
Submitted:
14 January 2026
Posted:
15 January 2026
You are already at the latest version
Abstract
Shaded coffee plantations are increasingly recognized as important agroecosystems for biodi-versity conservation in the tropical Andes. 1) Background: Dung beetles (Scarabaeinae) play a fundamental role in ecosystem functioning; however, their diversity and ecological roles within shaded coffee systems in southern Ecuador remain poorly understood. 2) Methods: This study assessed dung beetle diversity in 14 shaded coffee plantations located in the provinces of Loja, Zamora Chinchipe, and El Oro, using pitfall traps baited with pig dung. Community structure was analyzed using Hill numbers, non-metric multidimensional scaling, and generalized linear models. 3) Results: A total of 209 individuals belonging to 14 genera and 42 species were rec-orded. Loja exhibited the highest species richness and abundance, representing 73% of all indi-viduals, whereas El Oro showed the lowest diversity. Onthophagus curvicornis (Latreille, 1817) was the dominant species, accounting for 20% of the total abundance. Habitat type and altitude significantly influenced dung beetle abundance, with a predominance of coprophagous and par-acoprid species. 4) Conclusions: Shaded coffee plantations maintain dung beetle communities comparable to those of natural ecosystems, emphasizing their contribution to ecosystem services such as nutrient cycling, soil improvement, and biological pest control, and supporting the pro-motion of sustainable coffee management practices in Andean agroecosystems.
Keywords:
Scarabaeinae
; coffee plantations
; agroecological management
; bioindicators
1. Introduction
Agriculture has played a fundamental role in satisfying nutritional and economic needs, generating significant changes in the compositions of flora and fauna in different ecosystems [1]. In the last 15 years, a notable increase has been observed in coffee cultivation in Ecuador, being among the nine crops with the largest harvested area [2]. The wide presence of coffee in all the provinces of Ecuador highlights the geographical diversity and adaptability of the crop in this country [3]. From the highlands of the Sierra to the coastal plains and the lush tropical forests of the Amazon, coffee has taken root in a variety of environments, demonstrating its ability to thrive in different climatic and soil conditions [4]. This geographic distribution reflects the genetic versatility of coffee varieties and the ability of farmers to take advantage of the unique characteristics of each region. In addition to its economic importance as a source of income, coffee plays a crucial role in the cultural identity of Ecuador [5].
Especially in the south of Ecuador, the country's hot spot, it has optimal climatic conditions for the cultivation of shade coffee (Coffea arabica L.). The combination of altitude, temperature and humidity creates a favorable environment for the growth of different coffee varieties [6,7]. Such as Bourbon, Caturra, Sidra, Geisha, Catucai and Typica are all appreciated for their unique and distinctive flavor [5]. Although coffee is grown throughout Ecuador, in recent years there has been a notable increase in coffee production in the south of the country. This is due to the fact that the southern region of Ecuador has fertile soils rich in minerals, which favors the development of healthy and productive coffee plants [8,9]. And mainly, for the growing international recognition of the quality of coffee produced in the region [10].
However, these plantations, along with associated shrub and tree species contribute shade to the growth of coffee crops, are not only crucial for economic development, but also have implications for local biodiversity [11]. Which has led to a further change of agricultural landscapes [12]. A widely applied strategy to evaluate these changes in biodiversity is through the use of bioindicators that allow quantifying the degree of modification of an ecosystem [13]. Therefore, dung beetles are one of the groups frequently used due to their well-known taxonomy, biology and ecology, as environmental bioindicators. Likewise, they have demonstrated sensitivity to changes in ecosystems, which is why they are very useful as indicators of the state of conservation of an ecosystem [14].
In this context, dung beetles play an important role in these agricultural ecosystems, since they accelerate the decomposition of organic substrates, especially animal excrement, thus facilitating the renewal of nutrients and increasing soil fertility [15]. Through the substantial removal of organic waste, they effectively curtail the dissemination of pathogens, mitigate fly populations, and enhance environmental hygiene [16]. Moreover, their subterranean activities foster soil porosity, hydraulic conductivity, and architectural integrity, thereby fostering favorable conditions for botanical proliferation and genetic diversity [11]. Certain dung beetle species serve as effective biocontrol agents by preying upon pestiferous insect larvae within dung repositories, a phenomenon particularly salient in agroecosystems [16]. The activities of dung beetles exert a discernible influence on carbon flux dynamics, potentially contributing to the amelioration of greenhouse gas emissions [17]. The rich diversity of dung beetles confers multifaceted benefits to ecosystems, including but not limited to provisioning of trophic resources, sculpting of microhabitats, and maintenance of ecological equilibrium [18].
The aim of present study was to know the species composition of Scarabaeinae assemblages from coffee crops by using a standardized sampling protocol. The development of an exhaustive list of dung beetles present in coffee plantations is proposed, in order to systematically document their richness and abundance in a specific geographical context. These results are considered of great relevance as a support resource for researchers, conservationists, farmers and policy makers interested in understanding and preserving biodiversity, as well as promoting sustainable agricultural practices.
2. Materials and Methods
Study area
The study was carried out in three provinces in the south of Ecuador Loja, Zamora Chinchipe and El Oro (Figure 1).
They are characterized by cultivated shadow coffee (Coffea arabica L.). Mainly, the varieties cultivates are Bourbon, Caturra, Sidra, Geisha, Catucai and Typica. These farms are distributed on terraces on the slopes of the hills, taking advantage of the optimal altitude and soil conditions for the production of high-quality grain [6,19,20,21]. We selected 14 coffee plantations within eight canton and three provinces (Table 1).
Methods
Dung beetle sampling. In each coffee plantation, three linear transects of 300 m were installed, with a separation of 100 m each. In each transect, six traps were installed, separated every 150m the pitfall traps that were used consisted of 1-liter plastic containers placed at ground level, with 30 g of pig manure as bait, these were active for 48 h [22,23]. Subsequently, the samples were placed in 80% ethyl alcohol and transported to the Colección de Invertebrados Sur del Ecuador (CISEC-MUTPL) for separation and taxonomic identification [24].
Data analysis
We analyze two principal components of taxonomic diversity, richness and abundance of the dung beetle community [25]. And take into account the color, diet (coprophagous, necrophagous or generalist) and behavior (telecoprids, endocoprids and paracoprids) components of functional diversity [26]. Besides, we evaluated the sampled effort through Chao 1 index [27].
The dung beetle community was analyzed using hill numbers for each land use for data based on richness (q=0) and diversity (q=1 and q=2) [28]. We also performed non-metric multidimensional analysis (NMDS), based on the Bray-Curtis index, using the abundance data obtained to graphically express the grouping patterns and differences in species composition. We verified the differentiation of the communities according to habitats with different land uses through a multivariate analysis of variances (Permanova), using the function adonis, with 999 permutations using the vegan package [29].
