1. Introduction
Photosynthesis is the fundamental biological process sustaining almost all life on Earth by converting light energy into chemical energy. Yet, it harbors a paradox, as harvesting light energy renders plants highly susceptible to damage from excess light [
1,
2,
3,
4,
5,
6,
7]. When light absorption surpasses the capacity of carbon assimilation, the photosynthetic apparatus becomes over-excited, triggering reactive oxygen species (ROS) generation through photo-oxidation. This photo-oxidative stress severely restricts plant productivity and survival, acting as a critical bottleneck in the global carbon cycle and agricultural yield.
This paradigm has shifted dramatically. Research reveals ROS, particularly H₂O₂ and ¹O₂, as integral components of sophisticated signaling networks [
8,
9]. This review synthesizes recent advances in understanding the molecular mechanisms, damage, and adaptive strategies associated with photo-oxidative stress in plants.
2. ROS Generation, Spatiotemporal Signaling, and Retrograde Communication
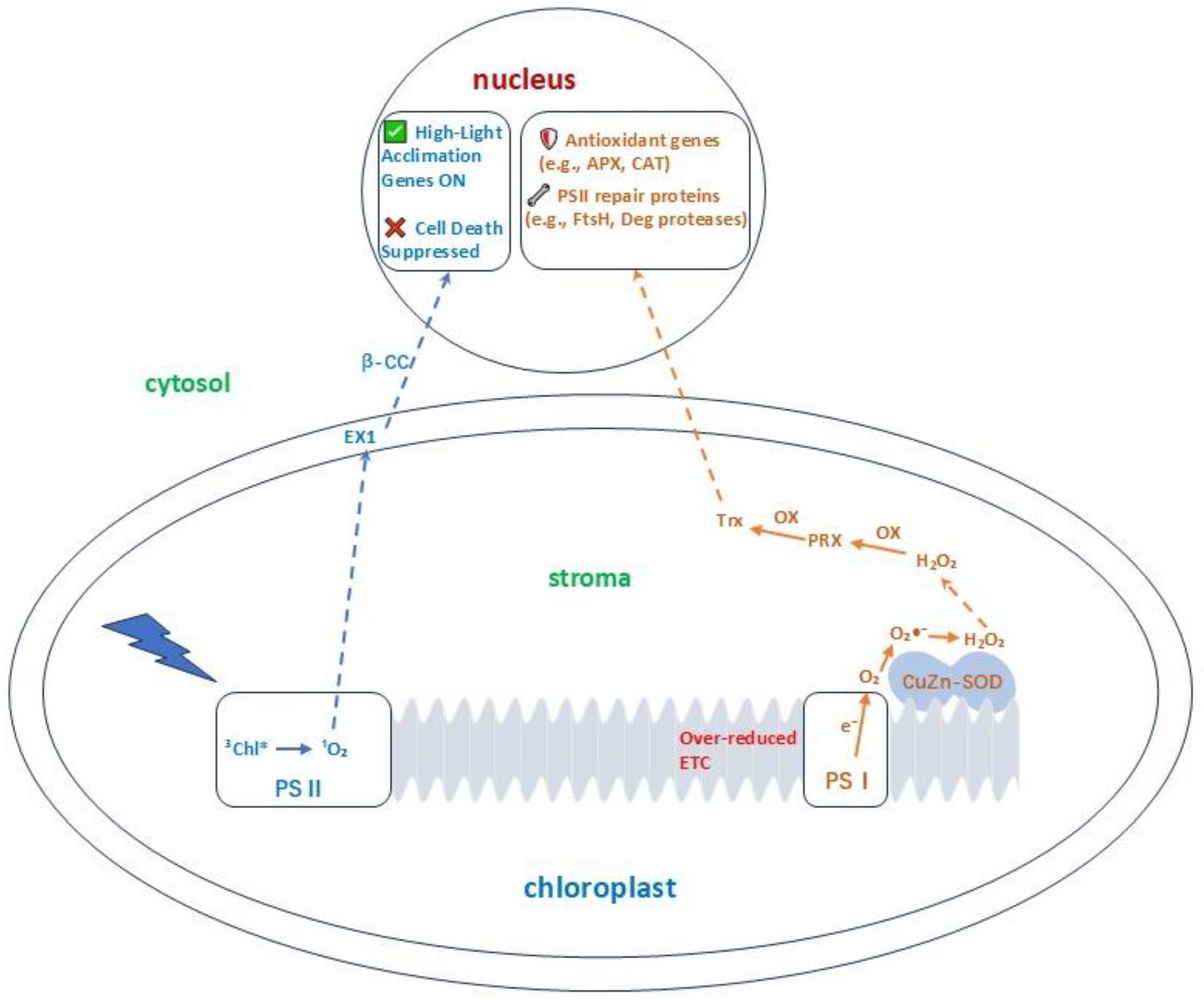
The phenomenon of photo-oxidation is fundamentally rooted in the generation and management of ROS. Once considered merely toxic byproducts of aerobic metabolism, ROS are now recognized as pivotal signaling molecules that coordinate plant responses to environmental challenges. Understanding their precise sites of production, the specificity of their signals, and their role in systemic communication is essential to appreciate the complexity of photo-oxidative stress management (
Figure 1).
2.1. Primary ROS and Their Specific Production Sites
ROS are inevitable by-products of electron transport in multiple cellular compartments, with chloroplasts as the primary source under photo-oxidative conditions. In the photosynthetic electron transport chain, PSII produces singlet oxygen (¹O₂) via the interaction of O₂ with the excited triplet state of chlorophyll (³Chl) when absorbed energy cannot be used for photochemistry [
10,
11]. ¹O₂ is highly reactive with a short half-life, confining its effects to the thylakoid membranes. PSI generates superoxide anions (O₂•⁻) through the Mehler reaction, where electrons due to an over-reduced electron transport chain are transferred to O₂ instead of ferredoxin [
12,
13,
14,
15]. Superoxide is rapidly dismutated (spontaneously or by superoxide dismutase, SOD) to form H₂O₂, which is more stable and diffuses over longer distances, serving as a versatile signaling molecule.
Beyond chloroplasts, peroxisomes are major H₂O₂ sources during photorespiration, which is a process linked to photosynthesis that intensifies under CO₂-limiting conditions [
16]. The glycolate oxidase reaction in photorespiration is a key H₂O₂ generator. The apoplast (cell wall space) also produces ROS via plasma membrane-localized NADPH oxidases (Respiratory Burst Oxidase Homologs, RBOHs), which are activated by stress to produce O₂•⁻, later converted to H₂O₂ [
17]. This apoplastic ROS wave is critical for systemic signaling, alerting distant tissues to localized stress.
2.2. Specificity of ROS Signals
A key advancement in plant biology is recognizing that different ROS species trigger distinct genetic and metabolic programs, allowing plants to fine-tune responses to stress type and intensity.
