
Submitted:
20 December 2025
Posted:
23 December 2025
You are already at the latest version
Abstract
Outer membrane vesicles (OMVs) from gut Escherichia coli are emerging postbiotic candidates, yet how probiotic versus commensal strains differentially program human dendritic cells (DCs) remains poorly defined. Here we compared OMVs from probiotic E. coli Nissle 1917 (EcN) and commensal strains ECOR12 and ECOR63 in human monocyte-derived DCs, integrating cytokines and microRNAs (miRNAs). All OMVs drove DC maturation, increasing CD83 and secretion of IL-6, IL-10 and TNF-α, but with strain-specific patterns: ECOR12 preferentially increased IL-10, whereas EcN and ECOR63 induced stronger IL-6/TNF-α outputs. EcN and ECOR12 OMVs upregulated miR-155-5p and let-7i-3p, while ECOR12 selectively boosted miR-146b-5p and EcN preferentially induced miR-29a-5p, linking vesicle origin to regulatory circuits controlling TLR signalling and epithelial barrier–related pathways. Principal component analysis showed that the first two components captured 96.2% of total variance, clearly segregating strains. MANOVA confirmed a robust multivariate strain effect (Pillai = 1.99, p < 2.2×10⁻¹⁶), and univariate models showed that strain explained 78–91% of cytokine variance and 98–99% of miRNA variance (R² up to 0.993). These data support OMVs as nanoscale conveyors of strain-encoded information that imprint distinct cytokine/miRNA fingerprints on DCs and provide mechanistic biomarkers for microbiota–immune crosstalk.
Keywords:
outer membrane vesicles
; Escherichia coli Nissle 1917
; dendritic cells
; microRNAs
; mucosal immunology
1. Introduction
Inflammatory disorders of the gut represent a growing biomedical and pub-lic-health challenge, not only because of their increasing prevalence, but also because they emerge from a complex network of host–microbe interactions that remains only partially resolved. Inflammatory bowel disease (IBD) alone has been described as a rising global burden with substantial impacts on quality of life, healthcare costs, and long-term complications [1,2]. Beyond IBD, chronic intestinal inflammation and barrier dysfunction are increasingly linked to al-tered microbial ecology and abnormal immune tone at mucosal surfaces [3,4]. A central difficulty is that many mechanistic mod-els still treat the microbiota as a diffuse “black box,” while the intestinal immune sys-tem responds to discrete molecular cues and particulate structures that can be mapped, measured, and potentially leveraged therapeutically [5,6]. This gap limits our ability to explain why some microbial com-munities support homeostasis while others precipitate inflammatory cascades, and it constrains the development of targeted microbiota-derived interventions [7,8].
The intestinal microbiota is now widely recognized as a functional organ that contributes to host defense, epithelial integrity, and metabolic balance [4,9]. These contributions are not merely indirect; commensal organisms can competitively exclude pathogens, shape mucosal immunity, and reinforce epithelial tight junctions, while also producing metabolites and vitamins that influence cellular differentiation and energy homeostasis [3,8]. Such functions are especially relevant in the gut, where epithelial cells coordinate barrier architecture and immune communication through pattern recognition receptors (PRRs), cytokine signaling, and antimicrobial programs [6,10]. In parallel, dendritic cells (DCs) operate as gate-keepers that sample luminal information (directly or indirectly), integrate danger and tolerance cues, and instruct downstream T-cell polarization—ranging from effector responses to regulatory programs [11,12]. Thus, intestinal homeostasis depends on tightly coupled signaling between epithelial cells and DCs, and disruption of this crosstalk is a plausible mechanistic route to chronic inflammation [6,10] (Figure 1).
A critical insight in recent years is that microbiota–host communication does not require whole bacterial cells. Bacterial membrane vesicles act as mediators of microbe–microbe and microbe–host community interactions [13]. Bacte-ria release membrane-bound extracellular vesicles including outer membrane vesicles (OMVs) in Gram-negative species that can traverse mucus, interact with epithelial and immune cells, and deliver complex cargos such as lipopolysaccharide (LPS), outer membrane proteins, peptidoglycan fragments, nucleic acids, and small RNAs [14,15]. OMVs are increasingly viewed as organized “packages” of microbial information rather than passive debris: they can promote colonization, modulate barrier properties, shape inflammatory pathways, and influ-ence adaptive immunity [16,17]. Importantly, vesicle biology is not restricted to Gram-negative organisms—vesicle-mediated signaling also occurs in Gram-positive bacteria and fun-gi—reinforcing the concept that extracellular vesicles (EVs) represent a conserved strategy for cross-kingdom communication [18]. In the gut environ-ment, where the immune system must discriminate between danger, commensal tol-erance, and beneficial immunoregulation, OMVs provide a mechanistically tractable interface to study how microbial cues are translated into host transcriptional and functional outcomes [19,20].
OMVs can also exert divergent effects depending on their origin and context. Ves-icles derived from pathogenic bacteria may amplify systemic inflammation and con-tribute to severe inflammatory phenotypes [21,22]. Conversely, OMVs from commensal or probiotic strains can engage PRRs in ways that promote barrier reinforcement and balanced immune activation rather than uncon-trolled inflammation [24,70]. Mechanistically, OMVs can enter intestinal epithelial cells via endocytic routes such as clathrin-dependent in-ternalization, with downstream consequences ranging from DNA damage responses to immune pathway activation, depending on cargo composition and the responding cell type [25]. OMV–host interactions therefore sit at the intersection of in-nate recognition (e.g., TLRs and NOD-like receptors), inflammasome pathways, cyto-kine networks, and epithelial stress programs [5,26]. These pathways are also entangled with autophagy-related pro-cesses that regulate antigen handling, inflammation, and cellular homeosta-sis—particularly under conditions of “signal 0” sensing driven by pathogen-associated molecular patterns (PAMPs) and damage-associated molecular patterns (DAMPs) [28] (Figure 2).
Among Gram-negative organisms, Escherichia coli provides a particularly in-formative framework because strains can occupy positions along a spectrum from harmless commensals to opportunistic pathogens, and some are used clinically as pro-biotics. The probiotic strain E. coli Nissle 1917 (EcN) is notable for its long history of therapeutic use and for mechanistic traits that support gut persistence and host bene-fit, including colonization factors, microcin production, and immunomodulatory po-tential [29,30]. Specific components such as fimbriae can contribute to biofilm formation and intestinal colonization [31], and capsule-associated features can shape epithelial PRR responses and down-stream MAPK-dependent cytokine induction [32]. At the same time, EcN is not simply “non-inflammatory”: its immunological impact is nuanced and con-text-dependent, emphasizing the need to characterize how EcN-derived structures (including OMVs) tune immune pathways rather than assuming uniform an-ti-inflammatory effects [29,33]. Phylogenetic frame-works for E. coli strains, including classic reference sets and rapid grouping methods, are essential to interpret how commensal background influences vesicle content and host responses [34,35].
Proteomic and functional studies support the idea that EcN OMVs carry immu-nologically active material and can elicit measurable mucosal responses. Proteomic profiling has revealed complex OMV compositions, reflecting selective cargo loading rather than random shedding [36,37]. In vivo and ex vivo work indicates that OMVs from commensal and probiotic E. coli strains can acti-vate immune and defense programs in the intestinal mucosa, including pathways as-sociated with innate sensing and barrier regulation [38]. In experimental colitis models, EcN OMVs have been linked to anti-inflammatory effects, highlighting their potential as biologically stable, cell-free effectors with therapeutic implications [33]. Yet these benefits coexist with a broader literature showing that OMVs can also trigger strong inflammatory outputs depending on their cargo and the responding immune compartment [17,21]. This duality strengthens the rationale for resolving OMV-driven responses at the level of specific host cell programs, rather than relying on coarse phenotypic endpoints.
Dendritic cells are a central hub for this resolution. Positioned at the interface of innate sensing and adaptive instruction, DCs integrate PRR engagement, inflam-masome activity, and cytokine signals to guide T-cell differentiation, B-cell help, and tolerance induction [11,12]. DC activation states are often discussed through surface markers and cytokines (e.g., IL-6, IL-12, TNF-α, IL-10), but a parallel regulatory layer is mediated by microRNAs (miRNAs) that can amplify, buffer, or reprogram inflammatory signaling [39,40]. Canonical examples include miR-155, which is induced during inflamma-tory activation and modulates cytokine production [41], and also shapes IL-1 signaling in activated monocyte-derived DCs [42]. Likewise, IL-10-dependent miR-146b can suppress TLR4 signaling, serving as a negative feed-back mechanism to prevent excessive inflammation [43]. Broader frameworks describe miRNAs as fine-tuners of TLR pathways and innate immune setpoints, reinforcing their relevance for mucosal immunology [44,45,46,47,48].
Crucially, miRNA regulation in the gut is not only an “immune-cell story.” miR-NAs also shape epithelial barrier function and intestinal permeability—key pheno-types in both functional bowel disorders and inflammatory disease. miR-29a has been associated with increased intestinal membrane permeability in clinical and experi-mental contexts, with downstream impacts on tight junction regulation [49,50]. Inhibition of miR-29a can restore barrier-associated proteins (e.g., ZO-1 and claudins) in diarrhea-predominant models, underscoring a mechanistic link between miRNA expression and epithelial integrity [51]. At the in-tersection of miRNA biology and PRR signaling, let-7 family members can regulate TLR4 expression and influence epithelial immune responses to infection-related stimu-li [52]. Collectively, these findings motivate a model in which OMVs may reshape mucosal outcomes not only by triggering cytokines through PRRs [5], but also by reprogramming miRNA networks that control the magnitude, duration, and tissue-level consequences of immune activation [53,39,40].
Evidence is[39,40,53 rapidly accumulating that microbiota-derived vesicles can drive pre-cisely this kind of miRNA-linked immune modulation. Transcriptomic profiling has shown that dendritic cells respond to gut microbiota-secreted vesicles with distinct miRNA signatures, suggesting that vesicle exposure can “write” immunological in-formation into post-transcriptional regulatory circuits [54]. Complementary work indicates that vesicles from EcN and gut-resident E. coli strains can differentially modulate human DCs and shape subsequent T-cell responses, high-lighting strain specificity at the level of adaptive instruction [55]. More broadly, reviews now position microbiota EVs as emerging players in gut homeostasis and disease, with a strong emphasis on immunoregulatory signaling, bar-rier effects, and cargo-driven mechanisms [20,56,57]. These advances, however, also expose a key limitation: many studies describe individual cytokines or selected miRNAs in isolation, while OMV responses are inherently multivariate and may involve coordinated patterns across inflammato-ry and regulatory axes [58,59]. In this setting, multivari-ate statistical strategies become more than a reporting choice—they are required to resolve immune “fingerprints” that would be missed by univariate comparisons.
Accordingly, dimensionality reduction and clustering approaches provide a prin-cipled route to interpret OMV-driven response profiles. Principal component analysis (PCA) can summarize correlated cytokine/miRNA variation into interpretable axes, while biplots allow loadings to be visualized as vectors that clarify which variables structure separation among conditions [60,61,62]. Hierarchical clustering and heatmap visualization can reveal coherent modules of response across conditions, and modern annotation-rich heatmap frame-works facilitate biologically meaningful pattern discovery [63,64,65]. When outcomes are multivariate by nature—as in combined cytokine and miRNA panels—MANOVA-based inference offers a formal test of condition effects, with classical work highlighting robustness properties of dif-ferent multivariate test statistics and modern work providing robust alternatives when assumptions are challenged [66,67]. Finally, careful effect-size reporting is essential in biological systems where statistical significance may not translate directly into biological relevance, and interpretive cautions around R² further motivate transparent reporting practices [68,69]. Embedding these tools within OMV immunobiology can transform complex da-tasets into mechanistic hypotheses, candidate biomarkers, and rational targets for pro-biotic-derived interventions.
Within this conceptual and methodological framework, EcN OMVs are especially compelling because they concentrate multiple probiotic attributes into a nanoscale, cell-free format: colonization-associated factors, antimicrobial microcins, and immu-nologically active surface components can be delivered to epithelial and immune compartments without requiring live bacterial replication [29,36]. At the same time, commensal E. coli strains offer a critical comparator to disentangle which vesicle-driven effects are truly “probiotic-specific” versus part of a broader commensal signaling landscape [34,35]. Prior work indicates that OMVs from probiotic and commensal E. coli activate innate pathways such as NOD1-mediated responses in epithelial cells [23], and that EcN-derived vesicles can protect against epithelial barrier dysfunction induced by enteropathogenic E. coli [70,71]. Meanwhile, evi-dence that probiotic-derived OMVs can influence macrophage polarization and anti-microbial activity further supports the idea that vesicles are active immunological agents with cell-type-specific outputs [72,73]. The unresolved question is not whether OMVs “do something,” but how vesicle origin and cargo translate into distinct epithelial–DC–macrophage programs that collectively deter-mine whether the net effect is inflammatory escalation, balanced defense, or toler-ance-supporting regulation [16,17,74] (Figure 3).
In this study, we address this mechanistic gap by examining how OMVs derived from probiotic and commensal E. coli strains shape mucosal-relevant immune signal-ing, with an explicit focus on dendritic-cell reprogramming and miRNA-linked regula-tion. We leverage the current understanding of innate recognition pathways [5], mucosal crosstalk between epithelial cells and DCs [10,11], and the emerging role of microbiota EVs in gut homeostasis [56,57] to frame OMVs as modular immune “signals” capable of driving coordinated cytokine–miRNA response states. The revised sche-matic figures included in this work are intentionally more specific and explanatory for the present experimental context and are introduced in the same order as the concep-tual argument: they synthesize (i) the protective, structural, and metabolic functions of the microbiota relevant to barrier integrity and immune tone (Figure 1); (ii) the PAMP/DAMP–PRR axis that initiates autophagy-linked immune programs (Figure 2); (iii) the strain-attribute-to-OMV-cargo framework used here to connect probi-otic/commensal traits to epithelial and dendritic-cell outcomes (Figure 3); and (iv) the cell-type-resolved OMV interaction pathways that link vesicle cargo to macrophage activation, DC polarization, and epithelial responses (Figure 4).
Therefore, the objectives of this study were to (1) characterize and compare den-dritic-cell immune activation patterns elicited by OMVs from probiotic EcN and com-mensal E. coli strains, integrating cytokine outputs with miRNA-associated regulatory signatures [36,54,55]; (2) contextualize these responses within epithelial and macrophage interaction path-ways relevant to barrier reinforcement, inflammasome-linked inflammation, and tol-erance induction [6,25,72]; and (3) apply multivariate analytical strategies (PCA/biplots, clustering/heatmaps, and mul-tivariate inference) to identify response fingerprints that clarify strain-specific im-munomodulation and support mechanistic interpretation [60,61,64,66].
2. Materials and Methods
2.1. Experimental Design
This study used an in vitro, controlled, comparative design to determine whether outer membrane vesicles (OMVs) derived from probiotic Escherichia coli Nissle 1917 (EcN) and a commensal E. coli strain (ECOR12) induce distinct maturation and im-munoregulatory programs in human monocyte-derived dendritic cells (Mo-DCs). The design follows the current framework that OMVs are biologically active, nanoscale carriers of microbial signals capable of shaping innate sensing and downstream im-mune outputs through cell-type–dependent mechanisms [14,15,16,17].
The experimental groups and replication scheme are summarized in Table 1.
2.2. Evaluated Variables
Outcome selection was guided by (i) the established role of dendritic cells as inte-grators of microbial sensing and T-cell instruction [10,11,12], and (ii) evidence that microbiota- and E. coli–derived vesicles can reprogram DC cytokines and microRNA (miRNA) networks [54,55]. Cytokines (IL-6, IL-10, TNF-α) were quantified to capture canonical maturation-associated out-puts, while flow cytometry markers (CD14, CD83, CD209) were used to phenotype the iDC→mDC transition [5,27]. miRNA targets were quantified by RT-qPCR to reflect key regulatory layers of innate signaling [41,42,43,44,45,47,48]. The complete panel of evaluated variables and their measurement modality is listed in Table 2.
