
Submitted:
01 January 2026
Posted:
02 January 2026
You are already at the latest version
Abstract
Antimicrobial resistance [AMR] is a silent yet intensifying global threat, with particularly severe consequences in tropical and subtropical ecosystems, where high ecological connectivity, widespread antimicrobial use, and inadequate sanitation create ideal conditions for the persistence and spread of antimicrobial resistance genes [ARGs]. Within the One Health framework, migratory birds warrant special attention because they traverse tropical AMR hotspots, linking contaminated aquatic, agricultural, and peri-urban environments along established flyways. Evidence from tropical regions demonstrates that migratory birds frequently carry clinically meaningful ARGs, including extended-spectrum β-lactamases [ESBLs], carbapenemases, and colistin resistance [mcr] genes, highlighting their role as biological connectors that redistribute resistant bacteria between human-dominated and natural ecosystems and contribute to the expansion of the global resistome. Addressing the complex interface among AMR, migratory birds, and ARGs requires integrative surveillance strategies that explicitly incorporate wildlife into existing health systems. Genomic and metagenomic monitoring of migratory bird populations, combined with cross-sectoral data sharing, can provide early warning signals of emerging resistance patterns and inform evidence-based interventions. Understanding the ecological role of migratory birds in tropical ecosystems is therefore essential for designing effective One Health strategies to curb transboundary AMR dissemination and preserve the long-term efficacy of antimicrobial therapies.
Keywords:
antimicrobial resistance [AMR]
; antimicrobial resistance genes [ARGs]
; migratory birds
; tropical ecosystem
; gene transfer
; zoonotic pathogens
; One Health
1. Introduction
Antimicrobial resistance [AMR] has emerged as one of the most critical global health challenges of the 21st century, undermining the efficacy of antibiotics that revolutionized modern medicine [1]. The World Health Organization has characterized AMR as a ‘silent pandemic’ with the potential to reverse decades of therapeutic progress and drive global mortality rates beyond those of tuberculosis, HIV/AIDS, and malaria by 2050 [2,3]. Although AMR is a global phenomenon, its effects are particularly severe in tropical and subtropical regions, where high population density, unregulated antimicrobial use, and weak sanitation infrastructure create ideal conditions for the emergence and dissemination of resistant pathogens [4,5]. In addition, tropical ecosystems sustain warm temperatures, high microbial diversity, and year-round aquatic productivity—environmental characteristics known to accelerate bacterial growth, horizontal gene transfer [HGT], and the persistence of antimicrobial resistance genes [ARGs] [6,7].
The persistence and global spread of ARGs are driven by complex ecological and evolutionary processes that extend beyond clinical environments [8]. Mobile genetic elements [MGEs], including plasmids, transposons, integrons, and bacteriophages, facilitate HGT and enable bacteria to acquire resistance traits across taxonomic and ecological boundaries[8,9]. Through this genetic mobility, hospitals, farms, wastewater systems, and natural habitats become linked within a single, interconnected global resistome [10]. Importantly, tropical regions often exhibit higher ARG and MGE abundance than temperate regions, reflecting climatic and ecological conditions that favour rapid microbial turnover and gene exchange [11].
Within this interconnected resistome, wildlife represents an often overlooked yet critical component of AMR ecology [12,13,14]. Among wildlife taxa, migratory birds warrant particular attention because they routinely traverse multiple ecological zones, including many of the world’s most contaminated tropical wetlands, rice paddies, deltas, and coastal lagoons—environments where exposure to ARGs is consistently high [12,15,16]. Through the ingestion of contaminated food or water, the gastrointestinal tracts of migratory birds may become colonized by antimicrobial-resistant bacteria [ARB], allowing these species to function as mobile reservoirs and dispersal agents of ARGs [17].
Each year, billions of migratory birds travel along major flyways spanning Africa, Asia, Europe, and the Americas, frequenting wetlands, agricultural landscapes, and urban waste sites often contaminated with antibiotic residues and resistant microorganisms [18,19,20]. Studies conducted along tropical flyways report widespread detection of clinically relevant ARGs through fecal shedding, environmental contamination, or direct interface with human-dominated ecosystems [18]. Through their transcontinental movements, migratory birds link distant ecosystems and facilitate the long-range redistribution of resistant bacteria and genetic material across ecological and geopolitical boundaries [17,19,20].
Despite this growing body of evidence, the role of migratory birds in the dissemination of AMR remains inadequately represented in global surveillance systems. While the One Health paradigm emphasizes the interconnectedness of human, animal, and environmental health [21], wildlife, particularly migratory birds traversing heavily impacted tropical ecosystems, is largely excluded from national and international AMR reporting frameworks such as WHO-GLASS [14]. At key tropical interfaces, including wetlands, rice paddies, and coastal lagoons, ecological and anthropogenic pressure converge, promoting bidirectional transfer of resistant bacteria and genes among wildlife, livestock, and human populations [22]. Surveillance efforts addressing these interfaces remain fragmented, geographically uneven, and disproportionately focused on high-income regions, despite the fact that many major migratory flyways intersect tropical areas where ARG contamination is most intense [14,23]
Against this backdrop, the present review consolidates current evidence on the role of migratory birds in the global dissemination of antimicrobial-resistant bacteria and ARGs, with a specific focus on tropical and subtropical flyways [19]. It addresses three guiding questions: [i] what evidence demonstrates that migratory birds in tropical ecosystems carry clinically relevant ARGs; [ii] which ecological interfaces and anthropogenic drivers facilitate ARG acquisition and dispersal; and [iii] how migratory birds can be effectively integrated into One Health–oriented AMR surveillance frameworks [24,25]. By synthesizing findings from surveillance studies and molecular epidemiology, this review highlights critical knowledge gaps and underscores the urgent need for harmonized, wildlife-inclusive AMR monitoring strategies that capture the transboundary flow of resistance across tropical ecosystems.
2. Antimicrobial Resistance [AMR] and Antimicrobial Resistance Genes [ARGs]
2.1. Mechanisms of Resistance
Antimicrobial resistance in bacteria associated with migratory birds results from a limited set of well-characterized mechanisms, including enzymatic inactivation of antibiotics, modification of antimicrobial targets, decreased membrane permeability, and active efflux [22,23,24,25,26,27]. In tropical aquatic environments, where resistant bacteria are prevalent, these mechanisms support colonization of avian gastrointestinal tracts and promote the persistence and dissemination of antimicrobial resistance genes [ARGs] across ecological boundaries ([28]; Table 1).
2.1.1. Enzymatic Degradation and β-Lactamases
Enzymatic degradation mediated by β-lactamases represents one of the most frequently documented resistance mechanisms in bacteria isolated from migratory birds [29,30]. These enzymes confer resistance to widely used β-lactam antibiotics and have been commonly reported in Enterobacteriaceae recovered from avian fecal samples, particularly in birds inhabiting or frequenting contaminated aquatic environments [31]. From an ecological perspective, the relevance of β-lactamases in migratory birds lies primarily in their environmental mobility rather than in their biochemical diversity [32]. β-lactamase–producing bacteria, including those carrying extended-spectrum β-lactamases [ESBLs] and carbapenemases, are consistently detected in tropical wetlands, wastewater-impacted waters, and agricultural runoff—key foraging and roosting habitats for migratory birds [33]. These environments facilitate the acquisition, persistence, and subsequent long-distance dispersal of clinically relevant resistance genes along migratory flyways.
2.1.2. Target Modification and Efflux Systems
Resistance in bacteria associated with migratory birds can also arise through modification of antimicrobial target sites and activation of efflux systems, mechanisms that reduce antibiotic susceptibility by limiting effective drug binding or intracellular accumulation [34]. Such resistance traits have been reported in bacteria recovered from avian fecal samples and from contaminated aquatic environments frequented by migratory birds, where persistent antibiotic pressure favors their maintenance [35,36]. In polluted tropical habitats, including wetlands, wastewater-impacted waters, and agricultural runoff, these mechanisms enhance bacterial persistence and contribute to the acquisition and retention of multidrug-resistant strains within avian hosts. Through repeated exposure at key foraging and roosting sites, migratory birds may subsequently facilitate the dispersal of these resistance traits along established flyways.
2.1.3. Mobile Genetic Elements: Plasmids, Integrons, and Transposons
Mobile genetic elements [MGEs] play a central role in shaping antimicrobial resistance profiles in bacteria associated with migratory birds by facilitating the transfer of antimicrobial resistance genes [ARGs] across environmental and host-associated microbial communities [37,38]. In tropical aquatic and terrestrial habitats frequently used by migratory birds, plasmid- and transposon-mediated gene exchange supports the persistence of resistance traits within bacterial populations exposed to sustained antimicrobial inputs [39]. Warm temperatures, high microbial diversity, and the presence of antibiotic residues in soil and water further enhance ARG retention and recombination, increasing the likelihood that migratory birds acquire and subsequently disseminate ARG-bearing bacteria along their migratory flyways [40,41,42].
2.2. Diminished Permeability and Limited Uptake
Reduced membrane permeability contributes to intrinsic antimicrobial resistance by limiting antibiotic entry into bacterial cells [43]. Such traits have been reported in Enterobacteriaceae isolated from tropical aquatic environments impacted by human and livestock wastewater, habitats commonly used by migratory birds for foraging and roosting [44]. Exposure to these contaminated systems may therefore facilitate the acquisition and persistence of intrinsically resistant bacteria within avian hosts, supporting their continued circulation along migratory flyways.
2.3. Horizontal Gene Transfer [HGT]
HGT is a key driver of antimicrobial resistance gene [ARG] dissemination within environmental bacterial communities and underlies the emergence of multidrug-resistant bacteria encountered by migratory birds [10,56,57,58,59]. In tropical aquatic and sedimentary environments frequented by migratory birds, conditions such as high nutrient loads, warm temperatures, and dense microbial assemblages promote frequent gene exchange and long-term ARG persistence [6,60,61,62,63]. These settings increase the likelihood that migratory birds acquire bacteria carrying transferable resistance determinants, thereby facilitating the redistribution of ARGs across geographic regions through migratory movement.

Table 1.
Central mechanisms of antimicrobial resistance [AMR], molecular basis, mechanism of action, and representative antibiotics affected.
Table 1.
