Submitted:
19 December 2025
Posted:
22 December 2025
You are already at the latest version
Abstract
Transposable elements (TEs) impact genes and genomes in different ways. In Drosophila, TEs play a critical role in establishing heterochromatic loci, such as centromeric and pericentromeric regions, and contribute to ectopic heterochromatin formation especially when found in a tandem repeat configuration. The Bari transposon family belongs to the Tc1/mariner superfamily. Bari1 is featured by a unique tandem repeat arrangement in the heterochromatin of D. melanogaster. Despite its conservation could suggest a structural role in both natural and laboratory populations, little is known about the role of the Bari1 repeat in the heterochromatin. In this paper, we have studied the impact of the Bari1 transposon clusters on chromatin. We obtained transgenic strains of D. melanogaster carrying arrays of either three or seven copies of the Bari1 transposon in a typical heterochromatic tandem repeat configuration. We performed qualitative and quantitative phenotypic assays to evaluate the impact of these arrays on the chromatin status. Besides, we performed Southern Blot Hybridization analyses to evaluate the stability of Bari1 repeats over multiple generations. Our results suggest that Bari1 arrays are very stable over time and can poorly act as heterochromatin seeding sites. Our findings expand the knowledge on the structural and functional impact of TEs, whose potentially neutral, beneficial, or detrimental effect cannot be envisioned without experimental validation.
Keywords:
1. Introduction
Transposable Elements and Heterochromatin
Bari1 Transposon
2. Materials and Methods
Plasmids Construction and Molecular Methods
Drosophila Genetics Methods
Fly Stocks Were Maintained at 25 °C on Standard Culturing Medium.