¹O₂, primarily from PSII, initiates unique signaling pathways leading to programmed cell death or acclimation, depending on flux [
11]. Nuclear-encoded chloroplast proteins EXECUTER1 (EX1) and EXECUTER2 (EX2) are central mediators. ¹O₂ oxidation of EX1 triggers retrograde signals that reprogram nuclear gene expression, activating genes involved in cell death suppression and high-light acclimation, which is distinct from H₂O₂-induced responses [
18,
19].
H₂O₂, with greater stability and mobility, acts as a broader signal. Its specificity stems from multiple mechanisms, including the oxidation of specific cysteine residues on target proteins (e.g., TGA and NPR1 transcription factors) which alters their function [
9]; spatial origin influences transcriptional profiles (e.g., chloroplast-derived H₂O₂ vs. apoplastic H₂O₂ from RBOHs); and signal intensity/duration matters, with transient pulses activating acclimation, while sustained bursts cause irreversible damage [
17,
20].
2.3. Retrograde Signaling
Retrograde signaling enables chloroplasts to communicate metabolic status to the nucleus, with ROS as pivotal messengers relaying photo-oxidative stress information to orchestrate nuclear gene expression.
¹O₂ from PSII, due to high reactivity and short half-life, does not reach the nucleus directly. Instead, it triggers a chloroplast-localized signaling cascade mediated by EX1 and EX2 [
18]. ¹O₂ oxidation of EX1 (embedded in thylakoid membranes) induces conformational changes or degradation, initiating downstream signaling involving chloroplast kinases and secondary messengers like β-cyclocitral (β-CC), a carotenoid oxidation derivative that accumulates under ¹O₂ stress and modulates nuclear stress-acclimation genes [
21]. This ¹O₂-EX1 module activates a specific genetic program suppressing programmed cell death and promoting photoacclimation [
10].
However, H₂O₂ converted by PSI-generated superoxide rapidly can diffuse through chloroplast envelope aquaporins. Genetically encoded H₂O₂ sensors show high light stress induces chloroplast H₂O₂ accumulation, followed by nuclear increases, indicating direct cytosolic traversal to influence nuclear components [
3]. H₂O₂ signaling involves oxidation of thiol groups on cytosolic and nuclear target proteins. Peroxiredoxins (PRXs) transfer oxidative equivalents to transcription factors [
22], with EX2 as a key downstream component modulating stress-responsive gene expression. This network regulates nuclear-encoded chloroplast proteins (e.g., antioxidant enzymes, PsbS) to restore chloroplast homeostasis [
23].
ROS signals also interact with classic retrograde signals, such as the plastoquinone pool redox state and metabolites like 3’-phosphoadenosine 5’-phosphate (PAP) and methylerythritol cyclodiphosphate (MEcPP). For example, MEcPP induced by high light activates stress-response genes and intersects with ROS pathways to fine-tune acclimation, ensuring nuclear gene expression matches the specific photo-oxidative stress intensity [
23,
24].
2.4. Integration with Hormonal Signaling Networks
The intricate crosstalk between ROS and phytohormone pathways forms a sophisticated, integrated signaling network essential for tailoring plant stress responses. This system is highly dynamic, with the nature of the interaction varying significantly depending on the specific stress combination, allowing the plant to prioritize and fine-tune its acclimation strategies [
17].
In abiotic stress responses, a potent positive feedback loop between abscisic acid (ABA) and hydrogen peroxide (H₂O₂) is central. ABA enhances the stability of systemic ROS waves by upregulating Respiratory Burst Oxidase Homolog (RBOH) gene expression, thereby amplifying the acclimation signal. Conversely, H₂O₂ can influence ABA biosynthesis and signaling. This reciprocal reinforcement is critical under conditions like drought and high light, where ABA-induced stomatal closure limits CO₂ intake. This promotes photosynthetic electron transport chain (PETC) over-reduction and subsequent H₂O₂ production at PSI, which further activates ABA pathways, reinforcing stomatal closure and shared stress-gene expression [
1,
17].
Biotic stress responses are orchestrated by more specific ROS-hormone pairings. Salicylic acid (SA)-mediated systemic acquired resistance (SAR) against biotrophic pathogens is closely linked to H₂O₂ signaling [
25,
26]. In contrast, ¹O₂ has been shown to activate jasmonic acid (JA)-mediated defenses, which are crucial against necrotrophs and herbivores. This specificity ensures an appropriate defense response; for instance, ¹O₂ generated during photo-oxidative stress can pre-emptively prime JA-dependent defenses, anticipating potential opportunistic necrotrophic infection following cellular damage [
1].
The integration of these pathways reveals a context-dependent hierarchy. Under combined stress, such as high light with pathogen attack, initial ¹O₂ from damaged photosystem II may prime JA signaling against necrotrophs. A subsequent biotrophic infection, however, could trigger a dominant SA response that antagonistically suppresses the JA pathway, illustrating the complex and sometimes conflicting priorities within the network [
27]. This regulatory hierarchy ensures the plant’s limited resources are allocated toward the most immediate threat, demonstrating the precision and adaptability of the interconnected ROS-phytohormone communication system.
