
Submitted:
13 January 2026
Posted:
14 January 2026
You are already at the latest version
Abstract
Central nervous system (CNS) disorders constitute a significant global health challenge; however, the development of therapeutic agents is considerably impeded by the difficulty in delivering effective concentrations within the brain. This comprehensive review delineates the current landscape of computational modeling techniques employed to address the formidable challenges associated with CNS drug delivery, with a particular emphasis on the anatomical barriers and physiological transport mechanisms pertinent to major neurological diseases. We categorize modeling approaches ranging from the atomistic scale, including molecular dynamics simulations of drug-blood-brain barrier (BBB) interactions, to macroscopic continuum and Physiologically Based Pharmacokinetic (PBPK) models that elucidate systemic distribution and overall brain exposure. We critically assess these models concerning established delivery routes, such as intranasal and intrathecal administration, as well as emerging methods, including focused ultrasound-mediated BBB opening and targeted nanoparticle delivery. This review underscores the growing importance of integrating complex physiological phenomena, such as glymphatic flow and cerebrospinal fluid (CSF) dynamics, into predictive models. Finally, we explore the emerging opportunities involving multiscale digital twins of the CNS that integrate molecular interactions, vascular hemodynamics, CSF and perivascular flow, and parenchymal transport within patient-specific anatomical geometries. The role of machine learning and surrogate modeling in expediting the prediction of drug transport parameters and optimizing delivery strategies is also examined. By providing a structured overview of current computational tools, this review aims to guide researchers in the design of more robust computational platforms for CNS drug delivery.
Keywords:
CNS
; blood brain barrier
; alzheimer’s
; nanoparticle
; drug delivery
; computational modeling
; continuum
; molecular dynamics
; PBPK
; machine learning
1. Introduction
Central nervous system (CNS) disorders represent an escalating and expensive global health issue. Neuropsychiatric and neurodegenerative conditions are among the most pressing health challenges worldwide in this century [1]. Additionally, neurological disorders like Alzheimer’s disease (AD) and Parkinson’s disease (PD) predominantly impact the swiftly growing elderly demographic. Patients with primary brain tumors or brain metastases have limited treatment options, primarily depending on surgical removal, systemic chemotherapy, or radiation therapy. As a result, many pharmaceutical companies have decreased their investment in CNS drug development due to high costs, lengthy regulatory processes, and historically low clinical success rates [1].
Biomaterial-based delivery systems present a promising path for rejuvenating CNS therapeutics. Recent advances in precision biomaterial synthesis allow for the creation of carriers that can be selectively functionalized, engineered to react to physiological or external stimuli, and designed with adjustable degradation properties [2]. Despite that, delivering therapeutic agents to the CNS remains one of the greatest challenges in drug development because of the multilayered barriers protecting the brain and spinal cord, including the blood-brain barrier (BBB), nasal mucosa, and complex cerebrospinal fluid (CSF) circulation patterns. Moreover, inter-species differences and disease effects on transport barriers add to the translation challenges. Mathematical and computational models offer great potential by simulating active targeting in the human neurovascular environment, thus paving the way for optimizing drug design and in-silico clinical trials leading to accelerated drug discovery and reduced development costs [3,4].
In this Review, we explore how both normal and diseased CNS physiology affects drug delivery to the brain. We discuss the pathophysiological barriers that complicate delivery and summarize biomaterial systems that can be administered systemically, intrathecally, intranasally, or peripherally. We then examine various classes of computational models for these CNS drug delivery approaches, detailing their key assumptions, strengths, and limitations—including physiologically based pharmacokinetic (PBPK) modeling [5,6], atomistic simulations [7,8], hybrid multiscale approaches [4,9], continuum fluid–structure and solute-transport frameworks [10,11], and machine-learning–driven prediction [12,13]. The Review concludes with a perspective on emerging opportunities for multiscale digital twins, hybrid physics–ML modeling, and clinical translation challenges.
2. Blood Brain Barrier
Delivering pharmaceuticals to the CNS presents a significant challenge due to the intricate physiological barriers, particularly the BBB. The BBB functions as a highly selective, semipermeable interface between the bloodstream and the CNS, primarily composed of brain microvascular endothelial cells connected by tight junctions [14,15]. It is crucial for maintaining CNS homeostasis by regulating nutrient flow, ionic balance, and preventing the entry of potentially harmful substances. The physical barrier of the BBB is constituted by tight junction protein complexes among endothelial cells, such as occludin, claudins, and junctional adhesion molecules (JAMs), which restrict the paracellular movement of polar solutes and large molecules [15]. Endothelial cells are supported by a basement membrane, extracellular matrix (ECM), pericytes, and astrocytic endfeet, forming the neurovascular unit (NVU), a cohesive multicellular interface that governs barrier permeability and communication between the CNS and peripheral blood. The ECM presents significant transport resistance at different levels. The vascular basement membrane, which is rich in collagen, laminins, and heparan sulfate proteoglycans, constitutes a dense, highly charged barrier that impedes diffusion and obstructs the movement of macromolecules and nanoparticles in the vicinity of CNS microvessels. Beyond the vasculature, the bulk parenchymal ECM, comprising hyaluronan, chondroitin sulfate proteoglycans, and components of astrocytic scars, further restricts solute mobility by increasing tortuosity and reducing effective diffusivity, particularly under neurodegenerative or inflammatory conditions. (Figure 1).
In addition to its physical barrier properties, the BBB also possesses a metabolic barrier composed of highly expressed efflux pumps and enzymes that metabolize or expel xenobiotics. Key transporters include P-glycoprotein (P-gp/ABCB1) and breast cancer resistance protein (BCRP/ABCG2), which prevent the accumulation of many therapeutics and endogenous metabolites in the CNS [15]. This selectivity is further reinforced by the minimal pinocytotic activity of BBB endothelial cells, which limits the nonspecific uptake of circulating molecules.
Successful CNS drug delivery depends on the ability of compounds to traverse this barrier, which is determined by their molecular size, lipophilicity, charge, and structural similarity to endogenous substrates. In healthy brain tissue, the hydrophilic cleft between adjacent endothelial cells measures approximately 0.8 nm, which is significantly narrower than the 10–25 nm intercellular gaps typically observed in non-BBB microvascular endothelium [16]. Molecules with molecular weights below approximately 400–500 Da and high lipophilicity are more likely to traverse across the BBB, although this is necessary but not sufficient for brain penetration [1]. For larger or hydrophilic molecules, entry into the CNS may occur through selective transport mechanisms, including carrier-mediated transport (e.g., glucose, amino acids) or receptor-mediated transcytosis (e.g., insulin, transferrin). Among the available receptor targets, low-density lipoprotein receptor–related protein-1 (LRP-1) is particularly attractive because it is abundantly expressed on brain capillary endothelium and frequently overexpressed in glioblastoma, enabling dual-site receptor engagement for therapeutic delivery [1,2,14]. Conversely, certain functional groups or inappropriate charge states at physiological pH can impede passive BBB transport. Additionally, the activity of efflux pumps like P-gp significantly limits brain accumulation even for molecules with favorable passive diffusion properties [15]. Due to the complexity of BBB permeability determinants, computational modeling and in silico prediction tools have increasingly been employed to screen and optimize CNS drug candidates in the early stages of discovery. Quantitative structure-activity relationships (QSAR), PBPK models, and machine-learning frameworks are routinely utilized to assess BBB permeability, prioritize compounds for synthesis, and guide structural modifications [1,17].
Although the BBB is known for its restrictive properties, it does allow certain peptides and proteins to pass through via adsorptive-mediated transcytosis, especially larger cationic molecules like albumin, immunoglobulins, leptin, and growth factors. Recent advancements in drug delivery have utilized receptor- or adsorptive-mediated transcytosis with ligands such as transferrin or Angiopep-2 to facilitate the transport of therapeutic agents into the brain [2]. Nonetheless, these approaches encounter obstacles, including endosomal entrapment, receptor saturation, and unintended effects on non-target areas.
3. Pathophysiological Barriers to Effective Drug Delivery in Major CNS Diseases
The diverse pathophysiological changes associated with CNS diseases significantly alter the extracellular space, extracellular matrix, and neurovascular unit, resulting in a microenvironment that is distinctly different from that of a healthy brain. These disease-induced pathological remodeling mechanisms, which include vascular dysfunction, enzymatic remodeling, neuroinflammation, and cellular disruption, affect the structure and biophysical properties of the interstitial space, thereby impacting the penetration and distribution of drug delivery systems within the parenchyma. Such disturbances may either impede therapeutic transport by increasing tortuosity and barrier resistance or temporarily enhance permeability through the breakdown of vascular or matrix integrity. Consequently, comprehending how disease physiology reconfigures these microdomains is crucial for developing delivery strategies that remain effective across various pathological contexts.
3.1. Neurodegenerative Diseases
Neurodegenerative diseases are characterized by the progressive loss of neuronal structure and function, often accompanied by pathological changes within the neurovascular unit [18,19]. Evidence of endothelial degeneration and diminished BBB function has been reported in amyotrophic lateral sclerosis (ALS), Parkinson’s disease (PD), and Alzheimer’s disease (AD), underscoring the central role of neurovascular dysfunction in aging-related neurodegeneration [15]. BBB breakdown often precedes clinical symptoms and results from the compounding effect of increased reactive gliosis, neurovascular uncoupling, heightened neuroinflammation, and progressive neuronal decline [20]. These processes reflect the collapse of CNS homeostasis and pose substantial challenges for the systemic and local delivery of therapeutics. Neurovascular alterations also manifest in psychiatric conditions such as schizophrenia and chronic stress disorders. These include basement membrane thickening, astrocytic end-foot retraction, microglial activation, and chronic inflammation in the perivascular compartments (Figure 2). Emerging evidence indicates that vascular dysfunction in these disorders often precedes overt neuronal damage, supporting its causative role in pathology and cognitive dysfunction [21,22]. Chronic BBB dysregulation contributes to secondary pathologies that impede drug penetration into the CNS. Downregulation of tight junction proteins increases endothelial permeability, leading to perivascular space (PVS) enlargement and the accumulation of plasma-derived neurotoxic proteins, such as fibrinogen. Concurrently, vascular dysfunction drives ECM modifications within both the vascular basement membrane and the parenchymal interstitium. The modifications include amyloid deposition, sulfated proteoglycan accumulation, and glial scarring, which impair neurovascular integrity and transport properties. Pericytes, which are pivotal for maintaining BBB structure and function, are progressively lost in angiopathy associated with neurodegeneration, resulting in microvascular rarefaction and compromised perfusion [23,24,25,26]. These microenvironmental adaptations create barriers that challenge traditional drug delivery strategies. Formulations optimized for intact BBB penetration are hindered by altered vascular architecture, dysfunctional glymphatic clearance, and expanded perivascular spaces (PVS), necessitating innovative strategies that exploit or circumvent these pathological changes.
3.1.1. Alzheimer’s Disease
Alzheimer’s disease (AD) is characterized by extracellular amyloid-β (Aβ) plaque formation, tau hyperphosphorylation, synaptic loss, and progressive cognitive impairment. Microvascular dysfunction is increasingly recognized not only as a consequence but also as a driver of AD pathology [27]. BBB compromise, characterized by tight junction disruption, pericyte loss, and vessel fragmentation, often arises years before the onset of clinical symptoms [28]. Vascular and metabolic dysfunction impair perivascular clearance of Aβ, exacerbating its deposition and accelerating neurodegeneration [29]. Impaired clearance pathways and ECM stiffening associated with amyloid accumulation adversely affect the transport and diffusion of therapeutics, particularly biologicals and nanoparticles. In response, advanced delivery platforms, such as receptor-mediated transcytosis systems, transferrin-targeted nanocarriers, and focused ultrasound-assisted BBB opening, have been developed to enhance drug penetration [14]. However, disease heterogeneity, particularly with regional variations in vascular and glymphatic integrity, underscores the need for precision delivery strategies tailored to pathological profiles of the patients.
3.1.2. Multiple Sclerosis
Multiple sclerosis (MS) is a chronic autoimmune demyelinating condition characterized by neuroinflammation and neurodegeneration. Disruption of BBB integrity is an early and critical pathological event that allows larger solutes, as well as autoreactive T-cells and monocytes to infiltrate the CNS and propagate demyelinating damage [30]. Neuroinflammation in MS amplifies glial activation, upregulates matrix metalloproteinases (MMPs), and disrupts tight junction proteins, impairing tissue repair and exacerbating the axonal injury. Drug delivery in MS is challenged by the coexistence of inflamed, permeable BBB regions and intact barriers, requiring strategies that can selectively target pathological zones. Nanoparticle carriers functionalized to bind inflammatory endothelial markers, anti-CD49d-modified microparticles, and intrathecal administration bypassing systemic circulation are being investigated for the effective delivery of immunomodulatory molecules to the CNS [31,32]. However, beyond immunomodulation, significant challenges remain in achieving durable neuroprotection and remyelination, including limited targeting of oligodendrocyte precursor cells, an inhospitable inflammatory lesion microenvironment, and poor integration with endogenous repair pathways [33,34]. In addition, intrathecal delivery introduces distinct barriers related to compartmentalized neuroinflammation, altered CSF dynamics, and nonuniform drug distribution across focal and diffuse lesions [35]. Emerging theranostic strategies aim to address these limitations by integrating imaging-guided delivery with lesion-specific therapeutic interventions.
3.1.3. Parkinson’s Disease
Parkinson’s disease (PD), historically regarded as a dopaminergic disorder of the substantia nigra, is now recognized to involve considerable vascular pathologies. Early stages of PD exhibit BBB disruption, endothelial dysfunction, and altered angiogenesis, contributing to neuronal vulnerability. Reduced expression of tight junction proteins and transformation of PVS morphology foster neuroinflammation and diminish neuroprotection [36]. Aggregated alphasynuclein, a PD biomarker, accumulates in the perivascular regions, further impairing glymphatic clearance pathways [37]. Therapeutic delivery to PD-affected brain regions is hindered by BBB disruption and inflammation and microvascular stress. Focused ultrasound (FUS) combined with microbubbles has been used experimentally to transiently open the BBB, facilitating the targeted delivery of neuroprotective agents, including GDNF and anti-alpha-synuclein antibodies [38]. Meanwhile, biomaterial-based approaches — such as injectable hydrogels and cell-laden scaffolds — offer local and sustained delivery options aimed at enhancing substantia nigra resilience and neuroregeneration.
3.2. Stroke
Stroke is a complex cerebrovascular disorder encompassing both ischemic and hemorrhagic etiologies, with a lifetime risk of approximately one in four individuals worldwide [39]. Each year, an estimated 14 million people experience stroke globally, of which approximately 70% are ischemic and approximately 30% hemorrhagic in origin [40]. The acute and chronic phases of stroke are marked by distinct pathophysiological changes that significantly affect drug delivery strategies both systemically and locally. These include dynamic alterations of BBB integrity, neuroinflammation involving multiple pathways and cell types, ECM remodeling, oxidative stress, and changes in cell–cell communication (Figure 3). Acute ischemic stroke occurs due to the occlusion of cerebral vessels responsible for regional perfusion, leading to neuronal hypoxia, excitotoxicity, and potentially irreversible damage within minutes [41]. The current first-line pharmacological therapy involves the intravenous administration of recombinant tissue plasminogen activator (tPA), which catalyzes plasmin-mediated clot dissolution [42]. While tPA has demonstrated functional benefits in several randomized controlled trials [EAST, NINDS, and ECASS III], its use is limited by a narrow therapeutic time window (<4.5 hours post-onset) and an increased risk of hemorrhagic transformation, especially at the standard systemic dose of 0.9 mg/kg [43,44].
To overcome these limitations, biomaterial-based drug delivery systems are being developed to enhance tPA specificity and modulate its mechanical release. Examples include targeted nanocarriers [45], fibrin-binding polymers [46], ultrasound-triggerable microbubbles, and matrix-anchored drug depots. These approaches aim to extend the therapeutic window while reducing systemic exposure and the associated hemorrhagic risks. Endovascular thrombectomy has revolutionized ischemic stroke management, offering significant improvements in functional outcomes for patients with large vessel occlusions. Using intra-arterial catheters, mechanical devices can retrieve obstructing thrombi up to 24 h post-stroke in selected patients with favorable imaging profiles (e.g., perfusion–diffusion mismatch) [47,48]. Moreover, these interventions enable the locoregional delivery of neuroprotective agents, anticoagulants, and anti-inflammatory molecules directly at the site of injury, bypassing systemic circulation and minimizing off-target effects. Combining thrombectomy with localized drug delivery systems, such as intra-arterial microspheres, polymeric nanocarriers, or responsive hydrogels, is being explored to modulate reperfusion injury, curb inflammatory cascades, and augment neuronal survival following recanalization.
Following the acute period, stroke enters the subacute and chronic phases, marked by complex neuroinflammation, glial activation, angiogenesis, and ECM remodeling. These processes often culminate in a hostile micromilieu characterized by glial scar formation, hindering axonal regeneration and plasticity. Chondroitin sulfate proteoglycans (CSPGs), major components of the glial scar, act as physical and biochemical inhibitors of neural repair. The localized delivery of chondroitinase ABC, an enzyme that degrades CSPGs, has shown potential in promoting neuroplasticity and functional recovery in preclinical models [49,50]. Matrix metalloproteinases (MMPs) are another class of ECM-modifying enzymes relevant to stroke therapy. While their premature activation may exacerbate secondary injury in the acute phase, controlled MMP activity (>1 week post-stroke) can support remodeling, angiogenesis, and repair [51]. Drug delivery systems based on hydrogels [49], biodegradable microspheres [52], or nanoparticle formulations [53] have been engineered for the timed and localized release of MMPs or MMP inhibitors based on specific physiological cues. Numerous agents targeting neuroinflammation, oxidative stress, and immune modulation are being evaluated for stroke therapy. Candidates such as minocycline, natalizumab, fingolimod, and uric acid have shown promise in modulating microglial activation, leukocyte infiltration, and oxidative damage [54,55]. Additionally, the delivery of growth factors (e.g., BDNF, VEGF), stem cell-supportive cues, or gene-regulating microRNAs is being explored using depot materials or injectable scaffolds to support neural recovery and stem cell integration [56]. Innovative delivery strategies based on physiology-responsive materials, such as pH-sensitive polymers or enzyme-responsive carriers, and minimally invasive methods, such as intranasal or intra-arterial administration, have shown the capacity to bypass compromised vasculature and target specific brain regions affected by stroke [57,58,59].