Southern Blot Hybridization
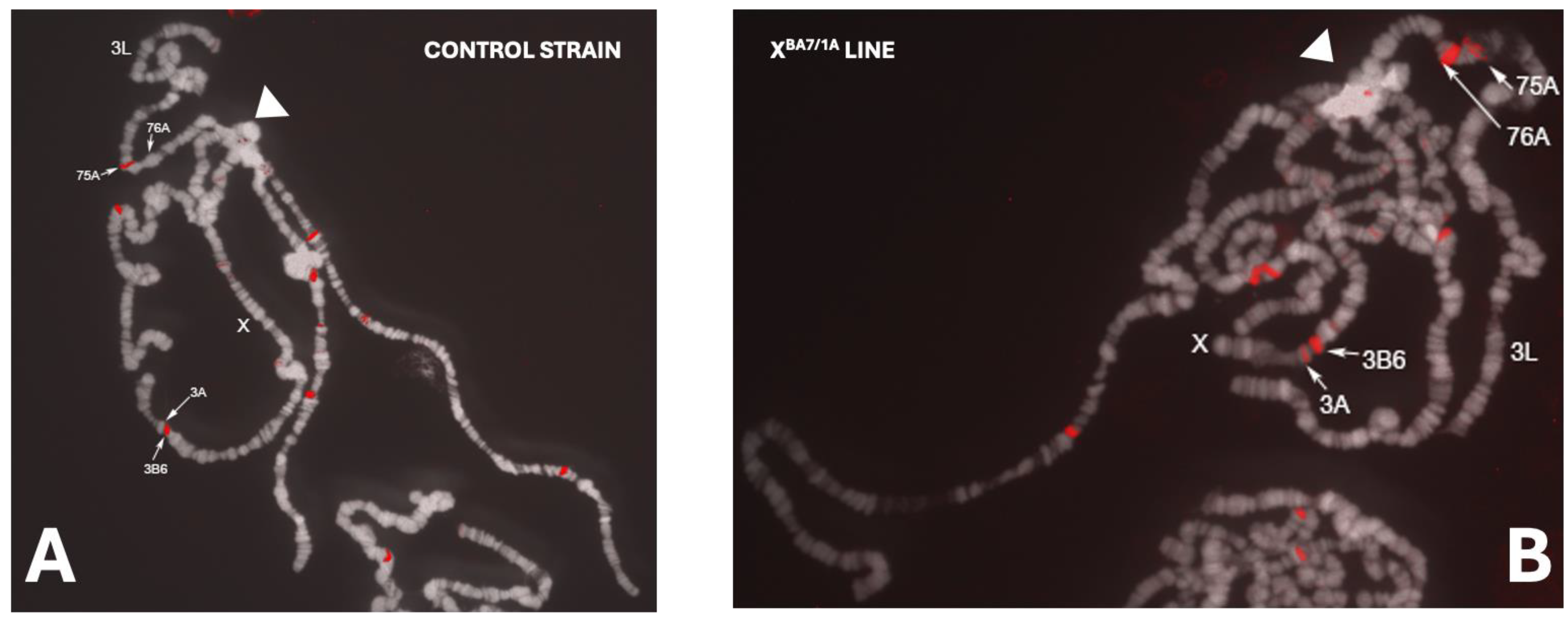
FISH (Fluorescence In Situ Hybridization) Experiments
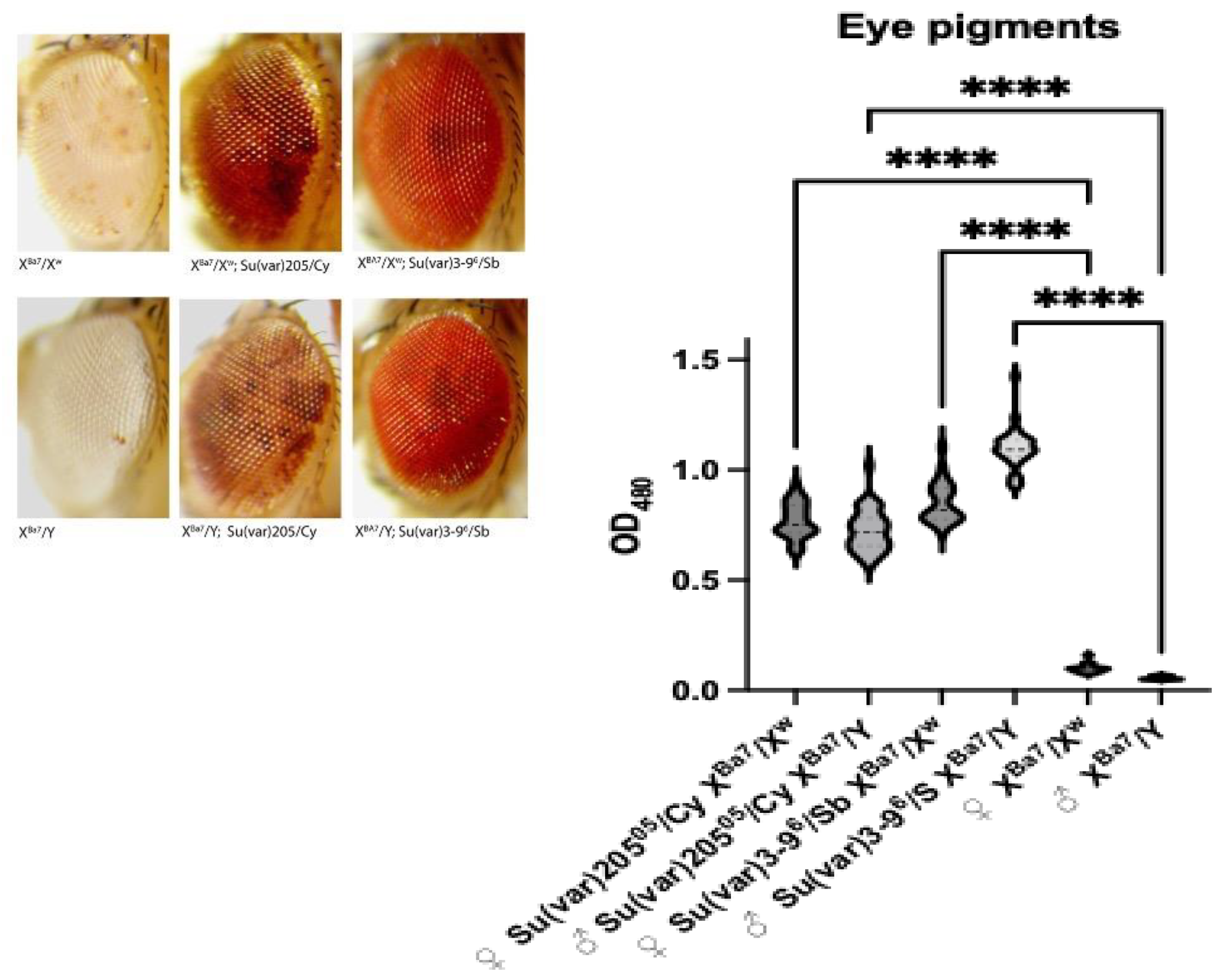
PEV (Position Effect Variegation) and Eye Pigment Quantification
3. Results
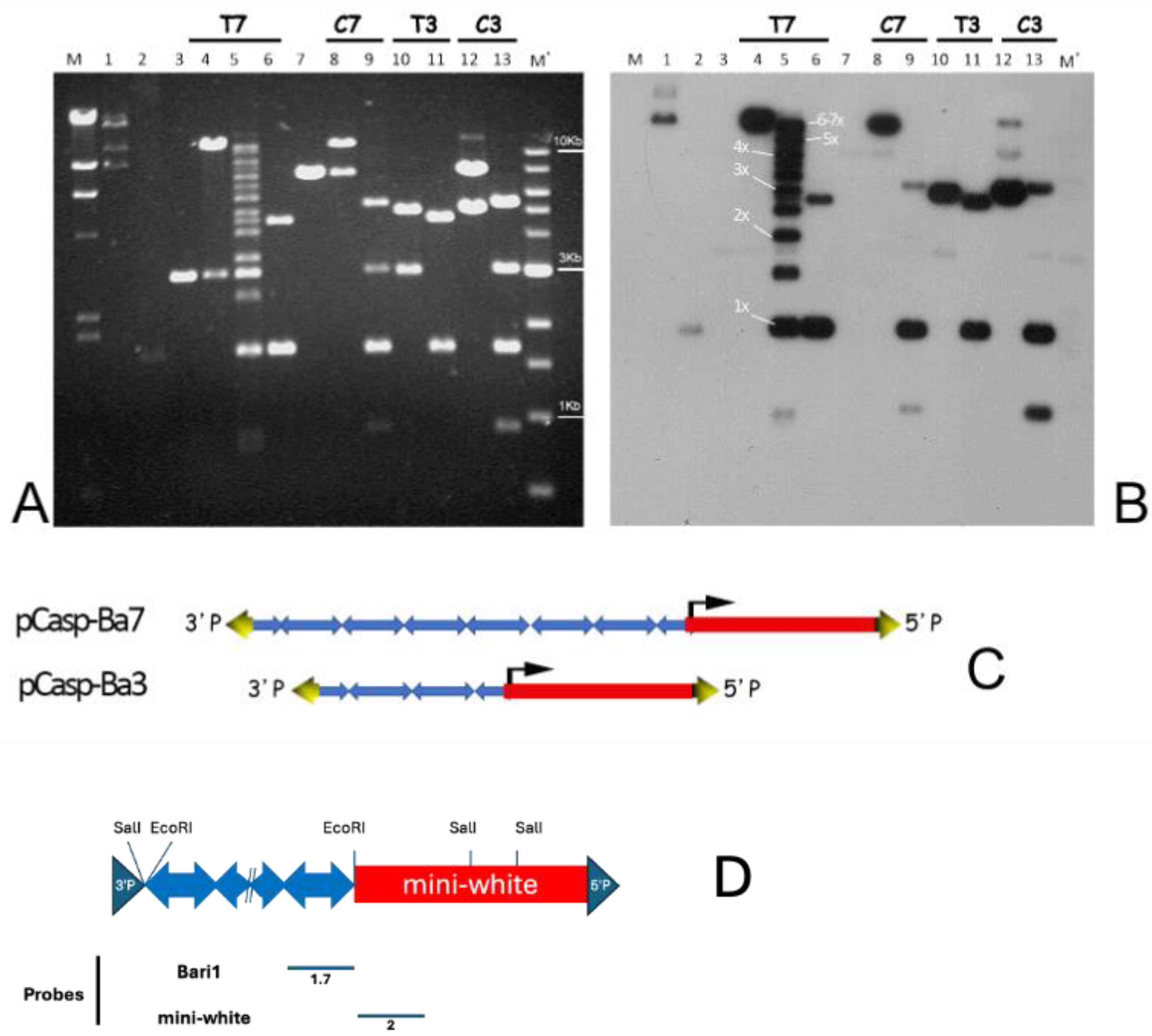
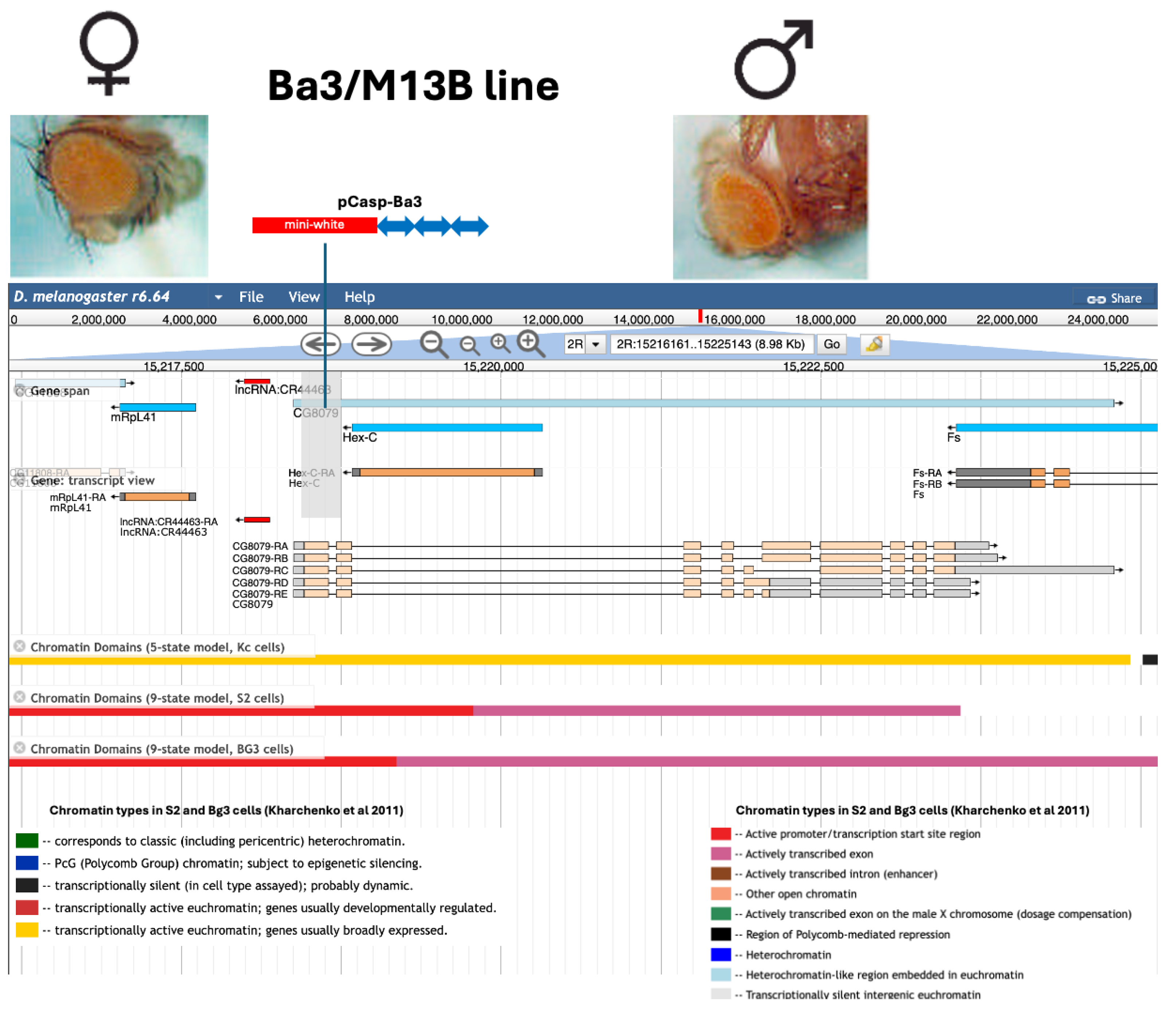
3.1. Strategy for Constructing Artificial Bari1 Arrays
3.2. Analysis of Two Transgenic Lines that Display Uneven Eye Pigmentation.
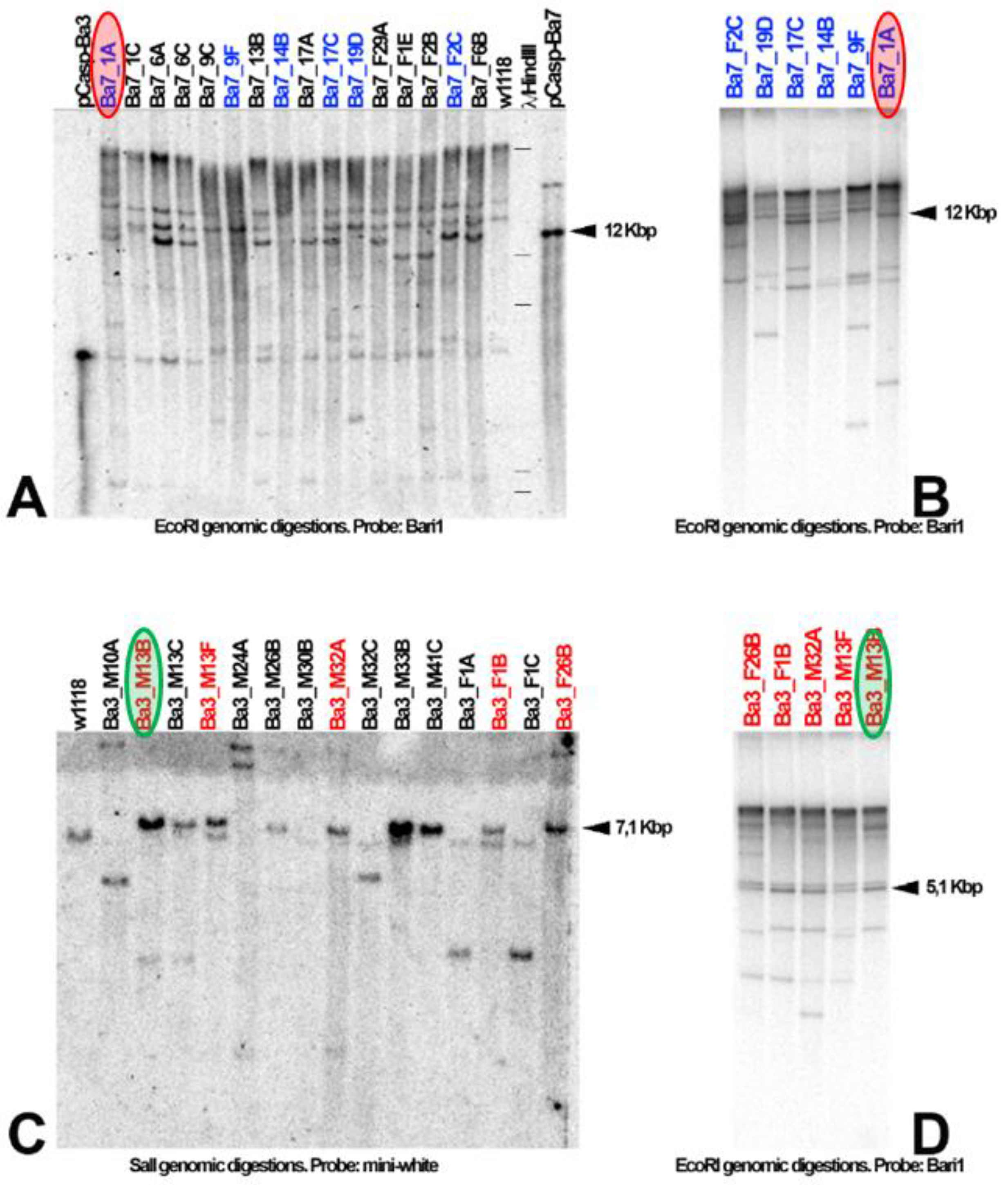
3.3. Bari1 Repeats Stability Over Time
4. Discussion
Bari1 Arrays do not Induce Heterochromatin Formation
Bari1 Arrays Are Stable over Time
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| PEV | Position Effect Variegation |
| piRNA | Piwi-interacting RNA |
| TE | Transposable Element |
| Rsp | Responder |
| iPCR | inverse PCR |
References
- Wells, J.N.; Feschotte, C. A Field Guide to Eukaryotic Transposable Elements. Annu Rev Genet 2020, 54, 539–561. [Google Scholar] [CrossRef]
- Kazazian, H.H., Jr. Mobile elements: drivers of genome evolution. Science 2004, 303, 1626–1632. [Google Scholar] [CrossRef]
- Bohne, A.; Brunet, F.; Galiana-Arnoux, D.; Schultheis, C.; Volff, J.N. Transposable elements as drivers of genomic and biological diversity in vertebrates. Chromosome Res 2008, 16, 203–215. [Google Scholar] [CrossRef]
- Bourque, G.; Burns, K.H.; Gehring, M.; Gorbunova, V.; Seluanov, A.; Hammell, M.; Imbeault, M.; Izsvák, Z.; Levin, H.L.; Macfarlan, T.S.; et al. Ten things you should know about transposable elements. Genome Biol 2018, 19, 199. [Google Scholar] [CrossRef]
- Sundaram, V.; Wysocka, J. Transposable elements as a potent source of diverse cis-regulatory sequences in mammalian genomes. Philos Trans R Soc Lond B Biol Sci 2020, 375, 20190347–20190347. [Google Scholar] [CrossRef] [PubMed]
- Marsano, R.M.; Dimitri, P. Constitutive Heterochromatin in Eukaryotic Genomes: A Mine of Transposable Elements. Cells 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Marsano, R.M.; Giordano, E.; Messina, G.; Dimitri, P. A New Portrait of Constitutive Heterochromatin: Lessons from Drosophila melanogaster. Trends Genet 2019, 35, 615–631. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Hou, Y.; Ebina, H.; Levin, H.L.; Voytas, D.F. Chromodomains direct integration of retrotransposons to heterochromatin. Genome Res 2008, 18, 359–369. [Google Scholar] [CrossRef]
- Kent, T.V.; Uzunović, J.; Wright, S.I. Coevolution between transposable elements and recombination. Philos Trans R Soc Lond B Biol Sci 2017, 372, 20160458. [Google Scholar] [CrossRef]
- Rebollo, R.; Karimi, M.M.; Bilenky, M.; Gagnier, L.; Miceli-Royer, K.; Zhang, Y.; Goyal, P.; Keane, T.M.; Jones, S.; Hirst, M.; et al. Retrotransposon-induced heterochromatin spreading in the mouse revealed by insertional polymorphisms. PLoS Genet 2011, 7, e1002301–e1002301. [Google Scholar] [CrossRef]
- Muller, H.J. Types of visible variations induced by X-rays inDrosophila. J Genet 1930, 22, 299–334. [Google Scholar] [CrossRef]
- Tartof, K.D.; Hobbs, C.; Jones, M. A structural basis for variegating position effects. Cell 1984, 37, 869–878. [Google Scholar] [CrossRef] [PubMed]
- Wallrath, L.L.; Elgin, S.C. Position effect variegation in Drosophila is associated with an altered chromatin structure. Genes Dev 1995, 9, 1263–1277. [Google Scholar] [CrossRef]
