
Submitted:
06 January 2026
Posted:
08 January 2026
You are already at the latest version
Abstract
Penicillium expansum inflicts significant economic damage in the fruit and vegetable industry due to its wide distribution and ability to infect a diverse range of hosts. Therefore, developing safe and environmentally sustainable strategies to suppress the growth of this pathogen is of critical importance. Bacillus subtilis, recognized for its broad antimicrobial activity and widespread occurrence, has been widely utilized in the biological management of plant diseases. This research seeks to assess the inhibitory potential of B. subtilis against P. expansum. The cell-free supernatant (CFS) derived from B. subtilis significantly suppresses the germination spores, germ tube extension, and hyphal development of P. expansum. It also reduces disease incidence in grapes and citrus and suppresses the expansion of lesions. Further investigate had shown that it can induced mycelium reactive oxygen species (ROS) accumulation, destroyed the cell membrane integrity, led to leakage of cytoplasmic contents and induced membrane lipid peroxidation. Moreover, exposure to high concentrations of CFS results in mycelial contraction and morphological abnormalities, triggering the disintegration of intracellular organelles and markedly upregulating the expression of apoptosis-associated genes. The self-protective response elicited by 5% CFS is insufficient to counteract the extent of cellular damage, ultimately driving cells toward a dynamic, multistage, and disintegrative form of cell death. The findings of this study offer a theoretical foundation for managing P. expansum after harvest.
Keywords:
Bacillus subtilis
; Penicillium expansum
; antimicrobial activity
1. Introduction
Penicillium expansum is a ubiquitous necrotrophic fungus found in various natural habitats [1]. It has a wide host spectrum and can infect numerous kinds of berries, pomes, and drupes [2,3], including grapes [4], apples, pears, peaches, and cherries [5,6]. The disease caused by P. expansum, commonly referred to as blue mold decay, is considered one of the most widespread causes of fruit deterioration after harvest globally [3]. It causes significant economic losses in the fruit industry during storage, transportation, and marketing. During fruit infection, P. expansum is capable of producing patulin, a toxic fungal metabolite that can be harmful to human health. which is primarily synthesized in the necrotic regions of the infected tissue, and can also migrate to other parts of the fruit [7]. Managing the growth and progression of P. expansum continues to be a major challenge in maintaining the safety and quality of fruit-based products.
Traditionally, Chemical agents like myclobutanil (MYC), imazalil (IMA), and pyrimethanil (PYR) have been extensively used to control green mold [8]; Nevertheless, their extensive use has led to the development of pathogen resistance and caused environmental contamination. Growing public concern about reducing fungicide use and ensuring the production of healthy, safe fruits has driven increased research efforts into the development of chemical-free methods for managing postharvest diseases. In recent years, there has been increasing interest in biological methods for controlling P. expansum, as they pose fewer risks to human health and the environment. The application of microbial antagonists to suppress pathogenic organisms has emerged as a central area of research. Commonly used biological control agents include various yeast species such as Rhodotorula mucilaginosa [9], Candida sake [10], and Kluyveromyces marxianus [11], as well as bacterial agents such as Bacillus spp. [12,13,14], Pseudomonas spp. [15,16] and Lactobacillus spp. [17]. Among these, Bacillus has garnered significant attention due to its widespread distribution in soil and broad-spectrum inhibitory activity, leading to its successful application in various commercial and industrial settings. The primary Bacillus species that have been utilized in biological control research and practical applications include Bacillus subtilis [18], B. cereus [19,20] , B. megaterium [21] , B. pumilus [22], B. siamensis [23], B. amyloliquefaciens [24] and B. velezensis [25]. Numerous studies have demonstrated that Bacillus spp. can inhibit various plant pathogenic fungi including but not limited to Aspergillus flavus [26], Fusarium verticillioides [27], Botrytis cinerea [28], and Botryosphaeria dothidea [29]. The cell-free supernatant (CFS) contains a variety of enzymes and secondary metabolites that enhance the growth of beneficial soil microorganisms, providing a sustainable, effective, and environmentally friendly solution to challenges associated with microbial inoculation. Its advantages, including low required concentrations and greater storage stability compared to live microbial cells, contribute to its broad applicability [30]. In this study, the CFS of B. subtilis was expanded to inhibit P. expansum and manage soft rot in grape, and citrus fruits. We primarily sought to investigate the inhibitory effects of CFS on P. expansum, while also delving into the mechanisms by which CFS damages P. expansum. This research aims to contribute to the development of an effective, environmentally friendly, and safe biological strategy for controlling P. expansum.
2. Materials and Methods
2.1. P. expansum and Preparation of Spore Suspension
P. expansum was cultured on potato dextrose agar (PDA) plates for 2 to 3 weeks at 25 °C temperature incubator. Spores were collected and resuspended in distilled water, then filtered through four layers of sterile cheesecloth to remove residual mycelium. Spore concentration was measured using a hemocytometer and adjusted to 1 × 106 cfu/mL.
2.2. Resuscitation of Bacillus subtilis and Preparation of Cell-Free Supernatant
Refer to Fan et al. [29], The Bacillus subtilis strain BS-1 was cryopreserved in the microbial culture collection. The strain was activated on a Luria - Bertani (LB) agar plate and incubated at 28 °C for 16 hours. Subsequently, a single colony was transferred to a liquid medium and cultured in a shaking flask at 28 °C until the optical density at 600 nm (OD600) reached 0.6. Thereafter, the culture was inoculated into a deodorizing fermentation medium (LB supplemented with 2% glucose) at an inoculum size of 0.5% and incubated at 28 °C for 72 hours under agitation at 180 rpm. After incubation, the bacterial suspension was centrifuged at 4 °C and 13,000 rpm for 20 min. The supernatant was filtered through a 0.22 μm membrane, supplemented with 50 mg/L streptomycin, and stored at 4 °C for later use.
2.3. Fruit Preparation and the Effect of CFS on Blue Mould Disease of Grape and Citrus Fruits
The Grape (Vitis vinifera L. cv. Kyoho) and citrus (Citrus sinensis cv. Navel) were harvested from the orchards in Yinchuan at commercial maturity stage without any treatments and immediately transferred to the laboratory. Fruits free from physical damage or microbial contamination were treated with 2% (v/v) sodium hypochlorite solution for 2 minutes, rinsed three times with sterile water, and then allowed to dry naturally. Disinfected fruit were injured at two evenly spaced locations in the equatorial area with a sterile stainless-steel probe. Each injury measured 2 mm in diameter and extended 3 mm in depth. 5 μL spore suspension of P. expansum and 5 uL 5% CFS were inoculated into each wounded. LB medium treated was as control. The processed fruits were stored in plastic containers and kept in a temperature-controlled incubation chamber set at 25 °C, with relative humidity maintained between 80% and 90%. The incidence rate was statistically analyzed and lesion diameters were recorded after 72 h of inoculation using a vernier caliper with the cross-crossing method.
2.4. Spore Germination, Germ Tube Elongation Test
For the spore germination and germ tube elongation assay, a 40 μL aliquot of P. expansum was uniformly spread on 1.5 × 3 cm sections cut from PDA plates containing 0, 1%, 2%, and 5% CFS. These sections were placed on sterile glass slides and incubated at 28 °C. After 8 and 10 hours of incubation, spore germination and the extension of germ tubes were observed, respectively. For each treatment, 50 spores were chosen at random to determine the germination percentage, and the lengths of the germ tubes were assessed using ImageJ software.
2.5. Mycelial Growth and Dry Weight
The mycelial development of P. expansum was evaluated by inoculating 20 μL of the fungal suspension into the center of PDA plates supplemented with 0, 1%, 2%, and 5% CFS, using an Oxford cup as the delivery method. The inverted plates were placed in a constant-temperature incubator at 28 °C for incubation. The colony diameter was measured every 24 hours using the cross-method measurement technique to evaluate the inhibitory effect.
A 1 mL sample of the spore suspension was inoculated into 20 mL of PDB medium and incubated at 28 °C with shaking at 180 rpm for 24 hours. Subsequently, CFS were added to the cultures adjust the concentration to 0, 1%, 2%, and 5%, which were further incubated for 48 h. In order to determine the dry weight, the mycelia were harvested and dried in an oven at 55 °C until a constant weight was achieved.
2.6. Mycelial Activity Measurement
Following Patel’s protocol [31] mycelia were harvested using the same method as described above, and their wet weight was recorded. The fragments were washed twice with PBS buffer (1 M, pH 7.0), then resuspended in 900 μL of PBS with 100 μL of MTT solution (5 mg/L). The samples were incubated in the dark at 28 °C on a shaker for 90 minutes, then centrifuged at 8000 × g for 3 minutes. 800 μL of DMSO was added to each sample, and the mixtures were gently shaken at 30 °C for 10 minutes to dissolve the formazan crystals completely. The absorbance was then recorded at a wavelength of 570 nm.