3.3. Brain Tumors
Gliomas are the most common type of primary brain tumor, with glioblastoma multiforme (GBM) being the most aggressive and prevalent subtype. GBM is characterized by rapid infiltrative growth of tumor cells, breakdown of BBB, profound genomic instability, and elevated interstitial pressure leading to resistance against conventional treatment (Figure 4). Despite multimodal clinical care, including surgical resection, temozolomide chemotherapy, and radiotherapy, the median survival for GBM remains approximately 12–15 months [60]. Beyond primary brain malignancies, approximately 9–17% of all cancer patients develop brain metastases, frequently originating from breast, lung, and melanoma cancers [61]. Similar to GBM, metastatic brain tumors face anatomical and physiological barriers that complicate effective drug delivery.
The ECM in gliomas undergoes extensive remodeling, distinguishing it from normal brain parenchyma. Glioma cells upregulate the deposition of ECM components, such as fibronectin, tenascin-C, and hyaluronan, as well as matrix-bound growth factors not found in healthy brain tissue [62]. These components create a stiffened tumor microenvironment that not only supports tumor proliferation and migration but also hinders the penetration of drugs. Proteolytic enzymes, including matrix metalloproteinases (MMPs), urokinase plasminogen activator (uPA), and cathepsins, are overexpressed by glioma cells to degrade ECM barriers and facilitate invasion into surrounding tissues [63]. This knowledge has inspired the design of protease-responsive drug delivery systems, in which nanocarriers or hydrogels are engineered to release therapeutics in response to enzyme activity within the tumor microenvironment [64].
Gliomas profoundly reshape the immune landscape within the CNS. Tumor-derived signals recruit tumor-associated macrophages (TAMs), myeloid-derived suppressor cells (MDSCs), and immune-inhibitory T cells, promoting an immunosuppressive niche. These cells not only facilitate tumor growth but also offer opportunities for cell-mediated drug delivery, where circulating immune cells are loaded with therapeutics or engineered ex vivo to penetrate the brain and target tumors [65]. Moreover, immunotherapies such as CAR-T cells, dendritic cell vaccines, and immune checkpoint blockade are being investigated for brain tumors and present further tumor-selective drug delivery options, despite the challenges posed by the immunosuppressive microenvironment [66].
A key challenge in treating brain tumors is the restrictive nature of the BBB, which acts as a selective interface to maintain cerebral homeostasis. However, in malignant gliomas, the BBB undergoes pathological modifications, creating what is referred to as the blood–tumor barrier (BTB) [67]. The BTB exhibits heterogeneous permeability, with regions of increased vessel leakiness interspersed with areas retaining an intact or partially intact BBB. This abnormal vasculature arises from tumor-induced angiogenesis driven by proangiogenic factors, such as vascular endothelial growth factor (VEGF) [68]. While increased permeability can facilitate drug access to certain tumor regions, it is insufficient for uniform drug distribution throughout the highly infiltrative tumor margins, where BBB function is often preserved [69]. These perfusion gradients lead to differential drug exposure, contributing to therapy resistance and tumor recurrence.
Despite BTB leakiness, systemic drug delivery remains inefficient because of heterogeneous distribution, drug efflux pumps (e.g., ABC transporters), and elevated intracranial pressure. Locoregional strategies, including convection-enhanced delivery (CED), ultrasound-mediated BBB opening, and implantable polymer wafers (e.g., Gliadel), have been developed to bypass these vascular barriers and deliver high local drug concentrations directly into or near the tumor [70,71]. Focused ultrasound (FUS) with microbubbles has emerged as a promising tool for transiently opening the BBB or BTB at precise anatomical sites, enhancing drug penetration without permanent barrier damage [72]. Biodegradable hydrogels and nanoparticles are being further refined for use in postoperative resection cavities or as systemic carriers that exploit tumor-specific cues.
4. Approaches for CNS Drug Delivery
Systemic administration to the CNS remains challenging due to the restrictive properties of the BBB. Recent advancements in nanotechnology have enhanced systemic delivery by incorporating small molecules, peptides, and nucleic acids into biocompatible lipid nanoparticles (LNPs), polymeric nanoparticles (PLGA, PEG-PCL), and liposomes, which protect the drugs from degradation and extend circulation time [55]. Hydrophobic small molecules are particularly well-suited to liposomal or lipid-based carriers due to strong bilayer interactions, whereas nucleic acids (siRNA, mRNA, plasmids) benefit from ionizable lipids or polymeric carriers capable of endosomal escape [73]. Exosomes have emerged as promising natural carriers for proteins and RNA, offering superior biocompatibility, although they are limited by low loading efficiency and scalability [74]. While these carriers enhance drug stability and circulation, their systemic use can result in off-target distribution, rapid clearance by the mononuclear phagocyte system, and variability in BBB penetration. Local delivery strategies, including CED, ultrasound-mediated BBB opening, and Gliadel wafers, have been developed to bypass these systemic barriers [71]. Below we provide a schematic diagram (Figure 5) and a brief comparison of major approaches for CNS drug delivery (Table 1).
4.1. Intravenous Drug Delivery
Intravenous (IV) administration remains the predominant method for CNS drug delivery; however, its efficacy is inherently constrained by the restrictive nature of the BBB. While small lipophilic compounds may traverse passively, most biologics and nucleic-acid therapies rely on receptor-mediated transcytosis pathways, such as those targeting the transferrin (TfR) receptor [75]. Advanced carriers, including PEGylated immunoliposomes, lipid nanoparticles (LNPs), and polymeric nanoparticles, enhance circulation time, stability, and ligand-specific uptake; nonetheless, their overall brain penetration remains limited [76,77]. Engineered exosomes have emerged as promising natural carriers for proteins and RNA, offering superior biocompatibility, although they are limited by low loading efficiency and scalability [78]. To augment IV delivery, pharmacological BBB modulation has been investigated using hyperosmotic agents (e.g., mannitol), bradykinin analogs, and histamine derivatives to transiently widen tight junctions and increase paracellular permeability. This controlled opening facilitates the passage of large molecules, including antibodies, enzymes, and nanoparticle-encapsulated therapeutics, and can be synergistic with nanoformulations whose size and surface properties are optimized to exploit these transient windows of permeability [79,80]. However, both IV delivery and pharmacological BBB disruption encounter limitations, including systemic toxicity, heterogeneous permeability, and risks of cerebral edema or neuroinflammation, underscoring the necessity for more targeted, reversible, and disease-aware strategies for CNS drug delivery.
4.2. Intra-Arterial Drug Delivery
Intra-arterial (IA) administration delivers therapeutics directly into brain-supplying arteries, achieving higher local concentrations while limiting systemic exposure [81]. This approach is suitable for chemotherapeutics, neuroprotective small molecules, nanoparticles, and stem cells in stroke and brain tumors. Polymeric nanoparticles (e.g., PLGA, PEG-NPs) and liposomes perform well due to controlled release and vascular interactions [82]. Pharmaceutical BBB disruption is again used to transiently open tight junctions, enabling enanced IA delivery of antibodies and nanoparticle-based drugs. IA delivery of resveratrol-loaded nanoparticles enhances neuroprotection after ischemia–reperfusion injury [83]. Limitations include the risk of embolism (notably with cells), BBB permeability constraints, and hemodynamic variability.
4.3. Focused Ultrasound
Focused ultrasound (FUS) is an enabling BBB-modulation strategy used in conjunction with systemic (typically IV) drug administration. Along with microbubbles, it produces spatially targeted, reversible BBB opening, facilitating delivery of antibodies, chemotherapeutics, nucleic acids, and stem cells [84]. Liposomal doxorubicin, polymeric nanoparticles, and LNP-siRNA formulations show significantly enhanced uptake with FUS [85]. Microbubble–nanoparticle hybrids offer integrated imaging and delivery functionalities [86]. Challenges include variability in BBB opening, microhemorrhage risk at high intensities, and the need for MRI guidance for targeted delivery. However, commercial products are available for image (MRI) - guided FUS BBB disruption [87,88].
4.4. Transnasal (Intranasal) Delivery
Intranasal delivery exploits the olfactory and trigeminal neural pathways to circumvent the BBB, enabling direct access to the CNS and allowing administration of proteins, peptides, gene therapies, and stem cells without systemic exposure [89]. Peptides such as vasoactive intestinal peptide (VIP), brain-derived neurotrophic factor (BDNF), and nucleic acids (siRNA, mRNA) have been successfully delivered via this route due to short transport distances and avoidance of hepatic metabolism [90,91]. Suitable carriers include mucoadhesive polymeric nanoparticles (chitosan, PEG-PCL), lipid nanoparticles, nanoemulsions, and engineered exosomes [92]. Exosomes provide excellent biological compatibility and neuronal tropism, although manufacturing challenges persist [93]. The limitations of intranasal delivery include small dosing volumes, mucociliary clearance, and potential epithelial toxicity at high nanoparticle concentrations.
4.5. Intrathecal and Intraventricular Delivery
The intrathecal (IT) and intraventricular (IVT) routes bypass the BBB by administering therapeutics directly into cerebrospinal fluid (CSF), ideal for macromolecules such as enzymes, nucleic acids, peptides, and chemotherapeutics [31]. Hydrogels, liposomes, and polymeric nanoparticles serve as effective carriers, enabling sustained release and reduced dosing frequency [94]. Exosomes show promise for gene and protein delivery with minimal immunogenicity, though CSF flow limits distribution [95]. Risks include infection, catheter malfunction, and uneven drug distribution.
4.6. Intracerebral Injection
Intracerebral injection facilitates the highly localized delivery of adeno-associated virus (AAV) vectors, CRISPR systems, neurotrophic factors, and oncolytic viruses, with viral vectors remaining the predominant modality due to their high transduction efficiency, despite concerns regarding immunogenicity [96,97]. Hydrogels, dendrimers, and other biomaterial carriers further support the local deposition of proteins, enzymes, engineered cells, and nucleic acid therapies, although this method is inherently invasive and offers limited tissue coverage beyond the injection site. CED builds on the same principle of direct parenchymal access but utilizes pressure-driven bulk flow to achieve a significantly broader distribution of chemotherapeutics, proteins, nucleic acids, and viral or cellular therapeutics [98]. Although traditionally performed through intracerebral cannulation, similar pressure-assisted infusion principles have also been applied in intratumoral, intraventricular, and intrathecal contexts; nevertheless, the most robust experimental and clinical evidence for CED remains anchored in parenchymal delivery. Nanoformulations, such as liposomes and polymeric nanoparticles, are particularly well-suited for CED due to their size control and structural stability, whereas hydrogels and exosomes enable sustained release and efficient movement through extracellular matrix microchannels [99]. Despite these advantages, both intracerebral injection and CED encounter procedural challenges, including catheter backflow, mechanical tissue disruption, and heterogeneous tissue resistance, necessitating careful optimization of infusion parameters and delivery geometry.
4.7. Polymeric Wafer Implants for Local Drug Delivery
Gliadel wafers represent one of the earliest clinically implemented and FDA-approved strategies for localized chemotherapeutic delivery to brain tumors, specifically engineered to overcome the significant transport limitations imposed by the BBB [100]. These biodegradable polyanhydride implants, containing the alkylating agent carmustine (BCNU), are strategically placed directly into the surgical resection cavity during glioma surgery. As the polymer matrix undergoes hydrolytic degradation, it releases high concentrations of BCNU into the surrounding parenchyma, facilitating penetration into infiltrative tumor margins while minimizing systemic exposure and toxicity. This localized approach entirely bypasses vascular barriers and provides sustained drug release over a period of days to weeks, thereby offering therapeutic access to regions that are poorly perfused or inaccessible through intravascular routes [71,101]. In contrast, systemically administered chemotherapeutics encounter substantial challenges in reaching brain tumors. The intact BBB at infiltrative margins restricts passive drug entry, while the abnormal, heterogeneous vasculature within tumor cores results in inconsistent perfusion, elevated interstitial fluid pressure, and rapid drug efflux. Diffusion through a dense extracellular matrix, metabolic degradation, and poor transcytosis further limit effective intratumoral concentrations. Collectively, these features create a highly restrictive microenvironment for drug transport, underscoring the necessity for localized delivery platforms such as Gliadel wafers or convection-enhanced delivery in contemporary neuro-oncology. Gliadel wafers are limited by their shallow drug penetration (only 1–3 mm), which prevents BCNU from reaching infiltrative tumor cells that extend beyond the resection margin. They can only be implanted during surgery, carry risks of local toxicity and edema, and provide fixed, non-adjustable dosing. As a result, their clinical benefit is modest and insufficient for highly invasive gliomas like GBM.

Table 1.
Comparison of major CNS drug-delivery approaches, including carriers, disease applications, strengths, limitations, and references.
Table 1.
Comparison of major CNS drug-delivery approaches, including carriers, disease applications, strengths, limitations, and references.
| Approach | Suitable Drug Carriers | Disease Targets | Strengths | Limitations | Key References |
|---|---|---|---|---|---|
| Intravenous (IV) Delivery |
|
|
|
|
[76,77,80] |
| Intra-arterial (IA) Delivery |
|
|
|
|
[81,83] |
| Focused Ultrasound (FUS) |
|
|
|
|
[84,85,86] |
| Intranasal Delivery |
|
|
|
|
[90,91,92] |
| Intrathecal / Intraventricular Delivery |
|
|
|
|
[31,94] |
| Intracerebral Injection |
|
|
|
|
[97,98,99] |
| Polymeric Wafer Implants (Gliadel) |
|
|
|
|
[71,100,101] |
5. Modeling Approaches
The diversity of delivery routes, including systemic intravenous administration across the BBB, intrathecal delivery into CSF spaces, intranasal nose-to-brain pathways, and FUS-mediated BBB opening, necessitates computational frameworks capable of capturing processes across the molecular, cellular, tissue, and organ scales. Computational modeling efforts have historically developed along distinct methodological lines, including pharmacokinetic/pharmacodynamic (PK/PD) models, molecular dynamics (MD) simulations, hybrid and multiscale frameworks, and continuum biophysical models based on computational fluid dynamics (CFD). Recently, machine learning (ML) and data-driven approaches have become prominent, complementing mechanistic models and enabling predictions from high-dimensional datasets. This review organizes CNS drug delivery modeling into five major paradigms and examines their core assumptions, mathematical structures, applications to each delivery route, and persistent challenges (Table 2). The goal is to provide a unified picture of the multi-scale modeling landscape and outline integrative paths forward for next-generation CNS drug-delivery design.
5.1. Physiologically Based Pharmacokinetic (PBPK) Models
PBPK and related pharmacokinetic/pharmacodynamic (PK/PD) models conceptualize organisms as networks of interconnected compartments, which correspond to the plasma, peripheral organs, subregions of the brain, and, in certain instances, explicit CSF spaces. Within each compartment, drug concentrations are assumed to be spatially homogeneous, with intercompartmental transfer driven by blood flow, membrane permeability, or empirically estimated rate constants. In the context of CNS drug delivery, PBPK models typically differentiate between blood, brain tissue, and CSF, aiming to mechanistically separate BBB permeation from tissue binding, metabolism, and clearance.
One of the earliest mechanistic PBPK frameworks for CNS delivery was developed by Ball et al., who modeled BBB exchange in rats by decomposing brain uptake into permeability-driven transport and reversible binding within brain tissue [102]. This framework was subsequently extended and generalized to preclinicsl CNS drug development by the authors [103]. Subsequent work by Yamamoto et al. formalized CNS PBPK structures capable of predicting concentration-time profiles across multiple brain and CSF compartments [5]. More recent works have highlighted PBPK as a model-informed drug development (MIDD) tool for CNS therapeutics, summarizing its strengths and limitations in capturing BBB transport, CSF turnover, and species-specific differences [104,105].
Mathematically, PBPK models consist of coupled ordinary differential equations that enforce mass balance in each compartment. For a two-compartment blood–brain model, the governing equations can be written as
where C, Q, and V are the drug concentration, flow rate, and volume in each compartment, respectively. is the arterial input concentration of drug. is the permeability of the BBB accounting for the passive, is the BBB surface area, and denotes systemic and brain elimination rate constants.
In many CNS applications, Equations (1)–() are augmented with a permeation flux J that models an additional exchange pathway. For the case of a saturable receptor-mediated transcytosis (RMT), the flux takes the form
where and are the maximum transport capacity of the RMT pathway and the Michaelis–Menten constant, respectively. Equation (3) is first converted into volumetric concentration rate through multiplication by , and then the resulting term is incorporated as a loss in the blood compartment (Equation (1)) and a corresponding gain in the brain compartment (Equation ()). Sato et al. recently leveraged such a formulation in a translational PBPK model of transferrin receptor (TfR)-mediated brain delivery, demonstrating a bell-shaped dependence between ligand affinity () and brain exposure arising from the competition between binding and recycling kinetics [6].