- Hazelrigg, T.; Levis, R.; Rubin, G.M. Transformation of white locus DNA in Drosophila: Dosage compensation, zeste interaction, and position effects. Cell 1984, 36, 469–481. [Google Scholar] [CrossRef] [PubMed]
- Kelsey, K.J.P.; Clark, A.G. Variation in Position Effect Variegation Within a Natural Population. Genetics 2017, 207, 1157–1166. [Google Scholar] [CrossRef]
- Wang, S.H.; Elgin, S.C.R. The impact of genetic background and cell lineage on the level and pattern of gene expression in position effect variegation. Epigenetics Chromatin 2019, 12, 70. [Google Scholar] [CrossRef] [PubMed]
- Schotta, G.; Ebert, A.; Dorn, R.; Reuter, G. Position-effect variegation and the genetic dissection of chromatin regulation in Drosophila. Semin Cell Dev Biol 2003, 14, 67–75. [Google Scholar] [CrossRef]
- Elgin, S.C.R.; Reuter, G. Position-Effect Variegation, Heterochromatin Formation, and Gene Silencing in Drosophila. Cold Spring Harb Perspect Biol 2013, 5. [Google Scholar] [CrossRef]
- Li, Y.; Kirschmann, D.A.; Wallrath, L.L. Does heterochromatin protein 1 always follow code? Proc Natl Acad Sci U S A 2002, 99 Suppl 4, 16462–16469. [Google Scholar] [CrossRef]
- Greil, F.; van der Kraan, I.; Delrow, J.; Smothers, J.F.; de Wit, E.; Bussemaker, H.J.; van Driel, R.; Henikoff, S.; van Steensel, B. Distinct HP1 and Su(var)3-9 complexes bind to sets of developmentally coexpressed genes depending on chromosomal location. Genes Dev 2003, 17, 2825–2838. [Google Scholar] [CrossRef]
- Reuter, G.; Wolff, I.; Friede, B. Functional properties of the heterochromatic sequences inducing wm4position-effect variegation in Drosophila melanogaster. Chromosoma 1985, 93, 132–139. [Google Scholar] [CrossRef]
- Howe, M.; Dimitri, P.; Berloco, M.; Wakimoto, B.T. Cis-effects of heterochromatin on heterochromatic and euchromatic gene activity in Drosophila melanogaster. Genetics 1995, 140, 1033–1045. [Google Scholar] [CrossRef]
- Dorer, D.R.; Henikoff, S. Expansions of transgene repeats cause heterochromatin formation and gene silencing in Drosophila. Cell 1994, 77, 993–1002. [Google Scholar] [CrossRef] [PubMed]
- Pirrotta, V. CHAPTER 22 - Vectors for P-Mediated Transformation in Drosophila. In Vectors, Rodriguez, R.L., Denhardt, D.T., Eds.; Butterworth-Heinemann: 1988; pp. 437-456.
- Sabl, J.F.; Henikoff, S. Copy Number and Orientation Determine the Susceptibility of a Gene to Silencing by Nearby Heterochromatin in Drosophila. Genetics 1996, 142, 447–458. [Google Scholar] [CrossRef] [PubMed]
- Riddle, N.C.; Shaffer, C.D.; Elgin, S.C.R. A lot about a little dot — lessons learned from Drosophila melanogaster chromosome 4. Biochemistry and Cell Biology 2009, 87, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.-L.; Haynes, K.; Simpson, C.L.; Lee, S.D.; Collins, L.; Wuller, J.; Eissenberg, J.C.; Elgin, S.C.R. cis-Acting Determinants of Heterochromatin Formation on Drosophila melanogaster Chromosome Four. Mol Cell Biol 2004, 24, 8210–8220. [Google Scholar] [CrossRef]
- Sentmanat, M.F.; Elgin, S.C. Ectopic assembly of heterochromatin in Drosophila melanogaster triggered by transposable elements. Proc Natl Acad Sci U S A 2012, 109, 14104–14109. [Google Scholar] [CrossRef]
- Ahmad, K.; Wooten, M.; Takushi, B.N.; Vidaurre, V.; Chen, X.; Henikoff, S. Cell-cycle-dependent repression of histone gene transcription by histone H4. bioRxiv 2025, 2024.2012.2023.630206. [Google Scholar] [CrossRef]
