myocardial fibrosis. Additional findings in these animals included: cerebral perivascular edema (animal no. 2), hepatic periportal fibrosis with ductular reaction (animal no. 5), and hydrothorax (animal no. 2). A ventricular septal defect was present in animal no. 35. Overall, 31 animals (68.9%) displayed gross and/or histologic features consistent with CMP (Appendix E Table). Of these, 15 were males, 14 females, and 2 of undetermined sex. Most affected individuals were adults (n=19), followed by juveniles (n=6), and calves (n=6). Acute cardiomyocyte degeneration and/or contraction band necrosis were identified in 20 cases (44%), and cardiomegaly was observed in 14 individuals (31%), including 9 males, 4 females, and 1 animal of undetermined sex; among these, 9 were adults, 4 juveniles, and 1 calf. Pulmonary edema (26/45; 57.8%) and hepatic congestion (18/45; 40%) were common in this group, particularly those with cardiomegaly, in which pulmonary edema was present in almost all cases.
Animal no. 8 exhibited marked emaciation with diffuse skeletal muscle atrophy and nearly complete lack of ingesta. Mild multiorgan parasitism, including suppurative cervical lymphadenitis, and the presence of an early-term fetus were also documented.
Fetal distress was diagnosed in animal no. 29, a newborn with typical findings of neonatal development e.g., folded fins and fetal folds, open umbilicus, non- erupted teeth, non-ossified cranial sutures. Histologically, pulmonary oedema and fetal atelectasis, and moderate, acute myodegeneration and necrosis were seen in the skeletal and diaphragmatic muscles.
3.3. Anthropogenic pathologic categories
VC involved 7/45 (%) K. breviceps (animals no. 7, 9, 15, 18, 19, 27, 43). Main gross findings in these animals, usually severe, included: sharp and blunt trauma, polyostotic fractures, more prominently in the cephalocervical region (e.g., skull, mandible, vertebrae; animals no. 7, 9, 18, 19, 27, 43) (Fig. 1H)., cutaneous lacerations (animals no. 9, 10, 19), sectioning or amputation (animals no. 15, 27 and 43), pulmonary hemorrhage (animals no. 9, 10, 18, 19, 27, 43), cavitary and multiorgan hemorrhage (animals no. 18 and 27), and evisceration (animal no. 27). Microscopically, pulmonary and CNS hemorrhage, contraction band necrosis, acute flocculent, granular, or discoid skeletal myodegeneration were common. Animal no. 19 exhibited multiorgan intravascular gas emboli, while chromic acid (H2CrO4) demonstrated intravascular fat emboli in animal no. 27. Gastrointestinal parasitism by Anisakis spp. and/or Pholeter gastrophilus was identified in animals no. 9 and 19. Dilated cardiomyopathy with fibrosis, hypertrophy, and contraction band necrosis was present in animals no. 19 and 27.
Fatal IFA involved 2/45 animals, including one K. breviceps and one K. sima (animal no. 1, 4). Trauma was the main etiologic diagnosis in these animals. Major gross findings associated with by-catch included: comminute fracture of the base of the skull and occipital bone with extensive subdural hematoma (animal no. 1) and fractures of the skull, mandible, and tympanic bulla together with cutaneous marks compatible with entanglement (animal no. 4). Microscopically, acute skeletal muscle degeneration and necrosis, pulmonary oedema and hemorrhage, and CNS hemorrhage, were described. Animal no. 1 exhibited marked parasitism of the skeletal epaxial and hypaxial muscles by Crassicauda spp., hydroureter, and dilated cardiomyopathy.
FBAP was determined in 2/45 K. breviceps (animals no. 30 and 41). Macroscopically, the glandular stomach of both animals was filled with moderate, heterogeneous, hard and firm plastic-based fragments along with ulcerative gastritis and myriads of Anisakis spp. (Fig. 1I).
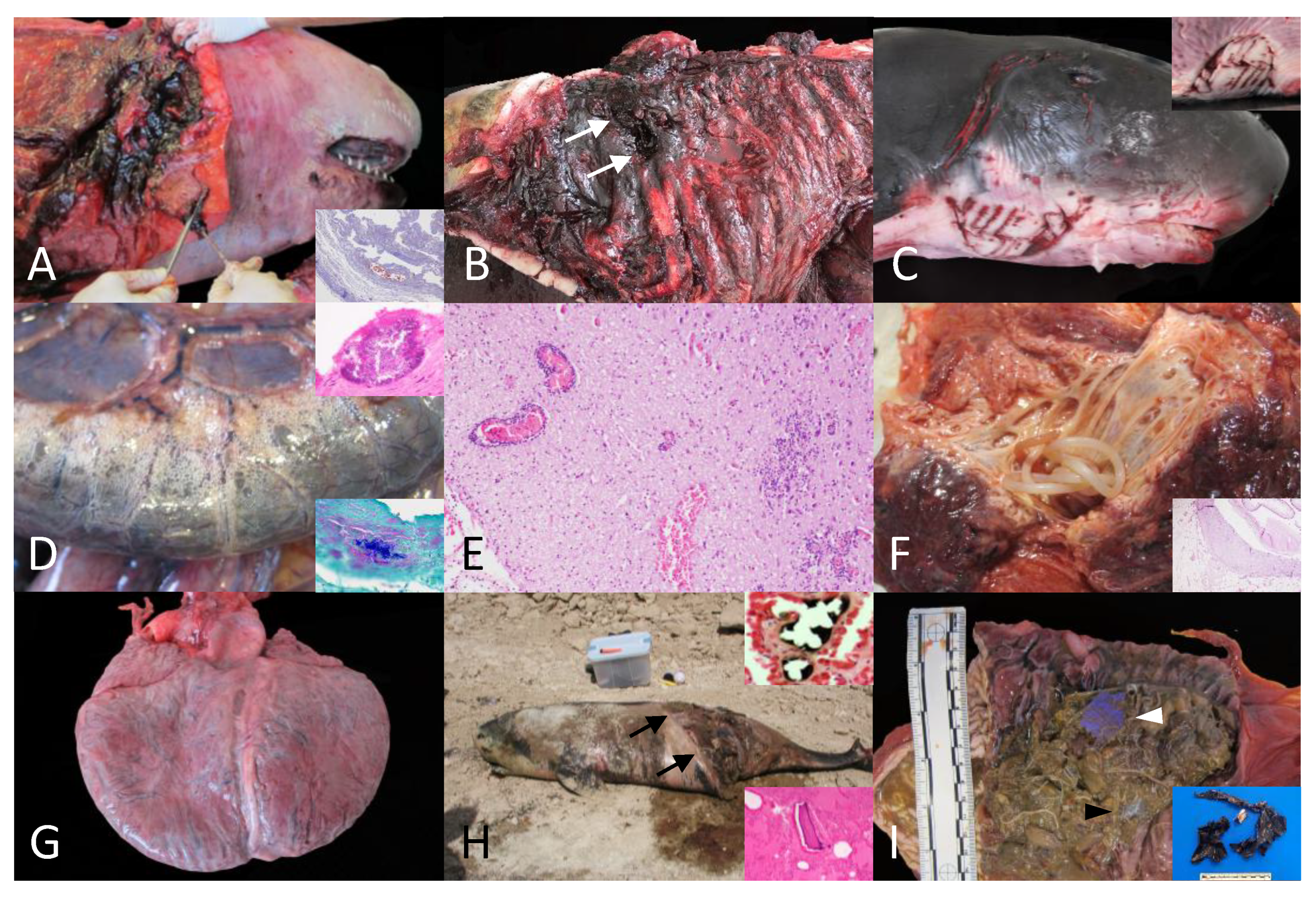
Figure 1.