2.7. Morphology and Microstructure Examination
The mycelium of P. expansum, was treated with 5% CFS for 72 h, rinsed two times using 0.1 M PBS buffer (pH 7.4) and subsequently fixed in 2.5% glutaraldehyde at 4 °C for 12 hours. Following fixation, the sample underwent three 15-minute rinses with the same PBS buffer. Dehydration was performed using an ascending concentration gradient of ethanol (30%, 50%, 70%, 80%, 90%, 95%, and 100%), after which the specimen was immersed in isopentyl acetate for 15 minutes and subsequently dried using a critical point drying method. Finally, the dried sample was attached to conductive carbon film with double-sided tape, mounted on an ion sputtering instrument, and coated with gold for about 30 seconds to make it electrically conductive. Morphological of mycelium were observed using a scanning electron microscope (SEM) (HITACHI, SU8100).
For transmission electron microscope (TEM) observation, mycelium samples were fixed and dehydrated as previously described. The samples were infiltrated and embedded in EMBed 812 resin using acetone as a transitional solvent. Fresh EMBed 812 solution was added every 2 hours to gradually increase resin concentration. After overnight infiltration at 37 °C, the samples were polymerized in molds at 65 °C for over 48 hours. Ultrathin sections of 60–80 nm was cut from the resin-embedded samples using an ultramicrotome, and these sections were subsequently mounted on 150-mesh copper grids pre-coated with a formvar support film. Sections were stained with 2% uranyl acetate in alcohol avoid light and 2.6% lead citrate CO₂-free (each 8 min), then observed and imaged under a HITACHI HT780 TEM.
2.8. ROS Detection
Referring to the method of Li et al. [32] , The mycelium was collected 16 h after treatment with 0, 1%, 2%, and 5% CFS. After removed the medium and rinsed three times with 0.05 M PBS buffer (pH 7.0). The mycelium was stained with 25 μM 2’,7’-dichlorohydrofluorescein diacetate (H2DCF-DA, Sigma) and incubated for 60 minutes in a water bath maintained at 30 °C. The mycelium was resuspended, rinsed using 0.05 M PBS buffer (pH 7.0) and subjected to centrifugation at 10,000 rpm for 2 minutes until the dye solution was completely eliminated. ROS accumulation was examined on a laser confocal microscopy (LSM710, Zeiss, Oberkochen, Germany) at 488 nm excitation and 520 nm emission. Fluorescence intensity was analyzed with ImageJ software.
2.9. Cell Leakage Detection
Referring to the method of Zhao et al. [28], cell membrane permeability and leakage were assessed by measuring conductivity, nucleic acid content, protein content and malondialdehyde (MDA) content. A 0.1 mL sample of P. expansum spore suspension was added to 5 mL of PDB medium and mixed uniformly. The cultures were incubated in a rotary shaker at 28 °C and 80 rpm for three days. The CFS was added to adjust the concentration to 0, 1%, 2%, and 5%, and samples were collected at 0, 3, 6, 9 h. The obtained cultures were filtered and subsequently utilized for measuring the electrolyte, nucleic acids and soluble proteins content. The concentration of electrolytes in the solution was assessed using a conductivity meter, while protein levels were quantified using the Coomassie Brilliant Blue assay [33], and the extent of nucleic acid leakage was evaluated by determining the absorbance at a wavelength of 260 nm. The mycelial samples were thoroughly homogenized into a fine powder with the aid of liquid nitrogen prior to conducting the MDA analysis. The MDA level was measured using the thiobarbituric acid (TBA) assay [34].
2.10. Cell Membrane Integrity Detection
The sample for detecting cell membrane integrity was prepared as described in Section 2.9. Mycelium was collected by centrifugation at 8,000 r/min for 2 min following 24 h of treatment. Following the method of Wang et al. [35], The mycelium was treated with a 5 mM propidium iodide (PI) solution, followed by incubation at 30 °C for 20 minutes in a water bath, and subsequently rinsed using 0.05 M PBS buffer (pH 7.0). The integrity of the mycelial cell membrane was assessed using laser confocal microscopy (LSM710, Zeiss, Oberkochen, Germany) at 535 nm excitation emission.
2.11. RNA Extracted and Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR) Analysis
After the mycelium was treated with 0, 5% CFS at 24 h and 48 h, the mycelium was collected and stored at -80 °C, respectively. Total RNA extraction was extracted using the OMEGA R6840-01 kit. Reverse transcription of RNA was carried out using the HiScript® III RT SuperMix for qPCR kit (Vazyme Biotech, Nanjing), and fluorescence quantitative PCR was carried out with Vazyme ChanQTM SYBR qPCR Master Mix. The above experiments were conducted according to the kit instructions. The primer sequences and corresponding gene IDs used in this study are listed as Table 1.
2.12. Data Statistics and Analysis
The experimental data were statistically analyzed and visualized using GraphPad Prism 8. In the significance analysis, a significant difference was observed when P < 0.05 and present with *, and an extremely significant difference was observed when P < 0.01 present with **. In the figures, samples that share the same letter are not significantly different from one another, whereas those labeled with different letters exhibit statistically significant differences.
3. Result
3.1. CFS Inhibited Spore Germination and Germ Tube Elongation of P. expansum
To evaluate the inhibitory effect of CFS on P. expansum, observations were conducted on spore germination and germ tube extension. As shown in Figure 1, the spores showed varying degrees of germination after 8 and 10 hours of incubation. Various concentrations of CFS markedly suppressed spore germination and germ tube growth, with the observed reduction in activity showing a concentration-dependent trend. Treatment with 5% CFS could reduce the spore germination rate from 94.3% to 47.1% (Figure 1A, B). while the germ tube lengths decreased from 22.4 μm to 8.8 μm, reducing by 61.1% respectively (Figure 1A, C). At 10 hours, while the control group showed a germination rate of 96.3%, the 5% CFS treatment group had a significantly lower rate of 84.8%. Germ tube elongation remained suppressed 10 hours after treatment. Germ tube lengths were 33.34 μm, 25.91 μm, 19.47 μm, and 16.70 μm at concentrations of 0, 1%, 2%, and 5%, respectively. At a concentration of 5%, the germ tube length was reduced by 49.9% relative to the control group. Germ tube elongation remained suppressed 10 hours after treatment. Germ tube lengths were 33.34 μm, 25.91 μm, 19.47 μm, and 16.70 μm at concentrations of 0, 1%, 2%, and 5%, respectively. At 5%, the germ tube length decreased by 49.9% compared to the control. The results indicate that CFS markedly decreased the germination rate of P. expansum spores (Figure 1B) as well as the length of germ tubes (Figure 1C), with a more pronounced inhibitory effect observed at higher concentrations.
3.2. CFS Inhibit Mycelial Growth and Activity
When investigating the effect of CFS on P. expansum on PDA medium, it was observed that mycelial growth was inhibited to varying degrees on PDA medium supplemented with different concentrations of CFS. The diameter of the colony diminished as the concentration of CFS increased. Low-concentration CFS exhibited a limited inhibitory effect on mycelial growth; however, with the concentration of CFS increasing, 5% CFS significantly suppressed colony expansion (Figure 2A, B). Compared with the control group, in the group containing 5% CFS, the bacteriostatic rates from the second day to the sixth day were 25.1%, 26.8%, 23.1%, 21.9%, and 19.9%, respectively. The effect of CFS on mycelial growth was assessed by measuring the dry weight of the mycelium. After 3 days of incubation, the dry weights of mycelium were 3.38 g, 2.78 g, 2.05 g, and 1.28 g, respectively, under different concentrations of CFS (Figure 2C, D). Compared with the control group, the 1%, 2%, and 5% CFS reduced the dry weight by 18.2%, 39.3%, and 62.6%, respectively. The mycelium activity was measured using the MTT method. It was observed that the mycelium activity decreased significantly following treatment with varying concentrations of CFS, and the decrease in activity showed a pattern dependent on concentration. After 48 h of treatment, the mycelium activity in the presence of 1%, 2%, and 5% CFS decreased by 0.08%, 27.7%, and 36.4%, respectively, while after 96 h, it decreased to 16.9%, 32%, and 41.2%, respectively (Figure 2E). The findings suggest that CFS is capable of markedly suppressing the mycelial development and physiological activity of P. expansum.
3.3. Inhibitory Effect of CFS on the Pathogenicity of P. expansum in Inoculated Grape and Citrus Fruits
To further investigate the effects of CFS on the pathogenicity of P. expansum in grape and citrus fruits, 5% CFS and P. expansum were co-inoculated into grape and citrus fruits. After 3 days of inoculation, the incidence rate significantly decreased 22.5% than the control group on grape (Figure 3A, B). On citrus (Figure 3C, D, E), the incidence rate significantly decreased 42.1%, 16.7% than the control group at 3,4 days. All fruits in the control group had developed complete disease symptoms, whereas the treated group exhibited an 83% disease incidence rate at 5 days. The lesion diameter was reduced by 25.4% compared to the control group at 3 days. The results demonstrated that 5% CFS could significantly suppress the alleviated the pathogenicity of P. expansum on grape and citrus.
