
Submitted:
16 December 2025
Posted:
09 January 2026
You are already at the latest version
Abstract
Results showed the presence of 425 species of macro fungi and 96 families in natural forest, Ascomycetes had 19 families and 41 species, 4 species are edible and 1 medicinal; the Basidiomycetes had 78 families and 384 species, and 50 species are edible, 6 medicinal, 65 toxic, 4 hallucinogenic and 3 bioluminescent. Regarding life forms Ascomycetes had 24 species saprotroph, 13 parasites and 1 mycorrhizal. Basidiomycetes had 229 saprotroph species, 119 mycorrhizal and 34 parasitic. Pure culture growth of 110 species was measured, saprotroph species grew 3.5 cm, mycorrhizal 0.7 cm and parasitic species 0.4 cm at 7 days from incubation. The Kruskall-Wallis analysis showed significant differences in the average growth of the species groups (p< 0.05). A pairwise analysis, after the Kruskall-Wallis, showed that growth of saprotrophs was significantly greater than mycorrhizal and parasitic species; the last two groups were not significantly different. Native edible strains of Pleurotus dejamour and Hericium erinaceus were grown and had statistically significant differences (P<0.05) for fruiting bodies production.
Keywords:
macro fungi
; diversity
; culture
; biotechnology
; edible species
Introduction
Mushrooms play an important role in the forests as they act as decomposer organisms, transforming organic matter in forest ecosystems and helping to convert it into simpler compounds and thus releasing nutrients that are essential for the growth of other microorganisms and plants that transform it and make it available for insects and many species of mammals e.g. rodents [23,36]. Many macro fungi species are linked with trees forming mutualistic symbiotic associations with e.g. oaks and conifers in temperate forests and some are edible and medicinal and are widely distributed in forests of the north of Mexico in the states of Baja California, Sonora, Chihuahua, Durango, Nuevo León, Coahuila, and Tamaulipas [35]. Temperate forests and macro fungi are abundant and diverse in the states of Nuevo León, Coahuila, and Tamaulipas, and many species have been reported as edible e.g. Cantharellus spp., Amanita caesarea group, Tuber lyonii and Boletus edulis [10,12,14,16]. Mexico is a multi-ethnic, multicultural country, and since pre-Columbian times, many ethnic groups have consumed edible wild mushrooms such as Boletus edulis, Boletus pinophilus, Amanita caesarea, Hydnum repandum, Lactarius deliciosus, Cantharellus cibarius and Craterellus cornucupioides [28]. The tradition related to forest mushrooms has been passed down from generation to generation, and during the rainy season, adults teach children how and where to find and harvest edible and medicinal wild mushrooms [20]. People who are already familiar with the taste of different species of edible mushrooms, eagerly await this season so they can enjoy the flavors of the different edible species. In the case of edible fruiting bodies producing fungi, their annual demand is increasing dramatically worldwide as health-conscious consumers seek them as beneficial foods with low carbohydrate and hygienic cultivated products ([1,34]. The nutritional properties of edible mushroom species are well known, as are those with medicinal uses. In the north of the country, only a few studies have been conducted on the diversity of toxic and medicinal macro fungi species, and it is important to understand their diversity and distribution, as well as the hosts with which they are associated [8]. These studies are needed as some species might be used to carry out biomedical research, either with a focus on the treatment of illness e.g. liver or colon cancer, to improve the immune system, or to reduce cholesterol or blood sugar levels [5,7,17,36]. Agro-industrial and forestry residues e.g. coffee husk are often accumulated in large quantities in the field or in sawmills and they become an environmental pollution problem when burned for disposal in different parts of the world [29,30]. Production of fruiting bodies of edible and medicinal mushrooms can be caried out in agricultural residues at low costs and sawmill residues can also be used for these purposes [1,3,30]. In Mexico, mushroom consumption per capita has increased 209% from 1990 to 1997, from 0.112 kg to 0.346 kg, which if compared to approximately 1 kg per capita per year in the United States and more than 2 kg per capita per year in Canada and England, this consumption is low in our country ([30]. In Mexico, fresh mushrooms are preferentially purchased in public markets (67.2%), supermarkets (14.7%), street markets (8.6%), green stores (2.7%), and other locations (6.8%). Most Mexican consumers are not well informed about the nutritional and health benefits of mushrooms, so they are not likely to buy mushrooms compared to other foods considered healthy [30,31]. Recent studies report that the richness of mycorrhizal fungi is unprotected worldwide; despite knowing the benefits they have for the world's vegetation and for obtaining goods and services for man [37]. The alteration of the availability of soil, water and the cycling of mineral nutrients in forests leads to significant changes in the carbon cycle that affect their development and escalate global climate change [21,37]. This research is needed due to the lack of information on diversity, ecology, edibility and biotechnological uses of macro fungi from this region of the country. The results of this study could be of useful for generating information and knowledge about mushroom species diversity, as well as for decision-making in forest management as well as for treatment of intoxication e.g. cases of poisoning from the consumption of wild mushrooms. Strains of some fungal species e.g. Trametes versicolor, Ganoderma spp., collected in the state of Nuevo León have been shown to have medicinal potential and can generate research interest as related to the production of secondary metabolites and their effects, e.g., in controlling liver cancer [7,17]. Other species are of great interest for forestry practices e.g. Pisolithus tinctorius, Laccaria laccata, Astraeus hygrometricus, Tuber lyonii as they benefit trees growth and health, as well as to produce plantations using edible mushrooms for sustainable management of natural resources in northeastern Mexico [19,37], This study aims to contribute to the knowledge of diversity of macro fungi associated to the forests in different altitudinal levels, soils and climates in the state of Nuevo León as well as to isolate pure cultures of the species and to measure their growth in culture as well as to produce fruiting bodies of selected edible and medicinal species from native strains.
Methodology
Collection of Fruiting Bodies
Field trips to the temperate forests located in different municipalities of the state of Nuevo Leon at the Sierra Madre Oriental in altitudes ranging from 550 to 3750 m at the top of Cerro El Potosí were carried out for 35 years. Oaks and conifers forests form either pure or mixed associations, and both genera Quercus and Pinus have a high diversity of species [4,14]. The sampling method for fruiting bodies collection was that used by [27]. The identification of species was carried out with the support of guides like [5,32] and websites like Index Fungorum (www.indexfungorum.org), and for the classification of the species was followed the studies by [26]. Fresh specimens taken to the laboratory were dehydrated at 450C. Sections were made from dehydrated samples, 10% KOH and Congo red were used as staining agents, and microscopic structures such as basidiospores, basidia, cystidia, pileipellis, and stipitipellis were described using an optical microscope. The specimens were deposited in the CFNL mycology herbarium and laboratory collection. This study was carried out in the Department of Silviculture and Management of Natural Resources of the Faculty of Forestry Sciences of the U.A.N.L.
Culture In Vitro
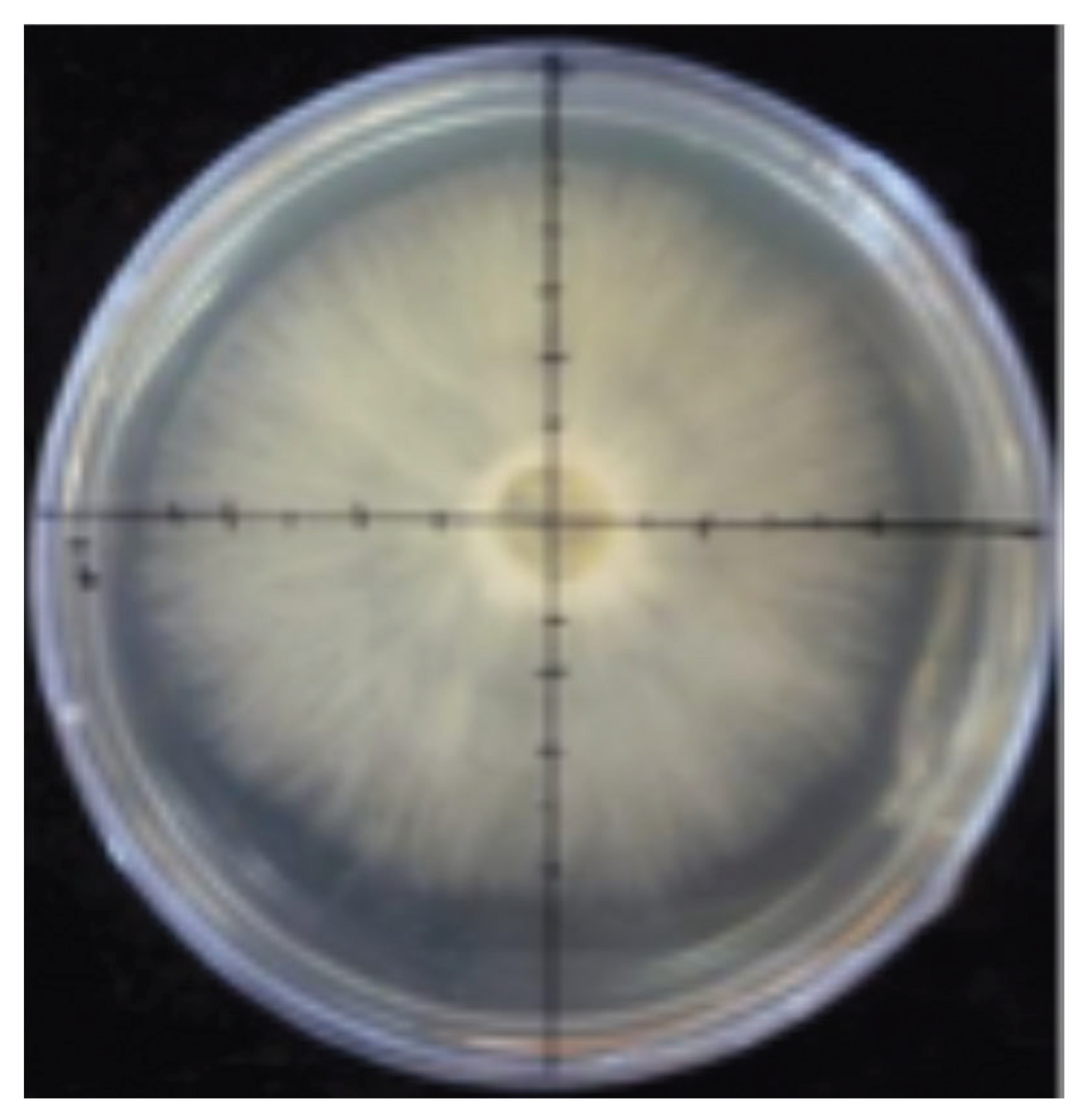
Once in the laboratory, young, fresh specimens of selected saprotrophic, parasitic and mycorrhizal species that were not damaged during the transport and showed no evidence of insect larvae were selected to obtain aseptic cultures in malt extract agar (3%) and modified Melin Norkrans culturing media. With the aseptic cultures obtained, an experiment to measure their average linear growth range in vitro using 6 replicates per species for 7 days was carried out. The daily growth of the 110 strains was measured in four orthogonal lines marked at the base of the Petri dishes and an analysis de variance ANOVA and the Kruskall-Wallis analysis was performed looking for possible differences in the growth rates of saprotrophic, mycorrhizal and parasitic species.
Production of Pleurotus dejamour and Hericium erinaceus in Residual Substrates
The strains of the species used in this study were obtained from wild fruiting bodies collected by the main author in forests in the state of Nuevo Leon. Six 6 strains of Hericium erinaceus and 8 strains of Pleurotus dejamour were selected and investigated. They were cultivated under laboratory and greenhouse conditions to evaluate their productivity using agricultural residues.
Strains
The species were identified in the laboratory using routine techniques and specialized keys. The strains were isolated in pure culture on malt extract agar (3%) added with streptomycin 120 ppm, to eliminate contaminants.
Preparation of Inoculum in Agar
Strains of both species were grown on malt extract agar in vitro. A 1 cm disk of mycelium growing on agar was used for each replicate. Petri dishes were incubated at 25°C and 6 replicates per strain were used. The daily growth of the strains was measured for 7 days in four orthogonal lines marked at the base of the Petri dishes. The values of developmental efficiency over time were treated by ANOVA analysis and Tukey's test (a=0.05%) was applied to determine the existence of significant differences between the development of the strains.
Preparation of Inoculant Seed Treatments
Cultures with active growth were used to inoculate the seed substrates used as inoculant (treatment 1: corn, wheat, sorghum, sunflower seed; treatment 2: canary seed, millet, turnip, linseed; and treatment 3: wheat). The grain mixtures were cleaned and weighed in 90 g portions, then hydrated for 48 h in 250 ml bottles. Excess water was removed from the jars by draining once the seeds were hydrated. The flasks with the seeds were sterilized at 120ºC for 30 min. Subsequently, the flasks were left to cool for 24 hours and under aseptic conditions were inoculated with 2 cm of mycelium from the aseptic cultures in Petri dish for each strain and species. The inoculated flasks were incubated at a temperature of 25ºC for two weeks and they were used as inoculant.
Figure 1.
Map showing the Great folded Sierra (in dark green) where temperate forests are distributed in the state of Nuevo Leon.
Figure 1.
Map showing the Great folded Sierra (in dark green) where temperate forests are distributed in the state of Nuevo Leon.
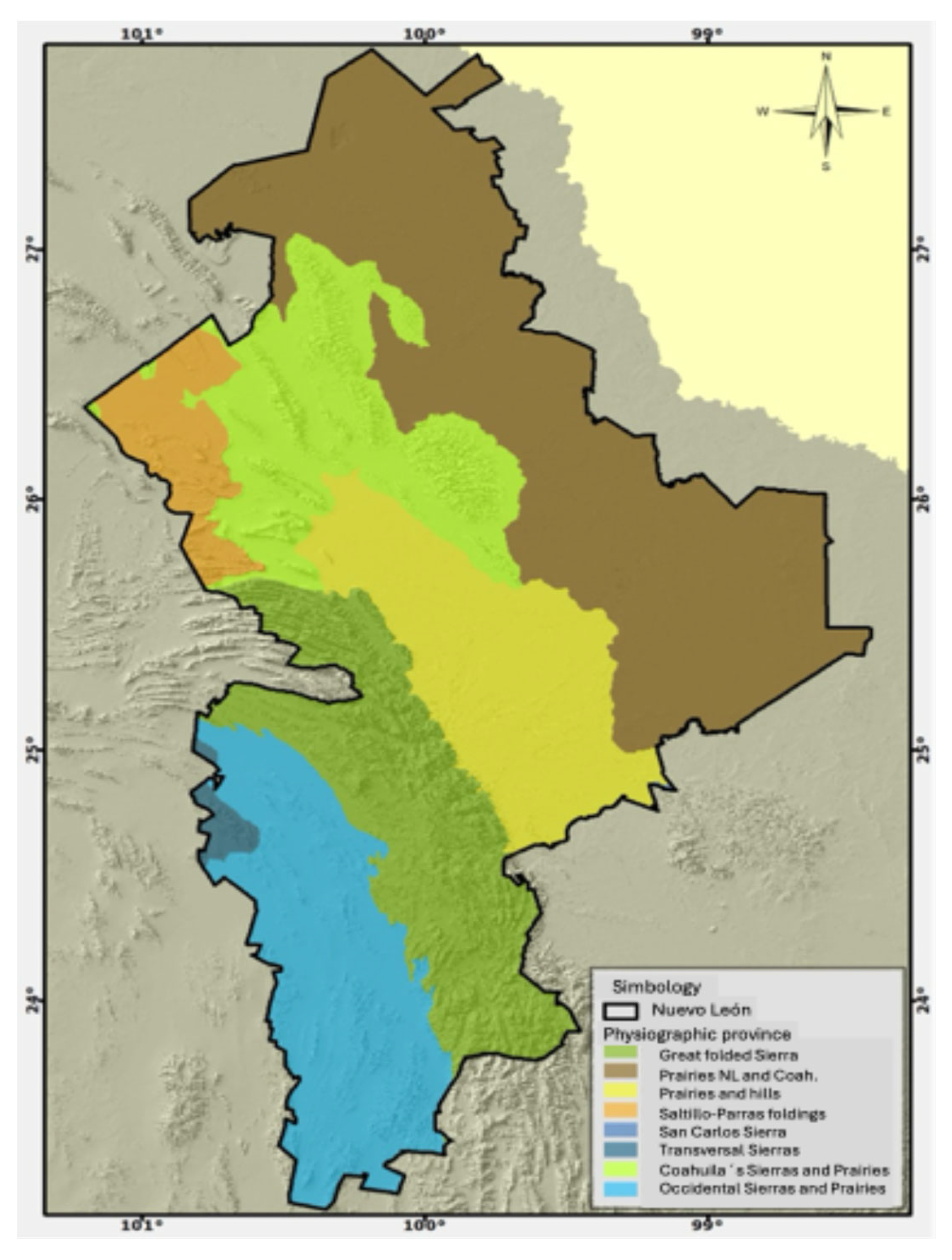
Figure 2.
Linear growth measurement of mycelium in vitro.