A. Animal no. 28. Intra-interspecific traumatic interaction. Extensive subcutaneous cervical circumferential hemorrhage extending deep into muscle planes. Inset: Animal no. 27: Intraspecific interaction with killer whale. Fat emboli (Oil red). A focal pulmonary subpleural vessel is partially obliterated by a fat emboli (red). B. Animal no. 34: Intra-interspecific traumatic interaction. Focal perforation with peripheral hemorrhages within the left dorso-lateral aspect of the thoracic cavity (white arrows) and associated lung perforation (non-visible). C. Animal no. 33. Intraspecific traumatic interaction with shark. Inset: Multifocal cutaneous intraspecific rake marks with dermal hemorrhages and often displaying clean and sharp edges and half-moon disposition (left and ventro-lateral cervical region). D. Animal no. 32. Clostridium perfringens infection. The serosa of the small intestine and the mesenterium are multifocally expanded by gas (emphysema; Pneumatosis intestinalis). Upper inset, H&E: Fibrinosuppurative peritonitis within the visceral diaphragmatic surface with focal intralesional bacterial aggregate. Lower inset, Gram stain: Note the gram positive intralesional bacteria. E. Animal no. 31. Infectious encephalitis, unknown etiology. Lymphoplasmacytic encephalitis with multiple perivascular cuffing formation (upper left), multiple areas of gliosis, and hemorrhage. F. Animal no. 32. Cervical gill adenitis. The cervical gland is expanded by numerous long nematodes (Crassicauda sp.). Inset, H&E: Detail of the pyogranulomatous panniculitis with intralesional transverse section of Crassicadua sp. displaying a light eosinophilic cuticle, hypodermis, pseudocoelom, coelomyarian-polymyarian musculature, digestive system lined by cuboidal to columnar cells, and paired uteri (non-visible). G. Animal no. 35. Dilated cardiomyopathy. The heart exhibits a globous shape with marked dilation of the right ventricular chamber and thin right free-ventricular wall (non-visible). Note doble apex. H. Animal no. 7. Vessel collision. Animal at necropsy spot. The abdomen exhibits a longitudinal and obliquus section with clear edges and partial evisceration (black arrows). Upper inset, Animal no. 19. Osmium tetraoxide (OsO₄). Detail of a pulmonary fat emboli within the pulmonary vasculature. Lower inset, Animal 18, H&E: Intravascular osseous fragments. I. Animal no. 30. Foreign body associated pathology. The keratinized stomach is filled with multiple transparent (black arrowhead) and blue (white arrowhead) plastic materials embedded in abundant Anisakis sp. Inset, animal no. 41: Display of gastric foreign plastic material.
Figure 1.
A. Animal no. 28. Intra-interspecific traumatic interaction. Extensive subcutaneous cervical circumferential hemorrhage extending deep into muscle planes. Inset: Animal no. 27: Intraspecific interaction with killer whale. Fat emboli (Oil red). A focal pulmonary subpleural vessel is partially obliterated by a fat emboli (red). B. Animal no. 34: Intra-interspecific traumatic interaction. Focal perforation with peripheral hemorrhages within the left dorso-lateral aspect of the thoracic cavity (white arrows) and associated lung perforation (non-visible). C. Animal no. 33. Intraspecific traumatic interaction with shark. Inset: Multifocal cutaneous intraspecific rake marks with dermal hemorrhages and often displaying clean and sharp edges and half-moon disposition (left and ventro-lateral cervical region). D. Animal no. 32. Clostridium perfringens infection. The serosa of the small intestine and the mesenterium are multifocally expanded by gas (emphysema; Pneumatosis intestinalis). Upper inset, H&E: Fibrinosuppurative peritonitis within the visceral diaphragmatic surface with focal intralesional bacterial aggregate. Lower inset, Gram stain: Note the gram positive intralesional bacteria. E. Animal no. 31. Infectious encephalitis, unknown etiology. Lymphoplasmacytic encephalitis with multiple perivascular cuffing formation (upper left), multiple areas of gliosis, and hemorrhage. F. Animal no. 32. Cervical gill adenitis. The cervical gland is expanded by numerous long nematodes (Crassicauda sp.). Inset, H&E: Detail of the pyogranulomatous panniculitis with intralesional transverse section of Crassicadua sp. displaying a light eosinophilic cuticle, hypodermis, pseudocoelom, coelomyarian-polymyarian musculature, digestive system lined by cuboidal to columnar cells, and paired uteri (non-visible). G. Animal no. 35. Dilated cardiomyopathy. The heart exhibits a globous shape with marked dilation of the right ventricular chamber and thin right free-ventricular wall (non-visible). Note doble apex. H. Animal no. 7. Vessel collision. Animal at necropsy spot. The abdomen exhibits a longitudinal and obliquus section with clear edges and partial evisceration (black arrows). Upper inset, Animal no. 19. Osmium tetraoxide (OsO₄). Detail of a pulmonary fat emboli within the pulmonary vasculature. Lower inset, Animal 18, H&E: Intravascular osseous fragments. I. Animal no. 30. Foreign body associated pathology. The keratinized stomach is filled with multiple transparent (black arrowhead) and blue (white arrowhead) plastic materials embedded in abundant Anisakis sp. Inset, animal no. 41: Display of gastric foreign plastic material.
4. Discussion
This study provides the most comprehensive pathology-based assessment of kogiids from the Atlantic Ocean to date. Epidemiologic data from 67 stranded whales were compiled, and detailed autopsy and histopathologic examinations were performed on 45 individuals (35 K. breviceps, 10 K. sima). By integrating gross, microscopic, and ancillary laboratory test results, we determined causes of death and characterized the pathogenesis of major natural and anthropogenic conditions. When applicable, traumatic and other lesions were interpreted using veterinary forensic criteria [33]. This approach provides a rigorous framework for evaluating health status, mortality drivers, and species-specific vulnerabilities in these poorly studied species [8,9,11,12,16,18,34,35].
Natural causes of death
Among natural CDs, traumatic intra-interspecific interaction was the most prevalent entity. The pathogenesis of fatal sharp-blunt trauma in cetaceans reflects the general principles of mechanical injury; the sudden application of kinetic energy exceeding tissue tolerance results in acute disruption of soft tissues, vascular rupture, and rapid hemodynamic and respiratory compromise, with high-energy ramming and compression producing severe internal polytrauma even when external lesions are limited [33,36,37]. The distribution and sex predilection of these events mirror patterns previously documented for cetaceans stranded in the same region [37]. In these animals, the main postmortem findings were consistent with sharp and blunt force polytrauma; numerous animals exhibited abundant undigested ingesta and frequent cutaneous rake marks, in agreement with previous studies [37]. Cutaneous tooth rake marks may be the most representative finding in this entity, as the other findings could be found in other categories involving fatal trauma [37]. Predation by killer whales (Orcinus orca) was identified based on deep, circular rake marks spaced 2-3 cm apart, and by massive loss of soft and bone tissues in two animals. Killer whales are versatile top predators known to prey upon numerous odontocetes and mysticetes [38,39,40,41,42,43,44] with documented fatal attacks on kogiids in different geographic regions [45,46]. These species are seasonal visitors in the Canary Islands, following tuna migration and frequently interacting with pygmy sperm whales [37,47]. Our findings align with the proposed higher susceptibility of deep-diving cetaceans to killer whale attacks [37].
Other less frequent but relevant findings included hepatocellular intracytoplasmic hyaline globules, reported in acute systemic stress responses [48], hyaline casts and pigmentary tubulonephrosis, documented in life stranding events and compatible with hypoxia, hematuria and/or myoglobinuria [24,25]. Intravascular and parenchymal gas embolism in animal no. 14 was interpreted as compatible with a decompression syndrome secondary to the traumatic event and abrupt alteration of the diving pattern [37,49]. Gas analysis could not be performed in this animal due to logistical impediments [50].
Parasitic gastritis by Anisakis spp. and P. gastrophilus, was the most prevalent comorbidity in 60% of evaluated animals, likely contributing to a reduced overall health status and a potentially increased vulnerability to traumatic interactions. Chronic mechanical injury from these parasites promotes ulceration, fibrosis, and protein-losing gastropathy, changes well documented in helminth-infected cetaceans [51,52,53]. Gastric anisakiasis is widely recognized in cetaceans and shows increasing global incidence [51,52,53]. An extensive study on short-beaked common dolphins (Delphinus delphis) reported an incidence of ulcerative gastritis with intralesional Anisakis spp. in 74% of examined animals, with molecular identification of Anisakis simplex and A. pegreffii [51]. Similar high morbidity has been described in other cetacean species [52,53]. In the present study, identification of the species or genotype of Anisakis involved was not performed.
Animal no. 10 exhibited a fibrinosuppurative peritonitis following rupture of a hepatic abscess. Implication of Brachychladiidae trematodes is presumed, as trematode-associated abscess rupture have been previously reported in odontocetes [11,54,55].
A herpesviral dermatitis was diagnosed in animal no. 33. Cutaneous herpesvirosis has been described and characterized in a variety of cetacean species [56,57,58,59,60], but it remains largely uncharacterized in kogiids [58,61,62]. This case therefore provides additional evidence of herpesviral infection in this family. A lymphoplasmacytic balanitis was identified in animal no. 34. Although a viral agent is highly suspected, immunohistochemical or molecular techniques are required for final confirmation.