2.3. Biological Material and Dendritic-Cell Generation
Peripheral blood from healthy donors was used as the source of peripheral blood mononuclear cells (PBMCs). PBMCs were isolated by density-gradient separation using Hytopaque® 1077, followed by washing and cell counting. CD14⁺ monocytes were iso-lated by positive selection (MidiMACS system; LS columns; CD14 Microbeads), then differentiated to immature Mo-DCs (iDCs) in Mo-DC differentiation medium supple-mented with recombinant human IL-4 and GM-CSF under standard incubator condi-tions (37 °C, 5% CO₂) for approximately 6–7 days. This differentiation strategy aligns with established approaches for generating monocyte-derived DCs for microbi-al/vesicle immunomodulation studies [21,27], and supports a biologically relevant platform for studying OMV-driven DC programming [24,55].
2.4. OMV Production, Isolation, STERILITY Control, and Pre-Use Characterization
OMVs were produced from E. coli strains grown on LB agar and incubated at 37 °C. After growth, bacteria were removed by centrifugation (10,000 × g, 30 min, 4 °C) and the supernatant was sterile-filtered (0.2 µm) under cold-chain conditions. The fil-trate was concentrated using Centricon® Plus-70 devices (10 kDa cutoff; swing-ing-bucket rotor at 3,500 × g, 20 min, 4 °C) to reduce ~1 L to ~8 mL. Residual bacteria were removed by 0.2 µm syringe filtration, and OMVs were pelleted by ultracentrifu-gation (1 h, 45,000 rpm, 4 °C). Pellets were resuspended in sterile PBS, sterility was verified by plating on LB agar (37 °C, 48 h), OMV protein was quantified by the Lowry method, and aliquots were stored at −20 °C until use. This workflow is consistent with the principle that OMVs represent a structured output of Gram-negative bacteria, with bioactivity that depends on clean isolation and standardized input dosing [15,36,37,75].
For structural and compositional quality control prior to stimulation assays, OMVs were processed for Cryo-TEM in a dedicated microscopy facility using cryogen-ic conditions (approximately −170 to −175 °C) and electron acceleration voltage (200 kV), following the study’s microscopy preparation pipeline [22]. In addition, OMV protein profiles were assessed by SDS-PAGE (10% acrylamide; ~10 µg protein loaded; run at 20 mA; Coomassie staining) based on prior laboratory protocols used for OMV protein profiling [25].
2.5. Dendritic-Cell Stimulation with OMVs
After Mo-DC differentiation, iDCs were washed with PBS and resuspended in Mo-DC medium. Cells were then stimulated with OMVs derived from intestinal E. coli strains at 10 µg/mL for 24 h (37 °C; 5% CO₂). This exposure paradigm was chosen to mimic physiologically relevant vesicle–immune cell contact at mucosal interfaces, where OMVs can deliver immunostimulatory and immunoregulatory cargos [5,16,20,57,74].
2.6. Data Acquisition Equipment and Measurement Settings
To ensure transparency in data acquisition, the main instruments and key settings are summarized in Table 3.
2.7. Flow Cytometry Phenotyping of Dendritic-Cell Maturation
DC maturation status was evaluated by flow cytometry using fluorescent mono-clonal antibodies targeting CD14 (monocyte marker), CD83 (maturation marker), and CD209/DC-SIGN (immature DC-associated marker). Data were acquired in a dedi-cated cytometry facility, and iDCs served as the phenotypic baseline control. Marker selection is consistent with established immunophenotyping approaches used to sepa-rate immature and mature Mo-DC states [27], and supports mechanis-tic interpretation of OMV-driven maturation alongside cytokine/miRNA readouts [11,55].
2.8. Cytokine Quantification by ELISA
Culture supernatants were collected after 24 h stimulation and assessed for IL-6, IL-10, and TNF-α secretion using BD Biosciences ELISA kits, following manufacturer protocols and previously established laboratory procedures. Cytokine selection reflects the central role of these mediators in DC maturation and inflammatory/regulatory po-larization downstream of pattern recognition receptor activation [5,6].
2.9. miRNA Extraction and RT-qPCR
Total RNA enriched for small RNAs was extracted from dendritic cells after OMV exposure using the miRNeasy® Mini Kit (Qiagen). RNA quantity and purity were as-sessed by NanoDrop® ND-1000 spectrophotometry (A260/A280). Samples were stored at −80 °C until analysis. Reverse transcription was performed using miRCURY® LNA® RT Kit (Qiagen), and qPCR was conducted using miRCURY LNA miRNA PCR assays. UNISP was used as an internal control, and selected housekeeping/reference targets (including U6 and additional reference miRNAs) supported normalization; relative expression was computed using the 2^-ΔΔCt method. This approach is widely used for miRNA quantification in immune-cell activation contexts, where miRNAs are established regulators of TLR/IL-1 signaling and inflammatory feedback control [41,42,43,44,45,47,48].
2.10. Data Analysis
2.11. Null and Working Hypotheses (Pre-Specified)
Hypotheses were defined at both multivariate and variable-specific levels, con-sistent with the concept that OMV exposure can shift coordinated immune programs rather than a single endpoint [15,57,74].
Null hypothesis (H0, multivariate): The joint response vector (cytokines + miRNA expression profile ± phenotypic markers) does not differ among groups (control iDC vs. EcN OMVs vs. ECOR12 OMVs). Operationally, H0 is evaluated by MANOVA and rejected when p < 0.05 using Pillai’s trace, given its documented robustness [66], with robust alternatives considered under assumption violations [67].
Working/alternative hypothesis (H1): At least one OMV condition produces a sta-tistically detectable shift in the multivariate immune response profile relative to con-trol, and/or EcN and ECOR12 OMVs elicit distinguishable response fingerprints. This expectation is biologically grounded in the strain-specific immunomodulatory capacity of E. coli OMVs and microbiota-derived vesicles [24,25,54,55] and in miRNA-mediated tuning of innate signaling circuits [42,43,45].
3. Results
3.1. Characterization of OMVs: Cryo-TEM and SDS-PAGE
Before assessing OMV biological effects, OMV preparations were characterized. Isolated OMVs were quantified using the Lowry protein assay [75], and the resulting preparations were subsequently processed for Cryo-TEM visualization.
The protein-based quantification (and the volumes used to standardize sample preparation to a final volume of 20 µL) are summarized in Table 5.
Representative Cryo-TEM micrographs of OMVs from each strain are presented in Figure 5.
As shown in Figure 5, vesicles obtained from the probiotic strain EcN and from the two commensal strains (ECOR12 and ECOR63) exhibited a round morphology with a clearly defined membrane, and in some cases double-membrane structures were observed. OMVs typically range from ~20–200 nm [21].
3.1.1. Protein Profile of OMVs (SDS-PAGE)
In addition to Cryo-TEM, the OMV protein profile of each sample was analyzed to relate banding patterns to the phylogenetic background of commensal vs. probiotic strains. A BenchMark™ Pre-Stained Protein Standard (Catalog #10747-012) was used as the molecular-weight marker.
The SDS-PAGE separation of OMV-associated proteins is shown in Figure 6.
The gel banding patterns differed among EcN, ECOR12, and ECOR63. EcN displayed a profile distinct from the commensal strains and showed a set of lower-molecular-weight bands (group A) relative to the other two strains. ECOR12 showed a pattern distinct from EcN and ECOR63 (group B2), which could plausibly reflect its different phylogenetic group assignment.
3.2. Maturation Profile of Dendritic Cells Stimulated with OMVs from Commensal and Probiotic Strains
DC maturation was evaluated using two complementary approaches: (i) ELISA quantification of secreted cytokines and (ii) flow cytometry assessment of surface maturation markers.
3.2.1. Cytokine Analysis by ELISA
To assess maturation-associated cytokine release after stimulation with OMVs from different strains, cytokines were measured by ELISA, focusing on IL-6, IL-10, and TNF-α. The maturation profile of immature DCs (iDC; control) was compared against DCs exposed to OMVs (mature DC condition) from EcN, ECOR12, and ECOR63. The ELISA quantifications are shown in Figure 7.
As shown in Figure 7, iDC controls exhibited very low cytokine release compared with OMV-stimulated DCs. For IL-10, ECOR12 OMVs induced a significantly higher release relative to EcN and ECOR63 OMVs. For IL-6, ECOR12 OMVs induced lower release than EcN and ECOR63, with ECOR63 showing the highest IL-6. For TNF-α, ECOR12 OMVs induced lower release than EcN and ECOR63, with ECOR63 again showing the strongest significance.
3.2.2. DC Maturation by Flow Cytometry
Immature monocyte-derived DCs were stimulated with 10 µg/mL OMVs from EcN and ECOR12 for 24 h at 37 °C and 5% CO₂. To verify maturation, cells were stained using a fluorochrome antibody cocktail (anti-CD14-FITC, anti-CD209-PE, anti-CD83-APC). The flow cytometry results are shown in Figure 8.
As indicated in Figure 8, the maturation marker CD83 exhibited higher fluorescence in OMV-treated conditions relative to the control, supporting successful DC maturation after OMV exposure.
3.3. Identification of miRNA Expression in DCs Stimulated with OMVs from Commensal and Probiotic Strains
DCs were stimulated with OMVs from EcN and ECOR12 to extract RNA and evaluate expression of selected miRNAs. The expression results for miR-155-5p and let-7i-3p are shown in Figure 9.
For miR-155-5p and let-7i-3p, OMVs from both EcN and ECOR12 induced significant expression relative to unstimulated controls. No statistically significant differences were detected between strains for miR-155-5p; for let-7i-3p, ECOR12 showed a higher trend without reaching significance. Differential expression results for miR-146b-5p and miR-29a-5p are shown in Figure 10.
3.4. PCA Biplot with k-Means Clustering (k = 3)
A principal component analysis (PCA) was applied to the combined dataset of cytokines (IL-6, IL-10, TNF-α; pg/mL) and miRNA expression (log2 miR-155-5p, let-7i-3p, miR-146b-5p, miR-29a-5p). The first two components captured most of the total variance, with PC1 = 75.0% and PC2 = 21.2% (cumulative 96.2%), indicating a strong low-dimensional structure suitable for visualization. In the PCA biplot (Figure 11), samples separated clearly across the PC1–PC2 space, and k-means (k = 3) produced three clusters that were largely concordant with strain-level structure. Vector loadings showed that IL-6 and TNF-α aligned with the upper-right region of the ordination (positive contribution in the direction of higher inflammatory response), whereas IL-10 loaded toward the lower portion of the plot, consistent with a distinct axis of variation driven by anti-inflammatory signaling. miRNA vectors (notably miR-29a-5p, miR-155-5p, miR-146b-5p, let-7i-3p) projected strongly away from the origin, supporting their high discriminatory contribution to sample separation.
3.5. Hierarchical Clustering Heatmap (Integrated Cytokines + miRNAs)
To evaluate global patterning and similarity structure across samples, we generated a heatmap using row-scaled (z-scored) values and performed hierarchical clustering (complete linkage) for both samples and variables. The resulting heatmap (Figure 12) revealed three dominant blocks aligned with the strain annotation bar, showing consistent within-strain profiles and strong between-strain contrasts. Controls exhibited broadly lower scaled values across most biomarkers (predominantly blue), whereas EcN and ECOR12 displayed elevated signatures across multiple variables (predominantly red), with clear differences in which markers were most enhanced. Variable clustering further indicated that biomarkers co-varied in biologically interpretable modules (e.g., inflammatory cytokines vs. regulatory miRNA patterns), supporting the PCA separation and motivating a formal multivariate test.
3.6. MANOVA (Global Effect of Strain) and Univariate Follow-Up with R² Integration
Given the coordinated behavior among biomarkers (as evidenced by PCA and hierarchical clustering), we tested whether strain produced a significant multivariate shift in the combined cytokine + miRNA response using MANOVA. The multivariate model was highly significant using the robust Pillai trace criterion: Pillai = 1.9882, F(14, 104) = 1246.5, p < 2.2×10⁻¹⁶, confirming that the biomarker profile differed strongly across strains (Table 6).
To interpret the multivariate effect, we performed post-MANOVA univariate ANOVAs (Table 7). All biomarkers showed strong strain effects (all p < 2.2×10⁻¹⁶), including cytokines (IL-6: F = 206.58; IL-10: F = 283.40; TNF-α: F = 102.94) and miRNAs (miR-155-5p: F = 1751.9; let-7i-3p: F = 1864.8; miR-146b-5p: F = 2836.5; miR-29a-5p: F = 4220.8). Importantly, the R² values (variance explained by strain) were high across biomarkers and especially pronounced for miRNAs (R² = 0.984–0.993), indicating that miRNA expression provided the strongest discriminatory power between strains, while cytokines also contributed substantially (IL-6 R² = 0.879; IL-10 R² = 0.909; TNF-α R² = 0.783) (Table 7).
We additionally tested cytokines and miRNAs as separate multivariate families. Cytokines alone remained strongly strain-dependent (Pillai = 1.8579, F(6, 112) = 243.98, p < 2.2×10⁻¹⁶) and miRNAs alone showed an even larger multivariate separation (Pillai = 1.9797, F(8, 110) = 1339.2, p < 2.2×10⁻¹⁶) (Table 6), consistent with the higher R² values observed for miRNA outcomes.
4. Discussion
Inflammatory disorders of the gastrointestinal tract continue to rise in prevalence and societal impact, and they remain difficult to treat precisely because disease trajec-tories emerge from multilayered interactions among microbial communities, epithelial barrier physiology, and innate–adaptive immune crosstalk [1,2,3,4]. In this context, the pre-sent study is relevant because it focuses on a mechanistically “addressable” unit of microbiota–host communication: bacterial extracellular vesicles, specifically Gram-negative outer membrane vesicles (OMVs). OMVs can package and deliver im-munologically active cargo (e.g., lipopolysaccharide, outer membrane proteins, pepti-doglycan fragments, DNA/RNA, and small RNAs) to epithelial and immune cells, ena-bling bacteria to shape host signaling without direct bacterial translocation [14,15]. This concept aligns with a growing consensus that microbiota-derived vesicles are not incidental byproducts but structured mediators of gut homeostasis and disease, with effects that depend strongly on vesicle origin, cargo composition, and the responding host compartment [13,16,17,19,20,56,57]. Accordingly, by con-trasting OMVs from a clinically used probiotic strain (Escherichia coli Nissle 1917, EcN) with those from commensal E. coli strains (ECOR12 and ECOR63), our work ad-dresses a persistent gap: why some microbiota-derived cues support immune balance and barrier integrity, whereas others favor inflammatory amplification.
4.1. OMV Heterogeneity Is Measurable and Biologically Meaningful
A necessary premise for interpreting immunological effects is that vesicle prepa-rations reflect biologically plausible OMV populations. Our Cryo-TEM images (Figure 5) supported the expected morphology of OMVs—rounded vesicles with a clearly de-fined membrane and occasional double-membrane structures—consistent with re-ported OMV architectures and with descriptions of outer–inner membrane vesicles in Gram-negative bacteria [21,22]. The observed na-noscale size range (classically ~20–200 nm, though broader distributions occur across species and preparation conditions) is also compatible with prior work and reinforces that OMVs represent particulate signals capable of traversing mucus and contacting epithelial or immune interfaces [15,21]. Importantly, vesicle-mediated signaling is not restricted to Gram-negative organisms; extracellular vesicles are also produced by Gram-positive bacteria and fungi, empha-sizing that vesiculation is a conserved communication strategy [18]. This broader framework strengthens the interpretation that the vesicles observed here plausibly act as cross-kingdom information carriers rather than inert debris [13,74].
Our protein-based quantification approach (Table 1), based on the Lowry assay, provided standardized inputs for downstream comparisons and follows a widely used biochemical method for protein measurement [75]. Nevertheless, pro-tein mass alone cannot capture vesicle number, size distribution, or cargo stoichiome-try, and thus should be interpreted as a practical normalization strategy rather than a complete physical characterization. This limitation is widely recognized in OMV/EV studies, where particle-based quantification and orthogonal characterization methods are encouraged to strengthen cross-study comparability [14,15].