Substrates for Fungal Growth
The substrates selected for experimentation in relation to fruiting bodies production of the fungal species under study were: 1) Coffee husk, 2) Sorghum harvest, 3) Mixture of straw from different wild grass species. The substrates were ground to a size of approximately 5 cm, placed in plastic containers to moisten them for 48 hours, and then used to fill nylon bags with a capacity of 3 kg, with a weight of 500 g (wet weight). The bags used were sterilized at 120°C for 30 minutes to eliminate the microorganisms and were allowed to cool prior to inoculation with the seed inoculant treatments.
Inoculation and Incubation on Final Substrate
The mycelium obtained in the flasks with the seed mixtures was used to inoculate the bags containing the pasteurized substrate. The inoculation of the polyethylene bags was carried out by adding 3% of the inoculants obtained in seeds per kg of substrate and 5 gr. of calcium carbonate was used per bag to lower pH. At the end of the inoculation, the bags were closed and incubated at a temperature of 25ºC. Once the mycelium had developed covering all the substrate, perforations were made homogeneously distributed with a sterile pocketknife on the surface of each polyethylene bag sown.
Figure 3.
Oak, Pine-Oak and pine forests located from 1600 - 3750 m.

Results
The results of this study showed the presence of 425 species of macro fungi belonging to 96 families both in the Basidiomycetes and Ascomycetes. Ascomycetes had 19 families with 41 species, 4 species have been reported as edible and 1 as medicinal. The Basidiomycetes had 78 families and 384 species, and 50 species have been reported as edible and 6 as medicinal and 65 as toxic, 4 as hallucinogenic and 3 as bioluminescent Omphalotus olivascens, Omphalotus subilludens and Panellus stipticus all the other species are unknown. Regarding life forms 24 species of Ascomycetes are saprotroph 13 parasites and 1 mycorrhizal and for Basidiomycetes 229 species are saprotroph and 119 are mycorrhizal and 34 are parasitic.

Table 1.
Macro fungi diversity in oak and pine forests of Nuevo Leon.
| SPECIES | FAMILY | EDIBILITY | LIFE FORM |
|---|---|---|---|
| Ascomycetes | |||
| Chlorociboria aeruginascens (Nyl.) Kanouse. | Chlorociboriaceae | S | |
| Calycina citrina (Hedw.) Gray. | Pezizellaceae | S | |
| Bulgaria inquinans (Pers.) Fr. | Phacidiaceae | S | |
| Leotia lubrica (Scop.) Pers. | Leotiaceae | S | |
| Cudonia circinans (Pers.) Fr. | Cudoniaceae | S | |
| Spathularia flavida Pers. | Cudoniaceae | S | |
| Gyromitra esculenta Pers. ex Fr. | Discinaceae | E | S |
| Paragyromitra infula (Schaeff.) X.C. Wang & W.Y. Zhuang. | Discinaceae | S | |
| Helvella acetabulum (L.) Quél. | Helvellaceae | S | |
| Helvella dryophila Vellinga & N.H. Nguye | Helvellaceae | S | |
| Helvella elastica Bull. | Helvellaceae | S | |
| Helvella lacunosa Fr. | Helvellaceae | S | |
| Helvella macropus (Pers.) P. Karst. | Helvellaceae | S | |
| Morchella conica Krombh. | Morchellaceae | E | M |
| Morchella sp. | Morchellaceae | M | |
| Otidea onotica (Pers.) Fuckel. | Otideaceae | S | |
| Peziza vesiculosa Pers. | Pezizaceae | S | |
| Sarcosphaera coronaria (Jacq.) J. Schröt. | Pezizaceae | S | |
| Aleuria aurantia (Pers.) Fuckel. | Pyronemataceae | S | |
| Humaria hemisphaerica (F.H. Wigg.) Fuckel. | Pyronemataceae | S | |
| Scutellinia scutellata (L.) Lambotte. | Pyronemataceae | S | |
| Phillipsia sp. | Sarcoscyphaceae | S | |
| Phillipsia domingensis (Berk.) Berk. ex Denison. | Sarcoscyphaceae | S | |
| Sarcoscypha coccinea (Scop.) Sacc. ex E.J. Durand. | Sarcoscyphaceae | S | |
| Sarcoscypha occidentalis (Schwein.) Sacc. | Sarcoscyphaceae | S | |
| Cordyceps militaris (L.) Fr. | Cordycipitaceae | M | P |
| Hypomyces chrysospermus Tul. & C. Tul. | Hypocreaceae | P | |
| Hypomyces hyalinus (Schwein.) Tul. & C. Tul. | Hypocreaceae | P | |
| Hypomyces lactifluorum (Schwein.) Tul. & C. Tul. | Hypocreaceae | E | P |
| Hypomyces sp. | Hypocreaceae | P | |
| Tolypocladium capitatum (Holmsk.) C.A. Quandt, Kepler & Spatafora. | Ophiocordycipitaceae | P | |
| Biscogniauxia atropunctata (Schwein.) Pouzar. | Graphostromataceae | P | |
| Annulohypoxylon thouarsianum (Lév.) Y.M. Ju, J.D. Rogers & H.M. Hsieh. | Hypoxylaceae | P | |
| Daldinia childiae J.D. Rogers & Y.M. Ju. | Hypoxylaceae | ||
| Hypoxylon fragiforme (Pers.) J. Kickx f. | Hypoxylaceae | ||
| Ustulina deusta (Hoffm.) Maire | Xylariaceae | P | |
| Xylaria hypoxylon (L.) Grev. | Xylariaceae | P | |
| Xylosphaera poitei (Lév.) Dennis. | Xylariaceae | P | |
| Xylaria polymorpha (Pers.) Grev. | Xylariaceae | P | |
| Xylaria titan Berk. & M.A. Curtis. | Xylariaceae | P | |
| Trichoglossum hirsutum (Pers.) Boud. | Geoglossaceae | S | |
| Basidiomycetes | |||
| Dacrymyces chrysospermus Berk. & M.A. Curtis. | Dacrymycetaceae | S | |
| Dacryopinax spathularia (Schwein.) G.W. Martin. | Dacrymycetaceae | S | |
| Myxarium nucleatum Wallr. | Hyaloriaceae | S | |
| Phaeotremella foliacea (Pers.) Wedin, J.C. Zamora & Millanes. | Tremellaceae | S | |
| Tremella fuciformis Berk. | Tremellaceae | S | |
| Tremella mesenterica (Schaeff.) Pers. | Tremellaceae | S | |
| Asproinocybe sp. | Agaricaceae | M | |
| Baeospora myosura (Fr.) Singer. | Agaricaceae | S | |
| Clitocybe fragrans (With.) P. Kumm. | Agaricaceae | S | |
| Clitocybe sordida Velen. | Agaricaceae | S | |
| Crucibulum laeve (Huds.) Kambly. | Agaricaceae | S | |
| Cyathus olla (Batsch) Pers. | Agaricaceae | S | |
| Cyathus stercoreus (Schwein.) De Toni. | Agaricaceae | S | |
| Cyathus striatus Willd. | Agaricaceae | S | |
| Cystodermella cinnabarina (Alb. & Schwein.) Harmaja. | Agaricaceae | S | |
| Cystoderma amianthinum (Scop.) Fayod. | Agaricaceae | S | |
| Cystodermella granulosa (Batsch) Harmaja. | Agaricaceae | S | |
| Gerronema strombodes (Berk. & Mont.) Singer. | Agaricaceae | S | |
| Infundibulicybe gibba (Pers.) Harmaja. | Agaricaceae | S | |
| Lepista nuda (Bull.) Cooke. | Agaricaceae | S | |
| Leucopholiota decorosa (Peck) O.K. Mill., T.J. Volk & Bessette. | Agaricaceae | S | |
| Melanoleuca cognata (Fr.) Konrad & Maubl. | Agaricaceae | S | |
| Melanoleuca dryophila Murrill. | Agaricaceae | S | |
| Nidula candida (Peck) V.S. White. | Agaricaceae | S | |
| Panaeolina foenisecii (Pers.) Maire. | Agaricaceae | S | |
| Pleurocybella porrigens (Pers.) Singer. | Agaricaceae | S | |
| Tricholomopsis decora (Fr.) Singer. | Agaricales | M | |
| Tricholomopsis rutilans (Schaeff.) Singer. | Agaricales | S | |
| Agaricus arvensis Schaeff. | Agaricaceae | E | S |
| Agaricus augustus Fr. | Agaricaceae | E | S |
| Agaricus campestris L. | Agaricaceae | E | S |
| Agaricus cantharellus Batsch. | Agaricaceae | S | |
| Agaricus hondensis Murrill. | Agaricaceae | S | |
| Agaricus placomyces Peck. | Agaricaceae | S | |
| Agaricus xanthodermus Genev. | Agaricaceae | T | S |
| Battarrea phalloides (Dicks.) Pers. | Agaricaceae | S | |
| Chlorophyllum molybdites (G. Mey.) Massee. | Agaricaceae | T | S |
| Chlorophyllum olivieri (Barla) Vellinga. | Agaricaceae | T | S |
| Chlorophyllum rhacodes (Vittad.) Vellinga. | Agaricaceae | T | S |
| Coprinus comatus (O.F. Müll.) Pers. | Agaricaceae | S | |
| Lepiota cristata (Bolton) P. Kumm. | Agaricaceae | T | S |
| Leucoagaricus leucothites (Vittad.) Wasser. | Agaricaceae | T | S |
| Leucocoprinus birnbaumii (Corda) Singer | Agaricaceae | T | S |
| Leucocoprinus cepistipes (Sowerby) Pat. | Agaricaceae | T | S |
| Leucocoprinus fragilissimus (Ravenel ex Berk. & M.A. Curtis) Pat. | Agaricaceae | S | |
| Leucocoprinus rubrotinctus (Peck) Redhead. | Agaricaceae | S | |
| Macrolepiota procera (Scop.) Singer. | Agaricaceae | E | S |
| Montagnea arenaria (DC.) Zeller. | Agaricaceae | S | |
| Phellorinia herculeana (Pers.) Kreisel. | Agaricaceae | S | |
| Podaxis pistillaris (L.) Fr. | Agaricaceae | E | S |
| Tulostoma pulchellum Sacc. | Agaricaceae | S | |
| T. mohavensis | Agaricaceae | S | |
| Amanita amerivirosa Tulloss, L.V. Kudzma & M. Tulloss. | Amanitaceae | T | M |
| Amanita bisporigera G.F. Atk. | Amanitaceae | T | M |
| Amanita calyptroderma G.F. Atk. & V.G. Ballen. | Amanitaceae | E | M |
| Amanita flavoconia G.F. Atk. | Amanitaceae | T | M |
| Amanita flavorubescens G.F. Atk. | Amanitaceae | T | M |
| Amanita fulva Fr. | Amanitaceae | T | M |
| Amanita jacksonii Pomerl. | Amanitaceae | E | M |
| Amanita magniverrucata Thiers & Ammirati | Amanitaceae | T | M |
| Amanita muscaria (L.) Lam. | Amanitaceae | T | M |
| Amanita novinupta Tulloss & J. Lindgr. | Amanitaceae | T | M |
| Amanita pantherina (DC.) Krombh. | Amanitaceae | T | M |
| Amanita phalloides (Vaill. ex Fr.) Link. | Amanitaceae | T | M |
| Amanita rubescens Pers. | Amanitaceae | T | M |
| Amanita smithiana Bas. | Amanitaceae | T | M |
| Amanita spreta (Peck) Sacc. | Amanitaceae | T | M |
| Amanita vaginata (Bull.) Lam. | Amanitaceae | T | M |
| Amanita velosa (Peck) Lloyd. | Amanitaceae | T | M |
| Bolbitius titubans (Bull.) Fr. | Bolbitiaceae | T | S |
| Conocybe apala (Fr.) Arnolds. | Bolbitiaceae | T | S |
| Conocybe tenera (Schaeff.) Kühner. | Bolbitiaceae | T | S |
| Clavaria fragilis Holmsk. | Clavariaceae | S | |
| Clavulinopsis fusiformis (Sowerby) Corner. | Clavariaceae | S | |
| Ramariopsis kunzei (Fr.) Corner. | Clavariaceae | S | |
| Collybia nuda (Bull.) Z.M. He & Zhu L. Yang | Clitocybaceae | S | |
| Cortinarius armillatus (Fr.) Fr. | Cortinariaceae | T | M |
| Cortinarius caperatus (Pers.) Fr. | Cortinariaceae | T | M |
| Cortinarius cinnamomeus (L.) Gray. | Cortinariaceae | T | M |
| Cortinarius corrugatus Peck. | Cortinariaceae | T | M |
| Cortinarius iodes Berk. & M.A. Curtis. | Cortinariaceae | T | M |
| Cortinarius semisanguineus (Fr.) Gillet. | Cortinariaceae | T | M |
| Cortinarius traganus (Fr.) Fr. | Cortinariaceae | T | M |
| Cortinarius violaceus (L.) Gray. | Cortinariaceae | T | M |
| Phlegmacium glaucopus (Schaeff.) Wünsche. | Cortinariaceae | T | M |
| Crepidotus crocophyllus (Berk.) Sacc. | Crepidotaceae | S | |
| Crepidotus mollis (Schaeff.) Staude. | Crepidotaceae | S | |
| Chondrostereum purpureum (Pers.) Pouzar. | Cyphellaceae | S | |
| Clitopilus prunulus (Scop.) P. Kumm. | Entolomataceae | S | |
| Entoloma sp. | Entolomataceae | S | |
| Entoloma sericeum Quél. | Entolomataceae | S | |
| Entoloma rhodopolium (Fr.) P. Kumm. | Entolomataceae | S | |
| Entoloma velutinum Hesler. | Entolomataceae | S | |
| Panaeolus antillarum (Fr.) Dennis. | Galeropsidaceae | S | |
| Panaeolus cinctulus (Bolton) Sacc. | Galeropsidaceae | S | |
| Panaeolus papilionaceus (Bull.) Quél. | Galeropsidaceae | S | |
| Laccaria sp. | Hydnangiaceae | M | |
| Laccaria amethystina Cooke. | Hydnangiaceae | M | |
| Laccaria bicolor (Maire) P.D. Orton. | Hydnangiaceae | E | M |
| Laccaria laccata (Scop.) Cooke. | Hydnangiaceae | E | M |
| Laccaria ochropurpurea (Berk.) Peck. | Hydnangiaceae | M | |
| Arrhenia sp. | Hygrophoraceae | T | S |
| Arrhenia epichysium (Pers.) Redhead, Lutzoni, Moncalvo & Vilgalys. | Hygrophoraceae | T | S |
| Cuphophyllus colemannianus (A. Bloxam) Bon. | Hygrophoraceae | S | |
| Cuphophyllus pratensis (Pers.) Bon. | Hygrophoraceae | S | |
| Cuphophyllus virgineus (Wulfen) Kovalenko. | Hygrophoraceae | S | |
| Gliophorus psittacinus (Schaeff.) Herink. | Hygrophoraceae | S | |
| Hygrocybe cantharellus (Schwein.) Murrill. | Hygrophoraceae | S | |
| Hygrocybe coccinea (Schaeff.) P. Kumm. | Hygrophoraceae | S | |
| Hygrocybe conica (Schaeff.) P. Kumm. | Hygrophoraceae | S | |
| Hygrocybe flavescens (Kauffman) Singer. | Hygrophoraceae | S | |
| Hygrocybe miniata (Fr.) P. Kumm. | Hygrophoraceae | S | |
| Hygrophorus chrysodon (Batsch) Fr. | Hygrophoraceae | S | |
| Hygrophorus eburneus (Bull.) Fr. | Hygrophoraceae | S | |
| Hygrophorus erubescens (Fr.) Fr. | Hygrophoraceae | S | |
| Hygrophorus pudorinus (Fr.) Fr. | Hygrophoraceae | S | |