In the context of systemic delivery across the BBB, PBPK models are typically integrated into comprehensive whole-body frameworks. These frameworks incorporate perfusion-limited or permeability-limited transport terms for various tissues. Brain compartments may be further divided into vascular, interstitial, and intracellular spaces, or into anatomically distinct regions such as the cortex, CSF, and tumor tissue, each characterized by unique permeabilities, blood flows, or binding capacities [106,106]. These models have been applied to both small molecules and biologics, with particular emphasis on interspecies differences in transporter systems and receptor expression levels.
Intrathecal and intra-CSF delivery are incorporated into PBPK structures by introducing dedicated CSF compartments (e.g., lumbar, cisternal, and ventricular spaces) and specifying the exchange rate constants that describe CSF turnover and CSF-to-brain transfer. In these models, complex CSF hydrodynamics, including pulsatility and mixing, are typically simplified into effective clearance or transfer coefficients. Monine et al. described such CNS-expanded PBPK frameworks and highlighted their utility in predicting CSF drug exposure and brain penetration following intrathecal administration [107]. Other translational studies have integrated CSF pharmacokinetic (PK) profiles with intracerebral microdialysis or imaging to infer effective tissue permeabilities [108].
For intranasal nose-to-brain delivery, PK models generally introduce a nasal dosing compartment with two parallel uptake routes: (i) systemic absorption followed by BBB transport and (ii) direct transport via olfactory and trigeminal pathways. Drath et al. summarized preclinical evidence supporting these parallel pathways and described PK analyses that attempt to deconvolve direct brain uptake from indirect systemic contributions [109]. Rygg and colleagues have proposed combining such PK structures with computational fluid dynamics (CFD)-derived regional deposition estimates to evaluate bioequivalence for nose-to-brain drug products [110].
Focused ultrasound (FUS)-mediated BBB opening is naturally incorporated into PBPK models by allowing the BBB permeability to become a time-dependent function that increases transiently following sonication. Burgess et al. reviewed the underlying mechanism through which microbubble-assisted FUS disrupts the BBB, providing an experimental basis for such PBPK extensions [111]. Subsequent preclinical and clinical studies have quantified the magnitude and duration of BBB opening with neuronavigation-guided FUS, enabling the incorporation of temporally resolved permeability profiles into PBPK models [112].
Overall, PBPK models are adept at capturing systemic disposition, inter-organ exchange, and intersubject variability at a relatively low computational cost. They are highly valuable for dose selection, sensitivity analyses, and species scaling. However, their assumptions of well-mixed compartments and the lack of explicit anatomical geometry limit their ability to capture spatially heterogeneous processes that are critical in intrathecal, intranasal, and FUS-modulated delivery. Parameter uncertainty remains a key challenge, particularly in BBB permeability, CSF-to-brain exchange rates, and nanoparticle-specific transport kinetics.
5.2. Continuum Models
Continuum models represent one of the most advanced classes of computational methodologies for CNS drug delivery, particularly for pathways where fluid dynamics and anatomical geometry significantly influence solute distribution. Within these frameworks, blood or CSF is modeled as an incompressible continuum, the brain parenchyma as a porous medium, and therapeutics as solutes transported through advection, diffusion, and reaction processes. The governing equations comprise the incompressible Navier–Stokes equations for CSF or blood flow, coupled with advection–diffusion–reaction equations for solute transport, and in porous tissues, Darcy or Darcy–Brinkman constitutive relations.
For an incompressible fluid, the Navier–Stokes equations take the form
where is the velocity field, p is the pressure, is the density, and is the dynamic viscosity. Solute transport in the same domain is governed by
where C represents the solute concentration, refers to the effective diffusivity that accounts for extracellular tortuosity, signifies an effective clearance rate arising from cellular uptake, efflux across barriers, and elimination through CSF ouflow. The term captures nonlinear binding processes, including reversible binding to ECM, receptor–ligand interactions, or enzyme-mediated degradation. S is a source term representing injection of drug, controlled release from an implant, or drug uptake from systemic blood circulation. For the brain tissue, a porous medium formulation was employed:
where K indicates the hydraulic permeability of the extracellular matrix.
Continuum models and partial differential equations (PDE) were foundational to understanding brain microphysiology [113] and have become the prevailing standard for intrathecal and CSF-based drug delivery systems. Continuum modeling has also played a pivotal role in explaining the barrier roles of diffusion, convection, binding and capillary clearance. Additionally, Peclet number explains why diffusion dominates for small molecules and convection for macromolecules, NPs etc. [114,115]. Khani et al. implemented a realistic cervical subarachnoid space geometry to investigate the impact of injection parameters on intrathecal solute transport, utilizing pulsatile CSF flow simulations in conjunction with advection-diffusion transport [3]. They presented a framework through comprehensive parameter sweeps involving catheter placement, injection waveforms, and physiological conditions, thereby introducing the concept of "human in silico trials" for optimizing intrathecal delivery (Figure 6). Their results were corroborated by in vitro flow-phantom experiments, demonstrated that minor alterations in injection location or timing can significantly influence drug dispersion along the spinal canal and cisternal/ventricular CSF spaces. Kouzehgarani et al. reviewed these fluid-dynamic models alongside pharmacokinetic analyses and clinical data, positing that CSF circulation can be harnessed to deliver large biologics into deeper brain structures when infusion protocols are meticulously calibrated [116].
Continuum modeling plays a pivotal role in elucidating glymphatic and perivascular transport mechanisms, with significant implications for both intrathecal and systemic drug delivery. Mestro et al. investigated periarterial solute movement driven by arterial pulsations [117], while Vinje et al. conducted simulations of solute transport within periarterial spaces and the parenchymal extracellular space, examining the interaction between diffusion and convection [10]. They integrated multimodal MRI with forward and inverse modeling to quantify subject-specific CSF tracer transport parameters in humans, highlighting the close relationship between imaging and continuum mechanics. Bohr et al. provided a comprehensive review of glymphatic models utilizing advection–diffusion–reaction equations in MRI-informed geometries [118]. Quirk et al. developed a brain-wide model incorporating explicit pial perivascular spaces, demonstrating that CSF-driven solute penetration is highly sensitive to the size and connectivity of PVS [119]. Nose-to-brain delivery has also been extensively studied using CFD and particle-tracking simulations. Models constructed in patient-specific nasal geometries have quantified device-dependent olfactory deposition, identified breathing patterns that optimize upper nasal delivery, and evaluated the impact of surgical modifications on deposition efficiency [11]. Vishnumurthy et al. synthesized these studies in a systematic review and meta-analysis, identifying robust CFD predictors of olfactory deposition and emphasizing parameters crucial for nose-to-brain targeting [120]. Boyuklieva et al. combined computational, in vitro, and in vivo measurements to assess the predictive power of CFD for nose-to-brain exposure, underscoring both its utility and current limitations [121]. In the context of FUS-mediated BBB opening, continuum models typically simulate drug transport downstream of the sonication event, with the FUS action represented as a localized, transient increase in BBB permeability. Burgess and Hynynen’s seminal review outlined how microbubble-assisted FUS enhances BBB permeability for chemotherapy and biologics [111]. Subsequent studies have incorporated spatially varying permeability maps derived from imaging or acoustic models into advection–diffusion simulations to predict regional drug accumulation following FUS [122]. Continuum and CFD-based models have also been employed to simulate drug transport from polymeric wafer implants by solving coupled diffusion–reaction–convection equations within realistic brain-tumor geometries reconstructed from MRI data [123]. These simulations incorporate Darcy’s law to account for interstitial fluid flow, enabling quantitative comparisons of various chemotherapeutic agents. The models predict how tissue heterogeneity, drug degradation kinetics, and physicochemical properties influence local concentration profiles and penetration depths. The modeling demonstrates that wafer-delivered drugs distribute unevenly across the tumor–cavity interface and underscores transport limitations that are not experimentally observable, thereby informing the optimization of local delivery strategies.
Overall, the strengths of continuum models lie in their ability to incorporate anatomical realism, spatial heterogeneity, and complex CSF or vascular-flow patterns. They are particularly effective for addressing route-specific questions, such as optimal intrathecal injection protocols or nasal device design. However, their limitations include the simplified treatment of barrier crossing and cellular uptake via effective permeability coefficients, as well as the high computational cost associated with whole-brain or whole-spine simulations. Predictions can also be sensitive to uncertain tissue parameters, such as permeability, porosity, and effective diffusivity.
5.3. Molecular Dynamics and Atomistic Simulations
Molecular dynamics (MD) and related atomistic methodologies operate at the opposite extreme to whole-body PBBK models, from nanometer and nanosecond to microsecond scales, aiming to elucidate the molecular mechanisms underlying barrier crossing, ligand binding, nanoparticle deformation, and carrier-membrane interactions. Within the context of the CNS, MD has been predominantly applied to investigate small-molecule permeation across lipid bilayers that mimic the BBB, peptide or ligand interactions with receptor targets involved in receptor-mediated transcytosis (RMT), and nanoparticle interactions with endothelial membranes [7,55].
In classical MD, atomic or coarse-grained trajectories are derived by integrating Newton’s equations of motion under a specified force field. The total potential energy is typically decomposed into bonded contributions, including bond stretching, angle bending, and dihedral rotations, as well as non-bonded interactions, comprising Lennard-Jones van der Waals potentials and Coulombic electrostatics. Coarse-grained models, such as Martini force fields, aggregate several atoms into beads with effective interaction potentials, facilitating simulations over larger spatial and temporal scales at a reduced computational cost. MD outputs encompass structural observables, diffusion coefficients, permeation-free energies, residence times, and metrics of lipid or nanoparticle deformation.
Numerous studies have employed MD simulations to explore BBB permeation at the molecular scale. Pedram et al. utilized all-atom and coarse-grained MD to simulate magnetic nanoparticle translocation across model endothelial bilayers under externally applied magnetic fields, quantifying the impact of nanoparticle size, hydrophobicity, and surface charge on the likelihood of membrane penetration [124]. Ivanczi et al. performed MD simulations of several cell-penetrating peptides (CPPs) and their drug-conjugates interacting with explicit lipid-bilayer models (Figure 7). They found that Linear CPPs show major conformational changes at membranes while cyclic CPPs stay structurally stable, demonstrating that peptide architecture strongly influences membrane penetration [8]. Ouyang et al. provided a review of MD simulations of BBB-crossing and tumor-homing peptides, illustrating how free-energy calculations, umbrella sampling, and conformational ensemble analysis can identify peptide motifs with high affinity and favorable insertion profiles for CNS drug delivery [7].
More broadly, MD studies have been integrated into the larger field of nanomedicine for brain delivery. Reviews by Saraiva et al. and others emphasize how MD, in conjunction with imaging and in vitro data, elucidates the influence of nanoparticle size, shape, ligand density, and surface charge on membrane wrapping, endocytosis, and intracellular trafficking [55]. These studies underscore the role of atomistic simulations in guiding the rational design of brain-targeting nanocarriers.
Although less common, MD and mesoscale simulations have been employed to investigate the biophysical effects of ultrasound-induced cavitation on biological membranes. These models represent lipid bilayers and either explicit or implicit gas bubble interfaces, applying oscillatory perturbations representative of focused ultrasound exposure. The resulting membrane thinning, pore nucleation, and transient defect formation have been characterized in several computational studies, providing mechanistic insights into FUS-mediated increases in BBB permeability [125].
MD has also been applied to intranasal and mucosal delivery. For instance, Gao et al. utilized MD simulations to compare mucoadhesive versus mucus-penetrating nanoparticle designs by simulating all-atom interactions between various NPs and the main nasal mucin. They found that PEG-PLGA-based particles exhibited minimal interaction (favoring penetration), whereas chitosan-derived particles showed strong mucin binding (mucoadhesion) [126]. Similar principles apply to nanoparticle behavior in CSF-like environments, although MD studies explicitly modeling the ionic composition and protein content of CSF remain sparse.
The principal advantage of MD lies in its high mechanistic fidelity, which effectively captures molecular recognition, conformational dynamics, and nanoscale transport events that are beyond the reach of continuum-scale or pharmacokinetic/pharmacodynamic (PK/PD) models. Nevertheless, MD is limited by constraints related to time and length scales. Even with the use of coarse-grained models and enhanced sampling techniques, the simulation of complete transcytosis pathways, long-distance diffusion across endothelial thickness, or vesicular trafficking remains impractical. Additionally, the actual geometries of the BBB and epithelial tissues are significantly more complex than the planar bilayers typically employed in MD simulations. The parameterization of force fields for novel nanoparticles continues to be a challenging task. To impact organ-level predictions, properties derived from MD, such as permeability, free energy, and diffusion coefficients, must be meticulously upscaled and integrated into hybrid or PBPK models.
5.4. Hybrid and Multiscale Models
Hybrid and multiscale models endeavor to integrate microscopic mechanisms with macroscopic transport behavior by incorporating molecular or cellular kinetics into tissue- and organ-scale transport frameworks. This methodology is particularly advantageous for CNS drug delivery, where the translocation of a therapeutic agent across the BBB or nasal mucosa is dictated by molecular-level recognition and membrane interactions. The binding kinetics and transcytosis of these therapeutic carriers (NPs and exosomes) are stochastic in nature and thus require molecular scale models. In contrast, subsequent dispersion through CSF, perivascular pathways, or brain parenchyma relies on continuum-scale flow and diffusion phenomena. In this context, multiscale models provide a comprehensive mathematical description that synthesizes the physics of molecular binding, cellular trafficking, fluid mechanics, and tissue-level heterogeneity [9,127].
Conceptually, multiscale CNS drug-delivery frameworks consist of at least two hierarchical tiers. At the microscopic level, molecular dynamics, Monte Carlo, agent-based simulations, or stochastic models can be utilized to quantify binding affinities, receptor-mediated transcytosis, vesicular trafficking, nanoparticle endocytosis, and intracellular transit. Such simulations reveal how particle size, surface charge, PEGylation, and hydrophobicity modulate biological interactions relevant to systemic circulation, mucus penetration, and transcytosis [126,128]. These molecular-level parameters, in turn, inform mesoscale or continuum transport models through effective permeabilities, binding rate constants, and release kinetics derived from bottom-up simulation. The outputs of these simulations and stochastic models, including binding and unbinding rates, internalization rate constants, receptor recycling times, and binding probabilities, are subsequently coarse-grained into effective kinetic parameters. In [4,129], the total solution of the PDEs is split into coarse (resolved) and fine (subgrid) scales. The fine scales account for microscopic adhesion kinetics governed by a stochastic particle adhesion model. These fine scales are variationally projected into coarse scales to yield a stabilized and computationally robust formulation (Figure 8). At the macroscopic scale, drug or nanoparticle concentration fields are represented by advection-diffusion-reaction equations, whose reaction terms incorporate these coarse-grained kinetic processes. A representative tissue-level transport equation takes the form
In this context, the uptake coefficient encapsulates the influence of receptor density, cellular uptake kinetics, and microvascular surface area. The negative sign indicates drug consumption in the cellular uptake process. In perfused tissues, these equations are integrated with vascular-side flow and transport models, alongside interface conditions that delineate solute exchange across vascular or perivascular boundaries.
Hybrid particle–continuum models integrate discrete dynamics at the nanoparticle or cell scale with continuum fluid flow and reaction–diffusion transport (Figure 9). For instance, Akalin et al. developed a hybrid scheme that couples continuum hemodynamics with discrete nanoparticle trajectories to predict vascular margination, wall adhesion, and transmural transport [130]. In the field of oncology, multiscale tumor models integrate evolving microvascular networks, interstitial flow, anti-angiogenic responses, and convection–diffusion–reaction drug transport to capture heterogeneous intratumoral exposure [131]. These models facilitate the simultaneous representation of vascular remodeling, transport barriers, and drug–tissue interactions, which cannot be resolved by single-scale methods.
Recently, multiscale modeling has been extended to nanoparticle-mediated intravascular and brain-targeted drug delivery, where molecular-scale binding, vascular flow physics, and tissue perfusion must be concurrently represented. A 2024 physics-based in silico framework integrated nanoparticle design, blood flow, and tissue-level transport to evaluate systemic nanocarrier delivery [132]. Similarly, the combination of advanced imaging with computational modeling has enabled multiscale prediction of nanoparticle accumulation in the brain [133]. Emerging approaches also incorporate machine learning as a bridge across scales, utilizing data-driven models trained on CFD, MD, or PK simulations to accelerate parameter exploration in complex delivery systems [12].
The primary advantage of hybrid and multiscale modeling lies in its capacity to represent mechanistic insights from molecular and cellular scales while still predicting organ-scale drug distribution, regional accumulation, or glymphatic clearance patterns. However, these models also entail substantial complexity, often involving numerous parameters that are challenging to measure. Model validation across scales remains difficult, and workflow standardization is limited in this field. Despite these challenges, the increasing availability of cross-scale experimental data—from microfluidic BBB platforms, high-field MRI, single-cell transcriptomics, and in vivo imaging—is anticipated to render hybrid and multiscale modeling approaches integral to future CNS drug-delivery research.
5.5. Machine-Learning and Data-Driven Models
Machine learning (ML) methodologies offer a complementary approach to mechanistic modeling by leveraging empirical patterns in extensive and often high-dimensional datasets. These techniques have proven valuable across various CNS drug-delivery challenges, including predicting BBB permeability, nanoparticle brain uptake, extracting imaging-derived biomarkers, and constructing surrogate models that approximate computationally intensive CFD simulations.