Beyond morphology, SDS-PAGE profiles (Figure 6) demonstrated that OMVs from EcN, ECOR12, and ECOR63 contain distinguishable protein patterns, consistent with the concept of selective cargo loading and strain-dependent OMV composition [36,37]. Such differences are expected because OMV bi-ogenesis and protein sorting can depend on outer membrane remodeling, stress re-sponses, and envelope composition, which vary across strains and growth contexts [15]. The proteomic literature further supports that OMV cargo is non-random and often enriched for membrane-associated and immunologi-cally active components, consistent with roles in host interaction and immune modu-lation [36,37]. However, our comparison also highlights an important interpretive caution: while protein banding patterns can reflect biologi-cal differences, they are not a reliable proxy for phylogeny. Although classical E. coli phylogenetic grouping (e.g., A, B1, B2, D) provides a valuable framework for inter-preting strain background and potential functional tendencies, SDS-PAGE alone can-not resolve phylogeny, particularly when strains share major phylogroups or when vesicle cargo is shaped by environmental regulation rather than lineage alone [34,35]. This reinforces the methodological point that “omics” resolution (proteomics/transcriptomics) and genetic typing are needed to map vesicle composition to strain background with confidence [35,36,37].
4.2. OMVs Can Mature Dendritic Cells, but the Direction of the Response Is Strain-Dependent
A central finding of this work is that OMVs from commensal and probiotic E. coli strains can drive dendritic cell (DC) maturation, as supported by cytokine readouts (Figure 7) and by flow cytometric changes in maturation markers (Figure 8). This out-come is mechanistically plausible because DCs are designed to integrate microbial sig-nals and translate them into adaptive immune instruction [11,12]. In the gut, DCs operate in continuous dialogue with epithelial cells, sampling luminal information directly or indirectly, and shaping T-cell polariza-tion and tolerance programs that maintain homeostasis [6,10]. Disruption of this epithelial–DC crosstalk is therefore a credible route to chronic inflammation and barrier dysfunction [6,10].
From a mechanistic standpoint, OMVs are well positioned to engage pattern recognition receptors (PRRs) because they can carry pathogen-associated molecular patterns (PAMPs) such as LPS, lipoproteins, and peptidoglycan fragments. PRR activa-tion (e.g., TLRs and NOD-like receptors) initiates innate immune cascades and can also intersect with autophagy-linked immune programs, shaping antigen handling and in-flammatory outcomes [5,28]. Consistent with this, prior studies in diverse Gram-negative contexts show that OMVs can modulate epithelial and immune signaling, including NOD1-dependent pathways and DC-mediated T helper polarization [23,24,26]. Importantly, not all OMVs drive the same immunological “tone”: pathogenic OMVs may amplify inflammation and systemic responses, whereas commensal or probiotic OMVs may support balanced defense or immunoregulation depending on cargo and context [16,17,21,22].
Our cytokine profiling (Figure 7) is consistent with this duality. IL-6 and TNF-α are classically associated with inflammatory activation and myeloid signaling cas-cades, whereas IL-10 is a key immunoregulatory cytokine that helps restrain excessive inflammation and supports tolerance programs in mucosal environments [6,10]. The observation that ECOR12-stimulated DCs exhib-ited comparatively higher IL-10, while ECOR63 tended toward higher IL-6 and TNF-α, suggests that commensal strains are not immunologically interchangeable and that OMVs may encode “strain-specific immunological instructions.” This aligns with work showing that vesicles from different E. coli backgrounds distinctly modulate human DCs and subsequently shape T-cell responses [55]. It also fits broader models in which microbiota-derived vesicles can act as immunoregulatory agents, potentially contributing to tolerance and homeostasis rather than indiscrimi-nate activation [20,56,57].
Flow cytometry results (Figure 8) provide an orthogonal validation that DCs un-derwent maturation-associated changes. The increased expression of maturation marker CD83 and the concurrent shifts in monocyte/DC-associated markers are con-sistent with DC activation in response to microbial stimulation. This is mechanistically coherent with DC biology, where maturation is accompanied by phenotypic repro-gramming that supports antigen presentation, costimulatory capacity, and cytokine production [12]. Notably, DC polarization is not fully captured by a single marker, and literature emphasizes that phenotype and functional polarization can dissociate depending on stimulation context—an important interpretive point when comparing vesicle-driven maturation across strains [27]. Addi-tionally, vesicles from other beneficial microbes have been reported to induce regula-tory immune outcomes, including Treg-associated signatures, supporting the broader plausibility that bacterial vesicles can promote tolerance-linked DC programs rather than exclusively inflammatory ones.
4.3. EcN OMVs as Postbiotic-like Immunomodulators—and Why “Probiotic” Is Not Equivalent to “Anti-Inflammatory”
EcN is a particularly informative model because it occupies a unique position as a non-pathogenic, clinically used probiotic with documented colonization ability and immunomodulatory potential [29,30]. EcN’s persistence and functional interactions can involve adhesion/colonization factors and fimbriae, which support biofilm formation and intestinal colonization [31]. Additional surface and capsule-associated features can shape epithelial PRR ac-tivation and downstream signaling, including MAPK-dependent cytokine induction [32]. This background makes EcN an ideal strain to test the hypothesis that probiotic effects can be mediated not only by live bacteria but also by secreted vesicles as cell-free effectors [14,15].
Consistent with this “postbiotic-like” model, EcN OMVs have been shown to ac-tivate mucosal defense programs and to yield anti-inflammatory effects in experi-mental colitis settings, supporting therapeutic relevance in inflammatory disease con-texts [33]. EcN vesicles can also protect epithelial barrier integrity against enteropathogenic insult, reinforcing the concept that vesicles can contribute to barrier resilience [70]. At the same time, EcN is not uni-formly anti-inflammatory; rather, its effects can be context-dependent, reflecting the nuanced reality that immune balance often requires controlled activation rather than suppression [29,30]. This nuance also matches broader clinical and nutritional literature on probiotics and IBD, which emphasizes variability across strains, patient contexts, and disease states [7,8]. Our findings—that EcN OMVs mature DCs and alter cytokine/miRNA signatures—fit well within this framework: EcN may contribute to “trained balance” rather than simple anti-inflammatory silencing, consistent with mucosal immunology principles [6,11].
4.4. miRNAs Provide a Mechanistic Layer That Can Reconcile Cytokine Differences with Durable Immune Programming
One of the most important conceptual advances in mucosal immunology is that immune outputs are controlled not only by receptor–ligand interactions and transcrip-tion factors, but also by post-transcriptional regulation mediated by microRNAs (miRNAs) [39,40,46]. miRNAs are now understood as essential modulators of DC differentiation and function, shaping the magnitude and duration of innate signaling and cytokine programs [48]. Inflammatory and regulatory circuits can be tuned by miRNA-mediated control of PRR signaling components, cytokine feedback loops, and antigen presentation pathways [44,45]. This is highly relevant for DC biology because DCs are “decision-making” cells that must translate microbial cues into either tolerance-supporting programs or effector responses [11,12].
Our miRNA results (Figure 9 and Figure 10) can be interpreted coherently through this lens. miR-155 is a canonical inflammation-associated miRNA induced during macrophage and DC activation; it modulates cytokine production and has been directly implicated in IL-1 pathway regulation in activated monocyte-derived DCs [41,42]. The observation that miR-155-5p increases with OMV stimula-tion (EcN and ECOR12) is therefore consistent with OMVs acting as im-mune-activating signals that engage innate pathways and reprogram DC regulatory circuits [5,28]. Similarly, miR-146 family members are widely described as negative feedback regulators that restrain excessive TLR signaling; IL-10-dependent miR-146b, for example, suppresses TLR4 pathway activity, providing a plausible mechanism by which regulatory cytokine environments stabilize homeo-static immune tone [43,47]. Our finding that miR-146b-5p is induced and may differ between strains aligns with a model where vesicles can simultaneously trigger activation and encode braking mecha-nisms—precisely the kind of balanced programming required at mucosal surfaces [6,10].
The let-7 family provides an additional bridge between innate sensing and miRNA regulation. let-7i can regulate TLR4 expression and thereby influence epithelial im-mune responsiveness to microbial stimuli [52]. Trends toward differen-tial let-7i-3p expression across strains may therefore reflect differences in vesicle cargo that alter PRR pathway setpoints. This possibility is consistent with the broader litera-ture on miRNAs as “fine-tuners” of TLR signaling, shaping inflammatory thresholds rather than simply turning pathways on or off [44,45,46].
Crucially, miRNA biology in gut disease extends beyond immune cells: epithelial permeability and barrier integrity are also regulated by miRNAs. miR-29a, in particu-lar, has been associated with increased intestinal permeability in human and experi-mental contexts, and manipulation of miR-29a can influence tight junction-related proteins such as ZO-1 and claudins [49,50,51]. Thus, our observation that miR-29a-5p is induced and differs between EcN and ECOR12 conditions is biologically meaningful because it suggests that OMVs might in-fluence mucosal outcomes by simultaneously programming DC immunity and barri-er-associated regulatory pathways. This integrative view is supported by reviews em-phasizing that miRNAs participate in gastrointestinal cell signaling and are implicated in inflammatory bowel disease pathophysiology and immune regulation [58,59].
An especially relevant emerging theme is that bacterial vesicles themselves can carry small RNAs capable of influencing host responses. Small RNAs within OMVs have been proposed as mediators of host–microbe interactions, expanding the mecha-nistic repertoire beyond proteins and classical PAMPs [53]. Our findings are consistent with this broader hypothesis, particularly when interpreted alongside transcriptomic evidence that gut microbiota-secreted vesicles can induce distinct miRNA signatures in DCs [54]. In other words, vesicles may not merely trigger immediate cytokine outputs; they may “write” durable regulatory states into immune cells via miRNA-linked circuits.
4.5. Linking Strain-Specific OMV Signaling to Epithelial Interfaces and Disease Relevance
The gut environment is characterized by continuous epithelial exposure to micro-bial products, requiring robust mechanisms to preserve barrier function while enabling rapid defense. Intestinal epithelial cells regulate barrier architecture, PRR signaling, antimicrobial peptide production, and immune communication, and they therefore represent a key site where OMVs can exert disease-relevant effects [6]. OMV uptake routes such as clathrin-dependent endocytosis have been demon-strated for E. coli vesicles in epithelial models, and vesicle internalization can produce divergent outcomes (including DNA damage responses) depending on cargo and con-text [25]. These mechanisms are not merely technical details: they imply that vesicle effects may be cell-type specific and may differ between epithelial and immune compartments, a point reinforced by broader OMV–host interaction literature [16,17,74].
From a translational perspective, OMVs are therefore plausible candidates for next-generation microbiota-derived interventions (e.g., postbiotics) in gut inflamma-tory conditions, provided that immunological directionality and safety constraints are established. Clinical and mechanistic reviews emphasize that while probiotics can benefit gastrointestinal disorders, efficacy is strain-specific and context-dependent, and inappropriate immune activation remains a concern [7,8]. This caution becomes especially relevant when one considers that OMVs from some Gram-negative contexts can induce systemic inflammatory phenotypes [21] and that pathogenic vesicles can support virulence and host cell manipulation [22,26]. Importantly, Sat has been reported not to be-have as a virulence factor in EcN. Our results support a more pre-cise framing: the question is not whether OMVs are “good” or “bad,” but which vesicle programs (defined by strain, cargo, and recipient cell type) promote balanced mucosal defense versus inflammatory escalation [19,20,57].
Recent work also supports the plausibility of cell-type-resolved OMV effects in immune systems beyond DCs. For example, EcN-derived OMVs have been reported to enhance immunomodulation and antimicrobial activity in macrophage contexts [72] and to influence macrophage polarization and colitis outcomes [73]. At epithelial levels, microbiota EVs can modulate nutrient transporters and ser-otonin-related genes via miRNA-associated pathways, indicating that vesicles can re-shape homeostasis-relevant gene networks [38]. These data strengthen the interpretation that vesicles can coordinate immune and epithelial axes—precisely the kind of multi-compartment reg-ulation implicated in IBD and related inflammatory conditions [1,2,6].
4.6. Why Multivariate Statistics Are Essential for OMV–Immune Datasets
Because OMV responses involve coordinated changes across cytokines, markers, and miRNA panels, multivariate approaches are essential to avoid over-interpreting isolated endpoints. PCA provides a principled way to compress correlated variables into interpretable axes, and biplots allow loadings to be visualized as vectors that clar-ify which variables drive group separation [60,61,62]. In parallel, clustering and heatmap-based pattern discovery can identify coherent response modules across conditions, supporting mechanistic inter-pretation rather than single-marker narratives [63,64,65]. Our use of these approaches is therefore aligned with best practices for interpreting complex immunological profiles, particularly when vesicle responses may differ subtly across strains.
Formal multivariate inference also matters. When outcomes are multivariate by nature, MANOVA-based tests provide a framework to evaluate condition effects, and classical work highlights that certain test statistics (e.g., Pillai’s trace) can show favor-able robustness properties under assumption deviations [66]. Robust MANOVA alternatives further extend inference reliability when distributions, covar-iance structures, or outliers challenge classical assumptions [67]. Finally, biological interpretation requires effect-size thinking: statistical signifi-cance does not automatically imply biological relevance, and careful reporting of effect sizes and uncertainty is recommended in experimental biology [68]. Even common summary metrics such as R² require interpretive caution; R² is sensitive to model structure and does not alone validate causality or mechanistic ade-quacy [69]. Together, these statistical principles justify the analytic strat-egy used in this study and support more reliable mechanistic claims about strain-specific immunomodulation.
Taken together, our findings support an integrated model in which OMVs from probiotic and commensal E. coli strains act as structured immune stimuli capable of driving DC maturation and reprogramming miRNA-linked regulatory circuits. The strain-specific divergence in cytokine patterns (e.g., IL-10 vs. IL-6/TNF-α tendencies) and miRNA signatures (miR-155, miR-146b, let-7i, miR-29a) is consistent with the concept that vesicles encode “immune instruction sets” rather than triggering uniform activation. This interpretation aligns with mechanistic literature on OMV biogenesis and host interaction [15,74], with mucosal immunology principles emphasizing epithelial–DC crosstalk [10,11], and with emerging evidence that microbiota vesicles can shape cytokine and miRNA landscapes relevant to gut homeostasis and inflammatory disease [20,54,55,56,57]. In the long term, these results reinforce the rationale for moving beyond “microbiota as a black box” toward testable, cell-free vesicle mechanisms that can be quantified, modeled, and potentially engi-neered for targeted mucosal immunomodulation [5,6,14].
5. Limitations and Future Directions
Several limitations should guide future work. First, while OMV protein normali-zation is practical [75], particle-level quantification (e.g., nanoparticle tracking analysis) and cargo-specific assays would strengthen conclusions about dose–response relationships and mechanistic drivers [14,15]. Second, SDS-PAGE provides a coarse view of vesicle composition; proteomics and RNA profiling would better connect cargo differences to immunological outputs [36,37,53]. Third, in vitro DC maturation models capture important biology but cannot fully represent the intestinal microenvi-ronment, where epithelial conditioning, mucus, metabolites, and multi-cellular inter-actions shape immune thresholds [4,6,10]. Future studies could integrate epithelial–DC co-cultures, macrophage compartments, and barrier readouts to test whether miRNA changes observed here translate into epithelial integrity and tolerance outcomes under inflammatory stress [25,38,72].
Finally, translational relevance requires careful framing within clinical realities of gut disease and exposure routes. IBD and chronic inflammatory disorders have multi-factorial etiologies and can be influenced by infections, diet, and public-health condi-tions that shape microbial exposures [1,3]. The global food safety perspective underscores that microbial exposures are not purely individualized but occur within population-level sys-tems—an angle that becomes increasingly important when considering microbio-ta-directed interventions or vesicle-based therapeutics. Thus, OMV-based strategies must be evaluated not only for immunological efficacy but also for safety, reproducibility, and context-dependent effects [16,17,19].
6. Conclusions
In conclusion, this study provides mechanistic insight into how outer membrane vesicles (OMVs) released by probiotic and commensal Escherichia coli strains modulate human monocyte-derived dendritic cells (DCs) at both the phenotypic and post-transcriptional levels. We show that OMVs from EcN, ECOR12 and ECOR63 are not inert by-products of bacterial growth but structured stimuli that consistently promote DC maturation, as evidenced by increased CD83 expression and the secretion of IL-6, IL-10 and TNF-α. Beyond this shared activation profile, our data reveal clear strain-specific signatures: OMVs from ECOR12 preferentially drive an IL-10–high, miR-146b-5p–enriched response compatible with negative feedback regulation of TLR signalling, whereas EcN OMVs favour relatively higher TNF-α, IL-6 and miR-29a-5p levels, linking probiotic vesicles to regulatory nodes involved in inflammatory control and epithelial barrier homeostasis. The coordinated induction of miR-155-5p and let-7i-3p across conditions, together with hierarchical clustering and PCA, further supports the concept that the OMV “origin” imprints distinct cytokine/miRNA fingerprints on DCs rather than eliciting a uniform probiotic or commensal response.