| Hygrophorus russula (Schaeff. ex Fr.) Bataille | Hygrophoraceae | S | |
| Lichenomphalia umbellifera (L.) Redhead, Lutzoni, Moncalvo & Vilgalys, | Hygrophoraceae | S | |
| Galerina marginata (Batsch) Kühner. | Hymenogastraceae | T | S |
| Gymnopilus luteofolius (Peck) Singer. | Hymenogastraceae | H | S |
| Gymnopilus sapineus (Fr.) Murrill. | Hymenogastraceae | S | |
| Psilocybe coronilla (Bull.) Noordel. | Hymenogastraceae | H | S |
| Psilocybe cyanescens Wakef. | Hymenogastraceae | H | S |
| Psilocybe semilanceata (Fr.) P. Kumm. | Hymenogastraceae | H | S |
| Inosperma calamistratum (Fr.) Matheny & Esteve-Rav. | Inocybaceae | T | M |
| Inocybe geophylla P. Kumm. | Inocybaceae | T | M |
| Inocybe hystrix (Fr.) P. Karst. | Inocybaceae | T | M |
| Pseudosperma sororium (Kauffman) Matheny & Esteve-Rav. | Inocybaceae | T | M |
| Apioperdon pyriforme (Schaeff.) Vizzini. | Lycoperdaceae | S | |
| Bovista plumbea Pers. | Lycoperdaceae | S | |
| Calbovista subsculpta Morse ex M.T. Seidl. | Lycoperdaceae | S | |
| Calvatia cyathiformis (Bosc) Morgan. | Lycoperdaceae | E | S |
| Lycoperdon marginatum Vittad. | Lycoperdaceae | S | |
| Lycoperdon perlatum Pers. | Lycoperdaceae | S | |
| Utraria pulcherrima (Berk. & M.A. Curtis) R.L. Zhao & J.Xin Li | Lycoperdaceae | S | |
| Utraria umbrina (Pers.) R.L. Zhao & J.Xin Li | Lycoperdaceae | S | |
| Hypsizygus ulmarius (Bull.) Redhead. | Lyophyllaceae | E | S |
| Lyophyllum decastes (Fr.) Singer. | Lyophyllaceae | E | S |
| Macrocystidia cucumis (Pers.) Joss. | Macrocystidiaceae | S | |
| Marasmius plicatulus Peck. | Marasmiaceae | S | |
| Marasmius rotula (Scop.) Fr. | Marasmiaceae | S | |
| Marasmius siccus (Schwein.) Fr. | Marasmiaceae | S | |
| Tetrapyrgos nigripes (Fr.) E. Horak. | Marasmiaceae | S | |
| Favolaschia sp. | Mycenaceae | S | |
| Hemimycena lactea (Pers.) Singer. | Mycenaceae | S | |
| Mycena acicula (Schaeff.) P. Kumm. | Mycenaceae | S | |
| Mycena amicta (Fr.) Quél. | Mycenaceae | S | |
| Mycena epipterygia (Scop.) Gray. | Mycenaceae | S | |
| Mycena galericulata (Scop.) Gray. | Mycenaceae | S | |
| Mycena leptocephala (Pers.) Gillet. | Mycenaceae | S | |
| Mycena pura (Pers.) P. Kumm. | Mycenaceae | S | |
| Panellus stipticus (Bull.) P. Karst. | Mycenaceae | T, BIO | S |
| Xeromphalina campanella (Batsch) Kühner & Maire. | Mycenaceae | S | |
| Xeromphalina cauticinalis (Fr.) Kühner & Maire. | Mycenaceae | S | |
| Xeromphalina fulvipes (Murrill) A.H. Sm. | Mycenaceae | S | |
| Marasmiellus candidus (Fr.) Singer. | Omphalotaceae | S | |
| Collybiopsis quercophila (Pouzar) R.H. Petersen. | Omphalotaceae | S | |
| Collybiopsis subnuda (Ellis ex Peck) R.H. Petersen. | Omphalotaceae | S | |
| Gymnopus dryophilus (Bull.) Murrill. | Omphalotaceae | S | |
| Mycetinis opacus (Berk. & M.A. Curtis) A.W. Wilson & Desjardin. | Omphalotaceae | S | |
| Omphalotus olivascens H.E. Bigelow, O.K. Mill. & Thiers. | Omphalotaceae | T | P |
| Omphalotus subilludens (Murrill) H.E. Bigelow. | Omphalotaceae | S | |
| Rhodocollybia butyracea (Bull.) Lennox. | Omphalotaceae | S | |
| Rhodocollybia maculata (Alb. & Schwein.) Singer. | Omphalotaceae | S | |
| Phyllotopsis nidulans (Pers.) Singer. | Phyllotopsidaceae | T | S |
| Armillaria mellea (Vahl) P. Kumm. | Physalacriaceae | E | P |
| Desarmillaria tabescens (Scop.) R.A. Koch & Aime. | Physalacriaceae | E, T | P |
| Cyptotrama asprata (Berk.) Redhead & Ginns. | Physalacriaceae | S | |
| Flammulina velutipes (Curtis) Singer. | Physalacriaceae | E | S |
| Hymenopellis furfuracea (Peck) R.H. Petersen. | Physalacriaceae | S | |
| Hymenopellis radicata (Relhan) R.H. Petersen. | Physalacriaceae | S | |
| Strobilurus conigenoides (Ellis) Singer. | Physalacriaceae | S | |
| Strobilurus trullisatus (Murrill) Lennox. | Physalacriaceae | S | |
| Hohenbuehelia angustata (Berk.) Singer. | Pleurotaceae | S | |
| Hohenbuehelia petaloides (Bull.) Schulzer. | Pleurotaceae | S | |
| Pleurotus cornucopiae (Paulet) Quél. | Pleurotaceae | E | S |
| Pleurotus djamor (Rumph. ex Fr.) Boedijn. | Pleurotaceae | E | S |
| Pleurotus dryinus (Pers.) P. Kumm. | Pleurotaceae | E | S |
| Resupinatus alboniger (Pat.) Singer. | Pleurotaceae | S | |
| Resupinatus applicatus (Batsch) Gray. | Pleurotaceae | S | |
| Pluteus americanus (P. Banerjee & Sundb.) Justo, E.F. Malysheva & Minnis. | Pluteaceae | S | |
| Pluteus cervinus (Schaeff.) P. Kumm. | Pluteaceae | S | |
| Pluteus chrysophlebius (Berk. & M.A. Curtis) Sacc. | Pluteaceae | S | |
| Pluteus petasatus (Fr.) Gillet. | Pluteaceae | S | |
| Volvariella bombycina (Schaeff.) Singer. | Pluteaceae | E | S |
| Volvopluteus gloiocephalus (DC.) Vizzini, Contu & Justo. | Pluteaceae | S | |
| Candolleomyces candolleanus (Fr.) D. Wächt. & A. Melzer. | Psathyrellaceae | S | |
| Coprinellus disseminatus (Pers.) J.E. Lange. | Psathyrellaceae | S | |
| Coprinellus domesticus (Bolton) Vilgalys, Hopple & Jacq. Johnson. | Psathyrellaceae | S | |
| Coprinellus micaceus (Bull.) Vilgalys, Hopple & Jacq. Johnson. | Psathyrellaceae | S | |
| Coprinopsis atramentaria (Bull.) Redhead, Vilgalys & Moncalvo. | Psathyrellaceae | S | |
| Coprinopsis lagopides (P. Karst.) Redhead, Vilgalys & Moncalvo. | Psathyrellaceae | S | |
| Coprinopsis lagopus (Fr.) Redhead, Vilgalys & Moncalvo. | Psathyrellaceae | S | |
| Parasola plicatilis (Curtis) Redhead, Vilgalys & Hopple. | Psathyrellaceae | S | |
| Psathyrella bipellis (Quél.) A.H. Sm. | Psathyrellaceae | S | |
| Psathyrella piluliformis (Bull.) P.D. Orton. | Psathyrellaceae | S | |
| Radulomyces sp. | Radulomycetaceae | S | |
| Schizophyllum commune Fr. | Schizophyllaceae | NR | S, P |
| Schizophyllum fasciatum Pat. | Schizophyllaceae | NR | S, P |
| Schizophyllum umbrinum Berk. | Schizophyllaceae | NR | S, P |
| Deconica coprophila (Bull.) P. Karst. | Strophariaceae | T | S |
| Deconica montana (Pers.) P.D. Orton. | Strophariaceae | T | S |
| Hypholoma capnoides (Fr.) P. Kumm. | Strophariaceae | T | S |
| Hypholoma fasciculare (Huds.) P. Kumm. | Strophariaceae | T | S |
| Hypholoma lateritium (Schaeff.) P. Kumm. | Strophariaceae | T | S |
| Pholiota adiposa (Batsch) P. Kumm. | Strophariaceae | T | S |
| Pholiota squarrosa (Vahl) P. Kumm. | Strophariaceae | T | S |
| Pholiota squarrosoides (Peck) Sacc. | Strophariaceae | T | S |
| Protostropharia semiglobata (Batsch) Redhead, Moncalvo & Vilgalys. | Strophariaceae | T | S |
| Stropharia coronilla (Bull.) Quél.. | Strophariaceae | H | S |
| Tapinella atrotomentosa (Batsch) Šutara. | Tapinellaceae | T | S |
| Tapinella panuoides (Fr.) E.-J. Gilbert. | Tapinellaceae | T | S |
| Leucopaxillus albissimus (Peck) Singer. | Tricholomataceae | T | S |
| Leucopaxillus gentianeus (Quél.) Kotl. | Tricholomataceae | T | S |
| Tricholoma caligatum (Viv.) Ricken. | Tricholomataceae | E | M |
| Tricholoma equestre (L.) P. Kumm. | Tricholomataceae | M | |
| Tricholoma murrillianum Singer. | Tricholomataceae | M | |
| Tricholoma sejunctum (Sowerby) Quél. | Tricholomataceae | M | |
| Tricholoma terreum (Schaeff.) P. Kumm. | Tricholomataceae | M | |
| Ductifera pululahuana (Pat.) Donk. | Auriculariaceae | S | |
| Pseudohydnum gelatinosum (Scop.) P. Karst. | Auriculariaceae | S | |
| Baorangia bicolor (Kuntze) G. Wu, Halling & Zhu L. Yang. | Boletaceae | M | |
| Boletellus ananas (M.A. Curtis) Murrill. | Boletaceae | M | |
| Boletus barrowsii Thiers & A.H. Sm. | Boletaceae | E | M |
| Boletus edulis Bull. | Boletaceae | E | M |
| Neoboletus subvelutipes (Peck) Yang Wang, B. Zhang & Yu Li | Boletaceae | M | |
| Exsudoporus frostii (J.L. Russell) Vizzini, Simonini & Gelardi. | Boletaceae | E | M |
| Cyanoboletus pulverulentus (Opat.) Gelardi, Vizzini & Simonini. | Boletaceae | M | |
| Harrya chromipes (Frost) Halling, Nuhn, Osmundson & Manfr. Binder. | Boletaceae | E | M |
| Hortiboletus campestris (A.H. Sm. & Thiers) Biketova & Wasser. | Boletaceae | M | |
| Imleria badia (Fr.) Vizzini. | Boletaceae | M | |
| Leccinellum albellum (Peck) Bresinsky & Manfr. Binder. | Boletaceae | M | |
| Leccinum manzanitae Thiers. | Boletaceae | E | M |
| Leccinum aeneum Halling. | Boletaceae | M | |
| Phylloporus rhodoxanthus (Schwein.) Bres. | Boletaceae | S | |
| Porphyrellus cyaneotinctus (A.H. SM. & Thiers) Singer | Boletaceae | M | |
| Pulchroboletus rubricitrinus (Murrill) Farid & A.R. Franck. | Boletaceae | M | |
| Pulveroboletus ravenelii (Berk. & M.A. Curtis) Murrill. | Boletaceae | M | |
| Retiboletus griseus (Frost) Manfr. Binder & Bresinsky. | Boletaceae | M | |
| Rubroboletus eastwoodiae (Murrill) Vasquez, Simonini, Svetash., Mikšík & Vizzini. | Boletaceae | M | |
| Strobilomyces strobilaceus (Scop.) Berk. | Boletaceae | M | |
| Sutorius eximius (Peck) Halling, Nuhn & Osmundson. | Boletaceae | M | |
| Tylopilus felleus (Bull.) P. Karst. | Boletaceae | T | M |
| Tylopilus plumbeoviolaceus (Snell & E.A. Dick) Snell & E.A. Dick. | Boletaceae | M | |
| Tylopilus tabacinus (Peck) Singer. | Boletaceae | M | |
| Xerocomellus carmeniae Garza-Ocañas, J. García & de la Fuente. | Boletaceae | M | |
| Xerocomellus diffractus N. Siegel, C.F. Schwarz & J.L. Frank. | Boletaceae | M | |
| Xerocomus subtomentosus (L.) Quél. | Boletaceae | M | |
| Boletinellus merulioides (Schwein.) Murrill. | Boletinellaceae | E | S,M |
| Boletinellus rompelii (Pat. & Rick) Watling | Boletinellaceae | E | S,M |
| Calostoma cinnabarinum Desv. | Calostomataceae | S | |
| Astraeus hygrometricus (Pers.) Morgan. | Diplocystidiaceae | M | |
| Astraeus pteridis (Shear) Zeller. | Diplocystidiaceae | M | |
| Gomphidius roseus (Fr.) Oudem. | Gomphidiaceae | M | |
| Gyroporus castaneus (Bull.) Quél. | Gyroporaceae | E | M |
| Hygrophoropsis aurantiaca (Wulfen) Maire ex Martin-Sans. | Hygrophoropsidaceae | S | |
| Melanogaster variegatus (Vittad.) Tul. & C. Tul. | Paxillaceae | E | M |
| Rhizopogon sp. | Rhizopogonaceae | M | |
| Rhizopogon occidentalis Zeller & C.W. Dodge. | Rhizopogonaceae | E | M |
| Pisolithus arhizus (Scop.) Rauschert. | Sclerodermataceae | M | |
| Scleroderma areolatum Ehrenb. | Sclerodermataceae | T | M |
| Scleroderma cepa Pers. | Sclerodermataceae | T | M |
| Scleroderma citrinum Pers. | Sclerodermataceae | T | M |
| Scleroderma texense Berk. | Sclerodermataceae | T | M |
| Scleroderma verrucosum (Bull.) Pers. | Sclerodermataceae | T | M |
| Serpula himantioides (Fr.) P. Karst. | Serpulaceae | P | |
| Suillus americanus (Peck) Snell. | Suillaceae | M | |
| Suillus lakei (Murrill) A.H. Sm. & Thiers. | Suillaceae | M | |
| Suillus spraguei (Berk. & M.A. Curtis) Kuntze. | Suillaceae | M | |
| Suillus tomentosus Singer, Snell & E.A. Dick. | Suillaceae | E | M |
| Cantharellus cibarius Fr. | Hydnaceae | E | M |
| Cantharellus cinnabarinus (Schwein.) Schwein. | Hydnaceae | E | M |
| Cantharellus lateritius (Berk.) Singer. | Hydnaceae | E | M |
| Cantharellus minor Peck. | Hydnaceae | E | M |
| Craterellus cornucopioides (L.) Pers. | Hydnaceae | E | M |
| Craterellus ignicolor (R.H. Petersen) Dahlman, Danell & Spatafora. | Hydnaceae | E | M |
| Hericium erinaceus (Bull) Pers. | Hericiaceae | E | S,M |
| Hydnum albidum Peck. | Hydnaceae | E | M |
| Hydnum repandum L. | Hydnaceae | E | M |
| Geastrum floriforme Vittad. | Geastraceae | S | |
| Geastrum minimum Schwein. | Geastraceae | S | |
| Myriostoma coliforme (Dicks.) Corda. | Geastraceae | S | |
| Gloeophyllum sp. | Gloeophyllaceae | P | |
| Gloeophyllum sepiarium (Wulfen) P. Karst. | Gloeophyllaceae | P | |
| Heliocybe sulcata (Berk.) Redhead & Ginns. | Gloeophyllaceae | S | |
| Neolentinus lepideus (Fr.) Redhead & Ginns. | Gloeophyllaceae | S | |
| Gomphus clavatus (Pers.) Gray. | Gomphaceae | M | |
| Turbinellus floccosus (Schwein.) Earle ex Giachini & Castellano. | Gomphaceae | M | |
| Pallidohirschioporus biformis (Fr.) Y.C. Dai, Yuan Yuan & M. Zhou | Hirschioporaceae | P | |
| Fuscoporia gilva (Schwein.) T. Wagner & M. Fisch. | Hymenochaetaceae | P | |