Central mechanisms of antimicrobial resistance [AMR], molecular basis, mechanism of action, and representative antibiotics affected.
| Mechanism of Resistance | Representative Genes or Enzymes | Mechanism of Action | Representative Antibiotics | References |
|---|---|---|---|---|
| 1. Enzymatic inactivation | blaTEM, blaSHV, blaCTX-M[ESBLs]; blaNDM, blaKPC, blaOXA-48 | Hydrolysis of β-lactam ring or other structural modification that inactivates the antibiotic | Penicillins, Cephalosporins, Carbapenems | [31,32,33,45,46] |
| 2. Target modification | ermB [23S rRNA methylation]; qnrA/B/S [DNA gyrase protection]; tetM [ribosomal protection protein] | Alteration or protection of antibiotic binding sites reduces affinity for the drug target | Macrolides, Quinolones, Tetracyclines | [34] |
| 3. Efflux pump activation | Multidrug efflux systems — ABC, MFS, RND families [acrAB-tolC, norA, mexAB-oprM] | Active transport of antibiotics out of the cell, lowering intracellular concentration | Fluoroquinolones, Tetracyclines, Chloramphenicol | [47,48,49,50] |
| 4. Mobile genetic elements | Broad-host-range and conjugative plasmids [IncF, IncI, IncA/C types] | Horizontal transfer of ARGs via conjugation between bacteria | Multidrug resistance [across classes] | [51,52,53,54,55] |
| 5. Integrons | intI1 and associated gene cassettes | Site-specific recombination | Multiple antibiotic classes | [39,40] |
| 6. Transposons | Tn3, Tn21, and related insertion sequences | Movement of ARGs between plasmids and chromosomes | Multiple antibiotic classes. | [41,42] |
3. Environmental Interface and Migratory Birds in AMR Dissemination
Antimicrobial resistance [AMR] extends beyond clinical settings and has become a pervasive ecological challenge shaped by interactions among humans, animals, and the environment within the One Health continuum [71,72]. The natural environment functions as both a reservoir and conduit for ARGs, continually enriched by antibiotic residues, resistant bacteria, and mobile genetic elements released from hospitals, livestock operations, aquaculture systems, and municipal wastewater [6,35,64]. These anthropogenic inputs generate intense selective pressures that accelerate bacterial adaptation and horizontal gene transfer within environmental microbial communities [65,66,67,68]. Tropical wetlands, estuaries, and wastewater-impacted habitats are particularly vulnerable to AMR enrichment. Warm temperatures, high organic loads, and continuous water availability support dense microbial populations and enhance ARG persistence, allowing these ecosystems to function as ecological mixing zones where resistant strains emerge and circulate [70,73,74,75,76,77]. Such conditions amplify ARG mobility and increase exposure risk for wildlife species that rely on these habitats.
At the wildlife–environment interface, migratory birds serve as biological connectors, linking human-impacted environments across continents [78,79,80]; specifically, species that feed, roost, or nest near landfills, sewage lagoons, aquaculture ponds, and agricultural fields risk acquiring multidrug-resistant bacteria that harbor clinically relevant resistance determinants, such as β-lactamase and colistin resistance genes, reflecting exposure to anthropogenically contaminated environments [12,14,17,18,19,79,81,82,83,84,85,86,87,88]. Following acquisition, these birds may disseminate resistant microorganisms across ecological and geopolitical boundaries through fecal shedding during migration, thereby linking highly contaminated tropical sites with distant ecosystems [17,18,23,89].
Recent genomic studies conducted across Africa, Europe, and Asia have identified antimicrobial resistance gene [ARG] signatures in migratory birds that closely resemble those observed in local wastewater isolates, indicating active exchange between anthropogenic sources and wildlife along tropical flyways [12,79,90,91,92]. This genomic overlap reflects sustained exposure of migratory birds to contaminated tropical wetlands, wastewater-impacted waters, and other human-influenced aquatic environments, where resistant bacteria and ARGs are abundant. Species such as gulls, waterfowl, and shorebirds that forage in these habitats frequently harbor extended-spectrum β-lactamase–producing Escherichia coli and carbapenemase-producing Enterobacterales, mirroring resistance profiles reported in human and livestock populations [24,74,75,91,93,94,95,96,97].
Within avian gastrointestinal tracts, microbiome analyses indicate that ARGs can persist and undergo horizontal transfer, supporting long-distance carriage during migration and subsequent deposition into new environments [12,20,23,98]. The ecological plasticity of migratory birds, combined with broad habitat use, diverse foraging strategies, and long-distance movement, therefore enables the coupling of environmental reservoirs with transcontinental dispersal processes. In this way, tropical ecosystems function not only as hotspots for the emergence of antimicrobial resistance but also as critical nodes in a feedback loop linking environmental reservoirs, migratory birds, and anthropogenic AMR sources, particularly in regions where porous ecological boundaries, high human–wildlife contact, and limited biosecurity amplify opportunities for ARG transmission [99,100,101].
4. Evidence of AMR in Migratory Birds and Associated Pathogens
4.1. Extended-Spectrum β-Lactamase [ESBL] Producers
ESBL–producing Escherichia coli and other Enterobacteriaceae are among the most frequently reported antimicrobial-resistant organisms detected in migratory birds [14,20]. Since the first identification of ESBL-producing E. coli in pigeons in 1975, wild birds have increasingly been recognized as reservoirs of clinically relevant ESBL genes [14]. Exposure to agricultural fields treated with antimicrobials, wastewater-impacted wetlands, and urban refuse sites represents a significant pathway for migratory birds to acquire resistant strains [20,79]. As a widely used indicator organism for environmental AMR surveillance, E. coli has been extensively investigated in avian populations across multiple regions [80]. In Egypt, Ahmed et al. [2019] reported plasmid-mediated colistin resistance genes in migratory birds, with mcr-1 detected in 20% and mcr-2 in 3.6% of isolates [82]. In Pakistan, Mohsin et al. [2017] found that 17.3% of wild birds carried faecal ESBL-producing E. coli, of which 88.4% exhibited multidrug resistance [83]. Similarly, in Europe, Vergara et al. [2017] reported E. coli in 54.5% of faecal samples collected from gulls in Barcelona, with more than half of these isolates identified as ESBL producers [84].
4.2. Other Clinically Relevant Pathogens
4.2.1. Carbapenem-Resistant Enterobacteriaceae [CRE]
Although less frequently reported than ESBL producers, carbapenem-resistant Enterobacteriaceae [CRE] have been identified in migratory birds, raising concern regarding the dissemination of high-risk resistance determinants [102,103]. A notable example involved Salmonella enterica serovar Corvallis isolated from a black kite [Milvus migrans] in Germany carrying blaNDM-1, conferring resistance to cefotaxime and reduced susceptibility to carbapenems [104]. Phylogenetic analyses suggested dissemination from South or Southeast Asia to Europe via migratory routes, potentially involving stopover regions in North Africa and West Africa [104]. Supporting this hypothesis, Villa et al. [2015] demonstrated that plasmid pRH-1738 recovered from avian isolates closely resembled a human-associated plasmid from Afghanistan, indicating possible wildlife–anthropogenic exchange [103]. These findings underscore the ecological significance of migratory birds as potential long-distance carriers of carbapenemase-producing bacteria.
4.2.2. Colistin Resistance [mcr Genes]
Colistin represents a last-resort antimicrobial for the treatment of infections caused by carbapenem-resistant Gram-negative bacteria. The emergence of plasmid-mediated mcr genes therefore poses a substantial public-health concern. Multiple studies have reported mcr-positive Enterobacteriaceae in migratory birds, including detection of mcr-1 in European herring gulls [Larus argentatus] and in kelp gulls sampled in Ushuaia, Argentina [105,106]. In these cases, mcr-1 was frequently carried on IncI2 plasmids lacking ESBL genes, suggesting the circulation of wildlife-associated resistance lineages distinct from those commonly reported in livestock or clinical settings. The detection of mcr genes in migratory birds highlights their potential role in maintaining the environment and disseminating last-resort resistance determinants across geographic regions.
4.2.3. Zoonotic Pathogens
Salmonella enterica remains a zoonotic pathogen of major relevance to both human and livestock health and frequently exhibits antimicrobial-resistant phenotypes [107]. Migratory birds have been implicated in the dissemination of ESBL-producing and multidrug-resistant Salmonella during migration [85]. Prevalence studies have reported Salmonella isolation rates of 28.26% in migratory birds sampled in Egypt and 21.21% in Bangladesh [86]. Evidence of overlapping Salmonella serotypes between migratory birds and poultry farms in the Middle East further suggests potential cross-transmission at the wildlife–livestock interface [87].
Table 2.
Evidence of antimicrobial resistance [AMR] in migratory birds and associated pathogens.
| Pathogen | Host group | Resistance category | Antibiotics affected | Tropical region / flyway interface | References |
|---|---|---|---|---|---|
| Escherichia coli | Gulls, waterfowl, pigeons | ESBLs; MDR common; sporadic mcr | β-lactams, third-generation cephalosporins; colistin | Sub-Saharan Africa; South Asia [primary]; Europe [comparative] | [82,83,105,106,108] |
| Salmonella entericaserovar Corvallis | Raptors [black kite] | Carbapenemase [blaNDM-1] | Carbapenems | Africa–Eurasia flyway interface | [103,104] |
| Salmonellaspp. | Waterfowl, mixed wild birds | MDR; ESBL-associated | Multiple antibiotic classes | South Asia; Middle East | [85,86,87,158] |
| Campylobacterspp. | Shorebirds, waterfowl | Fluoroquinolone and macrolide resistance | Fluoroquinolones, macrolides | East Asia [tropical–subtropical interface] | [125,160] |
5. Risk Factors for Antimicrobial Resistance Genes [ARGs] Dissemination
The dissemination of ARGs by migratory birds is driven by an interconnected system of anthropogenic, agricultural, and environmental factors that create ecological hotspots for exposure, acquisition, and spread of ARB [111]. These interfaces are particularly prevalent in tropical and subtropical ecosystems, where inadequate sanitation, high antimicrobial use, and intense human–animal–environment interactions converge. Within such settings, migratory birds frequently encounter ARG-enriched habitats, acquire resistant microorganisms, and redistribute them across continents along major flyways [Figure 1].