3.4. CFS Upregulated the Expression of Genes and Induced ROS Accumulation in the Mycelium of P. expansum
Reactive oxygen species (ROS), which encompass both oxygen free radicals and non-radical reactive molecules, play a critical role in regulating essential biological processes, including growth, stress tolerance, and programmed cell death (PCD). To better understand the mechanism by which CFS induces hyphal inhibition, ROS levels in hyphal cells were monitored following exposure to CFS. As shown in Figure 4A, The intensity of ROS-associated green fluorescence in hyphal cells rose progressively with increasing CFS concentration, suggesting sustained ROS buildup after CFS application. Pathogenic fungi generate ROS through the catalytic activity of NADPH oxidases (Nox). NoxA is the only one that contains a complete catalytic core domain among the three Nox catalytic subunits in filamentous fungi. The regulatory subunit NoxR and the small GTPase RacA are essential for activating the function of fungal NoxA [37,38]. The PeNoxA, PeNoxR and PeRacA genes related to ROS synthesis were detected. As shown in Figure 4 C, D, and E. PeNoxA, PeNoxR, PeRacA were significantly upregulated by 64.18-fold, 30.91-fold, 40.46-fold at 48 h post-CFS treatment, respectively.
3.5. CFS Destroyed the Cell Membrane Integrity, Resulted in the Release of Cytoplasmic Components and Triggered Lipid Peroxidation in the Cell Membrane of P. expansum
To further investigate the impact of excessive ROS accumulation on the cell membrane, membrane integrity was assessed using PI staining, macromolecule secretion analysis, and measurement of MDA levels. Propidium iodide (PI) can bind to DNA and emit red fluorescence; however, because it cannot penetrate intact cell membranes, it is widely used as a reliable tool for assessing cell membrane integrity. As shown in Figure 5A, the control group displayed normal cellular morphology, with nearly negligible red fluorescence detected across the entire field of view. In the CFS treatment group, 1% and 2% CFS also produced faint red fluorescence, stronger red fluorescence was observed in 5% CFS treatment group. The intensity of red fluorescence gradually rose with increasing CFS concentration, further confirming a dose-dependent response. The alterations in cell membrane integrity were further demonstrated by measuring the conductivity, nucleic acid content, and protein concentration in the supernatant obtained from the co-culture of CFS and P. expansum following 3 h, 6 h, and 9 h of treatment with 5% CFS. The conductivity (Figure 5C), nucleic acid concentration (Figure 5D) and soluble protein content (Figure 5E), exhibited a sustained and statistically significant increase compared to the control group. MDA, resulting from oxidative degradation of lipids in the fungal hyphal membrane, also gradually accumulated with the treatment time and significantly higher than the control group (Figure 5F). In summary, CFS destroyed the integrity of the hyphal membrane resulting in the imbalance of ion and the leakage of intracellular substances, which simultaneously induced the lipid peroxidation reaction.
3.5. CFS Causes Mycelial Shrinkage and Deformation, Leading to the Breakdown of Cellular Organelles
To gain deeper insights into the mechanisms through which CFS suppresses the growth of P. expansum, scanning electron microscopy (SEM) and transmission electron microscopy (TEM) analyses were performed. As shown in Figure 6A, in the control group, the mycelium was smooth, intact, with clearly visible septa. The mucopolysaccharide layer on the cell wall surface was evenly distributed (white arrow). The mycelia treated with 5% CFS showed mild damage, such as slight localized shrinkage (red arrow), partial distortion, uneven surface thickness, and sparse mucus distribution. The mycelium diaphragm also had uneven thickness (white arrow) (Figure 6B). The microstructure of the mycelium was analyzed using transmission electron microscopy. The results revealed no significant shrinkage of the cell body and preserved the normal morphology of cellular organelles in the control (Figure 6C). The overall damage to the bacteria treated with 5% CFS was relatively pronounced, characterized by sparse cytoplasm, significantly reduced electron density, shrunken cell bodies, and most organelles displaying evident swelling and vacuolation. The thickness of the cell wall (CW) was irregular, with sparse and unevenly distributed mucilaginous polysaccharides on the surface. The plasma membrane (PM) showed extensive damage and signs of disintegration. The nuclear membrane (N) was absent, and the chromatin appeared to have dissolved. Mitochondria (M) exhibited marked swelling, with a sparse and partially dissolved matrix, reduced or absent cristae, and vacuolar changes. The rough endoplasmic reticulum (RER) displayed a relatively reduced profile along with noticeable dilation. Auto phagolysosomes (ASS) were observed as isolated structures (Figure 6D). In summary, treatment with 5% CFS induced notable structural damage to the mycelium, as evidenced by cytoplasmic rarefaction, a marked decrease in electron density, cellular atrophy, and pronounced swelling and vacuolation of most organelles.
3.6. Alterations in the Expression Levels of Genes Associated with Autophagy Following CFS Treatment
To further investigate the impact of CFS on cell apoptosis of P. expansum, qRT-PCR was employed to analyze the gene expression levels related to autophagy-related genes after treated with 5% CFS for 24 h and 48 h. Subtilisin-related peptidase S8 (PePRT ) is a proteolytic enzyme highly expressed in plants and is closely related to the autophagy process and pathogenicity of P. expansum [36]. Compared with the control group, the expression levels of PePRT showed no significant change at 24 h post-CFS treatment. After 48 h, PePRT was significantly upregulated by 16.75-fold (Figure 7A). Program cell death, commonly referred to as apoptosis, is regulated by metacaspase [39,40]. After 48 hours of treatment with CFS, PeMetacaspase1 was upregulated by 2.06-fold (Figure 7B), and PeMetacaspase2 was upregulated by 4.96-fold (Figure 7C).
4. Discussion
P. expansum is a fungal pathogen responsible for causing blue mold rot in fruits and is capable of producing patulin, a mycotoxin that raises significant food safety concerns. Utilizing environmentally friendly and safe biological control methods to prevent and control P. expansum represents a highly effective and sustainable strategy. Bacillus has garnered significant attention owing to its broad distribution and capacity to produce antibacterial compounds. B. subtilis is widely utilized as a biocontrol agent owing to its capacity to effectively inhibit the growth of various pathogenic microorganisms. The CFS exhibits potent inhibitory activity against plant pathogenic fungi, promotes plant growth, and contributes to environmental remediation, demonstrating various potential applications in agriculture [30]. In this study, the CFS of B. subtilis significantly suppressed spore germination and germ tube elongation (Figure 1), while significantly inhibiting the mycelial growth of P. expansum in a dose-dependent manner under in vitro conditions. (Figure 2). This evidence was similar to the B. subtilis can inhibit the Botrytis cinerea [41], Fusarium graminearum [42], Verticillium dahlia [43], and Botryosphaeria dothidea [29]. To evaluate the inhibitory effect of CFS in vivo, co-inoculation of CFS and spore suspensions was performed on grapes and citrus fruits. In accordance with previous findings observed in kiwifruit [29] and blueberry [44], CFS significantly reduced the incidence and the lesion expansion (Figure 3), demonstrating its potential as a biological control agent for managing blue mold in grapes and citrus fruits.
When exposed to biotic stresses, organisms frequently initiate a range of defensive responses, the production of ROS represents a prevalent and essential mechanism [45] . This study found significantly increased ROS accumulation (Figure 4A, B) and upregulation of related synthase genes PeNoxA, PeNoxR, PeRacA (Figure 4C, D, E) after 5% CFS treatment (Figure 4A, B). However, ROS plays a dual role in signal transduction [46]. It acts as a secondary messenger regulating growth, development, and defense under normal conditions, but excessive accumulation leads to oxidative stress, damaging cells, degrading biomolecules, and disrupting physiological functions [47] . The integrity of the cell membrane and the leakage of macromolecular substances were detected. As expected, in the 5% CFS group, the intensity of red fluorescence detected by PI staining was significantly 25% higher than that in the control group, indicating a substantial compromise in cell membrane integrity (Figure 5A, B). Meanwhile, the increased conductivity of the supernatant (Figure 5C), elevated nucleic acid content (Figure 5D), and higher levels of soluble proteins (Figure 5E) further confirm the disruption of cell membrane integrity, resulting in the leakage of intracellular components. These findings confirm that the plasma membrane in P. expansum is a primary target of CFS. MDA accumulation (Figure 5F) also demonstrated excessive ROS induce lipid peroxidation, alter membrane permeability, which was similar to the results reported by Zhao et al. [28] and Fan et al. [29], as an indirect indicator of impaired membrane integrity.