Bacteriologic analysis identified septicemia by Cl. Perfringens in animals no. 32 and 36, exhibiting multisystemic lesions. The pathogenesis of Cl. perfringens infection is largely toxin-driven, with rapid proliferation under anaerobic conditions and release of α-toxin, perfringolysin O, and β-toxin (type C), which induce endothelial injury, mucosal necrosis, hemorrhage, and systemic toxemia [63,64]. Clostridial diseases have been rarely documented, causing disease in both captive and free-ranging cetaceans [65,66,67,68,69]. Reported historical lesions in cetaceans included necrotizing myositis, emphysema, and dryness, mainly involving skeletal muscles, serofibrinous exudate, multiorgan hemorrhage, and myocarditis [65,66,67,68,69]. Notably, pneumatosis intestinalis has been associated with gas-producing Cl. perfringens (e.g., type A) in humans and several animal species [70,71,72,73]. Clostridial toxin characterization for strain typification was not performed in these animals. Clostridium tertium was isolated from a dwarf sperm whale (animal no. 3). Clostridium tertium has been rarely identified in cetacean species [12,74]. Our findings expand the body of knowledge of clostridial diseases in cetaceans, documenting the first Cl. perfringens infection in pygmy sperm whales and a rare Cl. tertium case in one dwarf sperm whale. A lymphoplasmacytic meningoencephalitis with myelitis was diagnosed in animal no. 31. PCR assessment for CeMV and CeHV yielded negative results. Further evaluations are required to elucidate the underlying etiology. Other cases without confirmed etiology included omphalophlebitis with peritonitis (animal no. 26), hepatitis, endometritis, nephritis, and pneumonia (animal no. 12 and no. 23).
Cervical slit adenitis by Crassicauda spp. was identified in most animals. A recent study provided an extensive morphologic and histologic description of the gill slit gland, only described in pygmy sperm whales [7]. The gill slit has been identified as a frequent site for infection by C. magna [6,8]. This giant nematode (~3 m-long in adults) reproduces inside this exocrine gland and uses it to release the eggs into the environment [7]. In our cases, gross and histological findings together with body location, align with a moderate to severe infestation by C. magna. Such severe parasitism of the gill split can alter social interaction, mating behaviors, or territoriality given the described functions for this gland [6,7,8]. Crassicaudiasis is a significant parasitic disease of cetaceans with predilection for vascular, renal, subcutaneous, and glandular (e.g., cervical gill slit, mammary gland) tissues, which may exhibit species-specific patterns of tissue tropism and serious health implications [6,75,76,77,78,79,80,81,82].
Cardiomyopathy was the most probable CD in 3 K. breviceps. In this condition, reduced myocardial function leads to congestive heart failure, with progressively impaired forward flow and venous congestion. Left-sided failure results in pulmonary venous congestion and alveolar edema, while right-sided or global failure causes chronic passive hepatic congestion and systemic congestion. Neurohumoral activation further increases circulating volume and capillary pressures, promoting interstitial and serous effusions [83]. In our study, the recurrent association of cardiomyopathic lesions with pulmonary edema and hepatic congestion is therefore consistent with terminal congestive cardiac failure rather than isolated myocardial injury. Cardiomyopathy is well documented in kogiids but the etiopathogenic mechanisms remain unknown [17,18,20,84]. Upregulation of metallothioneins and sequestering of transition metals (e.g., selenium) have been described in kogiids with CMP, arguably due to adaptive cardioprotection [85]. Bossart et al. [17] documented gross and histologic features of CMP and myocardial degeneration occurring in approximately 50% and 40 %, respectively, of examined kogiids, mainly adult males. In contrast, our cohort showed 31 (69%) individuals with lesions compatible with CMP, without evident sex distribution bias and a broader age range. The high prevalence of chronic cardiac lesions with remodeling, particularly among adult individuals, supports the interpretation of these as hallmarks of pathologic CMP rather than species-specific physiologic adaptations. Further investigations into the etiopathogenic mechanisms of CMP in kogiid whales, including genetic analyses, are warranted.
Animal no. 13 exhibited multiple sarcoplasmic masses within skeletal muscle, resembling lesions described in chronic myopathies of humans and domestic animals [86].
Malnutrition or starvation was the CD in animal no. 8. Prolonged energy deficiency leads to depletion of adipose and muscular reserves, impaired gluconeogenesis, and systemic metabolic stress with elevated pro-inflammatory cytokines, particularly TNF-α, which accelerates proteolysis and lipolysis, compounding tissue catabolism [87]. This animal had profound skeletal muscle atrophy, minimal ingesta, myositis and fasciitis by Crassicauda spp., and cutaneous lesions consistent with intra-interspecific interactions. Malnutrition in cetaceans is a critical health challenge linked to reduced reproductive capacity, increased susceptibility to comorbidities, and progression to multiorgan dysfunction [11,12,88,89,90,91,92,93]. Poor body condition is described in species undergoing prolonged periods of starvation during migration or breeding seasons [94,95,96]. In this case, early pregnancy likely imposed additional energetic demands that might have aggravated health decline [97].
Fetal distress was the CD in a neonate (animal no. 29). Fetal distress arises from impaired placental or fetal oxygenation, leading to hypoxia, redistribution of fetal blood flow, and increased intestinal peristalsis with meconium passage and aspiration, producing atelectasis, chemical pneumonitis, and metabolic injury [98,99]. Our findings in this case are consistent with previous cases of fetal distress associated with various etiologies in cetaceans [11,26,99,100].
Anthropogenic causes of death
Vessel collision is a major threat to cetaceans with a growing incidence driven by increasing maritime traffic and vessel speed [101,102,103]. Certain species, including the fin whale (Balaenoptera physalus), humpback whales (Megaptera novaeangliae), North Atlantic right whales (Eubalaena glacialis), or sperm whales (Physeter macrocephalus), are particularly vulnerable [36,101,103,104]. In our cohort, adult males were overrepresented among VC cases, contrasting with previous studies that describe higher susceptibility in pregnant females and neonates in some cetacean populations, providing novel epidemiologic data of ship collision in K. breviceps [11,101,103,104,105,106]. VC risk is strongly influenced by boat speeds and species-specific surface behaviors, including resting, foraging, nursing, and socializing on surface [101,102,104,107,108,109,110,111]. The Atlantic Ocean harbors the highest occurrence of documented ship strikes, with the Canary Islands identified as a major collision hotspot due to intense maritime traffic and high densities of cetacean populations [11,12,105,109,111]. Gross and histologic findings in our cases are consistent with propeller and hull impacts, leading to catastrophic tissue damage and rapid cardiovascular collapse [33,103,112,113,114]. With occasional pulmonary fat embolism further supporting antemortem trauma even in decomposed carcasses [31,105]. VC emerged as the leading anthropogenic CD in this study, underscoring its significance as a conservation concern for pygmy sperm whales for collision-related mortality could exceed recruitment and drive population decline, either alone or in synergy with other stressors [110,113].
IFA was the CD in animals no. 1 and 4. Bycatch is the leading anthropogenic threat to cetaceans worldwide, with gillnets, trawls, and longlines responsible for most entanglement-related mortality [115,116,117]. Small odontocetes are especially vulnerable, often dying from asphyxia, while larger mysticetes may endure long-term entanglement injuries [36,115,117,118,119]. In addition, fishing gear can also cause sharp injuries, including hook or harpoon punctures [118,120]. Underreporting is widespread, especially in gillnet fisheries, where observer-based estimates routinely exceed fisher logbook reports [121,122]. Although adult shallow divers are typically overrepresented in IFA records from the Canary Islands [118], the involvement of 2 juveniles in our cohort aligns with proposed increased susceptibility linked to behavioral immaturity, nearshore foraging, and longer surface intervals [117,118]. IFAs are rarely documented in kogiids due to their cryptic behavior and identification challenges at sea [18,123,124,125]. These findings contribute to the limited knowledge on the pathologic consequences of fisheries interactions in kogiids and highlight the need to consider such impacts on less-reported species.
FBAP was the cause of death in animals no. 30 and 41. Marine debris ingestion poses a widespread threat to cetaceans, with records in over 63% of species [126,127,128]. Deep-diving species and young age are recognized as risk factors for foreign body ingestion, likely due to limited foraging experience and reduced ability to discriminate prey from acoustically mistaken anthropogenic debris [11,89,129,130]. Plastic ingestion has been previously reported in kogiids [3,131]. In this study, both animals exhibited moderate to abundant firm and hard plastic fragments in the glandular stomach associated with ulcerative gastritis, a common concurrent lesion in FBAP [89,132]. In animal no. 30, numerous Anisakis spp. were embedded within the plastic mass, causing gastric obstruction that likely resulted in impaired digestion, malnutrition, and increased susceptibility to comorbidities [11,89,133]. Animal no. 41 had intralesional filamentous bacteria associated with Splendore-Hoeppli phenomenon within the gastric ulcer [53]. These findings reinforce early age and deep-diving behavior as major risk factors for foreign-body ingestion in kogiids.