| Hydnoporia olivacea (Schwein.) Teixeira. | Hymenochaetaceae | P | |
| Inonotus hispidus (Bull.) P. Karst. | Hymenochaetaceae | P | |
| Fulvifomes robiniae (Murrill) Murrill | Hymenochaetaceae | P | |
| Pseudoinonotus dryadeus (Pers.) T. Wagner & M. Fisch. | Hymenochaetaceae | P | |
| Trichaptum biforme (Fr.) Ryvarden. | Hymenochaetales | P | |
| Contumyces rosellus (M.M. Moser) Redhead, Moncalvo, Vilgalys & Lutzoni. | Rickenellaceae | P | |
| Lysurus mokusin (L. f.) Fr. | Phallaceae | S | |
| Climacocystis borealis (Fr.) Kotl. & Pouzar. | Polyporales | S | |
| Fabisporus sanguineus (L.) Zmitr. | Polyporales | S | |
| Cerrena hydnoides (Sw.) Zmitr. | Cerrenaceae | S | |
| Cerrena unicolor (Bull.) Murrill. | Cerrenaceae | S | |
| Jahnoporus hirtus (Cooke) Nuss. | Dacryobolaceae | S | |
| Postia ptychogaster (F. Ludw.) Vesterh. | Dacryobolaceae | S | |
| Daedalea quercina (L.) Pers. | Fomitopsidaceae | P | |
| Rhodofomes cajanderi (P. Karst.) B.K. Cui, M.L. Han & Y.C. Dai | Fomitopsidaceae | P | |
| Byssomerulius corium (Pers.) Parmasto. | Irpicaceae | S | |
| Byssomerulius incarnatus (Schwein.) Gilb. | Irpicaceae | S | |
| Irpex lacteus (Fr.) Fr. | Irpicaceae | S | |
| Leptoporus mollis (Pers.) Quél. | Irpicaceae | S | |
| Vitreoporus dichrous (Fr.) Zmitr. | Irpicaceae | S | |
| Ischnoderma resinosum (Schrad.) P. Karst. | Ischnodermataceae | P | |
| Phaeolus schweinitzii (Fr.) Pat. | Laetiporaceae | P | |
| Irpiciporus pachyodon (Pers.) Kotl. & Pouzar. | Meruliaceae | P | |
| Phlebia radiata Fr. | Meruliaceae | S | |
| Phlebia tremellosa (Schrad.) Nakasone & Burds. | Meruliaceae | S | |
| Panus conchatus (Bull.) Fr. | Panaceae | S | |
| Panus strigellus (Berk.) Chardón & Toro. | Panaceae | S | |
| Hapalopilus rutilans (Pers.) Murrill. | Phanerochaetaceae | P | |
| Terana coerulea (Lam.) Pers. | Phanerochaetaceae | S | |
| Abortiporus biennis (Bull.) Singer. | Podoscyphaceae | P | |
| Cryptoporus volvatus (Peck) Shear. | Polyporaceae | P | |
| Cubamyces lactineus (Berk.) Lücking. | Polyporaceae | P | |
| Cyanosporus caesius (Schrad.) McGinty. | Polyporaceae | P | |
| Daedaleopsis confragosa (Bolton) J. Schröt. | Polyporaceae | P | |
| Favolus tenuiculus P. Beauv. | Polyporaceae | E | S |
| Fomes fasciatus (Sw.) Cooke. | Polyporaceae | P | |
| Ganoderma applanatum (Pers.) Pat. | Polyporaceae | M | P |
| Ganoderma curtisii (Berk.) Murrill. | Polyporaceae | M | P |
| Lentinus arcularius (Batsch) Zmitr. | Polyporaceae | M | S |
| Lentinus crinitus (L.) Fr. | Polyporaceae | S | |
| Lentinus tigrinus (Bull.) Fr. | Polyporaceae | S | |
| Lentinus tricholoma (Mont.) Zmitr. | Polyporaceae | S | |
| Lenzites betulinus (L.) Fr. | Polyporaceae | P | |
| Neofavolus alveolaris (DC.) Sotome & T. Hatt. | Polyporaceae | S | |
| Picipes badius (Pers.) Zmitr. & Kovalenko. | Polyporaceae | S | |
| Polyporus umbellatus (Pers.) Fr. | Polyporaceae | S | |
| Trametes gibbosa (Pers.) Fr. | Polyporaceae | P | |
| Trametes hirsuta (Wulfen) Lloyd. | Polyporaceae | S | |
| Trametes versicolor (L.) Lloyd. | Polyporaceae | M | S |
| Sparassis radicata Weir. | Sparassidaceae | E | P |
| Steccherinum ochraceum (Pers. ex J.F. Gmel.) Gray. | Steccherinaceae | S | |
| Gymnosporangium juniperi-virginianae Schwein. | Gymnosporangiaceae | P | |
| Albatrellopsis ellisii (Berk. ex Cooke & Ellis) Teixeira. | Albatrellaceae | E | S |
| Albatrellopsis flettii (Morse ex Pouzar) Audet. | Albatrellaceae | S | |
| Albatrellus ovinus (Schaeff.) Kotl. & Pouzar. | Albatrellaceae | S | |
| Artomyces pyxidatus (Pers.) Jülich. | Auriscalpiaceae | S | |
| Auriscalpium vulgare Gray. | Auriscalpiaceae | S | |
| Lentinellus ursinus (Fr.) Kühner. | Auriscalpiaceae | S | |
| Bondarzewia sp. | Bondarzewiaceae | P | |
| Bondarzewia berkeleyi (Fr.) Bondartsev & Singer. | Bondarzewiaceae | P | |
| Heterobasidion annosum (Fr.) Bref. | Bondarzewiaceae | P | |
| Heterobasidion occidentale Otrosina & Garbel. | Bondarzewiaceae | P | |
| Peniophora albobadia (Schwein.) Boidin. | Peniophoraceae | S | |
| Lactarius argillaceifolius Hesler & A.H. Sm. | Russulaceae | M | |
| Lactifluus corrugis (Peck) Kuntze. | Russulaceae | M | |
| Laeticutis cristata (Schaeff.) Audet. | Russulales | M | |
| Lactarius deliciosus (L.) Gray. | Russulaceae | E | M |
| Lactarius indigo (Schwein.) Fr. | Russulaceae | E | M |
| Lactarius paradoxus Beardslee & Burl. | Russulaceae | M | |
| Lactifluus piperatus (L.) Roussel. | Russulaceae | T | M |
| Lactarius subplinthogalus Coker. | Russulaceae | M | |
| Lactifluus volemus (Fr.) Kuntze. | Russulaceae | E | M |
| Neoalbatrellus subcaeruleoporus Audet & B.S. Luther. | Russulales | S | |
| Russula adusta (Pers.) Fr. | Russulaceae | M | |
| Russula brevipes Peck. | Russulaceae | E | M |
| Russula cyanoxantha (Schaeff.) Fr. | Russulaceae | E | M |
| Russula emetica (Schaeff.) Pers. | Russulaceae | M | |
| Russula flavida Frost ex Peck. | Russulaceae | M | |
| Russula fragrantissima Romagn. | Russulaceae | M | |
| Russula mariae Peck. | Russulaceae | M | |
| Russula parvovirescens Buyck, D. Mitch. & Parrent. | Russulaceae | M | |
| Russula xerampelina (Schaeff.) Fr. | Russulaceae | M | |
| Stereum hirsutum (Willd.) Pers. | Stereaceae | S | |
| Stereum lobatum (Kunze ex Fr.) Fr. | Stereaceae | S | |
| Xylobolus frustulatus (Pers.) P. Karst. | Stereaceae | S | |
| Sebacina schweinitzii (Peck) Oberw. | Sebacinaceae | S | |
| Helvellosebacina concrescens (Schwein.) Oberw., Garnica & K. Riess. | Sebacinaceae | S | |
| Hydnellum spongiosipes (Peck) Pouzar. | Bankeraceae | S | |
| Sarcodon imbricatus (L.) P. Karst. | Bankeraceae | S | |
| Phellodon atratus K.A. Harrison. | Thelephoraceae | S | |
| Polyozellus multiplex (Underw.) Murrill. | Thelephoraceae | S | |
| Thelephora palmata (Scop.) Fr. | Thelephoraceae | M | |
| Thelephora terrestris Ehrh. ex Fr. | Thelephoraceae | M |
Abbreviations: Edibility: E = Edible, M = Medicinal, T = Toxic, H = Hallucinogenic, NR = Not recomended, BIO = Bioluminiscent. Life forms: S = Saprotrophic, P = Parasitic; M = Mycorrhizal.
Figure 4.
1.- Hypomyces lactifluorum, 2.- H. hialinus, 3.- H. chrysospermus, and 4.- H. microspermus, parasitic species on fruiting bodies of Russula spp., and Lactairus indigo, 5.- Fomitpsis pinicola, 6.- Ganoderma spp., 7.- Fomes fasciatus, 8.- Rhodofomes subfeei, 9.-Exudoporus frostii, 10.- Hortiboletus rubellus, 11.- Suillellus luridus, 12.- Hydnum repandum, 13.- Armillaria mellea, 14.- Pleurotus dryinus, 15.- Stereum ostrea, 16.- Bisomerulius corium.
Figure 4.
1.- Hypomyces lactifluorum, 2.- H. hialinus, 3.- H. chrysospermus, and 4.- H. microspermus, parasitic species on fruiting bodies of Russula spp., and Lactairus indigo, 5.- Fomitpsis pinicola, 6.- Ganoderma spp., 7.- Fomes fasciatus, 8.- Rhodofomes subfeei, 9.-Exudoporus frostii, 10.- Hortiboletus rubellus, 11.- Suillellus luridus, 12.- Hydnum repandum, 13.- Armillaria mellea, 14.- Pleurotus dryinus, 15.- Stereum ostrea, 16.- Bisomerulius corium.

Figure 5.
Example of some edible, parasitic and medicinal species from temperate forests of Nuevo León: 1.- Ganoderma. applanatum 2.-Trametes versicolor 3.-Morchella conica, 4.-Hericium erinaceus, 5.-Ganoderma resinaceum and 6.-Tuber lyonii.
Figure 5.
Example of some edible, parasitic and medicinal species from temperate forests of Nuevo León: 1.- Ganoderma. applanatum 2.-Trametes versicolor 3.-Morchella conica, 4.-Hericium erinaceus, 5.-Ganoderma resinaceum and 6.-Tuber lyonii.

Growth In Vitro of Selected Species from Nuevo Leon
Pure cultures from 110 species were obtained in vitro and their lineal growth was measured in solid culture media and compared according to their growth habit. Saprotroph species showed the greatest growth at 7 days of incubation in culture medium (3.5 cm), followed by mycorrhizal (0.7) and parasitic species (0.4). The Kruskall-Wallis analysis showed significant differences in the average growth of the species groups (p< 0.05). A pairwise analysis, after the Kruskall-Wallis, showed that the growth of saprotrophic species was significantly greater than that of mycorrhizal and parasitic species; these last two groups were not significantly different.
Figure 6.
Results from Kruskall-Wallis analysis showed significant differences in the average growth of the species groups (p< 0.05).
Figure 6.
Results from Kruskall-Wallis analysis showed significant differences in the average growth of the species groups (p< 0.05).
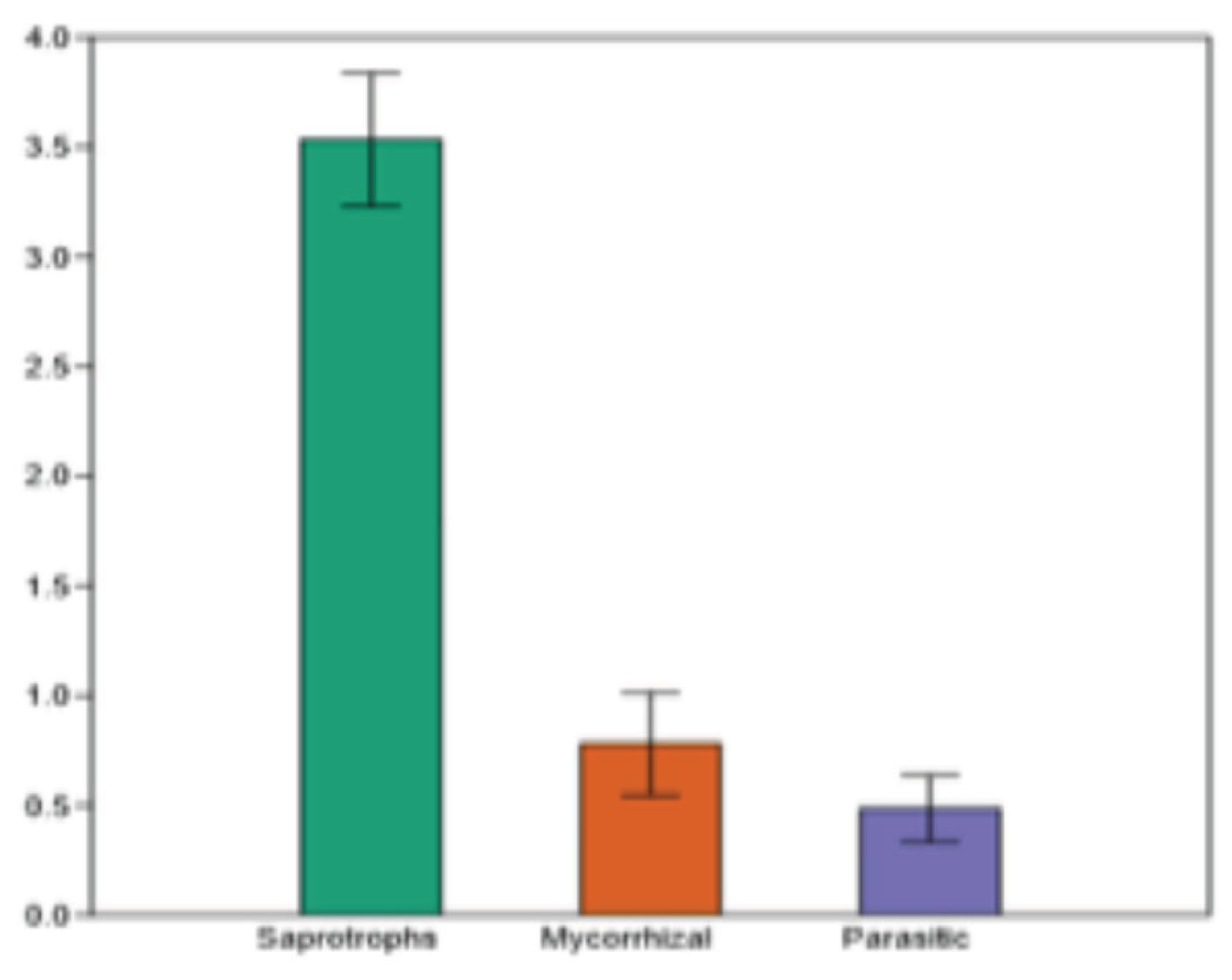
Table 2.
Mean growth in vitro of selected species after 7 days.
| Species | Family | Habit | Growth in vitro (cm) x week |
|---|---|---|---|
|
Agaricaceae | S | 3.5 |
|
Agaricaceae | S | 2.1 |
|
Agaricaceae | S | 2.4 |
|
Agaricaceae | S | 1.2 |
|
Amanitaceae | M | 9.5 |
|
Physalacriaceae | M | 0.9 |
|
Auriculariaceae | P | 1.3 |
|
Auriscalpiaceae | S | 2.3 |
|
Hydnaceae | M | 2.3 |
|
Hydnaceae | M | 2.3 |
|
Hydnaceae | M | 3.4 |
|
Omphalotaceae | S | 0.3 |
|
Omphalotaceae | S | 4.3 |
|
Agaricaceae | S | 4 |
|
Agaricaceae | S | 3.4 |
|
Agaricaceae | S | 9 |
|
Psathyrellaceae | S | 0.7 |
|
Polyporaceae | S | 4.3 |
|
Cortinariaceae | M | 0.4 |
|
Agaricaceae | S | 3.9 |
|
Fomitopsidaceae | P | 1.9 |
|
Fomitopsidaceae | S | 2.5 |
|
Entolomataceae | S | 2.4 |
|
Hericiaceae | S | 2 |
|
Hericiaceae | S | 2.1 |
|
Hericiaceae | S | 0.9 |
|
Hericiaceae | S | 1.7 |
|
Hericiaceae | S | 6 |
|
Physalacriaceae | S | 2.3 |
|
Physalacriaceae | S | 1.3 |
|
Polyporaceae | P | 2.2 |
|
Fomitopsidaceae | P | 7.5 |
|
Fomitopsidaceae | P | 2.3 |
|
Polyporaceae | P | 5.4 |
|
Polyporaceae | P | 6.1 |
|
Polyporaceae | P | 1.8 |
|
Polyporaceae | P | 1.5 |
|
Polyporaceae | P | 3.7 |
|
Polyporaceae | P | 2.3 |
|
Polyporaceae | P | 2.4 |
|
Polyporaceae | P | 6.9 |
|
Polyporaceae | S | 3.9 |
|
Polyporaceae | S | 1.3 |
|
Polyporaceae | S | 3.4 |
|
Gloeophyllaceae | P | 6.4 |
|
Hymenogastraceae | S | 3.7 |
|
Hymenogastraceae | S | 4.6 |
|
Hymenogastraceae | S | 3.5 |
|
Stereaceae | S | 2 |
|
Schizoporaceae | S | 0.8 |
|
Hypocreaceae | P | 1.9 |
|
Hymenochaetaceae | P | 1.8 |
|
Hymenochaetaceae | P | 5.5 |
|
Hydnangiaceae | M | 9.6 |
|
Hydnangiaceae | M | 9.8 |
|
Boletaceae | M | 6.5 |
|
Omphalotaceae | S | 0.6 |
|
Omphalotaceae | S | 0.4 |
|
Gloeophyllaceae | S | 3.5 |
|
Gloeophyllaceae | S | 1.5 |
|
Gloeophyllaceae | S | 2.9 |
|
Gloeophyllaceae | S | 1.7 |
|
Tricholomataceae | M | 3.5 |
|
Phallaceae | S | 0.5 |
|
Phallaceae | S | 1.3 |
|
Phallaceae | S | 0.3 |
|
Strophariaceae | P | 4.7 |
|
Strophariaceae | S | 3.9 |
|
Physalacriaceae | S | 2.8 |
|
Physalacriaceae | S | 1.8 |
|
Polyporaceae | S | 6.3 |
|
Paxillaceae | M | 0.8 |
|
Laetiporaceae | P | 1.6 |
|
Hymenochaetaceae | P | 2.6 |
|
Hymenochaetaceae | P | 2 |
|
Sclerodermataceae | M | 0.08 |
|
Pleurotaceae | S | 1.9 |
|
Pleurotaceae | S | 1.3 |
|
Pleurotaceae | S | 2.5 |
|
Pleurotaceae | S | 4.1 |
|
Pleurotaceae | S | 3.6 |
|
Pleurotaceae | S | 4.1 |
|
Pleurotaceae | S | 6.2 |
|
Pleurotaceae | S | 2.2 |
|
Pleurotaceae | S | 5.1 |
|
Pleurotaceae | S | 3.1 |
|
Pleurotaceae | S | 2.4 |
|
Pluteaceae | S | 2.4 |
|
Polyporaceae | S | 6.5 |
|
Polyporaceae | S | 11.1 |
|
Polyporaceae | S | 14.2 |
|
Polyporaceae | S | 9 |
|
Gelatoporiaceae | S | 5.8 |
|
Dacryobolaceae | S | 7.5 |
|
Polyporaceae | S | 3.5 |
|
Russulaceae | M | 8.4 |
|
Schizophyllaceae | S | 2.5 |
|
Schizophyllaceae | S | 2.3 |
|
Boletaceae | M | 1.6 |
|
Suillaceae | M | 2.6 |
|
Tapinellaceae | S | 0.7 |
|
Thelephoraceae | M | 5.6 |
|
Thelephoraceae | M | 3 |
|
Polyporaceae | S | 5.7 |
|
Polyporaceae | S | 4.7 |
|
Polyporaceae | S | 6 |
|
Polyporaceae | P | 6 |
|
Polyporaceae | P | 5.8 |
|
Polyporaceae | P | 9.8 |
|
Mycenaceae | S | 1.3 |
Growth In Vitro
Pleurotus dejamour
The strain number 3 produced primordia and fruiting bodies 24, 27 and 30 days after inoculation in the different seed mixtures (mixture #1, #2 and wheat).