Microscopic observations provide more direct and compelling evidence to support the earlier assertion that CFS compromises the structural integrity of cell membranes. The plasma membrane exhibited severe disruption and disintegration (Figure 6D). Meanwhile, novel insights into the underlying mechanisms of cell death were offered by SEM, TEM observations and qRT-PCR. CFS induced mycelial damage, shrinkage, and deformation, resulting in an irregular surface morphology (Figure 6B). Notably, significant cell body atrophy (Figure 6B) serves as an early morphological hallmark of apoptosis [48] , while the presence of autophagic lysosomes (Figure 6D) and upregulation of autophagy-related genes of PePRT (Figure 7A), PeMetacaspase1 (Figure 7B), PeMetacaspase2 (Figure 7C) further confirm concurrent autophagy activation. We propose that early stress induced by CFS trigger fungi to activate systemic resistance through apoptosis induction. However, as stress continues, intense ROS signaling gradually disrupts intracellular membrane integrity, leading to MDA accumulation and impairment of critical organelles such as mitochondria structures essential for sustained energy production. Concurrently, most organelles exhibit pronounced swelling and vacuolization, the cytoplasm becomes less dense, and electron density markedly decreases, culminating in organelle rupture and disruption of the regulated apoptotic cascade. Moreover, numerous organelles involved in biosynthetic processes undergo vacuolization and functional decline, resulting in widespread protein degradation, leakage of intracellular contents, and loss of membrane integrity. Collectively, these findings demonstrate that the CFS of B. subtilis effectively inhibits the growth of P. expansum and induces reactive oxygen species (ROS) accumulation. On one hand, ROS trigger systemic resistance by promoting apoptosis; on the other hand, excessive ROS levels compromise cell membrane integrity, leading to leakage of intracellular components. The self-protective response elicited by 5% CFS is insufficient to counteract the extent of cellular damage, ultimately driving cells toward a dynamic, multistage, and disintegrative form of cell death.
Funding
The present study was financially supported by the youth talent cultivation project of North Minzu university (2021KYQD41),the Key Research and Development Plan of Ningxia (2025BEH04064), National Natural Science Foundation of China (32260794) .
References
- Ackah, M.; Boateng, N.A.S.; Foku, J.M.; Ngolong Ngea, G.L.; Godana, E.A.; Zhang, H.; Yang, Q. Genome-wide analysis of the PG gene family in Penicillium expansum: Expression during the infection stage in pear fruits (Pyrus bretschneideri). Physiol. Mol. Plant Pathol. 2024, 131, 102270. [Google Scholar] [CrossRef]
- Wang, K.; Zheng, X.; Su, Y.; Lu, Y.; Yang, Q.; Shi, Y.; Lanhuang, B.; Zhang, X.; Zhao, L.; Godana, E.A.; et al. A glycoside hydrolase superfamily gene plays a major role in Penicillium expansum growth and pathogenicity in apples. Postharvest Biol. Technol. 2023, 198, 112228. [Google Scholar] [CrossRef]
- Luciano-Rosario, D.K.; Nancy, P.; Jurick, II; Wayne, M. Penicillium expansum: biology, omics, and management tools for a global postharvest pathogen causing blue mould of pome fruit. Mol. Plant Pathol 2020, 21, 1391–1404. [Google Scholar] [CrossRef]
- Alimadadi, N.p.; Zahra Nasr; Shaghayegh, Fazeli; Seyed Abolhassan, Shahzadeh. Screening of antagonistic yeast strains for postharvest control of Penicillium expansum causing blue mold decay in table grape. Fungal Biol. 2023, 127, 901–908. [Google Scholar] [CrossRef]
- Lu, M.; Chen, Y.; Li, L.; Ma, Y.; Tong, Z.; Guo, D.; Sun, P.; An, D. Analysis and Evaluation of the Flagellin Activity of Bacillus amyloliquefaciens Ba168 Antimicrobial Proteins against Penicillium expansum. Molecules 2022, 27, 4259. [Google Scholar] [CrossRef] [PubMed]
- Droby, S. Improving quality and safety of fresh fruits and vegetables after harvest by the use of biocontrol agents and natural materials. Acta Hortic. 2006, 709, 45–51. [Google Scholar] [CrossRef]
- Al Riachy, R.; Strub, C.; Durand, N.; Chochois, V.; Lopez-Lauri, F.; Fontana, A.; Schorr-Galindo, S. The Influence of Long-Term Storage on the Epiphytic Microbiome of Postharvest Apples and on Penicillium expansum Occurrence and Patulin Accumulation. Toxins 2024, 16, 102. [Google Scholar] [CrossRef] [PubMed]
- Costa, J.H.; Bazioli, J.M.; de Moraes Pontes, J.G.; Fill, T.P. Penicillium digitatum infection mechanisms in citrus: What do we know so far? Fungal Biol. 2019, 123, 584–593. [Google Scholar] [CrossRef]
- Zheng, X.; Yang, Q.; Zhang, X.; Apaliya, M.T.; Ianiri, G.; Zhang, H.; Castoria, R. Biocontrol Agents Increase the Specific Rate of Patulin Production by Penicillium expansum but Decrease the Disease and Total Patulin Contamination of Apples. Front. Microbiol. 2017, 8–2017, 1240. [Google Scholar] [CrossRef]
- Alvarez, A.; Gelezoglo, R.; Garmendia, G.; González, M.L.; Magnoli, A.P.; Arrarte, E.; Cavaglieri, L.R.; Vero, S. Role of Antarctic yeast in biocontrol of Penicillium expansum and patulin reduction of apples. Environ. Sustainability 2019, 2, 277–283. [Google Scholar] [CrossRef]
- Zheng, X.; Zheng, L.; Xia, F.; Li, J.; Zhou, W.; Yuan, L.; Rao, S.; Yang, Z. Biological control of blue mold rot in apple by Kluyveromyces marxianus XZ1 and the possible mechanisms of action. Postharvest Biol. Technol. 2023, 196, 112179. [Google Scholar] [CrossRef]
- Song, C.; Zhang, Y.; Zhao, Q.; Chen, M.; Zhang, Y.; Gao, C.; Jia, Z.; Song, S.; Guan, J.; Shang, Z. Volatile organic compounds produced by Bacillus aryabhattai AYG1023 against Penicillium expansum causing blue mold on the Huangguan pear. Microbiol. Res. 2024, 278, 127531. [Google Scholar] [CrossRef]
- Ding, Y.; Liu, F.; Yang, J.; Fan, Y.; Yu, L.; Li, Z.; Jiang, N.; An, J.; Jiao, Z.; Wang, C. Isolation and identification of Bacillus mojavensis YL-RY0310 and its biocontrol potential against Penicillium expansum and patulin in apples. Biol. Control 2023, 182, 105239. [Google Scholar] [CrossRef]
- Yu, L.; Qiao, N.; Zhao, J.; Zhang, H.; Tian, F.; Zhai, Q.; Chen, W. Postharvest control of Penicillium expansum in fruits: A review. Food Biosci. 2020, 36, 100633. [Google Scholar] [CrossRef]
- Ren, Y.; Yao, M.; Chang, P.; Sun, Y.; Li, R.; Meng, D.; Xia, X.; Wang, Y. Isolation and characterization of a Pseudomonas poae JSU-Y1 with patulin degradation ability and biocontrol potential against Penicillium expansum. Toxicon 2021, 195, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ashrafi-Saiedlou, S.R.-S.; Mirhassan, Fattahi; Mohammad Ghosta, Youbert. Biosynthesis and characterization of iron oxide nanoparticles fabricated using cell-free supernatant of Pseudomonas fluorescens for antibacterial, antifungal, antioxidant, and photocatalytic applications. Sci. Rep. 2025, 15, 1018. [Google Scholar] [CrossRef]
- Luz, C.; Saladino, F.; Luciano, F.B.; Mañes, J.; Maca, G. In vitro antifungal activity of bioactive peptides produced by Lactobacillus plantarum against Aspergillus parasiticus and Penicillium expansum. LWT Food Sci. Technol. 2017, 81, 128–135. [Google Scholar] [CrossRef]
- Zhang, S.; Zheng, Q.; Xu, B.; Liu, J. Identification of the Fungal Pathogens of Postharvest Disease on Peach Fruits and the Control Mechanisms of Bacillus subtilis JK-14. Toxins 2019, 11, 322–337. [Google Scholar] [CrossRef]