5. Conclusions
In conclusion, this study provides a comprehensive pathologic assessment of stranded Kogia breviceps and K. sima over a two-decade period in the Canary Islands. Natural causes of death accounted for most deaths, with intra-interespecific trauma representing the most frequent pathologic category, followed by infectious and parasitic disease, cardiomyopathy, malnutrition, and fetal distress. Anthropogenic causes were also relevant and included primarily vessel collisions, fishing interactions and foreign body-associated pathology. Intra-interspecific traumatic interactions were overrepresented in this cohort. Cardiomyopathic lesions were highly prevalent (69%) without any evident sex distribution bias. Cervical gill adenitis by Crassicauda spp. and gastritis by Anisakis sp. were prevalent parasitic diseases. Septicemia caused by Clostridium perfringens and Cl. tertium were novel findings for these species. Collectively, these findings enhance current knowledge on kogiid pathology and provide valuable information for conservation efforts, emphasizing the importance of ongoing stranding surveillance and systematic post-mortem investigations to better understand and mitigate natural and anthropogenic threats.
Author Contributions
Conceptualization, P. D.-S., J.D.-D., M.A. Methodology, P. D.-S , J.D.-D. Validation, A.F., E.S.,N.C. Formal analysis, P. D.-S., J.D.-D. Investigation, P. D.-S., J.D.-D., M.A., A.F. Resources, A.F., M.A., N.C.,V.M. Data curation, P. D.-S., J.D.-D. Writing—original draft preparation, P. D.-S., J.D.-D., C.S.-S. Writing—review and editing, P. D.-S., J.D.-D., M.A., O.Q.-C., C.S.-S., E.S., A.F. Supervision, J.D.-D., C.S.-S., M.A., O.Q.-C., E.S., A.F. Project administration, A.F. Funding acquisition, A.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author
Acknowledgments
All authors of this study would like to thank our laboratory technicians and administrative staff, as well as all members and volunteers of the Cetacean Stranding Network of the Canary Islands, for their invaluable support. We also extend our gratitude to everyone contributing to the faithful scientific veracity. Special acknowledgement is given to Víctor Torres for his altruistic and dedicated collaborative effort.
Conflicts of Interest
The authors declare no potential conflict of interest with respect to the research, authorship and/or publication of this article.
References
- Committee on Taxonomy. List of marine mammal species and subspecies. Society for Marine Mammalogy. Available online: www.marinemammalscience.org (accessed on 29/05/2025).
- Plön, S. Pygmy Sperm Whale Kogia breviceps (de Blainville, 1838). In Handbook of the Mammals of Europe; Springer: Cham, Switzerland, 2023; pp. 1–27. [Google Scholar]
- McAlpine, D.F. Pygmy and dwarf sperm whales. In Encyclopedia of Marine Mammals, 3rd ed.; Würsig, B., Thewissen, J.G.M., Kovacs, K.M., Eds.; Academic Press: London, United Kingdom, 2018; pp. 786–788. [Google Scholar]
- McAlpine, D.F. Pygmy and dwarf sperm whales. In Encyclopedia of Marine Mammals, 2nd ed.; Perrin, W.F., Würsig, B., Thewissen, J.G.M., Eds.; Academic Press: Amsterdam, The Netherlands, 2009; pp. 936–938. [Google Scholar]
- West, K.L.; Walker, W.A.; Baird, R.W.; White, W.; Levine, G.; Brown, E.; et al. Diet of pygmy sperm whales (Kogia breviceps) in the Hawaiian Archipelago. Mar. Mamm. Sci. 2009, 25, 931–943. [Google Scholar] [CrossRef]
- Keenan-Bateman, T.F.; McLellan, W.A.; Costidis, A.M.; Harms, C.A.; Gay, D.M.; Rotstein, D.S.; et al. Habitat use pattern of the giant parasitic nematode Crassicauda magna within the pygmy sperm whale Kogia breviceps. Dis. Aquat. Organ. 2018, 127, 163–175. [Google Scholar] [CrossRef]
- Keenan, T.F.; McLellan, W.A.; Rommel, S.A.; Costidis, A.M.; Harms, C.A.; Thewissen, J.G.M.; et al. Gross and histological morphology of the cervical gill slit gland of the pygmy sperm whale (Kogia breviceps). Anat. Rec. 2022, 305. 688–703. [Google Scholar] [CrossRef]
- Keenan-Bateman, T.F.; McLellan, W.A.; Harms, C.A.; Piscitelli, M.A.; Barco, S.G.; Thayer, V.G.; et al. Prevalence and anatomic site of Crassicauda sp. infection, and its use in species identification, in kogiid whales from the mid-Atlantic United States. Mar. Mamm. Sci. 2016, 32, 868–883. [Google Scholar] [CrossRef]
- Bennati-Madureira, L.; Gomes, G.L.; Daros, K.A.C.; Casas, A.L. da S. Anatomical description of a pygmy sperm whale, Kogia breviceps (Cetacea: Kogiidae), pre-term calf using CT scan and 3D reconstructions. Anat. Rec. 2024, 307, 1043–1058. [Google Scholar] [CrossRef]
- Bloodworth, B.E.; Odell, D.K. Kogia breviceps (Cetacea: Kogiidae). Mamm. Species 2008, 819, 1–12. [Google Scholar] [CrossRef]
- Díaz-Delgado, J.; Fernández, A.; Sierra, E.; Sacchini, S.; Andrada, M.; Vela, A.I.; et al. Pathologic findings and causes of death of stranded cetaceans in the Canary Islands (2006–2012). PLoS ONE 2018, 13, e0193025. [Google Scholar] [CrossRef] [PubMed]
- Arbelo, M.; de los Monteros, A.E.; Herráez, P.; Andrada, M.; Sierra, E.; Rodríguez, F.; et al. Pathology and causes of death of stranded cetaceans in the Canary Islands (1999–2005). Dis. Aquat. Organ. 2013, 103, 87–99. [Google Scholar] [CrossRef]
- Stamper, M.A.; Whitaker, B.R.; Schofield, T.D. Case study: Morbidity in a pygmy sperm whale Kogia breviceps due to ocean-borne plastic. Mar. Mamm. Sci. 2006, 22, 719–722. [Google Scholar] [CrossRef]
- Shamsi, S.; Spröhnle-Barrera, C.; Hossen, M.S. Occurrence of Anisakis spp. (Nematoda: Anisakidae) in a pygmy sperm whale Kogia breviceps (Cetacea: Kogiidae) in Australian waters. Dis. Aquat. Organ. 2019, 134, 65–74. [Google Scholar] [CrossRef]
- Rousselet, E.; Stolen, M.; Durden, W.N.; Jablonski, T.; Stacy, N.I.; Rotstein, D.S. Bilateral polycystic kidneys and focal renal cystadenoma in a pygmy sperm whale (Kogia breviceps). J. Wildl. Dis. 2019, 55, 258–261. [Google Scholar] [CrossRef] [PubMed]
- Plön, S.; Best, P.B.; Duignan, P.; Lavery, S.D.; Bernard, R.T.F.; Van Waerebeek, K.; et al. Population structure of pygmy (Kogia breviceps) and dwarf (Kogia sima) sperm whales in the Southern Hemisphere may reflect foraging ecology and dispersal patterns. In Advances in Marine Mammal Science; Springer: Cham, Switzerland, 2023; pp. 85–114. [Google Scholar]
- Bossart, G.D.; Hensley, G.; Goldstein, J.D.; Kroell, K.; Manire, C.A.; Defran, R.H.; et al. Cardiomyopathy and myocardial degeneration in stranded pygmy (Kogia breviceps) and dwarf (Kogia sima) sperm whales. Aquat. Mamm. 2007, 33, 113–122. [Google Scholar] [CrossRef]
- National Marine Fisheries Service (NMFS). Dwarf and pygmy sperm whale (Kogia spp.) best practices. 2021. [Google Scholar]
- Plön, S.; Andra, K.; Auditore, L.; Gegout, C.; Hale, P.J.; Hampe, O.; et al. Marine mammals as indicators of Anthropocene ocean health. npj Biodivers. 2024, 3, 24. [Google Scholar] [CrossRef]
- Li, W.T.; Chou, L.S.; Chiou, H.Y.; Chen, I.H.; Yang, W.C. Analyzing 13 years of cetacean strandings: Multiple stressors to cetaceans in Taiwanese waters and their implications for conservation and future research. Front. Mar. Sci. 2021, 8, 633056. [Google Scholar] [CrossRef]