- Watase, G.J.; Nelson, J.O.; Yamashita, Y.M. Nonrandom sister chromatid segregation mediates rDNA copy number maintenance in Drosophila. Science Advances 2022, 8, eabo4443. [Google Scholar] [CrossRef]
- Gu, T.; Gracheva, E.; Lee, M.; Leung, W.; Bieser, S.; Nielsen, A.; Smiley, A.T.; Vuong, N.N.T.; Walther, M.; Reuter, G.; et al. Heterochromatin-based silencing of a foreign tandem repeat in Drosophila melanogaster shows unusual biochemistry and temperature sensitivity. bioRxiv 2025, 2025.2007.2031.667933. [Google Scholar] [CrossRef]
- Estécio, M.R.H.; Gallegos, J.; Dekmezian, M.; Lu, Y.; Liang, S.; Issa, J.-P.J. SINE retrotransposons cause epigenetic reprogramming of adjacent gene promoters. Mol Cancer Res 2012, 10, 1332–1342. [Google Scholar] [CrossRef]
- Lippman, Z.; Gendrel, A.-V.; Black, M.; Vaughn, M.W.; Dedhia, N.; Richard McCombie, W.; Lavine, K.; Mittal, V.; May, B.; Kasschau, K.D.; et al. Role of transposable elements in heterochromatin and epigenetic control. Nature 2004, 430, 471–476. [Google Scholar] [CrossRef]
- Evtushenko, E.V.; Levitsky, V.G.; Elisafenko, E.A.; Gunbin, K.V.; Belousov, A.I.; Šafář, J.; Doležel, J.; Vershinin, A.V. The expansion of heterochromatin blocks in rye reflects the co-amplification of tandem repeats and adjacent transposable elements. BMC Genomics 2016, 17, 337. [Google Scholar] [CrossRef]
- Zemach, A.; McDaniel, I.E.; Silva, P.; Zilberman, D. Genome-Wide Evolutionary Analysis of Eukaryotic DNA Methylation. Science 2010, 328, 916. [Google Scholar] [CrossRef]
- Jeon, J.; Choi, J.; Lee, G.-W.; Park, S.-Y.; Huh, A.; Dean, R.A.; Lee, Y.-H. Genome-wide profiling of DNA methylation provides insights into epigenetic regulation of fungal development in a plant pathogenic fungus, Magnaporthe oryzae. Scientific reports 2015, 5, 8567–8567. [Google Scholar] [CrossRef] [PubMed]
- Montanini, B.; Chen, P.-Y.; Morselli, M.; Jaroszewicz, A.; Lopez, D.; Martin, F.; Ottonello, S.; Pellegrini, M. Non-exhaustive DNA methylation-mediated transposon silencing in the black truffle genome, a complex fungal genome with massive repeat element content. Genome Biol 2014, 15, 411–411. [Google Scholar] [CrossRef]
- Minervini, C.F.; Marsano, R.M.; Casieri, P.; Fanti, L.; Caizzi, R.; Pimpinelli, S.; Rocchi, M.; Viggiano, L. Heterochromatin protein 1 interacts with 5’UTR of transposable element ZAM in a sequence-specific fashion. Gene 2007, 393, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Shpiz, S.; Ryazansky, S.; Olovnikov, I.; Abramov, Y.; Kalmykova, A. Euchromatic transposon insertions trigger production of novel Pi- and endo-siRNAs at the target sites in the Drosophila germline. PLoS Genet 2014, 10, e1004138–e1004138. [Google Scholar] [CrossRef]
- Leung, D.C.; Dong, K.B.; Maksakova, I.A.; Goyal, P.; Appanah, R.; Lee, S.; Tachibana, M.; Shinkai, Y.; Lehnertz, B.; Mager, D.L.; et al. Lysine methyltransferase G9a is required for de novo DNA methylation and the establishment, but not the maintenance, of proviral silencing. Proc Natl Acad Sci U S A 2011, 108, 5718–5723. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Wang, L.; Bennett, B.D.; Wang, J.; Li, J.; Qin, Y.; Takaku, M.; Wade, P.A.; Wong, J.; Hu, G. Rif1 promotes a repressive chromatin state to safeguard against endogenous retrovirus activation. Nucleic Acids Res 2017, 45, 12723–12738. [Google Scholar] [CrossRef]