To evaluate the abundance distribution, we performed a rank abundance curve, which shows the relative abundance of species and their uniformity within each land use, providing essential information for predicting changes in community patterns and allowing us to better explain how the dominance and distribution of species behaved in each land use [30].
To evaluate the effects of habitat (Loja, El Oro, Zamora Chinchipe) on the abundance, food preference, functional group and dung beetle species, Generalized Linear Models (GLMs) were applied using the Poisson distribution in the R software package [31], which helped measure the incidence of the three land uses to the number of coprophagous beetles found in the sampling.
3. Results
We found 209 individuals distributed in 14 genera and 42 species (Figure 2).
One species, Onthophagus curvicornis, represents 20% of the total abundance, while the species Aphodius sp. 1, Canthidium sp. 11, Canthon sp. 1 Canthon sp. 2 Canthon sp. 5, Deltochilum robustus, Dichotomius quinquelobatus, Dichotomius sp. 2, Ontherus brevicollis, Oxysternon silenus, Phanaeus meleagris, Uroxys lojanus, Uroxys sp. 5, Uroxys sp. 7 only represent 0.5% of the total abundance sampled (Table 2). Between provinces, Loja showed the highest abundance (n= 149) and richness (n= 31) representing the 73 % of dung beetles fauna and El Oro was the least abundant (n= 18) and richness (n= 8) representing only the 19 % of coprophagous fauna (Table 2). And according to estimator no parametric Chao 1 in Loja province we have collected 73 %, in El Oro the 63% and Zamora Chinchipe the 69% of estimated dung beetle fauna (Table 2, Figure 2).
According to the rarefaction and extrapolation curves, species richness (q= 0) was higher in the Loja province. Shannon diversity (q= 1) was higher in Loja province. Simpson diversity (q= 2) were higher in the Loja and Zamora Chinchipe province (Figure 2).
The abundance distribution was more uniform in El Oro and Zamora Chinchipe provinces. In Loja province was dominated by O. curvicornis, Canthidium sp. 9 and Canthon balteatus. Eurysternus lanuginosus and Dichotomius prietoi were found only in the El Oro province. In the same way as Canthidium opacum was found only in Zamora Chinchipe province (Figure 3).
With the analysis of non-parametric multidimensional scaling (NMDS), the coprophagous beetle community in Loja was relatively different for El Oro and Zamora Chinchipe (r2= 0.27; p= 0.781) (Figure 4).
In Loja province (Z= 19.164, p<0.001) had a positive effect on dung beetle abundance (Table 3). Zamora Chinchipe (Z= -3.042, p= 0.002) and El Oro (Z= -3.093, p= 0.002) had a negative effect on dung beetle abundance (Table 3). Coprophagous was the food preference significant (Z= 19.76, p<0.001). The functional group did not have a significant effect (Table 3). And the species Canthidium sp. 9, Canthon balteatus, Onthophagus curvicornis, Onthophagus nabelecki and Onthophagus rubrescens had a positive effect (Table 3).
4. Discussion
The high richness of dung beetles (n = 42) recorded in coffee plantations indicates that diversity is remarkably high, being comparable to or even exceeding that reported for other natural ecosystems such as dry grasslands (n = 6 spp.) [32], dry forests (n = 4–6 spp.) [33], Andean forests (35 spp.) [34], and Amazonian forests (n = 54 spp [24,35]. Within these plantations, a variety of microhabitats can coexist, including shaded areas [36], open spaces, and zones with different soil types [33,37]. This habitat heterogeneity likely promotes the coexistence of multiple dung beetle species, contributing to the high overall richness [38,39] and to the maintenance of local biodiversity and associated ecosystem services [37]).
However, it is important to note that a single species, Onthophagus curvicornis, dominated the sampled assemblages, accounting for 20% of the total abundance. This dominance suggests that certain species may play key roles in structuring dung beetle communities within agricultural systems [38].
Dung beetle abundance and richness varied markedly among provinces. Loja exhibited the highest abundance and richness, representing 73% of the sampled dung beetle fauna, whereas El Oro showed the lowest values, accounting for only 19%. These results indicate pronounced spatial heterogeneity in dung beetle communities, likely driven by local environmental conditions, land-use practices, and habitat characteristics. Among the predictor variables evaluated, elevation emerged as the most significant factor across all three models.
The presence of Canthonini varied as a function of elevation, annual precipitation, and precipitation during the coldest quarter. According to the classification tree model, all precipitation-related variables were important predictors, with a validation error of 21%. The probability of occurrence of this tribe decreased with increasing elevation and, more markedly, with increasing annual precipitation, highlighting the importance of rainfall regime and seasonality. In contrast, the occurrence of Coprini increased slightly with elevation, precipitation of the driest month, and precipitation seasonality, with the latter two variables being the most influential according to the classification model. Finally, the presence of Phanaeini was primarily influenced by elevation and mean annual temperature.
Environmental gradients, particularly precipitation and altitude, were identified as key determinants of the distribution of Canthonini and Coprini, whereas temperature exerted a stronger influence on Phanaeini. This pattern is consistent with previous studies emphasizing the role of topography and climate as major drivers of dung beetle diversity and distribution in the Andes and the Amazon basin [37,40,41,42].
The broad probability of occurrence observed for Phanaeini suggests a generalist climatic niche, with occurrence increasing with mean annual temperature and decreasing with elevation, in agreement with documented trends for larger-bodied species with higher thermal tolerance [43]. Conversely, the higher occurrence of Coprini under conditions of strong precipitation seasonality suggests specialization toward environments characterized by temporally variable resource availability [41]. Collectively, these findings demonstrate that dung beetles respond differentially to macroclimatic and topographic gradients, reflecting their ecological sensitivity and reinforcing their value as bioindicators of environmental heterogeneity [37,40].
At fine spatial scales, a more pronounced increase in taxonomic beta diversity was observed compared to functional turnover. This pattern is consistent with observations from tropical communities, where functional structure remains relatively stable despite changes in species composition [41,42,44]. Such stability suggests the presence of functional redundancy, whereby different species perform similar ecological roles [11]. The persistence of a limited number of functional groups across environmental gradients, despite taxonomic turnover, supports the hypothesis that stochastic processes such as dispersal limitation and ecological drift play a key role in structuring local communities [45]. These mechanisms generate spatial heterogeneity and promote high beta diversity even under functional stability [46].