Pleurotus dejamour Strains
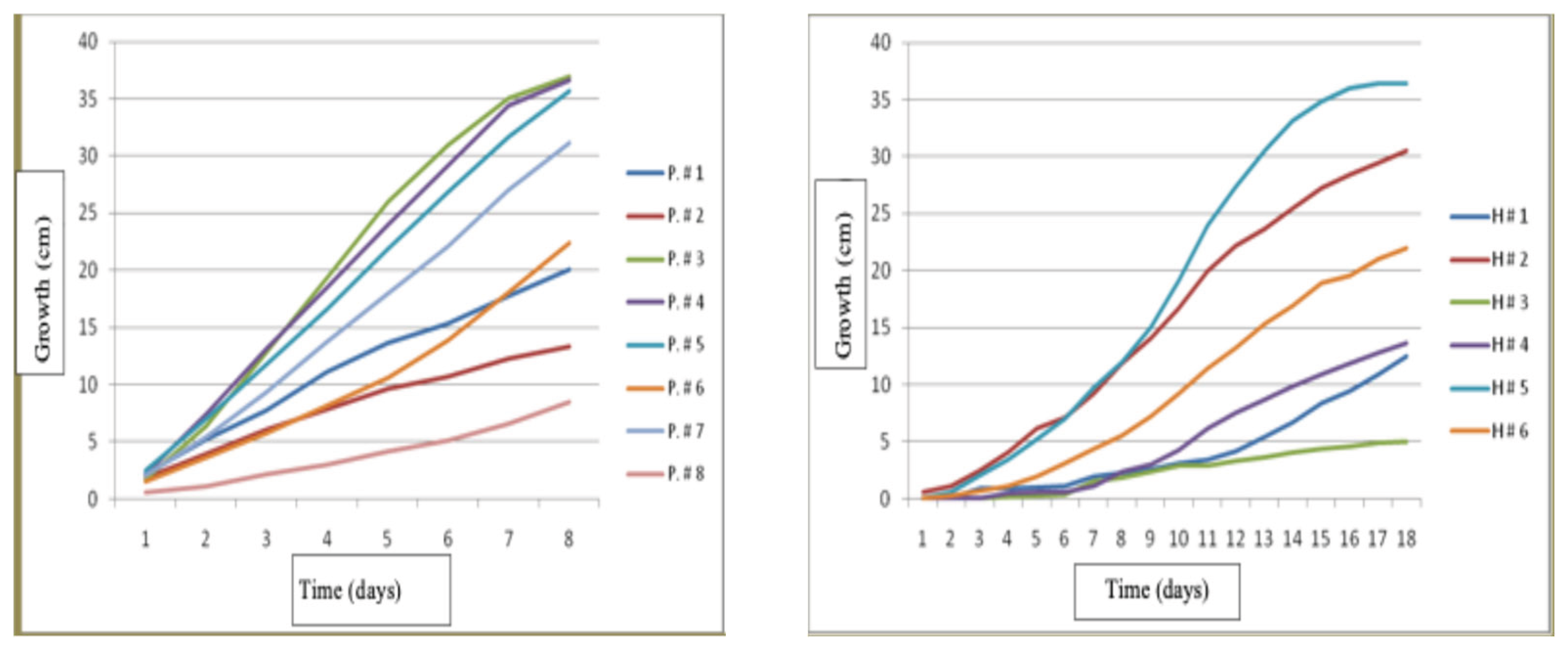
The best growth for Pleurotus dejamour was for strains P. #3, P. #4 and P. #5, at nine days from inoculation and the slowest growing strain was #8. An analysis of variance (ANOVA) was performed and the results showed statistically significant differences (P<0.05) in the growth for the eight strains of Pleurotus. However, no significant difference in the fast-growing strains P. #3, P. #4 and P. #5 was found, on the other hand, the slow growing strains i.e.1, 2, 6, 7, 8 had significant differences in their growth.
Hericium erinaceus
The best growth for Hericium erinaceus strains was for H#5 and it was followed by strains H#2 and H #6 since day 9 and it continued like that until day 18 and the slowest growth occurred for strain H #3. The results of the ANOVA showed significant differences of (P<0.05) for the growth of the six strains.
Figure 7.
- Growth of Pleurotus dejamour and Hericium erinaceus strains in vitro.
Hericium erinaceus
All the strains of Hericium erinaceus produced primordia in vitro on malt extract agar medium as well as in the jars filled with the different seeds. All strains also produced fruiting bodies in bags filled with straw in the different treatments.
Figure 8.
Primordia of Hericium erinaceus in vitro.

Inoculant Production in Treatments with a Blend of Seeds
Treatment 1 (Blend: Corn, Wheat, Sorghum)
The growth of Pleurotus dejamour strains on seed mixtures was evaluated. Results show that strain P3 had the best growth 24 days after inoculation, followed by strains P4 and P5, respectively (Figure 9).
Treatment 2 (Blend of Canary seed, Millet, Turnip, Flaxseed)
The results show that the best growth was produced by strain P4, followed by strains P3 and P5, and the lowest growth was obtained by strain P8 (Figure 10).
Treatment 3 (Wheat)
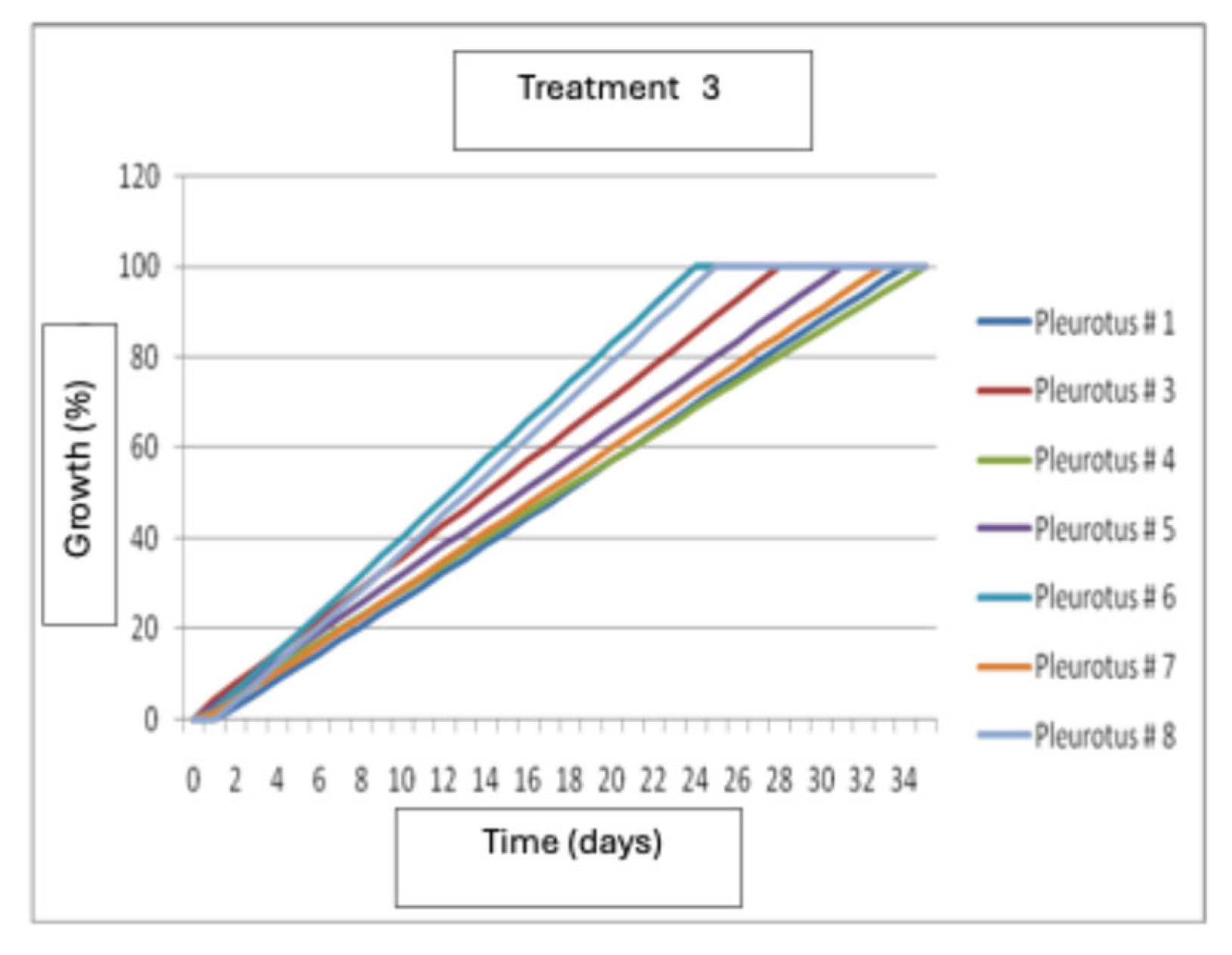
The results show that wheat seeds induced the fastest growth in strain P6 at 24 days after inoculation. This was followed by strains P8 and P3, respectively, while strain P4 was the slowest, obtaining its maximum growth at 34 days after inoculation (Figure 11).
Figure 9.
Mycelial growth of Pleurotus dejamour in seed Treatment 1.
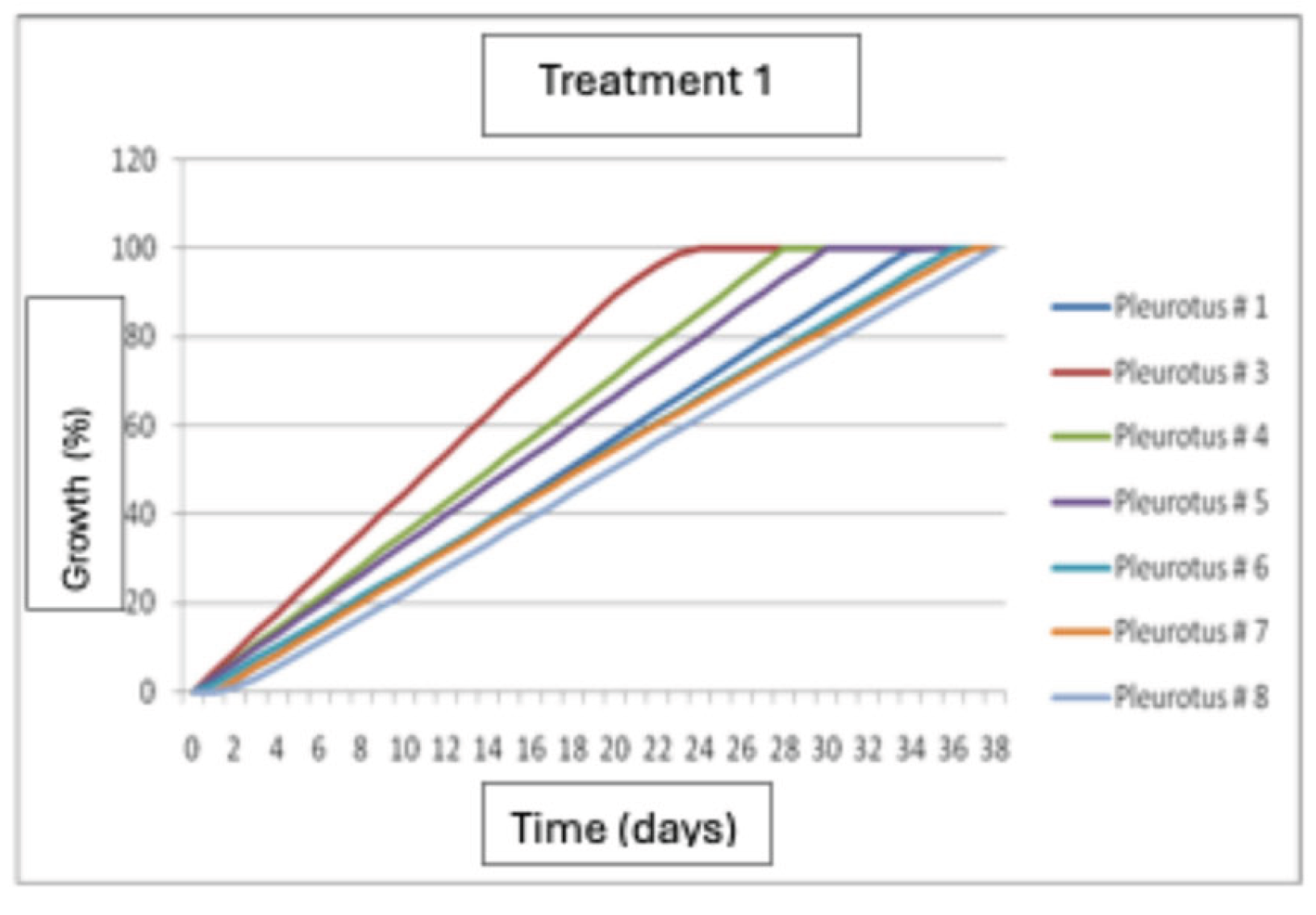
Figure 10.
Mycelial growth of Pleurotus dejamour in seed Treatment 2.
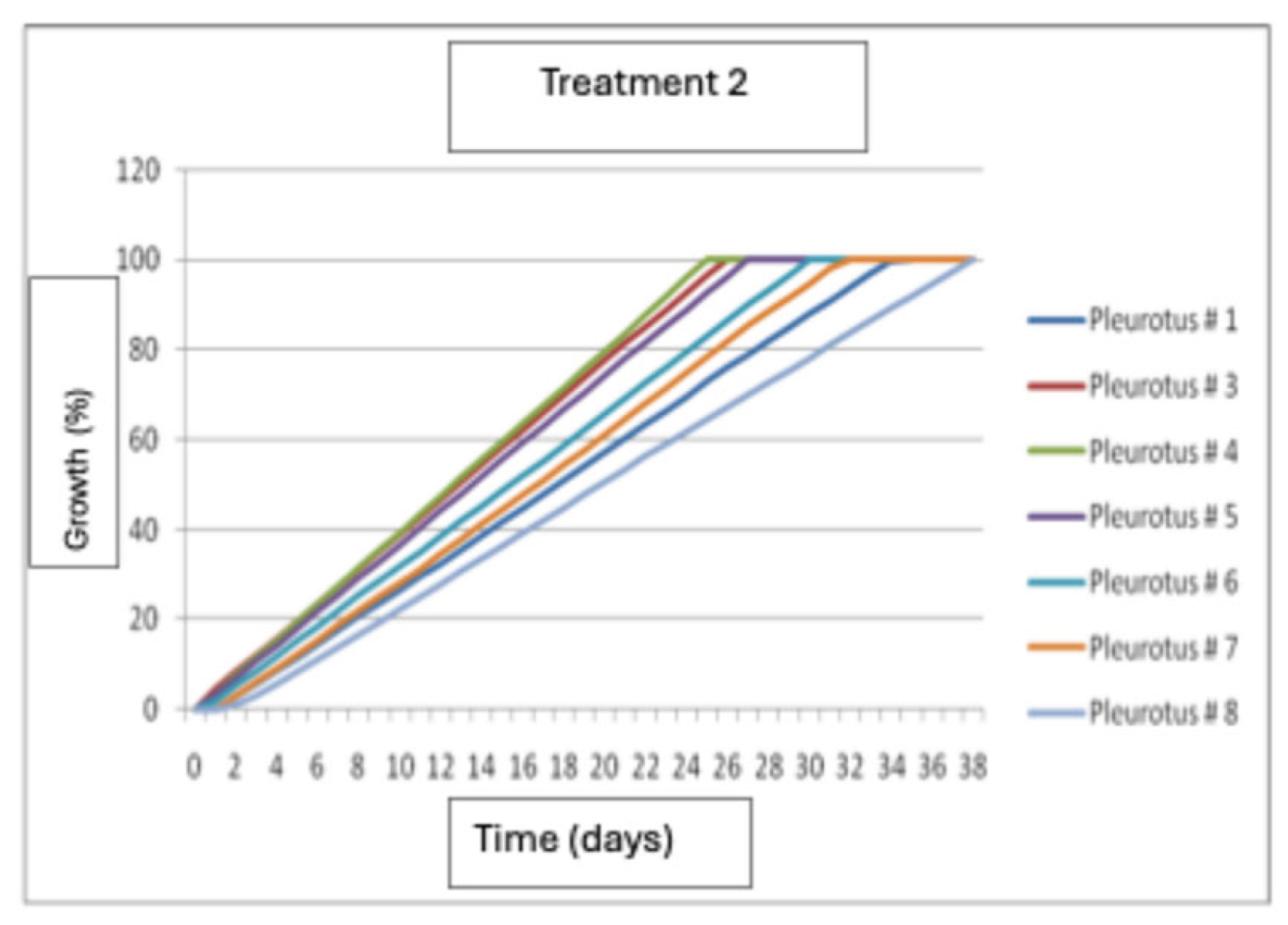
Figure 11.