By validating microarray-derived candidates at the level of individual miRNAs, this work strengthens the evidence that DC–OMV interactions are tightly coupled to miRNA-dependent regulatory circuits. The integration of protein readouts and miRNA profiles delineates a set of miRNAs—particularly miR-155-5p, let-7i-3p, miR-146b-5p and miR-29a-5p—that emerge as central hubs linking OMV sensing to pathways controlling NF-κB activation, TLR tolerance, cytokine output and, potentially, antigen presentation. These strain-specific miRNA signatures provide a plausible molecular basis for the differential immunomodulatory properties described for EcN and commensal strains in vivo, and suggest that OMVs act as nanoscale conveyors of bacterial “information” capable of fine-tuning DC activation thresholds rather than simply switching immune responses on or off.
Although our study is limited to an in vitro model based on monocyte-derived DCs and a restricted panel of OMVs and miRNAs, the consistency of the patterns observed across donors and strains argues that these vesicle-induced programs are robust. Future work should address how these DC reprogramming events translate into T-cell priming, regulatory versus effector polarization, and epithelial integrity in more complex systems, including intestinal organoids and in vivo models of mucosal inflammation. Moreover, expanding the analysis to OMVs from additional members of the gut microbiota, and to broader miRNA and transcriptomic panels, will help determine whether the cytokine/miRNA fingerprints identified here can be developed as mechanistic biomarkers of microbiota–immune crosstalk. Overall, our findings support a model in which gut-derived OMVs, particularly those from probiotic EcN and selected commensals, represent promising non-viable “post-biotic” candidates to rationally modulate DC function and, ultimately, to inform the design of OMV-based interventions for intestinal inflammatory and immune-mediated disorders.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, K.R-P, B.P-G., L. C-M. P. R-G, A. J-G & E. R-N.; methodology, K.R-P., B.P-G., L. C-M., M. A-F, P. R-G & E. R-N; software, K. R-P., S.V-A & P. F-A.; validation, K.R-P., B.P-G, P. R-G., A. J-G, L.C-M & E. R-N; formal analysis, K. R-P, B. P-G. S.V-A. & P. F-A.; investigation, K. R-P, B.P-G., S. A-G & E. R-N; resources, K.R-P. & E. R-N.; data curation, S.V-A. & P. F-A.; writing—original draft preparation, K.R-P.; writing—review and editing, K.R-P., B. P-G, L. C-M, A. J-G, S. A-G & S. V-A; visualization, K. R-P., L. C-M & M. A-F; supervision, K.R-P., M A-F., P.R-G, A. J-G, S. A-G. & E. R-N; project administration, E.R-N; funding acquisition, UNEMI. All authors have read and agreed to the published version of the manuscript.”.
Funding
This research received no external founding.
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding author.
Acknowledgments
The authors are grateful to the Universidad Estatal de Milagro (UNEMI) for supporting our publication.
Conflicts of Interest
The authors declare no conflicts of interest.
Appendix A

Table A1.
Primers used for miRCURY® LNA® miRNA PCR.
| Group 1 | Group 2 | Group 3 | Normalizers | |||||
| Name | ID | Name | ID | Name | ID | Name | ID | |
| miR-146b-5p | MIMAT0002809 | hsa-miR-29a-5p | MIMAT0004503 | hsa-miR-155-5p | MIMAT0000646 | hsa−let−7f−5p | MIMAT0000067_1 | |
| hsa-miR-24-3p | MIMAT0000080_1 | hsa-miR-33a-3p | MIMAT0004506 | hsa-miR-146a-5p | MIMAT0000449 | hsa−miR−421 | MIMAT0003339 | |
| hsa-miR-10a-5p | MIMAT0000253 | hsa-miR-589-3p | MIMAT0003256 | hsa-let-7i-3p | MIMAT0004585 | |||
References
- Kaplan, G. G. The global burden of IBD: From 2015 to 2025. Nature Reviews Gastroenterology & Hepatology 2015, 12(12), 720–727. [Google Scholar] [CrossRef] [PubMed]
- Triantafillidis, J. K.; Merikas, E.; Georgopoulos, F. Current and emerging drugs for the treatment of inflammatory bowel disease. Drug Design, Development and Therapy 2011, 5, 185–210. [Google Scholar] [CrossRef] [PubMed]
- Purchiaroni, F.; Tortora, A.; Gabrielli, M.; Bertucci, F.; Gigante, G.; Ianiro, G.; Gasbarrini, A. The role of intestinal microbiota and the immune system. European Review for Medical and Pharmacological Sciences 2013, 17(3), 323–333. [Google Scholar] [PubMed]
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochemical Journal 2017, 474(11), 1823–1836. [Google Scholar] [CrossRef]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124(4), 783–801. [Google Scholar] [CrossRef]
- Peterson, L. W.; Artis, D. Intestinal epithelial cells: Regulators of barrier function and immune homeostasis. Nature Reviews Immunology 2014, 14(3), 141–153. [Google Scholar] [CrossRef]
- Oliveira, G.; Gonzalez, I. An update on probiotics, prebiotics and symbiotics in clinical nutrition. Endocrinología y Nutrición 2016, 63(9), 482–494. [Google Scholar] [CrossRef]
- Sanchez, B.; Delgado, S.; Blanco-Miguez, A.; Lourenco, A.; Gueimonde, M.; Margolles, A. Probiotics, gut microbiota, and their influence on host health and disease. Molecular Nutrition & Food Research 2017, 61(1), e1600240. [Google Scholar] [CrossRef]
- O’Hara, A. M.; Shanahan, F. The gut flora as a forgotten organ. EMBO Reports 2006, 7(7), 688–693. [Google Scholar] [CrossRef]
- Rimoldi, M.; Chieppa, M.; Salucci, V.; Avogadri, F.; Sonzogni, A.; Sampietro, G. M.; Rescigno, M. Intestinal immune homeostasis is regulated by the crosstalk between epithelial cells and dendritic cells. Nature Immunology 2005, 6(5), 507–514. [Google Scholar] [CrossRef]
- Rescigno, M.; Chieppa, M. Gut-level decisions in peace and war. Nature Medicine 2005, 11(3), 254–255. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, M. B.; Sureda, M.; Rebollo, J. Células dendríticas I: aspectos básicos de su biología y funciones. Inmunología 2012, 31(1), 21–30. [Google Scholar] [CrossRef]
- Caruana, J. C.; Walper, S. A. Bacterial membrane vesicles as mediators of microbe–microbe and microbe–host community interactions. Frontiers in Microbiology 2020, 11, 432. [Google Scholar] [CrossRef] [PubMed]
- Jan, A. T. Outer membrane vesicles (OMVs) of Gram-negative bacteria: A perspective update. Frontiers in Microbiology 2017, 8, 1053. [Google Scholar] [CrossRef]
- Schwechheimer, C.; Kuehn, M. J. Outer-membrane vesicles from Gram-negative bacteria: Biogenesis and functions. Nature Reviews Microbiology 2015, 13(10), 605–619. [Google Scholar] [CrossRef]
- Kaparakis-Liaskos, M.; Ferrero, R. L. Immune modulation by bacterial outer membrane vesicles. Nature Reviews Immunology 2015, 15(6), 375–387. [Google Scholar] [CrossRef]
- Pathirana, R. D.; Kaparakis-Liaskos, M. Bacterial membrane vesicles: Biogenesis, immune regulation and pathogenesis. Cellular Microbiology 2016, 18(11), 1518–1524. [Google Scholar] [CrossRef]
- Brown, L.; Wolf, J. M.; Prados-Rosales, R.; Casadevall, A. Through the wall: Extracellular vesicles in Gram-positive bacteria, mycobacteria and fungi. Nature Reviews Microbiology 2015, 13(10), 620–630. [Google Scholar] [CrossRef]
- Charpentier, L. A.; Pita-Grisanti, V.; Valvano, M. A. Bacterial outer membrane vesicles and immune modulation of the host. Membranes 2023, 13(9), 752. [Google Scholar] [CrossRef]
- Meng, R.; Zeng, M.; Ji, Y.; Huang, X.; Xu, M. The potential role of gut microbiota outer membrane vesicles in colorectal cancer. Frontiers in Microbiology 2023, 14, 1270158. [Google Scholar] [CrossRef]
- Park, K. S.; Choi, K. H.; Kim, Y. S.; Hong, B. S.; Kim, O. Y.; Kim, J. H.; Gho, Y. S. Outer membrane vesicles derived from Escherichia coli induce systemic inflammatory response syndrome. PLOS ONE 2010, 5(6), e11334. [Google Scholar] [CrossRef] [PubMed]
- Perez-Cruz, C.; Delgado, L.; Lopez-Iglesias, C.; Mercade, E. Outer–inner membrane vesicles naturally secreted by Gram-negative pathogenic bacteria. PLOS ONE 2015, 10(1), e0116896. [Google Scholar] [CrossRef] [PubMed]
- Cañas, M. A.; Fábrega, M. J.; Giménez, R.; Badia, J.; Baldomà, L. Outer membrane vesicles from probiotic and commensal Escherichia coli activate NOD1-mediated immune responses in intestinal epithelial cells. Frontiers in Microbiology 2017, 8, 498. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Giardino Torchia, M. L.; Lawson, G. W.; Karp, C. L.; Ashwell, J. D.; Mazmanian, S. K. Outer membrane vesicles of a human commensal mediate immune regulation and disease protection. Cell Host & Microbe 2012, 12(4), 509–520. [Google Scholar] [CrossRef]
- Cañas, M. A.; Gimenez, R.; Fabrega, M. J.; Toloza, L.; Baldoma, L.; Badia, J. Outer membrane vesicles from the probiotic Escherichia coli Nissle 1917 and the commensal ECOR12 enter intestinal epithelial cells via clathrin-dependent endocytosis and elicit differential effects on DNA damage. PLOS ONE 2016, 11(8), e0160374. [Google Scholar] [CrossRef]
- Chatterjee, D.; Chaudhuri, K. Vibrio cholerae O395 outer membrane vesicles modulate intestinal epithelial cells in a NOD1 protein-dependent manner and induce dendritic cell-mediated Th2/Th17 cell responses. Journal of Biological Chemistry 2013, 288(6), 4299–4309. [Google Scholar] [CrossRef]
- Mester, B.; Bauer, E.; Wood, C. E.; Hermans, I. F.; Gasser, O. Expression of CD1a and type-1 polarization are dissociated in human monocyte-derived dendritic cells. PLOS ONE 2015, 10(10), e0140432. [Google Scholar] [CrossRef]
- Tang, D.; Kang, R.; Coyne, C. B.; Zeh, H. J.; Lotze, M. T. PAMPs and DAMPs: Signal 0s that spur autophagy and immunity. Immunological Reviews 2012, 249(1), 158–175. [Google Scholar] [CrossRef]
- Sonnenborn, U. Escherichia coli strain Nissle 1917—from bench to bedside and back: History of a special Escherichia coli strain with probiotic properties. FEMS Microbiology Letters 2016, 363(19), fnw212. [Google Scholar] [CrossRef]
- Trebichavsky, I.; Splichal, I.; Rada, V.; Splichalova, A. Modulation of natural immunity in the gut by Escherichia coli strain Nissle 1917. Nutrition Reviews 2010, 68(8), 459–464. [Google Scholar] [CrossRef]
- Lasaro, M. A.; Salinger, N.; Zhang, J.; Wang, Y.; Zhong, Z.; Goulian, M.; Zhu, J. F1C fimbriae play an important role in biofilm formation and intestinal colonization by the Escherichia coli commensal strain Nissle 1917. Applied and Environmental Microbiology 2009, 75(1), 246–251. [Google Scholar] [CrossRef] [PubMed]
- Hafez, M.; Hayes, K.; Goldrick, M.; Grencis, R. K.; Roberts, I. S. The K5 capsule of Escherichia coli strain Nissle 1917 is important in stimulating expression of Toll-like receptor 5, CD14, MyD88, and TRIF together with the induction of interleukin-8 expression via the mitogen-activated protein kinase pathway in epithelial cells. Infection and Immunity 2010, 78(5), 2153–2162. [Google Scholar] [CrossRef] [PubMed]
- Fabrega, M. J.; Rodriguez-Nogales, A.; Garrido-Mesa, J.; Algieri, F.; Badia, J.; Gimenez, R.; Baldoma, L. Intestinal anti-inflammatory effects of outer membrane vesicles from Escherichia coli Nissle 1917 in DSS-experimental colitis in mice. Frontiers in Microbiology 2017, 8, 1274. [Google Scholar] [CrossRef] [PubMed]