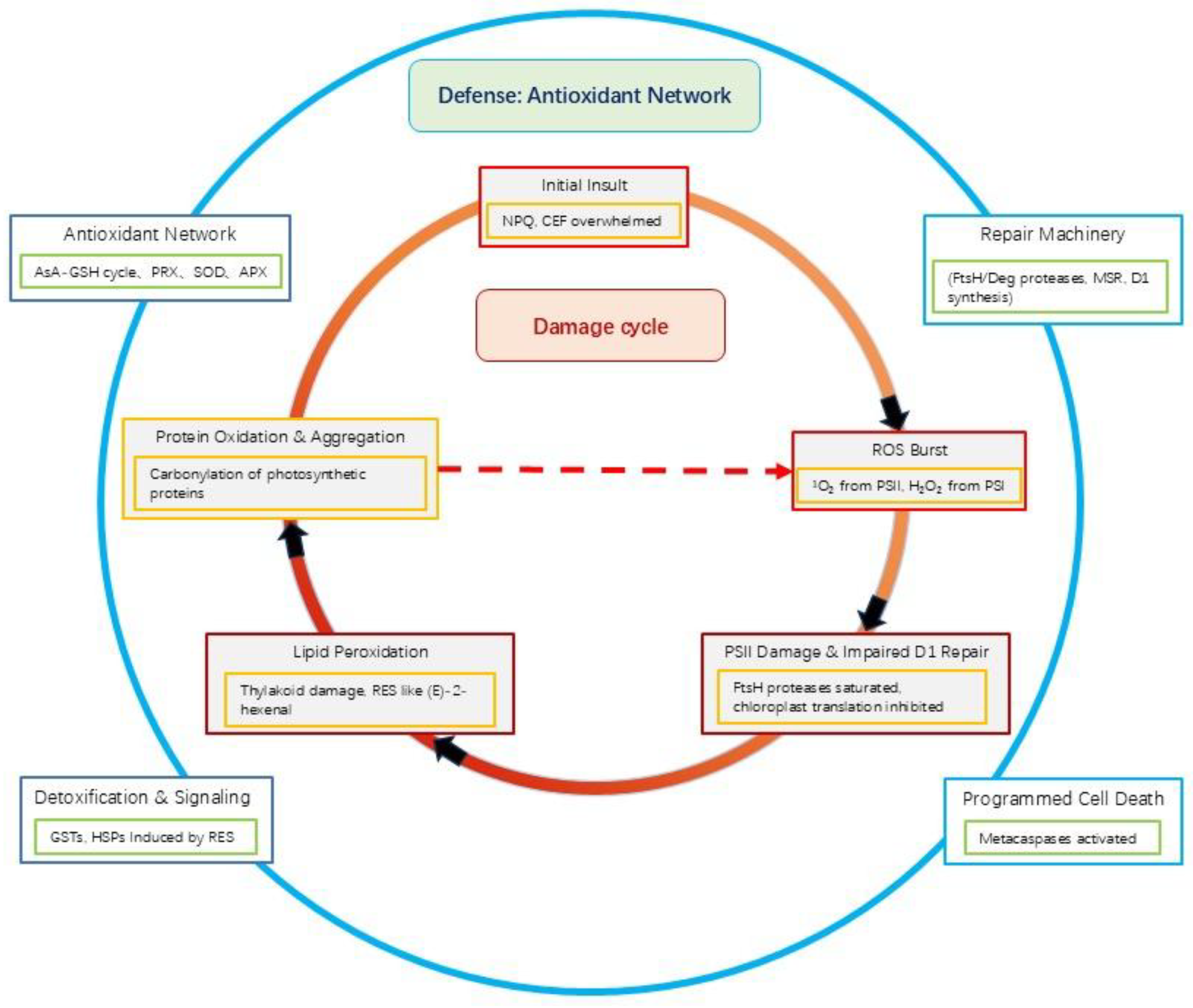
3. Mechanisms of Photo-oxidative Damage
When photoprotective and antioxidant capacities are overwhelmed, ROS cause widespread cellular damage. This damage is both a passive degenerative process and an active driver of stress signaling and metabolic reprogramming, determining cell fate through repair or irreversible collapse.
3.1. An Interconnected Damage Network
The initial site of photo-oxidative damage often depends on the stress type and the predominant ROS species. However, the ensuing effects are highly synergistic, creating a self-propagating cycle of deterioration. A central node in this network is the impairment of the PSII repair cycle.
When the D1 protein repair machinery (e.g., FtsH protease) becomes saturated or itself oxidized under sustained stress [
10,
28,
29], the accumulation of non-functional PSII centers leads to a direct increase in ¹O₂ production [
11]. This ¹O₂ burst has two major consequences that propagate the damage.
The first is direct exacerbation of lipid peroxidation. The thylakoid membranes, rich in polyunsaturated fatty acids (PUFAs), are prime targets for ¹O₂ and hydroxyl radicals. Increased lipid peroxidation compromises membrane integrity, further destabilizing the photosynthetic apparatus and potentially inactivating membrane-bound protein complexes [
30,
31].
The second is the inhibition of de novo protein synthesis. The oxidative by-products of lipid peroxidation, such as reactive electrophilic species (RES) like (E)-2-hexenal, as well as H₂O₂ itself, can diffuse to the stromal compartment and inhibit chloroplast translation [
32,
33]. This creates a critical bottleneck, preventing the synthesis of new D1 and other proteins essential for repair, thereby locking the cycle in a destructive feedback loop.
Simultaneously, damage to PSI, which is often caused by acceptor-side limitation and over-reduction, can be irreversible due to the destruction of its Fe-S clusters [
15,
34]. PSI photoinhibition cripples cyclic electron flow (CEF), a key photoprotective mechanism for regulating the proton motive force and preventing over-reduction [
12,
35]. The failure of CEF further exacerbates the over-reduced state of the electron transport chain, increasing ROS production at both photosystems and accelerating the damage to lipids and proteins [
36].
This convergence of damage on the proteostasis network represents a final common pathway. The accumulation of carbonylated and aggregated proteins, despite the efforts of peroxiredoxins and the ubiquitin-proteasome system, ultimately leads to the activation of metacaspases and the initiation of programmed cell death [
37]. This organized death removes cells that are net sinks of energy and potential sources of propagating oxidative signals, protecting the rest of the organism.
3.2. Impact on Photosystem Stability and Repair
Sustained photoinhibitory stress directly targets the stability and repair mechanisms of both photosystems, with distinct consequences. The repair cycle of PSII, initiated by the rapid ¹O₂-mediated damage to its D1 protein, is particularly vulnerable to failure under prolonged stress. Impairment occurs at multiple points, with the FtsH protease complex responsible for degrading damaged D1 becoming saturated or oxidized [
28], while concurrent damage to the chloroplast translational apparatus limits the de novo synthesis of replacement D1 proteins [
33,
38]. This leads to the accumulation of inactive PSII centers and a subsequent increase in ROS production. In contrast, damage to PSI is often more detrimental and irreversible. Over-reduction conditions can destroy the Fe-S clusters within PSI, for which no efficient repair mechanism exists, necessitating the energy-costly and slow de novo synthesis of the entire complex [
39]. The vulnerability of PSI is further heightened under fluctuating light when the protective PGR5/PGRL1-mediated cyclic electron flow is inhibited [
40].
3.3. Lipid Peroxidation and Oxylipin Signaling
Thylakoid membranes, rich in PUFAs, are prime ROS targets. Hydroxyl radicals (•OH) and ¹O₂ initiate lipid peroxidation, forming unstable lipid hydroperoxides that decompose into RES like malondialdehyde (MDA) and (E)-2-hexenal [
31,
41,
42,
43].
This degradation has dual effects. It compromises membrane structure and photosynthetic efficiency, and RES (oxylipins) act as signaling molecules. Oxylipins like (E)-2-hexenal covalently modify proteins (protein carbonylation) and activate defense-related gene expression. For instance, (E)-2-hexenal directly induces the expression of genes encoding GLUTATHIONE S-TRANSFERASE (GST) and HEAT SHOCK PROTEINS (HSPs) [
42,
44,
45]. GSTs are crucial for detoxifying electrophiles and ROS, while HSPs protect and refold damaged proteins. Furthermore, transcriptomic studies reveal that (E)-2-hexenal and related RES activate a specific suite of transcripts that overlaps with, but is distinct from, H₂O₂ -induced genes, including those involved in the synthesis of protective flavonoids and glucosinolates [
46]. This demonstrates that membrane lipid peroxidation is not merely a degenerative process but is transduced into a coordinated genetic defense program for acclimation.
3.4. Proteostasis and Protein Degradation
Maintaining proteome integrity (proteostasis) is critical under stress, as photo-oxidation causes irreversible protein carbonylation, leading to loss of function, aggregation, and cytotoxicity.