- Wang, L.; Jin, P.; Wang, J.; Jiang, L.; Zhang, S.; Gong, H.; Liu, H.; Zheng, Y. In vitro inhibition and in vivo induction of defense response against Penicillium expansum in sweet cherry fruit by postharvest applications of Bacillus cereus AR156. Postharvest Biol. Technol. 2015, 101, 15–17. [Google Scholar] [CrossRef]
- Wang, X.; Xu, F.; Wang, J.; Jin, P.; Zheng, Y. Bacillus cereus AR156 induces resistance against Rhizopus rot through priming of defense responses in peach fruit. Food Chem. 2013, 136, 400–406. [Google Scholar] [CrossRef]
- Marzouk, T.; Chaouachi, M.; Sharma, A.; Jallouli, S.; Mhamdi, R.; Kaushik, N.; Djébali, N. Biocontrol of Rhizoctonia solani using volatile organic compounds of solanaceae seed-borne endophytic bacteria. Postharvest Biol. Technol. 2021, 181, 111655. [Google Scholar] [CrossRef]
- Dobrzynski, J.; Jakubowska1, Z.; Kulkova, I.; Kowalczyk, P.; Kramkowski, K. Biocontrol of fungal phytopathogens by Bacillus pumilus. Front. Microbiol. 2023, 14, 1194606. [Google Scholar] [CrossRef] [PubMed]
- Pedretti, N.; Iseppi, R.; Condò, C.; Spaggiari, L.; Messi, P.; Pericolini, E.; Di Cerbo, A.; Ardizzoni, A.; Sabia, C. Cell-Free Supernatant from a Strain of Bacillus siamensis Isolated from the Skin Showed a Broad Spectrum of Antimicrobial Activity. Microorganisms 2024, 12, 718. [Google Scholar] [CrossRef]
- Li, Z.L.; Jiang, J.M.; Sun, K.Y.; Ye, S.H. Bacillus amyloliquefaciens LJ1 from Nanguo Pear: Suppressing Penicillium expansum colonization and degrading patulin in postharvest disease management. Food Control 2026, 179, 111564. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, L.; Liang, W.; Liu, M. Biocontrol potential of endophytic Bacillus velezensis strain QSE-21 against postharvest grey mould of fruit. Biol. Control 2021, 161, 104711. [Google Scholar] [CrossRef]
- Li, K.Y.; Cheng, S.; Liu, Z.; Pan, Q.Y.; Zuo, X.F.; Guo, A.L.; Lv, J. Characteristics of inhibition of Aspergillus flavus growth and degradation of aflatoxin B1 by cell-free fermentation supernatant of Bacillus velezensis 906. Food Biosci. 2024, 61, 104954. [Google Scholar] [CrossRef]
- Hirozawa, M.T.; Ono, M.A.; Suguiura, I.M.D.; Bordini, J.G.; Hirooka, E.Y.; Ono, E.Y.S. Antifungal effect and some properties of cell-free supernatants of two Bacillus subtilis isolates against Fusarium verticillioides. Braz. J. Microbiol. 2024, 55, 2527–2538. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Liu, K.; Fan, Y.; Cao, J.; Li, H.; Song, W.; Liu, Y.; Miao, M. Cell-free supernatant of Bacillus velezensis suppresses mycelial growth and reduces virulence of Botrytis cinerea by inducing oxidative stress. Front. Microbiol. 2022, 13, 980022. [Google Scholar] [CrossRef]
- Fan, Y.; Liu, K.; Lu, R.; Gao, J.; Song, W.; Zhu, H.; Tang, X.; Liu, Y.; Miao, M. Cell-Free Supernatant of Bacillus subtilis Reduces Kiwifruit Rot Caused by Botryosphaeria dothidea through Inducing Oxidative Stress in the Pathogen. J. Fungi 2023, 9, 127. [Google Scholar] [CrossRef]
- Alori, E.T.; Onaolapo, A.O.; Ibaba, A.L. Cell free supernatant for sustainable crop production. Front. Sustainable Food Syst. 2025, 9, 1549048. [Google Scholar] [CrossRef]
- Patel, N.; Oudemans, P.; Hillman, B.; Kobayashi, D. Use of the tetrazolium salt MTT to measure cell viability effects of the bacterial antagonist Lysobacter enzymogenes on the filamentous fungus Cryphonectria parasitica. Anton. Leeuw. Int. J. G. 2013, 103, 1271–1280. [Google Scholar] [CrossRef]
- Li, H.; Su, Q.; Huang, Z.; Yu, S.; Liu, Y.; Miao, M. Calcium Chloride Enhances Defense Response Against Diaporthe nobilis by Eliciting Reactive Oxygen Species in Kiwifruit. J. Food Process. Preserv. 2025, 2025, 8135751. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Jiao, W.; Liu, X.; Chen, Q.; Du, Y.; Li, Y.; Yue, F.; Dong, X.; Fu, M. Epsilon-poly-l-lysine (ε-PL) exhibits antifungal activity in vivo and in vitro against Botrytis cinerea and mechanism involved. Postharvest Biol. Technol. 2020, 168, 111270. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, X.; Chen, T.; Xu, Y.; Tian, S. Antifungal effects of hinokitiol on development of Botrytis cinerea in vitro and in vivo. Postharvest Biol. Technol. 2020, 159, 111038. [Google Scholar] [CrossRef]
- Levin, E.; Kishore, A.; Ballester, A.R.; Raphael, G.; Feigenberg, O.; Liu, Y.; Norelli, J.; Gonzalez-Candelas, L.; Wisniewski, M.; Droby, S. Identification of pathogenicity-related genes and the role of a subtilisin-related peptidase S8 (PePRT) in authophagy and virulence of Penicillium expansum on apples. Postharvest Biol. Technol. 2019, 149, 209–220. [Google Scholar] [CrossRef]
- Zhang, X.; Zong, Y.; Gong, D.; Yu, L.; Sionov, E.; Bi, Y.; Prusky, D. NADPH Oxidase Regulates the Growth and Pathogenicity of Penicillium expansum. Front. Plant Sci. 2021, 12, 696210. [Google Scholar] [CrossRef]
- Li, H.; Tian, S.; Qin, G. NADPH Oxidase Is Crucial for the Cellular Redox Homeostasis in Fungal Pathogen Botrytis cinerea. Mol Plant Microbe Interact 2019, 32, 1508–1516. [Google Scholar] [CrossRef]
- Tsiatsiani, L.; Van Breusegem, F.; Gallois, P.; Zavialov, A.; Lam, E.; Bozhkov, P.V. Metacaspases. Cell Death and Differentiation 2011, 18, 1279–1288. [Google Scholar] [CrossRef]
- Yue, J.-y.; Wang, W.-w.; Jie, X.-r.; Gao, Z.-x.; Wang, H.-z. The metacaspase TaMCA1-mediated crosstalk between autophagy and PCD contributes to the defense response of wheat seedlings against powdery mildew. Int. J. Biol. Macromol. 2025, 292, 139265. [Google Scholar] [CrossRef] [PubMed]
- Alkilayh, O.A.; Hamed, K.E.; Sayyed, R.Z.; Abdelaal, K.; Omar, A.F. Characterization of Botrytis cinerea, the causal agent of tomato grey mould, and its biocontrol using Bacillus subtilis. Physiol. Mol. Plant Pathol. 2024, 133, 102376. [Google Scholar] [CrossRef]
- Lu, Z.; Chen, M.; Long, X.; Yang, H.; Zhu, D. Biological potential of Bacillus subtilis BS45 to inhibit the growth of Fusarium graminearum through oxidative damage and perturbing related protein synthesis. Front. Microbiol. 2023, 14–2023, 1064838. [Google Scholar] [CrossRef]
- Song, J.; Chen, J.Y.; Zhang, D.D.; Li, R.; Kong, Z.Q.; Zhu, H.; Dai, X.F.; Han, D.F.; Wang, D. Genome Resource of Bacillus subtilis KRS015, a Potential Biocontrol Agent for Verticillium dahliae. Phytofrontiers 2024, 4, 443–448. [Google Scholar] [CrossRef]
- Lu, Y.Y.; Ma, D.T.; He, X.; Wang, F.; Wu, J.R.; Liu, Y.; Jiao, J.Y.; Deng, J. Bacillus subtilis KLBC BS6 induces resistance and defence-related response against Botrytis cinerea in blueberry fruit. Physiol. Mol. Plant Pathol. 2021, 114, 101599. [Google Scholar] [CrossRef]
- Castro, B.; Citterico, M.; Kimura, S.; Stevens, D.M.; Coaker, G.L. Stress-induced reactive oxygen species compartmentalization, perception and signalling. Nat. Plants 403–412. [CrossRef] [PubMed]
- Nigam, M.; Punia, B.; Dimri, D.B.; Mishra, A.P.; Radu, A.-F.; Bungau, G. Reactive oxygen species: A double-edged sword in the modulation of cancer signaling pathway dynamics. Cells 2025, 14, 1207. [Google Scholar] [CrossRef]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Biodivers. 2020, 21, 363–383. [Google Scholar] [CrossRef] [PubMed]
- Monier, B.; Suzanne, M. The Morphogenetic Role of Apoptosis. Curr. Top. Dev. Biol. 2015, 114, 335–362. [Google Scholar]
Figure 1.
Effects of varying CFS concentrations on spore germination. (A) Effects of different concentrations of CFS on spore germination (B) Statistical chart of the effect of different concentrations of CFS on spore germination rate; (C) Graph showing the statistical analysis of germ tube length influenced by varying concentrations of CFS; different letters indicated statistically significant differences between groups (P < 0.05).
Figure 1.