- Ochman, H.; Selander, R. K. Standard reference strains of Escherichia coli from natural populations. Journal of Bacteriology 1984, 157(2), 690–693. [Google Scholar] [CrossRef]
- Clermont, O.; Bonacorsi, S.; Bingen, E. Rapid and simple determination of the Escherichia coli phylogenetic group. Applied and Environmental Microbiology 2000, 66(10), 4555–4558. [Google Scholar] [CrossRef]
- Aguilera, L.; Toloza, L.; Gimenez, R.; Odena, A.; Oliveira, E.; Aguilar, J.; Baldoma, L. Proteomic analysis of outer membrane vesicles from the probiotic strain Escherichia coli Nissle 1917. Proteomics 2014, 14(2–3), 222–229. [Google Scholar] [CrossRef]
- Lee, J.; Kim, O. Y.; Gho, Y. S. Proteomic profiling of Gram-negative bacterial outer membrane vesicles: Current perspectives. Proteomics—Clinical Applications 2016, 10(9–10), 897–909. [Google Scholar] [CrossRef]
- Olivo-Martínez, Y. A.; Díaz-Garrido, N.; Vera, R.; Badia, J.; Baldomà, L. Extracellular vesicles of the probiotic Escherichia coli Nissle 1917 reduce PepT1 levels in IL-1β-treated Caco-2 cells via upregulation of miR-193a-3p. Nutrients 2024, 16(16), 2719. [Google Scholar] [CrossRef]
- Lindsay, M. A. MicroRNAs and the immune response. Trends in Immunology 2008, 29(7), 343–351. [Google Scholar] [CrossRef]
- Sonkoly, E.; Ståhle, M.; Pivarcsi, A. MicroRNAs and immunity: Novel players in the regulation of normal immune function and inflammation. Seminars in Cancer Biology 2008, 18(2), 131–140. [Google Scholar] [CrossRef]
- O’Connell, R. M.; Taganov, K. D.; Boldin, M. P.; Cheng, G.; Baltimore, D. MicroRNA-155 is induced during the macrophage inflammatory response. Proceedings of the National Academy of Sciences of the United States of America 2007, 104(5), 1604–1609. [Google Scholar] [CrossRef] [PubMed]
- Ceppi, M.; Pereira, P. M.; Dunand-Sauthier, I.; Barras, E.; Reith, W.; Santos, M. A.; Pierre, P. MicroRNA-155 modulates the interleukin-1 signalling pathway in activated human monocyte-derived dendritic cells. Proceedings of the National Academy of Sciences of the United States of America 2009, 106(8), 2735–2740. [Google Scholar] [CrossRef] [PubMed]
- Curtale, G.; Mirolo, M.; Renzi, T. A.; Rossato, M.; Bazzoni, F.; Locati, M. Negative regulation of Toll-like receptor 4 signalling by IL-10-dependent microRNA-146b. Proceedings of the National Academy of Sciences of the United States of America 2013, 110(28), 11499–11504. [Google Scholar] [CrossRef] [PubMed]
- Bayraktar, R.; Bertilaccio, M. T. S.; Calin, G. A. The interaction between two worlds: MicroRNAs and Toll-like receptors. Frontiers in Immunology 2019, 10, 1053. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, L. A. J.; Sheedy, F. J.; McCoy, C. E. MicroRNAs: The fine-tuners of Toll-like receptor signalling. Nature Reviews Immunology 2011, 11(3), 163–175. [Google Scholar] [CrossRef]
- Rebane, A.; Akdis, C. A. MicroRNAs: Essential players in the regulation of inflammation. Journal of Allergy and Clinical Immunology 2013, 132(1), 15–26. [Google Scholar] [CrossRef]
- Taganov, K. D.; Boldin, M. P.; Chang, K. J.; Baltimore, D. NF-κB-dependent induction of microRNA miR-146, an inhibitor targeted to signaling proteins of innate immune responses. Proceedings of the National Academy of Sciences of the United States of America 2006, 103(33), 12481–12486. [Google Scholar] [CrossRef]
- Turner, M. L.; Schnorfeil, F. M.; Brocker, T. MicroRNAs regulate dendritic cell differentiation and function. The Journal of Immunology 2011, 187(8), 3911–3917. [Google Scholar] [CrossRef]
- Zhou, Q.; Souba, W. W.; Croce, C. M.; Verne, G. N. MicroRNA-29a regulates intestinal membrane permeability in patients with irritable bowel syndrome. Gut 2010, 59(6), 775–784. [Google Scholar] [CrossRef]
- Chao, G.; Wang, Y.; Zhang, S.; Yang, W.; Ni, Z.; Zheng, X. MicroRNA-29a increases intestinal membrane permeability in a rat model of irritable bowel syndrome. Oncotarget 2017, 8(50), 85828–85837. [Google Scholar] [CrossRef]
- Zhu, H.; Xiao, X.; Shi, Y.; Wu, Y.; Huang, Y.; Li, D.; Wang, X. Inhibition of miRNA-29a regulates intestinal barrier function in diarrhea-predominant irritable bowel syndrome by upregulating ZO-1 and CLDN1. Experimental and Therapeutic Medicine 2020, 20(2), 1071–1080. [Google Scholar] [CrossRef] [PubMed]
- Chen, X. M.; Splinter, P. L.; O’Hara, S. P.; LaRusso, N. F. A cellular microRNA, let-7i, regulates Toll-like receptor 4 expression and contributes to cholangiocyte immune responses against Cryptosporidium parvum infection. Journal of Biological Chemistry 2007, 282(39), 28929–28938. [Google Scholar] [CrossRef] [PubMed]
- Badi, S. A.; Bruno, P. S.; Moshiri, A.; Tarashi, S.; Siadat, S.; Masotti, A. Small RNAs in outer membrane vesicles and their function in host–microbe interactions. Cellular Microbiology 2020, 22(7), e13277. [Google Scholar] [CrossRef]
- Díaz-Garrido, N.; Bonnin, S.; Riera, M.; Giménez, R.; Badia, J.; Baldomà, L. Transcriptomic microRNA profiling of dendritic cells in response to gut microbiota-secreted vesicles. Cells 2020, 9(6), 1534. [Google Scholar] [CrossRef]
- Díaz-Garrido, N.; Fábrega, M. J.; Vera, R.; Giménez, R.; Badia, J.; Baldomà, L. Membrane vesicles from the probiotic Nissle 1917 and gut resident Escherichia coli strains distinctly modulate human dendritic cells and subsequent T cell responses. Journal of Functional Foods 2019, 61, 103495. [Google Scholar] [CrossRef]
- Margutti, P.; D’Ambrosio, A.; Zamboni, S. Microbiota-derived extracellular vesicles as emerging players in gut homeostasis and disease. International Journal of Molecular Sciences 2024, 25(16), 8722. [Google Scholar] [CrossRef]
- Tian, C.; Gao, X.; Wang, Y.; Tian, Z. Emerging role of bacterial outer membrane vesicles in gastrointestinal diseases. Gut Pathogens 2023, 15, 51. [Google Scholar] [CrossRef]
- Jung, H.; Kim, J. S.; Lee, K. H.; Tizaoui, K.; Terrazzino, S.; Cargnin, S.; Kronbichler, A. Roles of microRNAs in inflammatory bowel disease. International Journal of Biological Sciences 2021, 17(8), 2112–2123. [Google Scholar] [CrossRef]
- Mishra, P.; Singh, D.; Ganju, L.; Kumar, B. MicroRNA in gastrointestinal cell signalling. Inflammopharmacology 2018, 26(1), 1–14. [Google Scholar] [CrossRef]
- Gabriel, K. R. The biplot graphic display of matrices with application to principal component analysis. Biometrika 1971, 58(3), 453–467. [Google Scholar] [CrossRef]
- Jolliffe, I. T.; Cadima, J. Principal component analysis: A review and recent developments. Philosophical Transactions of the Royal Society A 2016, 374(2065), 20150202. [Google Scholar] [CrossRef]
- Lever, J.; Krzywinski, M.; Altman, N. Principal component analysis. Nature Methods 2017, 14, 641–642. [Google Scholar] [CrossRef]
- Eisen, M. B.; Spellman, P. T.; Brown, P. O.; Botstein, D. Cluster analysis and display of genome-wide expression patterns. Proceedings of the National Academy of Sciences of the United States of America 1998, 95(25), 14863–14868. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32(18), 2847–2849. [Google Scholar] [CrossRef] [PubMed]
- Murtagh, F.; Legendre, P. Ward’s hierarchical agglomerative clustering method: Which algorithms implement Ward’s criterion? Journal of Classification 2014, 31, 274–295. [Google Scholar] [CrossRef]
- Olson, C. L. Comparative robustness of six tests in multivariate analysis of variance. Journal of the American Statistical Association 1974, 69(348), 894–908. [Google Scholar] [CrossRef]
- Van Aelst, S.; Willems, G. Robust and efficient one-way MANOVA tests. Journal of the American Statistical Association 2011, 106(494), 706–718. [Google Scholar] [CrossRef]
- Nakagawa, S.; Cuthill, I. C. Effect size, confidence interval and statistical significance: A practical guide for biologists. Biological Reviews 2007, 82(4), 591–605. [Google Scholar] [CrossRef]
- Kvålseth, T. O. Cautionary note about R². The American Statistician 1985, 39(4), 279–285. [Google Scholar] [CrossRef]
- Solé, M.; Giménez, R.; Cañas, M. A.; Badia, J.; Baldomà, L. Extracellular vesicles and soluble factors secreted by Escherichia coli Nissle 1917 and ECOR63 protect against enteropathogenic Escherichia coli-induced intestinal epithelial barrier dysfunction. BMC Microbiology 2019, 19, 166. [Google Scholar] [CrossRef]
- Riera, M. Efecto de las vesículas de membrana externa de cepas de Escherichia coli comensales y probióticas en la activación de células dendríticas a través del epitelio intestinal. In Universidad de Barcelona; 2019. [Google Scholar]
- Hu, R.; Lin, H.; Li, J.; Wang, W.; Wang, C.; Wang, Y.; Sun, X. Probiotic Escherichia coli Nissle 1917-derived outer membrane vesicles enhance immunomodulation and antimicrobial activity in RAW264.7 macrophages. BMC Microbiology 2020, 20, 268. [Google Scholar] [CrossRef]
- Ma, D.; Zhang, Y.; Zhang, J.; Shi, J.; Gao, S.; Long, F.; Wang, X.; Pu, X.; Sun, J.; Liang, S.; Cannon, R.; Boas-Villas, S.; Han, T-L. embrane vesicle–host cell interactions. Microbiology Spectrum 2019, 7(1), GPP3-0001-2018. [Google Scholar] [CrossRef]
- Lowry, O. H.; Rosebrough, N. J.; Farr, A. L.; Randall, R. J. Protein measurement with the Folin phenol reagent. Journal of Biological Chemistry 1951, 193(1), 265–275. [Google Scholar] [CrossRef]
Figure 1.
Functional roles of the intestinal microbiota in host homeostasis. Conceptual overview summarizing protective functions (pathogen displacement and antimicrobial factor production), structural functions (barrier fortification, IgA induction, tight-junction strengthening), and metabolic functions (vitamin synthesis, SCFA production, and energy salvage) that collectively shape epithelial–immune equilibrium.
Figure 1.
Functional roles of the intestinal microbiota in host homeostasis. Conceptual overview summarizing protective functions (pathogen displacement and antimicrobial factor production), structural functions (barrier fortification, IgA induction, tight-junction strengthening), and metabolic functions (vitamin synthesis, SCFA production, and energy salvage) that collectively shape epithelial–immune equilibrium.
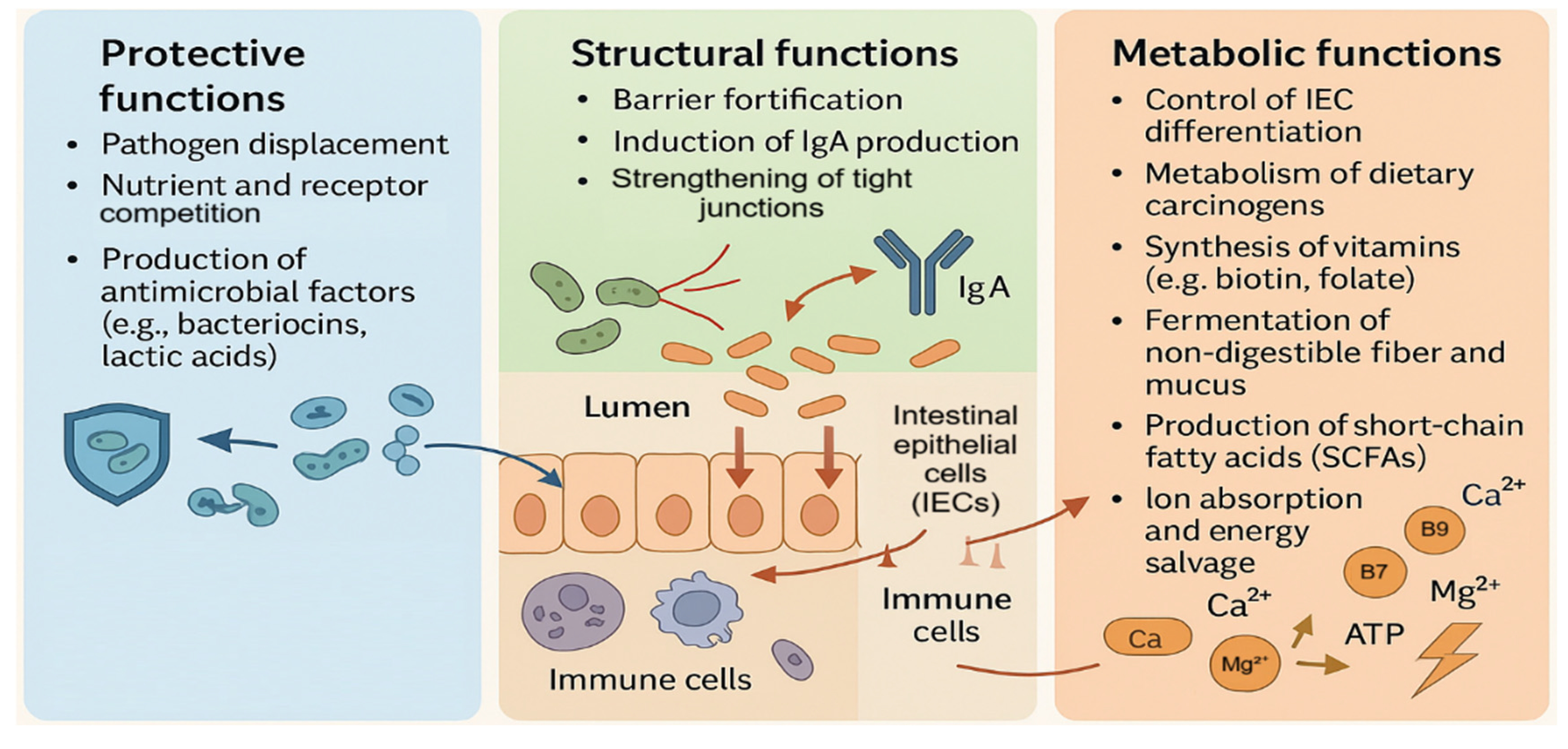
Figure 2.
“Signal 0” immune sensing links PAMPs/DAMPs to PRR activation and autophagy-associated immune programs. Diagram integrating microbial and damage signals (PAMPs/DAMPs), recognition by PRRs (e.g., TLRs, NLRs, RLRs, RAGE), downstream autophagy pathways (non-selective vs. selective), and the resulting effects on innate and adaptive immune responses (“immunophagy”).
Figure 2.
“Signal 0” immune sensing links PAMPs/DAMPs to PRR activation and autophagy-associated immune programs. Diagram integrating microbial and damage signals (PAMPs/DAMPs), recognition by PRRs (e.g., TLRs, NLRs, RLRs, RAGE), downstream autophagy pathways (non-selective vs. selective), and the resulting effects on innate and adaptive immune responses (“immunophagy”).
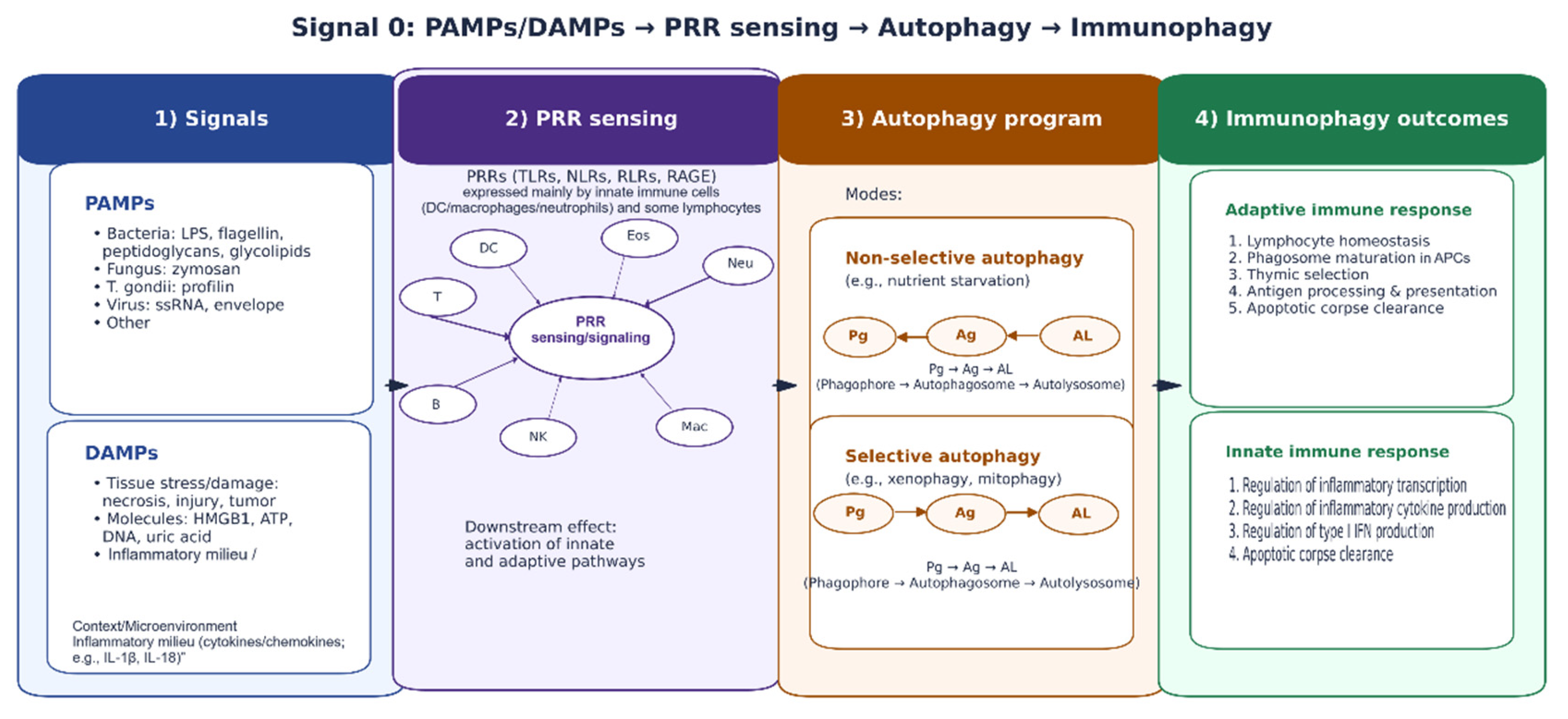
Figure 3.