Initial ML research concentrated on small-molecule BBB permeability, where models trained on extensive chemical libraries demonstrated improvements over traditional QSAR-based methods. Wang et al. demonstrated that support vector machines, random forests, and ensemble resampling strategies significantly enhance predictive accuracy by capturing nonlinear interactions among molecular descriptors and addressing the inherent class imbalance between BBB-permeable and impermeable compounds [17]. Deep learning models, such as graph neural networks, have also been applied to BBB permeability prediction, showing improved performance for complex chemotypes [13].
Recently, ML has been extended to nanoparticle-mediated brain delivery. Yousfan et al. compiled the largest nanoparticle CNS delivery dataset to date, with 403 nanocarriers characterized for brain uptake, and trained multiple ML models to identify the physicochemical determinants of CNS penetration [2]. Their feature-importance analyses revealed that surface charge, ligand density, and material composition often surpass particle size as predictors of brain accumulation, providing mechanistic hypotheses for subsequent experimental or multi-scale model validation. A comprehensive review by Alves et al. situates these ML methods within the broader modeling ecosystem, emphasizing their synergy with PBPK models, MD simulations, and multi-omics characterization of BBB transport [1].
In intranasal delivery, ML-based surrogate models trained on CFD outputs have been employed to rapidly predict nasal deposition patterns and guide device or formulation optimization [134]. In the intrathecal context, surrogate models have been developed to approximate CSF drug-dispersion outcomes across wide parameter ranges, significantly reducing the computational cost associated with thousands of CFD simulations [3].
ML plays an increasingly significant role in image-based modeling. Brain-wide glymphatic studies have integrated ML techniques to detect tracer propagation patterns, estimate subject-specific diffusion and convection parameters, and initialize continuum-scale solute transport models. Deep learning frameworks have been developed for the segmentation of PVS, ventricles, and CSF flow features, providing anatomical and geometric inputs for downstream continuum simulations [135]. Similar approaches have been explored for estimating glymphatic efficiency metrics from MRI time-series data [136].
Despite their potential, ML methods encounter challenges, including data heterogeneity, limited standardized datasets for nanoparticles and biologics, inconsistent experimental protocols, and difficulties in interpretability. Overfitting is common, and extrapolation beyond the training domains remains uncertain. Therefore, the most promising future direction is hybrid ML–mechanistic modeling, wherein ML is utilized to estimate uncertain parameters, discover latent features, or provide computational surrogates, while physics-based continuum or PBPK models ensure mechanistic interpretability and physically consistent extrapolation.
Table 2.
Comparison of major computational modeling approaches for CNS drug delivery.
| Modeling Approach | Major Assumptions | Strengths | Limitations | References |
|---|---|---|---|---|
| PBPK Models |
|
|
|
[5,6,105,110] |
| Continuum Models (CFD, Advection–Diffusion, Porous Media) |
|
|
|
[3,10,11,116,123] |
| Molecular Dynamics / Atomistic Simulations |
|
|
|
[7,8,125] |
| Hybrid and Multiscale Models |
|
|
|
[4,128,132,133] |
| Machine-Learning / Data-Driven Models |
|
|
|
[12,13,120] |
6. Future Perspective
Mathematical and computational modeling has frequently demonstrated remarkable efficacy in elucidating the structure, mechanisms, and design principles across a wide range of biological systems. Nevertheless, no single model serves as a universal solution, and the central nervous system (CNS), characterized by incomplete parameter knowledge, hierarchical interactions, and emergent collective phenomena, remains one of the most challenging domains for quantitative prediction. Consequently, the future of CNS drug delivery modeling hinges not only on methodological innovation but also on the clarity of assumptions, transparent limitations, and rigorous validation. Exemplary models are founded on explicit hypotheses, make falsifiable predictions, and remain open to refinement as new physiological data become available. Although they may be less realistic than in vivo studies, these models serve as essential complements that facilitate counterfactual experiments, optimization of drug design, and in silico clinical trials, thereby accelerating therapeutic development and reducing costs. To sustain this virtuous cycle, models should clearly differentiate between results and interpretations and delineate their domains of applicability as explicitly as possible.
A significant opportunity lies in the development of next-generation multiscale digital twins of the CNS. Such frameworks will integrate molecular-scale receptor binding and transcytosis, mesoscale endothelial mechanics, intravascular hemodynamics, CSF and perivascular flow, and parenchymal diffusion–reaction processes within a unified computational framework. Patient-specific digital twins incorporating MRI, CT, or optical imaging will enable validation and individualized predictions of drug dispersion for intrathecal biologics, intra-arterial nanoparticles, focused ultrasound–enabled therapies, or intranasal formulations. Realizing this vision will necessitate advances in image registration, parameter estimation, reduced-order modeling, uncertainty quantification, and machine learning–augmented inference.
Equally important is the need to model the dynamic and heterogeneous nature of the BBB and neurovascular unit. Single-cell atlases have revealed substantial spatial variations in endothelial, astrocytic, and pericyte phenotypes, including the zonation of transporters, tight junction proteins, and signaling receptors [137]. Future models must incorporate this heterogeneity, as well as age-related or disease-induced changes in stiffness, inflammation, oxidative stress, and vascular morphology. BBB transport should be considered an actively regulated process rather than a static barrier, responding to shear stress, neurovascular coupling, and metabolic demand.
Translating rodent models to human applications remains a significant challenge. Many existing formulations were developed for lissencephalic rodent brains, which differ from the human CNS in terms of geometry, vascular resistance, CSF turnover, sleep architecture, glymphatic topology, and arteriole–venule ratios. While scaling arguments based on the Peclet number, Womersley number, or dimensionless permeability offer guidance, they cannot fully account for species-specific neuroanatomy or biomechanics. Successful translation will necessitate models that accommodate these anatomical and physiological differences while integrating human microdialysis measurements, CSF sampling, positron emission tomography (PET) tracers, dynamic contrast-enhanced magnetic resonance imaging (DCE-MRI), and multimodal neuroimaging.
A promising parallel approach is the development of hybrid physics–machine learning frameworks. Machine learning can infer uncertain parameters (such as permeability, receptor density, and uptake rate constants) from imaging or omics data, learn surrogate mappings to accelerate CFD or PDE simulations, and classify CNS transport phenotypes. Physics-informed neural networks, graph neural networks for CNS geometry, and diffusion models for generating virtual cohorts will increasingly support large-scale in silico trials in the future. Crucially, hybrid models must incorporate physical constraints, such as mass conservation, boundary conditions, and transport limits, to avoid biologically implausible extrapolations.
Finally, future progress will depend on standardization, reproducibility, and the availability of shared data resources. Variations in geometry fidelity, boundary conditions, and parameter choices remain major obstacles to comparing models across studies. Publicly available reference geometries, curated nanoparticle delivery datasets, validated BBB permeability libraries, and benchmarking problems for intrathecal and perivascular CFD simulations would significantly accelerate research in this field. Model categories, such as those summarized in Table 2, help organize prior work and highlight opportunities for impactful advances; however, their utility depends on continued experimental grounding. Ultimately, each model should be applied where it is most informative, validated experimentally whenever possible, including in vivo, and used in combination with complementary approaches when necessary. Collectively, these directions point toward a future in which CNS drug delivery modeling becomes increasingly integrative, predictive, and clinically actionable. By merging mechanistic biophysics with data-driven inference and patient-specific neuroimaging, next-generation models have the potential to guide therapy design, personalize dosing strategies, and uncover new biological principles governing drug movement in the brain.
Conflicts of Interest
The authors declare no potential conflict of interest.
References
- Alves, P. A.; Camargo, L. C.; Souza, G. M. d.; Mortari, M. R.; Homem-de Mello, M. Computational modeling of pharmaceuticals with an emphasis on crossing the blood–brain barrier. Pharmaceuticals 2025, 18(2), 217. [Google Scholar] [CrossRef]
- Yousfan, A.; Al Rahwanji, M. J.; Hanano, A.; Al-Obaidi, H. A comprehensive study on nanoparticle drug delivery to the brain: application of machine learning techniques. Molecular Pharmaceutics 2023, 21(1), 333–345. [Google Scholar] [CrossRef]
- Khani, M.; Burla, G. K. R.; Sass, L. R.; Arters, O. N.; Xing, T.; Wu, H.; Martin, B. A. Human in silico trials for parametric computational fluid dynamics investigation of cerebrospinal fluid drug delivery: impact of injection location, injection protocol, and physiology. Fluids and Barriers of the CNS 2022, 19(1), 8. [Google Scholar] [CrossRef]
- Goraya, S. A.; Ding, S.; Miller, R. C.; Arif, M. K.; Kong, H.; Masud, A. Modeling of spatiotemporal dynamics of ligand-coated particle flow in targeted drug delivery processes. Proceedings of the National Academy of Sciences 2024, 121(22), e2314533121. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Valitalo, P. A.; Huntjens, D. R.; Proost, J. H.; Vermeulen, A.; Krauwinkel, W.; Beukers, M. W.; van den Berg, D.-J.; Hartman, R.; Wong, Y. C. Predicting drug concentration-time profiles in multiple cns compartments using a comprehensive physiologically-based pharmacokinetic model. CPT: pharmacometrics & systems pharmacology 2017, 6(11), 765–777. [Google Scholar]
- Sato, S.; Liu, S.; Goto, A.; Yoneyama, T.; Okita, K.; Yamamoto, S.; Hirabayashi, H.; Iwasaki, S.; Kusuhara, H. Advanced translational pbpk model for transferrin receptor-mediated drug delivery to the brain. Journal of Controlled Release 2023, 357, 379–393. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, J.; Sheng, Y.; Wang, W. Recent advances of studies on cell-penetrating peptides based on molecular dynamics simulations. Cells 2022, 11(24), 4016. [Google Scholar] [CrossRef] [PubMed]
- Ivanczi, M.; Balogh, B.; Kis, L.; Mandity, I. Molecular dynamics simulations of drug-conjugated cell-penetrating peptides. Pharmaceuticals 2023, 16(9), 1251. [Google Scholar] [CrossRef]
- Clancy, C. E.; An, G.; Cannon, W. R.; Liu, Y.; May, E. E.; Ortoleva, P.; Popel, A. S.; Sluka, J. P.; Su, J.; Vicini, P. Multiscale modeling in the clinic: drug design and development. Annals of biomedical engineering 2016, 44(9), 2591–2610. [Google Scholar] [CrossRef]
- Vinje, V.; Zapf, B.; Ringstad, G.; Eide, P. K.; Rognes, M. E.; Mardal, K.-A. Human brain solute transport quantified by glymphatic mri-informed biophysics during sleep and sleep deprivation. Fluids and Barriers of the CNS 2023, 20(1), 62. [Google Scholar] [CrossRef]
- Sundstrom, E.; Talat, R.; Sedaghat, A. R.; Khosla, S.; Oren, L. Computational modeling of nasal drug delivery using different intranasal corticosteroid sprays for the treatment of eustachian tube dysfunction. Journal of Engineering and Science in Medical Diagnostics and Therapy 2022, 5(3), 031103. [Google Scholar] [CrossRef]
- Debnath, G.; Vasu, B.; Gorla, R. S. R. Current state-of-the-art in multi-scale modeling in nano-cancer drug delivery: role of ai and machine learning. Cancer Nanotechnology 2025, 16(1), 45. [Google Scholar] [CrossRef]
- Pereira, T.; Abbasi, M.; Oliveira, J. L.; Ribeiro, B.; Arrais, J. Optimizing blood–brain barrier permeation through deep reinforcement learning for de novo drug design. Bioinformatics 2021, 37 Supplement_1, i84–i92. [Google Scholar] [CrossRef]
- Wu, D.; Chen, Q.; Chen, X.; Han, F.; Chen, Z.; Wang, Y. The blood–brain barrier: Structure, regulation and drug delivery. Signal transduction and targeted therapy 2023, 8(1), 217. [Google Scholar] [CrossRef]
- Profaci, C. P.; Munji, R. N.; Pulido, R. S.; Daneman, R. The blood–brain barrier in health and disease: Important unanswered questions. Journal of Experimental Medicine 2020, 217(4), e20190062. [Google Scholar] [CrossRef]
- Abbott, N. J.; Rönnbäck, L.; Hansson, E. Astrocyte–endothelial interactions at the blood–brain barrier. Nature reviews neuroscience 2006, 7(1), 41–53. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, H.; Wu, Z.; Wang, T.; Li, W.; Tang, Y.; Liu, G. In silico prediction of blood–brain barrier permeability of compounds by machine learning and resampling methods. ChemMedChem 2018, 13(20), 2189–2201. [Google Scholar] [CrossRef] [PubMed]
- Troili, F.; Cipollini, V.; Moci, M.; Morena, E.; Palotai, M.; Rinaldi, V.; Romano, C.; Ristori, G.; Giubilei, F.; Salvetti, M. Perivascular unit: this must be the place. the anatomical crossroad between the immune, vascular and nervous system. Frontiers in neuroanatomy 2020, 14 17. [Google Scholar]
- Troili, F.; Cipollini, V.; Moci, M.; Morena, E.; Palotai, M.; Rinaldi, V.; Romano, C.; Ristori, G.; Giubilei, F.; Salvetti, M. Corrigendum: Perivascular unit: This must be the place. the anatomical crossroad between the immune, vascular and nervous system. Frontiers in Neuroanatomy 2020, 14 51. [Google Scholar] [CrossRef] [PubMed]
- Montagne, A.; Barnes, S. R.; Sweeney, M. D.; Halliday, M. R.; Sagare, A. P.; Zhao, Z.; Toga, A. W.; Jacobs, R. E.; Liu, C. Y.; Amezcua, L. Blood-brain barrier breakdown in the aging human hippocampus. Neuron 2015, 85(2), 296–302. [Google Scholar] [CrossRef] [PubMed]
- Iturria-Medina, Y.; Sotero, R. C.; Toussaint, P. J.; Mateos-Pérez, J. M.; Evans, A. C. Early role of vascular dysregulation on late-onset alzheimer’s disease based on multifactorial data-driven analysis. Nature communications 2016, 7(1), 11934. [Google Scholar] [CrossRef] [PubMed]