Mycelial growth of Pleurotus dejamour in seed Treatment 3.
Treatment 1: Birdseed, millet, turnip, flaxseed, mustard
The results of this study showed that there were no statistically significant differences (P<0.05) in the growth of the six strains in this treatment. The best growth was produced by strain H # 4 at 20 days after inoculation, it was followed by strain H #1 and the slowest growth occurred in strain H #3.
Treatment 2: Corn, wheat, sorghum
The results of this study showed that there is no statistically significant difference (P<0.05) in the growth of the six strains in this treatment. Results showed that strain H #2 had the best growth at 19.5 days from inoculation, followed by strain H #6, and the with the lowest growth occurred was for strain H #4.
Treatment 3: Wheat
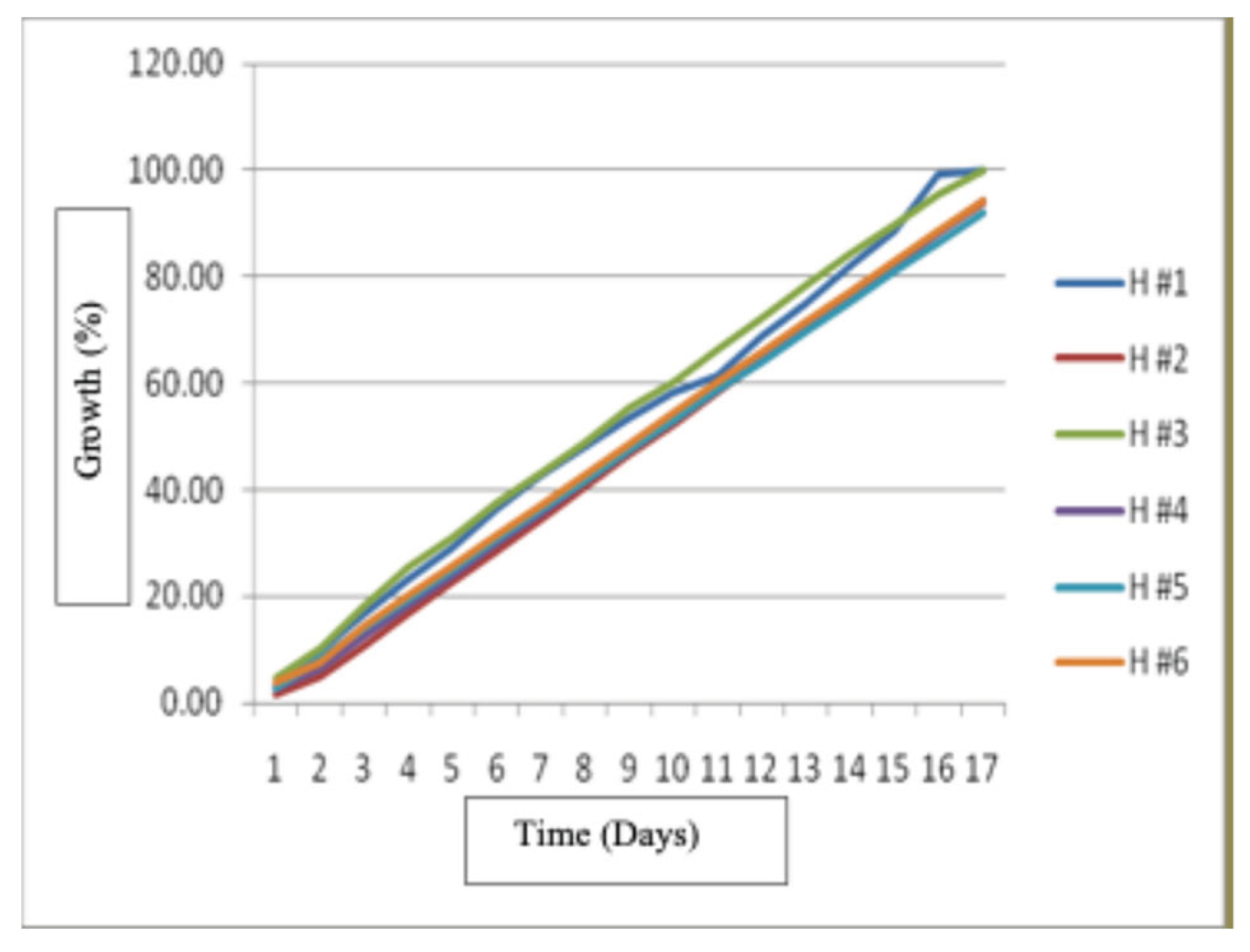
Results showed that strains H # 1 and H # 3 had the best growth at 16.5 and 17 days from inoculation, they were followed by strains H # 2, H # 4, and H # 6 and the strain with the lowest growth was H #5. The strains had similar homogeneous growth, and had only small differences (P<0.05)
Figure 12.
Growth of Hericium erinaceus strains in treatment 1.
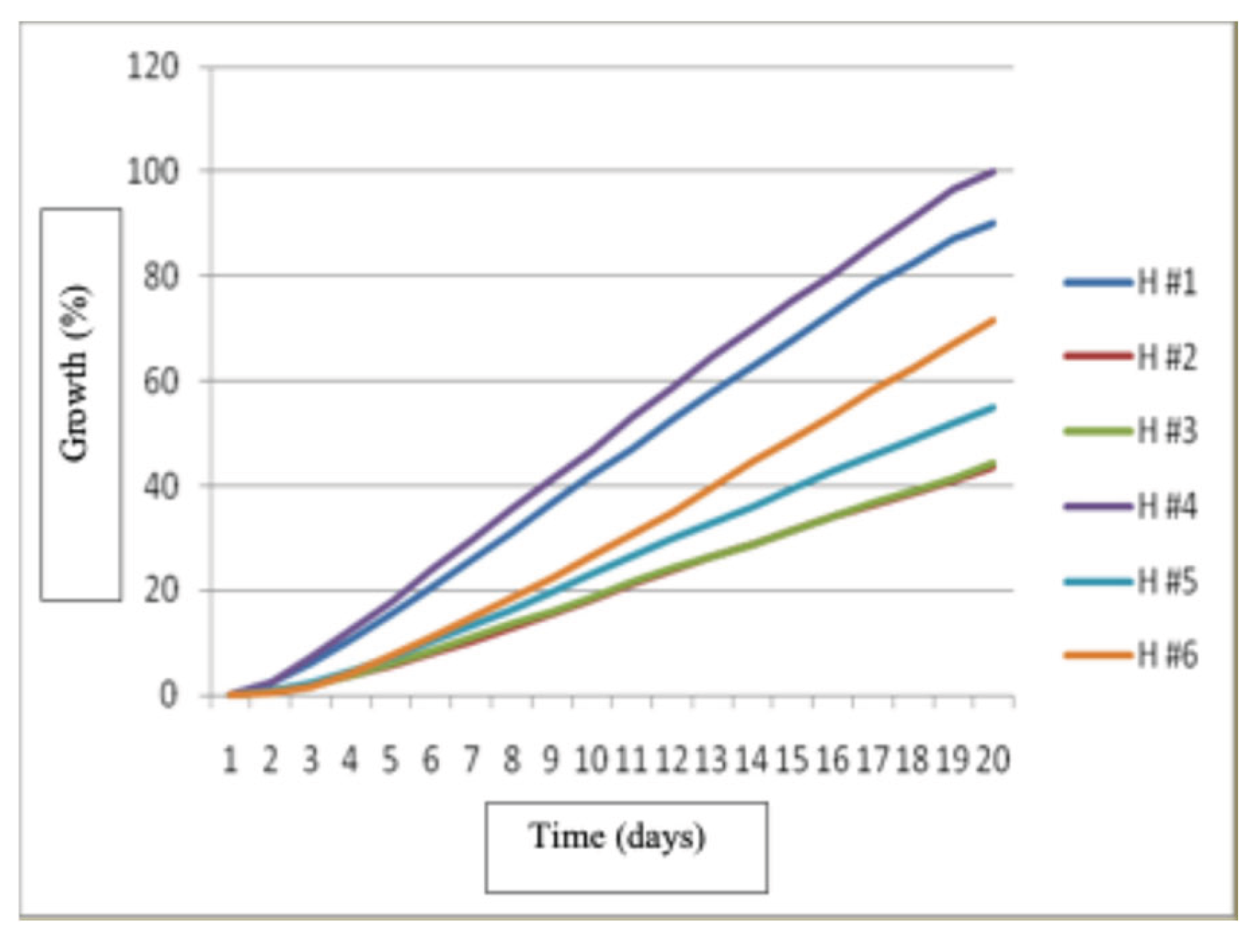
Figure 13.
- Growth of Hericium erinaceus strains in treatment 2.
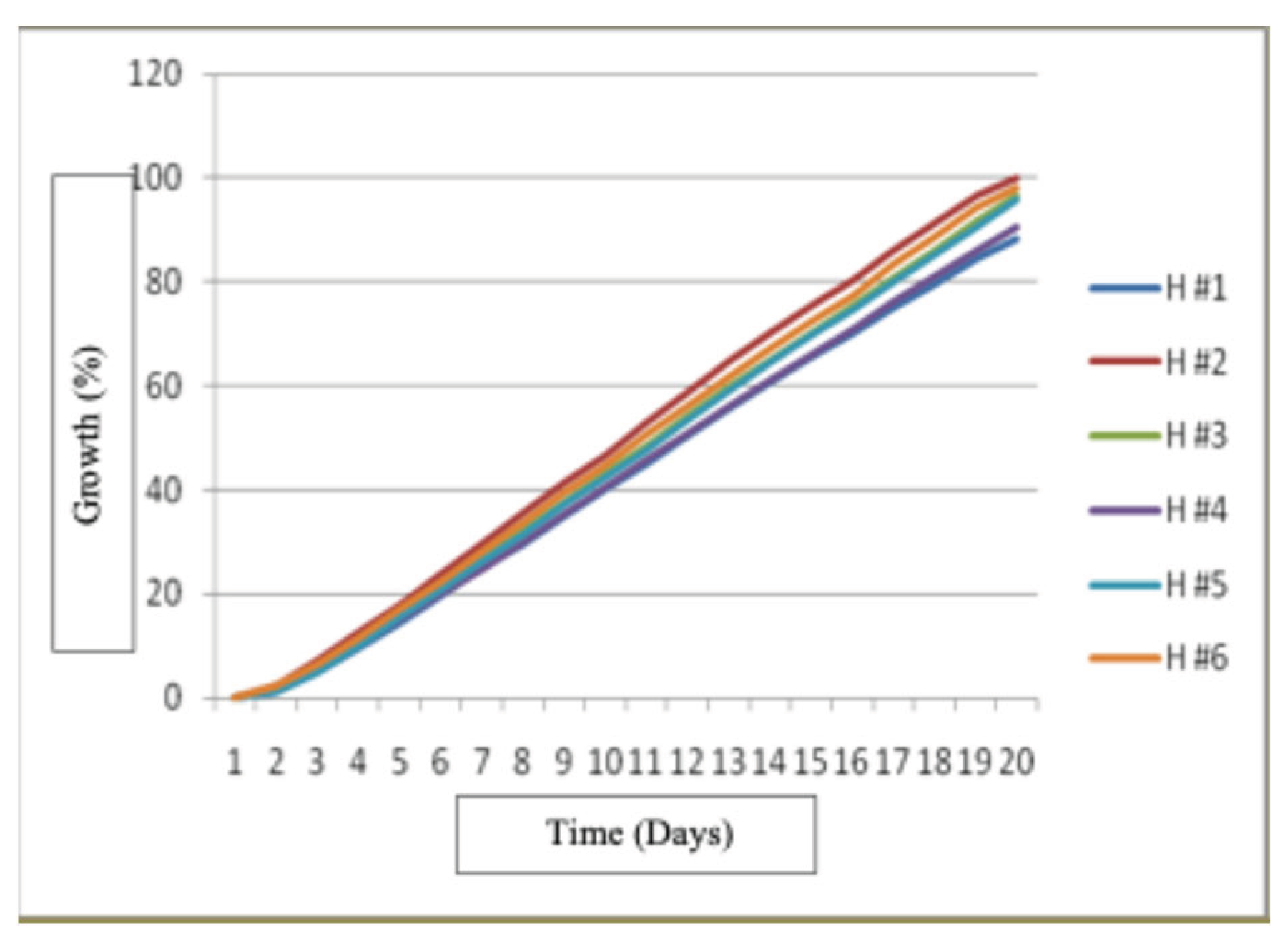
Figure 14.
Growth of Hericium erinaceus strains in treatment 3.
Fruit Bodies Production of Pleurotus dejamour: Treatment No. 1: Sorghum Straw
Results of this study showed that there was a statistically significant difference (P<0.05) in the production of fruiting bodies of the eight strains. The strain that showed the highest fruiting body production was P# 5 at the dose of molasses and humic acids at 8%, with a production of 410.53 and 405.36 g. Strain P#2 at doses of molasses and humic acids at 8% had the lowest production with 40.2 and 33.20 g. The strains that did not produce fruiting bodies were P# 3 and P# 6 at a dose of 8% humic acids and P# 1, P# 7, and P# 8 at all three doses used.
Fruit Bodies Production of Pleurotus dejamour in Treatment No. 2: Pasture Straw
The results of this study showed that there is a statistically significant difference (P<0.05) in the production of fruiting bodies of the eight strains evaluated with three doses used. The strain with the highest fruit body production was P#4 at the molasses dose, with a production of 342.77 g, followed by strain P#4 at the 6% humic acid dose, with an average production of 267.61 g. Strain P#4 with the 8% humic acid dose had the lowest production, with 11.71 g. In strains P# 1, P# 2, P# 7, and P# 8 at all three doses used, and strains P# 3, P# 5, and P# 6 at the 8% humic acid dose, no fruiting was obtained in this treatment.
Fruit Bodies Production of Pleurotus dejamour in Treatment No. 3: Coffee Husks
The results of this study indicate that there is a statistically significant difference (P<0.05) in the production of fruiting bodies of the eight strains evaluated with three doses used. The strain with the highest fruit body production was P# 3 at a dose of 6% humic acids with a production of 93.52 g, followed by strain P# 3 at a dose of molasses with a production of 79.5 g. Strain P#4 at the molasses dose had the lowest production, with 15.31 g. In strains P# 1, P# 2, P# 5, P# 6, P# 7, and P# 8 in the three doses evaluated, and strains P# 3 and P# 4 at a dose of 8% humic acids, no fruiting bodies were obtained for this treatment.
Figure 15.
Average production of Pleurotus dejamour fruiting bodies in treatment 1.
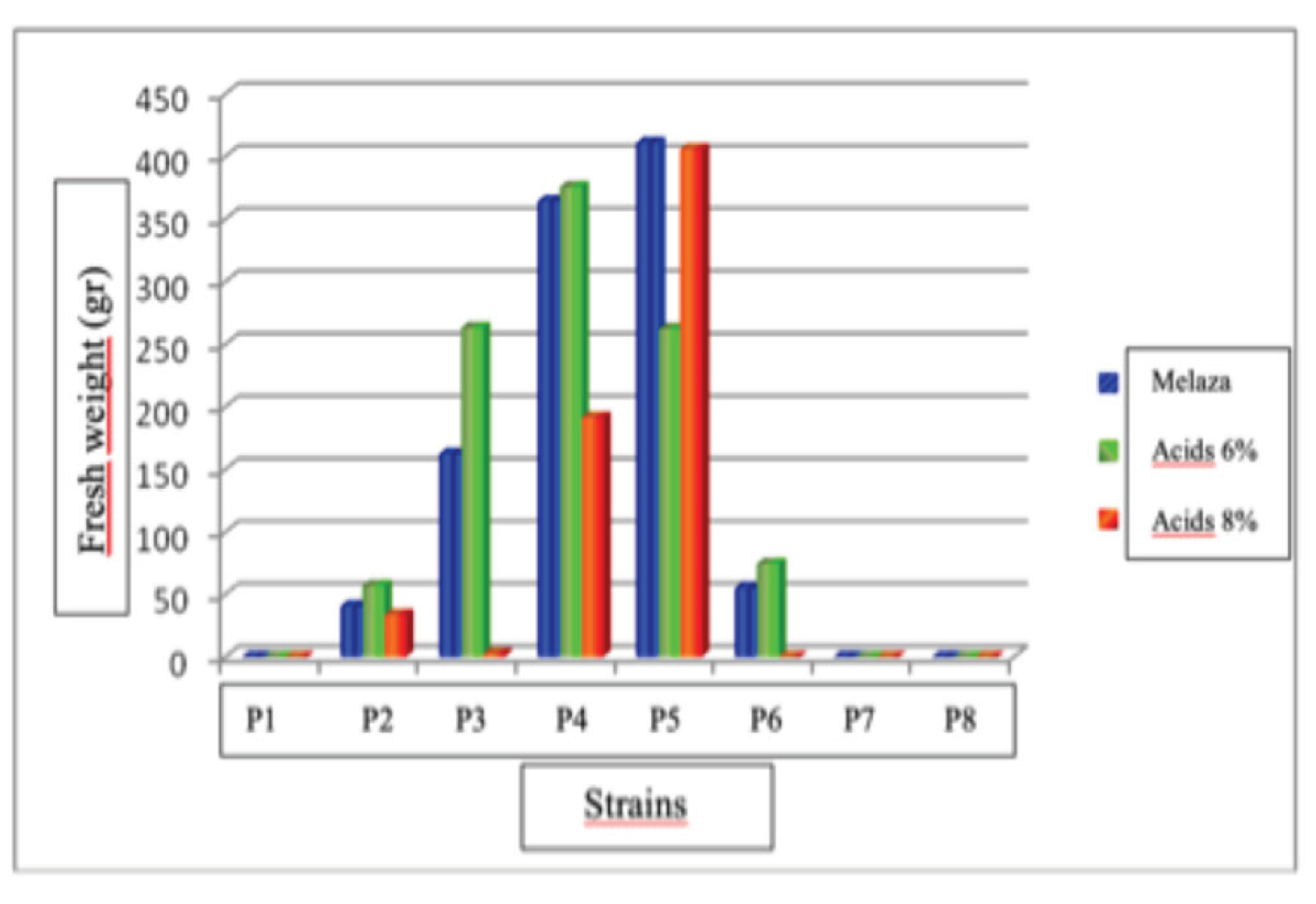
Figure 16.