At the regional scale, both taxonomic and functional beta diversity were considerably higher in the Amazon basin than in the coastal region, a pattern consistent with the greater structural complexity and productivity of humid forests [42]. This increased heterogeneity can be attributed to higher availability of trophic resources, particularly dung, which is a key determinant of dung beetle abundance and richness [47,48]. In addition, understory structure and litter accumulation influence nesting activities of functional guilds such as rollers, which depend on humid and shaded microhabitats [14,42].
In contrast, the coastal region exhibited lower levels of species heterogeneity, likely associated with its drier climate, reduced vegetation cover, and limited availability of microhabitats [40,49]. Nevertheless, both regions supported a comparable number of functional groups (12 in the coastal region and 14 in the Amazon basin), with similar responses of functional beta diversity across spatial scales. This convergence suggests that deterministic processes such as niche partitioning [50], interact with stochastic processes, with the latter playing a crucial role in maintaining the high species richness characteristic of Amazonian ecosystems [22].
From a conservation perspective, these results highlight the importance of protecting landscapes that encompass a broad range of environmental gradients, as both taxonomic and functional diversity are essential for ecosystem stability [37]. Dung beetle communities are highly sensitive to habitat degradation, exhibiting reductions in species richness and shifts in community composition in landscapes altered by land-use change [40]. The high beta diversity observed at small spatial scales in the Amazon basin indicates that regional richness is maintained through high species turnover, driven by well preserved vegetation, healthy soils, and favorable conditions for dung burial and reproduction [51].
This study was limited by sampling conducted during a single period of the year, which prevented the assessment of seasonal variation in dung availability, reproductive activity, and community responses to climatic fluctuations. Additionally, key agroecosystem attributes such as understory composition, shade intensity, agricultural management intensity, livestock presence, and land use history were not incorporated, despite their potential influence on dung beetle abundance and species turnover. The exclusive use of pitfall traps baited with pig dung may also have underrepresented species with different trophic preferences or lower attraction to this substrate. Finally, the functional analysis could be strengthened by incorporating quantitative morphological traits such as body size, burial capacity, or dung removal biomass which would allow a more precise estimation of the contribution of each functional guild to ecosystem services within agroecosystems.
5. Conclusions
Shaded coffee plantations in southern Ecuador support a diverse community of dung beetles, whose richness highlights the capacity of these agroecosystems to maintain microhabitats comparable to those found in forest systems. The simultaneous presence of multiple functional guilds and feeding strategies indicates that these crops promote essential ecological processes, particularly those related to organic matter decomposition and nutrient recycling.
Differences in species richness and community composition among provinces reflect the strong influence of environmental variables such as elevation, moisture, and landscape structure, emphasizing that these agroecosystems function as environmental mosaics in which resource availability is shaped by both microclimatic conditions and agricultural management.
Furthermore, the coexistence of rollers, tunnellers, and endocoprids reveals a high degree of functional redundancy, a characteristic that enhances the resilience of the agricultural system. This complementarity of functions facilitates dung removal, improves soil aeration, and contributes to natural pest regulation, thereby strengthening the productive stability of shaded coffee plantations.
Author Contributions
Conceptualization, DMA.; methodology DMA.; formal analysis, AMR and DMA.; investigation, AMR.; data curation, AMR.; writing—original draft preparation, AMR and DMA; writing—review and editing, AMR and DMA funding acquisition, AMR and DMA. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by VIII call for funding for final degree and postgraduate projects 2025 ”of Universidad Técnica Particular de Loja.
Acknowledgments
We are infinitely grateful to: Holger Jumbo, Alex Pardo, Patricio Rodriguez, Luis Alvear, Luis Montaño, Antonio Flores, José Aguirre, Andres Muñoz, Ronald Martínez, Willian Abarca, Lucia Murillo, Edith Añazco, Leonardo Valarezo, and the Kawsana farm, for kindly lending us their farms in the different cantons south of Ecuador. To the students of Biology, Agriculture and Ivonne Gonzalez Coronel of Universidad Técnica Particular de Loja. And, Romina Donoso, for collaborating with us with field and laboratory work.
References
- Cadotte, M.W.; Carscadden, K.A.; Mirotchnick, N. Beyond Species: Functional Diversity and the Maintenance of Ecological Processes and Services. Journal of Applied Ecology 2011, 48, 1079–1087. [Google Scholar] [CrossRef]
- Visscher, A.M.; Chavez, E.; Caicedo, C.; Tinoco, L.; Pulleman, M. Biological Soil Health Indicators Are Sensitive to Shade Tree Management in a Young Cacao (Theobroma Cacao L.) Production System. Geoderma Regional 2024. [Google Scholar] [CrossRef]
- Torres; Torres, J.J.; Mena-Mosquera, V.E.; Rueda-Sánchez, M.N. Influencia de La Altitud Sobre La Composición Florística, Estructura y Carbono de Bosques Del Chocó. UNED Research Journal 2021, 14, e3746–e3746. [Google Scholar] [CrossRef]
- Montero, M.O.V.; Montero, K.E.V. Tipificación de Los Sistemas de Cultivo de Café, Cacao y Ganadero, En La Amazonía Ecuatoriana. Revista Alfa 2024. [Google Scholar] [CrossRef]