Effects of varying CFS concentrations on spore germination. (A) Effects of different concentrations of CFS on spore germination (B) Statistical chart of the effect of different concentrations of CFS on spore germination rate; (C) Graph showing the statistical analysis of germ tube length influenced by varying concentrations of CFS; different letters indicated statistically significant differences between groups (P < 0.05).
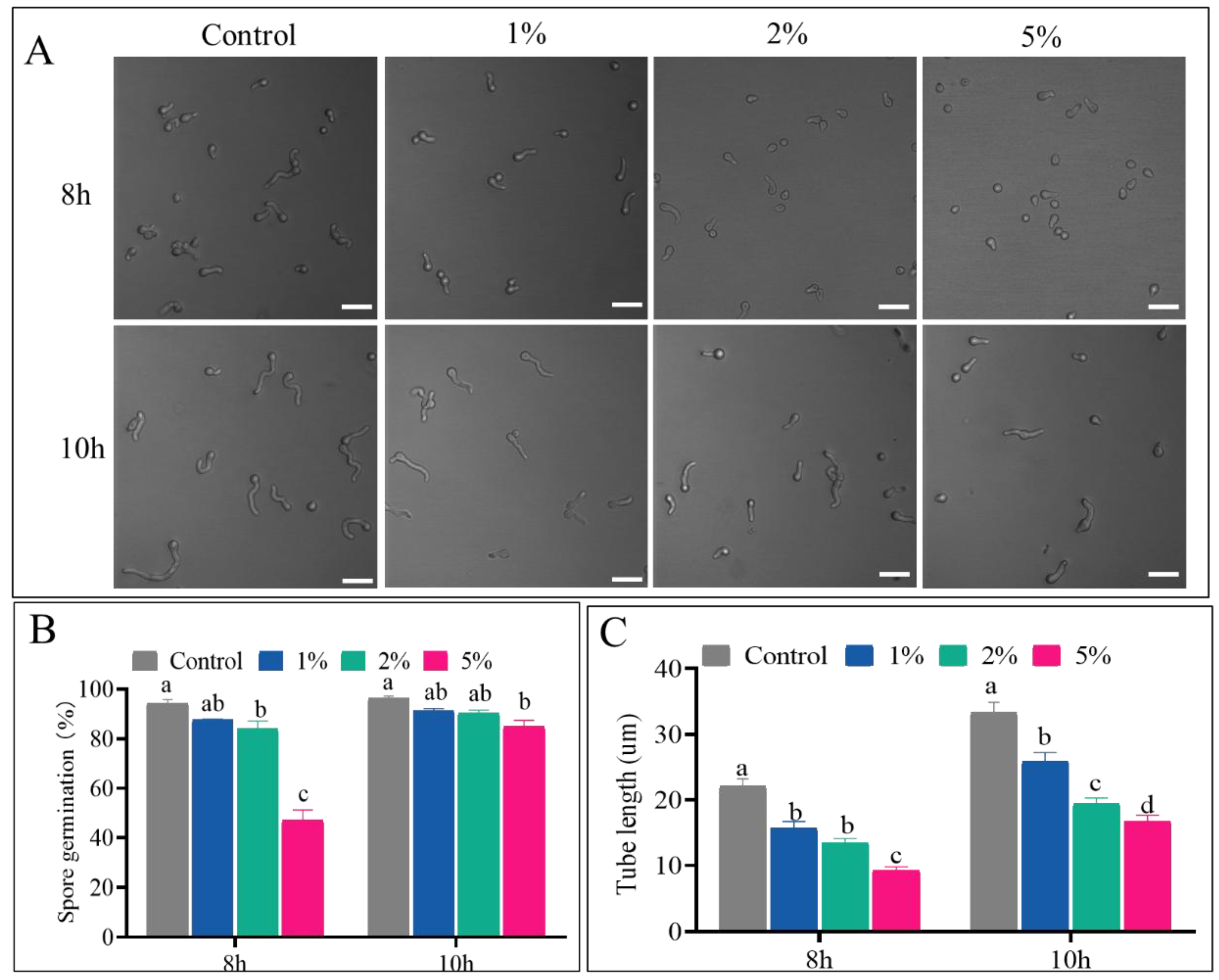
Figure 2.
Effects of CFS on colony growth, dry weight and mycelial activity. (A) colony growth recorded at 48 h-144 h; (B) statistics analysis of colony diameter; (C, D) Mycelium dry weight recorded and statistics analysis; (E) mycelial activity analysis; different letters indicated significant differences between groups (P < 0.05).
Figure 2.
Effects of CFS on colony growth, dry weight and mycelial activity. (A) colony growth recorded at 48 h-144 h; (B) statistics analysis of colony diameter; (C, D) Mycelium dry weight recorded and statistics analysis; (E) mycelial activity analysis; different letters indicated significant differences between groups (P < 0.05).
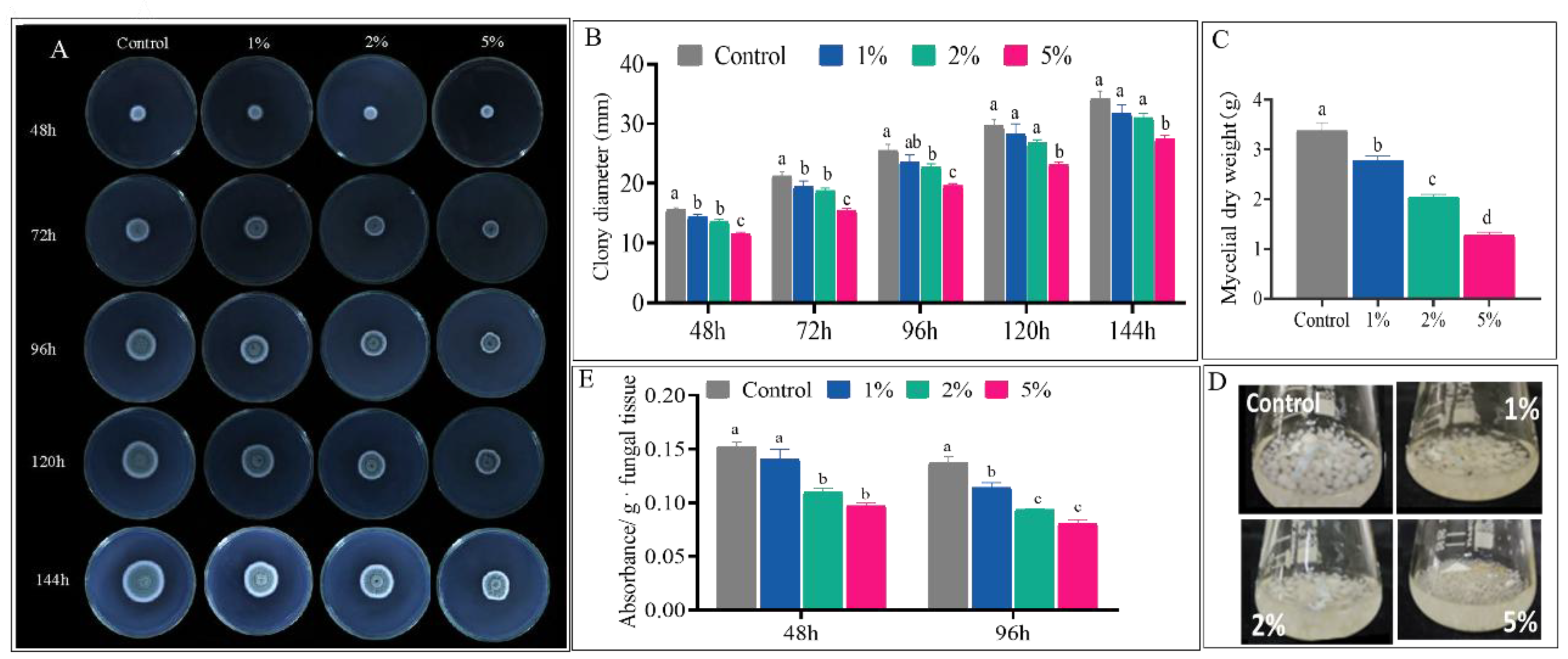
Figure 3.
Effect of 5% CFS on pathogenicity of P. expansum on grape and citrus. (A) Disease symptoms on grape after 3 days; (B) Statistics of grape incidence after 3 days treatment with 5% CFS; (C) Disease symptoms on citrus after 3, 4, 5, 6 days; (D) and (E) were disease incidence and lesion diameter statistics. ** indicated significant difference between groups (P < 0.01).
Figure 3.
Effect of 5% CFS on pathogenicity of P. expansum on grape and citrus. (A) Disease symptoms on grape after 3 days; (B) Statistics of grape incidence after 3 days treatment with 5% CFS; (C) Disease symptoms on citrus after 3, 4, 5, 6 days; (D) and (E) were disease incidence and lesion diameter statistics. ** indicated significant difference between groups (P < 0.01).