- IJsseldijk, L.; Brownlow, A.C.; Mazzariol, S. Best practice on cetacean post mortem investigation and tissue sampling. In ACCOBANS–ASCOBANS Joint Guidelines; ACCOBANS: Monaco, 2019; pp. 1–73. [Google Scholar]
- Geraci, J.R.; Lounsbury, V.J. Marine Mammals Ashore: A Field Guide for Strandings, 2nd ed.; National Aquarium in Baltimore: Baltimore, MD, USA, 2005. [Google Scholar]
- Cowan, D.F.; Curry, B.E. Histopathology of the alarm reaction in small odontocetes. J. Comp. Pathol. 2008, 139, 24–33. [Google Scholar] [CrossRef]
- Herráez, P.; Sierra, E.; Arbelo, M.; Jaber, J.R.; Espinosa de los Monteros, A.; Fernández, A. Rhabdomyolysis and myoglobinuric nephrosis (capture myopathy) in a striped dolphin. J. Wildl. Dis. 2007, 43, 770–774. [Google Scholar] [CrossRef]
- Herráez, P.; Espinosa de los Monteros, A.; Fernández, A.; Edwards, J.F.; Sacchini, S.; Sierra, E. Capture myopathy in live-stranded cetaceans. Vet. J. 2013, 196, 181–188. [Google Scholar] [CrossRef]
- Câmara, N.; Sierra, E.; Fernández, A.; Suárez-Santana, C.M.; Puig-Lozano, R.; Arbelo, M.; et al. Skeletal and cardiac rhabdomyolysis in a live-stranded neonatal Bryde’s whale with fetal distress. Front. Vet. Sci. 2019, 6, 239. [Google Scholar] [CrossRef] [PubMed]
- Sierra, E.; Zucca, D.; Arbelo, M.; García-Álvarez, N.; Andrada, M.; Déniz, S.; et al. Fatal systemic morbillivirus infection in bottlenose dolphin, Canary Islands, Spain. Emerg. Infect. Dis. 2014, 20, 269–271. [Google Scholar] [CrossRef]
- Sierra, E.; Sánchez, S.; Saliki, J.T.; Blas-Machado, U.; Arbelo, M.; Zucca, D.; et al. Retrospective study of etiologic agents associated with nonsuppurative meningoencephalitis in stranded cetaceans in the Canary Islands. J. Clin. Microbiol. 2014, 52, 2390–2397. [Google Scholar] [CrossRef] [PubMed]
- Sierra, E.; Fernández, A.; Zucca, D.; Câmara, N.; Felipe-Jiménez, I.; Suárez-Santana, C.; et al. Morbillivirus infection in Risso’s dolphin Grampus griseus: A phylogenetic and pathological study of cases from the Canary Islands. Dis. Aquat. Organ. 2018, 128, 123–136. [Google Scholar] [CrossRef]
- Vandevanter, D.R.; Warrener, P.; Bennett, L.; Schultz, E.R.; Coulter, S.; Garber, R.L.; et al. Detection and analysis of diverse herpesviral species by consensus primer PCR. J. Clin. Microbiol. 1996, 34, 1666–1671. [Google Scholar] [CrossRef]
- Sierra, E.; Fernández, A.; Espinosa de los Monteros, A.; Arbelo, M.; Díaz-Delgado, J.; Andrada, M.; et al. Histopathological muscle findings may be essential for a definitive diagnosis of suspected sharp trauma associated with ship strikes in stranded cetaceans. PLoS ONE 2014, 9, e88780. [Google Scholar] [CrossRef]
- Arregui, M.; Fernández, A.; Paz-Sánchez, Y.; Santana, Á.; Sacchini, S.; Sierra, E.; et al. Comparison of three histological techniques for fat emboli detection in lung cetacean tissue. Sci. Rep. 2020, 10, 18518. [Google Scholar] [CrossRef] [PubMed]
- Ressel, L.; Hetzel, U.; Ricci, E. Blunt force trauma in veterinary forensic pathology. Vet. Pathol. 2016, 53, 941–961. [Google Scholar] [CrossRef]
- West, K.; Silva-Krott, I.; Rotstein, D.; Levine, G. Causes of mortality and pathologic findings in Pacific Island cetaceans: A review of strandings from 2006–2022. In Annual Marine Species Monitoring Report for the Pacific; U.S. Navy Marine Species Monitoring Program: San Diego, CA, USA, 2022. [Google Scholar]
- Cipriani, P.; Palomba, M.; Giulietti, L.; Aco-Alburqueque, R.; Andolfi, R.; Ten Doeschate, M.; et al. Anisakid parasite diversity in a pygmy sperm whale, Kogia breviceps (Cetacea: Kogiidae), stranded at the edge of its distribution range in the Northeast Atlantic Ocean. Parasite 2024, 31, 43. [Google Scholar] [CrossRef]
- Moore, M.; van der Hoop, J.; Barco, S.; Costidis, A.; Gulland, F.; Jepson, P.; et al. Criteria and case definitions for serious injury and death of pinnipeds and cetaceans caused by anthropogenic trauma. Dis. Aquat. Organ. 2013, 103, 229–264. [Google Scholar] [CrossRef] [PubMed]
- Puig-Lozano, R.; Fernández, A.; Saavedra, P.; Tejedor, M.; Sierra, E.; de la Fuente, J.; et al. Retrospective study of traumatic intra-interspecific interactions in stranded cetaceans, Canary Islands. Front. Vet. Sci. 2020, 7, 85. [Google Scholar] [CrossRef] [PubMed]
- Baumgartner, C.D.; Selbmann, A.; Middel, H.; Schulze, J.; Bellon, G.; Samarra, F.I.P. Interactions between killer whales (Orcinus orca) and neonate long-finned pilot whales (Globicephala melas) off South Iceland. Ecol. Evol. 2025, 15, e11023. [Google Scholar] [CrossRef]
- Ochoa, G.M.; Ramos, E.A.; Gonzalez-Socoloske, D. Predation of a sperm whale (Physeter macrocephalus) by killer whales (Orcinus orca) in Guanaja, Honduras. Aquat. Mamm. 2025, 51, 31–36. [Google Scholar] [CrossRef]
- Ferguson, S.H.; Higdon, J.W.; Chmelnitsky, E.G. The rise of killer whales as a major Arctic predator. In A Little Less Arctic; Springer: Dordrecht, The Netherlands, 2010; pp. 117–136. [Google Scholar]
- Ferguson, S.H.; Kingsley, M.C.S.; Higdon, J.W. Killer whale (Orcinus orca) predation in a multi-prey system. Popul. Ecol. 2012, 54, 31–41. [Google Scholar] [CrossRef]
- Pitman, R.L.; Ballance, L.T.; Mesnick, S.I.; Chivers, S.J. Killer whale predation on sperm whales: Observations and implications. Mar. Mamm. Sci. 2001, 17, 494–507. [Google Scholar] [CrossRef]
- Seyboth, E.; Bassoi, M.; de Lima, R.C.; do Prado, J.H.F. Killer whale predation on an Antarctic minke whale in the northern Antarctic Peninsula. An. Acad. Bras. Cienc. 2024, 96, e20231345. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, T.A.; Stacey, P.J.; Baird, R.W. A review of killer whale interactions with other marine mammals: Predation to co-existence. Mamm. Rev. 1991, 21, 151–180. [Google Scholar] [CrossRef]
- Olson, P.A.; Hancock-Hanser, B.L.; McCullough, J.L.K.; Dahlheim, M.; Schulman-Janiger, A.; Black, N.; et al. Killer whale predation on a pygmy sperm whale in an oceanic habitat. Mar. Mamm. Sci. 2023, 39, 281–288. [Google Scholar] [CrossRef]
- Dunphy-Daly, M.M.; Heithaus, M.R.; Claridge, D.E. Temporal variation in dwarf sperm whale (Kogia sima) habitat use and group size off Great Abaco Island, Bahamas. Mar. Mamm. Sci. 2008, 24, 171–184. [Google Scholar] [CrossRef]
- Martín, V.; Tejedor, M.; Carrillo, M.; Pérez-Gil, M.; Arbelo, M.; Servidio, A.; et al. Strandings and at-sea observations reveal the Canary Archipelago as an important habitat for pygmy and dwarf sperm whales. In Advances in Marine Mammal Research; Springer: Cham, Switzerland, 2024; pp. 21–64. [Google Scholar]
- Fernández, A.; Câmara, N.; Sierra, E.; Arbelo, M.; Bernaldo de Quirós, Y.; Jepson, P.D.; et al. Cetacean intracytoplasmic eosinophilic globules: A cytomorphological, histological, histochemical, immunohistochemical, and proteomic characterization. Animals 2023, 13, 2034. [Google Scholar] [CrossRef]
- Fernández, A.; Sierra, E.; Díaz-Delgado, J.; Sacchini, S.; Sánchez-Paz, Y.; Suárez-Santana, C.; et al. Deadly acute decompression sickness in Risso’s dolphins. Sci. Rep. 2017, 7, 17254. [Google Scholar] [CrossRef]
- de Quirós, Y.B.; González-Díaz, O.; Arbelo, M.; Sierra, E.; Sacchini, S.; Fernández, A. Decompression vs. decomposition: Distribution, amount, and gas composition of bubbles in stranded marine mammals. Front. Physiol. 2012, 3, 177. [Google Scholar] [CrossRef]