5.1. Human Activity
5.1.1. Antibiotic Misuse in Clinical Settings
Extensive and often inappropriate antibiotic use in clinical settings significantly contributes to the persistence and amplification of resistant bacteria in the environment [112]. Between 30% and 90% of administered antibiotics are excreted in biologically active forms and enter hospital wastewater systems [113]. Consequently, hospital effluents contain concentrated mixtures of antimicrobial residues, ARGs, and pathogenic bacteria [114,115]. Studies from China, the Netherlands, and Nepal have documented tetracycline, quinolone, carbapenemase, and last-resort resistance genes, including blaNDM and mcr, in untreated or partially treated hospital wastewater [114]. When discharged into municipal sewage systems or natural water bodies, these effluents create heavily contaminated aquatic zones that migratory birds frequently use for feeding and resting [116,117].
5.1.2. Urban Wastewater and Sewage as Hotspots
Industrial, domestic, and hospital effluents commonly converge in wastewater treatment plants [WWTPs] or are discharged directly into natural waterways, often with incomplete removal of ARB and ARGs despite multiple treatment stages [118]. ARGs and resistant bacteria have been detected throughout wastewater processing chains [113], and resistance determinants may persist for up to 20 km downstream of treated effluent discharge points [119]. Migratory birds frequently exploit these anthropogenic water bodies, where ARG concentrations remain elevated [18]. Studies from Europe and Asia report that gulls and other migratory species using wastewater-impacted habitats carry ESBL genes, carbapenemases, aminoglycoside resistance genes, and quinolone resistance determinants [18,70,120].
5.1.3. Ecotourism and Bird–Human Contact Zones
The expansion of ecotourism activities, including birdwatching, hiking, and nature-based recreation, has intensified direct and indirect interactions between humans and wild birds [121,122]. Human presence in previously undisturbed habitats can introduce resistant bacteria into wildlife environments and promote cross-species bacterial exchange [123]. In China and Egypt, Escherichia coli, Campylobacter, and Salmonella strains harboring ARGs have been isolated from migratory birds nesting near heavily visited tourist sites [124,125]. These contact zones increase opportunities for bidirectional ARG spillover between humans and wildlife, highlighting ecotourism as a subtle but increasingly relevant driver of AMR dissemination.
5.2. Livestock and Agricultural Practices
5.2.1. Antibiotics in Animal Feed and Aquaculture
Antimicrobial use in livestock production and aquaculture remains a major contributor to environmental contamination with AMR [99,126,127]. Up to 90% of administered antibiotics are excreted unmetabolized in manure and urine [128], introducing AMR residues and ARB into soils and aquatic systems. Manure application as fertilizer adds ARGs to agricultural fields, where they may spread to soil microbiota or enter surface waters through runoff [129,130,131]. Slaughterhouse effluents also contain clinically significant ARGs and persistent pathogens [132,133]. Downstream river systems have also been shown to harbor multidrug resistance determinants that accumulate in fish and other aquatic organisms [131,134,135].
5.2.2. Interaction Between Migratory Birds and Farm Environments
Farmlands frequently serve as stopover sites for migratory birds because of the availability of food and water [101]. Shared access to irrigation channels, harvested crop fields, and livestock watering points creates opportunities for ARG exchange among birds, domestic animals, and agricultural landscapes [101]. These interactions reinforce One Health linkages and facilitate the circulation of resistance determinants between wildlife and livestock populations.
5.2.3. Spillover of Resistance Genes into the Food Chain
ARGs carried by migratory birds pose food safety risks by contaminating crop and livestock environments. Bird fecal deposition can introduce resistant bacteria into soil, irrigation water, and animal feed [101,135]. Livestock exposed to ARG-contaminated water or feed may subsequently transmit resistant bacteria to humans through meat, milk, feces, or direct contact [126,137]. Fresh produce may also be affected; resistant E. coli and multidrug-resistant Acinetobacter baumannii have been isolated from fruits and vegetables irrigated with contaminated water [138,139]. These pathways highlight the role of migratory birds in linking environmental AMR reservoirs with food production systems.
6. One Health Implications
The dissemination of antimicrobial resistance by migratory birds exemplifies the interconnectedness of human, animal, and environmental health central to the One Health paradigm [4,5,12,21,23]. By moving between natural wetlands, tropical agricultural landscapes, coastal estuaries, and densely populated urban areas, migratory birds facilitate ecological mixing of antimicrobial-resistant bacteria [ARB] and antimicrobial resistance genes [ARGs] across geographic and political boundaries [18,70,96,120]. This transboundary connectivity highlights that AMR is not sustained within isolated sectors but through continuous biological exchange among wildlife, humans, livestock, and the environment [5,21,140,141,142]. Understanding these linkages is essential for developing One Health strategies that address the emerging role of migratory birds in the dissemination of resistance [8,14,143,144].
6.1. Human Health
6.1.1. Spillover of Multidrug-Resistant Pathogens
Migratory birds function as biological bridges connecting distant ecosystems and human populations [12]. Long-distance migrants using the American, African–Eurasian, and East Asian–Australasian flyways have been shown to harbor human-associated resistant pathogens, including Escherichia coli, Salmonella enterica, and Campylobacter jejuni [145,146,147,148,149]. Urban expansion intensifies bird–human interactions, particularly at peri-urban wetlands, garbage dumps, and wastewater-adjacent communities, increasing opportunities for pathogen spillover and indirect human exposure [150,151].
6.1.2. Public Health Surveillance Gaps
Current AMR surveillance systems remain heavily biased toward clinical and livestock settings, with limited integration of wildlife reservoirs. As climate change and land-use modification alter migratory routes and stopover sites, bird–human contact zones are shifting in ways that existing surveillance frameworks fail to capture [142]. Addressing these gaps requires wildlife-inclusive One Health surveillance, including AMR screening at significant migratory stopovers, coordinated public health alerts along flyways, genomic monitoring of avian isolates, and the integration of GPS tracking and remote-sensing data to identify high-risk ecological nodes [141].
6.2. Animal Health
6.2.1. Exposure of Domestic and Wild Animals
Migratory birds interact with resident birds, livestock, and wildlife at shared water sources, crop fields, and coastal wetlands, creating opportunities for interspecies exchange of ARB and ARGs [91,96,97]. Overlapping use of aquatic habitats and foraging grounds facilitates microbial exchange through direct contact and environmental contamination, promoting the local establishment of resistance within resident animal populations [91,152]. Over time, these processes blur traditional boundaries between wildlife and domestic animal reservoirs of AMR [152].
6.2.2. AMR Threat to Biodiversity and Veterinary Medicine
The introduction of ARGs into naïve wildlife populations can disrupt ecological equilibria and facilitate the emergence of novel pathogen lineages [143,153,154]. Widespread antimicrobial use in livestock, aquaculture, and companion-animal medicine contributes to environmental contamination with resistant bacteria [18,155], which are subsequently encountered by migratory birds foraging at waste sites, ponds, and agricultural interfaces [79]. Migratory birds may also introduce resistant parasites and associated microbiota into new wildlife populations, while limited therapeutic options for wildlife infections amplify AMR-related risks to biodiversity and veterinary medicine [150,154].
6.3. Environmental Health
6.3.1. Ecosystems as Persistent ARG Reservoirs
Each year, billions of migratory birds traverse diverse ecological zones while carrying bacteria that may harbor clinically relevant ARGs [19,140,156]. Owing to their ecological flexibility, many species, particularly waterbirds, act as effective environmental vectors and amplifiers of AMR [19,25,157]. Fecal shedding during migration and stopovers results in continuous deposition of ARB and ARGs into wetlands, estuaries, and agricultural landscapes [92,159]. Escherichia coli, a key indicator organism for AMR surveillance, is frequently isolated in these contexts due to its high capacity for gene exchange and environmental persistence [157,160].
6.3.2. Long-Term Cycling and Evolution of Resistance
Migratory birds contribute to the persistence and global cycling of ARGs, reinforcing a dynamic “global resistome” shaped by repeated introduction, amplification, and redistribution across continents [14,20,93,161]. Resident avian species further amplify these processes by sustaining ARGs within local environments [14]. Within these ecological networks, avian-associated microbial communities may undergo local adaptation, co-selection, and recombination, generating novel resistance profiles that can ultimately spill back into human and livestock microbiomes.
7. Perspective Piece: Recommendations and Future Directions
AMR has evolved from a primarily clinical challenge into an ecological and transboundary crisis, particularly across tropical regions where environmental contamination, dense human–animal interfaces, and shifting migratory pathways converge. Migratory birds, due to their intercontinental mobility and regular use of human-modified tropical habitats, function as both biological vectors and sentinels of ARGs within the One Health framework. Addressing this complexity requires integrated surveillance, genomic-scale monitoring, and coordinated policies linking environmental microbiology, veterinary medicine, conservation biology, and public health
7.1. Strengthening AMR Surveillance in Migratory Bird Populations
Current AMR surveillance systems remain heavily focused on human and livestock sectors, leaving major wildlife reservoirs, particularly migratory birds, poorly represented [14,94]. To address this gap, avian surveillance should be integrated into existing platforms such as the WHO Global Antimicrobial Resistance and Use Surveillance System [GLASS] and the OIE World Animal Health Information System [WAHIS]. Routine sampling at key ecological interfaces, including wetlands, rice paddies, landfills, sewage lagoons, and agricultural landscapes, would enable spatiotemporal mapping of ARG dynamics along tropical flyways [18,95]. Standardized protocols and harmonized molecular workflows are essential to ensure data comparability and effective early-warning systems.
7.2. Genomic and Metagenomic Approaches to Track ARGs
Advances in next-generation sequencing enable high-resolution characterization of resistomes in migratory birds and their environments. Whole-genome sequencing and metagenomic profiling provide insights into ARG diversity, plasmid mobility, and microbial community structure across tropical ecosystems [8,11,23]. When integrated with ecological metadata, migratory corridors, habitat characteristics, and climatic parameters, these data can support predictive models to identify emerging ARG dissemination routes [89,98].
7.3. Integrated One Health Policies for AMR Control
Effective AMR mitigation requires coordinated governance that explicitly incorporates wildlife into national and regional action plans [5,21]. Cross-sectoral collaboration among ministries of health, agriculture, and environment, supported by WHO, FAO, OIE, and UNEP, should prioritize integrated wildlife–environment–human AMR databases, shared early-warning tools, transboundary data exchange, and joint outbreak investigations in tropical regions [4]. Strengthening collaboration among microbiologists, veterinarians, ornithologists, ecologists, and public health authorities will ensure that AMR surveillance is ecologically grounded and globally harmonized [4].