Plants defend against protein oxidation via the antioxidant system, with 2-Cys PRXs as key H₂O₂ and organic hydroperoxide scavengers in chloroplasts [
47]. Under severe stress, 2-Cys PRXs become overoxidized, switching from peroxidases to molecular chaperones that prevent protein aggregation [
48].
When prevention fails, damaged proteins are degraded via the ubiquitin-proteasome system (UPS) and proteases like FtsH. Methionine sulfoxide reductases (MSRs) specifically repair oxidized methionine residues in photosynthetic proteins [
49], while metacaspases trigger controlled proteolysis or programmed cell death in extreme cases, removing heavily damaged cells to benefit the organism [
37].
4. Anti-Oxidation and Photoprotective Mechanisms
Plants have evolved multi-layered defense systems to survive fluctuating light and combinatorial stresses. Recent research has advanced understanding of these mechanisms, enabling predictive and engineering applications (
Figure 2).
4.1. A Multi-Layered Photoprotective Arsenal
To counteract photo-oxidative stress, plants employ an integrated, multi-tiered defense system that operates across temporal and functional scales. This coordinated arsenal includes the rapid dissipation of excess light energy, a sophisticated metabolic network for maintaining redox homeostasis, and the flexible redirection of photosynthetic electron flow. Together, these strategies function synergistically to prevent the over-reduction of PETC and mitigate cellular damage.
The first and fastest line of defense is dynamic energy dissipation through NPQ. In vascular plants, NPQ is activated by thylakoid lumen acidification under high light, which protonates the PSII Subunit S (PSBS) and stimulates the violaxanthin-to-zeaxanthin conversion [
50]. This process induces conformational changes in antenna complexes, safely dissipating excitation energy as heat [
51]. The kinetics of NPQ relaxation are critical; slow relaxation under fluctuating light can unnecessarily limit photosynthesis. Landmark research demonstrated that accelerating NPQ relaxation by overexpressing key components (xanthophyll cycle enzymes and PSBS) enhanced tobacco biomass yield by approximately 15%, validating the agricultural potential of optimizing this dynamic process [
52]. The full induction of NPQ is further supported by the proton gradient sustained by cyclic electron flow, illustrating the crosstalk between regulatory pathways [
12].
Underpinning these rapid responses is a robust metabolic network centered on redox homeostasis. The ascorbate-glutathione (AsA-GSH) cycle forms the core of the chloroplast antioxidant system, functioning as a central redox hub. Within this cycle, hydrogen peroxide is scavenged by ascorbate peroxidase (APX), while ascorbate is regenerated via glutathione and NADPH. The localization of specific APX isoforms, such as thylakoid-bound tAPX, is crucial for local membrane protection [
53]. This system is intrinsically linked to cellular metabolism through NADPH supply and is deeply integrated with the thioredoxin (Trx) system. Thioredoxins (Trx-f, -m, -x, -y, -z) regulate enzyme activity via dithiol-disulfide exchange, with Trx-x specifically activating peroxiredoxins. The Trx and GSH systems often operate synergistically, and their integration is mediated by key players like NADPH-thioredoxin reductase C (NTRC), which connects photosynthetic electron flow, antioxidant metabolism, and carbon assimilation [
54].
When light absorption exceeds carbon assimilation capacity, plants activate alternative electron sinks to alleviate PETC over-reduction. CEF plays a fundamental role by recycling electrons from ferredoxin back to the plastoquinone pool, primarily through the PGR5/PGRL1 and NDH-dependent pathways [
12]. This process is essential for generating the trans-thylakoid proton gradient that drives ATP synthesis and activates NPQ, thereby providing crucial protection to Photosystem I [
12]. The water-water cycle (WWC), which involves the Mehler reaction, serves as another electron sink by reducing oxygen to superoxide, which is subsequently detoxified to hydrogen peroxide, fulfilling roles in both electron dissipation and redox signaling [
55,
56]. Additionally, photorespiration, once considered a wasteful pathway, is now recognized as an indispensable electron sink under CO₂-limiting conditions. By consuming ATP and NADPH during the recycling of 2-phosphoglycolate, it effectively relieves over-reduction pressure on the PETC and is vital for plant survival under combined stress [
57].
In summary, the plant’s photoprotective strategy is a highly integrated system where dynamic energy quenching, metabolic redox buffering, and flexible electron partitioning operate in concert. This multi-layered defense enables plants to balance efficient light utilization with the avoidance of photo-oxidative damage. The effectiveness of this integrated system, however, is shaped by evolutionary trade-offs between growth and defense, a concept explored in the following section.
4.2. Enzymatic Detoxification
Prxs, particularly 2-Cys Prxs, are central to chloroplast antioxidant defense. These abundant thiol-based peroxidases efficiently reduce H₂O₂, organic hydroperoxides, and peroxynitrite. Under high peroxide flux, 2-Cys Prxs undergo hyperoxidation, switching from peroxidases to chaperones that prevent protein aggregation [
58], serving as both antioxidants and proteoprotective agents.
Dedicated repair systems include MSR, which restores oxidized methionine residues in photosynthetic proteins [
49]. Irreversibly damaged proteins are removed by chloroplast proteases (FtsH, Deg) and the cytosolic UPS, completing the surveillance-repair-degradation cycle essential for stress tolerance.
4.3. Ecological and Evolutionary Context of Photo-Oxidative Resilience
The molecular mechanisms governing photo-oxidative stress are not uniform across the plant kingdom but are instead evolutionary products fine-tuned by ecological niche and life history strategy. This natural variation reveals fundamental trade-offs and provides a crucial genetic reservoir for breeding resilient crops.
A primary evolutionary trade-off exists between maximizing growth under limiting light and minimizing damage under excess light. Species adapted to high-light environments, such as sun-acclimated plants, typically exhibit constitutively elevated capacities for NPQ, antioxidant pools, and CEF [
34,
36,
50,
56]. In contrast, shade-adapted species prioritize efficient light capture but possess minimal photoprotective reserves, leaving them highly vulnerable to sudden high-light exposure [
59]. This divergence in strategy underscores the high metabolic cost of photoprotection, which creates a pervasive growth-defense trade-off. Investment in antioxidants, photoprotective pigments like zeaxanthin, and protein repair cycles (e.g., D1 turnover) diverts resources from growth and reproduction. Consequently, genetic lines with enhanced stress tolerance, such as Arabidopsis mutants with elevated ¹O₂ signaling or antioxidant capacity, often exhibit reduced growth under non-stressful conditions [
18].