- Puyutaxi, F.M.A.; Guambi, L.D.; Corral, R.; Castillo, H.E.G.; Medina, Segundo Alfonso Vasco; Medina, S.A.V.; Motato, N.; Larrea, G.R.S.; L.Z., A.; T.A., Z.; et al. Variedades Mejoradas de Café Arábigo Una Contribución Para El Desarrollo de La Caficultura En El Ecuador; 2004. [Google Scholar]
- Álvarez, O.; Maldonado, J.; Montaño, T.; Caraballo, M.Á. Estabilidad Vertical de La Atmósfera En Las Provincias de Loja y Zamora Chinchipe, Ecuador. 2016, 6. [Google Scholar]
- Harvey, C.A.; Pritts, A.A.; Zwetsloot, M.J.; Jansen, K.; Pulleman, M.M.; Armbrecht, I.; Avelino, J.; Barrera, J.F.; Bunn, C.; García, J.H.; et al. Transformation of Coffee-Growing Landscapes across Latin America. A Review. Agron. Sustain. Dev. 2021, 41, 62. [Google Scholar] [CrossRef]
- Salazar, J.C.; Burgos, E.R.; Bautista, E.H.D. Efecto de Las Condiciones de Cultivo, Las Características Químicas Del Suelo y El Manejo de Grano En Los Atributos Sensoriales de Café (Coffea Arabica L.) En Taza. Acta Agronómica 2015, 64, 342–348. [Google Scholar] [CrossRef]
- Sadeghian, S. Nutrición Del Café. Consideraciones Para El Manejo de La Fertilidad Del Suelo. 2022. [Google Scholar] [CrossRef]
- Guambi, L.A.D.; Murillo, R.M.A.; Alonso, P.C.P. La Competitividad de Los Cafés Arábigos de Especialidades En Los Concursos “Taza Dorada” En Ecuador. Journal of Agricultural Sciences Resarch 2022, 2, 2–9. [Google Scholar] [CrossRef]
- deCastro-Arrazola, I.; Andrew, N.R.; Berg, M.P.; Curtsdotter, A.; Lumaret, J.-P.; Menéndez, R.; Moretti, M.; Nervo, B.; Nichols, E.S.; Sánchez-Piñero, F.; et al. A Trait-based Framework for Dung Beetle Functional Ecology. Journal of Animal Ecology 2022. [Google Scholar] [CrossRef]
- Pfiffner, L.; Balmer, O. La Agriculture Ecológica Fomenta La Biodiversidad; 2011. [Google Scholar]
- Newbold, T.; Hudson, L.N.; Hill, S.L.L.; Contu, S.; Lysenko, I.; Senior, R.A.; Börger, L.; Bennett, D.J.; Choimes, A.; Collen, B.; et al. Global Effects of Land Use on Local Terrestrial Biodiversity. Nature 2015, 520, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Braga, R.F.; Korasaki, V.; Andresen, E.; Louzada, J. Dung Beetle Community and Functions along a Habitat-Disturbance Gradient in the Amazon: A Rapid Assessment of Ecological Functions Associated to Biodiversity. PLOS ONE 2013, 8. [Google Scholar] [CrossRef]
- Noriega, J.; Hortal, J.; DeCastro-Arrazola, I.; Alves-Martins, F.; Ortega, J.; Bini, L.; Andrew, N.; Arellano, L.; Beynon, S.; Davis, A.; et al. Dung Removal Increases under Higher Dung Beetle Functional Diversity Regardless of Grazing Intensification. Nature communications 2023. [Google Scholar] [CrossRef]
- Slade, E.M.; Riutta, T.; Roslin, T.; Tuomisto, H. The Role of Dung Beetles in Reducing Greenhouse Gas Emissions from Cattle Farming. Scientific Reports 2016, 6, 18140–18140. [Google Scholar] [CrossRef]
- Piccini, I.; Arnieri, F.; Caprio, E.; Nervo, B.; Pelissetti, S.; Palestrini, C.; Roslin, T.; Rolando, A. Greenhouse Gas Emissions from Dung Pats Vary with Dung Beetle Species and with Assemblage Composition. PLOS ONE 2017, 12, 1–15. [Google Scholar] [CrossRef]
- Andriuzzi, W.S.; Wall, D.H. Soil Biological Responses to, and Feedbacks on, Trophic Rewilding. Philosophical Transactions of the Royal Society B 2018, 373, 20170448. [Google Scholar] [CrossRef] [PubMed]
- Yaguana, C.; Lozano Deicy; Aguirre, Z. Flora y Endemismo Del Bosque Humedo Tropical de La Quinta El Padmi, Zamora Chinchipe; 2010. [Google Scholar]
- Loayza, V.E.; Rojas, D.; Salas Tenesaca, E.E.; Samaniego Namicela, A. Fortalecimiento Organizacional de Asociaciones de Productores de Café En Las Provincias de Loja y Zamora Chinchipe, Ecuador. 2019, 5. [Google Scholar] [CrossRef]
- López-Dávila, A.J.; Escobar-Ramírez, S.; Armbrecht, I. Nesting of Arboreal Ants (Hymenoptera: Formicidae) in Artificial Substrates in Coffee Plantations in the Colombian Andes. Uniciencia 2021, 35, 1–17. [Google Scholar] [CrossRef]
- Nunes, C.A.; Barlow; França, J.; Filipe; Berenguer, E.; Solar, R.R.C.; Louzada, J.; Leitão, R.P.; Maia, L.F.; Oliveira, V.H.F.; Braga, R.F.; et al. Functional Redundancy of Amazonian Dung Beetles Confers Community-Level Resistance to Primary Forest Disturbance. Biotropica 2021. [Google Scholar] [CrossRef]
- Mora-Aguilar, E.F.; Arriaga-Jiménez, A.; Correa, C.M.A.; da Silva, P.G.; Korasaki, V.; López-Bedoya, P.A.; Hernández, M.I.M.; Pablo-Cea, J.D.; Salomão, R.P.; Valencia, G.; et al. Toward a Standardized Methodology for Sampling Dung Beetles (Coleoptera: Scarabaeinae) in the Neotropics: A Critical Review. Frontiers in Ecology and Evolution 2023, 11. [Google Scholar] [CrossRef]
- Chamorro, W.; Marín-Armijos, D.; Asenjo, A.; Vaz-de-Mello, F.Z. Scarabaeinae Dung Beetles from Ecuador: A Catalog, Nomenclatural Acts, and Distribution Records. ZooKeys 2019, 826, 1–343. [Google Scholar] [CrossRef]
- Callaghan, C.T.; Santini, L.; Spake, R.; Bowler, D.E. Population Abundance Estimates in Conservation and Biodiversity Research. Trends in Ecology & Evolution 2024. [Google Scholar] [CrossRef]
- Clusella-Trullas, S.; Nielsen, M.E. The Evolution of Insect Body Coloration under Changing Climates. Current opinion in insect science 2020, 41, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Silva, P.G.D. Annotated Checklist of Aphodiinae (Coleoptera: Scarabaeidae) from Rio Grande Do Sul and Santa Catarina, Brazil. EntomoBrasilis 2015, 8, 145–151. [Google Scholar] [CrossRef]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and Extrapolation with Hill Numbers: A Framework for Sampling and Estimation in Species Diversity Studies. Ecological Monographs 2014, 84, 45–67. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-6 2019. Available online: https://CRAN.R-project.org/package=vegan.
- Whittaker, R.J.; Willis, K.J.; Field, R. Scale and Species Richness: Towards a General, Hierarchical Theory of Species Diversity. Journal of Biogeography 2001, 28, 453–470. [Google Scholar] [CrossRef]
- R-Core-Team R: A Language and Environment for Statistical Computing. In R: A language and environment for statistical computing.; 2019.