- Winkler, E. A.; Sengillo, J. D.; Bell, R. D.; Wang, J.; Zlokovic, B. V. Blood–spinal cord barrier pericyte reductions contribute to increased capillary permeability. Journal of Cerebral Blood Flow & Metabolism 2012, 32(10), 1841–1852. [Google Scholar] [CrossRef]
- Yamazaki, Y.; Shinohara, M.; Shinohara, M.; Yamazaki, A.; Murray, M. E.; Liesinger, A. M.; Heckman, M. G.; Lesser, E. R.; Parisi, J. E.; Petersen, R. C. Selective loss of cortical endothelial tight junction proteins during alzheimer’s disease progression. Brain 2019, 142(4), 1077–1092. [Google Scholar] [CrossRef]
- Yamada, M.; Hamaguchi, T. The sulfation code for propagation of neurodegeneration. Journal of Biological Chemistry 2018, 293(27), 10841–10842. [Google Scholar] [CrossRef]
- Blanchard, J. W.; Bula, M.; Davila-Velderrain, J.; Akay, L. A.; Zhu, L.; Frank, A.; Victor, M. B.; Bonner, J. M.; Mathys, H.; Lin, Y.-T. Reconstruction of the human blood–brain barrier in vitro reveals a pathogenic mechanism of apoe4 in pericytes. Nature medicine 2020, 26(6), 952–963. [Google Scholar] [CrossRef]
- Kisler, K.; Nelson, A. R.; Rege, S. V.; Ramanathan, A.; Wang, Y.; Ahuja, A.; Lazic, D.; Tsai, P. S.; Zhao, Z.; Zhou, Y. Pericyte degeneration leads to neurovascular uncoupling and limits oxygen supply to brain. Nature neuroscience 2017, 20(3), 406–416. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, M. D.; Sagare, A. P.; Zlokovic, B. V. Blood–brain barrier breakdown in alzheimer disease and other neurodegenerative disorders. Nature Reviews Neurology 2018, 14(3), 133–150. [Google Scholar] [CrossRef]
- Nation, D. A.; Sweeney, M. D.; Montagne, A.; Sagare, A. P.; D’Orazio, L. M.; Pachicano, M.; Sepehrband, F.; Nelson, A. R.; Buennagel, D. P.; Harrington, M. G. Blood–brain barrier breakdown is an early biomarker of human cognitive dysfunction. Nature medicine 2019, 25(2), 270–276. [Google Scholar] [CrossRef]
- Zlokovic, B. V. Neurovascular pathways to neurodegeneration in alzheimer’s disease and other disorders. Nature Reviews Neuroscience 2011, 12(12), 723–738. [Google Scholar] [CrossRef]
- Lassmann, H. Multiple sclerosis pathology. Cold Spring Harbor perspectives in medicine 2018, 8(3), a028936. [Google Scholar] [CrossRef] [PubMed]
- Madadi, A. K.; Sohn, M.-J. Advances in intrathecal nanoparticle delivery: targeting the blood–cerebrospinal fluid barrier for enhanced cns drug delivery. Pharmaceuticals 2024, 17(8), 1070. [Google Scholar] [CrossRef]
- Kapate, N.; Dunne, M.; Kumbhojkar, N.; Prakash, S.; Wang, L. L.-W.; Graveline, A.; Park, K. S.; Chandran Suja, V.; Goyal, J.; Clegg, J. R. A backpack-based myeloid cell therapy for multiple sclerosis. Proceedings of the National Academy of Sciences 2023, 120(17), e2221535120. [Google Scholar] [CrossRef] [PubMed]
- Franklin, R. J.; Ffrench-Constant, C. Regenerating cns myelin—from mechanisms to experimental medicines. Nature Reviews Neuroscience 2017, 18(12), 753–769. [Google Scholar] [CrossRef] [PubMed]
- Stadelmann, C.; Timmler, S.; Barrantes-Freer, A.; Simons, M. Myelin in the central nervous system: structure, function, and pathology. Physiological reviews 2019. [Google Scholar] [CrossRef]
- Magliozzi, R.; Howell, O. W.; Nicholas, R.; Cruciani, C.; Castellaro, M.; Romualdi, C.; Rossi, S.; Pitteri, M.; Benedetti, M. D.; Gajofatto, A. Inflammatory intrathecal profiles and cortical damage in multiple sclerosis. Annals of neurology 2018, 83(4), 739–755. [Google Scholar] [CrossRef]
- Gray, M. T.; Woulfe, J. M. Striatal blood–brain barrier permeability in parkinson’s disease. Journal of Cerebral Blood Flow & Metabolism 2015, 35(5), 747–750. [Google Scholar]
- Elabi, O.; Gaceb, A.; Carlsson, R.; Padel, T.; Soylu-Kucharz, R.; Cortijo, I.; Li, W.; Li, J.-Y.; Paul, G. Human α-synuclein overexpression in a mouse model of parkinson’s disease leads to vascular pathology, blood brain barrier leakage and pericyte activation. Scientific reports 2021, 11(1), 1120. [Google Scholar] [CrossRef]
- Leinenga, G.; Götz, J. Scanning ultrasound removes amyloid-β and restores memory in an alzheimer’s disease mouse model. Science translational medicine 2015, 7(278), 278ra33–278ra33. [Google Scholar] [CrossRef] [PubMed]
- Campbell, B.; Khatri, P. Stroke lancet, 396 (10244)(2020), View PDF View article View in Scopus; 2020; pp. 129–142. [Google Scholar]
- Feigin, V. L.; Abate, M. D.; Abate, Y. H.; Abd ElHafeez, S.; Abd-Allah, F.; Abdelalim, A.; Abdelkader, A.; Abdelmasseh, M.; Abd-Elsalam, S.; Abdi, P. Global, regional, and national burden of stroke and its risk factors, 1990–2021: a systematic analysis for the global burden of disease study 2021. The Lancet Neurology 2024, 23(10), 973–1003. [Google Scholar] [CrossRef]
- Moskowitz, M. A.; Lo, E. H.; Iadecola, C. The science of stroke: mechanisms in search of treatments. Neuron 2010, 67(2), 181–198. [Google Scholar] [CrossRef]
- Wardlaw, J. M.; Murray, V.; Berge, E.; Del Zoppo, G.; Sandercock, P.; Lindley, R. L.; Cohen, G. Recombinant tissue plasminogen activator for acute ischaemic stroke: an updated systematic review and meta-analysis. The Lancet 2012, 379(9834), 2364–2372. [Google Scholar] [CrossRef]
- Davis, S. M.; Donnan, G. A. 4.5 hours: the new time window for tissue plasminogen activator in stroke. Stroke 2009, 40(6), 2266–2267. [Google Scholar] [CrossRef]
- Hacke, W.; Kaste, M.; Bluhmki, E.; Brozman, M.; Dávalos, A.; Guidetti, D.; Larrue, V.; Lees, K. R.; Medeghri, Z.; Machnig, T. Thrombolysis with alteplase 3 to 4.5 hours after acute ischemic stroke. New England journal of medicine 2008, 359(13), 1317–1329. [Google Scholar] [CrossRef] [PubMed]
- Jickling, G. C.; Liu, D.; Ander, B. P.; Stamova, B.; Zhan, X.; Sharp, F. R. Targeting neutrophils in ischemic stroke: translational insights from experimental studies. Journal of Cerebral Blood Flow & Metabolism 2015, 35(6), 888–901. [Google Scholar] [CrossRef]
- Li, S.; Wang, F.; Li, X.; Chen, J.; Zhang, X.; Wang, Y.; Liu, J. Dipole orientation matters: Longer-circulating choline phosphate than phosphocholine liposomes for enhanced tumor targeting. ACS applied materials & interfaces 2017, 9(21), 17736–17744. [Google Scholar]
- Nogueira, R. G.; Jadhav, A. P.; Haussen, D. C.; Bonafe, A.; Budzik, R. F.; Bhuva, P.; Yavagal, D. R.; Ribo, M.; Cognard, C.; Hanel, R. A. Thrombectomy 6 to 24 hours after stroke with a mismatch between deficit and infarct. New England Journal of Medicine 2018, 378(1), 11–21. [Google Scholar] [CrossRef]
- Campbell, B. C.; Mitchell, P. J.; Kleinig, T. J.; Dewey, H. M.; Churilov, L.; Yassi, N.; Yan, B.; Dowling, R. J.; Parsons, M. W.; Oxley, T. J. Endovascular therapy for ischemic stroke with perfusion-imaging selection. New England Journal of Medicine 2015, 372(11), 1009–1018. [Google Scholar] [CrossRef]
- Hettiaratchi, M. H.; O’Meara, M. J.; Teal, C. J.; Payne, S. L.; Pickering, A. J.; Shoichet, M. S. Local delivery of stabilized chondroitinase abc degrades chondroitin sulfate proteoglycans in stroke-injured rat brains. Journal of controlled release 2019, 297, 14–25. [Google Scholar] [CrossRef]
- Karimi-Abdolrezaee, S.; Eftekharpour, E.; Wang, J.; Morshead, C. M.; Fehlings, M. G. Delayed transplantation of adult neural precursor cells promotes remyelination and functional neurological recovery after spinal cord injury. Journal of Neuroscience 2006, 26(13), 3377–3389. [Google Scholar] [CrossRef] [PubMed]
- George, P. M.; Oh, B.; Dewi, R.; Hua, T.; Cai, L.; Levinson, A.; Liang, X.; Krajina, B. A.; Bliss, T. M.; Heilshorn, S. C. Engineered stem cell mimics to enhance stroke recovery. Biomaterials 2018, 178, 63–72. [Google Scholar] [CrossRef]
- Yamawaki-Ogata, A.; Hashizume, R.; Satake, M.; Kaneko, H.; Mizutani, S.; Moritan, T.; Ueda, Y.; Narita, Y. A doxycycline loaded, controlled-release, biodegradable fiber for the treatment of aortic aneurysms. Biomaterials 2010, 31(36), 9554–9564. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Du, Y.; Wang, Y.; Wang, M.; Wang, L.; Yu, N.; Luo, S.; Wu, F.; Yang, G. Targeted drug delivery systems for matrix metalloproteinase-responsive anoparticles in tumor cells: a review. International Journal of Biological Macromolecules 2024, 257, 128658. [Google Scholar] [CrossRef]
- Bolan, F.; Louca, I.; Heal, C.; Cunningham, C. J. The potential of biomaterial-based approaches as therapies for ischemic stroke: a systematic review and meta-analysis of pre-clinical studies. Frontiers in Neurology 2019, 10, 924. [Google Scholar] [CrossRef]
- Saraiva, C.; Praça, C.; Ferreira, R.; Santos, T.; Ferreira, L.; Bernardino, L. Nanoparticle-mediated brain drug delivery: Overcoming blood–brain barrier to treat neurodegenerative diseases. Journal of controlled release 2016, 235, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Chamorro, Á.; Dirnagl, U.; Urra, X.; Planas, A. M. Neuroprotection in acute stroke: targeting excitotoxicity, oxidative and nitrosative stress, and inflammation. The Lancet Neurology 2016, 15(8), 869–881. [Google Scholar] [CrossRef] [PubMed]
- Lochhead, J. J.; Thorne, R. G. Intranasal delivery of biologics to the central nervous system. Advanced drug delivery reviews 2012, 64(7), 614–628. [Google Scholar] [CrossRef]
- Hu, Q.; Katti, P. S.; Gu, Z. Enzyme-responsive nanomaterials for controlled drug delivery. Nanoscale 2014, 6(21), 12273–12286. [Google Scholar] [CrossRef]
- Chu, S.; Shi, X.; Tian, Y.; Gao, F. ph-responsive polymer nanomaterials for tumor therapy. Frontiers in oncology 2022, 12, 855019. [Google Scholar] [CrossRef]
- Sipos, D.; Raposa, B. L.; Freihat, O.; Simon, M.; Mekis, N.; Cornacchione, P.; Kovács, Á. Glioblastoma: clinical presentation, multidisciplinary management, and long-term outcomes. Cancers 2025, 17(1), 146. [Google Scholar] [CrossRef]
- Cagney, D. N.; Martin, A. M.; Catalano, P. J.; Redig, A. J.; Lin, N. U.; Lee, E. Q.; Wen, P. Y.; Dunn, I. F.; Bi, W. L.; Weiss, S. E. Incidence and prognosis of patients with brain metastases at diagnosis of systemic malignancy: a population-based study. Neuro-oncology 2017, 19(11), 1511–1521. [Google Scholar] [CrossRef]
- Miroshnikova, Y. A.; Mouw, J. K.; Barnes, J. M.; Pickup, M. W.; Lakins, J. N.; Kim, Y.; Lobo, K.; Persson, A. I.; Reis, G. F.; McKnight, T. R. Tissue mechanics promote idh1-dependent hif1α–tenascin c feedback to regulate glioblastoma aggression. Nature cell biology 2016, 18(12), 1336–1345. [Google Scholar] [CrossRef]
- Habic, A.; Novak, M.; Majc, B.; Lah Turnsek, T.; Breznik, B. Proteases regulate cancer stem cell properties and remodel their microenvironment. Journal of Histochemistry & Cytochemistry 2021, 69(12), 775–794. [Google Scholar] [CrossRef] [PubMed]
- Mura, S.; Nicolas, J.; Couvreur, P. Stimuli-responsive nanocarriers for drug delivery. Nature materials 2013, 12(11), 991–1003. [Google Scholar] [CrossRef]
- Xia, Y.; Rao, L.; Yao, H.; Wang, Z.; Ning, P.; Chen, X. Engineering macrophages for cancer immunotherapy and drug delivery. Advanced materials 2020, 32(40), 2002054. [Google Scholar] [CrossRef]
- Hossain, M. A.; Liu, G.; Dai, B.; Si, Y.; Yang, Q.; Wazir, J.; Birnbaumer, L.; Yang, Y. Reinvigorating exhausted cd8+ cytotoxic t lymphocytes in the tumor microenvironment and current strategies in cancer immunotherapy. Medicinal Research Reviews 2021, 41(1), 156–201. [Google Scholar] [CrossRef]
- Arvanitis, C. D.; Ferraro, G. B.; Jain, R. K. The blood–brain barrier and blood–tumour barrier in brain tumours and metastases. Nature Reviews Cancer 2020, 20(1), 26–41. [Google Scholar] [CrossRef]
- Vaupel, P. The role of hypoxia-induced factors in tumor progression. The oncologist 2004, 9(S5), 10–17. [Google Scholar] [CrossRef]
- Nance, E. A.; Woodworth, G. F.; Sailor, K. A.; Shih, T.-Y.; Xu, Q.; Swaminathan, G.; Xiang, D.; Eberhart, C.; Hanes, J. A dense poly (ethylene glycol) coating improves penetration of large polymeric nanoparticles within brain tissue. Science translational medicine 2012, 4(149), 149ra119–149ra119. [Google Scholar] [CrossRef] [PubMed]
- Bobo, R. H.; Laske, D. W.; Akbasak, A.; Morrison, P. F.; Dedrick, R. L.; Oldfield, E. H. Convection-enhanced delivery of macromolecules in the brain. Proceedings of the National Academy of Sciences 1994, 91(6), 2076–2080. [Google Scholar] [CrossRef]
- Gutenberg, A.; Lumenta, C.; Braunsdorf, W.; Sabel, M.; Mehdorn, H.; Westphal, M.; Giese, A. The combination of carmustine wafers and temozolomide for the treatment of malignant gliomas. a comprehensive review of the rationale and clinical experience. Journal of neuro-oncology 2013, 113(2), 163–174. [Google Scholar] [CrossRef] [PubMed]
- Hynynen, K.; McDannold, N.; Vykhodtseva, N.; Jolesz, F. A. Noninvasive mr imaging–guided focal opening of the blood-brain barrier in rabbits. Radiology 2001, 220(3), 640–646. [Google Scholar] [CrossRef]
- Kulkarni, J. A.; Witzigmann, D.; Chen, S.; Cullis, P. R.; Van Der Meel, R. Lipid nanoparticle technology for clinical translation of sirna therapeutics. Accounts of chemical research 2019, 52(9), 2435–2444. [Google Scholar] [CrossRef] [PubMed]
- Kamerkar, S.; LeBleu, V. S.; Sugimoto, H.; Yang, S.; Ruivo, C. F.; Melo, S. A.; Lee, J. J.; Kalluri, R. Exosomes facilitate therapeutic targeting of oncogenic kras in pancreatic cancer. Nature 2017, 546(7659), 498–503. [Google Scholar] [CrossRef]
- Haqqani, A. S.; Bélanger, K.; Stanimirovic, D. B. Receptor-mediated transcytosis for brain delivery of therapeutics: receptor classes and criteria. Frontiers in Drug Delivery 2024, 4, 1360302. [Google Scholar] [CrossRef] [PubMed]
- Patel, T.; Zhou, J.; Piepmeier, J. M.; Saltzman, W. M. Polymeric nanoparticles for drug delivery to the central nervous system. Advanced drug delivery reviews 2012, 64(7), 701–705. [Google Scholar] [CrossRef]
- Heidarzadeh, M.; Gursoy-Ozdemir, Y.; Kaya, M.; Eslami Abriz, A.; Zarebkohan, A.; Rahbarghazi, R.; Sokullu, E. Exosomal delivery of therapeutic modulators through the blood–brain barrier; promise and pitfalls. Cell & Bioscience 2021, 11(1), 142. [Google Scholar] [CrossRef]
- Luan, X.; Sansanaphongpricha, K.; Myers, I.; Chen, H.; Yuan, H.; Sun, D. Engineering exosomes as refined biological nanoplatforms for drug delivery. Acta Pharmacologica Sinica 2017, 38(6), 754–763. [Google Scholar] [CrossRef]
- Rapoport, S. I. Osmotic opening of the blood–brain barrier: principles, mechanism, and therapeutic applications. Cellular and molecular neurobiology 2000, 20(2), 217–230. [Google Scholar] [CrossRef]
- Pardridge, W. M. Receptor-mediated drug delivery of bispecific therapeutic antibodies through the blood-brain barrier. Frontiers in drug delivery 2023, 3, 1227816. [Google Scholar] [CrossRef] [PubMed]
- McDermott, J.; Ben-Shalom, N.; D’Amico, R. S.; Boockvar, J. A. Intra-arterial drug delivery for brain tumors; Current Status and Future Directions: Arterial Treatment of Neurological Diseases, 2025; pp. 109–117. [Google Scholar]