- Pons-Bordas, C.; Hazenberg, A.; Hernandez-Gonzalez, A.; Pool, R.V.; Covelo, P.; Sánchez-Hermosin, P.; et al. Recent increase of ulcerative lesions caused by Anisakis spp. in cetaceans from the north-east Atlantic. J. Helminthol. 2020, 94, e127. [Google Scholar] [CrossRef]
- Motta, M.R.A.; Pinheiro, D.C.S.N.; Carvalho, V.L.; Viana, D.A.; Vicente, A.C.P.; Iñiguez, A.M. Gastric lesions associated with the presence of Anisakis spp. Dujardin, 1845 (Nematoda: Anisakidae) in cetaceans stranded on the coast of Ceará, Brazil. Biota Neotrop. 2008, 8, 91–95. [Google Scholar] [CrossRef]
- Hrabar, J.; Bočina, I.; Gudan Kurilj, A.; Đuras, M.; Mladineo, I. Gastric lesions in dolphins stranded along the eastern Adriatic coast. Dis. Aquat. Organ. 2017, 125, 125–139. [Google Scholar] [CrossRef]
- Nakagun, S.; Kobayashi, Y. Histochemical and immunohistochemical characterizations of hepatic trematodiasis in odontocetes. Front. Vet. Sci. 2020, 7, 375. [Google Scholar] [CrossRef]
- Jaber, J.R.; Pérez, J.; Arbelo, M.; Andrada, M.; Hidalgo, M.; Gómez-Villamandos, J.C.; et al. Hepatic lesions in cetaceans stranded in the Canary Islands. Vet. Pathol. 2004, 41, 147–153. [Google Scholar] [CrossRef]
- Segura-Göthlin, S.; Fernández, A.; Arbelo, M.; Andrada Borzollino, M.A.; Felipe-Jiménez, I.; Colom-Rivero, A.; et al. Viral skin diseases in odontocete cetaceans: Gross, histopathological, and molecular characterization of selected pathogens. Front. Vet. Sci. 2023, 10, 1169301. [Google Scholar] [CrossRef] [PubMed]
- Martineau, D.; Lagacé, A.; Béland, P.; Higgins, R.; Armstrong, D.; Shugart, L.R. Pathology of stranded beluga whales (Delphinapterus leucas) from the St. Lawrence Estuary, Québec, Canada. J. Comp. Pathol. 1988, 98, 287–310. [Google Scholar] [CrossRef]
- Sacristán, C.; Esperón, F.; Ewbank, A.C.; Díaz-Delgado, J.; Ferreira-Machado, E.; Costa-Silva, S.; et al. Novel herpesviruses in riverine and marine cetaceans from South America. Acta Trop. 2019, 190, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Manire, C.A.; Smolarek, K.A.; Romero, C.H.; Kinsel, M.J.; Clauss, T.M.; Byrd, L. Proliferative dermatitis associated with a novel alphaherpesvirus in an Atlantic bottlenose dolphin (Tursiops truncatus). J. Zoo Wildl. Med. 2006, 37, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Felipe-Jiménez, I.; Fernández, A.; Andrada, M.; Arbelo, M.; Segura-Göthlin, S.; Colom-Rivero, A.; et al. Contribution to herpesvirus surveillance in beaked whales stranded in the Canary Islands. Animals 2021, 11, 3170. [Google Scholar] [CrossRef]
- Sacristán, C.; Ewbank, A.C.; Duarte-Benvenuto, A.; Sacristán, I.; Zamana-Ramblas, R.; Costa-Silva, S.; et al. Survey of selected viral agents (herpesvirus, adenovirus and hepatitis E virus) in liver and lung samples of cetaceans, Brazil. Sci. Rep. 2024, 14, 2689. [Google Scholar] [CrossRef]
- Smolarek Benson, K.A.; Manire, C.A.; Ewing, R.Y.; Saliki, J.T.; Townsend, F.I.; Ehlers, B.; et al. Identification of novel alpha- and gammaherpesviruses from cutaneous and mucosal lesions of dolphins and whales. J. Virol. Methods 2006, 136, 261–266. [Google Scholar] [CrossRef]
- Uzal, F.A.; Navarro, M.A.; Asin, J.; Henderson, E.E. Clostridial diseases of horses: A review. Vaccines 2022, 10, 1143. [Google Scholar] [CrossRef] [PubMed]
- Uzal, F.A.; Songer, J.G. Clostridial diseases. In Diseases of Swine, 11th ed.; Wiley: Hoboken, NJ, USA, 2019; pp. 792–806. [Google Scholar]
- Buck, J.D.; Shepard, L.L.; Spotte, S. Clostridium perfringens as the cause of death of a captive Atlantic bottlenosed dolphin (Tursiops truncatus). J. Wildl. Dis. 1987, 23, 488–491. [Google Scholar] [CrossRef]
- Driscoll, P.; Pierce, V.; Whitaker, B.; Kelly, K. Multi-agency case report: Clostridium perfringens disseminated infection in a live-stranded bottlenose dolphin (Tursiops truncatus) in Maryland. In Proceedings of the International Association for Aquatic Animal Medicine; IAAM: Seattle, WA, USA, 2007. [Google Scholar]
- Danil, K.; St. Leger, J.; Dennison, S.; Bernaldo de Quirós, Y.; Scadeng, M.; Nilson, E.; et al. Clostridium perfringens septicemia in a long-beaked common dolphin Delphinus capensis: An etiology of gas bubble accumulation in cetaceans. Dis. Aquat. Organ. 2014, 111, 183–190. [Google Scholar] [CrossRef]
- Gulland, F.; Dierauf, L.; Whitman, K. (Eds.) Handbook of Marine Mammal Medicine: Health, Disease, and Rehabilitation, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- St. Leger, J.; Raverty, S.; Mena, A. Cetacea. In Pathology of Wildlife and Zoo Animals; Elsevier: Amsterdam, The Netherlands, 2018; pp. 461–514. [Google Scholar]
- Hutchinson, K.M.; Tart, K.; Anderson, K.L.; Powell, L.L. Pneumatosis of the intestines, colon and liver in a young cat. Vet. Med. Sci. 2018, 4, 150–158. [Google Scholar] [CrossRef]
- Kiu, R.; Shaw, A.G.; Sim, K.; Acuna-Gonzalez, A.; Price, C.A.; Bedwell, H.; et al. Particular genomic and virulence traits associated with preterm infant-derived toxigenic Clostridium perfringens strains. Nat. Microbiol. 2023, 8, 1160–1175. [Google Scholar] [CrossRef]
- Neto, G.G.; Bueno, M.G.; Silva, R.O.S.; Lobato, F.C.F.; Guimarães, J.P.; Bossart, G.D.; et al. Acute necrotizing colitis with pneumatosis intestinalis in an Amazonian manatee calf. Dis. Aquat. Organ. 2016, 120, 189–194. [Google Scholar] [CrossRef]
- Rich, A.F.; Zendri, F.; Costa, T.; Timofte, D.; Drake, G.J.; Rowland, H.; et al. Nectarivorous bird emphysematous ingluvitis (NBEI): A novel disease in Loriinae birds associated with Clostridium perfringens infection. Front. Vet. Sci. 2020, 7, 606112. [Google Scholar] [CrossRef] [PubMed]
- Šeol, B.; Gomerčić, M.Đ.; Naglić, T.; Gomerčić, T.; Galov, A.; Gomerčić, H. Isolation of Clostridium tertium from a striped dolphin (Stenella coeruleoalba) in the Adriatic Sea. J. Wildl. Dis. 2006, 42, 709–711. [Google Scholar] [CrossRef]
- Díaz-Delgado, J.; Fernández, A.; Xuriach, A.; Sierra, E.; Bernaldo de Quirós, Y.; Mompeo, B.; et al. Verminous arteritis due to Crassicauda sp. in Cuvier’s beaked whales (Ziphius cavirostris). Vet. Pathol. 2016, 53, 1233–1240. [Google Scholar] [CrossRef] [PubMed]
- Jabbar, A.; Beveridge, I.; Bryant, M.S. Morphological and molecular observations on the status of Crassicauda magna, a parasite of the subcutaneous tissues of the pygmy sperm whale, with a re-evaluation of the systematic relationships of the genus Crassicauda. Parasitol. Res. 2015, 114, 835–841. [Google Scholar] [CrossRef] [PubMed]
- Zucca, P.; Di Guardo, G.; Pozzi-Mucelli, R.; Scaravelli, D.; Francese, M. Use of computer tomography for imaging of Crassicauda grampicola in a Risso’s dolphin (Grampus griseus). J. Zoo Wildl. Med. 2004, 35, 391–394. [Google Scholar] [CrossRef]
- Suárez-González, Z.; Fernández, A.; Colom-Rivero, A.; García-Álvarez, N.; González, J.F.; Sierra, E. Morphological and molecular characterization of Crassicauda anthonyi in Cuvier’s beaked whales from the Canary Islands. BMC Vet. Res. 2025, 21, 148. [Google Scholar] [CrossRef] [PubMed]
- Lempereur, L.; Delobelle, M.; Doom, M.; Haelters, J.; Levy, E.; Losson, B.; et al. Crassicauda boopis in a fin whale (Balaenoptera physalus) ship-struck in the eastern North Atlantic Ocean. Parasitol. Open 2017, 3, e12. [Google Scholar] [CrossRef]