Working model of probiotic/commensal E. coli traits converging on OMV-mediated host modulation. Schematic connecting strain attributes (iron uptake, adhesion/colonization, microcins) to OMV cargo delivery (LPS, outer membrane proteins, RNAs/small RNAs) and downstream outcomes in intestinal epithelial cells and dendritic cells, including tight-junction reinforcement and polarization toward inflammatory or regulatory programs shaped by cytokines and miRNA modulation.
Figure 3.
Working model of probiotic/commensal E. coli traits converging on OMV-mediated host modulation. Schematic connecting strain attributes (iron uptake, adhesion/colonization, microcins) to OMV cargo delivery (LPS, outer membrane proteins, RNAs/small RNAs) and downstream outcomes in intestinal epithelial cells and dendritic cells, including tight-junction reinforcement and polarization toward inflammatory or regulatory programs shaped by cytokines and miRNA modulation.
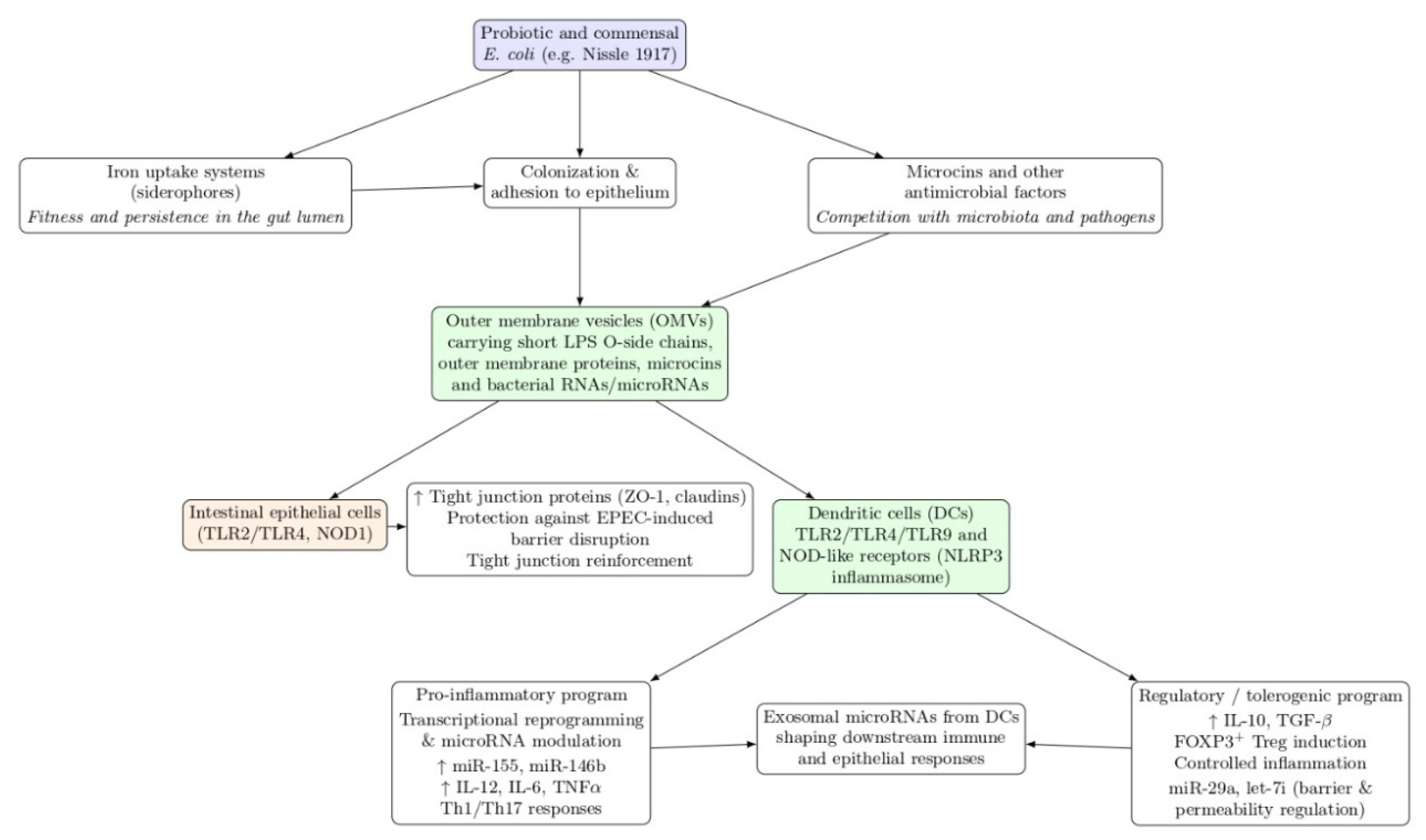
Figure 4.
Cell-type-resolved pathways of OMV interaction at the intestinal interface. Summary of OMV-driven signaling in macrophages (NF-κB/MAPK activation and inflammasome-linked inflammation), dendritic cells (transcriptional/miRNA reprogramming, antigen presentation, and tolerance vs. effector skewing), and epithelial cells (PRR signaling, antimicrobial programs, mucus, tight-junction reinforcement, and pyroptosis-related outcomes), highlighting how vesicle effects depend on the responding cell compartment.
Figure 4.
Cell-type-resolved pathways of OMV interaction at the intestinal interface. Summary of OMV-driven signaling in macrophages (NF-κB/MAPK activation and inflammasome-linked inflammation), dendritic cells (transcriptional/miRNA reprogramming, antigen presentation, and tolerance vs. effector skewing), and epithelial cells (PRR signaling, antimicrobial programs, mucus, tight-junction reinforcement, and pyroptosis-related outcomes), highlighting how vesicle effects depend on the responding cell compartment.
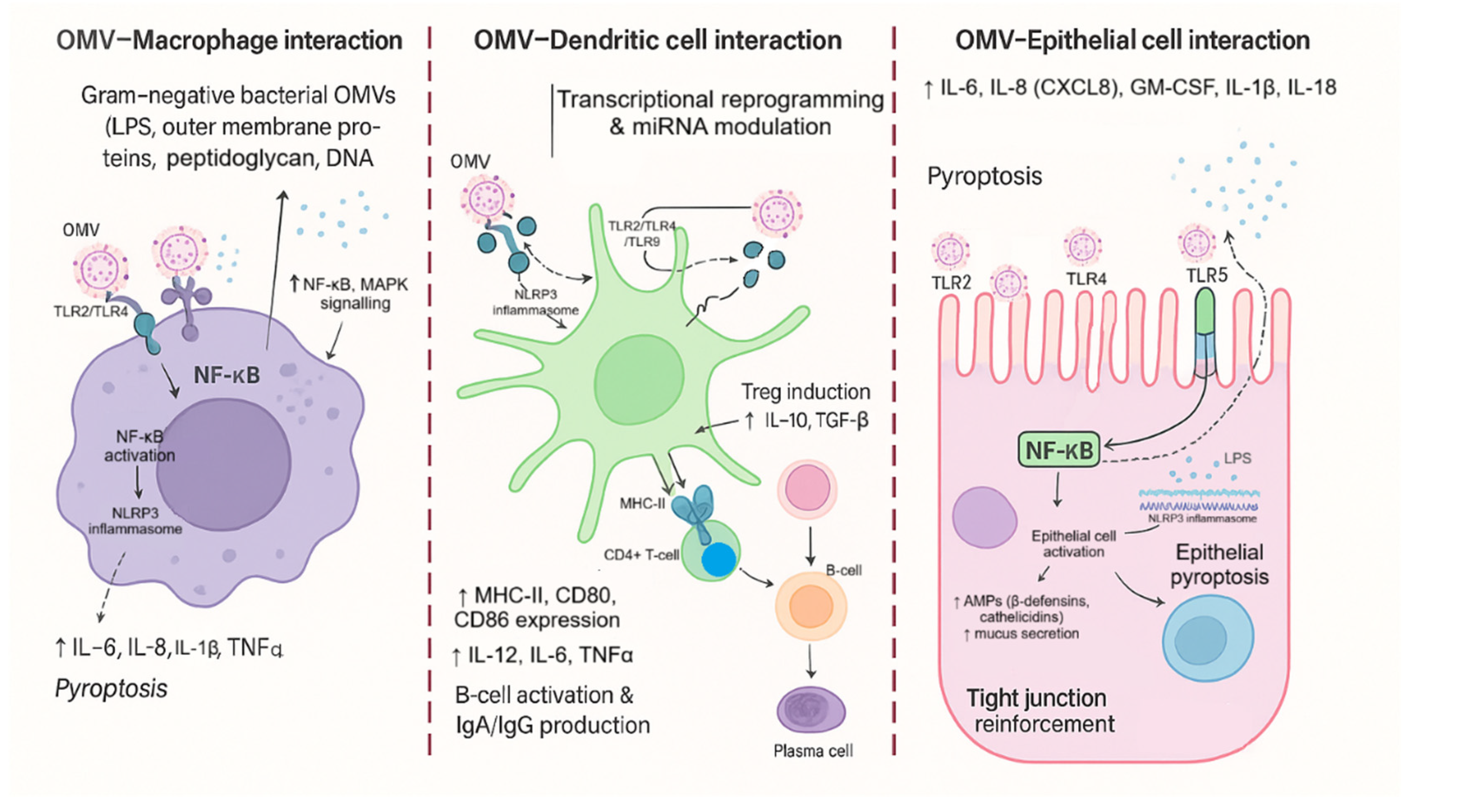
Figure 5.
Cryo-TEM images of OMVs isolated from different strains. (A) EcN OMVs, (B) ECOR12 OMVs, (C) ECOR63 OMVs. Vesicles are indicated by arrows.
Figure 5.
Cryo-TEM images of OMVs isolated from different strains. (A) EcN OMVs, (B) ECOR12 OMVs, (C) ECOR63 OMVs. Vesicles are indicated by arrows.
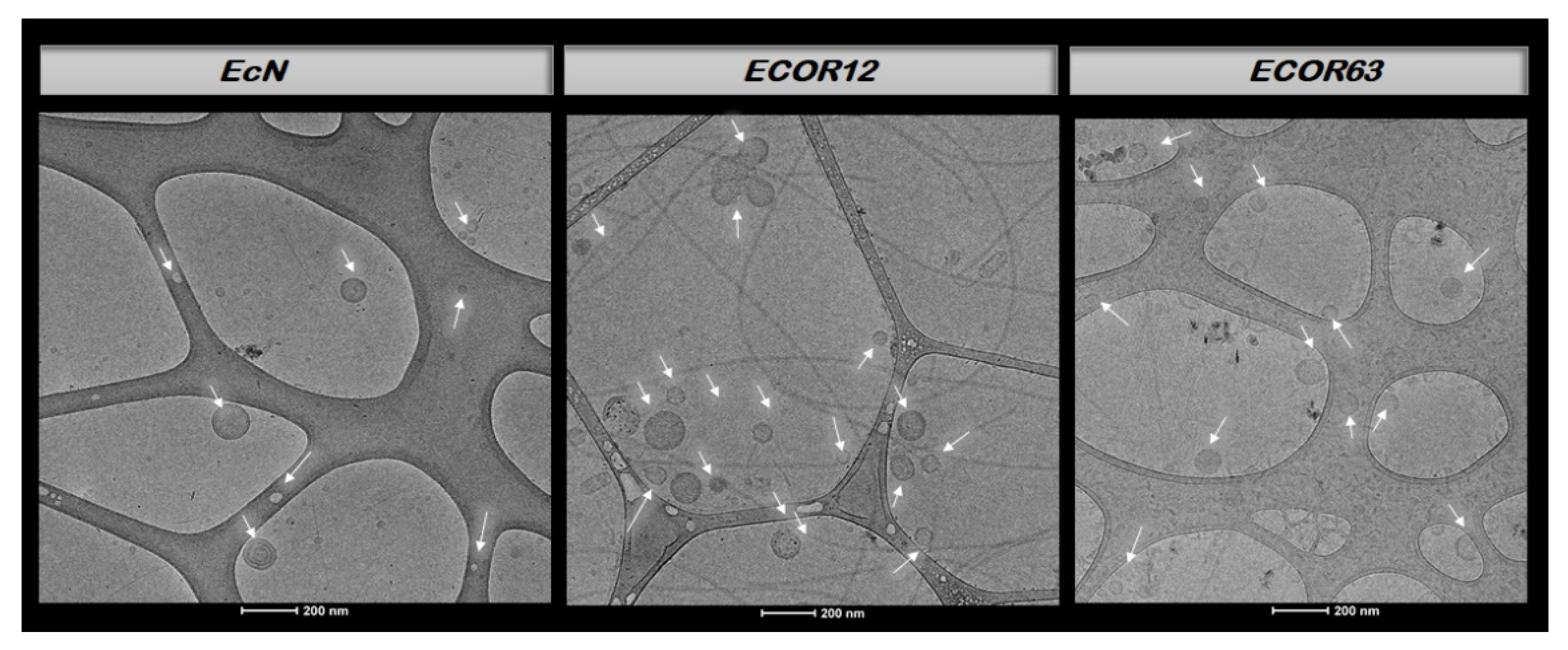
Figure 6.
Electrophoretic separation of proteins present in OMVs. OMVs (20 µL protein per strain) were separated on a 10% acrylamide gel and visualized by Coomassie Blue staining. The marker lane indicates molecular weight, with band masses labeled on the side.
Figure 6.
Electrophoretic separation of proteins present in OMVs. OMVs (20 µL protein per strain) were separated on a 10% acrylamide gel and visualized by Coomassie Blue staining. The marker lane indicates molecular weight, with band masses labeled on the side.
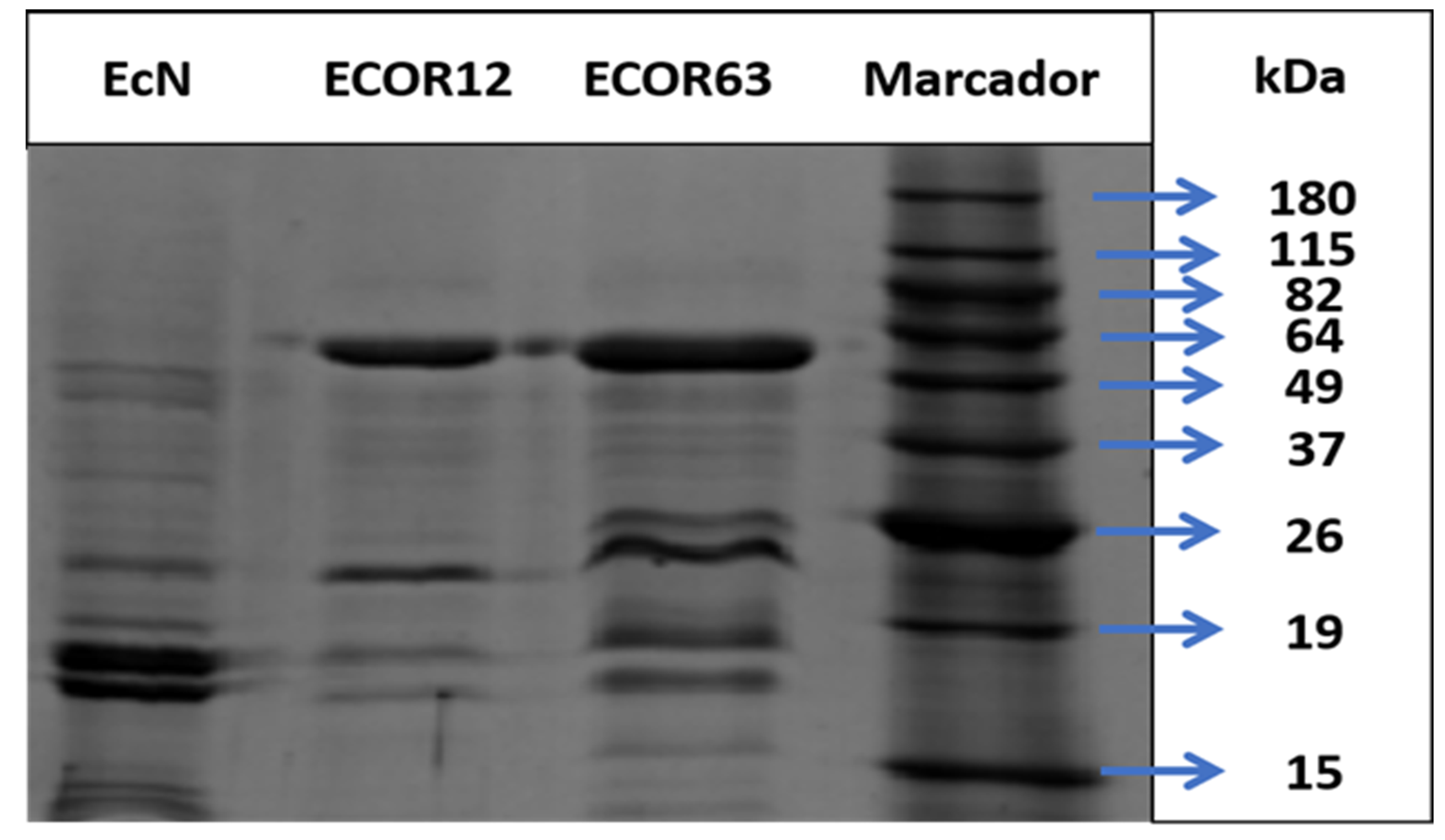
Figure 7.