- Caraway, C. A.; Gaitsch, H.; Wicks, E. E.; Kalluri, A.; Kunadi, N.; Tyler, B. M. Polymeric nanoparticles in brain cancer therapy: A review of current approaches. Polymers 2022, 14(14), 2963. [Google Scholar] [CrossRef]
- Lu, X.; Dong, J.; Zheng, D.; Li, X.; Ding, D.; Xu, H. Reperfusion combined with intraarterial administration of resveratrol-loaded nanoparticles improved cerebral ischemia–reperfusion injury in rats. Nanomedicine: Nanotechnology, Biology and Medicine 2020, 28, 102208. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Hynynen, K.; Lipsman, N. Applications of focused ultrasound in the brain: from thermoablation to drug delivery. Nature Reviews Neurology 2021, 17(1), 7–22. [Google Scholar] [CrossRef]
- Grasso, G.; Torregrossa, F.; Noto, M.; Bruno, E.; Feraco, P.; Buscemi, F.; Bartolotta, T. V.; Gagliardo, C. Mr-guided focused ultrasound–induced blood-brain barrier opening for brain metastasis: a review. Neurosurgical focus 2023, 55(2), E11. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Wang, P.; Sun, S.; Liang, X. Applications of micro/nanotechnology in ultrasound-based drug delivery and therapy for tumor. Current medicinal chemistry 2021, 28(3), 525–547. [Google Scholar]
- Lipsman, N.; Meng, Y.; Bethune, A. J.; Huang, Y.; Lam, B.; Masellis, M.; Herrmann, N.; Heyn, C.; Aubert, I.; Boutet, A. Blood–brain barrier opening in alzheimer’s disease using mr-guided focused ultrasound. Nature communications 2018, 9(1), 2336. [Google Scholar] [CrossRef]
- Chen, K.-T.; Chai, W.-Y.; Lin, Y.-J.; Lin, C.-J.; Chen, P.-Y.; Tsai, H.-C.; Huang, C.-Y.; Kuo, J. S.; Liu, H.-L.; Wei, K.-C. Neuronavigation-guided focused ultrasound for transcranial blood-brain barrier opening and immunostimulation in brain tumors. Science Advances 2021, 7(6), eabd0772. [Google Scholar] [CrossRef]
- Wu, H.; Zhou, Y.; Wang, Y.; Tong, L.; Wang, F.; Song, S.; Xu, L.; Liu, B.; Yan, H.; Sun, Z. Current state and future directions of intranasal delivery route for central nervous system disorders: a scientometric and visualization analysis. Frontiers in pharmacology 2021, 12, 717192. [Google Scholar] [CrossRef]
- Dufes, C.; Olivier, J.-C.; Gaillard, F.; Gaillard, A.; Couet, W.; Muller, J.-M. Brain delivery of vasoactive intestinal peptide (vip) following nasal administration to rats. International journal of pharmaceutics 2003, 255(1-2), 87–97. [Google Scholar] [CrossRef]
- Alcala-Barraza, S. R.; Lee, M. S.; Hanson, L. R.; McDonald, A. A.; Frey, W. H.; McLoon, L. K. Intranasal delivery of neurotrophic factors bdnf, cntf, epo, and nt-4 to the cns. Journal of drug targeting 2010, 18(3), 179–190. [Google Scholar] [CrossRef]
- Awad, R.; Avital, A.; Sosnik, A. Polymeric nanocarriers for nose-to-brain drug delivery in neurodegenerative diseases and neurodevelopmental disorders. Acta Pharmaceutica Sinica B 2023, 13(5), 1866–1886. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Wang, F.; Wang, K.; Zhong, Y.; Wei, X.; Wang, Q.; Zhang, H. Engineered exosomes: a promising drug delivery strategy for brain diseases. Current Medicinal Chemistry 2022, 29(17), 3111–3124. [Google Scholar] [CrossRef]
- Oliveira, J. M.; Carvalho, L.; Silva-Correia, J.; Vieira, S.; Majchrzak, M.; Lukomska, B.; Stanaszek, L.; Strymecka, P.; Malysz-Cymborska, I.; Golubczyk, D. Hydrogel-based scaffolds to support intrathecal stem cell transplantation as a gateway to the spinal cord: clinical needs, biomaterials, and imaging technologies. NPJ Regenerative medicine 2018, 3(1), 8. [Google Scholar] [CrossRef]
- Strazielle, N.; Ghersi-Egea, J.-F. Potential pathways for cns drug delivery across the blood-cerebrospinal fluid barrier. Current pharmaceutical design 2016, 22(35), 5463–5476. [Google Scholar] [CrossRef] [PubMed]
- Sudhakar, V.; Richardson, R. M. Gene therapy for neurodegenerative diseases. Neurotherapeutics 2019, 16(1), 166–175. [Google Scholar] [CrossRef] [PubMed]
- Arjomandnejad, M.; Dasgupta, I.; Flotte, T. R.; Keeler, A. M. Immunogenicity of recombinant adeno-associated virus (aav) vectors for gene transfer. BioDrugs 2023, 37(3), 311. [Google Scholar] [CrossRef] [PubMed]
- Nance, E.; Zhang, C.; Shih, T.-Y.; Xu, Q.; Schuster, B. S.; Hanes, J. Brain-penetrating nanoparticles improve paclitaxel efficacy in malignant glioma following local administration. ACS nano 2014, 8(10), 10655–10664. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, Y.; Ding, J.; Li, Z.; Tian, Y.; Zeng, M.; Wu, X.; Su, B.; Jiang, J.; Wu, C. Hydrogel-based multifunctional deep brain probe for neural sensing, manipulation, and therapy. ACS nano, 2025. [Google Scholar]
- Leong, K.; D’Amore, P.; Marletta, M.; Langer, R. Bioerodible polyanhydrides as drug-carrier matrices. ii. biocompatibility and chemical reactivity. Journal of biomedical materials research 1986, 20(1), 51–64. [Google Scholar] [CrossRef]
- Pena, E. S.; Graham-Gurysh, E. G.; Bachelder, E. M.; Ainslie, K. M. Design of biopolymer-based interstitial therapies for the treatment of glioblastoma. International journal of molecular sciences 2021, 22(23), 13160. [Google Scholar] [CrossRef]
- Ball, K.; Bouzom, F.; Scherrmann, J.-M.; Walther, B.; Decleves, X. Physiologically based pharmacokinetic modelling of drug penetration across the blood–brain barrier—towards a mechanistic ivive-based approach. The AAPS journal 2013, 15(4), 913–932. [Google Scholar] [CrossRef]
- Ball, K.; Bouzom, F.; Scherrmann, J.-M.; Walther, B.; Declèves, X. A physiologically based modeling strategy during preclinical cns drug development. Molecular pharmaceutics 2014, 11(3), 836–848. [Google Scholar] [CrossRef]
- Bloomingdale, P.; Bakshi, S.; Maass, C.; van Maanen, E.; Pichardo-Almarza, C.; Yadav, D. B.; van der Graaf, P.; Mehrotra, N. Minimal brain pbpk model to support the preclinical and clinical development of antibody therapeutics for cns diseases. Journal of pharmacokinetics and pharmacodynamics 2021, 48(6), 861–871. [Google Scholar] [CrossRef] [PubMed]
- Mehta, P.; Soliman, A.; Rodriguez-Vera, L.; Schmidt, S.; Muniz, P.; Rodriguez, M.; Forcadell, M.; Gonzalez-Perez, E.; Vozmediano, V. Interspecies brain pbpk modeling platform to predict passive transport through the blood–brain barrier and assess target site disposition. Pharmaceutics 2024, 16(2), 226. [Google Scholar] [CrossRef] [PubMed]
- Gaohua, L.; Neuhoff, S.; Johnson, T. N.; Rostami-Hodjegan, A.; Jamei, M. Development of a permeability-limited model of the human brain and cerebrospinal fluid (csf) to integrate known physiological and biological knowledge: Estimating time varying csf drug concentrations and their variability using in vitro data. Drug metabolism and pharmacokinetics 2016, 31(3), 224–233. [Google Scholar] [CrossRef]
- Monine, M.; Norris, D.; Wang, Y.; Nestorov, I. A physiologically-based pharmacokinetic model to describe antisense oligonucleotide distribution after intrathecal administration. Journal of Pharmacokinetics and Pharmacodynamics 2021, 48(5), 639–654. [Google Scholar] [CrossRef]
- Shannon, R. J.; Carpenter, K. L.; Guilfoyle, M. R.; Helmy, A.; Hutchinson, P. J. Cerebral microdialysis in clinical studies of drugs: pharmacokinetic applications. Journal of pharmacokinetics and pharmacodynamics 2013, 40(3), 343–358. [Google Scholar] [CrossRef] [PubMed]
- Drath, I.; Richter, F.; Feja, M. Nose-to-brain drug delivery: from bench to bedside. Translational neurodegeneration 2025, 14(1), 23. [Google Scholar] [CrossRef]
- Rygg, A.; Hindle, M.; Longest, P. W. Linking suspension nasal spray drug deposition patterns to pharmacokinetic profiles: A proof-of-concept study using computational fluid dynamics. Journal of Pharmaceutical Sciences 2016, 105(6), 1995–2004. [Google Scholar] [CrossRef]
- Burgess, A.; Shah, K.; Hough, O.; Hynynen, K. Focused ultrasound-mediated drug delivery through the blood–brain barrier. Expert review of neurotherapeutics 2015, 15(5), 477–491. [Google Scholar] [CrossRef]
- Wu, S.-K.; Chu, P.-C.; Chai, W.-Y.; Kang, S.-T.; Tsai, C.-H.; Fan, C.-H.; Yeh, C.-K.; Liu, H.-L. Characterization of different microbubbles in assisting focused ultrasound-induced blood-brain barrier opening. Scientific reports 2017, 7(1), 46689. [Google Scholar] [CrossRef]
- Nicholson, C.; Hrabetova, S. Brain extracellular space: the final frontier of neuroscience. Biophysical journal 2017, 113(10), 2133–2142. [Google Scholar] [CrossRef]
- Tzafriri, A. R.; Edelman, E. R. Convective and diffusive transport in drug delivery. Cancer Targeted Drug Delivery: An Elusive Dream 2013, 573–606. [Google Scholar]
- Krewson, C. E.; Klarman, M. L.; Saltzman, W. M. Distribution of nerve growth factor following direct delivery to brain interstitium. Brain research 1995, 680(1-2), 196–206. [Google Scholar] [CrossRef]
- Kouzehgarani, G. N.; Feldsien, T.; Engelhard, H. H.; Mirakhur, K. K.; Phipps, C.; Nimmrich, V.; Clausznitzer, D.; Lefebvre, D. R. Harnessing cerebrospinal fluid circulation for drug delivery to brain tissues. Advanced Drug Delivery Reviews 2021, 173, 20–59. [Google Scholar] [CrossRef]
- Mestre, H.; Tithof, J.; Du, T.; Song, W.; Peng, W.; Sweeney, A. M.; Olveda, G.; Thomas, J. H.; Nedergaard, M.; Kelley, D. H. Flow of cerebrospinal fluid is driven by arterial pulsations and is reduced in hypertension. Nature communications 2018, 9(1), 4878. [Google Scholar] [CrossRef]
- Bohr, T.; Hjorth, P. G.; Holst, S. C.; Hrabetova, S.; Kiviniemi, V.; Lilius, T.; Lundgaard, I.; Mardal, K.-A.; Martens, E. A.; Mori, Y. The glymphatic system: Current understanding and modeling. IScience 2022, 25(9). [Google Scholar] [CrossRef]
- Quirk, K.; Boster, K. A.; Tithof, J.; Kelley, D. H. A brain-wide solute transport model of the glymphatic system. Journal of the Royal Society Interface 2024, 21(219), 20240369. [Google Scholar] [CrossRef]
- Vishnumurthy, P.; Radulesco, T.; Bouchet, G.; Regard, A.; Michel, J. Computational fluid dynamics approach for direct nose-to-brain drug delivery: A systematic review and meta-analysis. Journal of Personalized Medicine 2025, 15(10), 447. [Google Scholar] [CrossRef]
- Boyuklieva, R.; Zagorchev, P.; Pilicheva, B. Computational, in vitro, and in vivo models for nose-to-brain drug delivery studies. Biomedicines 2023, 11(8), 2198. [Google Scholar] [CrossRef] [PubMed]
- Downs, M. E.; Buch, A.; Karakatsani, M. E.; Konofagou, E. E.; Ferrera, V. P. Blood-brain barrier opening in behaving non-human primates via focused ultrasound with systemically administered microbubbles. Scientific reports 2015, 5(1), 15076. [Google Scholar] [CrossRef]
- Arifin, D. Y.; Lee, K. Y. T.; Wang, C.-H. Chemotherapeutic drug transport to brain tumor. Journal of controlled release 2009, 137(3), 203–210. [Google Scholar] [CrossRef] [PubMed]
- Pedram, M. Z.; Shamloo, A.; Alasty, A.; Ghafar-Zadeh, E. Optimal magnetic field for crossing super-para-magnetic nanoparticles through the brain blood barrier: a computational approach. Biosensors 2016, 6(2), 25. [Google Scholar] [CrossRef]
- Müller, W. A.; Sarkis, J. R.; Marczak, L. D. F.; Muniz, A. R. Computational analysis of the simultaneous application of ultrasound and electric fields in a lipid bilayer. Biochimica et Biophysica Acta (BBA)-Biomembranes 2024, 1866(7), 184364. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Xiong, Y.; Chen, H.; Gao, X.; Dai, J.; Zhang, Y.; Zou, W.; Gao, Y.; Jiang, Z.; Han, B. Mucus adhesion vs. mucus penetration? screening nanomaterials for nasal inhalation by md simulation. Journal of Controlled Release 2023, 353, 366–379. [Google Scholar] [CrossRef] [PubMed]
- Jagger, B. R.; Kochanek, S. E.; Haldar, S.; Amaro, R. E.; Mulholland, A. J. Multiscale simulation approaches to modeling drug–protein binding. Current opinion in structural biology 2020, 61, 213–221. [Google Scholar] [CrossRef]
- Rim, J. E.; Pinsky, P. M.; Van Osdol, W. W. Multiscale modeling framework of transdermal drug delivery. Annals of biomedical engineering 2009, 37(6), 1217–1229. [Google Scholar] [CrossRef]
- Goraya, S. A. Computational modeling of nanoparticle transport and adhesion under shear-dependent flow: A multiscale digital-twin framework, Brain MultiphysicsInferred from uploaded PDF. Brain Multiphysics 2024. [Google Scholar]
- Akalın, A. A.; Dedekargınoğlu, B.; Choi, S. R.; Han, B.; Ozcelikkale, A. Predictive design and analysis of drug transport by multiscale computational models under uncertainty. Pharmaceutical research 2023, 40(2), 501–523. [Google Scholar] [CrossRef]
- Mohammadi, M.; Sefidgar, M.; Aghanajafi, C.; Kohandel, M.; Soltani, M. Computational multi-scale modeling of drug delivery into an anti-angiogenic therapy-treated tumor. Cancers 2023, 15(22), 5464. [Google Scholar] [CrossRef]
- Buchete, N.-V.; Cicha, I.; Dutta, S.; Neofytou, P. Multiscale physics-based in silico modelling of nanocarrier-assisted intravascular drug delivery. Frontiers in Drug Delivery 2024, 4, 1362660. [Google Scholar] [CrossRef]
- Huang, D.; Wang, Q.; Cao, Y.; Yang, H.; Li, M.; Wu, F.; Zhang, Y.; Chen, G.; Wang, Q. Multiscale nir-ii imaging-guided brain-targeted drug delivery using engineered cell membrane nanoformulation for alzheimer’s disease therapy. ACS nano 2023, 17(5), 5033–5046. [Google Scholar] [CrossRef]
- Tarki, F. E.; Sharbatdar, M.; Zarrabi, M.; Vafaee, F.; Khanbabaei, G. Computational insights into intranasal drug delivery: Enhancing outcomes in pediatric cystic fibrosis. Respiratory Medicine 2025, 108430. [Google Scholar] [CrossRef]
- Zhou, L.; Nguyen, T. D.; Chiang, G. C.; Keil, S. A.; Wang, X. H.; Hu, T.-W.; Lan, H.; Xi, K.; Costa, A. P.; Tanzi, E. B. Brain glymphatic fluid mapping in alzheimer’s disease: a human mri and pet study. Brain Communications 2025, 7(3), fcaf200. [Google Scholar] [CrossRef] [PubMed]
- Taoka, T.; Ito, R.; Nakamichi, R.; Nakane, T.; Kawai, H.; Naganawa, S. Diffusion tensor image analysis along the perivascular space (dti-alps): revisiting the meaning and significance of the method. Magnetic Resonance in Medical Sciences 2024, 23(3), 268–290. [Google Scholar] [CrossRef] [PubMed]
- Vanlandewijck, M.; He, L.; Mäe, M. A.; Andrae, J.; Ando, K.; Del Gaudio, F.; Nahar, K.; Lebouvier, T.; Laviña, B.; Gouveia, L. A molecular atlas of cell types and zonation in the brain vasculature. Nature 2018, 554(7693), 475–480. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Central nervous system and neurovascular unit. The neurovascular unit comprises endothelial cells, pericytes, astrocytes, microglia, neurons, smooth muscle cells, and the surrounding extracellular matrix, working together to tightly regulate BBB permeability and cerebral homeostasis.
Figure 1.
Central nervous system and neurovascular unit. The neurovascular unit comprises endothelial cells, pericytes, astrocytes, microglia, neurons, smooth muscle cells, and the surrounding extracellular matrix, working together to tightly regulate BBB permeability and cerebral homeostasis.
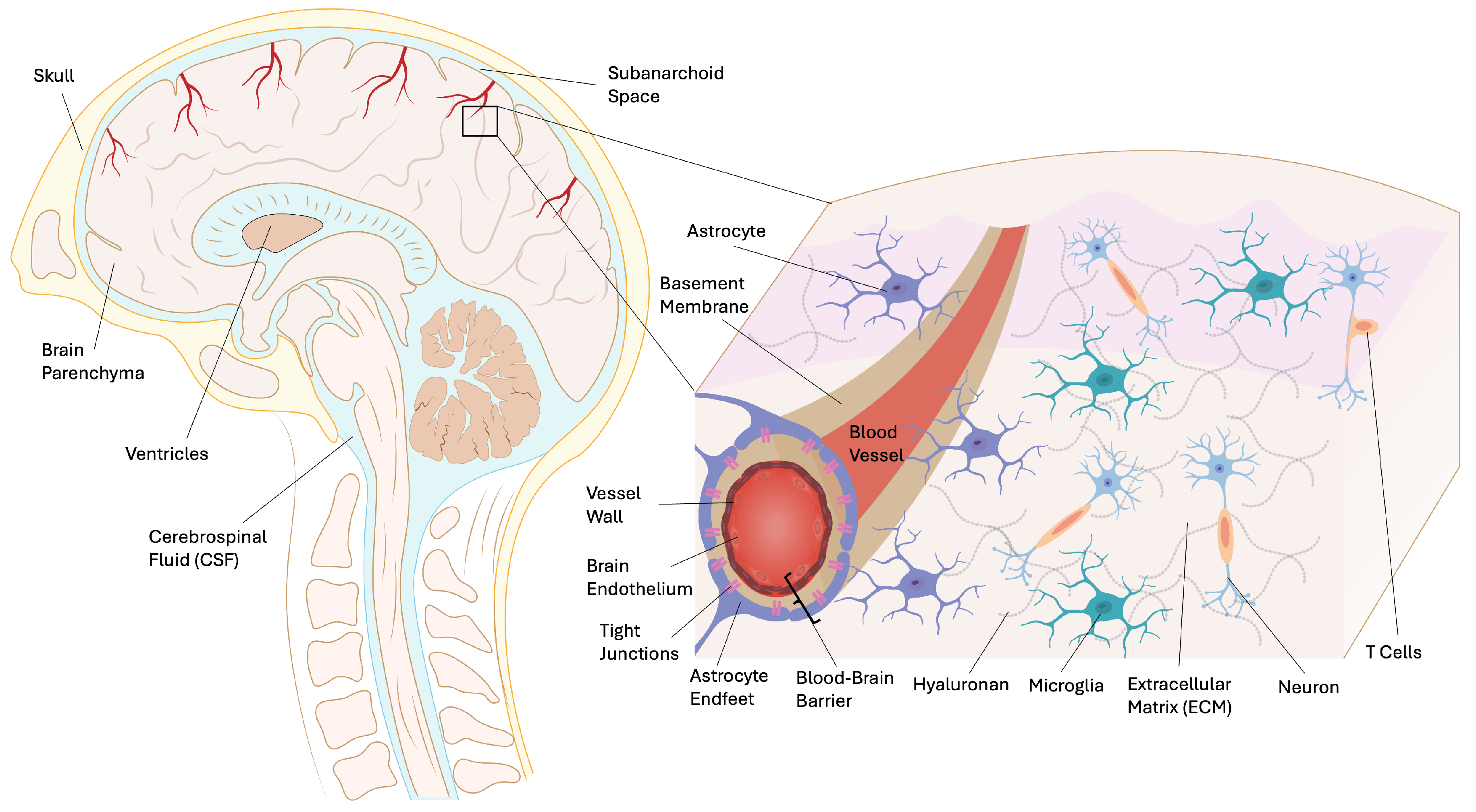
Figure 2.