7.4. Mitigation Strategies and Sustainable Interventions
Reducing selective pressure in tropical ecosystems requires coordinated interventions across agriculture, wastewater management, and habitat stewardship. Limiting non-therapeutic antibiotic use in livestock and aquaculture remains a central priority [126,127]. Upgrading wastewater treatment technologies, including membrane bioreactors, advanced oxidation processes, and biofiltration, can substantially reduce the release of ARGs into the environment [112,115,162]. Ecological restoration of wetlands and the establishment of buffer zones around migratory bird habitats can further reduce contamination and interspecies contact [79,144].
7.5. Practical, Low-Cost Veterinary and Farm Biosecurity Measures
Simple, affordable biosecurity interventions can significantly reduce opportunities for wildlife–livestock exchange of ARB and ARGs [22,79]. These include covering livestock feed and water troughs, restricting wild bird access to poultry areas, limiting shared water points, improving manure management through composting, and mitigating runoff from agricultural sites [152]. Such measures are particularly critical in tropical regions characterized by high wildlife–livestock overlap and have been shown to reduce environmental contamination and cross-species transmission [163].
7.6. Integrating Climate-Sensitive Monitoring into AMR Surveillance
Climate change is reshaping migratory bird phenology, habitat use, and flyway connectivity across tropical ecosystems, altering opportunities for ARG exchange [78,140]. To address these emerging risks, AMR surveillance should incorporate climate-based habitat suitability models, anomaly-driven alerts for drying or newly formed wetlands, and targeted sampling at climate-altered peri-urban and agricultural water bodies [22,152]. Early-warning systems combining climate anomaly detection with targeted microbiological sampling could help identify evolving ARG hotspots [112,115].
7.7. Ethical Considerations for Habitat-Level Interventions
Interventions aimed at mitigating AMR risks in wildlife habitats must be carefully designed to avoid unintended ecological harm. Habitat modification, avian removal, or environmental disinfection may disrupt migratory connectivity, affect non-target species, or damage sensitive ecosystems [78,143,153]. In line with One Health principles, habitat-level interventions should undergo rigorous environmental and conservation impact assessments, involve interdisciplinary expertise, and demonstrate clear, evidence-based benefits that outweigh potential ecological risks [21,22,142].
8. Limitations
This review highlights several limitations that reflect broader deficiencies in the global evidence base on antimicrobial resistance [AMR] in migratory birds [12,13,14]. First, most studies rely on detection-based approaches that document antimicrobial-resistant bacteria or resistance genes [ARGs] without demonstrating direct transmission, host-to-host spread, or long-distance dispersal mediated by birds. The predominance of cross-sectional and opportunistic study designs means that most findings remain correlational rather than causal [19,69,79]. In addition, DNA-based methods often fail to distinguish viable bacteria from residual genetic material, limiting their epidemiological and veterinary relevance and constraining inference about active transmission [79].
Second, avian AMR research is geographically uneven. Surveillance efforts are concentrated in Europe, East Asia, and parts of North America, where laboratory capacity and wildlife monitoring systems are well established [70,79,88,91]. In contrast, tropical low- and middle-income regions, including Southeast Asia, sub-Saharan Africa, and Latin America, remain markedly underrepresented due to limited microbiological infrastructure, weaker veterinary surveillance, and fewer coordinated monitoring programs [14,18,101]. This imbalance restricts the generalizability of existing findings and likely underestimates ARG diversity and circulation along tropical flyways. Finally, substantial taxonomic and flyway biases persist. Research focuses disproportionately on gulls and waterfowl, while passerines, raptors, and long-distance migrants, many of which rely on tropical stopover sites, are comparatively understudied [25]. Methodological heterogeneity across studies, including differences in sampling design, host selection, analytical workflows, genomic resolution, and metadata reporting, further complicates cross-study comparisons and synthesis [25,164].
9. Conclusion
AMR is a global ecological crisis that extends beyond clinical boundaries, encompassing environmental and geographic dimensions. Within the One Health framework, migratory birds act as both sentinels and potential vectors of ARGs, linking human settlements, agricultural systems, and natural ecosystems along continental flyways [19,90,140]. Their repeated use of human-modified habitats exposes them to resistant bacteria derived from wastewater effluents, agricultural runoff, and landfill waste, reinforcing connectivity between clinical and environmental resistomes. Evidence across regions shows that avian-associated bacteria frequently harbor extended-spectrum β-lactamases, carbapenemases, and mcr genes—resistance determinants of significant clinical concern [82,158]. These findings underscore that AMR is not solely a biomedical problem, but an ecological and evolutionary process shaped by interactions among species, ecosystems, and human activity. Effective responses require integrated molecular surveillance, environmental context, and coordinated policy action. Genomic and metagenomic tools should underpin global AMR monitoring, supported by strengthened antimicrobial stewardship and cross-sectoral collaboration. Recognizing migratory birds not merely as passive carriers but as ecological indicators of emerging resistance pathways is essential. Protecting avian and environmental health is therefore integral to sustaining microbial balance and safeguarding long-term human health [8,14,94].
Author Contributions
OBI: Conceptualization, Validation, Visualization, Supervision, Writing – original draft, Writing – review & editing. ZME: Validation, Visualization, Writing – original draft, Writing – review & editing. NOI: Writing – review & editing. AOA: Writing – review & editing. OTR: Writing – review & editing. OAA: Writing – review & editing. AAA: Writing – review & editing. AAA: Writing – review & editing. MHI: Writing – review & editing, Visualization. YJS: Writing – review & editing. SOO: Writing – review & editing.
Acknowledgments
We want to acknowledge all authors who have contributed with high-quality reviews.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author[s] declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or that its manufacturer may claim, is not guaranteed or endorsed by the publisher.
References
- Salam MA, Al-Amin MY, Salam MT, Pawar JS, Akhter N, Rabaan AA, et al. Antimicrobial Resistance: A Growing Serious Threat for Global Public Health. p. 1946. [CrossRef]
- Barron M. The antimicrobial resistance pandemic: Breaking the silence. https://asm.org/articles/2024/october/antimicrobial-resistance-pandemic-breaking-silence.
- Naghavi M, Vollset SE, Ikuta KS, Swetschinski LR, Gray AP, Wool EE, et al. Global burden of bacterial antimicrobial resistance 1990–2021: a systematic analysis with forecasts to 2050. The Lancet. [2024] .
- Horefti E. The Importance of the One Health Concept in Combating Zoonoses. [CrossRef]
- Horvat O, Kovačević Z. Human and Veterinary Medicine Collaboration: Synergistic Approach to Address Antimicrobial Resistance Through the Lens of Planetary Health. Antibiotics. [2025] 14:38. [CrossRef]
- Bengtsson-Palme J, Kristiansson E, Larsson DGJ. Environmental factors influencing the development and spread of antibiotic resistance. FEMS microbiology reviews. [2018] 42:68–80. [CrossRef]
- James R, Hardefeldt LY, Ierano C, Charani E, Dowson L, Elkins S, et al. Antimicrobial stewardship from a One Health perspective. Nature reviews. Microbiology. [2025] . [CrossRef]
- Bustamante M, Mei S, Daras IM, Doorn GS, Falcao Salles J, Vos MGJ. An eco-evolutionary perspective on antimicrobial resistance in the context of One Health. iScience. [2024] 28:111534. [CrossRef]
- Tokuda M, Shintani M. Microbial evolution through horizontal gene transfer by mobile genetic elements. Microbial biotechnology. [2024] 17:14408. [CrossRef]
- Goh YX, Anupoju SMB, Nguyen A. Evidence of horizontal gene transfer and environmental selection impacting antibiotic resistance evolution in soil-dwelling Listeria. Nat Commun. [2024] 15:10034. [CrossRef]
- Johansson MHK, Aarestrup FM, Petersen TN. Importance of mobile genetic elements for dissemination of antimicrobial resistance in metagenomic sewage samples across the world. PloS one. [2023] 18:0293169. [CrossRef]
- Jarma D, Sánchez MI, Green AJ, Peralta-Sánchez JM, Hortas F, Sánchez-Melsió A, et al. Faecal microbiota and antibiotic resistance genes in migratory waterbirds with contrasting habitat use. Science of The Total Environment. [2021] 783:146872. [CrossRef]
- Franklin AB, Ramey AM, Bentler KT, Barrett NL, McCurdy LM, Ahlstrom CA, et al. Gulls as Sources of Environmental Contamination by Colistin-resistant Bacteria. Scientific Reports. [2020] 10:1–10. [CrossRef]
- Mbuthia CW, Hoza AS. Wild birds as potential reservoirs of antimicrobial-resistant Escherichia coli: A systematic review. Frontiers in Microbiology. [2025] 16:1615826. [CrossRef]
- Ramey AM. Antimicrobial resistance: Wildlife as indicators of anthropogenic environmental contamination across space and through time. Current Biology. [2021] 31:R1385–R1387. [CrossRef]
- Łopińska A, Nowak-Zaleska A, Kosicki JZ, Jerzak L, Węgrzyn A, Węgrzyn G. Reassessing the role of wild birds in the spread of antibiotic resistance: the white stork as a model species in studying populations from Central European river valley. Microbiology Spectrum. [2025] . [CrossRef]
- Lisovski S, Hoye BJ, Conklin JR, Battley PF, Fuller RA, Gosbell KB, et al. Predicting resilience of migratory birds to environmental change. Proceedings of the National Academy of Sciences of the United States of America. [2024] 121:2311146121. [CrossRef]
- Elsohaby I, Samy A, Elmoslemany A, Alorabi M, Alkafafy M, Aldoweriej A, et al. Migratory Wild Birds as a Potential Disseminator of Antimicrobial-Resistant Bacteria around Al-Asfar Lake, Eastern Saudi Arabia. Antibiotics. [2021] 10:260. [CrossRef]
- Lin Y, Dong X, Sun R, Wu J, Tian L, Rao D, et al. Migratory birds-one major source of environmental antibiotic resistance around Qinghai Lake, China. Science of the Total Environment. [2020] 739:139758. [CrossRef]
- Yuan Y, Liang B, Jiang BW, Zhu LW, Wang TC, Li YG, et al. Migratory wild birds carrying multidrug-resistant Escherichia coli as potential transmitters of antimicrobial resistance in China. PloS one. [2021] 16:0261444. [CrossRef]
- Danasekaran R. One Health: A Holistic Approach to Tackling Global Health Issues. Indian journal of community medicine : official publication of Indian Association of Preventive & Social Medicine. [2024] 49:260–263. [CrossRef]
- Jiménez-Ruiz S, Santos N, Barasona JA, Fine AE, Jori F. Pathogen transmission at the domestic-wildlife interface: a growing challenge that requires integrated solutions. Frontiers in veterinary science. [2024] 11:1415335. [CrossRef]
- Guardia T, Varriale L, Minichino A, Balestrieri R, Mastronardi D, Russo TP, et al. Wild birds and the ecology of antimicrobial resistance: an approach to monitoring. Journal of Wildlife Management. [2024] 88:. [CrossRef]
- Skarżyńska M, C Z, M. B, A. B, Ł. K, W. P, et al. Antimicrobial Resistance Glides in the Sky-Free-Living Birds as a Reservoir of Resistant Escherichia coli With Zoonotic Potential. Frontiers in microbiology. [2021] 12:656223. [CrossRef]
- Zeballos-Gross D, Rojas-Sereno Z, Salgado-Caxito M, Poeta P, Torres C, Benavides JA. The Role of Gulls as Reservoirs of Antibiotic Resistance in Aquatic Environments: A Scoping Review. Frontiers in Microbiology. [2021] 12:. [CrossRef]
- Reygaert WC. An overview of the antimicrobial resistance mechanisms of bacteria. AIMS Microbiology. [2018] 4:482–501. [CrossRef]
- Christaki E, Marcou M, Tofarides A. Antimicrobial resistance in bacteria: mechanisms, evolution, and persistence. Journal of molecular evolution. [2020] Jan;88[1]:26-40:.