Convergent evolutionary adaptations to minimize photo-oxidative stress are exemplified by C4 and Crassulacean Acid Metabolism (CAM) photosynthesis. Both pathways function as preemptive solutions by concentrating CO₂ around Rubisco, thereby dramatically suppressing photorespiration (a major source of H₂O₂) and minimizing conditions that lead to PETC over-reduction [
60]. C4 plants, with their specialized Kranz anatomy, can maintain high photosynthetic rates with partially closed stomata under drought and heat, conferring superior resilience to combinatorial stresses that induce photo-oxidation. CAM plants achieve similar benefits by temporally separating CO₂ fixation (at night) from light capture (during the day), thereby avoiding transpirational water loss and minimizing photoinhibitory risk.
Understanding this ecological and evolutionary context is directly applicable to crop improvement. The vast natural variation between stress-tolerant wild relatives and high-yielding cultivars represents a genetic resource for identifying valuable alleles. Targeted screening of germplasm for traits such as rapid NPQ relaxation, robust CEF activity, and stable PSI function under fluctuating light can uncover these alleles [
52,
61]. The challenge for modern breeding lies in leveraging these evolved natural solutions informed by the ecological context from which they arose to engineer climate-resilient crops that mitigate the yield penalty traditionally associated with enhanced photoprotection.
4.4. Harnessing Photoprotection for Crop Improvement
The fundamental understanding of photoprotective mechanisms has opened a promising frontier for engineering climate-resilient crops. Moving beyond proof-of-concept in model plants, the focus is now on translating these mechanisms into agronomically viable traits that enhance yield under real-world field conditions. Several strategies beyond the landmark engineering of accelerated NPQ relaxation have shown significant promise. These include the targeted overexpression of chloroplast antioxidant enzymes, such as superoxide dismutase and ascorbate peroxidase, to mitigate photo-oxidative damage across multiple stresses [
62]. Rewiring photorespiration through synthetic glycolate metabolic pathways, which bypass the native H₂O₂-producing route, has successfully reduced energy losses and increased biomass in crops like tobacco and rice [
63,
64]. Furthermore, enhancing alternative electron sinks by manipulating components like PGR5 to boost CEF has proven effective in protecting Photosystem I and improving growth under fluctuating light [
12].
However, translating these successes from controlled environments to major crops faces significant translational hurdles. A primary challenge is the pleiotropic effects and inherent energy costs of introduced traits; for example, constitutive overexpression of protective proteins can divert resources from growth, often resulting in a yield penalty under optimal conditions, a clear manifestation of the growth-defense trade-off [
52]. Moreover, trait performance must be validated under dynamic, combinatorial field stresses, as advantages observed in growth chambers may not persist in real agricultural environments [
65]. Finally, given that resilience is a polygenic trait, engineering robust tolerance may require the simultaneous optimization of multiple pathways through sophisticated gene stacking and precise spatiotemporal control of transgene expression.
For integration into modern breeding programs, “photo-oxidative stress resilience” must be defined by quantifiable phenotypic markers. Key measurable parameters include the kinetics of NPQ induction and relaxation, the in vivo stability of PSI and PSII, established markers of oxidative damage like lipid peroxidation, and the redox states of antioxidant pools such as glutathione and ascorbate. The development of high-throughput phenotyping platforms, coupled with genome-wide association studies, is now enabling the identification of natural genetic variation underlying these physiological traits within crop germplasm [
61,
66]. By systematically linking these molecular and physiological metrics to yield performance under stress, photo-oxidative resilience can be effectively targeted and selected for, paving the way for the development of next-generation, climate-adapted cultivars.
5. Future Research Directions
Advances in understanding the molecular mechanisms of photo-oxidative stress have opened new frontiers for predictive modeling and targeted engineering of plant resilience under climate change. Future research will likely converge on several interdisciplinary approaches that bridge fundamental discovery with agricultural application.
The precision of genome editing tools, particularly CRISPR/Cas systems, offers unprecedented opportunities to fine-tune photoprotective and antioxidant pathways beyond simple gene knockouts. A key focus will be engineering cell-type-specific or stress-inducible promoters to drive the expression of crucial genes, e.g., PsbS or PGR5, thereby optimizing the trade-off between photoprotection and growth. Synthetic biology approaches may further introduce novel antioxidant circuits or rewire existing signaling networks to enhance stress tolerance [
63]. A significant translational challenge remains in effectively porting these genetic modifications from model plants like Arabidopsis to major crops and validating their efficacy under dynamic field conditions.
To fully decipher the complexity of plant stress responses, the integration of multi-omics data encompassing genomics, transcriptomics, proteomics and metabolomics will be essential. Temporal multi-omics profiling can capture the cascade of molecular events from initial stress perception to long-term acclimation. Coupled with advanced computational biology and machine learning, such integrated datasets will help identify key regulatory hubs and predict phenotypic outcomes, moving toward a systems-level understanding of resilience [
67]. For example, genome-wide association studies (GWAS) have proven powerful in linking genetic variation to stress traits, as demonstrated by the identification of the TaSnRK1α–TaCAT2 module that enhances wheat resistance to Fusarium crown rot by bolstering ROS-scavenging capacity [
66].
Importantly, plant stress responses are highly heterogeneous across tissues and cell types, necessitating investigation at single-cell and subcellular resolution. Genetically encoded biosensors, such as roGFP2-Orp1 for H₂O₂ and GRX1-roGFP2 for redox state, enable real-time, in vivo tracking of signaling dynamics within specific cellular compartments [
3]. These tools have already revealed, for instance, that systemic ROS waves travel preferentially through the apoplast of vascular bundle sheath cells to systemically alert distant tissues [
13]. Combining such biosensors with single-cell RNA sequencing will uncover distinct transcriptional programs in different cell types under stress [
68]. Applying these approaches to leaves experiencing photo-oxidation could clarify whether bundle sheath and mesophyll cells in C₃ plants, or the specialized cell types of C₄ plants, employ divergent photoprotective strategies, information critical for precise, tissue-targeted engineering without disrupting overall plant physiology.
Finally, harnessing natural variation present in crop wild relatives and landraces provides a complementary and evolutionarily informed route to resilience. High-throughput phenotyping will enable systematic screening of germplasm for desirable traits such as rapid NPQ relaxation, enhanced cyclic electron flow, or robust antioxidant capacity. Subsequent GWAS can link these phenotypes to underlying genetic loci, providing molecular markers to accelerate the development of climate-resilient cultivars [
61].
In conclusion, the synergy of precision genome editing, integrative multi-omics, spatially resolved dynamic analysis, and genomics-assisted breeding will be instrumental in unraveling the remaining complexities of photo-oxidative stress. Together, these approaches will equip future crops with the robust, multilayered defenses necessary to thrive in an increasingly variable and challenging climate.