- Domínguez, D.; Marín-Armijos, D.; Ruiz, C. Structure of Dung Beetle Communities in an Altitudinal Gradient of Neotropical Dry Forest. Neotrop Entomol 2015, 44, 40–46. [Google Scholar] [CrossRef]
- Carrión-Paladines, V.; Fries, A.; Muñoz, A.; Castillo, E.; García-Ruiz, R.; Marín-Armijos, D. Effects of Land-Use Change on the Community Structure of the Dung Beetle (Scarabaeinae) in an Altered Ecosystem in Southern Ecuador. Insects 2021, 12, 306. [Google Scholar] [CrossRef]
- Cárdenas-Bautista, J.S.; Parada-Alfonso, J.A.; Carvajal-Cogollo, J.E. Dung Beetles (Scarabaeidae, Scarabaeinae) of the Foothills–Andean Forest Strip of Villavicencio, Colombia. CheckList 2020, 16, 821–839. [Google Scholar] [CrossRef]
- Espinoza, V.R.; Noriega, J.A. Diversity of the Dung Beetles (Coleoptera: Scarabaeinae) in an Altitudinal Gradient in the East Slope of Los Andes, Napo Province, Ecuador. Neotropical Biodiversity 2018, 4, 145–151. [Google Scholar] [CrossRef]
- Arenas-Clavijo, A.; Armbrecht, I. Soil Ants (Hymenoptera: Formicidae) and Ground Beetles (Coleoptera: Carabidae) in a Coffee Agroforestry Landscape during a Severe-Drought Period. Agroforest Syst 2019, 93, 1781–1792. [Google Scholar] [CrossRef]
- Nichols, E.; Larsen, T.H.; Spector, S.; Davis, A.L.V.; Escobar, F.; Favila, M.E.; Vulinec, K. Global Dung Beetle Response to Tropical Forest Modification and Fragmentation: A Quantitative Literature Review and Meta-Analysis. Biological Conservation 2007, 137, 1–19. [Google Scholar] [CrossRef]
- Carvalho, E.C.; Maldaner, M.; Costa-Silva, V.; Sehn, H.; Franquini, C.; Campos, V.; Seba, V.; Maia, L.; Vaz-de-Mello, F.; França, F. Dung Beetles from Two Sustainable-Use Protected Forests in the Brazilian Amazon. BDJ 2023, 11, e96101. [Google Scholar] [CrossRef] [PubMed]
- Ratoni, B.; Ahuatzin, D.; Corro, E.J.; Salomão, R.P.; Escobar, F.; López-Acosta, J.C.; Dáttilo, W. Landscape Composition Shapes Biomass, Taxonomic and Functional Diversity of Dung Beetles within Human-Modified Tropical Rainforests. J Insect Conserv 2023, 27, 717–728. [Google Scholar] [CrossRef]
- Halffter, G.; Arellano, L. Response of Dung Beetle Diversity to Human-Induced Changes in a Tropical Landscape1. Biotropica 2002, 34, 144–154. [Google Scholar] [CrossRef]
- Horgan, F.G. Invasion and Retreat: Shifting Assemblages of Dung Beetles amidst Changing Agricultural Landscapes in Central Peru. Biodivers Conserv 2009, 18, 3519–3541. [Google Scholar] [CrossRef]
- Nichols, E.; Spector, S.; Louzada, J.; Larsen, T.; Amezquita, S.; Favila, M.E. Ecological Functions and Ecosystem Services Provided by Scarabaeinae Dung Beetles. Biological Conservation 2008, 141, 1461–1474. [Google Scholar] [CrossRef]
- Bremer, L.L.; Farley, K.A. Does Plantation Forestry Restore Biodiversity or Create Green Deserts? A Synthesis of the Effects of Land-Use Transitions on Plant Species Richness. Biodivers Conserv 2010, 19, 3893–3915. [Google Scholar] [CrossRef]
- Larsen, T.H.; Lopera, A.; Forsyth, A. Understanding Trait-Dependent Community Disassembly: Dung Beetles, Density Functions, and Forest Fragmentation. Conservation Biology 2008, 22, 1288–1298. [Google Scholar] [CrossRef]
- Cultid-Medina, C.A.; Escobar, F. Assessing the Ecological Response of Dung Beetles in an Agricultural Landscape Using Number of Individuals and Biomass in Diversity Measures. Environmental Entomology 2016, 45, 310–319. [Google Scholar] [CrossRef]
- Morales Alba, Andrés Felipe; Carvajal-Cogollo, Juan E.; Morales Irina. Diversidad de Escarabajos Coprófagos En Dos Periodos de Precipitación Anual En Un Fragmento de Bosque Andino, Santander, Colombia. Intropica 2023, 50–64. [Google Scholar] [CrossRef]
- Campos, R.C.; Hernández, M.I.M. Dung Beetle Assemblages (Coleoptera, Scarabaeinae) in Atlantic Forest Fragments in Southern Brazil. Rev. Bras. entomol. 2013, 57, 47–54. [Google Scholar] [CrossRef]
- Silva, P.G.D.; Hernández, M.I.M. Scale-Dependence of Processes Structuring Dung Beetle Metacommunities Using Functional Diversity and Community Deconstruction Approaches. PLoS ONE 2015, 10, e0123030. [Google Scholar] [CrossRef]
- Escobar-Ramírez, S.; Tscharntke, T.; Armbrecht, I.; Torres, W.; Grass, I. Decrease in B-diversity, but Not in A-diversity, of Ants in Intensively Managed Coffee Plantations. Insect Conserv Diversity 2020, 13, 445–455. [Google Scholar] [CrossRef]
- Da Silva, P.G.; Lobo, J.M.; Hernández, M.I.M. The Role of Habitat and Daily Activity Patterns in Explaining the Diversity of Mountain Neotropical Dung Beetle Assemblages. Austral Ecology 2019, 44, 300–312. [Google Scholar] [CrossRef]
- Maldonado, M.B.; Aranibar, J.N.; Serrano, A.M.; Chacoff, N.P.; Vázquez, D.P. Dung Beetles and Nutrient Cycling in a Dryland Environment. CATENA 2019, 179, 66–73. [Google Scholar] [CrossRef]
Figure 1.
Location of the 14 coffee plantations sampled in the provinces of El Oro, Loja and Zamora Chinchipe in the South of Ecuador.
Figure 1.
Location of the 14 coffee plantations sampled in the provinces of El Oro, Loja and Zamora Chinchipe in the South of Ecuador.