- Varela MF, Stephen J, Lekshmi M, Ojha M, Wenzel N, Sanford LM, et al. Bacterial Resistance to Antimicrobial Agents. Antibiotics. [2021] 10:593. [CrossRef]
- Liu Y, Luo J, Chen X, Liu W, Chen T. Cell Membrane Coating Technology: A Promising Strategy for Biomedical Applications. Nano-Micro Lett. [2019] 11:100. [CrossRef]
- Mora-Ochomogo M, Lohans CT. β-Lactam Antibiotic Targets and Resistance mechanisms: from Covalent Inhibitors to Substrates. RSC Medicinal Chemistry. [2021] 12:1623–1639. [CrossRef]
- Sawa T, Kooguchi K, Moriyama K. Molecular Diversity of extended-spectrum β-lactamases and carbapenemases, and Antimicrobial Resistance. Journal of Intensive Care. [2020] 8:. [CrossRef]
- Bush K, Jacoby GA. Updated Functional Classification of -Lactamases. Antimicrobial Agents and Chemotherapy. [2010] 54:969–976. [CrossRef]
- Philippon A, Arlet G, Labia R, Iorga BI. Class C β-Lactamases: Molecular Characteristics. Clinical Microbiology Reviews. [2022] 35:. [CrossRef]
- Lin J, Zhou D, Steitz TA, Polikanov YS, Gagnon MG. Ribosome-Targeting Antibiotics: Modes of Action, Mechanisms of Resistance, and Implications for Drug Design. Annual Review of Biochemistry. [2018] 87:451–478.
- Partridge SR, Kwong SM, Firth N, Jensen SO. Mobile Genetic Elements Associated with Antimicrobial Resistance. Clinical Microbiology Reviews. [2018] 31:. [CrossRef]
- Galgano M, Pellegrini F, Catalano E, Capozzi L, Sambro LD, Sposato A, et al. Acquired Bacterial Resistance to Antibiotics and Resistance Genes: From Past to Future. Antibiotics. [2025] 14:222. [CrossRef]
- Vrancianu CO, Popa LI, Bleotu C, Chifiriuc MC. Targeting Plasmids to Limit Acquisition and Transmission of Antimicrobial Resistance. Frontiers in Microbiology. [2020] 11:. [CrossRef]
- Castañeda-Barba S, Top EM, Stalder T. Plasmids, a molecular cornerstone of antimicrobial resistance in the One Health era. Nature Reviews Microbiology. [2023] 22:. [CrossRef]
- Bhat BA, Mir RA, Qadri H, Dhiman R, Almilaibary A, Alkhanani M, et al. Integrons in the development of antimicrobial resistance: critical review and perspectives. Frontiers in Microbiology. [2023] 14:. [CrossRef]
- Hegazy EE, ElNaghy WS, Shalaby MM, Shoeib SM, Abdeen NSM, Fouda MH, et al. Study of Class 1, 2, and 3 Integrons, Antibiotic Resistance Patterns, and Biofilm Formation in Clinical Staphylococcus aureus Isolates from Hospital-Acquired Infections. Pathogens. [2025] 14:705. [CrossRef]
- Babakhani S, Oloomi M. Transposons: the agents of antibiotic resistance in bacteria. Journal of Basic Microbiology. [2018] 58:905–917. [CrossRef]
- Shintani M, Vestergaard G, Milaković M, Kublik S, Smalla K, Schloter M, et al. Integrons, transposons and IS elements promote diversification of multidrug resistance plasmids and adaptation of their hosts to antibiotic pollutants from pharmaceutical companies. Environmental Microbiology. [2023] 25:3035–3051. [CrossRef]
- Sinha S, Upadhyay LS. Understanding antimicrobial resistance [AMR] mechanisms and advancements in AMR diagnostics. Diagnostic Microbiology and Infectious Disease. [2025] 16:.
- Nazir A, Nazir A, Zuhair V, Aman S, Sadiq SUR, Hasan AH, et al. The Global Challenge of Antimicrobial Resistance: Mechanisms. Case Studies, and Mitigation Approaches. Health Science Reports. [2025] 8:71077.
- Paudel R, Shrestha E, Chapagain B, Tiwari BR. Carbapenemase producing Gram negative bacteria: Review of resistance and detection methods. Diagnostic Microbiology and Infectious Disease. [2024] 110:116370. [CrossRef]
- Halat DH, Moubareck CA. The Current Burden of Carbapenemases: Review of Significant Properties and Dissemination among Gram-Negative Bacteria. Antibiotics. [2020] 9:186. [CrossRef]
- Nishino K, Yamasaki S, Nakashima R, Zwama M, Hayashi-Nishino M. Function and Inhibitory Mechanisms of Multidrug Efflux Pumps. Frontiers in Microbiology. [2021] 12:. [CrossRef]
- Huang YS, Zhou H. Breakthrough Advances in Beta-Lactamase Inhibitors: New Synthesized Compounds and Mechanisms of Action Against Drug-Resistant Bacteria. Pharmaceuticals. [2025] 18:206. [CrossRef]
- Gaurav A, Bakht P, Saini M, Pandey S, Pathania R. Role of bacterial efflux pumps in antibiotic resistance and strategies to discover novel efflux pump inhibitors. Microbiology. [2023] 169:001333. [CrossRef]
- Sharma A, Gupta VK, Pathania R. Efflux pump inhibitors for bacterial pathogens. Indian J Med Res. [2019] 149:129–145. [CrossRef]
- Brown CJ, Sen D, Yano H, Bauer ML, Rogers L, Van, et al. Diverse Broad-Host-Range Plasmids from Freshwater Carry Few Accessory Genes. Applied and Environmental Microbiology. [2013] 79:7684–7695. [CrossRef]
- Ramsay JP, Kwong SM, Murphy RJT, Yui Eto K, Price KJ, Nguyen QT, et al. An updated view of plasmid conjugation and mobilization in Staphylococcus. Mobile Genetic Elements. [2016] 6:. [CrossRef]
- Tokuda M, Suzuki H, Yanagiya K, Yuki M, Inoue K, Ohkuma M, et al. Determination of Plasmid pSN1216-29 Host Range and the Similarity in Oligonucleotide Composition Between Plasmid and Host Chromosomes. Frontiers in Microbiology. [2020] 11:. [CrossRef]
- Foley SL, Kaldhone PR, Ricke SC, Han J. Incompatibility Group I1 [IncI1] Plasmids: Their Genetics, Biology, and Public Health Relevance. Microbiology and Molecular Biology Reviews. [2021] 85:. [CrossRef]
- Virolle C, Goldlust K, Djermoun S, Bigot S, Lesterlin C. Plasmid Transfer by Conjugation in Gram-Negative Bacteria: From the Cellular to the Community Level. Genes. [2020] 11:1239. [CrossRef]
- Atmakuri A, Yadav B, Tiwari B, Drogui P, Tyagi RD, Wong JW. Nature’s architects: a comprehensive review of extracellular polymeric substances and their diverse applications. Waste Disposal & Sustainable Energy. [2024] Dec;6[4]:529-51:.