- Kot, B.C.; Ho, H.H.; Leung, E.K.; Chung, T.Y.; Tsui, H.C. Characterisation of Crassicauda fuelleborni nematode infection in Indo-Pacific finless porpoises (Neophocaena phocaenoides) using postmortem computed tomography. Int. J. Parasitol. Parasites Wildl. 2022, 18, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Kot, B.C.W.; Ho, H.H.N.; Leung, E.K.C.; Chung, T.YT.; Tsui, H.C.L. Characterisation of Crassicauda fuelleborni nematode infection in Indo-Pacific finless porpoises (Neophocaena phocaenoides) using postmortem computed tomography. Int. J. Parasitol. Parasites Wildl. 2022, 18, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Lambertsen, R.H. Taxonomy and distribution of a Crassicauda species (Nematoda: Spirurida) infecting the kidney of the common fin whale (Balaenoptera physalus Linné, 1758). J. Parasitol. 1985, 71, 485–488. [Google Scholar] [CrossRef]
- Robinson, W.F.; Robinson, N.A. Cardiovascular system. In Jubb, Kennedy & Palmer’s Pathology of Domestic Animals; Maxie, M.G., Ed.; Elsevier: St. Louis, MO, USA, 2016; Vol. 3, pp. 1–101.e1. [Google Scholar]
- Bossart, G.D.; Odell, D.K.; Altman, N.H. Cardiomyopathy in stranded pygmy and dwarf sperm whales. J. Am. Vet. Med. Assoc. 1985, 187, 1137–1140. [Google Scholar] [CrossRef]
- Bryan, C.E.; Bossart, G.D.; Christopher, S.J.; Davis, W.C.; Kilpatrick, L.E.; McFee, W.E.; et al. Selenium protein identification and profiling by mass spectrometry: A tool to assess progression of cardiomyopathy in a whale model. J. Trace Elem. Med. Biol. 2017, 44, 40–49. [Google Scholar] [CrossRef]
- Sierra, E.; de los Monteros, A.E.; Fernández, A.; Arbelo, M.; Caballero, M.J.; Rivero, M.; et al. Sarcoplasmic masses in the skeletal muscle of a stranded pigmy sperm whale (Kogia breviceps). J. Wildl. Dis. 2013, 49, 679–683. [Google Scholar] [CrossRef]
- Abbas, A.; Kumar, V.; Aster, J. Environmental and nutritional diseases. In Robbins & Cotran Pathologic Basis of Disease, 10th ed.; Elsevier: Philadelphia, PA, USA, 2021; pp. 391–436. [Google Scholar]
- IJsseldijk, L.L.; Hessing, S.; Mairo, A.; ten Doeschate, M.T.I.; Treep, J.; van den Broek, J.; et al. Nutritional status and prey energy density govern reproductive success in a small cetacean. Sci. Rep. 2021, 11, 19201. [Google Scholar] [CrossRef]
- Puig-Lozano, R.; Bernaldo de Quirós, Y.; Díaz-Delgado, J.; García-Álvarez, N.; Sierra, E.; de la Fuente, J.; et al. Retrospective study of foreign body-associated pathology in stranded cetaceans, Canary Islands (2000–2015). Environ. Pollut. 2018, 243, 1840–1852. [Google Scholar] [CrossRef]
- Pallin, L.J.; Garrigue, C.; Kellar, N.M.; Baker, C.S.; Bonneville, C.D.; Derville, S.; et al. Demographic and physiological signals of reproductive events in humpback whales on a southwest Pacific breeding ground. Conserv. Physiol. 2024, 12, coae002. [Google Scholar] [CrossRef]
- Raverty, S.; St. Leger, J.; Noren, D.P.; Burek Huntington, K.; Rotstein, D.S.; Gulland, F.M.D.; et al. Pathology findings and correlation with body condition index in stranded killer whales (Orcinus orca) in the northeastern Pacific and Hawaii from 2004 to 2013. PLoS ONE 2020, 15, e0242505. [Google Scholar] [CrossRef]
- Albrecht, S.; Minto, C.; Rogan, E.; Deaville, R.; O’Donovan, J.; Daly, M.; et al. Emaciated enigma: Decline in body conditions of common dolphins in the Celtic Seas ecoregion. Ecol. Evol. 2024, 14, e70325. [Google Scholar] [CrossRef] [PubMed]
- Kebke, A.; Samarra, F.; Derous, D. Climate change and cetacean health: Impacts and future directions. Philos. Trans. R. Soc. B 2022, 377, 20210147. [Google Scholar] [CrossRef]
- Owen, K.; Thompson, R.; Donnelly, D.; Noad, M.; Bury, S.; Pinkerton, M.; et al. Southern Ocean humpback whale trophic ecology. II. Influence of fasting and opportunistic feeding on skin stable isotope values of migrating whales. Mar. Ecol. Prog. Ser. 2024, 734, 157–171. [Google Scholar] [CrossRef]
- Van Aswegen, M.; Szabo, A.; Currie, J.J.; Stack, S.H.; West, K.L.; Hofmann, N.; et al. Energetic cost of gestation and prenatal growth in humpback whales. J. Physiol. 2025, 603, 529–550. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, F.; Madsen, P.T.; Andrews-Goff, V.; Double, M.C.; How, J.R.; Clapham, P.; et al. Extreme capital breeding for giants: Effects of body size on humpback whale energy expenditure and fasting endurance. Ecol. Modell. 2025, 501, 110994. [Google Scholar] [CrossRef]
- McHuron, E.A.; Adamczak, S.; Costa, D.P.; Booth, C. Estimating reproductive costs in marine mammal bioenergetic models: A review of current knowledge and data availability. Conserv. Physiol. 2023, 11, coac080. [Google Scholar] [CrossRef]
- Martínez-Burnes, J.; Mota-Rojas, D.; Villanueva-García, D.; Ibarra-Ríos, D.; Lezama-García, K.; Barrios-García, H.; et al. Meconium aspiration syndrome in mammals. CAB Rev. 2019, 14, 1–13. [Google Scholar] [CrossRef]
- Colegrove, K.M.; Venn-Watson, S.; Litz, J.; Kinsel, M.J.; Terio, K.A.; Fougeres, E.; et al. Fetal distress and in utero pneumonia in perinatal dolphins during the Northern Gulf of Mexico unusual mortality event. Dis. Aquat. Organ. 2016, 119, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Groch, K.; Catão-Dias, J.; Kolesnikovas, C.; de Castilho, P.; Moreira, L.; et al. Pathologic findings and causes of death in southern right whales Eubalaena australis, Brazil. Dis. Aquat. Organ. 2019, 137, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Laist, D.W.; Knowlton, A.R.; Mead, J.G.; Collet, A.S.; Podesta, M. Collisions between ships and whales. Mar. Mamm. Sci. 2001, 17, 35–75. [Google Scholar] [CrossRef]
- Ritter, F.; Panigada, S. Collisions of vessels with cetaceans—The underestimated threat. In World Seas: An Environmental Evaluation; Elsevier: Amsterdam, The Netherlands, 2019; pp. 531–547. [Google Scholar]
- Schoeman, R.P.; Patterson-Abrolat, C.; Plön, S. A global review of vessel collisions with marine animals. Front. Mar. Sci. 2020, 7, 292. [Google Scholar] [CrossRef]
- Manuel, C.; Ritter, F. Increasing numbers of ship strikes in the Canary Islands: Proposals for immediate action to reduce risk of vessel–whale collisions. J. Cetacean Res. Manag. 2010, 11, 131–136. [Google Scholar]
- Arregui, M.; de Quirós, Y.B.; Saavedra, P.; Sierra, E.; Suárez-Santana, C.M.; Arbelo, M.; et al. Fat embolism and sperm whale ship strikes. Front. Mar. Sci. 2019, 6, 379. [Google Scholar] [CrossRef]
- Peel, D.; Smith, J.N.; Childerhouse, S. Vessel strike of whales in Australia: The challenges of analysis of historical incident data. Front. Mar. Sci. 2018, 5, 69. [Google Scholar] [CrossRef]
- Izadi, S.; Johnson, M.; de Soto, N.A.; Constantine, R. Night-life of Bryde’s whales: Ecological implications of resting in a baleen whale. Behav. Ecol. Sociobiol. 2018, 72, 130. [Google Scholar] [CrossRef]
- Neilson, J.L.; Gabriele, C.M.; Jensen, A.S.; Jackson, K.; Straley, J.M. Summary of reported whale-vessel collisions in Alaskan waters. J. Mar. Biol. 2012, 2012, 106282. [Google Scholar] [CrossRef]
- Winkler, C.; Murphy, S.; Ritter, F.; Panigada, S.; Murphy, S. Global numbers of ship strikes: An assessment of collisions between vessels and cetaceans using available data in the IWC Ship Strike Database. Report to the International Whaling Commission, 2020. Available online: www.m-e-e-r.org (accessed on 12 January 2026).