- Brennecke, J.; Aravin, A.A.; Stark, A.; Dus, M.; Kellis, M.; Sachidanandam, R.; Hannon, G.J. Discrete small RNA-generating loci as master regulators of transposon activity in Drosophila. Cell 2007, 128, 1089–1103. [Google Scholar] [CrossRef] [PubMed]
- Fagegaltier, D.; Bougé, A.-L.; Berry, B.; Poisot, É.; Sismeiro, O.; Coppée, J.-Y.; Théodore, L.; Voinnet, O.; Antoniewski, C. The endogenous siRNA pathway is involved in heterochromatin formation in Drosophila. Proceedings of the National Academy of Sciences 2009, 106, 21258–21263. [Google Scholar] [CrossRef] [PubMed]
- Gu, T.; Elgin, S.C.R. Maternal Depletion of Piwi, a Component of the RNAi System, Impacts Heterochromatin Formation in Drosophila. PLoS Genet 2013, 9, e1003780. [Google Scholar] [CrossRef] [PubMed]
- Pal-Bhadra, M.; Leibovitch, B.A.; Gandhi, S.G.; Chikka, M.R.; Bhadra, U.; Birchler, J.A.; Elgin, S.C. Heterochromatic silencing and HP1 localization in Drosophila are dependent on the RNAi machinery. Science 2004, 303, 669–672. [Google Scholar] [CrossRef]
- Conte, C.; Calco, V.; Desset, S.; Leblanc, P.; Dastugue, B.; Vaury, C. Impact of multiple insertions of two retroelements, ZAM and Idefix at an euchromatic locus. Genetica 2000, 109, 53–59. [Google Scholar] [CrossRef]
- Palazzo, A.; Marconi, S.; Specchia, V.; Bozzetti, M.P.; Ivics, Z.; Caizzi, R.; Marsano, R.M. Functional Characterization of the Bari1 Transposition System. PLoS One 2013, 8, e79385. [Google Scholar] [CrossRef]
- Palazzo, A.; Lovero, D.; D’Addabbo, P.; Caizzi, R.; Marsano, R.M. Identification of Bari Transposons in 23 Sequenced Drosophila Genomes Reveals Novel Structural Variants, MITEs and Horizontal Transfer. PLoS One 2016, 11, e0156014. [Google Scholar] [CrossRef]
- Tellier, M.; Bouuaert Corentin, C.; Chalmers, R. Mariner and the ITm Superfamily of Transposons. Microbiology Spectrum 2015, 3. [Google Scholar] [CrossRef]
- Moschetti, R.; Caggese, C.; Barsanti, P.; Caizzi, R. Intra- and Interspecies Variation Among Bari-1 Elements of the Melanogaster Species Group. Genetics 1998, 150, 239–250. [Google Scholar] [CrossRef]
- Davies, D.R.; Goryshin, I.Y.; Reznikoff, W.S.; Rayment, I. Three-dimensional structure of the Tn5 synaptic complex transposition intermediate. Science 2000, 289, 77–85. [Google Scholar] [CrossRef]
- Caizzi, R.; Caggese, C.; Pimpinelli, S. Bari-1, a new transposon-like family in Drosophila melanogaster with a unique heterochromatic organization. Genetics 1993, 133, 335–345. [Google Scholar] [CrossRef]
- Wu, C.I.; Lyttle, T.W.; Wu, M.L.; Lin, G.F. Association between a satellite DNA sequence and the Responder of Segregation Distorter in D. melanogaster. Cell 1988, 54, 179–189 pii. [Google Scholar] [CrossRef] [PubMed]
- Larracuente, A.M. The organization and evolution of the Responder satellite in species of the Drosophila melanogaster group: dynamic evolution of a target of meiotic drive. BMC Evol Biol 2014, 14, 233. [Google Scholar] [CrossRef]
- Caggese, C.; Pimpinelli, S.; Barsanti, P.; Caizzi, R. The distribution of the transposable element Bari-1 in the Drosophila melanogaster and Drosophila simulans genomes. Genetica 1995, 96, 269–283. [Google Scholar] [CrossRef] [PubMed]