- Liu W, Huang Y, Zhang H, Liu Z, Huan Q, Xiao X, et al. Factors and Mechanisms Influencing Conjugation In Vivo in the Gastrointestinal Tract Environment: A Review. International Journal of Molecular Sciences. [2023] 24:5919. [CrossRef]
- Boyd SE, Holmes A, Peck R, Livermore DM, Hope WW. OXA-48-Like β-Lactamases: Global Epidemiology, Treatment Options, and Development Pipeline. Antimicrobial Agents and Chemotherapy. [2022] 66:. [CrossRef]
- Xia C, Yan R, Liu C, Zhai J, Zheng J, Chen W, et al. Epidemiological and genomic characteristics of global blaNDM-carrying Escherichia coli. Annals of Clinical Microbiology and Antimicrobials. [2024] 23:. [CrossRef]
- Baquero F, Martínez JL. Interventions on metabolism: Making antibiotic-susceptible bacteria. mBio. [2017] 8:. [CrossRef]
- Kraemer MUG, Reiner Jr RC, Brady OJ, Messina JP, Gilbert M, Pigott DM, et al. Past and future spread of the arbovirus vectors Aedes aegypti and Aedes albopictus. Nature microbiology. [2019] 4:854–863. [CrossRef]
- Zhuang M, Yan W, Xiong Y, Wu Z, Cao Y, Sanganyado E, et al. Horizontal plasmid transfer promotes antibiotic resistance in selected bacteria in Chinese frog farms. Environment International. [2024] 190:108905. [CrossRef]
- Allen R, Yokota T. Endosomal Escape and Nuclear Localization: Critical Barriers for Therapeutic Nucleic Acids. Molecules [Basel. [2024] 29:5997. [CrossRef]
- Wellington EMH, Boxall ABA, Cross P, Feil EJ, Gaze WH, Hawkey PM, et al. The role of the natural environment in the emergence of antibiotic resistance in Gram-negative bacteria. The Lancet Infectious Diseases. [2013] 13:155–165. [CrossRef]
- Zhang S, Huang J, Zhao Z, Cao Y, Li B. Hospital Wastewater as a Reservoir for Antibiotic Resistance Genes: A Meta-Analysis. Frontiers in Public Health. [2020] 8:574968. [CrossRef]
- Huijbers PMC, Blaak H, De Jong MCM, Graat EAM, Vandenbroucke-Grauls CMJE, De Roda Husman AM. Role of the Environment in the Transmission of Antimicrobial Resistance to Humans: A Review. Environmental science & technology. [2015] 49:11993–12004. [CrossRef]
- Rizzo L, Manaia C, Merlin C, Schwartz T, Dagot C, Ploy MC, et al. Urban wastewater treatment plants as hotspots for antibiotic resistant bacteria and genes spread into the environment: A review. Science of the Total Environment. [2013] 447:345–360. [CrossRef]
- Karkman A, Pärnänen K, Larsson DGJ. Fecal pollution can explain antibiotic resistance gene abundances in anthropogenically impacted environments. Nature Communications 2019 10:1. [2019] 10:1–8. [CrossRef]
- Zhao H, Sun R, Yu P, Alvarez PJJ. High levels of antibiotic resistance genes and opportunistic pathogenic bacteria indicators in urban wild bird feces. Environmental Pollution. [2020] 266:115200. [CrossRef]
- Atterby C, Börjesson S, Ny S, Järhult JD, Byfors S, Bonnedahl J. ESBL-producing Escherichia coli in Swedish gulls-A case of environmental pollution from humans? PloS One. [2017] 12:. [CrossRef]
- Liu Y, Li R, Xiao X, Wang Z. Molecules that Inhibit Bacterial Resistance Enzymes. Molecules. [2018] 24:43. [CrossRef]
- Muteeb G, Rehman T, Shahwan M, Aatif M. Origin of Antibiotics and Antibiotic Resistance, and Their Impacts on Drug Development: A Narrative Review. Pharmaceuticals. [2023] 16:1615. https://doi.org/https://pmc.ncbi.nlm.nih.gov/articles/PMC10675245/.
- Meradji S, Basher NS, Sassi A, Ibrahim NA, Idres T, Touati A. The Role of Water as a Reservoir for Antibiotic-Resistant Bacteria. Antibiotics. [2025] 14:763. [CrossRef]
- Adekanmbi AO, Akinpelu MO, Olaposi A V, Oyelade AA. Extended spectrum beta-lactamase encoding gene-fingerprints in multidrug resistant Escherichia coli isolated from wastewater and sludge of a hospital treatment plant in Nigeria. International Journal of Environmental Studies. [2020] 78:140–150. [CrossRef]
- Ibrahim E, Mkwanda C, Masoambeta E, Scudeller L, Kostyanev T, Twabi HH, et al. Prevalence of Extended-Spectrum β-Lactamase-Producing Escherichia coli, Klebsiella pneumoniae and Enterobacter cloacae in Wastewater Effluent in Blantyre, Malawi. Antibiotics. [2025] 14:562. [CrossRef]
- Nwankwoala HO, Okujagu DC. A REVIEW OF WETLANDS AND COASTAL RESOURCES OF THE NIGER DELTA: POTENTIALS, CHALLENGES AND PROSPECTS. Environment & Ecosystem Science. [2021] 5:37–46. [CrossRef]
- Monsalves N, Leiva AM, Gómez G, Vidal G. Antibiotic-Resistant Gene Behavior in Constructed Wetlands Treating Sewage: A Critical Review. Sustainability. [2022] 14:8524. [CrossRef]
- Mei X. Effect of Climate Change and Loss of Habitat on Migratory Birds. Transactions on Environment, Energy and Earth Sciences. [2024] 3:288–293. [CrossRef]
- Blanco G, López-Hernández I, Morinha F, López-Cerero L. Intensive farming as a source of bacterial resistance to antimicrobial agents in sedentary and migratory vultures: Implications for local and transboundary spread. The Science of the Total Environment. [2020] 739:140356. [CrossRef]
- Anjum MF, Schmitt H, Börjesson S, Berendonk TU, Donner E, Stehling EG, et al. The potential of using E. coli as an indicator for the surveillance of antimicrobial resistance [AMR] in the environment. Current Opinion in Microbiology. [2021] 64:152–158. [CrossRef]
- Anjum MF, Duggett NA, AbuOun M, Randall L, Nunez-Garcia J, Ellis RJ, et al. Colistin resistance in Salmonella and Escherichia coli isolates from a pig farm in Great Britain. Journal of Antimicrobial Chemotherapy. [2016] 71:2306–2313. [CrossRef]
- Ahmed ZS, Elshafiee EA, Khalefa HS, Kadry M, Hamza DA. Evidence of colistin resistance genes [mcr-1 and mcr-2] in wild birds and its public health implication in Egypt. Antimicrobial resistance and infection control. [2019] 8:. [CrossRef]
- Mohsin M, Raza S, Schaufler K, Roschanski N, Sarwar F, Semmler T, et al. High prevalence of CTX-M-15-Type ESBL-Producing E. coli from migratory avian species in Pakistan. Frontiers in Microbiology. [2017] 8:285845. [CrossRef]
- Vergara A, Pitart C, Montalvo T, Roca I, Sabaté S, Hurtado JC, et al. Prevalence of Extended-Spectrum-β-Lactamase- and/or Carbapenemase-Producing Escherichia coli Isolated from Yellow-Legged Gulls from Barcelona, Spain. Antimicrobial Agents and Chemotherapy. [2017] 61:e02071-16. [CrossRef]
- Card RM, Chisnall T, Begum R, Sarker MS, Hossain MS, Sagor MS, et al. Multidrug-resistant non-typhoidal Salmonella of public health significance recovered from migratory birds in Bangladesh. Frontiers in Microbiology. [2023] 14:1162657. [CrossRef]
- Shalaby AG, Erfan M, Nasef S. Surveillance for Salmonella in migratory, feral and zoo birds. Suez Canal Veterinary Medical Journal. SCVMJ. [2014] 19:85–101. [CrossRef]
- Al-baqir A, Hussein A, Ghanem I, Megahed M. Characterization of Paratyphoid Salmonellae Isolated from Broiler Chickens at Sharkia Governorate. Egypt. Zagazig Veterinary Journal. [2019] 47:183–192. [CrossRef]
- Dreyer S, Globig A, Bachmann L, Schütz AK, Schaufler K, Homeier-Bachmann T. Longitudinal Study on Extended-Spectrum Beta-Lactamase-E. coli in Sentinel Mallard Ducks in an Important Baltic Stop-Over Site for Migratory Ducks in Germany. Microorganisms. [2022] 10:. [CrossRef]
- Segawa T, Takahashi A, Kokubun N, Ishii S. Spread of antibiotic resistance genes to Antarctica by migratory birds. Science of The Total Environment. [2024] 923:171345. [CrossRef]
- Cheng Z, Chen Y, Li M, Lv C, Zhou N, Chen W, et al. An Unusual “Gift” from Humans: Third-Generation Cephalosporin-Resistant Enterobacterales in migratory birds along the East Asian-Australasian Flyway. Environment International. [2025] 197:109320. [CrossRef]
- Ahlstrom CA, Bonnedahl J, Woksepp H, Hernandez J, Olsen B, Ramey AM. Acquisition and dissemination of cephalosporin-resistant E. coli in migratory birds sampled at an Alaska landfill as inferred through genomic analysis. Scientific Reports. [2018] 8:7361. [CrossRef]
- Wang L, Jia R, Ma R, Li J, Wu S, Fan Y, et al. High throughput screening for human disease associated-pathogens and antimicrobial resistance genes in migratory birds at ten habitat sites in China. BMC Microbiology. [2025] 25:355. [CrossRef]