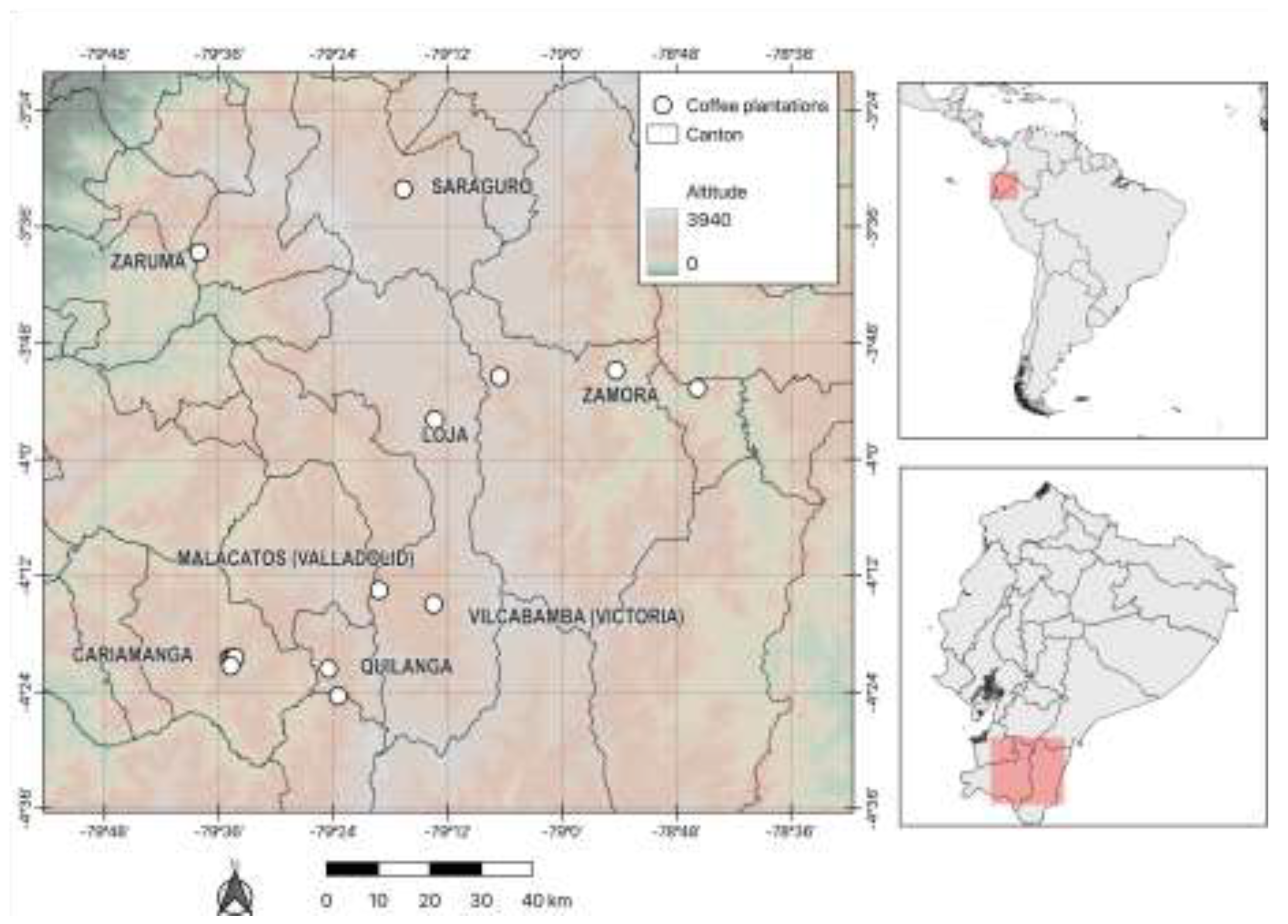
Figure 2.
Hill numbers for each province in southern Ecuador: LOJ (Loja), ORO (El Oro) and ZCH (Zamora Chinchipe).
Figure 2.
Hill numbers for each province in southern Ecuador: LOJ (Loja), ORO (El Oro) and ZCH (Zamora Chinchipe).
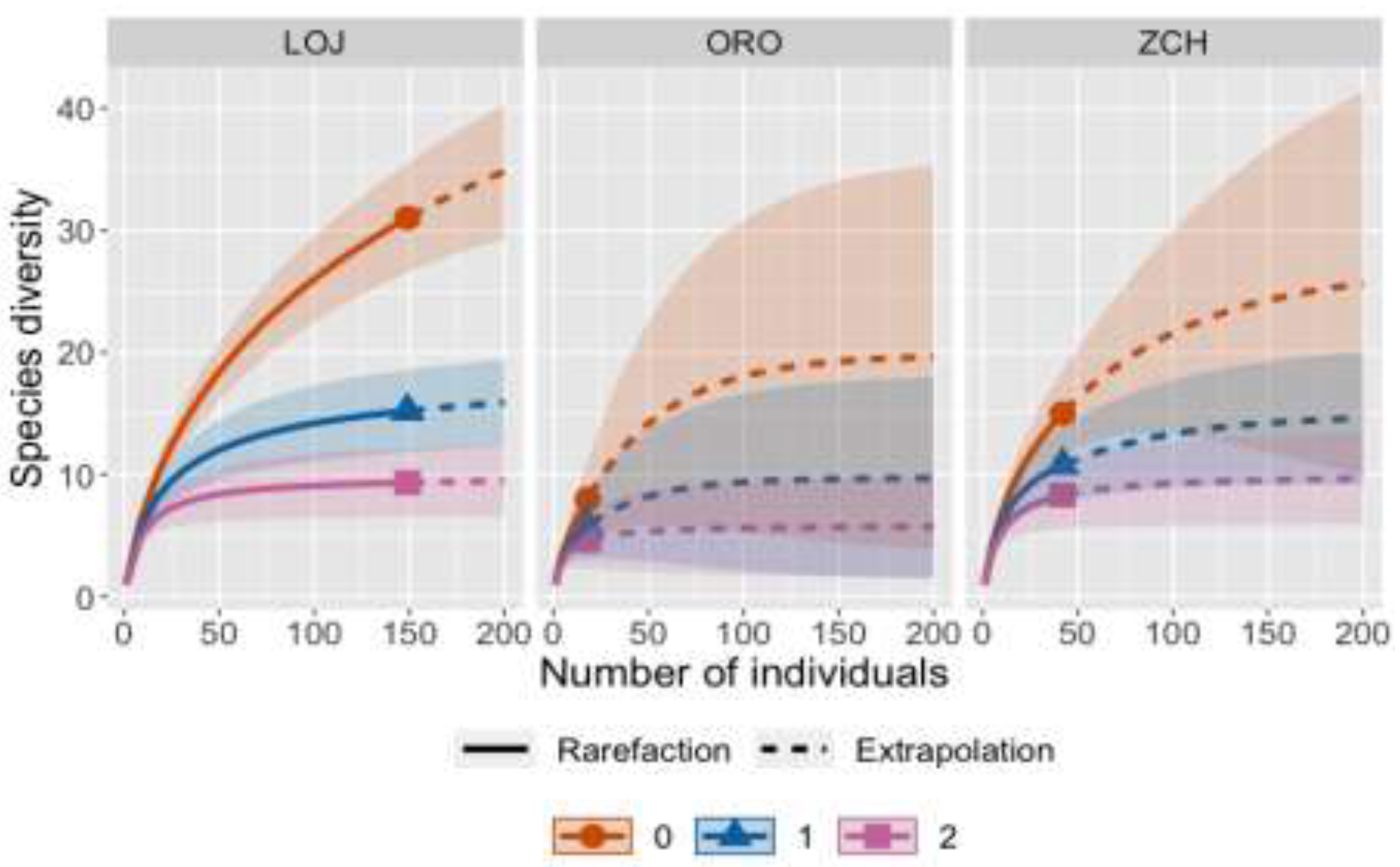
Figure 3.