Neurodgeneration physiology is characterized by increased amyloid and sulfated proteoglycan deposition. Together with disease-specific protease action, these are targets for drug delivery.
Figure 2.
Neurodgeneration physiology is characterized by increased amyloid and sulfated proteoglycan deposition. Together with disease-specific protease action, these are targets for drug delivery.
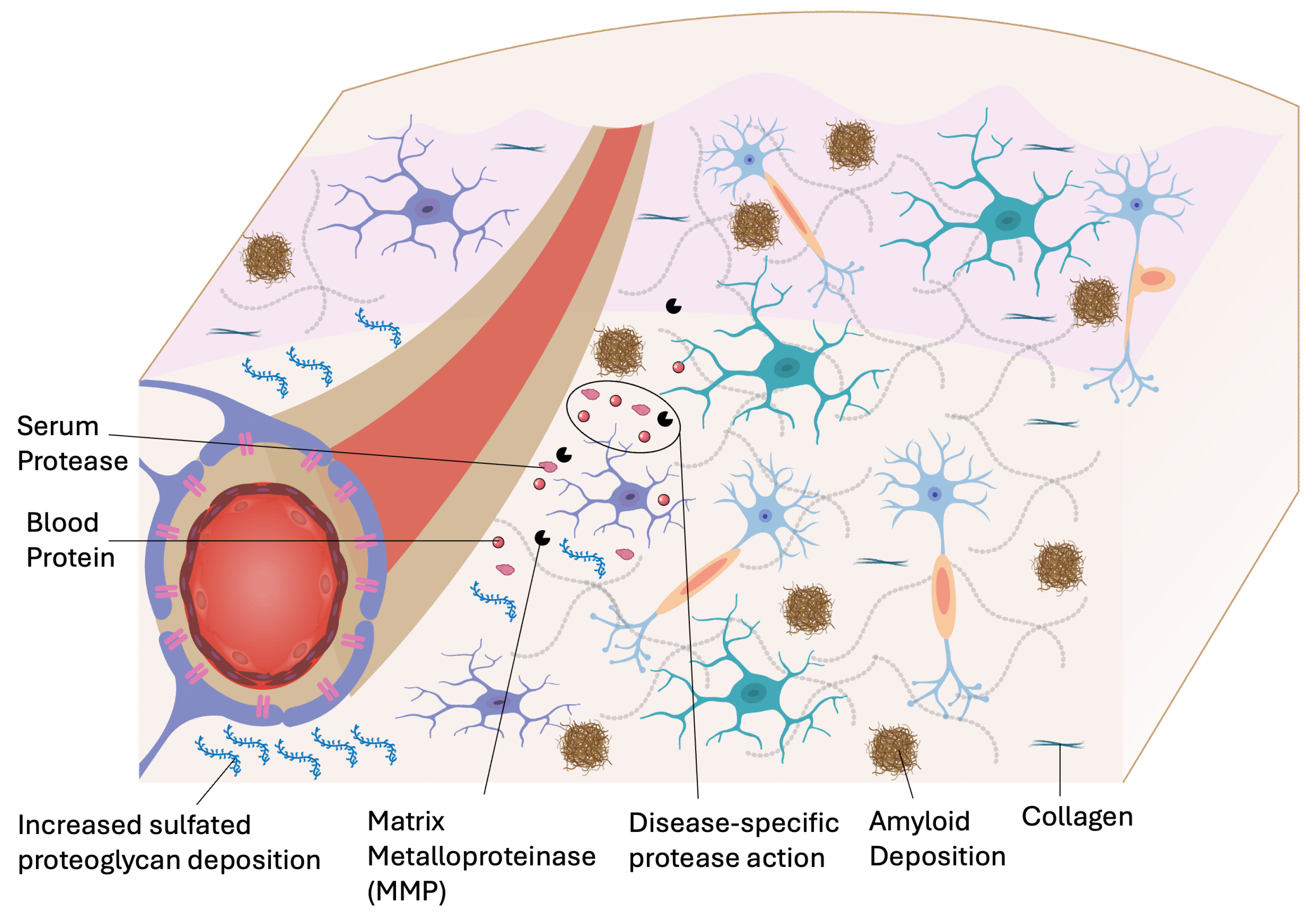
Figure 3.
In stroke, vascular breakdown leads to increased tissue permeability, edema, and tissue necrosis. Major targets for drug delivery include infiltrating monocytes and macrophages as well as increased protease and MMP activity.
Figure 3.
In stroke, vascular breakdown leads to increased tissue permeability, edema, and tissue necrosis. Major targets for drug delivery include infiltrating monocytes and macrophages as well as increased protease and MMP activity.
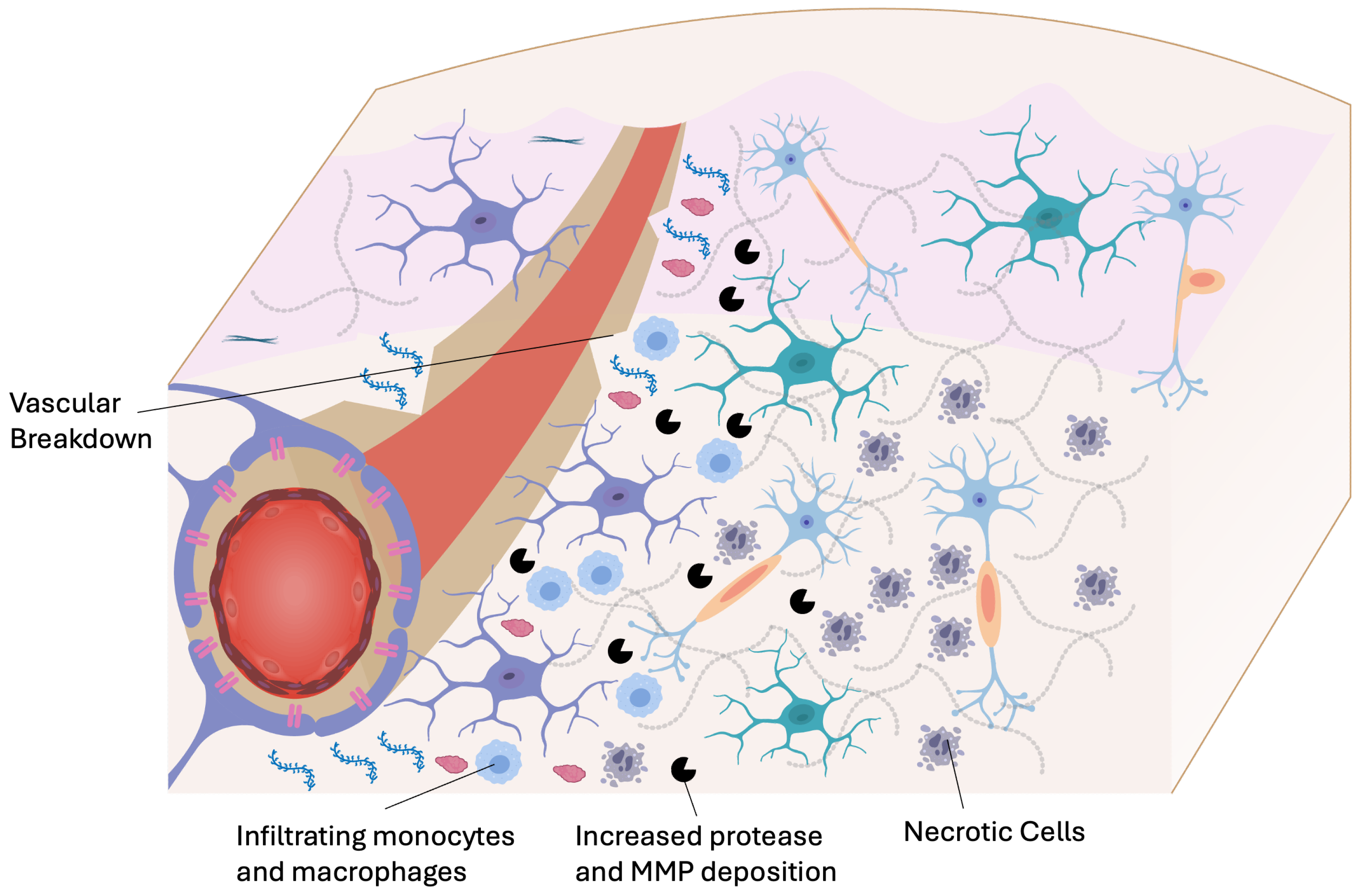
Figure 4.
Brain tumors disrupt CNS physiology through breakdown of the blood–brain barrier, abnormal and leaky angiogenesis, and elevated interstitial pressure that impairs perfusion and drug delivery. Tumors induce chronic neuroinflammation, remodel the extracellular matrix via proteases, and create hypoxic, metabolically reprogrammed microenvironments.
Figure 4.
Brain tumors disrupt CNS physiology through breakdown of the blood–brain barrier, abnormal and leaky angiogenesis, and elevated interstitial pressure that impairs perfusion and drug delivery. Tumors induce chronic neuroinflammation, remodel the extracellular matrix via proteases, and create hypoxic, metabolically reprogrammed microenvironments.
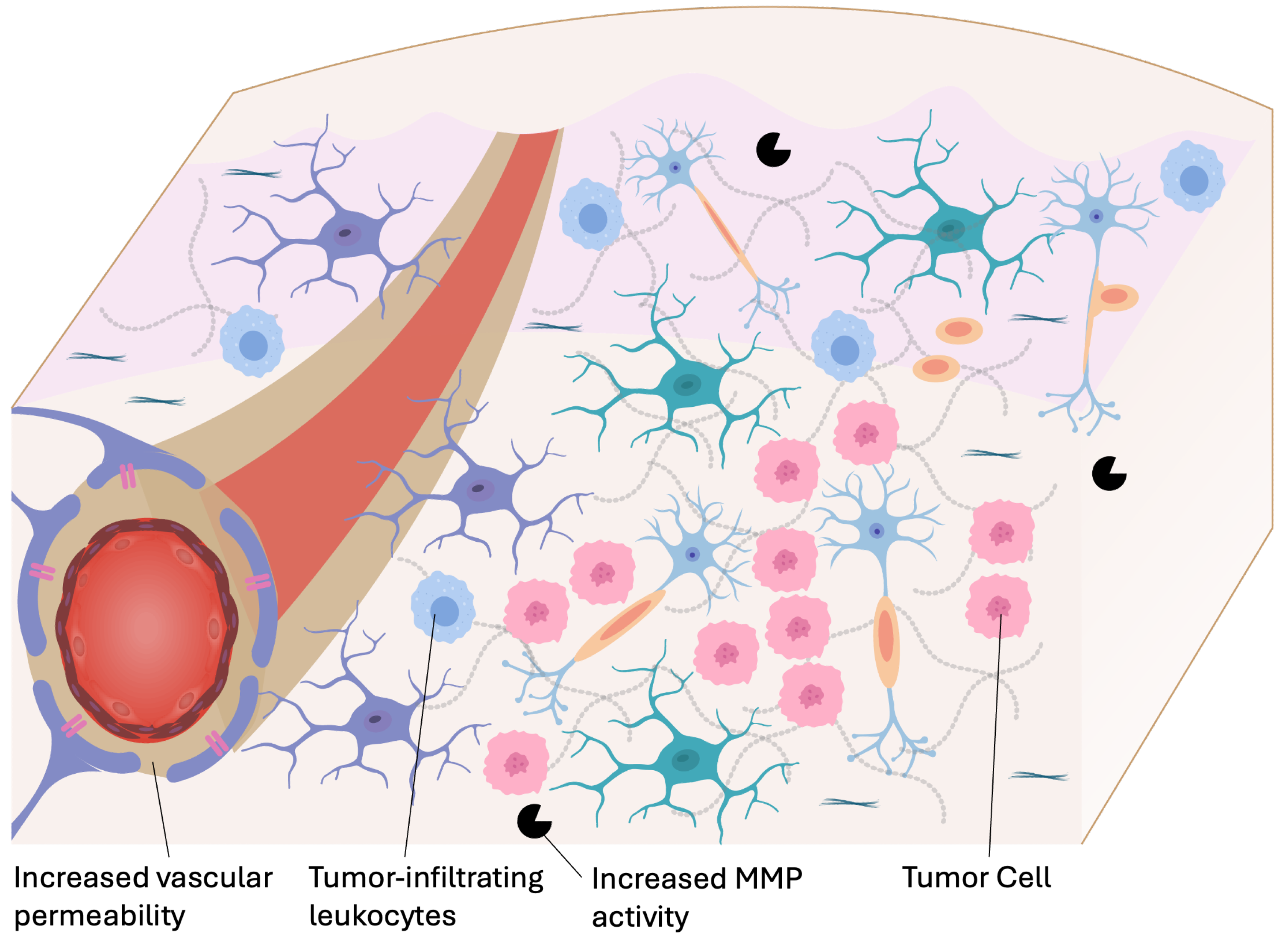
Figure 5.
Schematic diagram showing major drug delivery approaches to the CNS.
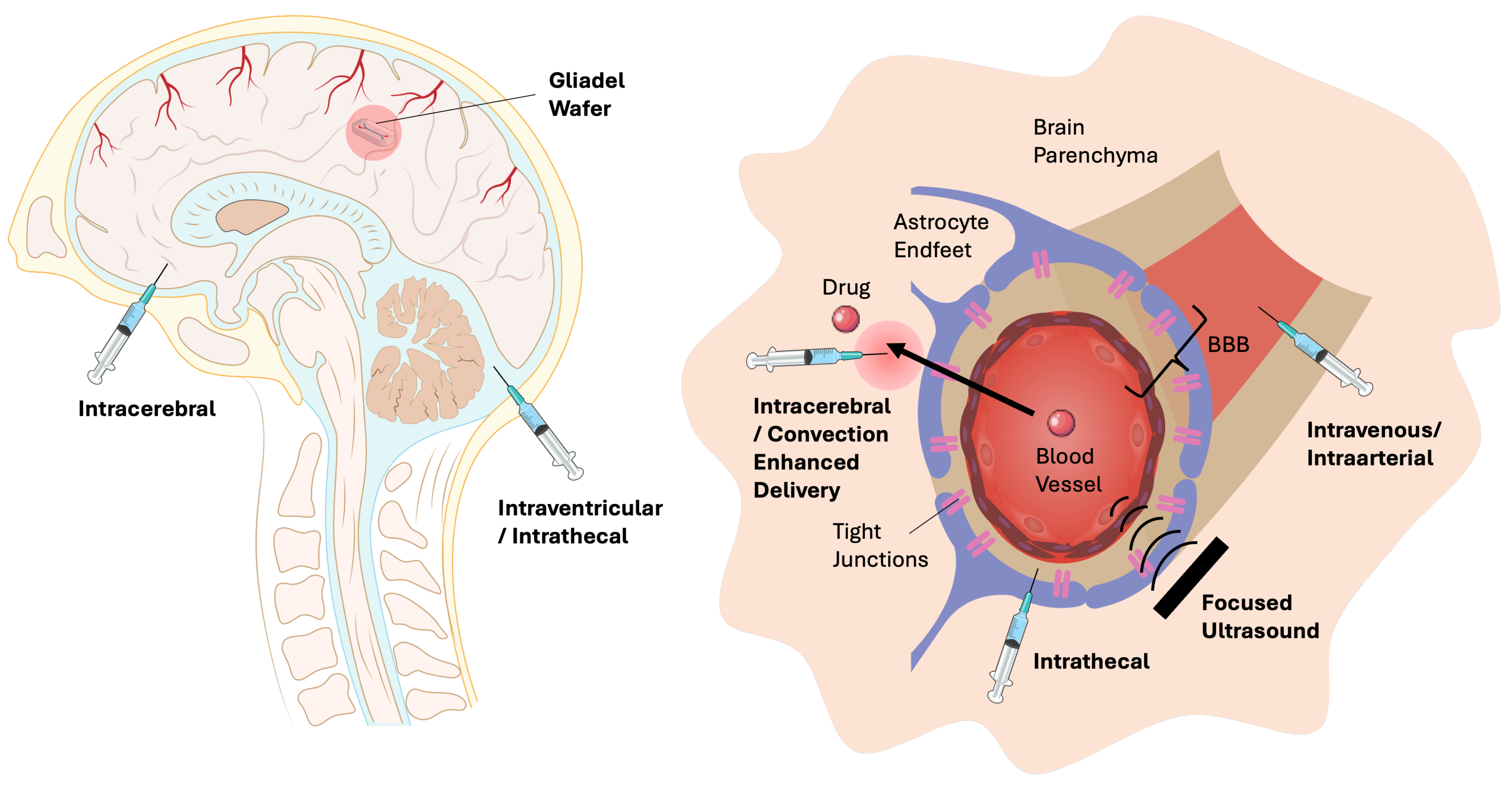
Figure 6.