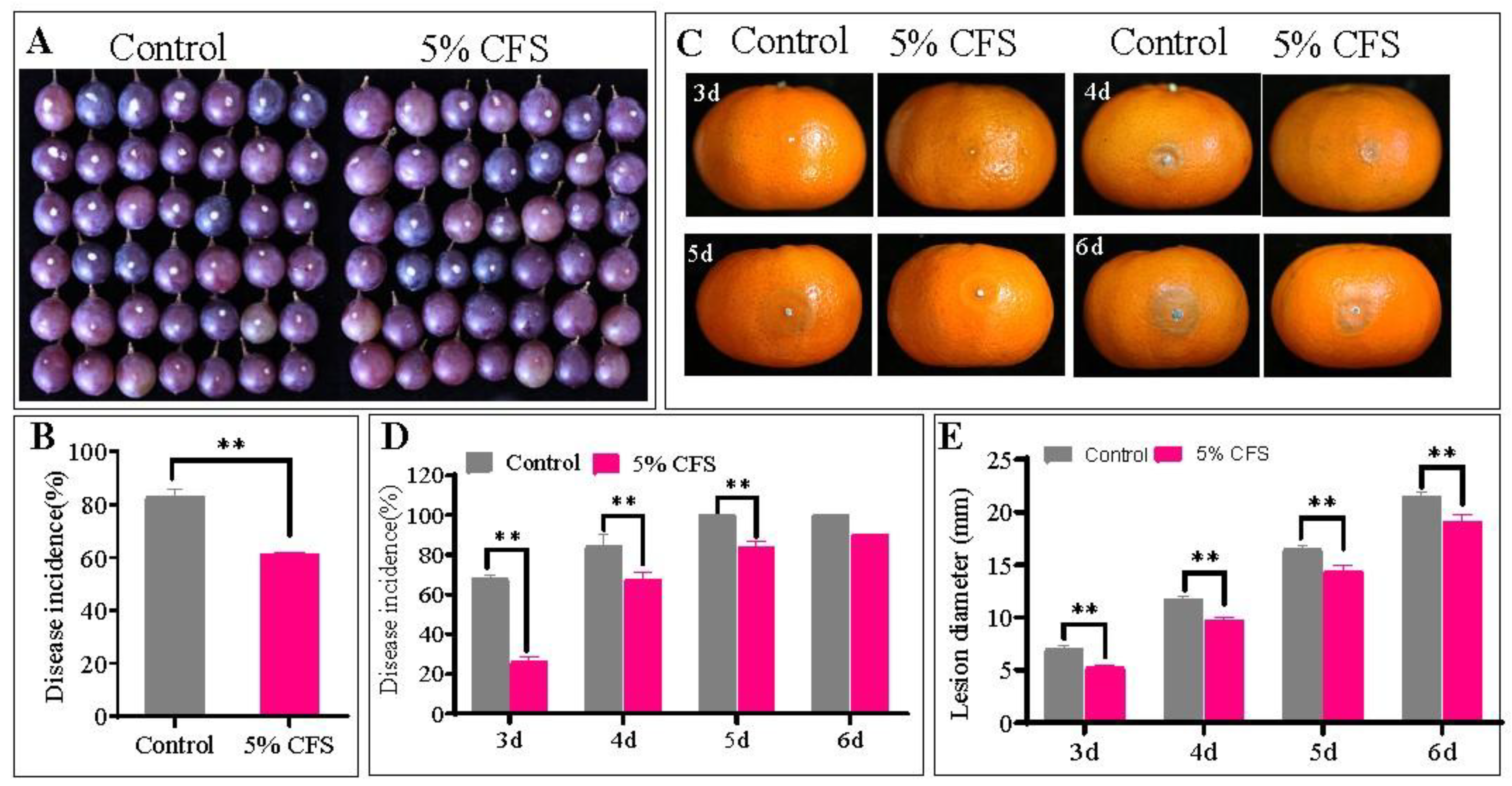
Figure 4.
CFS on ROS accumulation (A), the relative intensity of fluorescence (B), and relative gene expression of PeNoxA (C), PeNoxR (D), PeRacA (E). Different letters indicated significant differences between groups (P < 0.05). * indicates a significant difference between groups (P < 0.05), while ** indicates significant difference between groups (P < 0.01).
Figure 4.
CFS on ROS accumulation (A), the relative intensity of fluorescence (B), and relative gene expression of PeNoxA (C), PeNoxR (D), PeRacA (E). Different letters indicated significant differences between groups (P < 0.05). * indicates a significant difference between groups (P < 0.05), while ** indicates significant difference between groups (P < 0.01).
Figure 5.
CFS on PI staining (A), the relative intensity of fluorescence (B), and conductivity (C), nucleic acid (D), protein release (E) and MDA accumulation (F) of P. expansum. Bar = 20 μm. Different letters indicated significant differences between groups (P < 0.05). * means significant difference between groups (P < 0.05), ** means significant difference between groups (P < 0.01).
Figure 5.
CFS on PI staining (A), the relative intensity of fluorescence (B), and conductivity (C), nucleic acid (D), protein release (E) and MDA accumulation (F) of P. expansum. Bar = 20 μm. Different letters indicated significant differences between groups (P < 0.05). * means significant difference between groups (P < 0.05), ** means significant difference between groups (P < 0.01).
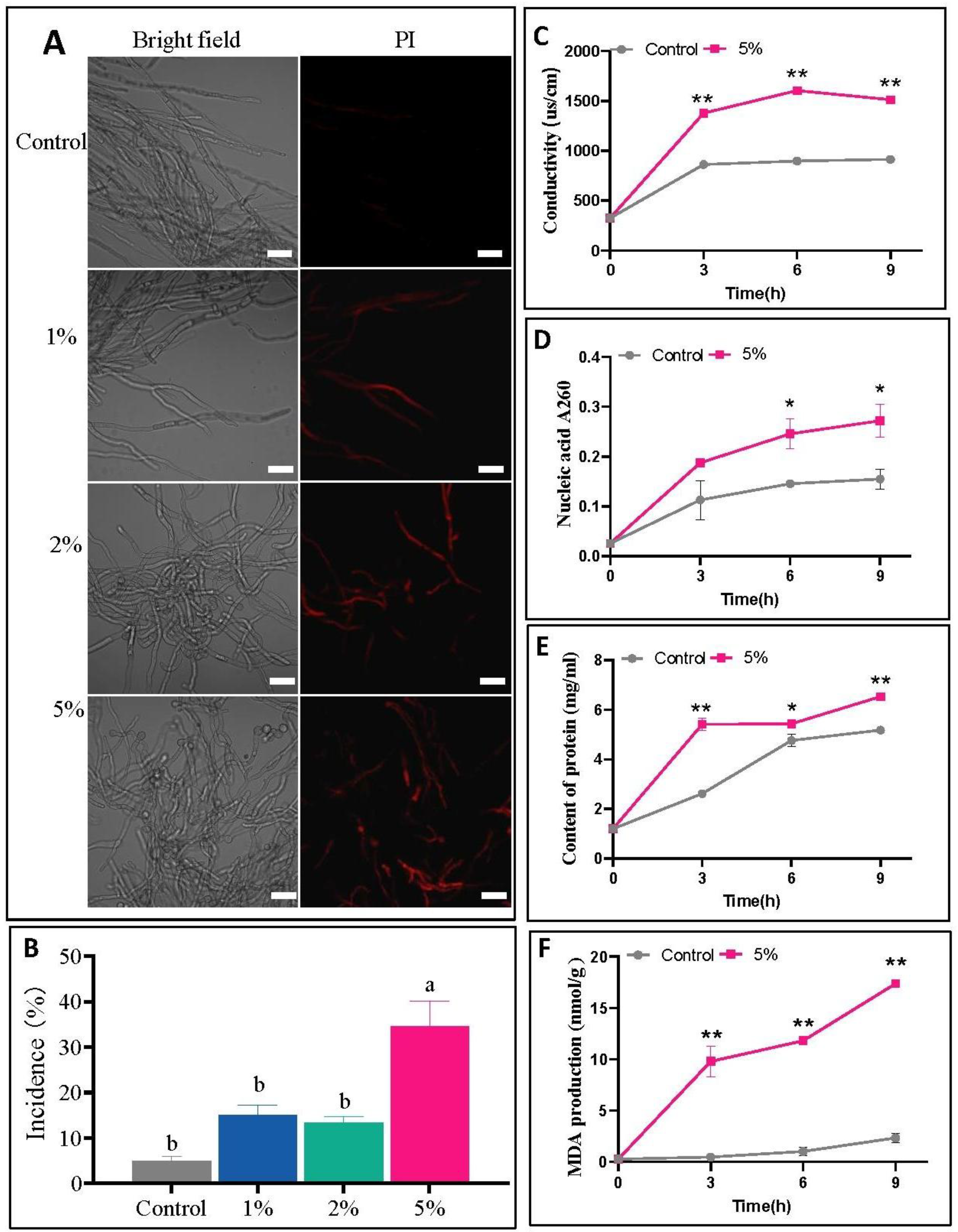
Figure 6.