Average production of fruiting bodies of Pleurotus dejamour in treatment 2.
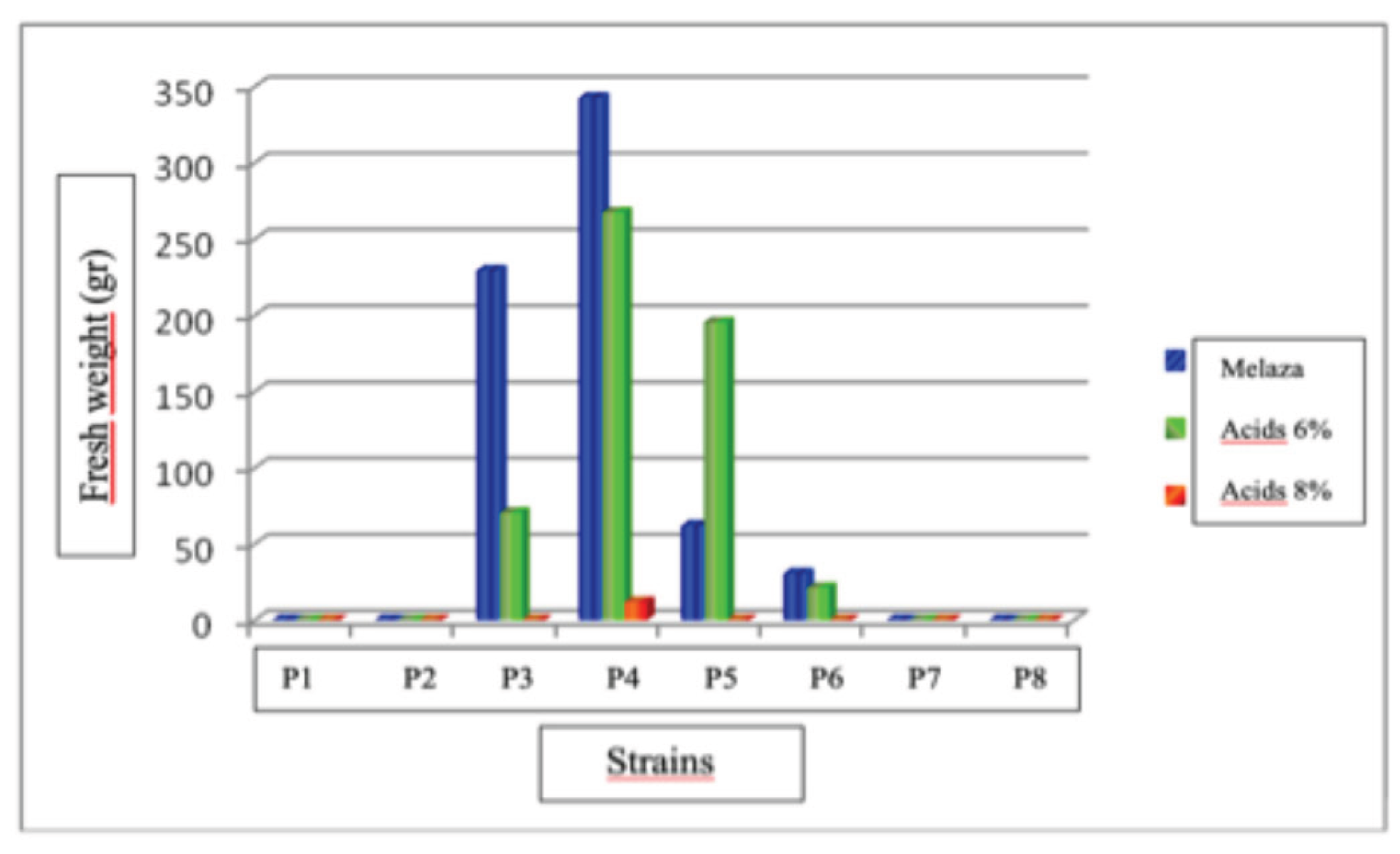
Figure 17.
Average production of Pleurotus dejamour fruiting bodies in treatment 3.
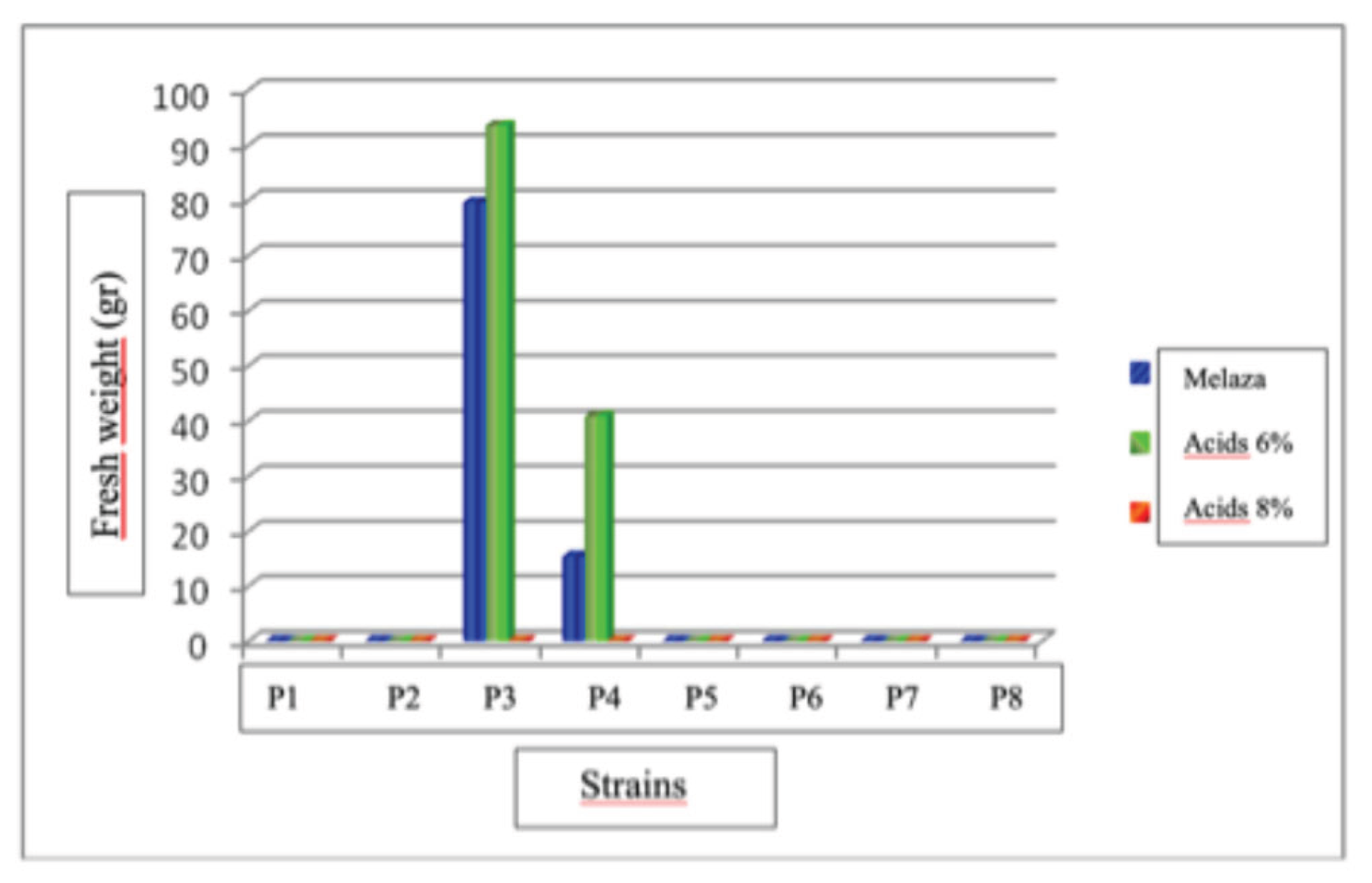
Production of Fruiting Bodies in different Substrates
Figure 18.
Fruiting bodies of Pleurotus dejamour growing in bags with substrate.
Fruiting bodies production of Hericium erinaceus in treatment No. 1
The results of this study indicate a statistically significant difference (P<0.05) in the production of fruiting bodies of the six strains of Hericium erinaceus evaluated with three doses used. Strain that showed the highest fruiting body production was H# 5 at a humic acid dose of 6%, with a production of 192.85 g, followed by H#3 at a humic acid dose of 6%, with a production of 172.85 g. Strain H#2 at a dose of 8% humic acids had the lowest production with 3.39 g.
Fruiting bodies production of Hericium erinaceus in treatment No. 1
The results of this study indicate a statistically significant difference (P<0.05) in the production of fruiting bodies of Hericium erinaceus for the six strains evaluated and with three doses used. The strain with the highest fruiting bodies production was H#1 at the molasses dose, with a production of 19.88 g, followed by strain H#5 at the 6% humic acid dose, with an average production of 17.46 g. Strain H#1 with a dose of 6% humic acids had the lowest production with1.02 g. No fruiting bodies were obtained in this treatment for strains H# 3, H# 4, H# 5, and H# 6 at the molasses dose, H# 4 and H# 6 at the humic acid dose, and H# 2, H# 3, and H# 6 at the 8% humic acid dose.
Figure 19.
Production of fruiting bodies of Hericium erinaceus strains in treatment No. 1.
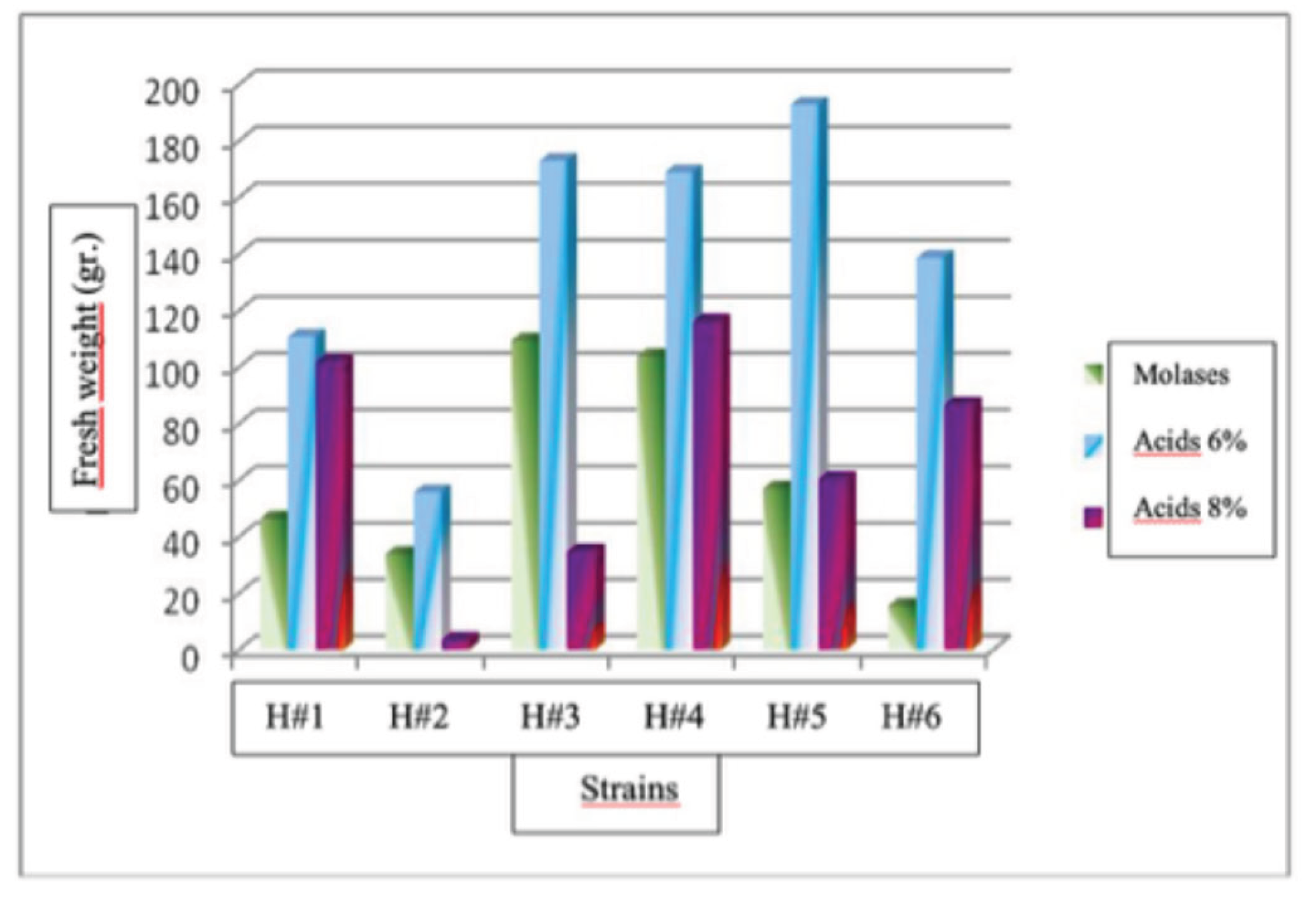
Figure 20.
Production of fruiting bodies of Hericium erinaceus strains in treatment No. 2.