Rank abundance curve of dung beetles in each province (LOJ= Loja, ORO= El Oro, and ZCH= Zamora Chinchipe).
Figure 3.
Rank abundance curve of dung beetles in each province (LOJ= Loja, ORO= El Oro, and ZCH= Zamora Chinchipe).
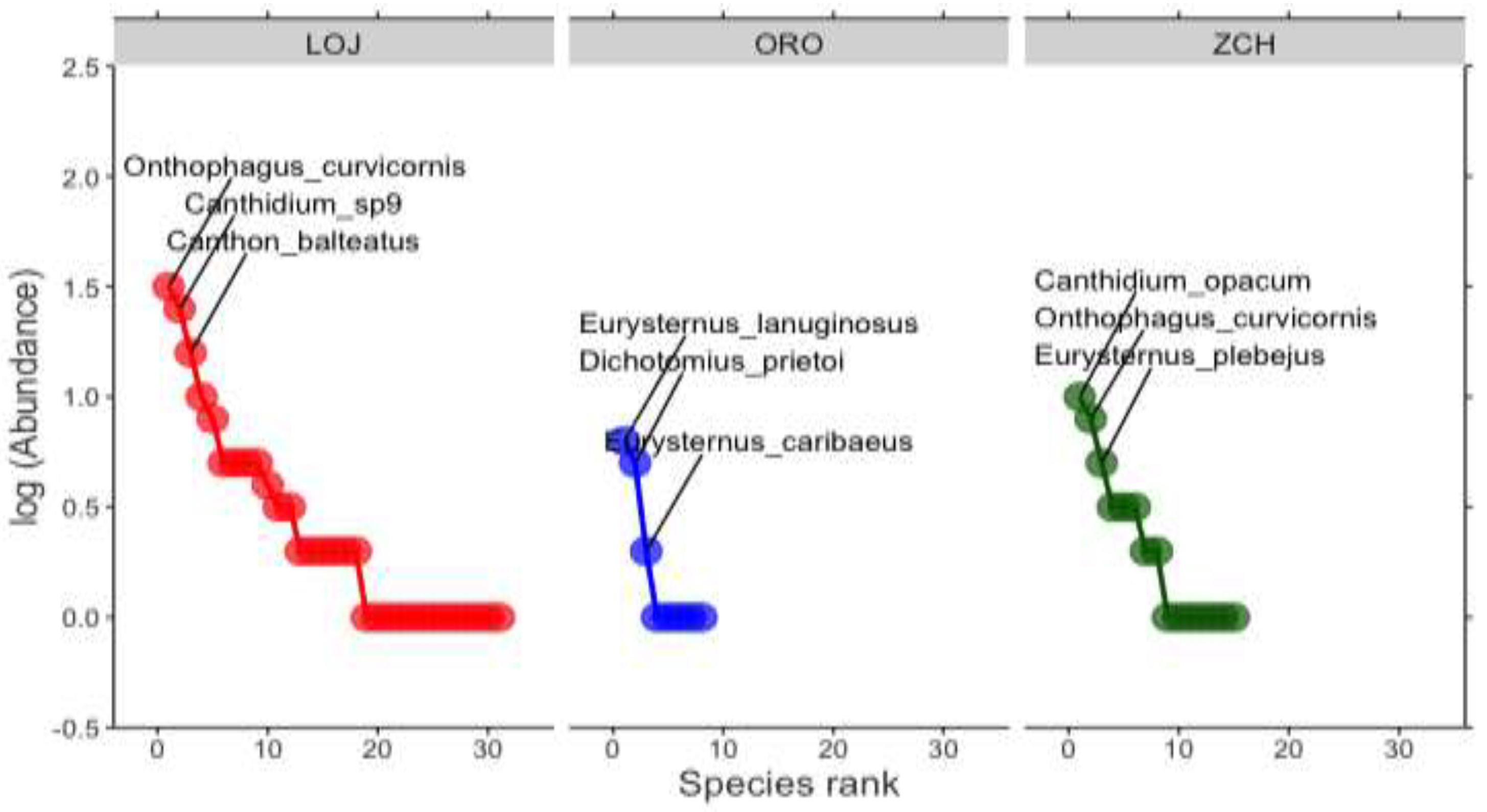
Figure 4.
NMDS ordination showed that the dung beetle communities clustered according to habitat: Loja, El Oro y Zamora Chinchipe. Colors: red, Loja; green, Zamora Chinchipe; blue, El Oro.
Figure 4.
NMDS ordination showed that the dung beetle communities clustered according to habitat: Loja, El Oro y Zamora Chinchipe. Colors: red, Loja; green, Zamora Chinchipe; blue, El Oro.

Table 1.
Details of the sampling sites, province, canton, latitude, longitude and altitude.
| Province | Canton | Latitude (S) | Longitude (W) | Altitude (m) |
|---|---|---|---|---|
| El Oro | Zaruma | -3.749 | -79.727 | 962 |
| -3.719 | -79.65 | 1123 | ||
| -3.644 | -79.635 | 1115 | ||
| Loja | Cariamanga | -4.342 | -79.582 | 1815 |
| -4.343 | -79.577 | 1865 | ||
| -4.34 | -79.571 | 2002 | ||
| -4.355 | -79.579 | 2095 | ||
| Loja | -3.931 | -79.222 | 2100 | |
| -4.224 | -79.319 | 1969 | ||
| -4.249 | -79.224 | 1700 | ||
| Quilanga | -4.360 | -79.408 | 1606 | |
| -4.406 | -79.391 | 1477 | ||
| Saraguro | -3.535 | -79.277 | 1180 | |
| Zamora Chinchipe | Zamora | -3.847 | -78.905 | 1012 |
| -3.858 | -79.109 | 1856 | ||
| -3.878 | -78.762 | 817 |
Table 2.
Details about the dung beetles sampled in coffee plantations in three provinces located south of Ecuador: LOJ (Loja), Oro (El Oro), ZCH (Zamora Chinchipe). Relevant characteristics of each species are provided, including food preference (C= Coprophagous, G= Generalist), functional group (E= Endocoprid, P= Paracoprid, T= Telecoprid).
Table 2.