Effect of CFS on the morphology (A, B) and microstructure (C, D) of mycelium. CW: cell wall; PM: cell membrane; LD: lipid droplets; M: mitochondria; RER: endoplasmic reticulum; AP: autophagosome; N: nucleus; ASS: Auto phagolysosomes.
Figure 6.
Effect of CFS on the morphology (A, B) and microstructure (C, D) of mycelium. CW: cell wall; PM: cell membrane; LD: lipid droplets; M: mitochondria; RER: endoplasmic reticulum; AP: autophagosome; N: nucleus; ASS: Auto phagolysosomes.
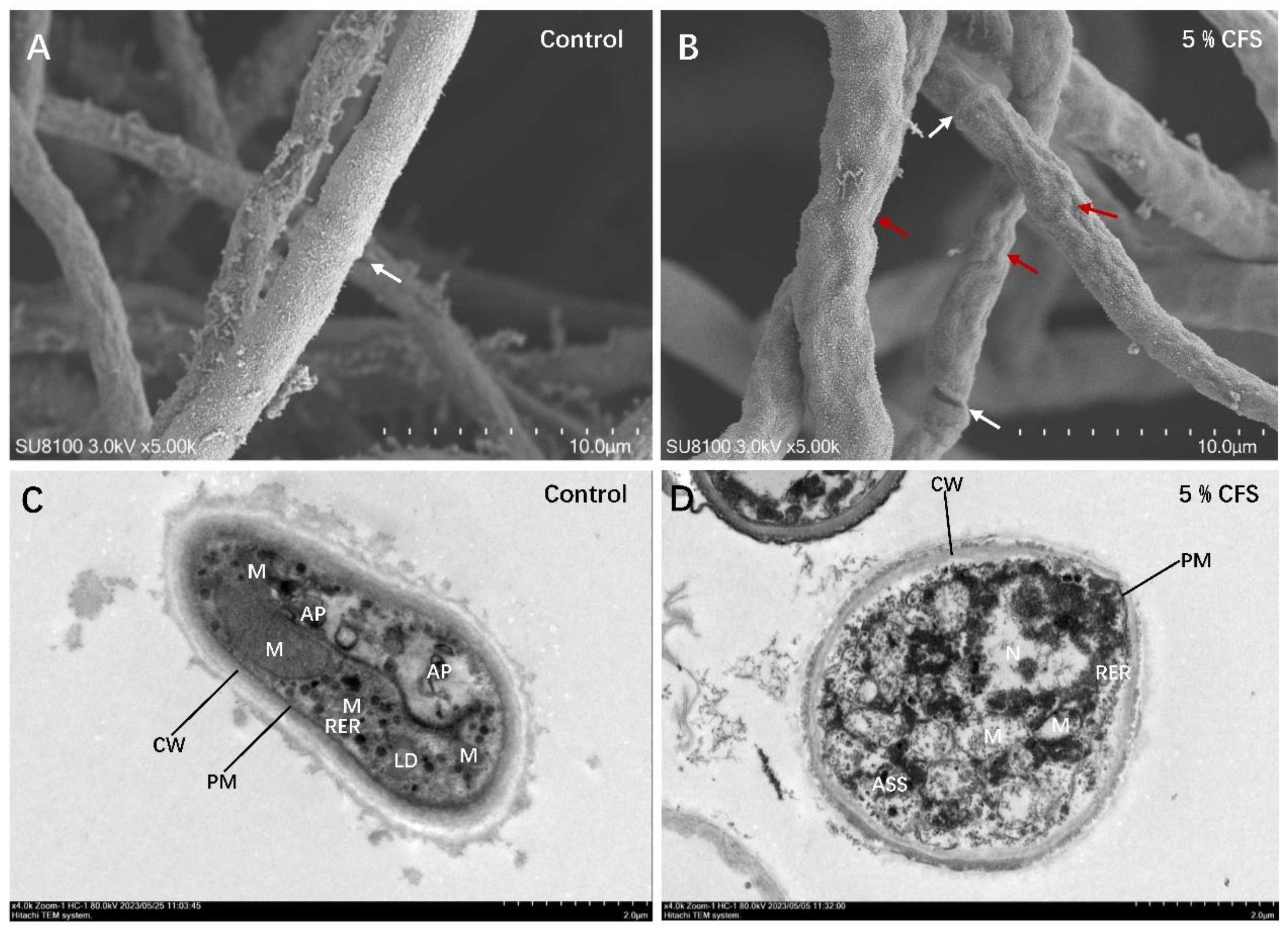
Figure 7.
Effect of 5% CFS on relative gene expression of PePRT (A), PeMetacaspase1 (B), and PeMetacaspase2 (C). ** indicates significant difference between groups (P < 0.01). Vertical bars represent the standard deviations of the mean values.
Figure 7.
Effect of 5% CFS on relative gene expression of PePRT (A), PeMetacaspase1 (B), and PeMetacaspase2 (C). ** indicates significant difference between groups (P < 0.01). Vertical bars represent the standard deviations of the mean values.
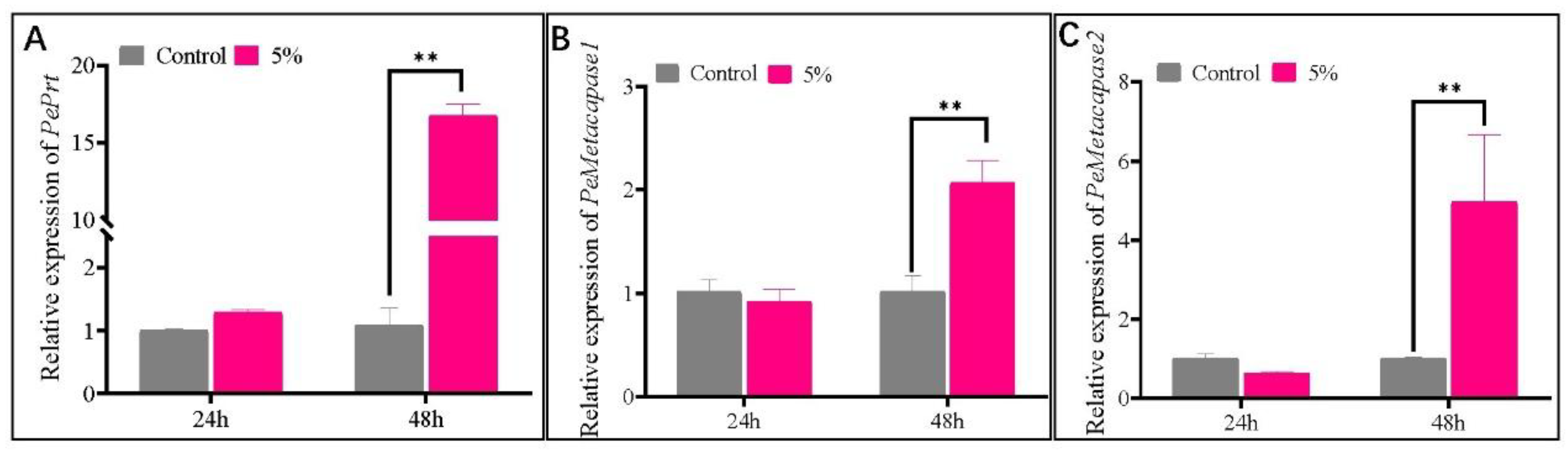

Table 1.
Primer sequences employed in RT-qPCR assays.
| Gene name | Accession No. | Primer name | Sequence of primer(5’-3’) |
| PePRT [36] | PEX2_027670 | Pe-prtF | CCGATGTTACCCCTAAGCAG |
| Pe-prtR | AGGATCTGAGTGTAATTGGCG | ||
| PeNoxA [37] | PEX2_053880 | Pe-NoxAF | CATTAGATGAGTCGGCGTGG |
| Pe-NoxAR | CAAGTTCTGGGCGGATATGG | ||
| PeNoxR [37] | PEX2_056490 | Pe-NoxRF | CTCTGAAGATGAAGGTGCAGG |
| Pe-NoxRR | AACGCTCTTCCACCCATATC | ||
| PeRacA [37] | PEX2_019970 | Pe-RacAF | GTACACAACGAATGCTTTCCC |
| Pe-RacAR | GATCGTAATCCTCTTGTCCAGC | ||
| PeMetacaspased1 | XM_016744186.1 | CASP1-F | TGATGTTTTCAGGGTCCAAGG |
| CASP1-R | CATTTCGGATCGTGTTCAGC | ||
| PeMetacaspased2 | XM_016742860.1 | CASP2-F | ACCAGCAAAACCCGATGAG |
| CASP2-R | TTCGTCACCATCCAAGTCG | ||
| Isy1 | PEX2-072240 | Isy1-F | CAAAGCCTGAGCGACTACCA |
| Isy1-R | CGCCCTTCATCGTCGTAAA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