ELISA quantification of IL-6, IL-10, and TNF-α (pg/mL) released by DCs stimulated with OMVs from commensal and probiotic strains.
Figure 7.
ELISA quantification of IL-6, IL-10, and TNF-α (pg/mL) released by DCs stimulated with OMVs from commensal and probiotic strains.
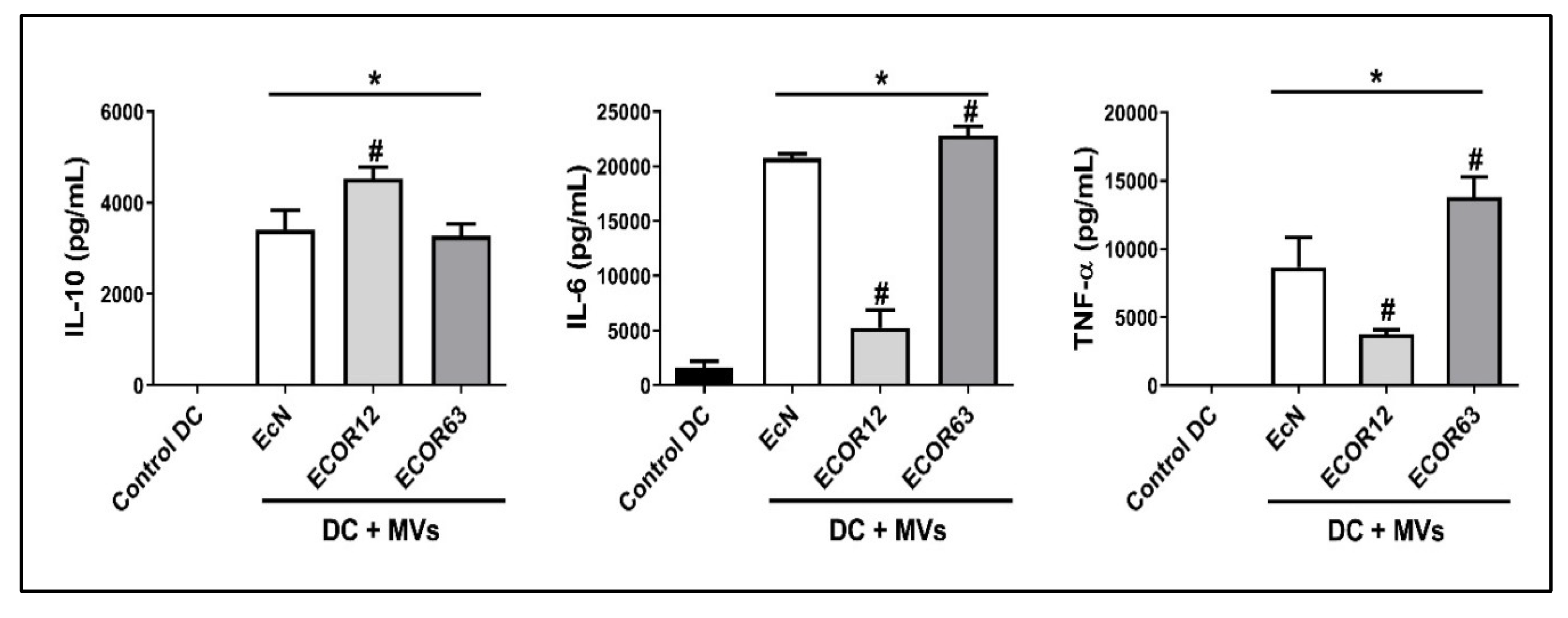
Figure 8.
Flow cytometry assessment of DC maturation markers. Three surface markers are shown (CD14-FITC, CD209-PE, CD83-APC) in DCs treated with 10 µg/mL OMVs from EcN and ECOR12; untreated iDCs were used as controls.
Figure 8.
Flow cytometry assessment of DC maturation markers. Three surface markers are shown (CD14-FITC, CD209-PE, CD83-APC) in DCs treated with 10 µg/mL OMVs from EcN and ECOR12; untreated iDCs were used as controls.
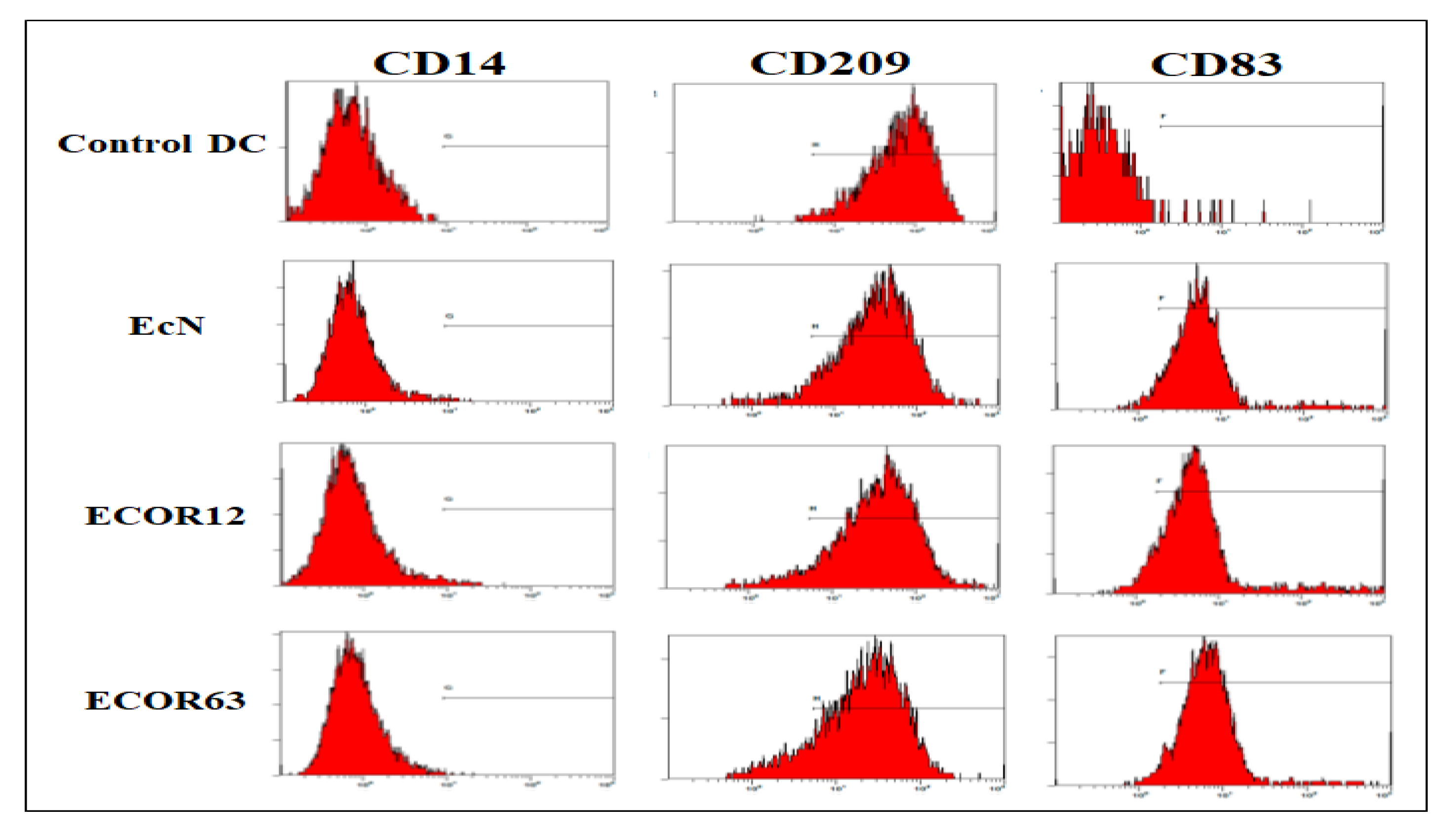
Figure 9.
Expression of miR-155-5p and let-7i-3p in DCs stimulated with EcN and ECOR12 OMVs.
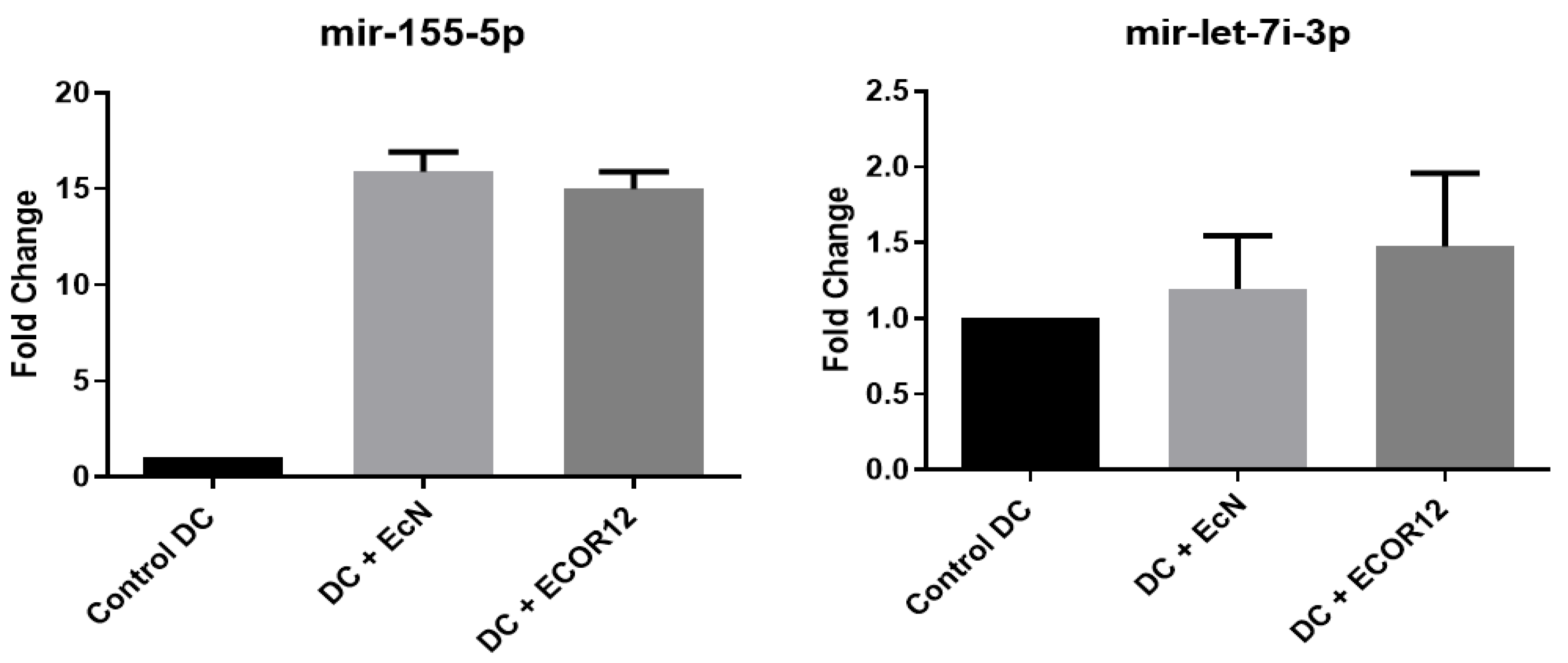
Figure 10.
Expression of miR-146b-5p and miR-29a-5p in DCs stimulated with EcN and ECOR12 OMVs.
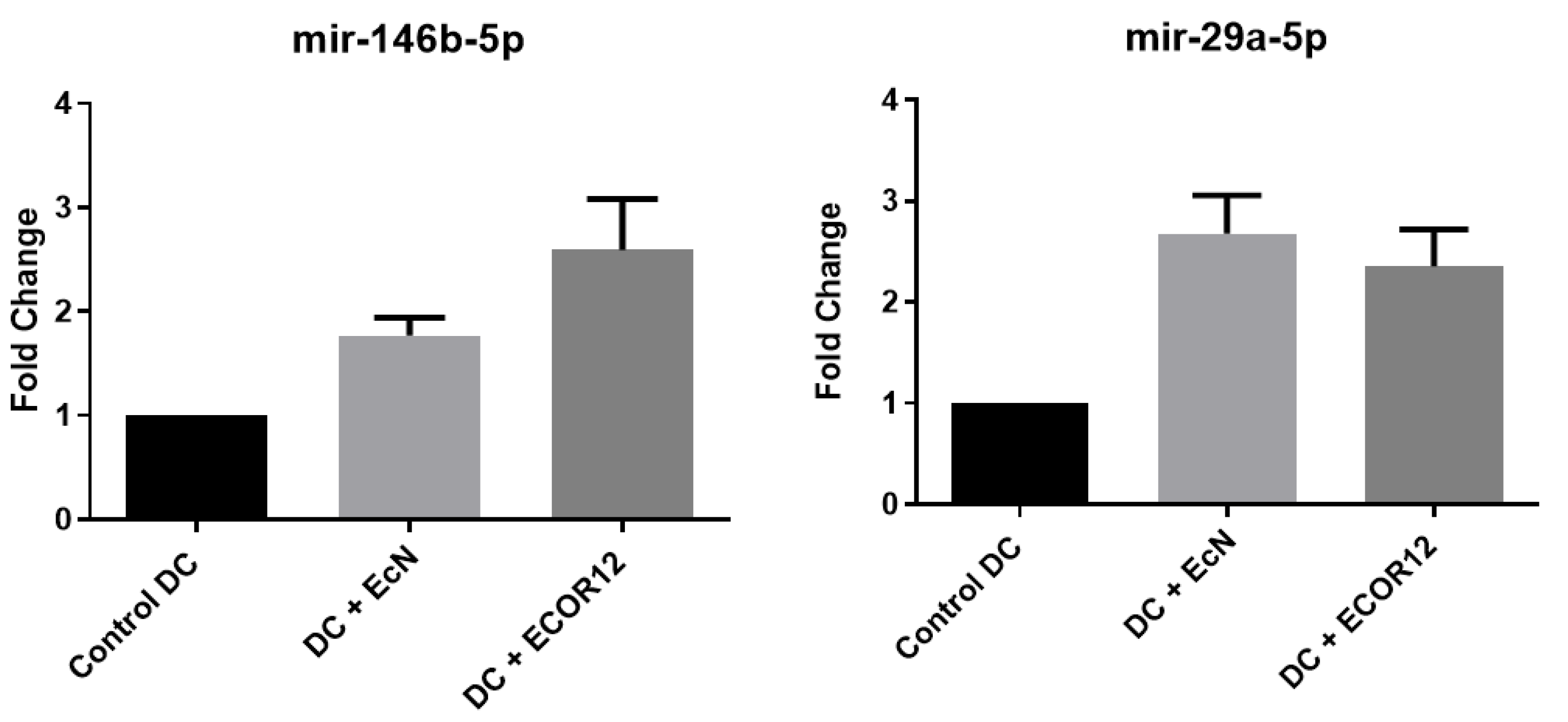
Figure 11.