Computational fluid dynamics simulation of intrathecal drug delivery in an anatomically idealized model of the human central nervous system. Spatio-temporal representation of tracer concentration as a percentage of injected dose over 1-hour for the (a) rigid bench-top model, (b) rigid digital model, and (c) compliant digital model. (Image reproduced from Khani et al. [3] under Creative Commons CC BY 4.0 License).
Figure 6.
Computational fluid dynamics simulation of intrathecal drug delivery in an anatomically idealized model of the human central nervous system. Spatio-temporal representation of tracer concentration as a percentage of injected dose over 1-hour for the (a) rigid bench-top model, (b) rigid digital model, and (c) compliant digital model. (Image reproduced from Khani et al. [3] under Creative Commons CC BY 4.0 License).
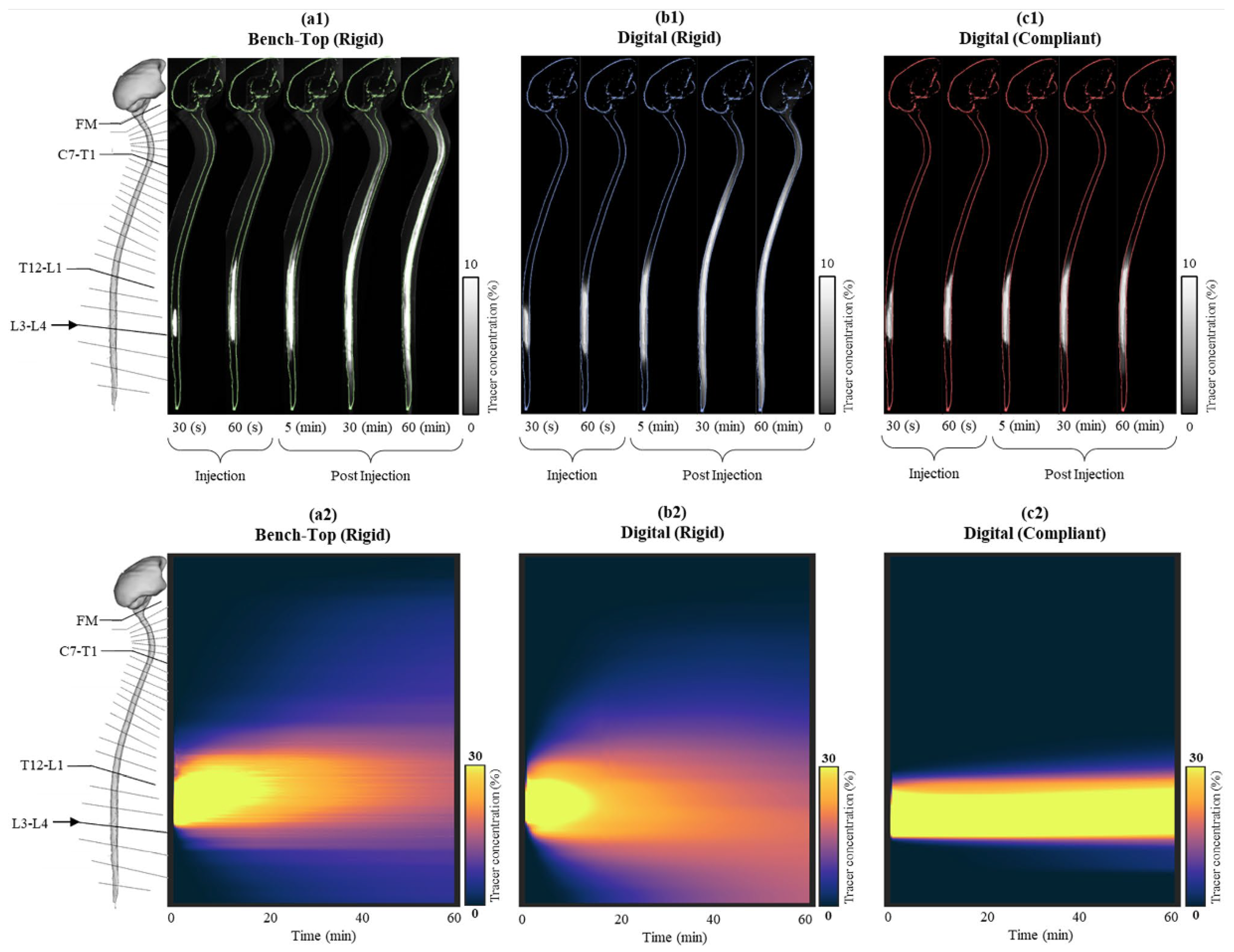
Figure 7.
MD simulation of initial and final positions of different unconjugated cell-penetrating peptides (CPP) conjugates during 1000 ns membrane simulations: (A) penetratin–doxorubicin, (B) 6,14-Phe-penetratin–doxorubicin, (C) dodeca-penetratin–doxorubicin, (D) penetratin–rasagiline, (E) 6,14-Phe-penetratin–rasagiline, (F) dodeca-penetratin–rasagiline, (G) penetratin–zidovudine, (H) 6,14-Phe-penetratin–zidovudine, (I) dodeca-penetratin–zidovudine. (Image reproduced from Ivanczi et al. under Creative Commons CC BY 4.0 license [8])
Figure 7.
MD simulation of initial and final positions of different unconjugated cell-penetrating peptides (CPP) conjugates during 1000 ns membrane simulations: (A) penetratin–doxorubicin, (B) 6,14-Phe-penetratin–doxorubicin, (C) dodeca-penetratin–doxorubicin, (D) penetratin–rasagiline, (E) 6,14-Phe-penetratin–rasagiline, (F) dodeca-penetratin–rasagiline, (G) penetratin–zidovudine, (H) 6,14-Phe-penetratin–zidovudine, (I) dodeca-penetratin–zidovudine. (Image reproduced from Ivanczi et al. under Creative Commons CC BY 4.0 license [8])
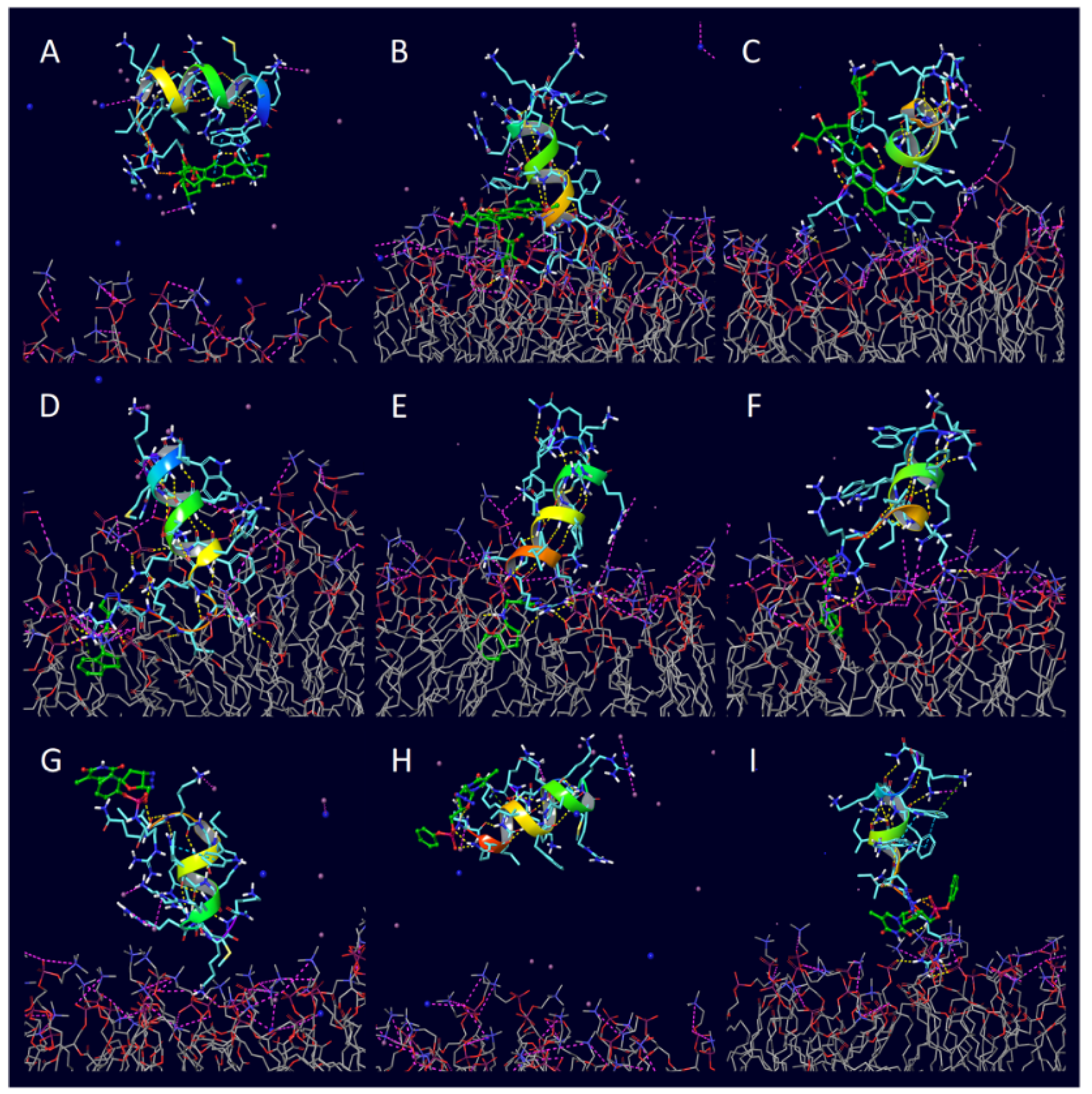
Figure 8.
Schematic diagram showing coarse-graining of microscale kinetics into macroscale fluid dynamics in hybrid models [4,129].
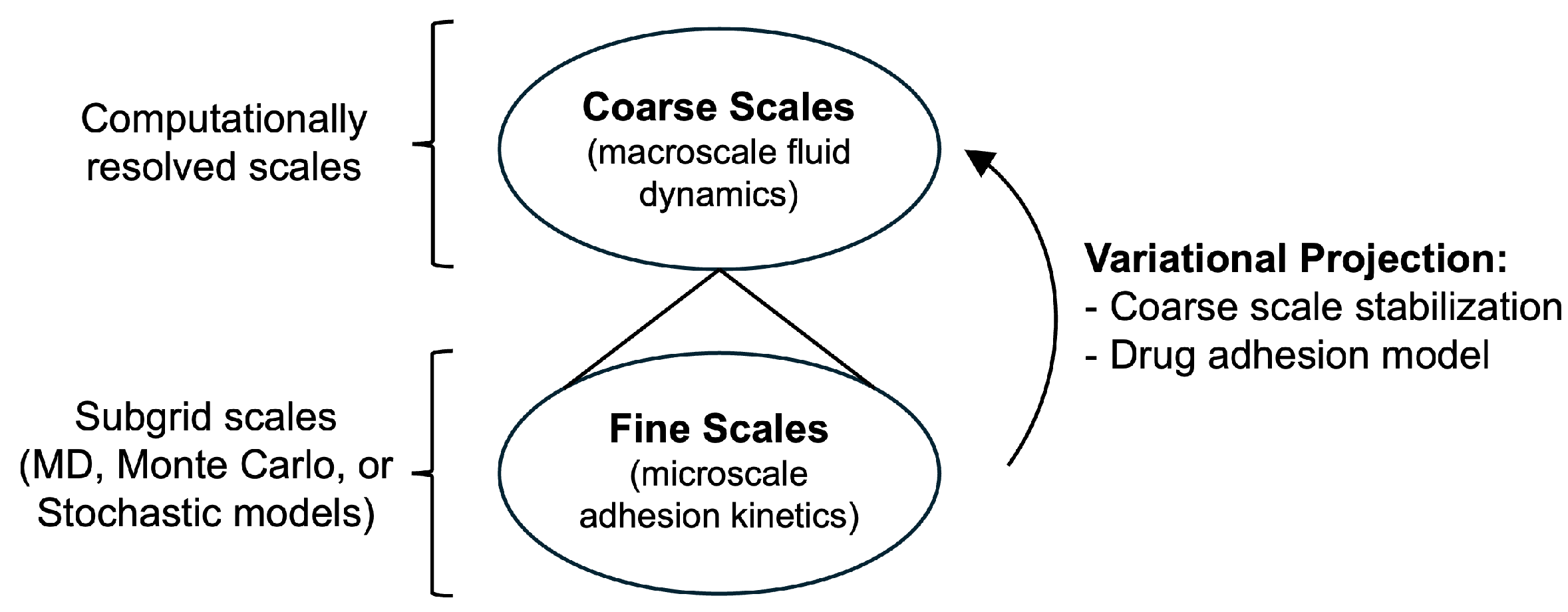
Figure 9.
Schematic diagram of drug delivery using nanocarriers designed for active targeting in circulation. The experimentally validated computational model serves as a digital twin, enabling investigation of the effects of hemodynamic forces and particle geometry on particle adhesion in patient-specific arterial models. A shear-rate dependent blood flow model along with resistance boundary conditions for downstream pressure is used to simulate pulsatile blood flow in the arterial system. Adhesion kinetics are simulated through various multiscale models that incorporate experimentally measured parameters in the boundary layer at the target sites [129].
Figure 9.
Schematic diagram of drug delivery using nanocarriers designed for active targeting in circulation. The experimentally validated computational model serves as a digital twin, enabling investigation of the effects of hemodynamic forces and particle geometry on particle adhesion in patient-specific arterial models. A shear-rate dependent blood flow model along with resistance boundary conditions for downstream pressure is used to simulate pulsatile blood flow in the arterial system. Adhesion kinetics are simulated through various multiscale models that incorporate experimentally measured parameters in the boundary layer at the target sites [129].
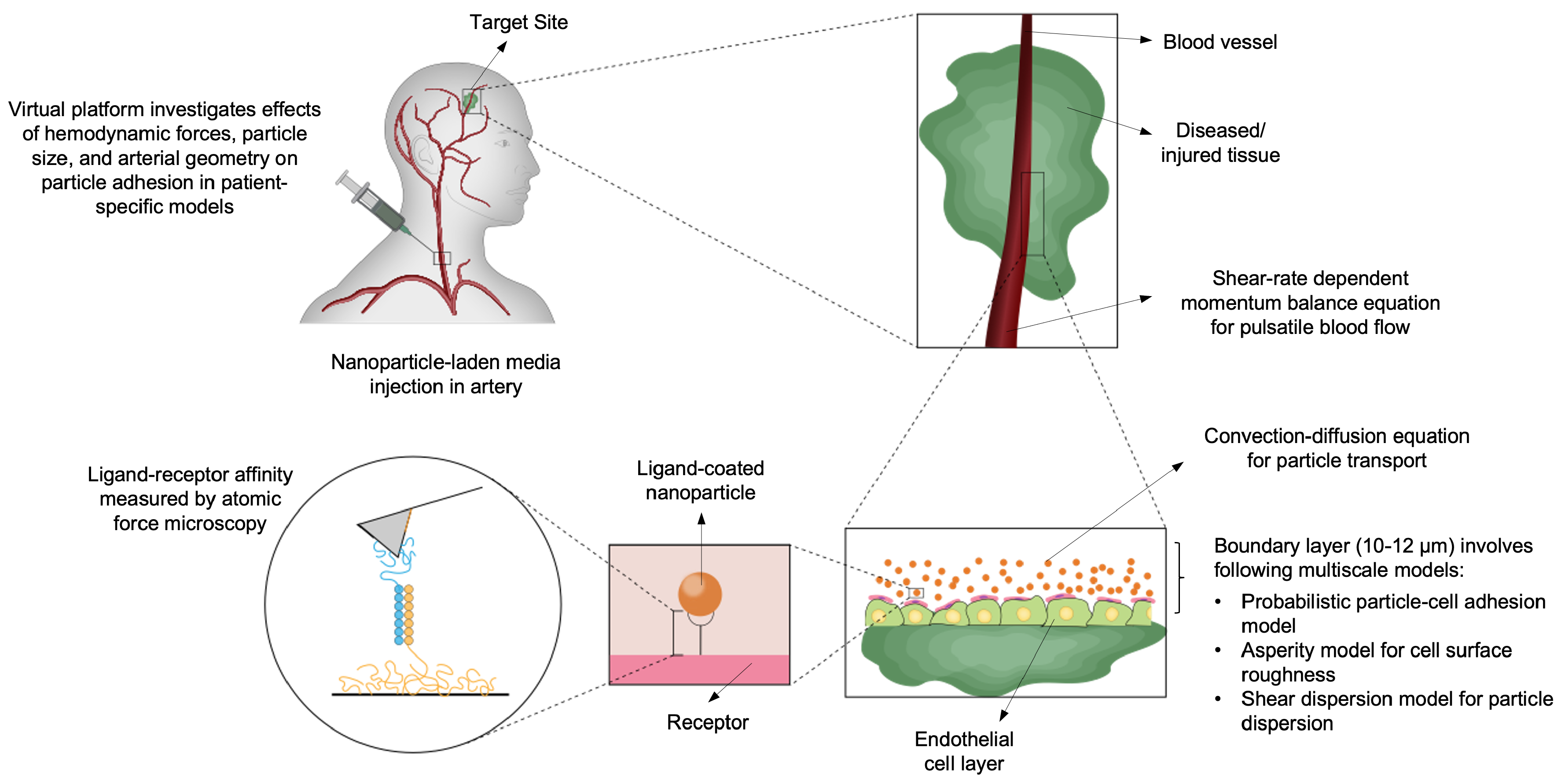
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