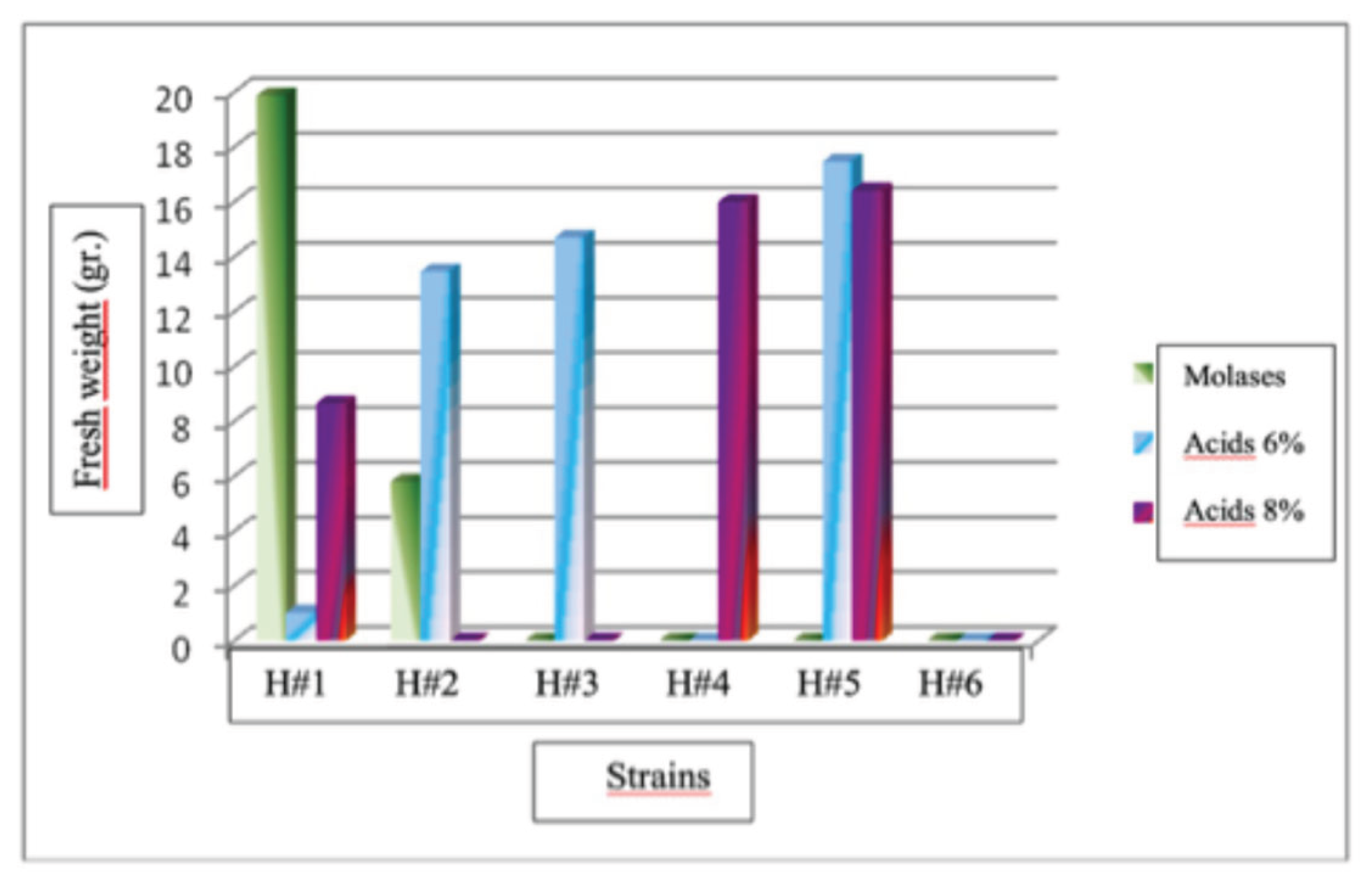
Analyzing the factors separately, it was found that for the strains of Hericium erinaceus had no significant differences in the production of fruiting bodies. Strains H#1, H#3, H#4, H#5, and H#6 showing no statistically significant differences. For the treatment factor, the best fruiting body production occurred in treatment 1 (i.e., sorghum), which had statistically significant differences (P<0.05) from the other two treatments with an average production of 21.91 g. The treatment with the lowest production was No. 2 (i.e., grass) with an average production of 1.55, and the treatment No. 3 (i.e., coffee) did not produce fruiting bodies. For the dose factor, the best fruit production was obtained with humic acid doses of 6%, with an average production of 7.82 g, with statistically significant differences (P<0.05) compared to the 8% humic acid and molasses doses.
Figure 21.
Fruitings bodies of Hericium erinaceus grown in bags.

Discussion
Results of this study showed that saprotrophic, parasitic and mycorrhizal macro fungi are widely distributed and very abundant in the natural oak, oak-pine and pine forests of the state of Nuevo León. Some species reported as edible, medicinal and mycorrhizal were found associated either with oak or pine species and some of them were isolated in pure culture in the laboratory and growth of 110 species was measured in vitro. Some strains of Pleurotus dejamour and Hericium erinaceus were cultivated and they grew very well in agricutural wastes and results suggests that they can be used in a larger scale. Recent studies report that the wealth of mycorrhizal fungi is unprotected worldwide, despite the known benefits they have for the world's vegetation and for obtaining goods and services for humans (Nuland, M., et al., 2025). Alterations in soil availability, water, and mineral nutrient cycles in forests lead to significant changes in the carbon cycle that affect their development and contribute to global climate change [21]. The protection of forests and their accompanying fungi in each locality, altitude, soil type, vegetation type, in the state of Nuevo León must be considered, as their importance is now recognized all over the world, and the state of Nuevo León should not be the exception. The health of forests promotes the integration of multiple species from different kingdoms, all of which are interconnected in symbiotic nutritional chains to fulfill their life cycles and thus promote a better climate, sufficient water and food supplies, and oxygen for humans on an annual basis. Our results represent a high percentage of fungal species but are only a small fraction of the species we have found over 45 years of study. The results indicate the presence of saprobic, parasitic, and mycorrhizal species, many of which are reported as edible and of very good quality, others as medicinal, and a few as hallucinogenic. The latter have recently been shown to have potential for improving the well-being of some patients with terminal illnesses, and their use in microdoses is rapidly increasing in demand worldwide. Our results on edible and medicinal mushrooms cultivation showed that they can be cultivated using relatively simple methods and residual agricultural products to produce high-quality organic food, as they are highly nutritious and low in sugar and fat [35]. We have conducted research related to the production of secondary metabolites from native strains of saprophytic and parasitic fungi, and the results have been encouraging, as they demonstrate the potential to reduce cholesterol, sugar, and uric acid levels, as well as being antioxidants and showing potential to reduce cancer levels in liver cells in vitro [7,17,38]. In the case of Schizophyllum commune and S. umbrinum they have been found to be both saprotrophic and parasitic with several species of native trees. Boletinellus merulioides is reported as mycorrhizal with Fraxinus spp., Gyrodon rompelli grows in thornscrubs in the absence of oaks, ash or pines with many legume trees in the surroundings belonging to different species and it might be possible that this species is facultative and either grows as saprotrophic or mycorrhizal. Desarmillaria tabescens has been reported as edible but there are also some reports of toxicity. The use of selected species of mycorrhizal fungi to inoculate oak and conifer seedlings produced in state nurseries using spores or ground fruits from these fungi. This practice ensures higher seedling survival, especially in areas that have been affected by forest fires, excessive logging, and water erosion [10] report excellent results using simple methods to inoculate native oaks with European truffles were reported by [19]. The results of this study also report at least one species of truffle that is edible and has the potential for in vitro cultivation for subsequent inoculation studies in nursery, greenhouse, and field conditions using native oak species that are highly diverse in the state of Nuevo León. Like truffles, the other species of mycorrhizal fungi reported in this study play important roles in the balance and maintenance of nutritional networks in the state's temperate oak, oak-pine, and coniferous forests. In addition to the above, many species are edible and of good quality and can be used in procedures known as mycorrhizal forestry [10]. Northeastern Mexico has potential for the development of mycological activities based on species that grow in forests, and we already have experience in implementing Mexico's first social and mycological project, carried out in 1999, the first Chihuahua Mushroom Fair. This project paved the way for mushroom fairs and festivals to be held with great success in different states of Mexico. In the state of Nuevo León, it was held in the municipality of Iturbide in 2007.
Conclusions
This study demonstrates that temperate forest in the state of Nuevo Leon have a high diversity of macro fungi species. Edible, medicinal and mycorrhizal fungi are abundant, and some strains have already been used in investigations showing promising results for human health and sustainable forest management. Results generate knowledge that can be used to promote future development in biotechnology and sustainable forest management as well as to promote social knowledge and use of macro fungi developing mushroom related activities with a positive social and economic impact.
References
- Anonymous. A mushrooming industry. Food production daily. Europe. 2007. Available online: http://foodproductiondaily.com/news/ng.asp?id=69917-mushrooms-shiitake.
- Claridge, AW; Trappe, JM; Hansen, K. Do fungi have a role as soil stabilizers and remediators after forest fire? Forest Ecology and Management 2009, 257, 1063–1069. [Google Scholar] [CrossRef]
- Chang Shu-Ting & Miles P. G., (1989). Edible mushroom and their cultivation. CRC. Press. pp. 27-40 and pp. 238–275.
- Estrada, E.; Villarreal, J.; Salinas, M.; Cantú, C.; González, H.; Jiménez, J., (2014) Coníferas de Nuevo León, México. Universidad Autónoma de Nuevo León. Facultad de Ciencias Forestales. 145 pp.
- Fergus, C.L.; Fergus, C. (2003). Common edible and poisonous mushrooms of the Northeast. (1st ed.) USA: Stackpole Books.
- Garza, F. Hongos ectomicorrícicos en el estado de Nuevo León. Rev. Mex. Mic. 1986, 2, 197–205. [Google Scholar]
- Garza, L.; Ramirez, X.; Garza, F.; Salinas, M.C.; Waksman, N.; Alcaraz, Y.; Torres, O. Evaluación de la actividad biológica de extractos acuosos de macromicetos del noreste de México. Ciencia UANL 2006, 9, 166–170. [Google Scholar]
- Garza, F.; Quiñónez, M.; Garza, L.; Carrillo, A.; Villalón, H.; González, H.; Valenzuela, R.; Guevara, G.; García, J.; García, M.; (2021) Some edible, toxic and medicinal mushrooms from temperate forests in the north of México. In Advances in macrofungi diversity, ecology and biotechnology. Edited by Kandrikere, R.S., and Deshmukh S. pp.145-198.
- Garza, F.; Quiñónez, M.; Guevara, G.; Carrillo, A.; de la Fuente, J.; García, J. Growth responses of Quercus fusiformis (Fagaceae) to ectomycorrhizal inoculation with Boletus luridellus Austrian. J. Mycol. 2022, 29, 193–202. [Google Scholar]
- Garza, F.; Quiñónez Martínez, M.; Carrillo Parra, A. (2023a) Diversity of Macrofungi in the Forest Ecosystems of the Cumbres National Park In: Garza F. (Ed.), Sustainable Management of Natural Resources. Diversity, Ecology, Taxonomy and Sociology. México, Linares: Springer. pp. 223–246. [CrossRef]
- Garza, F.; Guevara Guerrero, G.; Cuellar Rodríguez, G.; Garza Ocañas, L. (2023b) Diversity of Symbiosis Between Species of Macrofungi and Insects in the Temperate Forest of Iturbide Nuevo León In: Garza, F. (Ed.), Sustainable Management of Natural Resources. Diversity, Ecology, Taxonomy and Sociology. México, Linares: Springer. pp. 183–222. Garza, F., Ed.. [CrossRef]
- García, J.; y Garza, F. Conocimiento de los hongos de la Familia Boletaceae de México. Ciencia UANL 2001, 4, 336–343. [Google Scholar]
- García, J.; Ramírez, Y.; Castillo, S.; Moreno, A. Micofagia por roedores en los bosques templados de Tamaulipas. En Biodiversidad Tamaulipeca 2005, 1, 232–237. [Google Scholar]
- García, J.; Valenzuela, R.; Raymundo, T.; García, L.; Guevara, G.; Garza, F.; Cázares, E.; Ruiz, E. Macrohongos asociados a encinares (Quercus spp.,) en algunas localidades del estado de Tamaulipas, México. En Biodiversidad Tamaulipeca 2014, 2, 103–140. [Google Scholar]
- García, J.; Ayala, O.; Guevara, G.; Garza, G.; de la Fuente, J.I. Tuber mixtecorum (Tuberaceae, Pezizales) a new truffle in the Maculatum clade from Mexico. Phytotaxa 2021, 509, 113–120. Available online: https://www.mapress.com/j/pt/. [CrossRef]
- García, L.; Estrada, E.; Cantú, C.; Jurado, E.; Villarreal, J. Diversidad y conservación de los encinos (Fagaceae) del estado de Tamaulipas. En Biodiversidad Tamaulipeca 2014, 2, 3–14. [Google Scholar]
- González, P.; Garza, L.; Salinas, M.C.; Vera, L.; Garza, F.; Ramírez, X.; Torres, O. Producción de metabolitos antimicrobianos, alcaloides y taninos por Suillus lakei in vitro. Ciencia UANL 2009, XII, 62–70. [Google Scholar]
- Guevara, G.; García, J. Hongos comestibles de Tamaulipas. En Biodiversidad Tamaulipeca 2005, 1, 221–231. [Google Scholar]
- Guevara, G.; Giovanni, M.L.; Garza, F.; Gaitán, R. Mycorrhizal Synthesis of Périgord Black Truffle (Tuber melanosporum) with Mexican Oak Species. Microbiol Biotechnol. Lett. 2022, 50, 40–50. [Google Scholar] [CrossRef]
- Guzmán, G; Mata, G; Salmones, D; Soto, C; Guzmán, L. (1993) El cultivo de los hongos comestibles con especial atención a especies tropicales y subtropicales en esquilmos y residuos agroindustriales. Primera Edición. Instituto Politécnico Nacional. 245pp.
- Guo, W.; Wang, C.; Brunner, I.; Zhou, Y.; Wang, J.; Tang, Q.; Li, M.H. A global synthesis reveals that environmental factors drive the proportion of ectomycorrhizal exploration types. Geoderma 2025, 460, 117430. [Google Scholar] [CrossRef]
- Haro-Luna, M.H.; Guzmán-Dávalos, L.; Ruan-Soto, F. (2025) Cultural Importance of Wild Mushrooms: A Comparative Analysis Between Wixaritari and Mestizo Communities Established in Different Types of Vegetation.
- Hawksworth, D.L. The fungal dimension of biodiversity: magnitude, significance, and conservation. Mycological Research 1991, 95, 641–655. [Google Scholar] [CrossRef]
- Index Fungorum (2025) www.indexfungorum.org. Consultado el 21 octubre de 2024.
- Iordache, V.; Gherghel, F.; Kothe, E. Assessing the effect of disturbances on ectomycorrhiza diversity. International Journal of Environmental Research and Public Health 2009, 6, 414–432. [Google Scholar] [CrossRef] [PubMed]
- Kirk PM, Cannon PF, Minter DW, Stalpers JA (eds) (2008) Ainsworth & bisby’s dictionary of the fungi. CAB International, Wallingford.
- Lodge, D.J.; Ammirati, J.F.; Dell, T.O.; Mueller, G.M. (2004) Terrestrial and lignicolous macrofungi: collecting and describing macrofungi. In: Mueller G, Bills GF, Foster MS (Eds.) Biodiversity of Fungi. Inventory and monitoring methods. Elsevier Academic Press, New York, USA, 1–158.
- Marín, M.A.; Silva, V.; Linares, G.; Castagnino, A.M.; Ticante, J.A. La biodiversidad de los hongos ectomicorricicos y su importancia para la conservación del bosque en la zona poblana del Parque Nacional Malintzi. Estudios en Biodiversidad 2015, 1, 180–195. [Google Scholar]
- Madinaveitia, F.; Solis, A.; Medrano, H.; Quintero, J. (1984). "Project for obtaining microbial proteins and soil improvers from sawdust. Isolation of Trichoderma sp." In the utilization of cellulosic resources in animal feed by Mayra de la Torre. CINVESTAV. 253-258.
- Martínez; Carrera, D. Mushroom biotechnology in tropical America. The International Journal of Mushroom Sciences 2000, 3, 9–20. [Google Scholar]
- Mayett, Y.; Martínez, D.; Sánchez, M.; Macias, A.; Mora, S.; Estrada, A. Consumption of edible mushrooms in developing countries: The case of Mexico. J. Int. Fd & Agribus. Mar. 2006, 18, 151–176. [Google Scholar]
- Ostry, M.E.; Anderson, N.A.; O’Brien, J.G. Field guide to common macrofungi in eastern forests and their ecosystem functions. USDA Forest Service Northern Research Station General Technical Report. NRS- 2011, 79, 1–90. [Google Scholar] [CrossRef]
- Rivera, E.; Garza, F.; Quiroz, H.; Cuéllar, G.; Uvalle, J.I.; Valenzuela, R.; Guevara, G. (2023). Interactions between macrofungi and insects via sporocarps in three types of vegetation of the municipality of Linares, Nuevo León, Mexico. In: Garza, F. (Ed.), Sustainable Management of Natural Resources. Diversity, Ecology, Taxonomy and Sociology. México, Linares: Springer. pp. 223–246. [CrossRef]
- Siegel, N.; y Schwarz, C. (2024). Mushrooms of Cascadia, a comprehensive guide to fungi of the Pacific NorthwestBackcountry Press, Humboldt, California, 575pp.
- Solano Zavala, M.H.; Molinar Monsivais, I.; Reyes Ruvalcaba, D.; Quiñónez Martínez, M.; Enriquez Anchondo, I.D.; Garza-Ocañas, F. (2023) Edible Macromycetes of Chihuahua. In Diversity and Nutritional Properties In: Garza F. (Ed.), Sustainable Management of Natural Resources. Diversity, Ecology, Taxonomy and Sociology. México, Linares: Springer. pp. 131–142. [CrossRef]
- Zak JC & Willig MR (2004). Fungal biodiversity patterns. In: Mueller GM, Bills GF & Foster MS (Eds.), Biodiversity of fungi: Inventory and monitoring methods (pp. 59–75). London, England: Elsevier Academy Press.
- Van Nuland, M.E., Averill, C., Justin D., Stewart, Prylutskyi, O., Corrales, A., van Galen, L.G., 2, Bethan F. Manley, B.F., Qin, C., Lauber, T., Mikryukov, V., Dulia, O., Furci, G., Marín, C., Merlin Sheldrake, M., Weedon, J.T., Peay, K.G., Cornwallis, C.K., Větrovský, T., Kohout, P., Baldrian, P., Tedersoo, L., West, S.A., Crowther, T.W., Kiers, E.T. SPUN Mapping Consortium* & Johan van den Hoogen1,2" 2025. Global hotspots of mycorrhizal fungal richness are poorly protected. Nature 1-27.
- Castillo, D.M., Zanatta, M.T., Garza L., Garza, F., Barbosa, O., Villarreal, J. (2025). Insuline release by Fraction II of aquous extracts of Neolentinus lepideus in healthy and alloxan-induced diabetic rats. Journal of Medicinal Plants Research Vol. 19-(10). p.196-204. December 2025. Article number EFC 33837 4044. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