PCA biplot with k-means clustering (k = 3).PCA of cytokines (IL-6, IL-10, TNF-α) and miRNAs (log2 miR-155-5p, let-7i-3p, miR-146b-5p, miR-29a-5p). Points are samples (n = 60), shapes indicate strain (Control, EcN, ECOR12), and colors represent k-means clusters (k = 3). Arrows correspond to variable loadings (vectors). Axis labels report explained variance (PC1 = 75.0%, PC2 = 21.2%).
Figure 11.
PCA biplot with k-means clustering (k = 3).PCA of cytokines (IL-6, IL-10, TNF-α) and miRNAs (log2 miR-155-5p, let-7i-3p, miR-146b-5p, miR-29a-5p). Points are samples (n = 60), shapes indicate strain (Control, EcN, ECOR12), and colors represent k-means clusters (k = 3). Arrows correspond to variable loadings (vectors). Axis labels report explained variance (PC1 = 75.0%, PC2 = 21.2%).
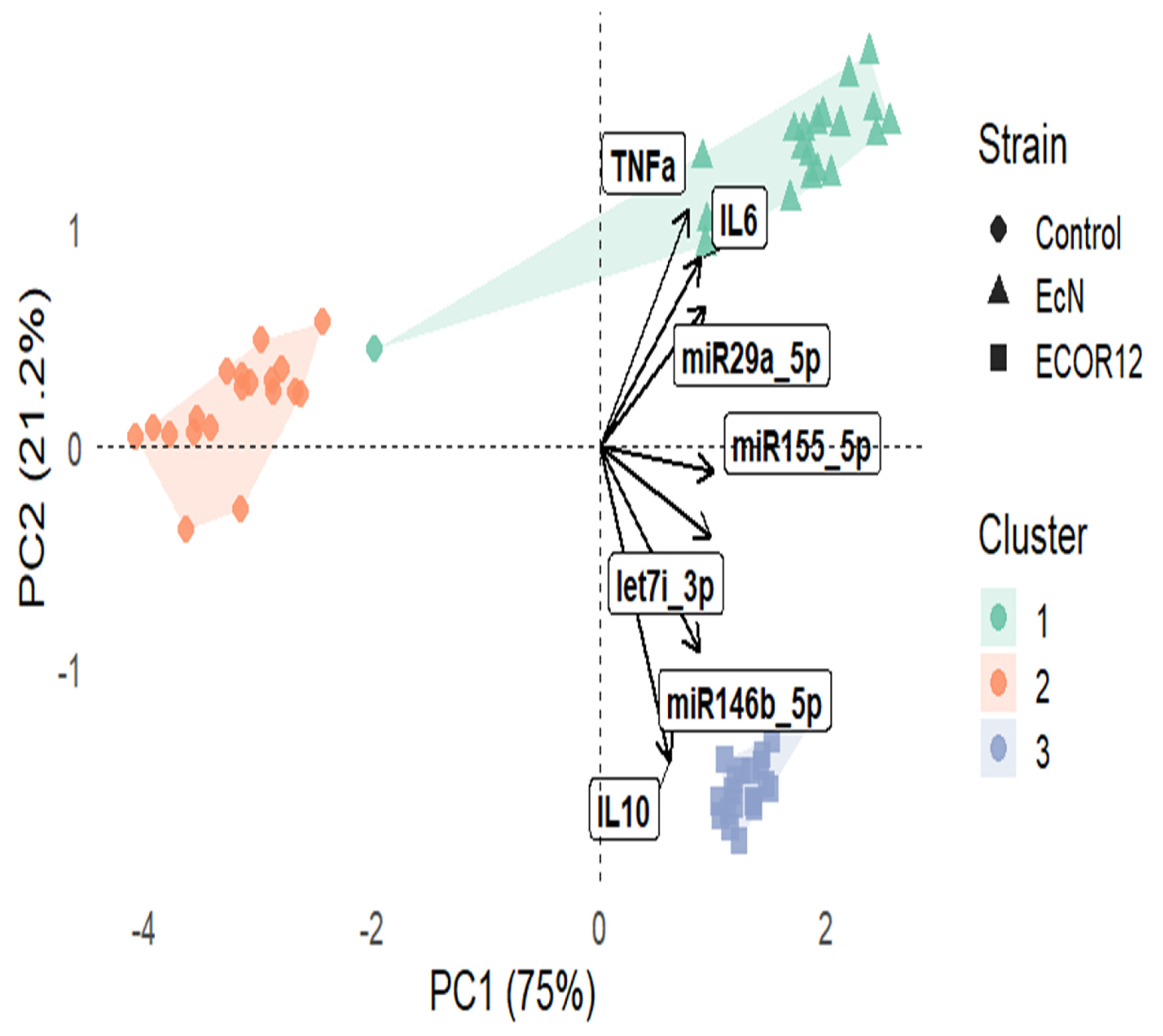
Figure 12.
Hierarchical clustering heatmap (strain-annotated). Heatmap of row-scaled biomarker levels (z-scores) for cytokines (IL-6, IL-10, TNF-α) and miRNAs (log2 miR-155-5p, let-7i-3p, miR-146b-5p, miR-29a-5p). Rows are samples (strain_rep), columns are variables. Both rows and columns were hierarchically clustered using complete linkage. The left annotation bar indicates strain (Control, EcN, ECOR12). Color scale denotes scaled value (blue = lower than mean; red = higher than mean).
Figure 12.
Hierarchical clustering heatmap (strain-annotated). Heatmap of row-scaled biomarker levels (z-scores) for cytokines (IL-6, IL-10, TNF-α) and miRNAs (log2 miR-155-5p, let-7i-3p, miR-146b-5p, miR-29a-5p). Rows are samples (strain_rep), columns are variables. Both rows and columns were hierarchically clustered using complete linkage. The left annotation bar indicates strain (Control, EcN, ECOR12). Color scale denotes scaled value (blue = lower than mean; red = higher than mean).
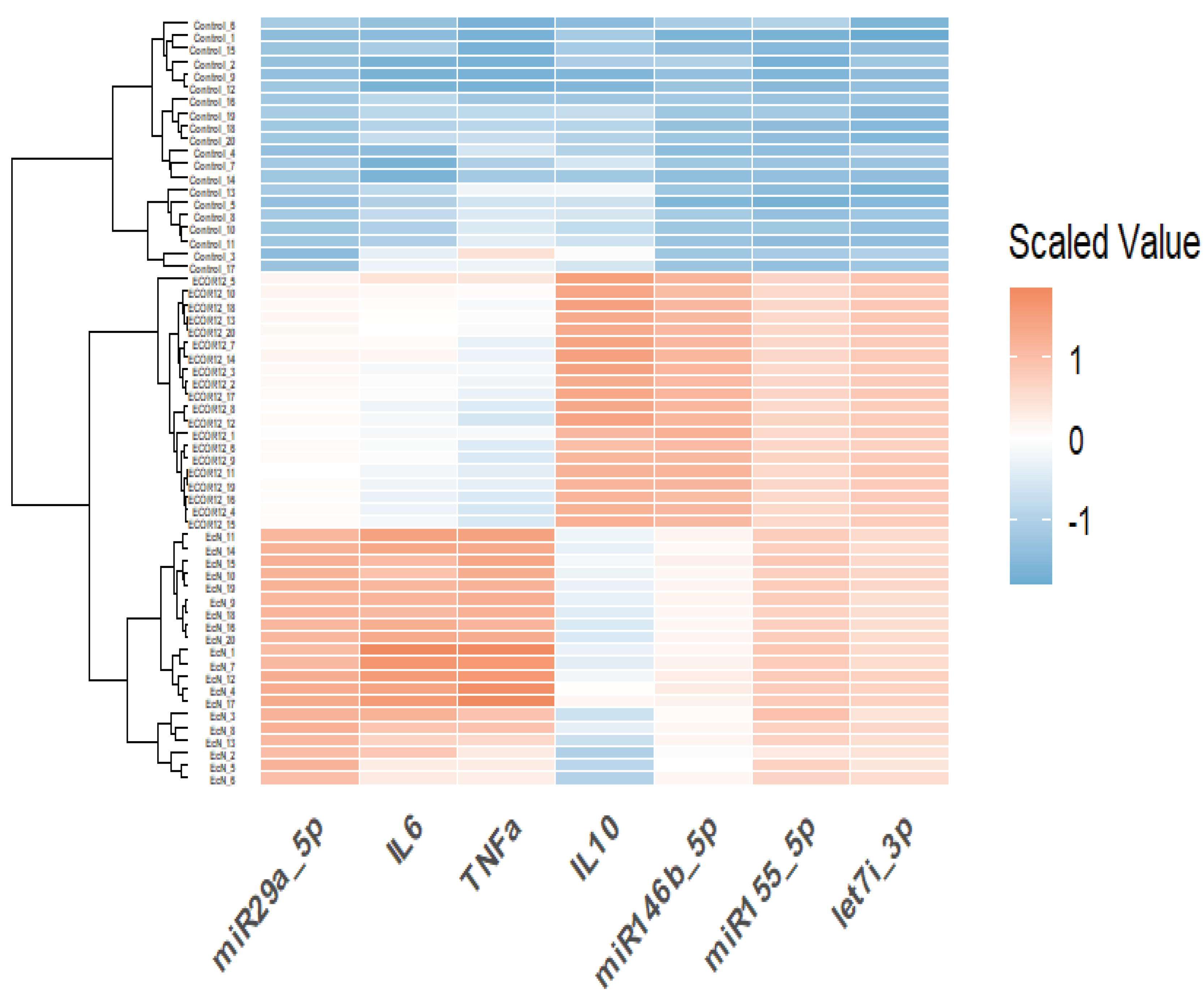
Table 1.
Experimental groups and exposure conditions for dendritic-cell stimulation with OMVs.
| Factor | Levels / description |
| Cell model | Human monocyte-derived dendritic cells (Mo-DCs) |
| Exposure groups | (i) Unstimulated iDC control; (ii) iDC + EcN OMVs; (iii) iDC + ECOR12 OMVs |
| OMV dose and duration | 10 µg/mL OMVs for 24 h |
| Biological replication | Independent experiments with technical processing in triplicate (as applicable) |
Note. Mo-DC differentiation and OMV stimulation conditions followed the laboratory workflow described in the study documentation.
Table 2.
Variables evaluated and measurement approaches.
| Domain | Variables | Measurement / output |
| DC phenotype | CD14, CD83, CD209 | Flow cytometry (fluorescent antibody labeling; % positive and/or intensity) |
| Secreted mediators | IL-6, IL-10, TNF-α | ELISA on culture supernatants (pg/mL) |
| Post-transcriptional regulation | miRNAs (panel assayed by RT-qPCR; normalized to internal controls/housekeeping) | Relative expression using 2^-ΔΔCt; log-transformed for multivariate analyses when required |
| OMV input control | Total OMV protein | Lowry protein assay (mg/mL equivalent), used to standardize dosing |
Note. The specific antibodies, cytokines, and miRNA RT-qPCR workflow correspond to the study’s established procedures for DC maturation and miRNA validation after OMV stimulation.
Table 3.
Equipment and acquisition settings used to generate cytokine, miRNA, and phenotyping data.
| Data type | Instrument / platform | Key settings / notes |
| Flow cytometry | Flow cytometer (CCiT-UB cytometry unit) | Antibodies: anti-CD14-FITC, anti-CD83-PE, anti-CD209-APC |
| Cytokines (ELISA) | ELISA kits (BD Biosciences) + microplate spectrophotometry | Readout per manufacturer instructions (typical absorbance at 450 nm) |
| RNA QC | NanoDrop® ND-1000 | A260/A280 ratio used to assess purity |
| miRNA qPCR | StepOnePlus™ Real-Time PCR System | 95 °C 2 min; 40 cycles of 95 °C 10 s and 56 °C 60 s |
| OMV isolation | Refrigerated centrifuges + ultracentrifugation | 10,000 × g (30 min, 4 °C); ultracentrifugation 45,000 rpm (1 h, 4 °C) |
| OMV imaging | Cryo-TEM (CCiT-UB microscopy unit) | Cryogenic imaging (~−170 to −175 °C), 200 kV |
Note. Instrument access and assay kits correspond to the laboratory workflow and facility resources described in the study documentation.
Table 4.
Statistical analysis workflow and inference criteria.
| Step | Method | Rationale / reference support |
| Data screening | Outlier inspection; distribution checks; transformation (when needed) | Ensures assumptions are evaluated before parametric inference |
| Global multivariate test | MANOVA with Pillai’s trace (α = 0.05) | Pillai’s trace is robust among MANOVA criteria [66]; robust MANOVA alternatives considered if as-sumptions are violated [67]. |
| Univariate follow-up | One- or two-way ANOVA + Tukey HSD post hoc | Standard approach for multiple-group contrasts; used in prior workflow |
| Dimension reduction | PCA + biplot visualization (centered/scaled features) | PCA summarizes correlated immune variables and biplots clarify loadings [60,61,62]. Clustering approaches are widely used to explore structure in multivariate bio-logical datasets. |
| Pattern discovery | Heatmaps + hierarchical clustering (distance-based); k-means as exploratory option | Hierarchical clustering and heatmaps reveal response modules [63,64,65]; k-means used as complementary clustering tool. |
| Reporting | Mean ± SD; effect sizes with CIs when applicable | Effect-size reporting supports biological interpretation beyond p-values [68]. |
Note. Inferential outcomes (p-values, group separations, cluster structures) are reported in the Results section; Table 4 specifies the a priori decision rules used to evaluate OMV effects.
Table 5.
Quantification of isolated OMVs by protein measurement (Lowry method).
| Strain | Concentration (mg/mL) | Sample volume (µL) | Loading buffer (µL) | H₂O (µL) | Final volume (µL) |
| EcN | 2.898 | 3.45 | 5.00 | 11.55 | 20 |
| ECOR12 | 2.744 | 3.64 | 5.00 | 11.36 | 20 |
| ECOR63 | 1.518 | 6.58 | 5.00 | 8.42 | 20 |
Note. Protein concentration was determined using the Lowry assay [75]. Reported volumes correspond to preparation of standardized 20 µL mixtures for downstream handling.
Table 6.
MANOVA summary (Pillai trace) for integrated biomarkers and biomarker families. Multivariate tests evaluating the effect of strain on (i) the combined cytokine+miRNA set, (ii) cytokines only, and (iii) miRNAs only. Pillai’s trace is reported as a robust multivariate statistic.
Table 6.
MANOVA summary (Pillai trace) for integrated biomarkers and biomarker families. Multivariate tests evaluating the effect of strain on (i) the combined cytokine+miRNA set, (ii) cytokines only, and (iii) miRNAs only. Pillai’s trace is reported as a robust multivariate statistic.
| Response set | df (effect) | Pillai | Approx. F | num df | den df | p-value |
| Cytokines + miRNAs (7 vars) | 2 | 1.9882 | 1246.5 | 14 | 104 | < 2.2e-16 |
| Cytokines only (3 vars) | 2 | 1.8579 | 243.98 | 6 | 112 | < 2.2e-16 |
| miRNAs only (4 vars) | 2 | 1.9797 | 1339.2 | 8 | 110 | < 2.2e-16 |
Table 7.
Univariate ANOVAs (post-MANOVA) and variance explained (R²) by strain. Univariate follow-up tests for each biomarker with corresponding R² values (proportion of variance explained by strain).
Table 7.
Univariate ANOVAs (post-MANOVA) and variance explained (R²) by strain. Univariate follow-up tests for each biomarker with corresponding R² values (proportion of variance explained by strain).
| Biomarker | F (2,57) | p-value | R² |
| IL-6 | 206.58 | < 2.2e-16 | 0.879 |
| IL-10 | 283.40 | < 2.2e-16 | 0.909 |
| TNF-α | 102.94 | < 2.2e-16 | 0.783 |
| miR-155-5p (log2) | 1751.9 | < 2.2e-16 | 0.984 |
| let-7i-3p (log2) | 1864.8 | < 2.2e-16 | 0.985 |
| miR-146b-5p (log2) | 2836.5 | < 2.2e-16 | 0.990 |
| miR-29a-5p (log2) | 4220.8 | < 2.2e-16 | 0.993 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