Author Contributions
writing—original draft preparation, review and editing, X.L., S.Y. and J.Z.; supervision, S.W.; funding acquisition, X.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Key Research and Development Project of Shandong Province (ZFJH202310), the Shandong Province Natural Science Foundation Project (ZR2023MC109) and International Science and Technology Cooperation Special Project of Shandong Academy of Agricultural Sciences.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| •OH |
Hydroxyl radicals |
| ¹O₂ |
Singlet oxygen |
| ³Chl |
Triplet state of chlorophyll |
| β-CC |
β-cyclocitral |
| ABA |
Abscisic acid |
| APX |
Ascorbate peroxidase |
| AsA-GSH |
Ascorbate-glutathione |
| CAM |
Crassulacean Acid Metabolism |
| CEF |
Cyclic electron flow |
| EX1 |
Nuclear-encoded chloroplast proteins EXECUTER1 |
| EX2 |
Nuclear-encoded chloroplast proteins EXECUTER2 |
| GWAS |
Genome-wide association studies |
| GST |
Glutathione S-transferase |
| H₂O₂ |
Hydrogen peroxide |
| HSPs |
Heat shock proteins |
| JA |
Jasmonic acid |
| MDA |
Malondialdehyde |
| MEcPP |
Methylerythritol cyclodiphosphate |
| MSRs |
Methionine sulfoxide reductases |
| NPQ |
Non-photochemical quenching |
| NTRC |
NADPH-thioredoxin reductase C |
| O₂•⁻ |
Superoxide anions |
| PAP |
3’-phosphoadenosine 5’-phosphate |
| PETC |
Photosynthetic electron transport chain |
| PRXs |
Peroxiredoxins |
| PSI |
Photosystem I |
| PSII |
Photosystem II |
| PSBS |
PSII Subunit S |
| PUFAs |
Polyunsaturated fatty acids |
| RBOH |
Respiratory burst oxidase homolog |
| RES |
Reactive electrophilic species |
| ROS |
Reactive oxygen species |
| SA |
Salicylic acid |
| SAA |
Systemic acquired acclimation |
| SAR |
Systemic acquired resistance |
| Trx |
Thioredoxin |
| UPS |
Ubiquitin-proteasome system |
| WWC |
Water-water cycle |
References
- Devireddy, A.R.; Zandalinas, S.I.; Fichman, Y.; Mittler, R. Integration of reactive oxygen species and hormone signaling during abiotic stress. Plant J 2021, 105(2), 459–476. [Google Scholar] [CrossRef]
- Dietz, K.J.; Turkan, I.; Krieger-Liszkay, A. Redox- and reactive oxygen species-dependent signaling into and out of the photosynthesizing chloroplast. Plant Phys 2016, 171(3), 1541–1550. [Google Scholar] [CrossRef] [PubMed]
- Exposito-Rodriguez, M.; Laissue, P.P.; Yvon-Durocher, G.; Smirnoff, N.; Mullineaux, P.M. Photosynthesis-dependent H2O2 transfer from chloroplasts to nuclei provides a high-light signalling mechanism. Nat Commun 2017, 8(1), 49. [Google Scholar] [CrossRef]
- Fichman, Y.; Mittler, R. Rapid systemic signaling during abiotic and biotic stresses: is the ROS wave master of all trades? Plant J 2020, 102(5), 887–896. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wakao, S.; Fischer, B.B.; Niyogi, K.K. Sensing and responding to excess light. Annu Rev Plant Bio 2009, 60, 239–260. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Meng, D.Y.; Hou, L.L.; Li, Y.; Guo, F.; Meng, J.J.; Wan, S.B.; Li, X.G. Peanut violaxanthin de-epoxidase alleviates the sensitivity of PSII photoinhibition to heat and high irradiance stress in transgenic tobacco. Plant Cell Rep 2015, 34, 1417–1428. [Google Scholar] [CrossRef]
- Yang, S.; Wang, F.; Guo, F.; Meng, J.J.; Li, X.G.; Wan, S.B. Calcium contributes to photoprotection and repair of photosystem II in peanut leaves during heat and high irradiance. J Integ Plant Bio 2015, 57(5), 486–495. [Google Scholar] [CrossRef]
- Mittler, R.; Blumwald, E. The roles of ROS and ABA in systemic acquired acclimation. Plant Cell 2015, 27(1), 64–70. [Google Scholar] [CrossRef]
- Waszczak, C.; Carmody, M.; Kangasjärvi, J. Reactive oxygen species in plant signaling. Annu Rev Plant Bio 2018, 69, 209–236. [Google Scholar] [CrossRef]
- D’Alessandro, S.; Havaux, M. Sensing β-carotene oxidation in photosystem II to master plant stress tolerance. New Phytol 2019, 223(4), 1776–1783. [Google Scholar] [CrossRef]
- Dall’Osto, L.; Cazzaniga, S.; Bressan, M.; Paleček, D.; Židek, K.; Niyogi, K.K.; Fleming, G.R.; Zigmantas, D.; Bassi, R. Two mechanisms for dissipation of excess light in monomeric and trimeric light-harvesting complexes. Nat Plants 2017, 3, 17033. [Google Scholar] [CrossRef] [PubMed]
- Yamori, W.; Shikanai, T. Physiological functions of cyclic electron transport around photosystem I in sustaining photosynthesis and plant growth. Annu Rev Plant Biol 2016, 67, 81–106. [Google Scholar] [CrossRef] [PubMed]
- Zandalinas, S.I.; Fichman, Y.; Mittler, R. Vascular bundles mediate systemic reactive oxygen signaling during light stress. Plant Cell 2020, 32(11), 3425–3435. [Google Scholar] [CrossRef] [PubMed]
- Zandalinas, S. I.; Fichman, Y.; Devireddy, A.R.; Sengupta, S.; Azad, R.K.; Mittler, R. Systemic signaling during abiotic stress combination in plants. Proc Nat Acad Sci 2020, 117(24), 13810–13820, e2125077119. [Google Scholar] [CrossRef]
- Tikkanen, M.; Grebe, S. Switching off photoprotection of photosystem I - a novel tool for gradual PSI photoinhibition. Physiol Plant 2018, 162(2), 156–161. [Google Scholar] [CrossRef]
- Noctor, G.; Reichheld, J.P.; Foyer, C.H. ROS-related redox regulation and signaling in plants. Semin Cell Dev Biol 2018, 80, 3–12. [Google Scholar] [CrossRef]
- Mittler, R.; Zandalinas, S.I.; Fichman, Y.; Van Breusegem, F. Reactive oxygen species signalling in plant stress responses. Nat Rev Mol Cell Biol 2022, 23(10), 663–679. [Google Scholar] [CrossRef]
- Dogra, V.; Kim, C. Singlet oxygen metabolism: from genesis to signaling. Front Plant Sci 2020, 10, 1640. [Google Scholar] [CrossRef]