- Rapi MC, Filipe J, Filippone Pavesi L, Raimondi S, Addis MF, Franciosini MP, et al. Resisting the Final Line: Phenotypic Detection of Resistance to Last-Resort Antimicrobials in Gram-Negative Bacteria Isolated from Wild Birds in Northern Italy. Animals. [2025] 15:2289. [CrossRef]
- Benavides JA, Salgado-Caxito M, Torres C, Godreuil S. Public Health Implications of Antimicrobial Resistance in Wildlife at the One Health Interface. Medical Sciences Forum. [2024] 25:1. [CrossRef]
- Ahlstrom CA, Toor ML, Woksepp H, Chandler JC, Reed JA, Reeves AB, et al. Evidence for continental-scale dispersal of antimicrobial resistant bacteria by landfill-foraging gulls. Science of The Total Environment. [2021] 764:144551. [CrossRef]
- Atterby C, Ramey AM, Hall GG, Järhult J, Börjesson S, Bonnedahl J. Increased prevalence of antibiotic-resistant E. coli in gulls sampled in Southcentral Alaska is associated with urban environments. Infection Ecology & Epidemiology. [2016] 6:32334. [CrossRef]
- Stedt J, Bonnedahl J, Hernandez J, McMahon BJ, Hasan B, Olsen B, et al. Antibiotic resistance patterns in Escherichia coli from gulls in nine European countries. Infection Ecology & Epidemiology. [2014] 4:21565. [CrossRef]
- Cao J, Hu Y, Liu F, Wang Y, Bi Y, Lv N, et al. Metagenomic analysis reveals the microbiome and resistome in migratory birds. Microbiome. [2020] 8:26. [CrossRef]
- Katale BZ, Misinzo G, Mshana SE, Chiyangi H, Campino S, Clark TG, et al. Genetic diversity and risk factors for the transmission of antimicrobial resistance across human, animals and environmental compartments in East Africa: A review. Antimicrobial Resistance and Infection Control. [2020] 9:1–20. [CrossRef]
- Laborda P, Sanz-García F, Ochoa-Sánchez LE, Gil-Gil T, Hernando-Amado S, Martínez JL. Wildlife and Antibiotic Resistance. Frontiers in Cellular and Infection Microbiology. [2022] 12:873989. [CrossRef]
- Blount JD, Horns JJ, Kittelberger KD, Neate-Clegg MHC, Şekercioğlu ÇH. Avian Use of Agricultural Areas as Migration Stopover Sites: A Review of Crop Management Practices and Ecological Correlates. Frontiers in Ecology and Evolution. [2021] 9:. [CrossRef]
- Carattoli A. Plasmids and the spread of resistance. International Journal of Medical Microbiology. [2013] 303:298–304. [CrossRef]
- Villa L, Guerra B, Schmoger S, Fischer J, Helmuth R, Zong Z, et al. IncA/C Plasmid Carrying blaNDM-1 , blaCMY-16 , and fosA3 in a Salmonella enterica Serovar Corvallis Strain Isolated from a Migratory Wild Bird in Germany. Antimicrobial Agents and Chemotherapy. [2015] 59:6597–6600. [CrossRef]
- Fischer J, Schmoger S, Jahn S, Helmuth R, Guerra B. NDM-1 carbapenemase-producing Salmonella enterica subsp. Enterica serovar Corvallis isolated from a wild bird in Germany. Journal of Antimicrobial Chemotherapy. [2013] 68:2954–2956. [CrossRef]
- Ruzauskas M, Vaskeviciute L. Detection of the mcr-1 gene in Escherichia coli prevalent in the migratory bird species Larus argentatus. The Journal of antimicrobial chemotherapy. [2016] 71:2333–2334. [CrossRef]
- Liakopoulos A, Mevius DJ, Olsen B, Bonnedahl J. The colistin resistance mcr-1 gene is going wild: Table 1. Journal of Antimicrobial Chemotherapy. [2016] 71:2335–2336. [CrossRef]
- Chatterjee P, Chauhan N, Jain U. Confronting antibiotic-resistant pathogens: Distinctive drug delivery potentials of progressive nanoparticles. Microbial Pathogenesis. [2024] 187:106499. [CrossRef]
- Fashae K, Engelmann I, Monecke S, Braun SD, Ehricht R. Molecular characterisation of extended-spectrum ß-lactamase producing Escherichia coli in wild birds and cattle, Ibadan, Nigeria. BMC Veterinary Research. [2021] 17:33. [CrossRef]
- Luangtongkum T, Jeon B, Han J, Plummer P, Logue CM, Zhang Q. Antibiotic Resistance in Campylobacter: Emergence, Transmission and Persistence. Future Microbiology. [2009] 4:189–200. [CrossRef]
- Smith JL, Fratamico PM. Fluoroquinolone Resistance in Campylobacter. Journal of Food Protection. [2010] 73:1141–1152. [CrossRef]
- Koutsoumanis K, Allende A, Álvarez-Ordóñez A, Bolton D, Bover-Cid S, Chemaly M, et al. Role played by the environment in the emergence and spread of antimicrobial resistance [AMR] through the food chain. EFSA Journal. [2021] 19:06651. [CrossRef]
- Lan L, Wang Y, Chen Y, Wang T, Zhang J, Tan B. A Review on the Prevalence and Treatment of Antibiotic Resistance Genes in Hospital Wastewater. Toxics. [2025] 13:263. [CrossRef]
- Gao P, Munir M, Xagoraraki I. Correlation of tetracycline and sulfonamide antibiotics with corresponding resistance genes and resistant bacteria in a conventional municipal wastewater treatment plant. Science of The Total Environment. [2012] 421–422:173–183. [CrossRef]
- Zhu L, Shuai XY, Lin ZJ, Sun YJ, Zhou ZC, Meng LX, et al. Landscape of genes in hospital wastewater breaking through the defense line of last-resort antibiotics. Water Research. [2022] 209:. [CrossRef]
- Huang J, Wang Z, Chen Z, Liang H, Li X, Li B. Occurrence and Removal of Antibiotic Resistance in Nationwide Hospital Wastewater Deciphered by Metagenomics Approach — China, 2018–2022. China CDC Weekly. [2023] 5:1023. [CrossRef]
- Majumder A, Gupta AK, Ghosal PS, Varma M. A review on hospital wastewater treatment: A special emphasis on occurrence and removal of pharmaceutically active compounds, resistant microorganisms, and SARS-CoV-2. Journal of Environmental Chemical Engineering. [2020] 9:104812. [CrossRef]
- Venter H, Henningsen ML, Begg SL. Antimicrobial resistance in healthcare, agriculture and the environment: the biochemistry behind the headlines. Essays in Biochemistry. [2017] 61:1. [CrossRef]
- Singer AC, Shaw H, Rhodes V, Hart A. Review of antimicrobial resistance in the environment and its relevance to environmental regulators. Frontiers in Microbiology. [2016] 7:219380. [CrossRef]
- Sabri NA, Schmitt H, Zaan B, Gerritsen HW, Zuidema T, Rijnaarts HHM, et al. Prevalence of antibiotics and antibiotic resistance genes in a wastewater effluent-receiving river in the Netherlands. Journal of Environmental Chemical Engineering. [2020] 8:102245. [CrossRef]
- Oteo J, Menciá A, Bautista V, Pastor N, Lara N, González-González F, et al. Colonization with Enterobacteriaceae-Producing ESBLs, AmpCs, and OXA-48 in Wild Avian Species, Spain 2015-2016. Microbial Drug Resistance. [2018] 24:932–938. [CrossRef]
- Ikuta LA, Blumstein DT. Do fences protect birds from human disturbance? Biological Conservation. [2003] 112:447–452. [CrossRef]
- Steven R, Pickering C, Guy Castley J. A review of the impacts of nature based recreation on birds. Journal of Environmental Management. [2011] 92:2287–2294. [CrossRef]
- Muehlenbein MP. Human-Wildlife Contact and Emerging Infectious Diseases. Human-Environment Interactions. [2012] 1:79. [CrossRef]
- Tawakol MM, Nabil NM, Samir A, Hassan HM, Reda RM, Abdelaziz O, et al. Role of migratory birds as a risk factor for the transmission of multidrug resistant Salmonella enterica and Escherichia coli to broiler poultry farms and its surrounding environment. BMC Research Notes. [2024] 17:314. [CrossRef]
- Wu S, Jia R, Wang Y, Li J, Li Y, Wang L, et al. Prevalence, Diversity, and Virulence of Campylobacter Carried by Migratory Birds at Four Major Habitats in China. Pathogens. [2024] 13:230. [CrossRef]
- Allcock S, Young EH, Holmes M, Gurdasani D, Dougan G, Sandhu MS, et al. Antimicrobial resistance in human populations: challenges and opportunities. Global Health, Epidemiology and Genomics. [2017] 2:4. [CrossRef]
- Rahman AU, Valentino V, Sequino G, Ercolini D, Filippis F. Comparative analysis of antibiotic-administered vs. antibiotic-free farming in meat production: Implications for health, environment, and antibiotic resistance. Food Microbiology. [2025] 133:104877. [CrossRef]
- Berendsen BJA, Wegh RS, Memelink J, Zuidema T, Stolker LAM. The analysis of animal faeces as a tool to monitor antibiotic usage. Talanta. [2015] 132:258–268. [CrossRef]
- Karami N, Martner A, Enne VI, Swerkersson S, Adlerberth I, Wold AE. Transfer of an ampicillin resistance gene between two Escherichia coli strains in the bowel microbiota of an infant treated with antibiotics. Journal of Antimicrobial Chemotherapy. [2007] 60:1142–1145. [CrossRef]