- Marsano, R.M.; Milano, R.; Minervini, C.; Moschetti, R.; Caggese, C.; Barsanti, P.; Caizzi, R. Organization and possible origin of the Bari-1 cluster in the heterochromatic h39 region of Drosophila melanogaster. Genetica 2003, 117, 281–289. [Google Scholar] [CrossRef]
- McGurk, M.P.; Barbash, D.A. Double insertion of transposable elements provides a substrate for the evolution of satellite DNA. Genome Res 2018, 28, 714–725. [Google Scholar] [CrossRef] [PubMed]
- Palazzo, A.; Caizzi, R.; Moschetti, R.; Marsano, R.M. What Have We Learned in 30 Years of Investigations on Bari Transposons? Cells 2022, 11. [Google Scholar] [CrossRef]
- Marsano, R.M.; Marconi, S.; Moschetti, R.; Barsanti, P.; Caggese, C.; Caizzi, R. MAX, a novel retrotransposon of the BEL-Pao family, is nested within the Bari1 cluster at the heterochromatic h39 region of chromosome 2 in Drosophila melanogaster. Mol Genet Genomics 2004, 270, 477–484. [Google Scholar] [CrossRef]
- Moschetti, R.; Chlamydas, S.; Marsano, R.M.; Caizzi, R. Conserved motifs and dynamic aspects of the terminal inverted repeat organization within Bari-like transposons. Mol Genet Genomics 2008, 279, 451–461. [Google Scholar] [CrossRef]
- van Steensel, B.; Delrow, J.; Henikoff, S. Chromatin profiling using targeted DNA adenine methyltransferase. Nat Genet 2001, 27, 304–308. [Google Scholar] [CrossRef]
- Guio, L.; Vieira, C.; Gonzalez, J. Stress affects the epigenetic marks added by natural transposable element insertions in Drosophila melanogaster. Sci Rep 2018, 8, 12197. [Google Scholar] [CrossRef]
- Quenerch’du, E.; Anand, A.; Kai, T. The piRNA pathway is developmentally regulated during spermatogenesis in Drosophila. Rna 2016, 22, 1044–1054. [Google Scholar] [CrossRef] [PubMed]
- Hartl, D.L.; Ochman, H. Inverse polymerase chain reaction. Methods in molecular biology (Clifton, N J ) 1996, 58, 293–301. [Google Scholar] [CrossRef]
- Larkin, A.; Marygold, S.J.; Antonazzo, G.; Attrill, H.; dos Santos, G.; Garapati, P.V.; Goodman, Joshua L.; Gramates, L S.; Millburn, G.; Strelets, V.B.; et al. FlyBase: updates to the Drosophila melanogaster knowledge base. Nucleic Acids Res 2021, 49, D899–D907. [Google Scholar] [CrossRef] [PubMed]
- Karess, R.E.; Rubin, G.M. Analysis of P transposable element functions in Drosophila. Cell 1984, 38, 135–146. [Google Scholar] [CrossRef]
- Rubin, G.M.; Spradling, A.C. Genetic transformation of Drosophila with transposable element vectors. Science 1982, 218, 348–353. [Google Scholar] [CrossRef] [PubMed]
- Eissenberg, J.C.; Morris, G.D.; Reuter, G.; Hartnett, T. The heterochromatin-associated protein HP-1 is an essential protein in Drosophila with dosage-dependent effects on position-effect variegation. Genetics 1992, 131, 345–352. [Google Scholar] [CrossRef]
- Reuter, G.; Dorn, R.; Wustmann, G.; Friede, B.; Rauh, G. Third chromosome suppressor of position-effect variegation loci in Drosophila melanogaster. Molecular and General Genetics MGG 1986, 202, 481–487. [Google Scholar] [CrossRef]
- Ashburner, M. Drosophila: A laboratory manual; Cold Spring Harbor Laboratory, 1989. [Google Scholar]
- Kennison, J.A. Dissection of Larval Salivary Glands and Polytene Chromosome Preparation. Cold Spring Harbor Protocols 2008, 2008, pdb.prot4708. [Google Scholar] [CrossRef]