- Ramel, F.; Birtic, S.; Ginies, C.; Soubigou-Taconnat, L.; Triantaphylidès, C.; Havaux, M. Carotenoid oxidation products are stress signals that mediate gene responses to singlet oxygen in plants. Proc Natl Acad Sci USA 2012, 109(14), 5535–40. [Google Scholar] [CrossRef]
- Li, X.G.; Guo, F.; Meng, J.J.; Yang, S.; Guo, S.J.; Wan, S.B.; Picimbon, J.F. Energy dissipation in photosystem II complexes of peanut (Arachis hypogaca L.) leaves subjected to light flashes. Plant Grow Reg 2014, 74(2), 131–138. [Google Scholar] [CrossRef]
- Deshpande, S.; Purkar, V.; Mitra, S. β-Cyclocitral, a master regulator of multiple stress-responsive genes in Solanum lycopersicum L. plants. Plants (Basel) 2021, 10(11), 2465. [Google Scholar] [CrossRef]
- Nietzel, T.; Mostertz, J.; Hochgräfe, F.; Schwarzländer, M. Redox regulation of mitochondrial proteins and proteomes by cysteine thiol switches. Mitochondrion 2017, 33, 72–83. [Google Scholar] [CrossRef]
- Shapiguzov, A.; Vainonen, J.P.; Hunter, K.; Tossavainen, H.; Tiwari, A.; Järvi, S.; Hellman, M.; Aarabi, F.; Alseekh, S.; Wybouw, B.; Van Der Kelen, K.; Nikkanen, L.; Krasensky-Wrzaczek, J.; Sipari, N.; Keinänen, M.; Tyystjärvi, E.; Rintamäki, E.; De Rybel, B.; Salojärvi, J.; Van Breusegem, F.; Fernie, A.R.; Brosché, M.; Permi, P.; Aro, E.M.; Wrzaczek, M.; Kangasjärvi, J. Arabidopsis RCD1 coordinates chloroplast and mitochondrial functions through interaction with ANAC transcription factors. Elife 2019, 8, e43284. [Google Scholar] [CrossRef]
- Chan, K.X.; Phua, S.Y.; Crisp, P.; McQuinn, R.; Pogson, B.J. Learning the languages of the chloroplast: retrograde signaling and beyond. Annu Rev Plant Bio 2016, 67, 25–53. [Google Scholar] [CrossRef]
- Cao, L.; Karapetyan, S.; Yoo, H.; Chen, T.; Mwimba, M.; Zhang, X.; Dong, X. H2O2 sulfenylates CHE, linking local infection to the establishment of systemic acquired resistance. Sci 2024, 385(6714), 1211–1217. [Google Scholar] [CrossRef] [PubMed]
- Janda, T.; Szalai, G.; Pál, M. Salicylic acid signalling in plants. Int J Mol Sci 2020, 21(7), 2655. [Google Scholar] [CrossRef] [PubMed]
- Berens, M.L.; Berry, H.M.; Mine, A.; Argueso, C.T.; Tsuda, K. Evolution of hormone signaling networks in plant defense. Annu Rev Phytopathol 2017, 55, 401–425. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Sakamoto, W. FtsH protease in the thylakoid membrane: physiological functions and the regulation of protease activity. Front Plant Sci 2018, 9, 855. [Google Scholar] [CrossRef]
- Qin, L.Q.; Li, L.; Bi, C.; Zhang, Y.L.; Wan, S.B.; Meng, J.J.; Meng, Q.W.; Li, X.G. Damaging mechanisms of chilling- and salt stress to Arachis hypogaea L. leaves. Photosynthetica 2011, 49(1), 37–42. [Google Scholar] [CrossRef]
- Triantaphylidès, C.; Havaux, M. Singlet oxygen in plants: production, detoxification and signaling. Trends Plant Sci 2009, 14(4), 219–28. [Google Scholar] [CrossRef]
- Yamori, W. Photosynthesis and respiration. In Plant Factory; (Second Edition), 2020; pp. 197–206. [Google Scholar]
- Nishiyama, Y.; Allakhverdiev, S.I.; Murata, N. A new paradigm for the action of reactive oxygen species in the photoinhibition of photosystem II. Biochim Biophys Acta 2006, 1757(7), 742–749. [Google Scholar] [CrossRef]
- Sun, Y.; Li, J.; Zhang, L.; Lin, R. Regulation of chloroplast protein degradation. J Genet Genomics 2023, 50(6), 375–384. [Google Scholar] [CrossRef]
- Li, X.G.; Xu, P.L.; Zhao, J.P.; Meng, J.J.; He, Q.W. Ferredoxin-quinone reductase benefits cyclic electron flow around ps1 in tobacco leaves upon exposure to chilling stress under low irradiance. Photosynthetica 2006, 44(3), 349–354. [Google Scholar] [CrossRef]
- Li, X.G.; Duan, W.; Meng, Q.W.; Zou, Q.; Zhao, S.J. The function of chloroplastic NAD (P) H dehydrogenase in tobacco during chilling stress under low irradiance. Plant Cell Phys 2004, 45(1), 103–108. [Google Scholar] [CrossRef] [PubMed]
- Li, X.G.; Zhao, J.P.; Xu, P.L.; Meng, J.J.; He, Q.W. Effects of cyclic electron flow inhibitor (antimycin A) on photosystem photoinhibition of sweet pepper leaves upon exposure to chilling stress under low irradiance. Agr Sci China 2006, 5(7), 101–105. [Google Scholar] [CrossRef]
- van Breusegem, F.; Dat, J.F. Reactive oxygen species in plant cell death. Plant Physiol 2006, 141(2), 384–90. [Google Scholar] [CrossRef]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J 2017, 90(5), 856–867. [Google Scholar] [CrossRef]
- Tikkanen, M.; Rantala, S.; Aro, E.M. Electron flow from PSII to PSI under high light is controlled by PGR5 but not by PSBS. Front Plant Sci 2015, 6, 521. [Google Scholar] [CrossRef]
- Järvi, S.; Suorsa, M.; Aro, E.M. Photosystem II repair in plant chloroplasts--Regulation, assisting proteins and shared components with photosystem II biogenesis. Biochim Biophys Acta 2015, 1847(9), 900–909. [Google Scholar] [CrossRef]
- Mueller, M.J.; Berger, S. Reactive electrophilic oxylipins: pattern recognition and signalling. Phytochemistry 2009, 70(13-14), 1511–1521. [Google Scholar] [CrossRef]
- Farmer, EE; Mueller, MJ. ROS-mediated lipid peroxidation and RES-activated signaling. Annu Rev Plant Biol 2013, 64, 429–50. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Ullah, F.; Zhou, D.X.; Yi, M.; Zhao, Y. Mechanisms of ros regulation of plant development and stress responses. Front Plant Sci 2019, 10, 800. [Google Scholar] [CrossRef] [PubMed]
- Alméras, E.; Stolz, S.; Vollenweider, S.; Reymond, P.; Mène-Saffrané, L.; Farmer, E.E. Reactive electrophile species activate defense gene expression in Arabidopsis. Plant J 2003, 34(2), 205–216. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.J.; Zhou, H.Y.; Zhang, X.S.; Li, X.G.; Meng, Q.W. Overexpression of CaHSP26 in transgenic tobacco alleviates photoinhibition of PSII and PSI during chilling stress under low irradiance. J Plant Phys 2007, 164(2), 126–136. [Google Scholar] [CrossRef]