- Wichmann F, Udikovic-Kolic N, Andrew S, Handelsman J. Diverse antibiotic resistance genes in dairy cow manure. MBio. [2014] 5:1017–1030. [CrossRef]
- Smith SD, Colgan P, Yang F, Rieke EL, Soupir ML, Moorman TB, et al. Investigating the dispersal of antibiotic resistance associated genes from manure application to soil and drainage waters in simulated agricultural farmland systems. PLOS ONE. [2019] 14:0222470. [CrossRef]
- Cong X, Krolla P, Khan UZ, Savin M, Schwartz T. Antibiotic resistances from slaughterhouse effluents and enhanced antimicrobial blue light technology for wastewater decontamionation. Environmental Science and Pollution Research International. [2023] 30:109315. [CrossRef]
- Savin M, Bierbaum G, Hammerl JA, Heinemann C, Parcina M, Sib E, et al. ESKAPE Bacteria and Extended-Spectrum-β-Lactamase-Producing Escherichia coli Isolated from Wastewater and Process Water from German Poultry Slaughterhouses. Applied and Environmental Microbiology. [2020] 86:. [CrossRef]
- Huang J, Mi J, Yan Q, Wen X, Zhou S, Wang Y, et al. Animal manures application increases the abundances of antibiotic resistance genes in soil-lettuce system associated with shared bacterial distributions. Science of The Total Environment. [2021] 787:147667. [CrossRef]
- Vezeau N, Kahn L. Spread and mitigation of antimicrobial resistance at the wildlife-urban and wildlife-livestock interfaces. Journal of the American Veterinary Medical Association. [2024] 262:741–747. [CrossRef]
- Osińska A, Korzeniewska E, Harnisz M, Felis E, Bajkacz S, Jachimowicz P, et al. Small-scale wastewater treatment plants as a source of the dissemination of antibiotic resistance genes in the aquatic environment. Journal of Hazardous Materials. [2020] 381:121221. [CrossRef]
- Founou LL, Founou RC, Essack SY. Antibiotic resistance in the food chain: A developing country-perspective. Frontiers in Microbiology. [2016] 7:232834. [CrossRef]
- O’Flaherty E, Solimini AG, Pantanella F, De Giusti M, Cummins E. Human exposure to antibiotic resistant-Escherichia coli through irrigated lettuce. Environment International. [2019] 122:270–280. [CrossRef]
- Carvalheira A, Silva J, Teixeira P. Lettuce and fruits as a source of multidrug resistant Acinetobacter spp. Food Microbiology. [2017] 64:119–125. [CrossRef]
- Ramey AM, Hernandez J, Tyrlöv V, Uher-Koch BD, Schmutz JA, Atterby C, et al. Antibiotic-Resistant Escherichia coli in Migratory Birds Inhabiting Remote Alaska. EcoHealth. [2018] 15:72–81. [CrossRef]
- Jindal M, Stone H, Lim S, MacIntyre CR. A Geospatial Perspective Toward the Role of Wild Bird Migrations and Global Poultry Trade in the Spread of Highly Pathogenic Avian Influenza H5N1. GeoHealth. [2025] 9:1296–2024. [CrossRef]
- Mourkas E, Valdebenito JO, Marsh H, Hitchings MD, Cooper KK, Parker CT, et al. Proximity to humans is associated with antimicrobial-resistant enteric pathogens in wild bird microbiomes. Current Biology. [2024] 34:3955-3965 4. [CrossRef]
- Perrin A, Glaizot O, Christe P. Migratory birds spread their haemosporidian parasites along the world’s major migratory flyways. Oikos. [2025] 2025:e11012. [CrossRef]
- Farkas K, Knight ME, Woodhall N, Williams RC, Seerung W, Silvester R, et al. Monitoring of antimicrobial resistance genes and influenza viruses in avian-populated water bodies. Sustainable Microbiology. [2025] 2:013. [CrossRef]
- Blagodatski A, Trutneva K, Glazova O, Mityaeva O, Shevkova L, Kegeles E, et al. Avian Influenza in Wild Birds and Poultry: Dissemination Pathways, Monitoring Methods, and Virus Ecology. Pathogens. [2021] 10:630. [CrossRef]
- Gómez C, Hobson KA, Bayly NJ, Rosenberg K V, Morales-Rozo A, Cardozo P, et al. Migratory connectivity then and now: A northward shift in breeding origins of a long-distance migratory bird wintering in the tropics. Proceedings of the Royal Society B: Biological Sciences. [2021] 288:2021–2188, 20210188. [CrossRef]
- Kiat Y, Izhaki I, Sapir N. The effects of long-distance migration on the evolution of moult strategies in Western-Palearctic passerines. Biological reviews of the Cambridge Philosophical Society. [2019] 94:700–720. [CrossRef]
- Ramey AM, Hill NJ, DeLiberto TJ, Gibbs SEJ, Camille Hopkins M, Lang AS, et al. Highly pathogenic avian influenza is an emerging disease threat to wild birds in North America. The Journal of Wildlife Management. [2022] 86:22171. [CrossRef]
- Shi X, Soderholm J, Chapman JW, Meade J, Farnsworth A, Dokter AM, et al. Distinctive and highly variable bird migration system revealed in Eastern Australia. Current Biology. [2024] 34:5359-5365 3. [CrossRef]
- Qiu Y, Lv C, Chen J, Sun Y, Tang T, Zhang Y, et al. The global distribution and diversity of wild-bird-associated pathogens: An integrated data analysis and modeling study. Med. [2025] 6:100553. [CrossRef]
- Zurfluh K, Albini S, Mattmann P, Kindle P, Nüesch-Inderbinen M, Stephan R, et al. Antimicrobial resistant and extended-spectrum β-lactamase producing Escherichia coli in common wild bird species in Switzerland. MicrobiologyOpen. [2019] 8:845. [CrossRef]
- Lee NH, You S, Taghizadeh A, Taghizadeh M, Kim HS. Cell Membrane-Cloaked Nanotherapeutics for Targeted Drug Delivery. Int J Mol Sci. [2022] 23:2223. [CrossRef]
- Llaberia-Robledillo M, Balbuena JA, Sarabeev V, Llopis-Belenguer C. Changes in native and introduced host–parasite networks. Biological Invasions. [2022] 24:543–555. [CrossRef]
- Poulin R, Angeli Dutra D. Animal migrations and parasitism: Reciprocal effects within a unified framework. Biological Reviews. [2021] 96:1331–1348. [CrossRef]
- Van Boeckel TP, Brower C, Gilbert M, Grenfell BT, Levin SA, Robinson TP, et al. Global trends in antimicrobial use in food animals. Proceedings of the National Academy of Sciences of the United States of America. [2015] 112:5649–5654. [CrossRef]
- Islam MS, Nayeem MMH, Sobur MA, Ievy S, Islam MA, Rahman S, et al. Virulence Determinants and Multidrug Resistance of Escherichia coli Isolated from Migratory Birds. Antibiotics. [2021] 10:190. [CrossRef]
- Esposito E, Scarpellini R, Celli G, Marliani G, Zaghini A, Mondo E, et al. Wild birds as potential bioindicators of environmental antimicrobial resistance: A preliminary investigation. Research in Veterinary Science. [2024] 180:105424. [CrossRef]
- Islam S, Md. P, A. T, M. R, K. AS, Md. I, et al. Migratory birds travelling to Bangladesh are potential carriers of multi-drug resistant Enterococcus spp., Salmonella spp., and Vibrio spp. Saudi Journal of Biological Sciences. [2021] 28:5963–5970. [CrossRef]
- Zeballos C, A. M, Gaj T. Next-Generation CRISPR Technologies and Applications. Trends Biotechnol. [2021] 39:692–705. [CrossRef]
- Ahmed NA, Gulhan T. Campylobacter in Wild Birds: Is It an Animal and Public Health Concern? Frontiers in Microbiology. [2022] 12:812591. [CrossRef]
- Nabil NM, Erfan AM, Tawakol MM, Haggag NM, Naguib MM, Samy A. Wild Birds in Live Birds Markets: Potential Reservoirs of Enzootic Avian Influenza Viruses and Antimicrobial Resistant Enterobacteriaceae in Northern Egypt. Pathogens. [2020] 9:196. [CrossRef]
- Sambaza SS, Naicker N. Contribution of wastewater to antimicrobial resistance: A review article. Journal of Global Antimicrobial Resistance. [2023] 34:23–29. [CrossRef]
- Esposito MM, Turku S, Lehrfield L, Shoman A. The Impact of Human Activities on Zoonotic Infection Transmissions. Animals : an open access journal from MDPI. [2023] 13:1646. [CrossRef]
- Pereira GQ, Gomes LA, Santos IS, Alfieri AF, Weese JS, Costa MC. Fecal microbiota transplantation in puppies with canine parvovirus infection. Journal of Veterinary Internal Medicine. [2018] 32:707–711. [CrossRef]
Figure 1.
Schematic representation of significant risk factors driving antimicrobial resistance gene [ARG] dissemination.
Figure 1.
Schematic representation of significant risk factors driving antimicrobial resistance gene [ARG] dissemination.
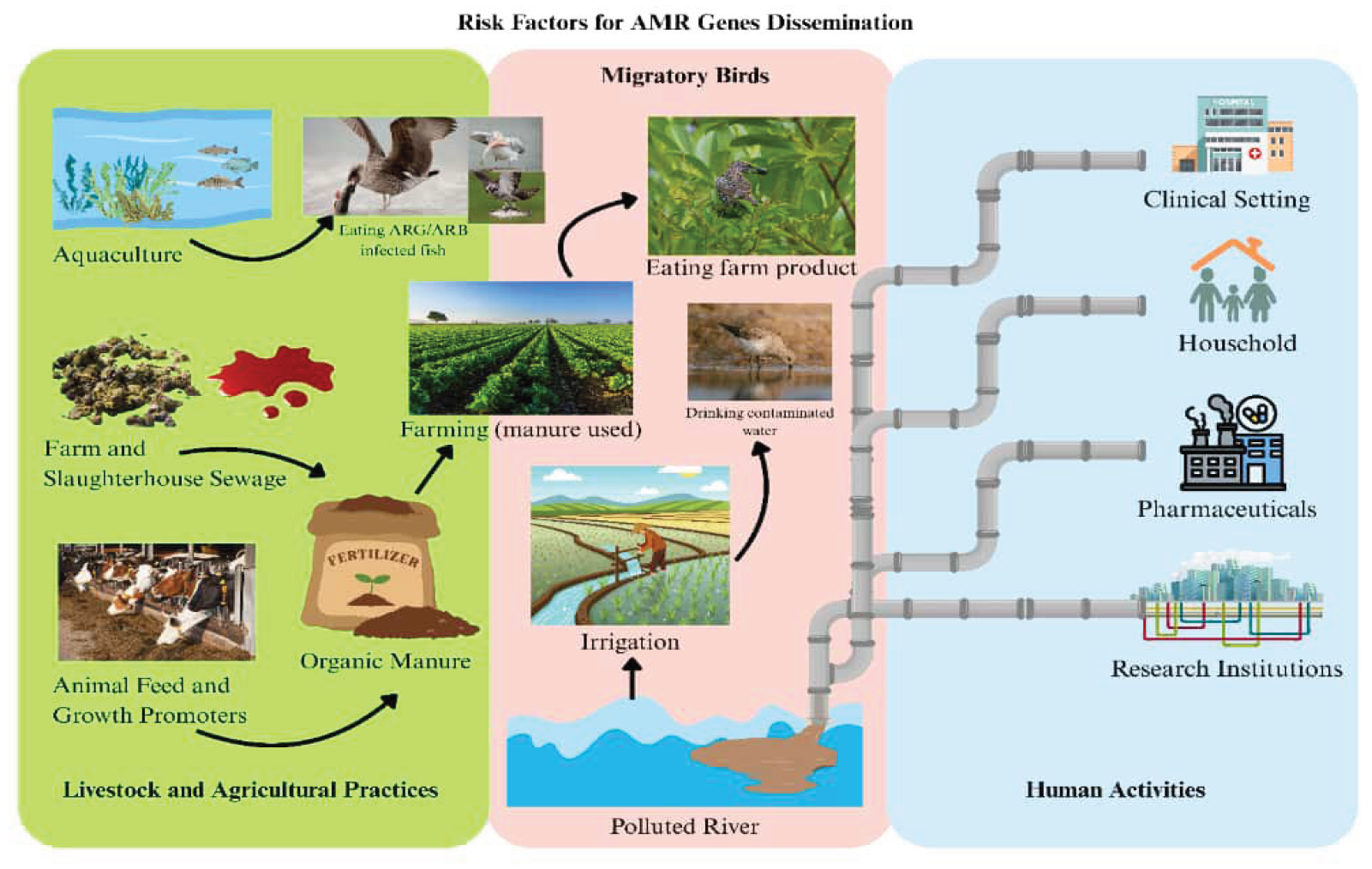
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