- Henikoff, S.; Dreesen, T.D. Trans-inactivation of the Drosophila brown gene: evidence for transcriptional repression and somatic pairing dependence. Proc Natl Acad Sci U S A 1989, 86, 6704–6708. [Google Scholar] [CrossRef]
- Herold, N.; Will, C.L.; Wolf, E.; Kastner, B.; Urlaub, H.; Luhrmann, R. Conservation of the protein composition and electron microscopy structure of Drosophila melanogaster and human spliceosomal complexes. Mol Cell Biol 2009, 29, 281–301. [Google Scholar] [CrossRef]
- Burkhart, B.D.; Montgomery, E.; Langley, C.H.; Voelker, R.A. Characterization of Allozyme Null and Low Activity Alleles from Two Natural Populations of Drosophila melanogaster. Genetics 1984, 107, 295–306. [Google Scholar] [CrossRef]
- Buels, R.; Yao, E.; Diesh, C.M.; Hayes, R.D.; Munoz-Torres, M.; Helt, G.; Goodstein, D.M.; Elsik, C.G.; Lewis, S.E.; Stein, L.; et al. JBrowse: a dynamic web platform for genome visualization and analysis. Genome Biol 2016, 17, 66. [Google Scholar] [CrossRef]
- Kharchenko, P.V.; Alekseyenko, A.A.; Schwartz, Y.B.; Minoda, A.; Riddle, N.C.; Ernst, J.; Sabo, P.J.; Larschan, E.; Gorchakov, A.A.; Gu, T.; et al. Comprehensive analysis of the chromatin landscape in Drosophila melanogaster. Nature 2011, 471, 480–485. [Google Scholar] [CrossRef]
- Filion, G.J.; van Bemmel, J.G.; Braunschweig, U.; Talhout, W.; Kind, J.; Ward, L.D.; Brugman, W.; de Castro, I.J.; Kerkhoven, R.M.; Bussemaker, H.J.; et al. Systematic Protein Location Mapping Reveals Five Principal Chromatin Types in Drosophila Cells. Cell 2010, 143, 212–224. [Google Scholar] [CrossRef]
- Donlin, M.J. Using the Generic Genome Browser (GBrowse). Current Protocols in Bioinformatics 2007, 17, 9.9.1–9.9.24. [Google Scholar] [CrossRef] [PubMed]
- Tritto, P.; Palumbo, V.; Micale, L.; Marzulli, M.; Bozzetti, M.P.; Specchia, V.; Palumbo, G.; Pimpinelli, S.; Berloco, M. Loss of Pol32 in Drosophila melanogaster Causes Chromosome Instability and Suppresses Variegation. PLoS One 2015, 10, e0120859. [Google Scholar] [CrossRef]
- Logsdon, G.A.; Vollger, M.R.; Eichler, E.E. Long-read human genome sequencing and its applications. Nature Reviews Genetics 2020, 21, 597–614. [Google Scholar] [CrossRef] [PubMed]
- Stark, R.; Grzelak, M.; Hadfield, J. RNA sequencing: the teenage years. Nature Reviews Genetics 2019, 20, 631–656. [Google Scholar] [CrossRef] [PubMed]
- Xie, N.; Zhou, Y.; Sun, Q.; Tang, B. Novel Epigenetic Techniques Provided by the CRISPR/Cas9 System. Stem Cells Int 2018, 2018, 7834175–7834175. [Google Scholar] [CrossRef]
- Caizzi, R.; Moschetti, R.; Piacentini, L.; Fanti, L.; Marsano, R.M.; Dimitri, P. Comparative Genomic Analyses Provide New Insights into the Evolutionary Dynamics of Heterochromatin in Drosophila. PLoS Genet 2016, 12, e1006212. [Google Scholar] [CrossRef] [PubMed]
- Clark, D.V.; Sabl, J.F.; Henikoff, S. Repetitive arrays containing a housekeeping gene have altered polytene chromosome morphology in Drosophila. Chromosoma 1998, 107, 96–104. [Google Scholar] [CrossRef]
- Eichten, S.R.; Ellis, N.A.; Makarevitch, I.; Yeh, C.-T.; Gent, J.I.; Guo, L.; McGinnis, K.M.; Zhang, X.; Schnable, P.S.; Vaughn, M.W.; et al. Spreading of Heterochromatin Is Limited to Specific Families of Maize Retrotransposons. PLoS Genet 2012, 8, e1003127. [Google Scholar] [CrossRef] [PubMed]