Details about the dung beetles sampled in coffee plantations in three provinces located south of Ecuador: LOJ (Loja), Oro (El Oro), ZCH (Zamora Chinchipe). Relevant characteristics of each species are provided, including food preference (C= Coprophagous, G= Generalist), functional group (E= Endocoprid, P= Paracoprid, T= Telecoprid).
| Species | LOJ | ORO | ZCH | Total | Food preference | Functional group |
|---|---|---|---|---|---|---|
| Aphodius sp. 1 Illiger, 1798 | 1 | 1 | C | E | ||
| Canthidium coerulescens Balthasar, 1939 | 3 | 1 | 1 | 5 | C | P |
| Canthidium opacum Balthasar, 1939 | 5 | 9 | 14 | C | P | |
| Canthidium sp. 11 Erichson, 1847 | 1 | 1 | C | P | ||
| Canthidium sp. 9 Erichson, 1847 | 24 | 24 | C | P | ||
| Canthon balteatus Boheman, 1858. | 17 | 17 | C | T | ||
| Canthon sp. 1 Hoffmannsegg, 1817 | 1 | 1 | C | T | ||
| Canthon sp. 2 Hoffmannsegg, 1817 | 1 | 1 | C | T | ||
| Canthon sp. 4 Hoffmannsegg, 1817 | 5 | 5 | C | T | ||
| Canthon sp. 5 Hoffmannsegg, 1817 | 1 | 1 | C | T | ||
| Canthon sp. 7 Hoffmannsegg, 1817 | 2 | 2 | C | T | ||
| Coprophanaeus ohausi Felsche, 1911 | 2 | 2 | G | P | ||
| Deltochilum robustus Molano & González, 2009 | 1 | 1 | G | T | ||
| Dichotomius talaus Blanchard, 1845 | 1 | 1 | 2 | C | P | |
| Dichotomius prietoi Martínez & Martínez, 1982 | 5 | 5 | C | P | ||
| Dichotomius problematicus Lüederwaldt, 1924 | 5 | 1 | 6 | C | P | |
| Dichotomius protectus Roze 1955 | 3 | 3 | C | P | ||
| Dichotomius quinquelobatus Felsche, 1901 | 1 | 1 | C | P | ||
| Dichotomius sp. 2 Hope, 1838 | 1 | 1 | C | P | ||
| Eurysternus caribaeus Herbst, 1789 | 2 | 2 | 4 | C | E | |
| Eurysternus lanuginosus Génier, 2009 | 6 | 6 | C | E | ||
| Eurysternus plebejus Harold, 1880 | 1 | 5 | 6 | C | E | |
| Onoreidium ohausi Arrow, 1931 | 2 | 2 | C | P | ||
| Ontherus brevicollis Kirsch, 1871 | 1 | 1 | C | P | ||
| Ontherus pubens Génier, 1996 | 4 | 2 | 6 | C | P | |
| Onthophagus confusus Boucomont, 1932 | 5 | 5 | C | P | ||
| Onthophagus curvicornis Latreille, 1811 | 34 | 8 | 42 | C | P | |
| Onthophagus dicranoides Balthasar, 1939 | 2 | 2 | C | P | ||
| Onthophagus nabelecki Balthasar, 1939 | 11 | 11 | C | P | ||
| Onthophagus rubrescens Blanchard, 184 | 8 | 8 | C | P | ||
| Onthophagus transisthmius Howden & Young, 1981 | 3 | 3 | C | P | ||
| Oxysternon conspicillatum Weber, 1801 | 1 | 1 | 2 | C | P | |
| Oxysternon silenus d’Olsoufieff, 1924 | 1 | 1 | C | P | ||
| Phanaeus achilles Boheman, 1858 | 3 | 3 | C | P | ||
| Phanaeus haroldi Kirsch, 1871 | 1 | 3 | 4 | C | P | |
| Phanaeus lunaris Taschenberg, 1870 | 1 | 1 | 2 | C | P | |
| Phanaeus meleagris Blanchard, 1843 | 1 | 1 | C | P | ||
| Scatimus mostrosus Balthasar, 1939 | 2 | 2 | C | P | ||
| Uroxys lojanus Arrow, 1933 | 1 | 1 | C | P | ||
| Uroxys sp. 5 Westwood, 1842 | 1 | 1 | C | P | ||
| Uroxys sp. 6 Westwood, 1842 | 2 | 2 | C | P | ||
| Uroxys sp. 7 Westwood, 1842 | 1 | 1 | C | P | ||
| Abundance | 149 | 18 | 42 | 209 | ||
| Richness | 31 | 8 | 15 | 42 | ||
| Chao1 | 42.1 | 12.7 | 21.8 |
Table 3.
Effects of habitat (Loja, El Oro, Zamora Chinchipe), food preference (Coprophagous, Generalist), functional group (Endocoprid, Paracoprid, Telecoprid) and species on the abundance of dung beetles. Significant values (p < 0.05) are highlighted in bold.
Table 3.
Effects of habitat (Loja, El Oro, Zamora Chinchipe), food preference (Coprophagous, Generalist), functional group (Endocoprid, Paracoprid, Telecoprid) and species on the abundance of dung beetles. Significant values (p < 0.05) are highlighted in bold.
| Responce Variable | Explanatory Variable | Standard error | Z-value | p-value |
|---|---|---|---|---|
| Abundance | Habitat | |||
| (Intercept) | 0.082 | 19.164 | <0.001 | |
| El Oro | 0.250 | -3.042 | 0.002 | |
| Zamora Chinchipe | 0.175 | -3.093 | 0.002 | |
| Chi test: | ||||
| Habitat | <0.001 | |||
| Abundance | Food preference | |||
| (Intercept) | 0.070 | 19.76 | <0.001 | |
| Generalist | 0.581 | -1.67 | 0.095 | |
| Chi test: | ||||
| Food preference | 0.048 | |||
| Abundance | Functional group | |||
| (Intercept) | 0.243 | 4.294 | <0.001 | |
| Paracoprid | 0.255 | 1.353 | 0.176 | |
| Telecoprid | 0.308 | 1.122 | 0.262 | |
| Chi test: | ||||
| Functional group | 0.360 | |||
| Abundance | Species | |||
| (Intercept) | 1 | 0 | 1 | |
| Canthidium sp. 9 | 1.021 | 3.114 | 0.002 | |
| Canthon balteatus | 1.029 | 2.753 | 0.006 | |
| Onthophagus curvicornis | 1.012 | 3.009 | 0.003 | |
| Onthophagus nabelecki | 1.044 | 2.296 | 0.022 | |
| Onthophagus rubrescens | 1.061 | 1.961 | 0.050 | |
| Chi test: | ||||
| Species | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