- Fais, A.; Lewis, T.P.; Zitterbart, D.P.; Álvarez, O.; Tejedor, A.; Soto, N.A. Erratum: Abundance and distribution of sperm whales in the Canary Islands: Can sperm whales in the archipelago sustain the current level of ship-strike mortalities? PLoS ONE 2016, 11, e0150660. [Google Scholar]
- Constantine, R.; Johnson, M.; Riekkola, L.; Jervis, S.; Kozmian-Ledward, L.; Dennis, T.; et al. Mitigation of vessel-strike mortality of endangered Bryde’s whales in the Hauraki Gulf, New Zealand. Biol. Conserv. 2015, 186, 149–157. [Google Scholar] [CrossRef]
- Campbell-Malone, R.; Barco, S.G.; Daoust, P.-Y.; Knowlton, A.R.; McLellan, W.A.; Rotstein, D.S.; et al. Gross and histologic evidence of sharp and blunt trauma in North Atlantic right whales (Eubalaena glacialis) killed by vessels. J. Zoo Wildl. Med. 2008, 39, 37–55. [Google Scholar] [CrossRef]
- Moore, M.J.; van der Hoop, J.; Barco, S.G.; Costidis, A.M.; Gulland, F.M.; Jepson, P.D.; et al. Criteria and case definitions for serious injury and death of pinnipeds and cetaceans caused by anthropogenic trauma. Dis. Aquat. Organ. 2013, 103, 229–264. [Google Scholar] [CrossRef]
- De Siqueira, A.; Cuevas, S.E.C.; Salvagni, F.A.; Maiorka, P.C. Forensic veterinary pathology. Vet. Pathol. 2016, 53, 979–987. [Google Scholar] [CrossRef]
- International Whaling Commission. Bycatch and entanglement of cetaceans in fishing. Available online: https://iwc.int/bycatch (accessed on 12 January 2026).
- Masere, C.; Baird, K. An initial exploration of cetacean bycatch and interactions in the WCPFC; Western and Central Pacific Fisheries Commission (WCPFC).
- Read, A.J.; Drinker, P.; Northridge, S. Bycatch of marine mammals in U.S. and global fisheries. Conserv. Biol. 2006, 20, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Puig-Lozano, R.; Fernández, A.; Sierra, E.; Saavedra, P.; Suárez-Santana, C.M.; de la Fuente, J.; et al. Retrospective study of fishery interactions in stranded cetaceans, Canary Islands. Front. Vet. Sci. 2020, 7, 571. [Google Scholar] [CrossRef]
- Reeves, R.; McClellan, K.; Werner, T. Marine mammal bycatch in gillnet and other entangling net fisheries, 1990 to 2011. Endang. Species Res. 2013, 20, 71–97. [Google Scholar] [CrossRef]
- Martino, L.; Leiva Forns, M.; Cid Cañete, M.; Pérez, L.; Pradas, C.; Domingo, M. Bycatch in cetaceans from the north-western Mediterranean Sea: Retrospective study of lesions and utility of bycatch criteria. Vet. Sci. 2025, 12, 711. [Google Scholar] [CrossRef]
- Elliott, B.; Kiszka, J.J.; Bonhommeau, S.; Shahid, U.; Nelson, L.; Read, A.J. Bycatch in drift gillnet fisheries: A sink for Indian Ocean cetaceans. Indian Ocean Tuna Commission (IOTC), 1992. Available online: https://iotc.org/cmms (accessed on 12 January 2026).
- Basran, C.J.; Sigurðsson, G.M. Using case studies to investigate cetacean bycatch/interaction under-reporting in countries with reporting legislation. Front. Mar. Sci. 2021, 8, 788562. [Google Scholar] [CrossRef]
- Segniagbeto, G.H.; Van Waerebeek, K.; Bowessidjaou, J.E.; Ketoh, K.; Kpatcha, T.K.; Okoumassou, K.; et al. Annotated checklist and fisheries interactions of cetaceans in Togo, with evidence of Antarctic minke whale in the Gulf of Guinea. Integr. Zool. 2014, 9, 1–13. [Google Scholar] [CrossRef]
- Moura, J.F.; Acevedo-Trejos, E.; Tavares, D.C.; Meirelles, A.C.O.; Silva, C.P.N.; Oliveira, L.R.; et al. Stranding events of Kogia whales along the Brazilian coast. PLoS ONE 2016, 11, e0146108. [Google Scholar] [CrossRef]
- Cardona-Maldonado, M.A.; Mignucci-Giannoni, A. Pygmy and dwarf sperm whales in Puerto Rico and the Virgin Islands, with a review of Kogia in the Caribbean. Caribb. J. Sci. 1999, 35, 29–37. [Google Scholar]
- Eisfeld-Pierantonio, S.M.; Pierantonio, N.; Simmonds, M.P. The impact of marine debris on cetaceans with consideration of plastics generated by the COVID-19 pandemic. Environ. Pollut. 2022, 300, 118967. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, Y.; Silva, M.A.; Pham, C.K.; Duncan, E.M. Cetaceans playing with single-use plastics (SUPs): A widespread interaction with likely severe impacts. Mar. Pollut. Bull. 2023, 194, 115428. [Google Scholar] [CrossRef] [PubMed]
- Alexiadou, P.; Foskolos, I.; Frantzis, A. Ingestion of macroplastics by odontocetes of the Greek Seas, Eastern Mediterranean: Often deadly! Mar. Pollut. Bull. 2019, 146, 67–75. [Google Scholar] [CrossRef]
- Zimmer-Correa, M.; Carneiro Proietti, M.; Couto Di Tullio, J.; Rodrigues, L.S.; Quadro Oreste, E.; Kessler, F.; et al. Plastic ingestion by odontocetes from the Western South Atlantic: A particular concern to a threatened species. Environ. Pollut. 2024, 360, 124659. [Google Scholar] [CrossRef]
- Redaelli, L.; Alcázar-Treviño, J.; Escánez, A.; Fernandez, M.; Aguilar de Soto, N.; Alves, F.; et al. Acoustic mimicry of plastics with the prey of deep-cetaceans: An experimental approach. In Proceedings of the International Conference on Underwater Acoustics, 2024; Institute of Acoustics. [Google Scholar]
- Whitaker, B.R.; Geraci, J.R.; St. Aubin, D.J. The near-fatal ingestion of plastic by a pygmy sperm whale, Kogia breviceps. VIN document, 1994. Available online: https://www.vin.com/doc/?id=3864102 (accessed on 12 January 2026).
- West, K.; Silva-Krott, I.; Rotstein, D.; Levine, G. U.S. Navy Marine Species Monitoring Program. Causes of mortality and pathologic findings in Pacific Island cetaceans. 2023. Available online: https://www.navymarinespeciesmonitoring.us/index.php/download_file/view/3092/ (accessed on 12 January 2026).
- Knowlton, A.R.; Kraus, S.D. Mortality and serious injury of northern right whales (Eubalaena glacialis) in the western North Atlantic Ocean. J. Cetacean Res. Manag. 2020, 21, 193–208. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