- Chlamydas, S.; Heun, P.; Dimitri, P.; Moschetti, R.; Barsanti, P.; Caizzi, R. The paracentric inversion In(2Rh)PL alters the centromeric organization of chromosome 2 in Drosophila melanogaster. Chromosome Res 2009, 17, 1–9. [Google Scholar] [CrossRef]
- Moshkin, Y.M.; Alekseyenko, A.A.; Semeshin, V.F.; Spierer, A.; Spierer, P.; Makarevich, G.F.; Belyaeva, E.S.; Zhimulev, I.F. The Bithorax Complex of Drosophila melanogaster: Underreplication and morphology in polytene chromosomes. Proceedings of the National Academy of Sciences 2001, 98, 570–574. [Google Scholar] [CrossRef]
- Kim, J.H.; Ebersole, T.; Kouprina, N.; Noskov, V.N.; Ohzeki, J.; Masumoto, H.; Mravinac, B.; Sullivan, B.A.; Pavlicek, A.; Dovat, S.; et al. Human gamma-satellite DNA maintains open chromatin structure and protects a transgene from epigenetic silencing. Genome Res 2009, 19, 533–544. [Google Scholar] [CrossRef]
- Bzymek, M.; Lovett, S.T. Instability of repetitive DNA sequences: the role of replication in multiple mechanisms. Proc Natl Acad Sci U S A 2001, 98, 8319–8325. [Google Scholar] [CrossRef]
- Heartlein, M.W.; Knoll, J.H.; Latt, S.A. Chromosome instability associated with human alphoid DNA transfected into the Chinese hamster genome. Mol Cell Biol 1988, 8, 3611–3618. [Google Scholar] [CrossRef]
- Stults, D.M.; Killen, M.W.; Williamson, E.P.; Hourigan, J.S.; Vargas, H.D.; Arnold, S.M.; Moscow, J.A.; Pierce, A.J. Human rRNA gene clusters are recombinational hotspots in cancer. Cancer Res 2009, 69, 9096–9104. [Google Scholar] [CrossRef]
- Warmerdam, Daniël O.; van den Berg, J.; Medema, René H. Breaks in the 45S rDNA Lead to Recombination-Mediated Loss of Repeats. Cell Reports 2016, 14, 2519–2527. [Google Scholar] [CrossRef] [PubMed]
- Depienne, C.; Mandel, J.L. 30 years of repeat expansion disorders: What have we learned and what are the remaining challenges? Am J Hum Genet 2021, 108, 764–785. [Google Scholar] [CrossRef]
- Houtchens, K.; Lyttle, T.W. Responder (Rsp) Alleles in the Segregation Distorter (SD) System of Meiotic Drive in Drosophila may Represent a Complex Family of Satellite Repeat Sequences. Genetica 2003, 117, 291–302. [Google Scholar] [CrossRef]
- Penagos-Puig, A.; Furlan-Magaril, M. Heterochromatin as an Important Driver of Genome Organization. Front Cell Dev Biol 2020, 8, 579137. [Google Scholar] [CrossRef] [PubMed]
- Palazzo, A.; Moschetti, R.; Caizzi, R.; Marsano, R.M. The Drosophila mojavensis Bari3 transposon: distribution and functional characterization. Mob DNA 2014, 5, 21. [Google Scholar] [CrossRef] [PubMed]
- Di Franco, C.; Terrinoni, A.; Dimitri, P.; Junakovic, N. Intragenomic Distribution and Stability of Transposable Elements in Euchromatin and Heterochromatin of Drosophila melanogaster: Elements with Inverted Repeats Bari 1, hobo, and pogo. J Mol Evol 1997, 45, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Westphal, T.; Reuter, G. Recombinogenic Effects of Suppressors of Position-Effect Variegation in Drosophila. Genetics 2002, 160, 609. [Google Scholar] [CrossRef]


| Construct | Injected embryos | G0 Adults | G0 Fertile individuals | G0 Sterile individuals | G0 w+ individuals | Progeny scored | Total w+ lines | X-linked |
Autosomal II III |
|
| pCasp-Ba7 | 186 | 71 | 63 | 8 | 18 | 20216 | 19* | 5* | 8 | 6 |
| pCasp-Ba3 | 220 | 112 | 75 | 37 | 17 | 16151 | 20 | 5 | 7 | 8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