- Gao, F.; Chen, J.; Ma, T.; Li, H.; Wang, N.; Li, Z.; Zhang, Z.; Zhou, Y. The glutathione peroxidase gene family in Thellungiella salsuginea: genome-wide identification, classification, and gene and protein expression analysis under stress conditions. Int J Mol Sci 2014, 15(2), 3319–3335. [Google Scholar] [CrossRef]
- Dietz, K.J. Thiol-based peroxidases and ascorbate peroxidases: why plants rely on multiple peroxidase systems in the photosynthesizing chloroplast? Mol Cells 2016, 39(1), 20–25. [Google Scholar] [CrossRef]
- Vaseghi, M.J.; Chibani, K.; Telman, W.; Liebthal, M.F.; Gerken, M.; Schnitzer, H.; Mueller, S.M.; Dietz, K.J. The chloroplast 2-cysteine peroxiredoxin functions as thioredoxin oxidase in redox regulation of chloroplast metabolism. Elife 2018, 7, e38194. [Google Scholar] [CrossRef]
- Roy, S.; Nandi, A.K. Arabidopsis thaliana methionine sulfoxide reductase B8 influences stress-induced cell death and effector-triggered immunity. Plant Mol Biol 2017, 93(1-2), 109–120. [Google Scholar] [CrossRef]
- Li, X.G.; Li, J.Y.; Zhao, J.P.; Xu, P.L.; He, Q.W. Xanthophyll cycle and inactivation of photosystem 2 reaction centers alleviating reducing pressure to photosystem 1 in morning glory leaves upon exposure to a short-term high irradiance. J Integ Plant Bio 2007, 49(7), 1047–1053. [Google Scholar] [CrossRef]
- Niyogi, K.K. Photoprotection and high light responses. Chlamydomonas Sourcebook 2009, 847–870. [Google Scholar] [CrossRef]
- Kromdijk, J.; Głowacka, K.; Leonelli, L.; Gabilly, S.T.; Iwai, M.; Niyogi, K.K.; Long, S.P. Improving photosynthesis and crop productivity by accelerating recovery from photoprotection. Sci 2016, 354(6314), 857–861. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Hanke, G. ROS production and signalling in chloroplasts: cornerstones and evolving concepts. Plant J 2022, 111(3), 642–661. [Google Scholar] [CrossRef] [PubMed]
- Zaffagnini, M.; Fermani, S.; Marchand, C.H.; Costa, A.; Sparla, F.; Rouhier, N.; Geigenberger, P.; Lemaire, S.D.; Trost, P. Redox homeostasis in photosynthetic organisms: novel and established thiol-based molecular mechanisms. Antioxid Redox Signal 2019, 31(3), 155–210. [Google Scholar] [CrossRef] [PubMed]
- Asada, K. The water-water cycle as alternative photon and electron sinks. Philos Trans R Soc Lond B Biol Sci 2000, 355(1402), 1419–1431. [Google Scholar] [CrossRef]
- Li, X.G.; Bi, Y.P.; Zhao, S.J.; Meng, Q.W.; Zou, Q.; He, Q.W. Cooperation of xanthophyll cycle with water-water cycle in the protection of PSII and PSI against inactivation during chilling stress under low irradiance. Photosynthetica 2005, 43(2), 261–266. [Google Scholar] [CrossRef]
- Busch, F.A. Photorespiration in the context of Rubisco biochemistry, CO2 diffusion and metabolism. Plant J 2020, 101(4), 919–939. [Google Scholar] [CrossRef]
- Liebthal, M.; Maynard, D.; Dietz, K.J. Peroxiredoxins and redox signaling in plants. Antioxid Redox Signal 2018, 28(7), 609–624. [Google Scholar] [CrossRef]
- Galmés, J.; Ochogavía, J.M.; Gago, J.; Roldán, E.J.; Cifre, J.; Conesa, M.À. Leaf responses to drought stress in Mediterranean accessions of Solanum lycopersicum: anatomical adaptations in relation to gas exchange parameters. Plant Cell Environ 2013, 36(5), 920–935. [Google Scholar] [CrossRef]
- Schuler, M.L.; Mantegazza, O.; Weber, A.P. Engineering C4 photosynthesis into C3 chassis in the synthetic biology age. Plant J 2016, 87(1), 51–65. [Google Scholar] [CrossRef]
- De Souza, A.P.; Wang, Y.; Orr, D.J.; Carmo-Silva, E.; Long, S.P. Photosynthesis across African cassava germplasm is limited by Rubisco and mesophyll conductance at steady state, but by stomatal conductance in fluctuating light. New Phytol 2020, 225(6), 2498–2512. [Google Scholar] [CrossRef]
- López-Calcagno, P.E.; Brown, K.L.; Simkin, A.J.; Fisk, S.J.; Vialet-Chabrand, S.; Lawson, T.; Raines, C.A. Stimulating photosynthetic processes increases productivity and water-use efficiency in the field. Nat Plants 2020, 6(8), 1054–1063. [Google Scholar] [CrossRef]
- South, P.F.; Cavanagh, A.P.; Liu, H.W.; Ort, D.R. Synthetic glycolate metabolism pathways stimulate crop growth and productivity in the field. Sci 2019, 363(6422), eaat9077. [Google Scholar] [CrossRef] [PubMed]
- Shen, B.R.; Wang, L.M.; Lin, X.L.; Yao, Z.; Xu, H.W.; Zhu, C.H.; Teng, H.Y.; Cui, L.L.; Liu, E.E.; Zhang, J.J.; He, Z.H.; Peng, X.X. Engineering a new chloroplastic photorespiratory bypass to increase photosynthetic efficiency and productivity in rice. Mol Plant 2019, 12(2), 199–214. [Google Scholar] [CrossRef] [PubMed]
- Long, S.P.; Taylor, S.H.; Burgess, S.J.; Carmo-Silva, E.; Lawson, T.; De Souza, A.P.; Leonelli, L.; Wang, Y. Into the shadows and back into sunlight: Photosynthesis in fluctuating light. Annu Rev Plant Biol 2022, 73, 617–648. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, L.; Wei, J.; Liu, L.; Liu, D.; Yan, X.; Yuan, M.; Zhang, L.; Zhang, N.; Ren, Y.; Chen, F. A TaSnRK1α-TaCAT2 model mediates resistance to Fusarium crown rot by scavenging ROS in common wheat. Nat Commun 2025, 16(1), 2549. [Google Scholar] [CrossRef]
- Weckwerth, W.; Ghatak, A.; Bellaire, A.; Chaturvedi, P.; Varshney, R.K. PANOMICS meets germplasm. Plant Biotechnol J 2020, 18(7), 1507–1525. [Google Scholar] [CrossRef]
- Shaw, R.; Tian, X.; Xu, J. Single-cell transcriptome analysis in plants: advances and challenges. Mol Plant 2021, 14(1), 115–126. [Google Scholar] [CrossRef]
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).